
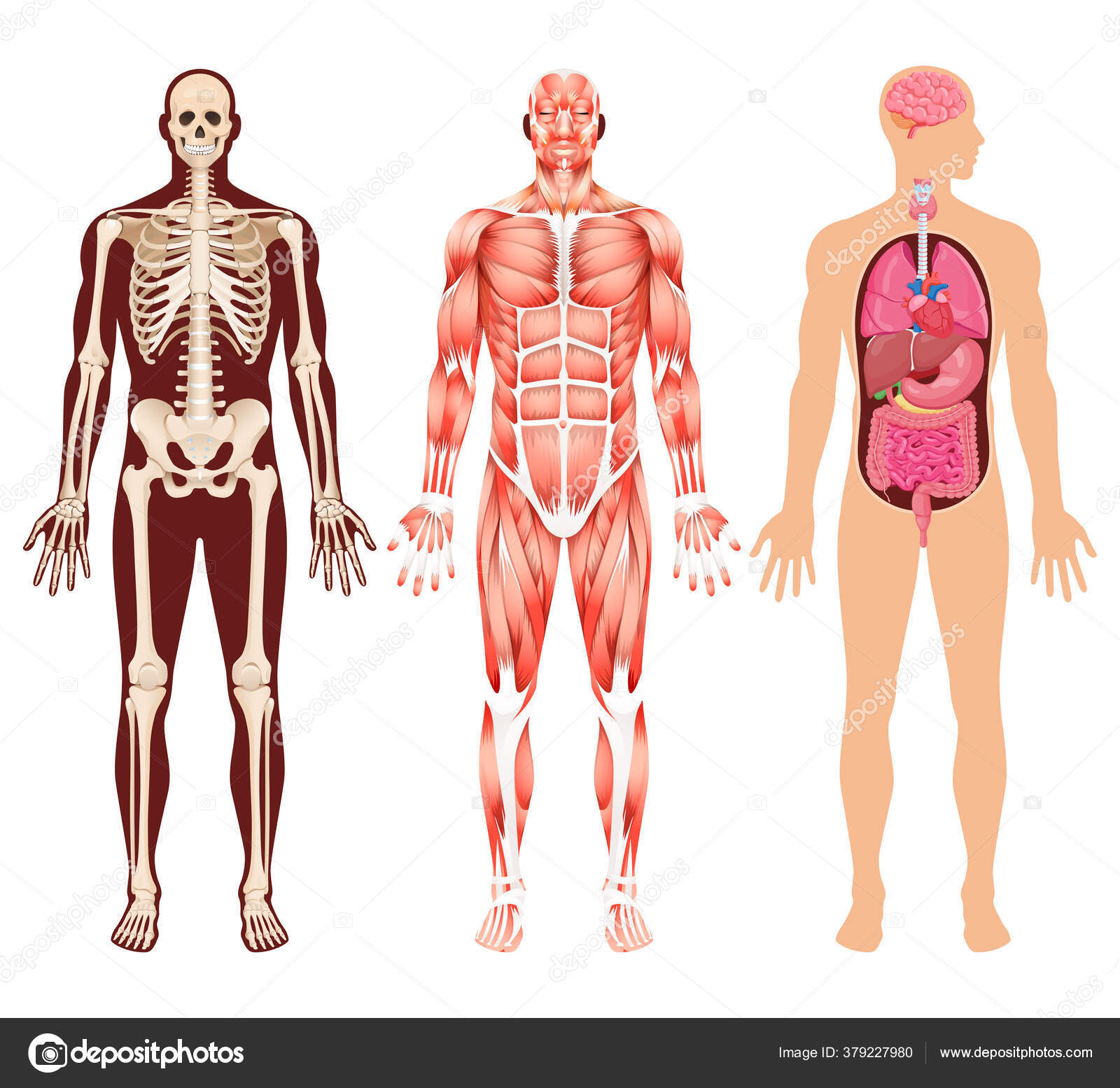
App Store: Скелет 3D анатомии
Bстроенные покупки (Полная версия)
«Скелет — 3D Атлас анатомии» – это атлас анатомии нового поколения в 3D, в котором вы найдете интерактивные и очень подробные анатомические модели!
Каждая кость человеческого скелета была реконструирована в 3D, вы можете вращать и приближать каждую модель, а также подробно рассматривать модели под любым углом.
При выборе моделей или значков вам будут показаны термины, относящиеся к любой специфической анатомической части, вы можете выбрать из 11 языков и показ терминов на двух языках одновременно.
«Скелет» – это полезный инструмент для студентов, которые получают образование в сфере медицины и физической культуры, для терапевтов, ортопедов, физиотерапевтов, психотерапевтов, медработников, кинезиологов, медсестер и спортивные тренеров.
ОЧЕНЬ ПОДРОБНЫЕ АНАТОМИЧЕСКИЕ 3D-МОДЕЛИ
• Костная система
• Точное 3D-моделирование
• Поверхность скелета с высоким разрешением текстур до стандарта 4K
ПРОСТОЙ И ПОНЯТНЫЙ ИНТЕРФЕЙС.
• Вращайте и приближайте каждую модель в трехмерном пространстве
• Разделение по районам для понятной и быстрой визуализации каждой структуры
• Умное вращение, автоматически двигает центр вращения для более легкой навигации
• Интерактивный значок позволяет визуализировать термин, который имеет отношение к каждой анатомической детали
• Скрыть / Показать интерфейс, идеально подходит для использования на смартфонах
МНОГОЯЗЫЧНОСТЬ
• Анатомические термины и интерфейс доступны 11 языках: Латинский, английский, французский, немецкий, итальянский, португальский, русский, китайский, японский и корейский
• Язык может быть выбран непосредственно через интерфейс приложения
• Анатомические термины могут быть показаны на двух языках одновременно
«Скелет» – это часть коллекции приложений для изучения анатомии человека «3D Atlas of Anatomy», новых приложений и обновлений, которые были разработаны.
Опорно-двигательная система. Скелет человека — урок.
 Окружающий мир, 3 класс.
Окружающий мир, 3 класс.Внутри нашего тела находится скелет. К нему прикрепляются мышцы.
Скелет и мышцы составляют опорно-двигательную систему.
Эта система:
- служит опорой телу;
- защищает внутренние органы;
- позволяет нашему организму двигаться.
Скелет
В скелете человека более \(200\) костей. У всех костей есть свои названия. Кости твёрдые и прочные. Каждая имеет определённые размеры и форму.
Скелет:
- является опорой тела;
- поддерживает форму тела;
- защищает все внутренние органы от повреждений.
Кости в скелете соединены так, что при движении могут изменять своё положение.
Заставляют кости двигаться прикреплённые к ним мышцы.
В скелете человека выделяют череп, позвоночник, грудную клетку, таз, верхние конечности (руки) и нижние конечности (ноги).
Череп защищает головной мозг от повреждений.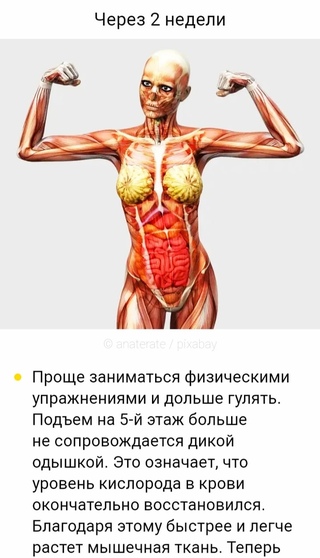 Он образован твёрдыми и прочными костями.
Он образован твёрдыми и прочными костями.
Позвоночник является главной опорой нашего тела. Без позвоночника человек не смог бы стоять и ходить. Позвоночник состоит из \(33\) позвонков. Позвонки имеют отверстия, которые образуют позвоночный канал. В нём расположен спинной мозг.
Грудная клетка образована рёбрами и надёжно защищает сердце и лёгкие.
К грудной клетке при помощи ключиц и лопаток прикреплены кости верхних конечностей — рук. Кости рук соединены подвижно. Поэтому мы можем их поднимать, опускать, сгибать в локтях и выполнять много других движений.
К нижней части позвоночника присоединены кости таза. Таз поддерживает и защищает внутренние органы. С тазом подвижно соединены кости нижних конечностей — ног. dbbdcQWU\b`KMLIEFxzyDDDgefKKK@DClpoHHH?CB?CBFJICEDBDCNNNY]\AEDLLLKKK[[[GKJAGEEIHAGE?ECNRQ;;;}{|6
dbbdcQWU\b`KMLIEFxzyDDDgefKKK@DClpoHHH?CB?CBFJICEDBDCNNNY]\AEDLLLKKK[[[GKJAGEEIHAGE?ECNRQ;;;}{|6
\\\JJJ777iiiaaa9=___465CGFQSR064JLK777-31HHHPRQ.0/jhi &$sop%»GGG 043576HLKJJJPNO yyySSS )'(AGE :::#»QWU
Анатомия. Скелет и мышцы — презентация онлайн
1. Анатомия.
Автор: Кириллов Александр ЮрьевичАнатомия.
СКЕЛЕТ И МЫШЦЫ.
2. Скелет человека.
4. Живая кость.
5. Схема строения скелета человека.
Скелет головы(череп)
Скелет верхних конечностей
Свободная верхняя
конечность
Плечевой
пояс
Мозговой
череп
Свод
черепа
Лицевой
череп
Ключица
Лопатка
Плечи
Предплечье
Основание
черепа
Кисть
Запястье
Пясть
Локтевая
Лучевая
Скелет нижних конечностей
Скелет туловища
Тазовый пояс
Позвоночный
столб
(33-34 позвонка)
Грудная клетка
(грудина,
12 пар рёбер)
Свободная нижняя конечность
Тазовая кость
Бедро
Голень
Стопа
Подвздошная
Лобковая
Седалищная
Бедренная
кость
Большеберцовая
Малоберцовая
Предплюсна
Плюсна
Фаланги
6.
 Рост костей. Гормон роста –
Рост костей. Гормон роста –соматотропный гормон,
вырабатывается в
гипофизе.
7. Остеопороз.
Рост и прочность костейопределяется интенсивностью
деятельности
прикрепляющихся к ним мышц.
(П. Ф. Лесгафт)
9. Остеофитоз.
10. Кости.
Плечевая кость: большой и малый бугорки,межбугорковая борозда, медиальный и латеральный
мыщелки, борозда лучевого нерва, блок плечевой кости,
дельтовидная бугристость.
11. Связки.
Паховаясвязка.
12. Виды суставов.
Соединениекостей.
Синартрозы
(не подвижные)
Синдесмозы
(соединит.
ткань)
Синхондрозы
(хрящи)
Гемиартрозы
(полуподвижные)
Диартрозы
(подвижные)
Синостозы
(костная
ткань)
Суставы
Лучезапястный
Одноосные
Двуосные
Многоосные
Цилиндрический
Блоковидный
Элипсовидный
Мыщелковый
Седловидный
Шаровидный
Плоский
(элипсовидный)
Запястно-пястный
сустав большого
пальца
(седловидный)
Суставы.

14. Коленный сустав.
Вид сустава:мыщелковый.
15. Мышцы
СТРОЕНИЕ И ФУНКЦИИ.17. Строение мышечного волокна.
18. Сокращение мышц.
19. Классификация мышц.
20. Мышцы шеи.
Короткие подзатылочные мышцы.Пальпация передних
сгибателей шеи.
Мышцы туловища.
24. Мышцы конечностей.
моделей человеческого скелета | Каролина.com
Когда вы покупаете модели человеческого скелета в Каролине, вы получаете 100% гарантию качества. Наши модели человеческого скелета анатомически точны и легко моются водой с мылом. Мы предлагаем в натуральную величину, настольные, сочлененные и разъединенные модели человеческого скелета для углубленного изучения каждой человеческой кости. Покупайте бренды, которые вы знаете и которым доверяете: Somso®, American 3B Scientific®, Altay® и другие.
- 60120240300За страницу
- Сортировать по Мы рекомендуемНовые поступленияБестселлерыЦена, от низкой до высокойЦена, от высокой до низкойНазвание — A-ZНазвание — Z-A
- 3B® Расчлененный человеческий скелет 2 продукта 547 долларов.
 00 — 681,50 долл. США Быстрый просмотр
00 — 681,50 долл. США Быстрый просмотр - Разобранный человеческий скелет 3B® с футляром для хранения Товар № 246777 $681,50 Быстрый просмотр
- Алтайская экономика Разобранный человеческий скелет Товар № 246765 436,00 долларов США Быстрый просмотр
- Разобранный полускелет человека Altay® Товар № 246728 353 доллара.70 Быстрый просмотр
- Скелет человека 3B® с опорой на стержень Товар № 246875 497,00 долларов США Быстрый просмотр
- Гибкий скелет человека 3B® Товар № 246877 $816,00 Быстрый просмотр
- 3B® Скелет человека, зависание Товар № 246809 625 долларов.00 Быстрый просмотр
- 3B® Мышечно-связочный скелет человека Товар № 246896 1228,50 долларов США Быстрый просмотр
- Экономичный настольный миниатюрный скелет Товар № 246915 Быстрый просмотр
- Мужской скелет человека Somso®, подвесной Товар № 246814 2600 долларов. 00 Быстрый просмотр
- Altay® Патология модели щитовидной железы человека Товар № 566746 $68,70 Быстрый просмотр
- Модель коленного сустава Алтай Эконом Товар № 247932 105,60 долларов США Быстрый просмотр
- Алтайский человеческий скелет нижней конечности с половиной таза Товар № 247930 109 долларов.70 Быстрый просмотр
- Модель скелетной руки человека Carolina® Товар № 569415 $83,00 Быстрый просмотр
- Набор «Скелет человека 3B®» Товар № 246875C $599,00 Быстрый просмотр
- Скелет человека Somso®, зависание 2 продукта 2600 долларов.00 — 2645,00 долларов США Быстрый просмотр
- Набор человеческих позвонков 3B®, 5 на нити Товар № 247536 $80.00 Быстрый просмотр
- 3B® Человеческая рука и кисть Товар № 247706 146,00 долларов США Быстрый просмотр
- Скелет человека Somso® с опорой на стержень 2 продукта 2150 долларов. 00 — 2165,00 долларов США Быстрый просмотр
- 3B® Стопа человека Товар № 247906 $85.00 Быстрый просмотр
- Модель плечевого сустава человека Carolina® Товар № 569406 105,00 долларов США Быстрый просмотр
- Расчлененный человеческий скелет Somso® 3 продукта 1580 долларов.00 — 1735,00 долларов США Быстрый просмотр
- GPI Anatomicals® Миниатюрный набор мускулистых суставов Товар № 247682 147,00 долларов США Быстрый просмотр
- 3B® Мышечный скелет человека Товар № 246891 1035,00 долларов США Быстрый просмотр
- Модель скелетной стопы человека Carolina® Товар № 569416 83 доллара.00 Быстрый просмотр
- Футляр для хранения человеческого скелета, черный пластик Товар № 249890A 165,00 долларов США Быстрый просмотр
- Altay® Настольный миниатюрный скелет Товар № 246925 246,00 долларов США Быстрый просмотр
- Модель локтевого сустава человека Товар № 247712 111 долларов.
- 3B® Разобранный полускелет человека Товар № 246725 459,00 долларов США Быстрый просмотр
- Скелетная рука Somso® Товар № 247684 158,00 долларов США Быстрый просмотр
- Экономика Разъединенный мышечный скелет человека Товар № 246776 709 долларов.30 Быстрый просмотр
- Скелетная стопа Somso® Товар № 247904 137,00 долларов США Быстрый просмотр
- Таз мужчины 3B® человека Товар № 247755 $89,50 Быстрый просмотр
- Altay® Крестец и копчик человека Товар № 566706 30 долларов.80 Быстрый просмотр
- Модель тазобедренного сустава человека Carolina® Товар № 569404 109,00 долларов США Быстрый просмотр
- Мышечный скелет человека Somso® Товар № 246882A 4075,00 долларов США Быстрый просмотр
- 3B® Расчлененный человеческий скелет Товар № 246775 547 долларов. 00 Быстрый просмотр
- Altay® Человеческие ребра с одной стороны, скелет, набор из 12 шт. Товар № 566738 $56,20 Быстрый просмотр
- Таз человека Altay® с нижними поясничными и бедренными головками Товар № 566737 $63,60 Быстрый просмотр
- Модель позвоночника человека Carolina® Miniature с головками бедренных костей Товар № 569414 110 долларов.00 Быстрый просмотр
- Колонка гибких позвонков Altay Economy Human, цветовая маркировка Товар № 247572 214,00 долларов США Быстрый просмотр
- Набор моделей патологии позвонков человека GPI Anatomicals®, 4 предмета Товар № 569200 205,00 долларов США Быстрый просмотр
- Модель стоматологического кариеса Altay® Товар № 566736 59 долларов.50 Быстрый просмотр
- Мышечный скелет человека Altay® Economy, с опорой на стержень Товар № 246887 $767,00 Быстрый просмотр
- Алтайский атлас, ось и набор затылочных костей Товар № 247535 47,20 долларов США Быстрый просмотр
- Головоломка со скелетом Товар № 246171 31 доллар. 25 Быстрый просмотр
- Алтайские кости кисти и предплечья Товар № 247705 $76,90 Быстрый просмотр
- Altay® Human Radius Skeleton Товар № 566739 $19,30 Быстрый просмотр
- Скелет человека, разобранный на части Somso®, с футляром для хранения, без числового кода Товар № 246787 1589 долларов.00 Быстрый просмотр
- Экономичный человеческий череп Товар № 566731 $94,00 Быстрый просмотр
- Алтайский гибкий позвоночник с головками бедренных костей и прикреплением мышц Товар № 247573 $184,50 Быстрый просмотр
- Разобранный мышечный скелет человека 3B® с футляром для хранения Товар № 246782 1040 долларов.00 Быстрый просмотр
- Мужской скелет человека Somso® с опорой на стержень Товар № 246867 2150,00 долларов США Быстрый просмотр
- Мышечные скелеты человека Altay® Economy 2 продукта 767,00–801,60 долларов США Быстрый просмотр
- Somso® Прозрачный футляр для хранения Товар № 247050 449 долларов. 00 Быстрый просмотр
- Altay® Economy Human Muscular Skeleton, Hang-Up Товар № 246885 $801,60 Быстрый просмотр
- Функциональный плечевой сустав Somso® Товар № 247709 409,00 долларов США Быстрый просмотр
- 3B® Скелет физиологии человека Товар № 246877A 1251 доллар.00 Быстрый просмотр
- Нога и стопа человека 3B® Товар № 247931 177,00 долларов США Быстрый просмотр
- Модели суставов человека Carolina® 4 продукта 102 – 109 долларов США Быстрый просмотр
 00 — 681,50 долл. США Быстрый просмотр
00 — 681,50 долл. США Быстрый просмотр 00 Быстрый просмотр
00 Быстрый просмотр 00 — 2165,00 долларов США Быстрый просмотр
00 — 2165,00 долларов США Быстрый просмотр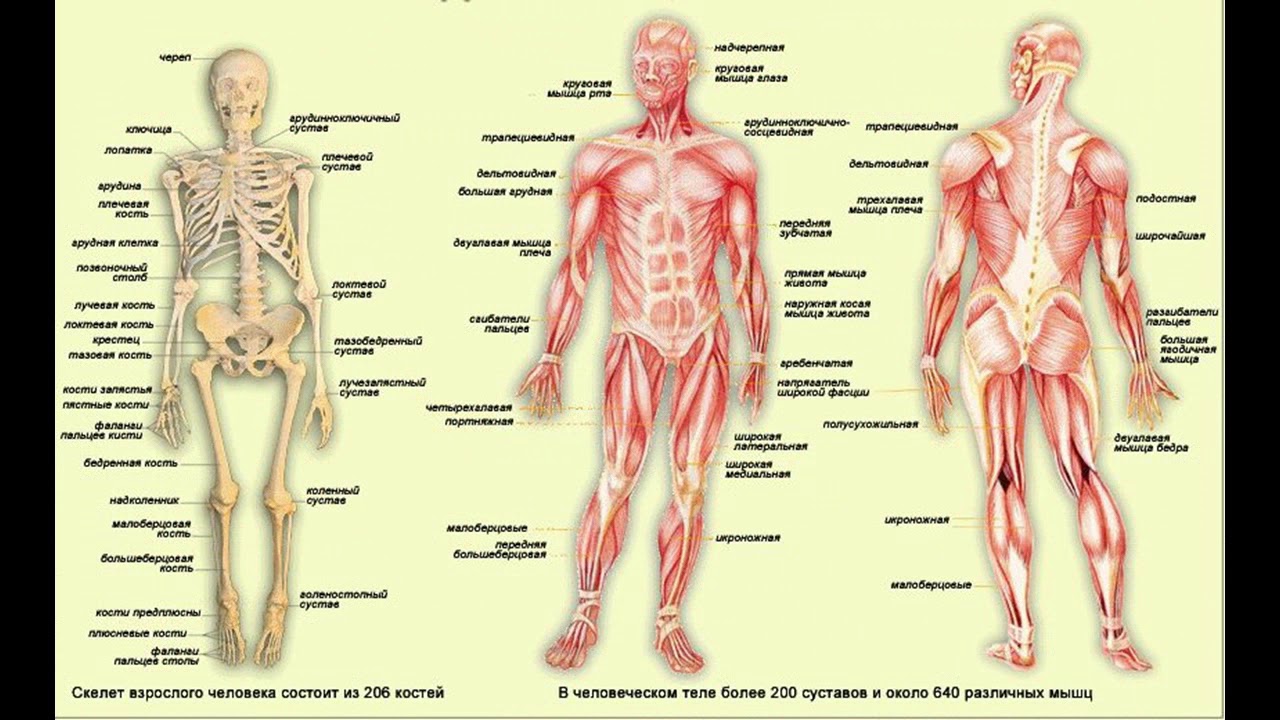
 00 Быстрый просмотр
00 Быстрый просмотр 25 Быстрый просмотр
25 Быстрый просмотр 00 Быстрый просмотр
00 Быстрый просмотр- 60120240300За страницу
- Сортировать по Мы рекомендуемНовые поступленияБестселлерыЦена, от низкой до высокойЦена, от высокой до низкойНазвание — A-ZНазвание — Z-A
Биохимическое разнообразие скелетных мышц человека | Журнал экспериментальной биологии
Данные представлены как среднее значение ± с. Эм. если не указано иное. Уровень значимости (α) был установлен на 0,05, и все статистические тесты проводились с использованием SPSS (версия 19, IBM, Armonk, NY, USA) или Matlab (Mathworks, Natick, MA, USA). Данные были проверены на нормальность (убедившись, что перекос и эксцесс находятся в диапазоне от –1,0 до 1,0), чтобы оправдать использование параметрических статистических тестов. Данные OH-P требовали использования логарифмического преобразования для восстановления приемлемого перекоса и эксцесса до <|1,0|. Для проверки различий между группами использовали одно- и двусторонний дисперсионный анализ (ANOVA).Мы измерили ряд переменных для каждой мышцы, но взаимосвязи (если есть) между переменными и анатомической организацией (если есть) неизвестны. Таким образом, мы использовали многофакторный метод пошагового дискриминантного функционального анализа (DFA) для определения переменных, которые лучше всего предсказывают группу мышц, используя F -ввести=3,84 и F -удалить=2,71.
Эм. если не указано иное. Уровень значимости (α) был установлен на 0,05, и все статистические тесты проводились с использованием SPSS (версия 19, IBM, Armonk, NY, USA) или Matlab (Mathworks, Natick, MA, USA). Данные были проверены на нормальность (убедившись, что перекос и эксцесс находятся в диапазоне от –1,0 до 1,0), чтобы оправдать использование параметрических статистических тестов. Данные OH-P требовали использования логарифмического преобразования для восстановления приемлемого перекоса и эксцесса до <|1,0|. Для проверки различий между группами использовали одно- и двусторонний дисперсионный анализ (ANOVA).Мы измерили ряд переменных для каждой мышцы, но взаимосвязи (если есть) между переменными и анатомической организацией (если есть) неизвестны. Таким образом, мы использовали многофакторный метод пошагового дискриминантного функционального анализа (DFA) для определения переменных, которые лучше всего предсказывают группу мышц, используя F -ввести=3,84 и F -удалить=2,71.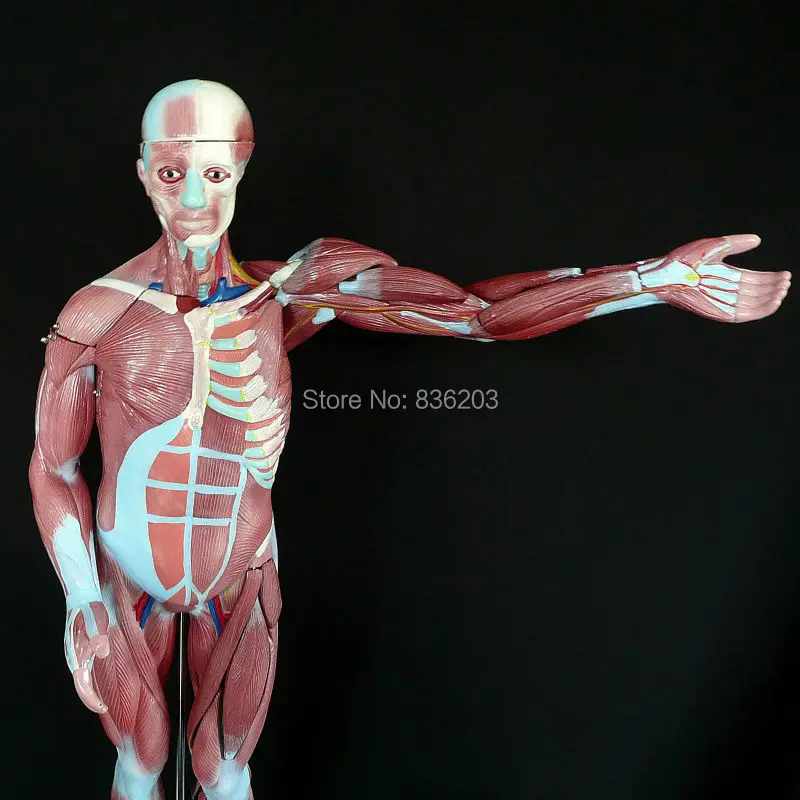 Эти пределы значений F использовались, поскольку они являются настройками по умолчанию для DFA в SPSS. Для DFA мышцы были классифицированы по анатомической области (аксиальные N =9, плечевые N =13, плечевые N =7, переднеплечевые N =20, кисти N =13, тазовые/ягодичные N =10, бедро N =15, нога N =13, по основной функции (сгибатели N =29, разгибатели N =34, отводящие N =9, приводящие N =1 , внутренний ротатор N =7, внешний ротатор N =8), по антигравитационному ( N =50) по сравнению с неантигравитационным ( N =50) состоянию и по скрещенным суставам (одиночный N =48, кратное N =52).Полный список мышц и их классификация доступны в дополнительном материале Таблица S1. Случаи без достоверных данных для всех независимых переменных не могут быть включены в DFA, поэтому размер выборки для DFA составил 586.
Эти пределы значений F использовались, поскольку они являются настройками по умолчанию для DFA в SPSS. Для DFA мышцы были классифицированы по анатомической области (аксиальные N =9, плечевые N =13, плечевые N =7, переднеплечевые N =20, кисти N =13, тазовые/ягодичные N =10, бедро N =15, нога N =13, по основной функции (сгибатели N =29, разгибатели N =34, отводящие N =9, приводящие N =1 , внутренний ротатор N =7, внешний ротатор N =8), по антигравитационному ( N =50) по сравнению с неантигравитационным ( N =50) состоянию и по скрещенным суставам (одиночный N =48, кратное N =52).Полный список мышц и их классификация доступны в дополнительном материале Таблица S1. Случаи без достоверных данных для всех независимых переменных не могут быть включены в DFA, поэтому размер выборки для DFA составил 586. Линейная регрессия использовалась для определения взаимосвязи между массой тайтина и процентным содержанием MHC.
Линейная регрессия использовалась для определения взаимосвязи между массой тайтина и процентным содержанием MHC.
Скелетная мышца, сконструированная из ткани человека: новая 3D-модель in vitro для определения распределения и токсичности лекарств после внутримышечной инъекции
Discher, D. E., Джанми П. и Ван Ю.-Л. Тканевые клетки чувствуют и реагируют на жесткость своего субстрата. Наука 310 , 1139–43 (2005).
ОБЪЯВЛЕНИЕ Статья пабмед КАС Google Scholar
Фалькенберг, Н. и др. . Трехмерные микроткани существенно способствуют доклиническим проверкам терапевтических мишеней при раке молочной железы. Рак Мед . 703–710 (2016).
Шмейхель, К. Л. Моделирование тканеспецифической передачи сигналов и функций органов в трех измерениях. J. Cell Sci. 116 , 2377–2388 (2003).
Артикул пабмед ПабМед Центральный КАС Google Scholar

Bell, C.C. и др. . Характеристика первичных сфероидов гепатоцитов человека как модельной системы для лекарственного поражения печени, функции печени и заболевания. науч. Респ. 6 , 25187 (2016).
ОБЪЯВЛЕНИЕ Статья пабмед ПабМед Центральный КАС Google Scholar
Эш, М. Б., Кинг, Т. Л. и Шулер, М. Л. Роль устройств типа «тело-на-чипе» в исследованиях лекарств и токсичности. год. Преподобный Биомед. англ. 13 , 55–72 (2011).
Артикул пабмед КАС Google Scholar
Кола, И. и Лэндис, Дж. Может ли фармацевтическая промышленность снизить процент отсева? Нац.Преподобный Друг Дисков. 3 , 711–715 (2004).
Артикул пабмед КАС Google Scholar
Олсон Х. и др. . Согласованность токсичности лекарственных средств для человека и животных. Регул. Токсикол. Фармакол. 32 , 56–67 (2000).
Регул. Токсикол. Фармакол. 32 , 56–67 (2000).
Артикул пабмед КАС Google Scholar
Дуйк Г.Истирание и перевод. Наука 302 , 603–605 (2003).
ОБЪЯВЛЕНИЕ Статья пабмед КАС Google Scholar
Козигу Г., Крозье Дж., Хендриксен К., Ману И. и Рамирез-Эрнандес Т. Европейское партнерство по альтернативным подходам к тестированию на животных (EPAA): продвижение альтернативных методов в Европе и далее. J Am Assoc Lab Anim Sci. 54 , 209–213 (2015).
ПабМед ПабМед Центральный Google Scholar
Юм К., Хонг С. Г. и Ли Л. П. Физиологически значимые органы на чипсах. Biotechnol J. 9 , 16–27 (2015).
Артикул КАС Google Scholar
Лангер, Р. и Ваканти, Дж. Тканевая инженерия. Наука 260 , 920–926 (1993).
и Ваканти, Дж. Тканевая инженерия. Наука 260 , 920–926 (1993).
ОБЪЯВЛЕНИЕ Статья пабмед КАС Google Scholar
Макрис, Э. А., Гомолл, А. Х., Мализос, К. Н., Ху, Дж. К. и Афанасиу, К. А. Методы восстановления и тканевой инженерии суставного хряща. Нац. Преподобный Ревматол. 11 , 21–34 (2014).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Чжан, Б.Г.Х. и др. . Последние достижения в области инженерии нервной ткани. Междунар. Дж. Артиф. Органы 37 , 277–291 (2014).
Артикул пабмед КАС Google Scholar
Вайнбергер, Ф., Маннхардт, И. и Эшенхаген, Т. Инженерия сердечной мышечной ткани. Обр. Рез. 120 , 1487–1500 (2017).
Артикул пабмед КАС Google Scholar

Klumpp, D., Horch, R.E. & Beier, J.P. In Tissue Engineering of Skeletal Muscle, Tissue Engineering for Tissue and Organ Regeneration (ed.Эберли, П. Д.) (InTech, 2011).
Клар А.С., Зимох Дж. и Бидерманн Т. Инженерия кожных тканей: применение стволовых клеток, полученных из жировой ткани. Биомед Рез. . 1–12 (2017).
Палаккан А.А., Дэвид Х., Анил К., Кумари Т. и Джеймс Р. Тканевая инженерия печени и источники клеток: проблемы и проблемы. Печень, внутр. 33 , 666–676 (2013).
Артикул пабмед КАС Google Scholar
Henkel, J. и др. . Костная регенерация на основе концепций тканевой инженерии — перспектива 21 века. Кость Res. 1 , 216–248 (2013).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Chua, A.W.C. и др. . Достижения инженерии тканей кожи при тяжелых ожогах: обзор и терапевтическое применение. Сжечь. Травма 4 , 3 (2016).
Достижения инженерии тканей кожи при тяжелых ожогах: обзор и терапевтическое применение. Сжечь. Травма 4 , 3 (2016).
Артикул Google Scholar
Бенам, К. Х. и др. . Разработано In Vitro моделей заболеваний. год. Преподобный Патол. мех. Дис. 10 , 195–262 (2015).
Артикул КАС Google Scholar
Spielmann, H., Hoffmann, S., Liebsch, M., Botham, P. & Fentem, J. Международное валидационное исследование ECVAM тестов in vitro на острое раздражение кожи: отчет о достоверности анализов EPISKIN и EpiDerm и функционального теста целостности кожи. Альтернативная лаборатория анимации. 35 , 559–601 (2007).
ПабМед КАС Google Scholar
Hendriks, D.F.G., Puigvert, L.F., Messner, S. & Mortiz, W. Трехмерные сфероидные модели печени для обнаружения и изучения соединений с холестатической активностью. Нац. Опубл. гр . 1–12 (2016).
Нац. Опубл. гр . 1–12 (2016).
Gecommentarierd Geneesmiddelenrepertorium. на, http://www.bcfi.be/nl/download (2018).
ДиПиро, Дж. Т. и др. . Фармакотерапия: патофизиологический подход . (McGraw-Hill Medical, 2014).
Николл, Л. Х. и Хесби, А. Внутримышечные инъекции: комплексный обзор исследований и руководство по доказательной практике. Заяв. Нурс. Рез. 15 , 149–62 (2002).
Артикул пабмед Google Scholar
Гелдерс Ю.G. Фармакология, фармакокинетика и клиническая разработка галоперидола деканоата. Междунар. клин. Психофармак. 1 , 1–11 (1986).
Артикул пабмед Google Scholar
Наяк, Р. К., Дуз, Д. Р. и Наир, Н. П. Биодоступность и фармакокинетика внутримышечного перорального и депо-галоперидола у пациентов с шизофренией. Дж. Клин. Фармакол. 27 , 144–150 (1987).
Дж. Клин. Фармакол. 27 , 144–150 (1987).
Артикул пабмед КАС Google Scholar
Саймон, А., де Алмейда Борхес, В. Р., Кабрал, Л. М. и де Соуза, В. П. Разработка и валидация дифференциального теста на растворение бетаметазона натрия фосфата и бетаметазона дипропионата внутримышечной инъекционной суспензии. AAPS Pharm Sci Tech 14 , 425–34 (2013).
Артикул КАС Google Scholar
Грей, В. и др. . Наука растворения USP 1 и 2: нынешние проблемы и актуальность в будущем. Фарм. Рез. 26 , 1289–302 (2009).
Артикул пабмед КАС Google Scholar
Seidlitz, A. & Weitschies, W. Методы растворения in-vitro для парентеральных препаратов с контролируемым высвобождением и их применимость для тестирования стентов с лекарственным покрытием. Дж. Фарм. Фармакол. 64 , 969–85 (2012).
Дж. Фарм. Фармакол. 64 , 969–85 (2012).
Артикул пабмед КАС Google Scholar
Алам, А., Ахуджа, А., Бабута, С., Гидвани, С. К. и Али, Дж. Состав и оценка фармацевтически эквивалентной парентеральной депо-суспензии ацетата метилпреднизолона. Индиан Дж. Фарм. науч. 71 , 30–4 (2009).
Артикул пабмед КАС Google Scholar
Ванденбург, Х. и др. . Тканеинженерные органоиды скелетных мышц для обратимой генной терапии. Гул. Джин Тер. 7 , 2195–2200 (1996).
Артикул пабмед КАС Google Scholar
Голобова Д. и др. . Формирование эндотелиальной сети в скелетных мышцах человека, созданных с помощью тканевой инженерии. Ткань Eng. Часть A 21 , 2548–58 (2015).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Пауэлл, К.А., Смайли, Б.Л., Миллс, Дж. и Ванденбург, Х.Х. Механическая стимуляция улучшает тканеинженерные скелетные мышцы человека. утра. Дж. Физиол. — Селл Физиол . 283 (2002).
Vandenburgh, H., Shansky, J. & Del Tatto, M.C. Органогенез скелетных мышц в культуре тканей. Методы Mol Med. 18 , 217–225 (1999).
ПабМед КАС Google Scholar
Wang, L., Shansky, J. & Vandenburgh, H. Индуцированное образование и созревание кластеров ацетилхолиновых рецепторов в определенной трехмерной биоискусственной мышце. Мол. Нейробиол. 48 , 397–403 (2013).
Артикул пабмед КАС Google Scholar
Ванденбург, Х. и др. . Автоматизированный скрининг лекарств с помощью сократительной мышечной ткани, созданной из дистрофических миобластов. FASEB J. 23 , 3325–34 (2009).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Ванденбург, Х. и др. . Платформа для скрининга лекарств, основанная на сократительной способности тканеинженерных мышц. Muscle Nerve 37 , 438–47 (2008).
Артикул пабмед КАС Google Scholar
Thorrez, L. и др. . Ангиогенез усиливает доставку и устойчивость фактора IX из биоинженерных мышечных имплантатов человека. Мол. тер. 14 , 442–51 (2006).
Артикул пабмед КАС Google Scholar
Шанский Дж. и др. . Тканевая инженерия скелетных мышц человека для клинического применения . Культура клеток для тканевой инженерии (2006).
Декруа, Л., Ван Мейлдер, В., Десендер, Л., Сампаолеси, М. и Торрез, Л. Очистка тканей для конфокальной визуализации нативных и биоискусственных скелетных мышц. Биотех. гистохим. 90 , 424–431 (2015).
Биотех. гистохим. 90 , 424–431 (2015).
Артикул пабмед КАС Google Scholar
Роша, Т., де Соуза, Б.М., Пальма, М.С. и да Круз-Хёфлинг, М.А. Миотоксические эффекты мастопарана из яда осы Polybia paulista (Hymenoptera, Epiponini) в скелетных мышцах мышей. Toxicon 50 , 589–99 (2007).
Артикул пабмед КАС Google Scholar
Rasband, W. S., ImageJ, Национальный институт здравоохранения США, Бетесда, Мэриленд, США, https://imagej.nih.gov/ij/, 1997–2016.
Pfaffl, M.W., Horgan, G.W. & Dempfle, L. Программный инструмент относительной экспрессии (REST) для группового сравнения и статистического анализа результатов относительной экспрессии в ПЦР в реальном времени. Рез. нуклеиновых кислот. 30 , 36 (2002).
Артикул Google Scholar

Брейди, М. А., Льюис, М. П. и Мудера, В. Синергия между миогенными и немиогенными клетками в трехмерной тканеинженерной конструкции черепно-лицевой скелетной мышцы. J. Tissue Eng. Реген. Мед. 2 , 408–417 (2008).
Артикул пабмед КАС Google Scholar
Карозио, С. и др. . Поколение eX vivo -васкуляризированной мышечно-инженерной ткани (X-MET). науч. Респ. 3 , 1420 (2013).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Abet, V., Filace, F., Recio, J., Alvarez-Builla, J. & Burgos, C. Пролекарственный подход: обзор недавних случаев. евро. Дж. Мед. хим. 127 , 810–827 (2017).
Артикул пабмед КАС Google Scholar
Замек-Глишинский, М.J. и др. . Дихлорфлуоресцеин и его диацетатная составляющая в печени. Фармакология 304 , 801–809 (2003).
Фармакология 304 , 801–809 (2003).
КАС Google Scholar
Дайв, К., Кокс, Х., Уотсон, Дж. В. и Уоркман, П. Полярные производные флуоресцеина в качестве улучшенных субстратных зондов для проточного цитоэнзимологического анализа клеточных эстераз. Мол. Клетка. Зонды 2 , 131–45 (1988).
Артикул пабмед КАС Google Scholar
Мерфи, М. М., Лоусон, Дж. А., Мэтью, С. Дж., Хатчесон, Д. А. и Кардон, Г. Сателлитные клетки, фибробласты соединительной ткани и их взаимодействие имеют решающее значение для регенерации мышц. Развитие 138 , 3625–3637 (2011).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Деннис, Р. Г., Косник, П. Е., Гилберт, М. Э. и Фолкнер, Дж. А. Возбудимость и сократимость скелетных мышц, полученных из первичных культур и клеточных линий. утра. Дж. Физиол. Клеточная физиол. 280 , C288–95 (2001).
утра. Дж. Физиол. Клеточная физиол. 280 , C288–95 (2001).
Артикул пабмед КАС Google Scholar
Фронтера В. Р. и Очала Дж. Скелетные мышцы: краткий обзор структуры и функции. Кальцин. Ткань внутр. 96 , 183–95 (2015).
Артикул пабмед КАС Google Scholar
Роман В.и Гомес, Э. Р. Позиционирование ядер в скелетных мышцах. Семин. Сотовый Дев. Биол . [Epub перед печатью] (2017).
Чемберлен, Дж. С., Джейнс, Дж. Б. и Хаушка, С. Д. Регуляция индукции креатинкиназы в дифференцирующихся миобластах мыши. Мол. Клетка. биол. 5 , 484–92 (1985).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Зуурвельд, Дж.Г., Виртц П., Лоерманс Х.М. и Виркамп Дж.Х. Постнатальный рост и дифференцировка трех мышц задних конечностей крысы. Характеристика биохимическими и ферментно-гистохимическими методами. Рез. клеточной ткани. 241 , 183–92 (1985).
Характеристика биохимическими и ферментно-гистохимическими методами. Рез. клеточной ткани. 241 , 183–92 (1985).
Артикул пабмед КАС Google Scholar
Дарвилл, Н. и др. . Влияние ингибирования макрофагов и ангиогенеза на высвобождение и абсорбцию лекарственного средства из внутримышечной суспензии палиперидона пальмитата с замедленным высвобождением. Дж. Контроль. Выпуск 230 , 95–108 (2016).
Артикул пабмед КАС Google Scholar
Breeuwer, P. и др. . Характеристика поглощения и гидролиза диацетата флуоресцеина и диацетата карбоксифлуоресцеина внутриклеточными эстеразами в Saccharomyces cerevisiae, что приводит к накоплению флуоресцентного продукта. Заяв. Окружающая среда. микробиол. 61 , 1614–9 (1995).
ПабМед ПабМед Центральный КАС Google Scholar

Sartorelli, A.C. и др. . Митомицин С: прототип биоредуктивного агента. Онкол. Рез. 6 , 501–8 (1994).
ПабМед КАС Google Scholar
Сигел, Д. и др. . Биоредуктивная активация митомицина С DT-диафоразой. Биохимия 31 , 7879–7885 (1992).
Артикул пабмед КАС Google Scholar
Neve, E.P.A. и др. . Экспрессия и функция mARC: роль в липогенезе и метаболической активации ксимелагатрана. PLoS One 10 (2015 г.).
Chen, Y., Jin, JY, Mukadam, S., Malhi, V. & Kenny, JR Применение IVIVE и моделирования PBPK в проспективном прогнозировании клинической фармакокинетики: стратегия и подход на этапе разработки лекарств с четыре тематических исследования. Биофарм. Утилизация наркотиков 33 , 85–98 (2012).
Артикул пабмед КАС Google Scholar
Yoon, M., Campbell, J.L., Andersen, J., Melvin, M.E. & Clewell, H. Количественный анализ in vitro на in vivo экстраполяция результатов анализа клеточной токсичности. Крит. Преподобный Токсикол. 42 , 633–652 (2012).
Артикул пабмед КАС Google Scholar
Пачеко, Д. П., Рейс, Р. Л., Коррело, В. М. и Маркес, А. П. Перекрестные помехи между тканевой инженерией и фармацевтической биотехнологией: последние достижения и будущие направления. Курс. фарм. Биотехнолог. 16 , 1012–23 (2015).
Артикул пабмед КАС Google Scholar
Черино, Г. и др. . Трехмерная многоклеточная мышечная тканевая инженерия в перфузионных биореакторах. Биотехнология. Биоэнг . 113 (2015).
Левенберг С. и др. . Инженерия васкуляризированной скелетной мышечной ткани. Нац. Биотехнолог. 23 , 879–84 (2005).
Нац. Биотехнолог. 23 , 879–84 (2005).
Артикул пабмед КАС Google Scholar
Coupin, G., Warter, J. & Poindron, P. Типы клеток, необходимые для эффективной иннервации мышечных клеток человека in Vitro Se. 212 , 204–212 (2000).
Cohen, L. S. et al . Уровень лидокаина в плазме после внутримышечного введения. утра. Дж. Кардиол. 29 , 520–523 (1972).
Артикул пабмед КАС Google Scholar
Эванс, Э. Ф., Проктор, Дж. Д., Фраткин, М. Дж., Веландия, Дж. и Вассерман, А. Дж. Кровоток в группах мышц и абсорбция лекарств. клин.Фармакол. тер. 17 , 44–47 (1975).
Артикул пабмед КАС Google Scholar
Юм К., Хонг С. Г., Хили К. Э. и Ли Л. П. Физиологически значимые органы на чипсах. Биотехнология. J. 9 , 16–27 (2014).
Биотехнология. J. 9 , 16–27 (2014).
Артикул пабмед КАС Google Scholar
Ито, А. и др. .Индукция функциональных тканеинженерных конструкций скелетных мышц с помощью определенной электрической стимуляции. науч. Респ. 4 , 4781 (2014).
Артикул пабмед ПабМед Центральный КАС Google Scholar
Rangarajan, S., Madden, L. & Bursac, N. Использование потока, электрической и механической стимуляции для развития поперечнополосатых мышц. Энн. Биомед. англ. 42 , 1391–405 (2014).
Артикул пабмед Google Scholar
Wickramasinghe, S.N. Доказательства метаболизма лекарств макрофагами: возможная роль макрофагов в патогенезе лекарственного повреждения тканей и в активации проканцерогенов окружающей среды. клин. лаборатория Гематол. 9 , 271–80 (1987).
лаборатория Гематол. 9 , 271–80 (1987).
Артикул пабмед КАС Google Scholar
Каплин, Дж. Д., Гранадос, Н. Г., Джеймс, М. Р., Монтазами, Р. и Хашеми, Н. Микрожидкостная технология «орган-на-чипе» для развития разработки лекарств и токсикологии. Доп. Здоровьеc. Матер. 4 , 1426–50 (2015).
Артикул пабмед КАС Google Scholar
Зона обучения: живое животное
Это простой факт, большинство животных двигаются. Люди (как и вы) могут двигаться, потому что ваше тело поддерживается внутренним скелетом — верно, вы просто мешок с костями!
Посмотрите на кости, из которых состоит скелет человека
Все позвоночные животные (рыбы, земноводные, рептилии, птицы и млекопитающие) имеют внутренний скелет.Хотя на первый взгляд они выглядят совершенно по-разному, у них есть некоторые общие характеристики.
Посмотрите на скелеты ниже и сравните их со скелетом человека. Можете ли вы узнать череп, кости ног и позвоночник у пеликана, лягушки и тигра?
Нажмите на картинку, чтобы узнать больше!
Посмотрите еще несколько скелетов животных, выставленных в музее
Итак, что делает твой скелет? Кости в вашем скелете действуют как якоря для всех ваших мышц.Мышцы работают, чтобы тянуть кости в разные стороны. Возьмитесь за мышцы руки. Как и большинство мышц, они работают парами: одна расширяется (увеличивается), а другая сокращается (уменьшается), что позволяет вам двигать рукой вокруг локтевого сустава. Ваш скелет также обеспечивает вам поддержку и защиту. Представьте себе ущерб, который вы могли бы нанести своему мозгу, если бы у вас не было черепа для его защиты!
Наведите курсор на картинки, чтобы увидеть скелет в действии.
Не все животные ходят и бегают, как люди — их скелеты приспособились к различным формам движения. Рыбы плавают, их длинные гибкие спины и крепкие плавники позволяют им легко скользить по воде. Лягушки прыгают, их сильные задние ноги и большие ступни помогают им прыгать. Птицы летают, их «руки» превратились в крылья.
Рыбы плавают, их длинные гибкие спины и крепкие плавники позволяют им легко скользить по воде. Лягушки прыгают, их сильные задние ноги и большие ступни помогают им прыгать. Птицы летают, их «руки» превратились в крылья.
Некоторые животные, например насекомые и крабы, имеют совершенно отличный от нашего тип скелета — их скелеты наружные (на внешней стороне тела). Их называют беспозвоночными, потому что у них нет позвоночника, состоящего из позвонков. Некоторые другие беспозвоночные, например медузы, вообще не имеют скелета!
Вы можете думать о других животных? Как они двигаются?
Как вы думаете, как изменились их скелеты, чтобы сделать возможным это движение?
Нажмите на картинку, чтобы узнать больше!
Узнайте больше о … М р С Н Е р G
Вернитесь к миссис Нерг на главную страницу или проверьте свои знания с помощью ее викторины!
7.1 Отделы скелетной системы – анатомия и физиология
Цели обучения
К концу этого раздела вы сможете:
- Обсудить функции костной системы
- Различают осевой скелет и добавочный скелет
- Определение осевого скелета и его компонентов
- Определение аппендикулярного скелета и его компонентов
Скелетная система включает все кости, хрящи и связки тела, которые поддерживают и придают форму телу и его структурам.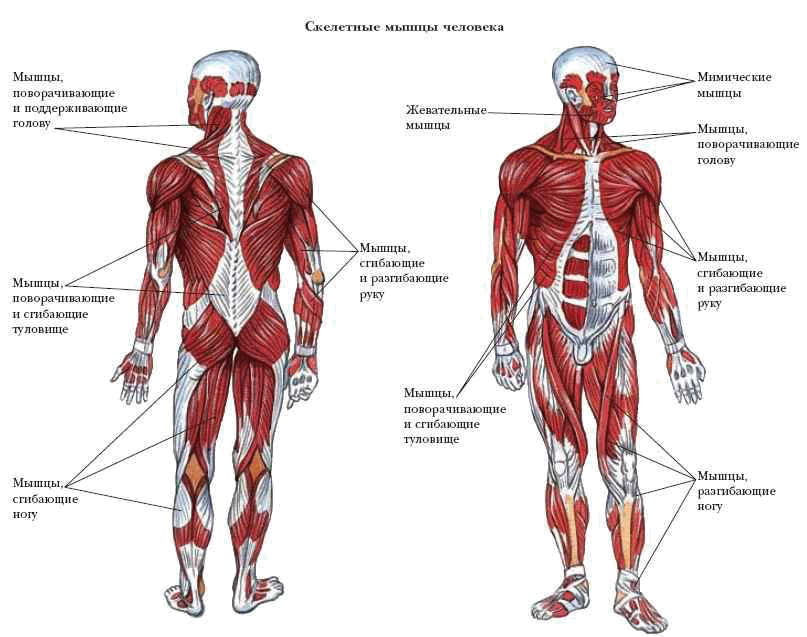 Скелет состоит из костей тела. У взрослых в скелете 206 костей. У более молодых людей количество костей больше, потому что некоторые кости сливаются вместе в детстве и подростковом возрасте, образуя кость взрослого человека. Основные функции скелета состоят в том, чтобы обеспечить жесткую внутреннюю структуру, которая может выдерживать вес тела против силы гравитации, и обеспечить структуру, на которую могут воздействовать мышцы, чтобы производить движения тела. Нижняя часть скелета предназначена для обеспечения устойчивости при ходьбе или беге.Напротив, верхний скелет имеет большую подвижность и диапазон движений, функции, которые позволяют вам поднимать и переносить предметы или поворачивать голову и туловище.
Скелет состоит из костей тела. У взрослых в скелете 206 костей. У более молодых людей количество костей больше, потому что некоторые кости сливаются вместе в детстве и подростковом возрасте, образуя кость взрослого человека. Основные функции скелета состоят в том, чтобы обеспечить жесткую внутреннюю структуру, которая может выдерживать вес тела против силы гравитации, и обеспечить структуру, на которую могут воздействовать мышцы, чтобы производить движения тела. Нижняя часть скелета предназначена для обеспечения устойчивости при ходьбе или беге.Напротив, верхний скелет имеет большую подвижность и диапазон движений, функции, которые позволяют вам поднимать и переносить предметы или поворачивать голову и туловище.
Помимо обеспечения опоры и движений тела, скелет выполняет защитную и накопительную функции. Он защищает внутренние органы, в том числе головной и спинной мозг, сердце, легкие и органы малого таза. Кости скелета служат основным местом хранения важных минералов, таких как кальций и фосфат.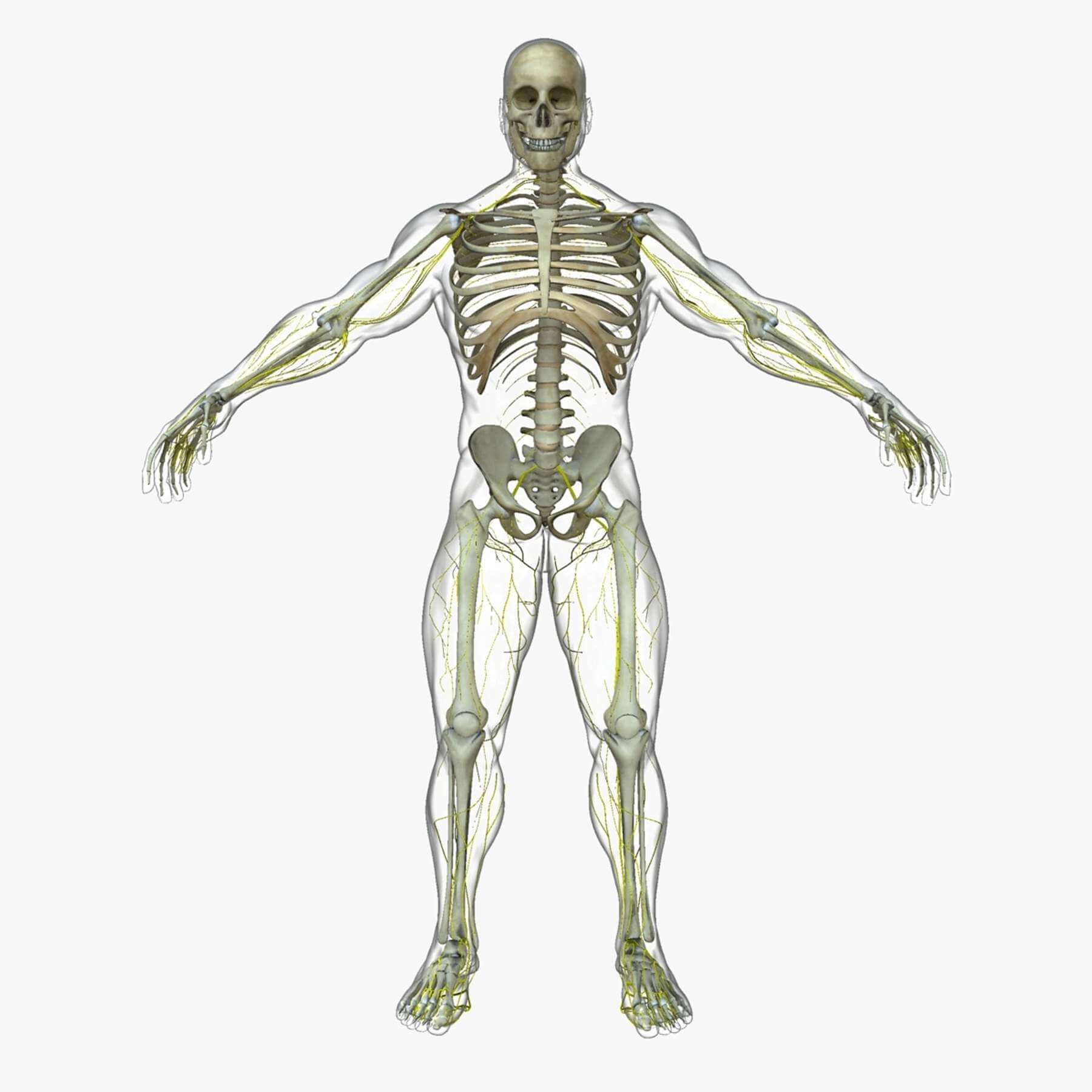 Костный мозг, находящийся в костях, хранит жир и содержит ткань тела, производящую клетки крови.
Костный мозг, находящийся в костях, хранит жир и содержит ткань тела, производящую клетки крови.
Скелет подразделяется на два основных отдела — осевой и аппендикулярный.
Осевой скелет
Скелет подразделяется на два основных отдела — осевой и аппендикулярный. Осевой скелет образует вертикальную центральную ось тела и включает в себя все кости головы, шеи, груди и спины (рис. 7.2). Он служит для защиты головного и спинного мозга, сердца и легких. Он также служит местом прикрепления мышц, которые двигают головой, шеей и спиной, и мышц, которые действуют через плечевые и тазобедренные суставы, чтобы двигать соответствующие конечности.
Осевой скелет взрослого человека состоит из 80 костей, включая череп, позвоночник и грудную клетку. Череп образован 22 костями. С головой также связаны еще семь костей, в том числе подъязычная кость и слуховые косточки (три маленькие кости в каждом среднем ухе). Позвоночный столб состоит из 24 костей, каждая из которых называется позвонком, а также крестца и копчика. Грудная клетка включает 12 пар ребер и грудину, уплощенную кость передней части грудной клетки.
Грудная клетка включает 12 пар ребер и грудину, уплощенную кость передней части грудной клетки.
Рис. 7.2 Осевой и аппендикулярный скелет Осевой скелет поддерживает голову, шею, спину и грудь и, таким образом, образует вертикальную ось тела. Он состоит из черепа, позвоночного столба (включая крестец и копчик) и грудной клетки, образованной ребрами и грудиной. Аппендикулярный скелет образован всеми костями верхних и нижних конечностей.
Аппендикулярный скелет
Аппендикулярный скелет включает все кости верхних и нижних конечностей, а также кости, прикрепляющие каждую конечность к осевому скелету.В аппендикулярном скелете взрослого человека 126 костей. Кости аппендикулярного скелета рассматриваются в отдельной главе.
Границы | Внеклеточный матрикс скелетных мышц – что мы знаем о его составе, регуляции и физиологических ролях? Повествовательный обзор
Введение
Скелетные мышцы являются важным компонентом строения тела человека, обычно на их долю приходится более 40% и 30% общей массы тела у мужчин и женщин соответственно (Kim et al. , 2002). Наиболее очевидной функцией скелетных мышц является создание сил, необходимых для поддержания вертикального положения тела и выполнения движения. Однако скелетные мышцы также играют важную роль во многих других физиологических процессах, включая термогенез (Rowland et al., 2015), метаболизм (Baskin et al., 2015) и секрецию многочисленных пептидов для связи с другими тканями (Pedersen and Febbraio). , 2012). Таким образом, укрепление и поддержание здоровья скелетных мышц имеет жизненно важное значение.Хотя в последние годы фармакологические миметики упражнений вызывают все больший научный интерес (Fan and Evans, 2017), именно физические упражнения считаются наиболее мощным и универсально применимым инструментом для этих целей.
, 2002). Наиболее очевидной функцией скелетных мышц является создание сил, необходимых для поддержания вертикального положения тела и выполнения движения. Однако скелетные мышцы также играют важную роль во многих других физиологических процессах, включая термогенез (Rowland et al., 2015), метаболизм (Baskin et al., 2015) и секрецию многочисленных пептидов для связи с другими тканями (Pedersen and Febbraio). , 2012). Таким образом, укрепление и поддержание здоровья скелетных мышц имеет жизненно важное значение.Хотя в последние годы фармакологические миметики упражнений вызывают все больший научный интерес (Fan and Evans, 2017), именно физические упражнения считаются наиболее мощным и универсально применимым инструментом для этих целей.
За последние десятилетия были проведены тысячи тренировочных исследований в попытке определить режимы упражнений, наиболее подходящие для увеличения размера мышц и улучшения их функциональных характеристик в различных когортах (например, на момент написания этой статьи Pubmed дал больше более 24 000 результатов для поисковых операторов «упражнения» и «мышечная сила»). Результаты этих исследований легли в основу различных руководств по назначению упражнений, наиболее известными из которых, вероятно, являются стенды по положению, опубликованные и периодически обновляемые Американским колледжем спортивной медицины (2009 г.), Garber et al. (2011). Большинство исследований основывают свою оценку эффективности тренировочных вмешательств на исследовании сократительных мышечных клеток. Часто изучаемые параметры включают размер мышц, измеренный на органном (Fisher et al., 2011) или клеточном уровне (Schoenfeld, 2010), распределение типов волокон (Adams et al., 1993), архитектуру (Aagaard et al., 2001), а также нервную стимуляцию мышц (Folland and Williams, 2007).
Результаты этих исследований легли в основу различных руководств по назначению упражнений, наиболее известными из которых, вероятно, являются стенды по положению, опубликованные и периодически обновляемые Американским колледжем спортивной медицины (2009 г.), Garber et al. (2011). Большинство исследований основывают свою оценку эффективности тренировочных вмешательств на исследовании сократительных мышечных клеток. Часто изучаемые параметры включают размер мышц, измеренный на органном (Fisher et al., 2011) или клеточном уровне (Schoenfeld, 2010), распределение типов волокон (Adams et al., 1993), архитектуру (Aagaard et al., 2001), а также нервную стимуляцию мышц (Folland and Williams, 2007).
Несмотря на обилие информации о податливости скелетных мышц, часто упускается из виду тот факт, что мышечные волокна встроены во внеклеточный матрикс (ВКМ), состоящий из сетки коллагеновых компонентов, а также смеси дополнительных макромолекул, таких как различные гликопротеины и протеогликаны. Недавние исследования показали, что внеклеточный матрикс играет важную роль в развитии (Thorsteinsdóttir et al., 2011), рост (Fry et al., 2017) и восстановление мышц (Calve et al., 2010), а также передачу силы сокращения (Street, 1983). Несмотря на то, что существуют доказательства гибкости ВКМ, лишь немногие исследования сообщают о его реакции на различные формы тренировок, что свидетельствует о том, что физиологическая роль ВКМ еще не полностью оценена специалистами по физическим упражнениям. Стремясь стимулировать дальнейшие исследования тренировочных реакций несократительных компонентов скелетных мышц, мы представляем обзор современного состояния знаний о составе, структуре и регуляции ВКМ, его физиологических ролях, нарушениях регуляции, связанных со старением и метаболическими нарушениями. нарушений, а также адаптации к физическим нагрузкам.
Недавние исследования показали, что внеклеточный матрикс играет важную роль в развитии (Thorsteinsdóttir et al., 2011), рост (Fry et al., 2017) и восстановление мышц (Calve et al., 2010), а также передачу силы сокращения (Street, 1983). Несмотря на то, что существуют доказательства гибкости ВКМ, лишь немногие исследования сообщают о его реакции на различные формы тренировок, что свидетельствует о том, что физиологическая роль ВКМ еще не полностью оценена специалистами по физическим упражнениям. Стремясь стимулировать дальнейшие исследования тренировочных реакций несократительных компонентов скелетных мышц, мы представляем обзор современного состояния знаний о составе, структуре и регуляции ВКМ, его физиологических ролях, нарушениях регуляции, связанных со старением и метаболическими нарушениями. нарушений, а также адаптации к физическим нагрузкам.
Состав и структура скелетных мышц ECM
ВКМ скелетных мышц представляет собой сложную сеть, состоящую из коллагенов, гликопротеинов, протеогликанов и эластина (Takala and Virtanen, 2000; Halper and Kjaer, 2014). Коллагены образуют сеть внутримышечной соединительной ткани (ВМСТ), т. е. центральные, волокнистые компоненты ВКМ. IMCT обычно изображается организованным в три слоя: (i) эндомизий, представляющий собой самый внутренний слой, который заключает в себе отдельные мышечные волокна, (ii) перимизий, объединяющий группы мышечных волокон, и (iii) эпимизий, покрывающий всю мышцу.Большая структурная сложность сети IMCT, подтвержденная сканирующими электронными микрофотографиями, предполагает, что эта традиционная классификация может быть упрощенной и что еще необходимо определить организацию ВКМ более высокого порядка (Gillies and Lieber, 2011). Исследования фасциальных тканей далее рассматривают слои IMCT как часть сложной системы взаимосвязанных и переплетающихся соединительных тканей, которая «окружает, переплетается между собой и пронизывает все органы, мышцы, кости и нервные волокна, придавая телу функциональную структуру, и создание среды, позволяющей всем системам организма работать комплексно» (Adstrum et al.
Коллагены образуют сеть внутримышечной соединительной ткани (ВМСТ), т. е. центральные, волокнистые компоненты ВКМ. IMCT обычно изображается организованным в три слоя: (i) эндомизий, представляющий собой самый внутренний слой, который заключает в себе отдельные мышечные волокна, (ii) перимизий, объединяющий группы мышечных волокон, и (iii) эпимизий, покрывающий всю мышцу.Большая структурная сложность сети IMCT, подтвержденная сканирующими электронными микрофотографиями, предполагает, что эта традиционная классификация может быть упрощенной и что еще необходимо определить организацию ВКМ более высокого порядка (Gillies and Lieber, 2011). Исследования фасциальных тканей далее рассматривают слои IMCT как часть сложной системы взаимосвязанных и переплетающихся соединительных тканей, которая «окружает, переплетается между собой и пронизывает все органы, мышцы, кости и нервные волокна, придавая телу функциональную структуру, и создание среды, позволяющей всем системам организма работать комплексно» (Adstrum et al.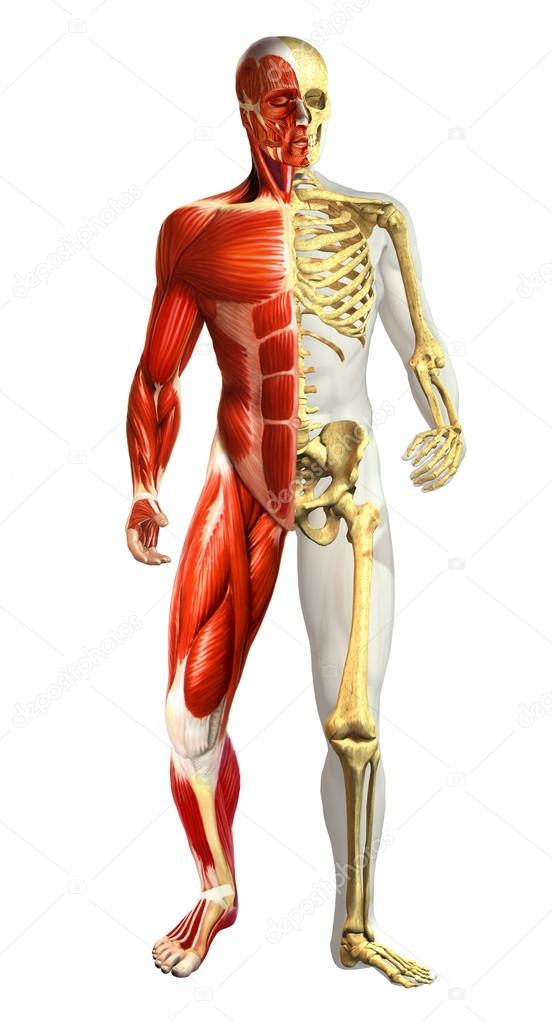 , 2017; Стекко и др., 2018). Эта система, которую обычно называют фасциальной системой, все чаще признается важной мишенью в спортивной медицине (Zügel et al., 2018).
, 2017; Стекко и др., 2018). Эта система, которую обычно называют фасциальной системой, все чаще признается важной мишенью в спортивной медицине (Zügel et al., 2018).
IMCT содержит различные формы коллагенов, наиболее распространены типы I и III (Duance et al., 1977; Light and Champion, 1984; Gillies and Lieber, 2011; McKee et al., 2019). Эндомизий взаимодействует с сарколеммой миофибрилл на специализированной базальной мембране, которая состоит в основном из коллагена IV типа и ламинина (Sanes, 1982; Martin and Timpl, 1987; Kjaer, 2004).Было обнаружено, что концентрация этих двух компонентов различается в зависимости от типа мышечного волокна, при этом медленно сокращающиеся волокна характеризуются значительно более высокими концентрациями коллагена IV, но более низкими концентрациями ламинина (Kovanen et al., 1988). Ламинин, в свою очередь, служит лигандом для двух сарколеммальных рецепторов – гликопротеинового комплекса, ассоциированного с дистрофином, и интегрина α7β1 (Grounds et al. , 2005) – расположенных в костамерах, которые представляют собой мембраносвязанные белковые структуры, выровненные в регистре с Z- диски миофибрилл.Считается, что интегрины действуют двунаправленно, позволяя внутриклеточным сигнальным молекулам регулировать внешнюю адгезию (передача сигналов «изнутри») и передавая внешние стимулы для воздействия на клеточные процессы («передача сигналов снаружи-внутрь») (Boppart and Mahmassani, 2019). . Таким образом, чистое напряжение цитоскелета индуцирует внутриклеточное связывание белков, таких как талин, винкулин или киндлин, что приводит к изменению конформации интегринового рецептора и позволяет внеклеточным доменам рецептора расширяться к белкам внутри внеклеточного матрикса.Кроме того, интегриновые лиганды из внеклеточного пространства, такие как ламинин, коллаген или фибронектин, облегчают формирование высокоаффинного вертикального состояния, что приводит к усилению связывания с белками ВКМ и кластеризации интегринов, особенно вдоль комплексов фокальной адгезии (Boppart and Mahmassani, 2019).
, 2005) – расположенных в костамерах, которые представляют собой мембраносвязанные белковые структуры, выровненные в регистре с Z- диски миофибрилл.Считается, что интегрины действуют двунаправленно, позволяя внутриклеточным сигнальным молекулам регулировать внешнюю адгезию (передача сигналов «изнутри») и передавая внешние стимулы для воздействия на клеточные процессы («передача сигналов снаружи-внутрь») (Boppart and Mahmassani, 2019). . Таким образом, чистое напряжение цитоскелета индуцирует внутриклеточное связывание белков, таких как талин, винкулин или киндлин, что приводит к изменению конформации интегринового рецептора и позволяет внеклеточным доменам рецептора расширяться к белкам внутри внеклеточного матрикса.Кроме того, интегриновые лиганды из внеклеточного пространства, такие как ламинин, коллаген или фибронектин, облегчают формирование высокоаффинного вертикального состояния, что приводит к усилению связывания с белками ВКМ и кластеризации интегринов, особенно вдоль комплексов фокальной адгезии (Boppart and Mahmassani, 2019). . Связанный с дистрофином гликопротеиновый комплекс является еще одним важным фактором в обеспечении механической связи между сократительными компонентами скелетных мышц (то есть актином) и взаимосвязанными слоями IMCT (Ervasti, 1993; Peter et al., 2011). Основные компоненты, связывающие сократительные элементы мышцы с интерстициальным матриксом, показаны на рисунке 1.
. Связанный с дистрофином гликопротеиновый комплекс является еще одним важным фактором в обеспечении механической связи между сократительными компонентами скелетных мышц (то есть актином) и взаимосвязанными слоями IMCT (Ervasti, 1993; Peter et al., 2011). Основные компоненты, связывающие сократительные элементы мышцы с интерстициальным матриксом, показаны на рисунке 1.
Рисунок 1. Основные компоненты внеклеточного матрикса скелетных мышц и его связь с сократительными компонентами мышц.
Суперсемейство коллагена содержит в общей сложности 28 различных членов, из которых было показано, что типы I, III, IV, V, VI, XII, XIII, XIV, XV, XVIII и XXII присутствуют в зрелых скелетных мышцах в гене и/или уровень белка (таблица 1).Фибриллообразующие типы I и III, безусловно, наиболее распространены, при этом протеомные исследования показывают, что на их совокупную долю приходится примерно 75% общего мышечного коллагена (McKee et al., 2019). Предполагается, что сильные параллельные волокна коллагена I типа, которые присутствуют в эндо-, пери- и эпимизии, придают мышце прочность на растяжение и жесткость, тогда как коллаген типа III образует рыхлую сеть волокон, которая придает эластичность мышцам. эндо- и перимизий (Кованен, 2002). Коллаген типа IV, спиральная молекула, образует сетчатую структуру, которая составляет основу базальной мембраны (Sanes, 2003).Коллаген типа VI был обнаружен в эпимизиальном, перимизиальном и эндомизиальном интерстиции, но особенно вблизи базальной мембраны, где он взаимодействует с карбоксиконцевым глобулярным доменом коллагена типа IV (Kuo et al., 1997). Интересно, что коллаген VI обладает нетипичными неколлагеновыми областями, формирующими отчетливую микрофибриллярную сеть в большинстве соединительных тканей (Maaß et al., 2016). Мутации коллагена VI приводят к нарушениям с комбинированным поражением мышц и соединительной ткани, включая врожденную мышечную дистрофию Ульриха, миопатию Бетлема, аутосомно-доминантную поясно-конечностную мышечную дистрофию и аутосомно-рецессивный миосклероз (Bushby et al., 2014).
эндо- и перимизий (Кованен, 2002). Коллаген типа IV, спиральная молекула, образует сетчатую структуру, которая составляет основу базальной мембраны (Sanes, 2003).Коллаген типа VI был обнаружен в эпимизиальном, перимизиальном и эндомизиальном интерстиции, но особенно вблизи базальной мембраны, где он взаимодействует с карбоксиконцевым глобулярным доменом коллагена типа IV (Kuo et al., 1997). Интересно, что коллаген VI обладает нетипичными неколлагеновыми областями, формирующими отчетливую микрофибриллярную сеть в большинстве соединительных тканей (Maaß et al., 2016). Мутации коллагена VI приводят к нарушениям с комбинированным поражением мышц и соединительной ткани, включая врожденную мышечную дистрофию Ульриха, миопатию Бетлема, аутосомно-доминантную поясно-конечностную мышечную дистрофию и аутосомно-рецессивный миосклероз (Bushby et al., 2014).
Таблица 1. Обзор коллагеновых компонентов внеклеточного матрикса скелетных мышц.
Коллаген типов XII, XIV, XIX и XXII относится к коллагенам, ассоциированным с фибриллами, с прерванными тройными спиралями (FACIT; Chiquet et al. , 2014; Calvo et al., 2020), при этом коллаген типа XXII, по-видимому, экспрессируется исключительно в тканях. соединения, такие как мышечно-сухожильное соединение в скелетной и сердечной мышцах (Koch et al., 2004).
, 2014; Calvo et al., 2020), при этом коллаген типа XXII, по-видимому, экспрессируется исключительно в тканях. соединения, такие как мышечно-сухожильное соединение в скелетной и сердечной мышцах (Koch et al., 2004).
Биоинформационные инструменты для скрининга протеома нормальных и пораженных тканей человека позволили охарактеризовать глобальный состав протеома ВКМ, или «матрисомы».Всего с ВКМ было связано 1027 генов, благодаря чему основные матрисомные белки (гликопротеины ВКМ, коллагены и протеогликаны) можно было отличить от белков, ассоциированных с матрисомой (белки, ассоциированные с ВКМ, регуляторы ВКМ и секретируемые факторы, которые могут взаимодействовать с матрисомами). основные белки ECM) (Naba et al., 2016). Учитывая сложность скелетной мышечной ткани человека, включающей многоядерные мышечные волокна, иммунные клетки, эндотелиальные клетки, мышечные стволовые клетки, немиогенные мезенхимальные клетки-предшественники и другие мононуклеарные клетки (Bentzinger et al. , 2013a), потребуются дальнейшие исследования для выяснения вклада каждой из этих клеток в структуру и ремоделирование IMCT. Сигнатуры генов, полученные, например, из секвенирования РНК изолированных мышечных волокон и других типов клеток, представляют собой многообещающий инструмент для деконволюции объемной ткани скелетных мышц (Rubenstein et al., 2020).
, 2013a), потребуются дальнейшие исследования для выяснения вклада каждой из этих клеток в структуру и ремоделирование IMCT. Сигнатуры генов, полученные, например, из секвенирования РНК изолированных мышечных волокон и других типов клеток, представляют собой многообещающий инструмент для деконволюции объемной ткани скелетных мышц (Rubenstein et al., 2020).
Физиологическая регуляция генов ECM
Гомеостаз внеклеточного матрикса поддерживается за счет точно настроенных анаболических и катаболических процессов, которые регулируются различными факторами роста, протеогликанами и ферментами, ответственными за деградацию коллагена.Было обнаружено, что после связывания с мембраносвязанными рецепторами факторы роста, принадлежащие к надсемейству трансформирующих факторов роста бета (TGF-β), индуцируют фосфорилирование белков Smad, которые передают внеклеточные сигналы в ядро, где они активируют транскрипцию нижестоящих генов, что приводит к продукции коллагена. (Макдональд и Кон, 2012 г. ). Другим, хотя и менее описанным, фактором аналогичной функции является фактор роста соединительной ткани (CTGF), избыточная экспрессия которого, как сообщалось, провоцирует дистрофический мышечный фиброз и функциональные нарушения (Morales et al., 2011).
). Другим, хотя и менее описанным, фактором аналогичной функции является фактор роста соединительной ткани (CTGF), избыточная экспрессия которого, как сообщалось, провоцирует дистрофический мышечный фиброз и функциональные нарушения (Morales et al., 2011).
Функция этих анаболических факторов в основном регулируется небольшими богатыми лейцином протеогликанами (SLRP). Декорин, прототип этого семейства, деактивирует профибротические TGF-β и CTGF (Zhu et al., 2007; Brandan and Gutierrez, 2013), а также ограничивает фибриллогенез путем прямого связывания с коллагеном I типа (Reese et al., 2013). . Другой SLRP представлен бигликаном, который конкурирует с декорином за тот же сайт связывания на коллагене (Schönherr et al., 1995) и, вероятно, играет роль как в формировании, так и в регенерации мышц (Brandan et al., 2008).
Транскрипционная регуляция образования белков, по-видимому, является важным фактором пластичности ВКМ. В этом отношении было показано, что экспрессия белка в скелетных мышцах слабо регулируется на уровне мРНК, что приводит к большим различиям в количестве мРНК и белка в различных тканях (Wang et al. , 2019). Интересно, что паттерн регуляции белка зависит от функции белка, при этом связь между мРНК и белком выше для внеклеточного матрикса и организации коллагеновых фибрилл (Makhnovskiy et al., 2020). Другим интересным аспектом регуляции количества белков ECM является тот факт, что индукция транскрипции, по-видимому, происходит довольно медленно для коллагена, поскольку для полной индукции транскрипции требуется почти 3 дня. Напротив, скорость секреции быстро адаптируется, так как она повышается менее чем за 1 час. В клетках с высоким уровнем выработки коллагена этот путь контролируется посттранскрипционной регуляцией, которая требует контроля с обратной связью между скоростью секреции и трансляции (rev. Schwarz, 2015).
, 2019). Интересно, что паттерн регуляции белка зависит от функции белка, при этом связь между мРНК и белком выше для внеклеточного матрикса и организации коллагеновых фибрилл (Makhnovskiy et al., 2020). Другим интересным аспектом регуляции количества белков ECM является тот факт, что индукция транскрипции, по-видимому, происходит довольно медленно для коллагена, поскольку для полной индукции транскрипции требуется почти 3 дня. Напротив, скорость секреции быстро адаптируется, так как она повышается менее чем за 1 час. В клетках с высоким уровнем выработки коллагена этот путь контролируется посттранскрипционной регуляцией, которая требует контроля с обратной связью между скоростью секреции и трансляции (rev. Schwarz, 2015).
Что касается ремоделирования тканей, то в регуляции гомеостаза внеклеточного матрикса участвуют два семейства ферментов, матриксные металлопротеиназы (ММР) и тканевые ингибиторы металлопротеиназ (ТИМП). MMP представляют собой протеолитические ферменты, которые расщепляют различные типы коллагенов и ингибируются TIMP (Visse and Nagase, 2003; Alameddine, 2012). В частности, ММП-1 и ММП-8 инициируют деградацию коллагенов I и III (преобладают в эндо-, пери- и эпимизии), тогда как ММП-2 и ММП-9 разрушают коллаген IV типа (основной коллагеновый компонент базальная мембрана) (Corcoran et al., 1996). ТИМП-1, -2 и -4 способны ингибировать все известные ММР (Christensen and Purslow, 2016).
В частности, ММП-1 и ММП-8 инициируют деградацию коллагенов I и III (преобладают в эндо-, пери- и эпимизии), тогда как ММП-2 и ММП-9 разрушают коллаген IV типа (основной коллагеновый компонент базальная мембрана) (Corcoran et al., 1996). ТИМП-1, -2 и -4 способны ингибировать все известные ММР (Christensen and Purslow, 2016).
ВКМ и сила скелетных мышц
Взаимодействие актина и миозина, а также многих других саркомерных белков приводит к укорочению мышечных волокон. Традиционные биомеханические модели часто изображают мышечно-сухожильные единицы как системы, в которых силы, возникающие при укорочении волокон, передаются продольно вдоль мышечного волокна и далее, в мышечно-сухожильном соединении, на сухожилие.Вблизи мышечно-сухожильного соединения миофибриллы имеют пальцевидные отростки, образованные инвагинациями плазматической мембраны (Knudsen et al., 2015). Эта структура увеличивает площадь поверхности, доступную для передачи силы. Ожидается, что передача силы будет происходить между пальцевидными отростками мышечных волокон и коллагеновыми волокнами, расположенными внутри инвагинаций, посредством сдвига базальной пластинки (Huijing, 1999). Содержащиеся здесь коллагены относятся к типу XXII, образующему внутренний слой, а также к типам III, VI, XII и XIV, лежащим дальше от мембраны мышечного волокна (Jakobsen et al., 2017). Хотя его точная роль до сих пор неясна, интересно отметить, что в мышцах коллаген XXII расположен исключительно в мышечно-сухожильном соединении. Было обнаружено, что у рыбок данио дефицит коллагена XXII приводит к мышечной дистрофии (Charvet et al., 2013), предполагая, что этот коллаген может служить для поддержания структурной целостности и стабилизации мышечно-сухожильного соединения.
Содержащиеся здесь коллагены относятся к типу XXII, образующему внутренний слой, а также к типам III, VI, XII и XIV, лежащим дальше от мембраны мышечного волокна (Jakobsen et al., 2017). Хотя его точная роль до сих пор неясна, интересно отметить, что в мышцах коллаген XXII расположен исключительно в мышечно-сухожильном соединении. Было обнаружено, что у рыбок данио дефицит коллагена XXII приводит к мышечной дистрофии (Charvet et al., 2013), предполагая, что этот коллаген может служить для поддержания структурной целостности и стабилизации мышечно-сухожильного соединения.
Учитывая тот факт, что значительная часть волокон в длинных мышцах оканчивается внутрипучково, не достигая непосредственно сухожилия (Barrett, 1962; Hijikata et al., 1993), однако ясно, что мышечно-сухожильный путь не может представлять собой единственный механизм передачи силы. Оканчивающиеся внутрипучковые волокна должны полагаться на среду, расположенную параллельно им, чтобы передавать свои усилия на пассивные компоненты опорно-двигательного аппарата (Sheard, 2000). Как впервые было установлено Street (1983), именно сеть IMCT внутри ECM способствует такой латеральной передаче сократительной силы. Передача силы по сети IMCT происходит от сократительных белков через костамер к эндомизию (Bloch and Gonzalez-Serratos, 2003; Peter et al., 2011) – как предполагают исследования моделирования, посредством сдвига (Sharafi and Blemker, 2011; Zhang and Gao, 2012) – и далее к перимизию, который в конечном итоге сливается с апоневрозами и сухожилиями (Passerieux et al., 2007). Первая информация о пропорциях передачи продольной и латеральной силы в поперечно-полосатых мышцах связана с элегантными экспериментами Huijing et al. (1998). После отделения прямых соединений нескольких головок длинного разгибателя пальцев крысы, что соответствует 55% общей мышечной массы, от сухожилия сустава, Huijing et al.(1998) наблюдали, что сила сохраняется на уровне 84% от силы интактной мышцы. Совсем недавно Рамасвами и соавт. (2011) использовали ярма для непосредственного измерения сил, передаваемых по продольному и поперечному пути.
Как впервые было установлено Street (1983), именно сеть IMCT внутри ECM способствует такой латеральной передаче сократительной силы. Передача силы по сети IMCT происходит от сократительных белков через костамер к эндомизию (Bloch and Gonzalez-Serratos, 2003; Peter et al., 2011) – как предполагают исследования моделирования, посредством сдвига (Sharafi and Blemker, 2011; Zhang and Gao, 2012) – и далее к перимизию, который в конечном итоге сливается с апоневрозами и сухожилиями (Passerieux et al., 2007). Первая информация о пропорциях передачи продольной и латеральной силы в поперечно-полосатых мышцах связана с элегантными экспериментами Huijing et al. (1998). После отделения прямых соединений нескольких головок длинного разгибателя пальцев крысы, что соответствует 55% общей мышечной массы, от сухожилия сустава, Huijing et al.(1998) наблюдали, что сила сохраняется на уровне 84% от силы интактной мышцы. Совсем недавно Рамасвами и соавт. (2011) использовали ярма для непосредственного измерения сил, передаваемых по продольному и поперечному пути. Их результаты не только подтвердили, что более 50% усилия передается латерально, но также показали, что латеральная передача силы была значительно снижена как у дистрофических, так и у старых грызунов. Их результаты позже подтвердили Чжан и Гао (2014).
Их результаты не только подтвердили, что более 50% усилия передается латерально, но также показали, что латеральная передача силы была значительно снижена как у дистрофических, так и у старых грызунов. Их результаты позже подтвердили Чжан и Гао (2014).
Некоторые аргументы предполагают, что латеральная передача силы является биомеханической необходимостью для поддержания целостности мышц и повышения эффективности сокращения.Во-первых, он помогает распределить сократительные силы по всей поверхности миофибрилл, что снижает механическое напряжение и предохраняет волокна от перерастяжения. Это может быть особенно важно в концевых участках волокон, которые обычно сужены и, следовательно, плохо приспособлены для переноса чрезмерных усилий (Monti et al., 1999). Косвенным подтверждением этой гипотезы являются исследования пожилых людей (Hughes et al., 2016) или пациентов, страдающих дистрофией Дюшенна (Virgilio et al., 2015), у которых дистрофин (т.э., костамерный белок, устанавливающий механическую связь между цитоскелетом, сарколеммой и ВКМ и, таким образом, облегчающий передачу латеральной силы), либо теряется, либо нарушается, и увеличивается восприимчивость к травмам растяжения мышц.
Кроме того, считается, что передача латеральной силы связывает волокна моста, сокращающиеся либо в разное время, либо в неодинаковой степени (Yucesoy et al., 2006), что помогает поддерживать выравнивание волокон и, таким образом, структурную целостность мышцы (Purslow, 2002).Недавно Дитрих и соавт. (2017) сравнили начало сокращения по результатам электромиографии и УЗИ в М-режиме. Вопреки интуиции, авторы обнаружили, что начало движения предшествует сигналу электромиографии примерно в 20% испытаний, что может быть объяснено переносом латеральной силы. Действительно, в то время как продольная передача сил может быть отложена из-за необходимости натягивать эластичные элементы, расположенные последовательно с мышцей (Nordez et al., 2009), трансламинарная сдвиговая связь между мышечными волокнами и сетью IMCT может обеспечить немедленную передачу силы. .Наконец, боковая передача силы обеспечивает механизм, посредством которого сила все еще может генерироваться и передаваться от мышечных волокон, которые прерываются из-за микротравмы или во время роста мышц (Purslow, 2010).
Помимо своей роли в латеральной передаче сократительной силы, внеклеточный матрикс также может влиять на укорочение мышечных волокон. Часто предполагается, что сократительная способность миофибрилл ограничена геометрией составляющих его саркомеров: саркомер и, таким образом, укорочение волокна прекращается, когда z -полосы вступают в контакт с миозиновыми филаментами.Однако эти идеи рассматривают только поведение саркомера как независимого исполнительного механизма. В условиях in vivo мышечные волокна встраиваются в сеть IMCT, что может препятствовать укорочению волокон. Действительно, принцип постоянного объема (Baskin, Paolini, 1967) диктует, что при укорочении мышечные волокна должны подвергаться радиальному расширению, что уже давно подтверждено экспериментально даже на саркомерном уровне (Brandt et al., 1967). Новые вычислительные модели и 90 196 измерений in situ 90 197 в мышцах лягушки, проведенные Azizi et al.(2017) продемонстрировали, что укорочение мышц затруднено, когда радиальное расширение ограничено физическими ограничениями.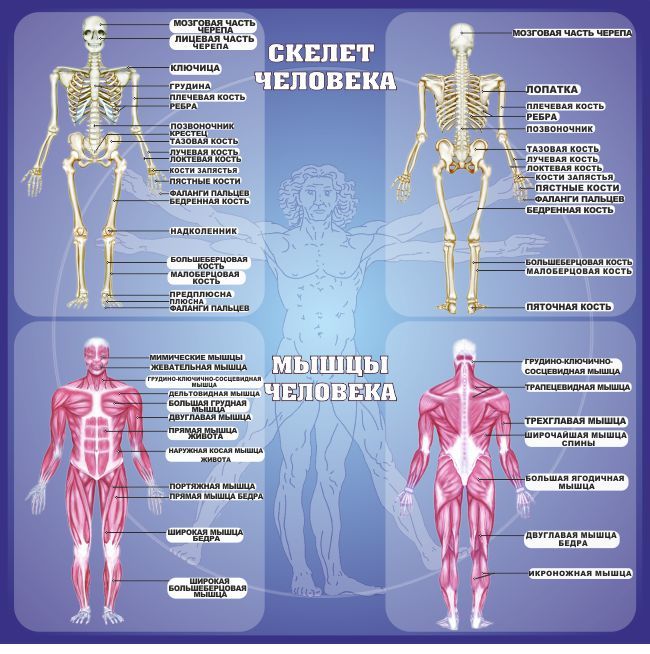 Следовательно, изменения количества и механических свойств сети IMCT, в которую встроены мышечные волокна, могут напрямую влиять на сократительную способность скелетных мышц. Такой сценарий может быть представлен мышечным фиброзом (Gillies et al., 2017).
Следовательно, изменения количества и механических свойств сети IMCT, в которую встроены мышечные волокна, могут напрямую влиять на сократительную способность скелетных мышц. Такой сценарий может быть представлен мышечным фиброзом (Gillies et al., 2017).
ECM в развитии, росте и восстановлении скелетных мышц
Помимо передачи силы, ВКМ скелетных мышц выполняет несколько важных функциональных функций.По-видимому, сеть IMCT обеспечивает механическую поддержку мышечных волокон, а также поддерживающих их нервов и кровеносных сосудов. Кровеносные капилляры проходят в интерстициях, занятых эндомизием, при этом их количество и плотность зависят от размера мышечных волокон (Janacek et al., 2009). Помимо этой наиболее очевидной роли, взаимодействие между миобластами, дифференцированными мышечными волокнами и компонентами ВКМ имеет центральное значение для эмбриогенного развития, дальнейшего роста и восстановления мышечной ткани.
Клеточным источником коллагеновых компонентов мышечного ВКМ являются специализированные IMCT фибробласты, которые происходят из различных эмбриогенных источников, включая сомиты (Nowicki et al. , 2003), мезодерму латеральной пластинки (Pearse et al., 2007) и нейральную клетки гребня (Olsson et al., 2001). Поскольку они продуцируют не только фибробласты, но и адипогенные клетки, фибробласты IMCT можно рассматривать как фиброадипогенные предшественники (Uezumi et al., 2010). Недавние исследования предоставили доказательства того, что, помимо этих очевидных ролей, фибробласты IMCT и продуцируемые ими соединительные ткани влияют как на миогенез (т.е., формирование мышечных предшественников и их дифференцировка в многоядерные миофибриллы) и морфогенез мышц (т. Эти сложные регуляторные процессы, происходящие во время эмбриогенного развития, здесь подробно не рассматриваются, но подробно рассмотрены в другом месте (Nassari et al., 2017; Sefton and Kardon, 2019). Вкратце, IMCT направляет мышечные предшественники к назначенным им областям-мишеням посредством комбинации сигналов притяжения (фактор роста гепатоцитов, фактор, полученный из стромальных клеток) и сигналов отталкивания (эфрин) (Dietrich et al.
, 2003), мезодерму латеральной пластинки (Pearse et al., 2007) и нейральную клетки гребня (Olsson et al., 2001). Поскольку они продуцируют не только фибробласты, но и адипогенные клетки, фибробласты IMCT можно рассматривать как фиброадипогенные предшественники (Uezumi et al., 2010). Недавние исследования предоставили доказательства того, что, помимо этих очевидных ролей, фибробласты IMCT и продуцируемые ими соединительные ткани влияют как на миогенез (т.е., формирование мышечных предшественников и их дифференцировка в многоядерные миофибриллы) и морфогенез мышц (т. Эти сложные регуляторные процессы, происходящие во время эмбриогенного развития, здесь подробно не рассматриваются, но подробно рассмотрены в другом месте (Nassari et al., 2017; Sefton and Kardon, 2019). Вкратце, IMCT направляет мышечные предшественники к назначенным им областям-мишеням посредством комбинации сигналов притяжения (фактор роста гепатоцитов, фактор, полученный из стромальных клеток) и сигналов отталкивания (эфрин) (Dietrich et al.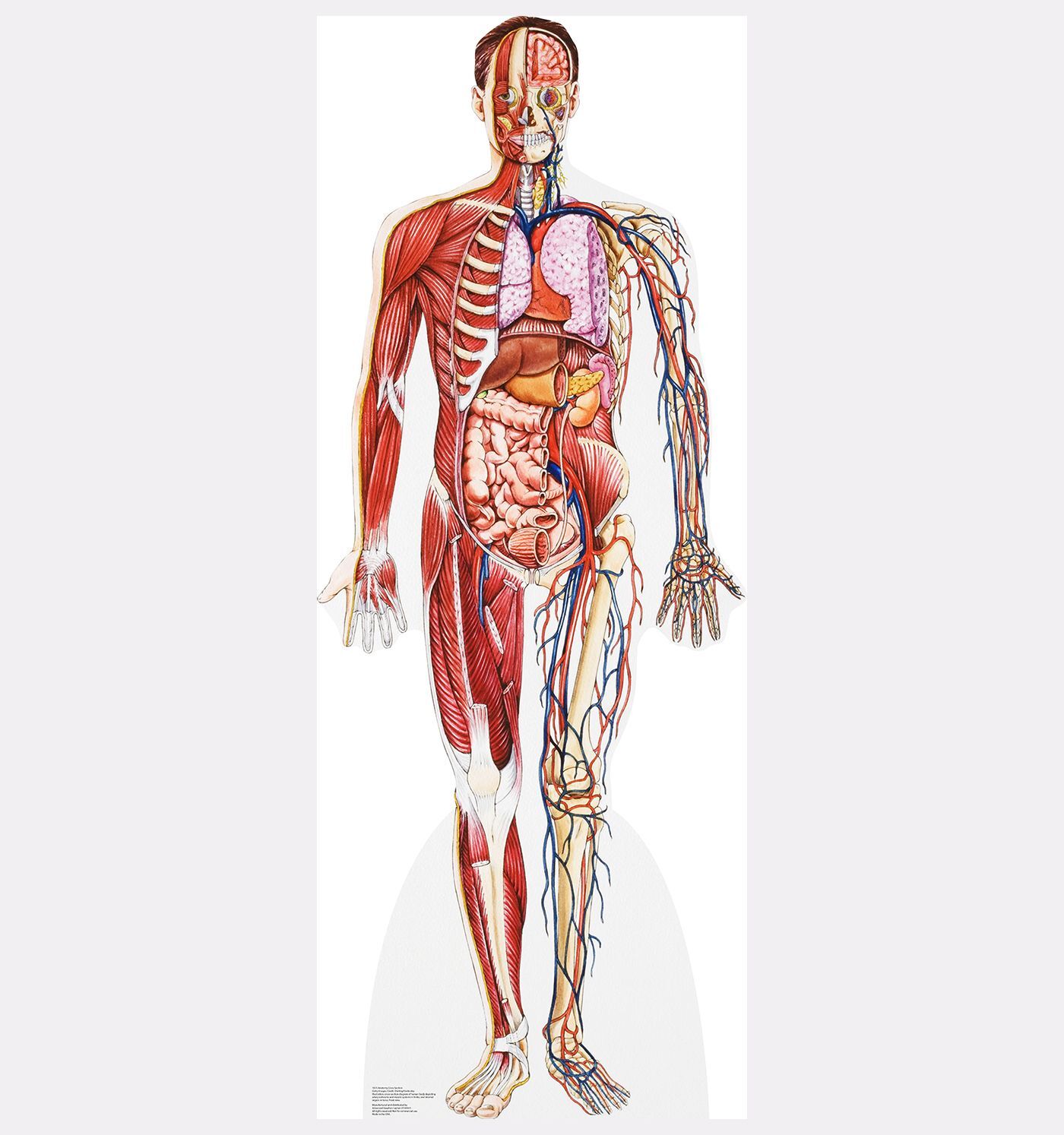 , 1999; Шварц и др., 2001). Через множество факторов транскрипции, экспрессируемых в фибробластах IMCT, IMCT затем способствует пролиферации, выживанию и дифференцировке соседних миобластов в зрелые миофибриллы (Kardon et al., 2003; Hasson et al., 2010; Iwata et al., 2013; Vallecillo). -Гарсия и др., 2017). Таким образом, можно предположить, что IMCT служит мезодермальным препаттерном, который контролирует участки дифференцировки миофибрилл и, следовательно, окончательное положение, размер и форму мышц.
, 1999; Шварц и др., 2001). Через множество факторов транскрипции, экспрессируемых в фибробластах IMCT, IMCT затем способствует пролиферации, выживанию и дифференцировке соседних миобластов в зрелые миофибриллы (Kardon et al., 2003; Hasson et al., 2010; Iwata et al., 2013; Vallecillo). -Гарсия и др., 2017). Таким образом, можно предположить, что IMCT служит мезодермальным препаттерном, который контролирует участки дифференцировки миофибрилл и, следовательно, окончательное положение, размер и форму мышц.
Будучи постмитотическими тканями, скелетные мышцы зависят от клеток-сателлитов, чтобы адаптироваться и регенерировать на протяжении всей жизни. Эти стволовые клетки располагаются в специализированных нишах между сарколеммой мышечных волокон и инкапсулирующими их базальными мембранами. Поддержание, активация и дифференцировка сателлитных клеток регулируются сложными каскадами транскрипционных факторов. Подробный обзор этих сотовых схем можно найти в недавнем обзоре Almada and Wagers (2016). Особое значение для этой рукописи имеет растущее количество данных, свидетельствующих о том, что на судьбу сателлитных клеток также сильно влияют взаимодействия с нишей ECM, в которой они находятся.Действительно, как динамическая среда, ниша стволовых клеток передает механические и химические сигналы, которые защищают покоящиеся стволовые клетки или вызывают активацию, пролиферацию и дифференцировку.
Особое значение для этой рукописи имеет растущее количество данных, свидетельствующих о том, что на судьбу сателлитных клеток также сильно влияют взаимодействия с нишей ECM, в которой они находятся.Действительно, как динамическая среда, ниша стволовых клеток передает механические и химические сигналы, которые защищают покоящиеся стволовые клетки или вызывают активацию, пролиферацию и дифференцировку.
В состоянии покоя сателлитные клетки экспрессируют канонический клеточный регулятор парного белка 7 (PAX7) (Olguin and Olwin, 2004). Исследования in vitro показали, что большая часть сателлитных клеток экспрессирует PAX7 при культивировании на матригеле, смеси белков ECM и факторов роста (Wilschut et al., 2010; Грефте и др., 2012). Дальнейшее подтверждение представления о том, что ECM активно участвует в поддержании покоя сателлитных клеток, происходит из сообщений о том, что сателлитные клетки, удаленные из своей ниши, быстро вступают в клеточный цикл и теряют способность к миогенной дифференцировке (Gilbert et al. , 2010). Интересно, что сателлитные клетки, по-видимому, также способны ощущать и реагировать на различные механические свойства ВКМ. Фактически, экспрессия PAX7 и выживаемость сателлитных клеток выше при культивировании на гидрогелях, имитирующих физиологическую жесткость мышц (Gilbert et al., 2010). Кроме того, сателлитные клетки, культивируемые на мягком гидрогеле, обладают большей функциональной способностью после трансплантации в реципиентную мышцу (Cosgrove et al., 2014).
, 2010). Интересно, что сателлитные клетки, по-видимому, также способны ощущать и реагировать на различные механические свойства ВКМ. Фактически, экспрессия PAX7 и выживаемость сателлитных клеток выше при культивировании на гидрогелях, имитирующих физиологическую жесткость мышц (Gilbert et al., 2010). Кроме того, сателлитные клетки, культивируемые на мягком гидрогеле, обладают большей функциональной способностью после трансплантации в реципиентную мышцу (Cosgrove et al., 2014).
Кроме того, было показано, что компоненты ECM влияют на деление стволовых клеток. В частности, белки фибронектин (Bentzinger et al., 2013b) и коллаген VI (Urciuolo et al., 2013), а также протеогликаны синдекан 3, синдекан 4, перлекан и декорин (Cornelison et al., 2001; Brack et al. ., 2008) были идентифицированы как составляющие ниши, влияющие на баланс между дифференцировкой и самообновлением и, таким образом, на поддержание регенеративной способности скелетных мышц.
При мышечной травме или в ответ на повышенную нагрузку обычно в основном покоящиеся сателлитные клетки активируются и дифференцируются в миобласты, которые в конечном итоге сливаются в зрелые миофибриллы. Хотя этот процесс требует своевременной экспрессии различных транскрипционных факторов, таких как миогенный фактор 5, миогенный детерминирующий белок или миогенин (Almada and Wagers, 2016), несколько исследований указывают на влияние ECM на каждый из этих этапов. Эксперименты с миобластами мыши (Grefte et al., 2012) или свиньи (Wilschut et al., 2010) показали, что на слияние миобластов положительно влияет матригель, но не отдельные субстраты, присутствующие в нише ECM. Вклад отдельных белков все еще плохо изучен, однако одновременное присутствие поли- D -лизина и ламинина (Boonen et al., 2009), гликозаминогликанов (Rønning et al., 2013) и протеогликанов сульфата гепарина (Gutiérrez and Brandan, 2010), по-видимому, играют заметную роль в пролиферации и дифференцировке сателлитных клеток. При активации стволовых клеток скелетных мышц локальное ремоделирование ВКМ сопровождается отложением ламинина-α1 и ламинина-α5 в базальной пластинке ниши сателлитных клеток (Rayagiri et al.
Хотя этот процесс требует своевременной экспрессии различных транскрипционных факторов, таких как миогенный фактор 5, миогенный детерминирующий белок или миогенин (Almada and Wagers, 2016), несколько исследований указывают на влияние ECM на каждый из этих этапов. Эксперименты с миобластами мыши (Grefte et al., 2012) или свиньи (Wilschut et al., 2010) показали, что на слияние миобластов положительно влияет матригель, но не отдельные субстраты, присутствующие в нише ECM. Вклад отдельных белков все еще плохо изучен, однако одновременное присутствие поли- D -лизина и ламинина (Boonen et al., 2009), гликозаминогликанов (Rønning et al., 2013) и протеогликанов сульфата гепарина (Gutiérrez and Brandan, 2010), по-видимому, играют заметную роль в пролиферации и дифференцировке сателлитных клеток. При активации стволовых клеток скелетных мышц локальное ремоделирование ВКМ сопровождается отложением ламинина-α1 и ламинина-α5 в базальной пластинке ниши сателлитных клеток (Rayagiri et al. , 2018). На мышах было показано, что сателлитные клетки мышц продуцируют коллагены ECM для поддержания покоя клеточно-автономным образом, при этом коллаген V является критическим компонентом ниши покоя, поскольку истощение приводит к аномальному входу в клеточный цикл и постепенному уменьшению количества стволовых клеток. бассейн (Багдади и др., 2018). Так же, как и для поддержания покоя, адекватные механические свойства ниши ECM также могут быть важны для созревания сателлитных клеток. Действительно, было обнаружено, что мышечные трубки оптимально дифференцируются на субстратах с мышечной жесткостью (Engler et al., 2004). В совокупности эти данные свидетельствуют о том, что жесткость ВКМ, сопровождающая как различные заболевания опорно-двигательного аппарата, так и процесс старения, может негативно влиять на регенеративную способность мышц.
, 2018). На мышах было показано, что сателлитные клетки мышц продуцируют коллагены ECM для поддержания покоя клеточно-автономным образом, при этом коллаген V является критическим компонентом ниши покоя, поскольку истощение приводит к аномальному входу в клеточный цикл и постепенному уменьшению количества стволовых клеток. бассейн (Багдади и др., 2018). Так же, как и для поддержания покоя, адекватные механические свойства ниши ECM также могут быть важны для созревания сателлитных клеток. Действительно, было обнаружено, что мышечные трубки оптимально дифференцируются на субстратах с мышечной жесткостью (Engler et al., 2004). В совокупности эти данные свидетельствуют о том, что жесткость ВКМ, сопровождающая как различные заболевания опорно-двигательного аппарата, так и процесс старения, может негативно влиять на регенеративную способность мышц.
Ремоделирование ВКМ мышц с возрастом
В более старшем возрасте скелетные мышцы обычно демонстрируют фиброзную морфологию (Lieber and Ward, 2013). В отличие от фасциального уплотнения, при котором общая структура коллагенов может сохраняться (Pavan et al., 2014), возрастной мышечный фиброз характеризуется утратой четкой двунаправленной решетчатой ориентации здоровых перимизиальных коллагеновых волокон и ее заменой. беспорядочной сетью волокон с уменьшенным образованием извитости (Järvinen et al., 2002). Кроме того, может быть увеличено абсолютное содержание коллагена и (неферментативное) поперечное связывание коллагеновых волокон (Haus et al., 2007b). Таким образом, модуль упругости внеклеточного матрикса может быть увеличен примерно в 35 раз (с ∼12 кПа у молодых до ∼418 кПа у старых мышц; Yin et al., 2013), причем этот эффект обусловлен накоплением плотно упакованных и сильно сшитый коллаген (Wood et al., 2014). У крупных долгоживущих животных, таких как тюлени Уэдделла, наблюдается увеличение внеклеточного пространства на 35–40% по мере увеличения общего и относительного содержания коллагена с возрастом.Однако это увеличение связано со сдвигом в сторону более высокого соотношения коллагена типа I и типа III (Hindle et al.
В отличие от фасциального уплотнения, при котором общая структура коллагенов может сохраняться (Pavan et al., 2014), возрастной мышечный фиброз характеризуется утратой четкой двунаправленной решетчатой ориентации здоровых перимизиальных коллагеновых волокон и ее заменой. беспорядочной сетью волокон с уменьшенным образованием извитости (Järvinen et al., 2002). Кроме того, может быть увеличено абсолютное содержание коллагена и (неферментативное) поперечное связывание коллагеновых волокон (Haus et al., 2007b). Таким образом, модуль упругости внеклеточного матрикса может быть увеличен примерно в 35 раз (с ∼12 кПа у молодых до ∼418 кПа у старых мышц; Yin et al., 2013), причем этот эффект обусловлен накоплением плотно упакованных и сильно сшитый коллаген (Wood et al., 2014). У крупных долгоживущих животных, таких как тюлени Уэдделла, наблюдается увеличение внеклеточного пространства на 35–40% по мере увеличения общего и относительного содержания коллагена с возрастом.Однако это увеличение связано со сдвигом в сторону более высокого соотношения коллагена типа I и типа III (Hindle et al. , 2009). Кроме того, концентрация коллагена IV типа увеличивается в базальной пластинке медленно сокращающихся мышц, тогда как концентрация ламинина, по-видимому, снижается с возрастом (Kovanen et al., 1988). Также было показано, что повышенное отложение белков базальной пластинки вытесняет сателлитные клетки из их ниш, что влияет на регуляцию деления сателлитных клеток (Snow, 1977) и может объяснить меньшее количество сателлитных клеток, обычно подсчитываемое в старых мышцах по сравнению с молодыми. (Брэк и др., 2007). Утрату и функциональную инактивацию стволовых клеток, негативно влияющую на гомеостаз тканей, можно считать общим признаком старения (López-Otín et al., 2013), который следует рассматривать как универсальную силу, способствующую старению мышц (Brack and Muñoz-Cánoves, 2016) и других тканей (Oh et al., 2014). Ожидается, что в дополнение к своему влиянию на сателлитные клетки дисрегуляция базальной мембраны также нарушит регенеративную способность мышц из-за неадекватной поддержки мышечных волокон и дезорганизованной ориентации каркаса (Sanes, 2003).
, 2009). Кроме того, концентрация коллагена IV типа увеличивается в базальной пластинке медленно сокращающихся мышц, тогда как концентрация ламинина, по-видимому, снижается с возрастом (Kovanen et al., 1988). Также было показано, что повышенное отложение белков базальной пластинки вытесняет сателлитные клетки из их ниш, что влияет на регуляцию деления сателлитных клеток (Snow, 1977) и может объяснить меньшее количество сателлитных клеток, обычно подсчитываемое в старых мышцах по сравнению с молодыми. (Брэк и др., 2007). Утрату и функциональную инактивацию стволовых клеток, негативно влияющую на гомеостаз тканей, можно считать общим признаком старения (López-Otín et al., 2013), который следует рассматривать как универсальную силу, способствующую старению мышц (Brack and Muñoz-Cánoves, 2016) и других тканей (Oh et al., 2014). Ожидается, что в дополнение к своему влиянию на сателлитные клетки дисрегуляция базальной мембраны также нарушит регенеративную способность мышц из-за неадекватной поддержки мышечных волокон и дезорганизованной ориентации каркаса (Sanes, 2003). Недавно Etienne et al. опубликовали обзор, включающий подробное описание влияния старения на внеклеточный матрикс скелетных мышц. (2020).
Недавно Etienne et al. опубликовали обзор, включающий подробное описание влияния старения на внеклеточный матрикс скелетных мышц. (2020).
Интересно, что данные профилирования транскрипции мышц, полученных от молодых и старых крыс, показывают, что из 682 наборов зондов, которые значительно различались между молодыми и старыми животными, 347 генов фактически уменьшились (а не увеличились) в старых/саркопенных мышцах по сравнению с молодыми мышцами. Было показано, что 24% из этих генов играют биологическую роль в ВКМ и клеточной адгезии (Pattison et al., 2003). Эти данные подтверждают гипотезу о том, что возрастные изменения ВКМ могут быть вызваны снижением способности к деградации, а не усилением синтеза коллагеновых структур. В частности, ММП, по-видимому, играют важную роль в этих процессах (de Sousa Neto et al., 2018). Это также подтверждается выводами, которые предполагают снижение способности к ремоделированию структур ВКМ в стареющих мышцах, вызванной упражнениями с отягощениями (Wessner et al.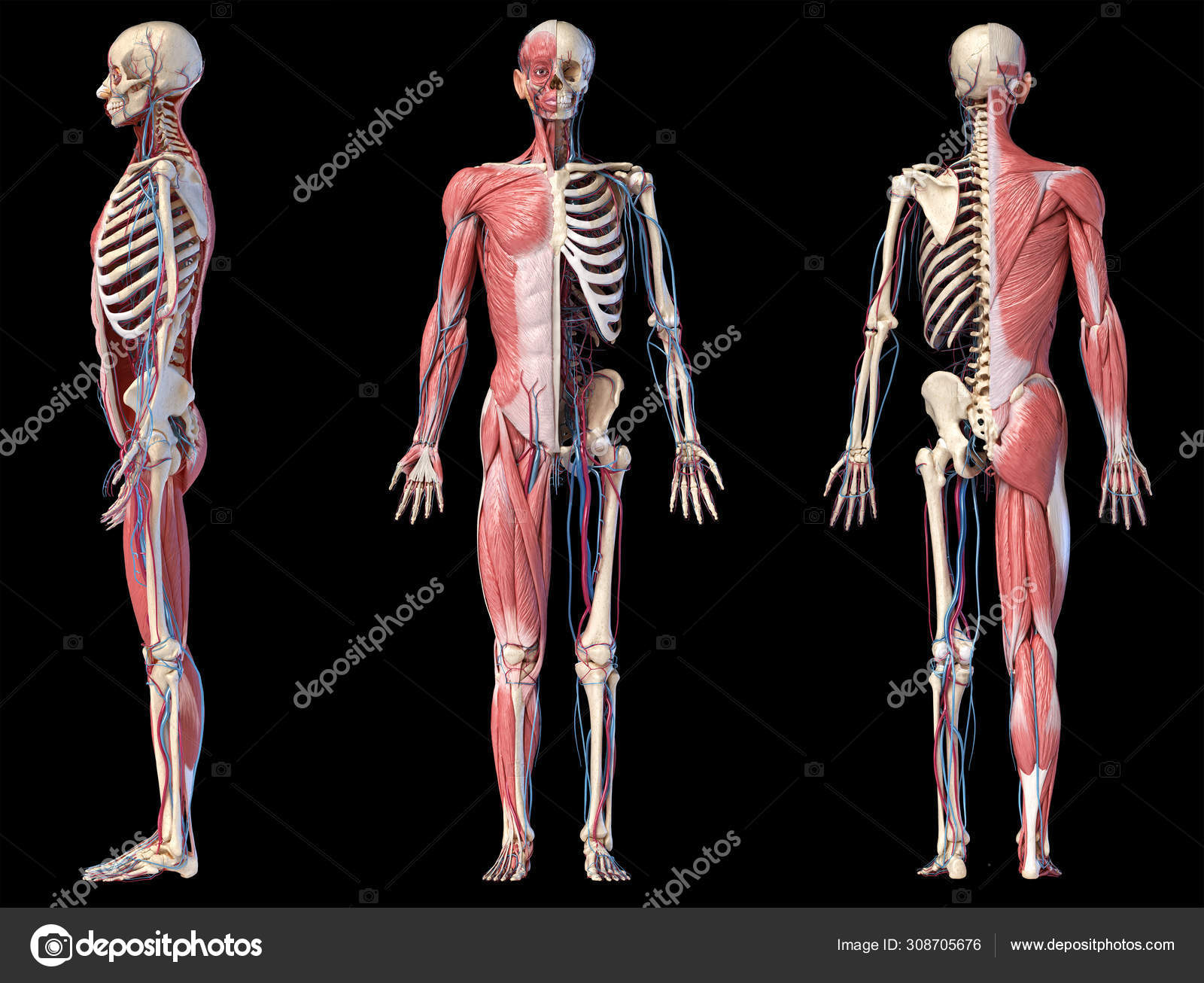 , 2019). Хотя механизмы еще полностью не изучены, считается, что эти изменения напрямую ухудшают функцию мышц, препятствуя сократительной способности волокон (Azizi et al., 2017) и передачу боковой силы (Sharafi and Blemker, 2011).
, 2019). Хотя механизмы еще полностью не изучены, считается, что эти изменения напрямую ухудшают функцию мышц, препятствуя сократительной способности волокон (Azizi et al., 2017) и передачу боковой силы (Sharafi and Blemker, 2011).
Нарушение регуляции ВКМ скелетных мышц в результате метаболических нарушений
Хорошо известно, что скелетные мышцы играют важную роль в стимулированном инсулином поглощении глюкозы (Richter and Hargreaves, 2013). Роль ECM в этом контексте может быть менее ясной. Мыши с дефицитом мышечно-специфического интегрина β1 демонстрируют снижение стимулированной инсулином скорости инфузии глюкозы и клиренса глюкозы, несмотря на отсутствие изменений в потреблении пищи, весе, уровне глюкозы натощак, уровне инсулина и экспрессии белка GLUT4 (Zong et al., 2009), предполагая взаимосвязь между аберрантной передачей сигналов интегрина и развитием диабета 2 типа. Кроме того, на модели диабета у животных было показано, что нарушение чувствительности к инсулину связано со снижением уровня белка изоформы дистрофина Dp427 и субкомплекса альфа/бета-дистрогликана (Mulvey et al. , 2005).
, 2005).
Повышенное количество коллагена типа I и III было обнаружено как у пациентов с диабетом 2 типа, так и у пациентов с ожирением, не страдающих диабетом (Berria et al., 2006), а переедание у людей было связано с увеличением экспрессии генов, связанных с IMCT (коллагены I, III, IV, V, SPARC, интегрин, Tam et al., 2014) и изменения в генных путях, связанных с взаимодействием с рецептором ЕСМ, фокальной адгезией и слипчивым соединением (Tam et al., 2017). Однако кормление мышей диетой с высоким содержанием жиров приводило к снижению уровня экспрессии генов COL1, COL3 и COL6, но не уровня белка (Tam et al., 2015).
Было показано, что деградация коллагенов посредством MMPs является важным компонентом ремоделирования ECM (Cui et al., 2017). Может ли это быть правдой в контексте диабета, было исследовано в исследовании на животных.Интересно, что генетическое истощение MMP9 не индуцировало резистентность к инсулину у худых мышей, несмотря на увеличение коллагена IV. Однако когда мышей кормили пищей с высоким содержанием жиров, делеция вызывала глубокую резистентность к инсулину.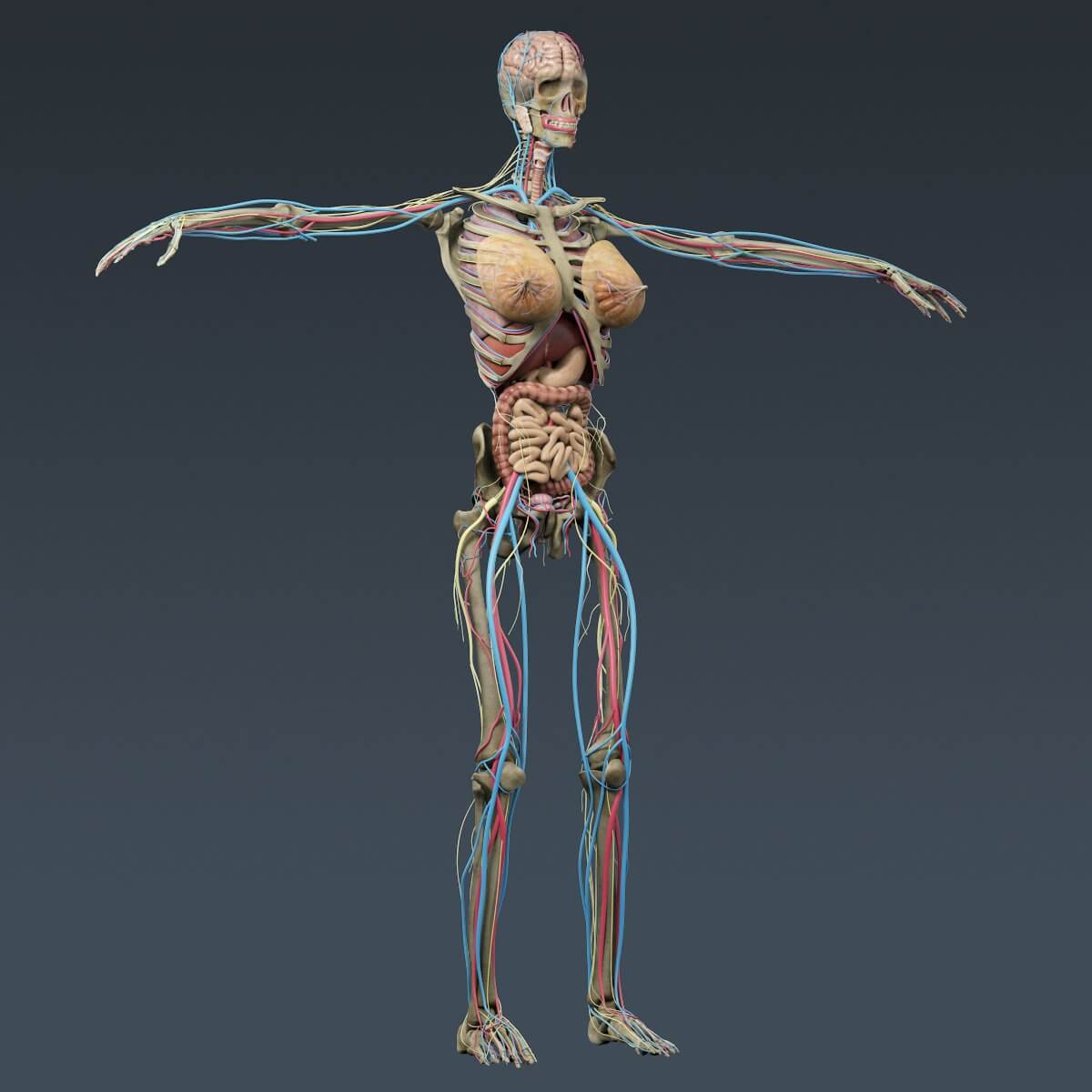 Эти результаты еще больше усиливают роль компонентов IMCT в прогрессировании мышечной инсулинорезистентности, особенно в состоянии переедания (Kang et al., 2014).
Эти результаты еще больше усиливают роль компонентов IMCT в прогрессировании мышечной инсулинорезистентности, особенно в состоянии переедания (Kang et al., 2014).
Наконец, гиалуронан, основной компонент внеклеточного матрикса, увеличивается при ожирении у мышей, вызванном диетой с высоким содержанием жиров.Лечение PEGPh30, которое дозозависимо снижает содержание гиалуроновой кислоты в мышечном ВКМ, предлагается для лечения резистентности к инсулину с сопутствующим уменьшением жировой массы, размера адипоцитов, а также резистентности к инсулину в печени и мышцах (Kang et al., 2013).
Подводя итог, можно сказать, что различные компоненты внеклеточного матрикса поражаются на разных стадиях диабета. Исследования о том, связан ли диабет с мышечной слабостью, противоречивы (Leong et al., 2015; Li et al., 2016), и еще предстоит выяснить, имеют ли непосредственное отношение к этому контексту изменения в путях, связанных с внеклеточным матриксом.
Адаптация к физической подготовке и неиспользованию
Первые данные, указывающие на пластичность IMCT в ответ на физическую активность, были опубликованы еще в 1970-х годах, когда Suominen and Heikkinen (1975) и Suominen et al. (1977) обнаружили повышенный уровень пролилгидроксилазы (фермента, способствующего биосинтезу коллагена) у спортсменов, тренирующихся на выносливость, а также в лонгитюдном исследовании после восьми недель аэробных тренировок. Позднее было обнаружено, что влияние упражнений на выносливость на активность проколлагеновых ферментов более выражено в красных мышцах по сравнению с белыми мышцами (Takala et al., 1983). Прямые измерения содержания коллагена, впервые проведенные в конце 1980-х годов, подтвердили, что содержание коллагена (типа IV) увеличилось в камбаловидной мышце крыс, устойчивой к утомлению, после продолжительных тренировок на выносливость (Kovanen et al., 1988). Несмотря на вызванное физической нагрузкой увеличение коллагена, Gosselin et al. (1998) обнаружили, что жесткости мышц, наблюдаемой с возрастом, можно противодействовать с помощью упражнений на выносливость, которые авторы связывают со снижением гидроксилизилпиридинолинового перекрестного связывания коллагеновых волокон.
(1977) обнаружили повышенный уровень пролилгидроксилазы (фермента, способствующего биосинтезу коллагена) у спортсменов, тренирующихся на выносливость, а также в лонгитюдном исследовании после восьми недель аэробных тренировок. Позднее было обнаружено, что влияние упражнений на выносливость на активность проколлагеновых ферментов более выражено в красных мышцах по сравнению с белыми мышцами (Takala et al., 1983). Прямые измерения содержания коллагена, впервые проведенные в конце 1980-х годов, подтвердили, что содержание коллагена (типа IV) увеличилось в камбаловидной мышце крыс, устойчивой к утомлению, после продолжительных тренировок на выносливость (Kovanen et al., 1988). Несмотря на вызванное физической нагрузкой увеличение коллагена, Gosselin et al. (1998) обнаружили, что жесткости мышц, наблюдаемой с возрастом, можно противодействовать с помощью упражнений на выносливость, которые авторы связывают со снижением гидроксилизилпиридинолинового перекрестного связывания коллагеновых волокон.
Эффекты иммобилизации на ВКМ скелетных мышц не совсем однозначны. Ранние исследования Karpakka et al. (1990, 1991) обнаружили у крыс снижение как активности гидроксилазы, так и содержания гидроксипролина (аминокислота, входящая в состав коллагена). Позднее было обнаружено, что изменения содержания коллагена в ответ на кратковременную иммобилизацию или неиспользование были довольно небольшими (Savolainen et al., 1988; Haus et al., 2007a), что можно объяснить относительно низкой скоростью обновления. Более недавнее исследование, напротив, показало, что содержание коллагена I и биомеханические свойства (модуль упругости, максимальное напряжение и предел текучести) фасции голени, покрывающей трехглавую мышцу голени крысы, значительно увеличились уже через 21 день разгрузки задних конечностей. (Хуанг и др., 2018). Интересно, что эти изменения можно было предотвратить, применяя вибрацию к задним лапам крыс два раза в день. У нетренирующихся людей иммуногистохимическое окрашивание показало отсутствие изменений в плотности сети коллагена I после 60 дней постельного режима. У испытуемых, выполняющих протокол контрмерных упражнений, состоящий из реактивных прыжков на системе саней, напротив, иммунореактивность коллагена I была снижена по сравнению с исходными уровнями (Schoenrock et al., 2018).
У испытуемых, выполняющих протокол контрмерных упражнений, состоящий из реактивных прыжков на системе саней, напротив, иммунореактивность коллагена I была снижена по сравнению с исходными уровнями (Schoenrock et al., 2018).
Еще одной моделью, позволяющей изучать адаптивность ВКМ мышц, является функциональная перегрузка, вызванная хирургическим устранением синергистов.В одном из первых соответствующих исследований Уильямс и Голдспинк (1981) перерезали сухожилия подошвенной и икроножной мышц самцов крыс, чтобы перегрузить камбаловидную мышцу. Гипертрофия мышц, наблюдаемая через три недели после тенотомии, сопровождалась повышением концентрации КИМТ (>45%) и отношения КМКТ к мышечной ткани. Гистологический анализ также показал, что увеличение IMCT было в основном связано с утолщением эндомизия. Сосредоточившись на мышечно-сухожильном соединении, Zamora и Marini (1988) провели аналогичные эксперименты и выделили подошвенную мышцу крысы посредством тенотомии камбаловидной мышцы и абляции икроножных мышц. По сравнению с контрольными животными фибробласты, расположенные в мышечно-сухожильном соединении, развивали более высокую степень активации цитоплазмы, ядра и ядрышка уже через одну-две недели функциональной перегрузки. В более позднем исследовании было проверено влияние IL-6 на индуцированное перегрузкой ремоделирование ECM путем сравнения мышей дикого типа и мышей с нокаутом IL-6 (White et al., 2009). В то время как увеличение площади поперечного сечения миофибрилл было аналогичным после 21 дня функциональной перегрузки, увеличение сырой массы мышц было значительно больше у мышей с нокаутом по IL-6.Гистологический анализ подтвердил, что это избыточное увеличение мышечной массы может быть объяснено значительно большим увеличением содержания несократительной ткани и концентрации гидроксипролина, что свидетельствует о содержании коллагена и фиброзе. В соответствии с этим наблюдением уровни мРНК проколлагена-1, IGF-1 и TGF-β были значительно выше у мышей с дефицитом IL-6. Наоборот, экспрессия мРНК MyoD, фактора транскрипции, необходимого для мио-, а не фиброгенной дифференцировки сателлитных клеток (Zammit, 2017), была значительно ослаблена у животных, лишенных IL-6.
По сравнению с контрольными животными фибробласты, расположенные в мышечно-сухожильном соединении, развивали более высокую степень активации цитоплазмы, ядра и ядрышка уже через одну-две недели функциональной перегрузки. В более позднем исследовании было проверено влияние IL-6 на индуцированное перегрузкой ремоделирование ECM путем сравнения мышей дикого типа и мышей с нокаутом IL-6 (White et al., 2009). В то время как увеличение площади поперечного сечения миофибрилл было аналогичным после 21 дня функциональной перегрузки, увеличение сырой массы мышц было значительно больше у мышей с нокаутом по IL-6.Гистологический анализ подтвердил, что это избыточное увеличение мышечной массы может быть объяснено значительно большим увеличением содержания несократительной ткани и концентрации гидроксипролина, что свидетельствует о содержании коллагена и фиброзе. В соответствии с этим наблюдением уровни мРНК проколлагена-1, IGF-1 и TGF-β были значительно выше у мышей с дефицитом IL-6. Наоборот, экспрессия мРНК MyoD, фактора транскрипции, необходимого для мио-, а не фиброгенной дифференцировки сателлитных клеток (Zammit, 2017), была значительно ослаблена у животных, лишенных IL-6. В совокупности эти результаты указывают на то, что элиминация синергистов вызывает увеличение содержания IMCT и, в частности, утолщение эндомизиальных структур в перегруженных мышцах. Эти адаптации могут служить для модуляции несократительных структур мышц в соответствии с повышенными функциональными требованиями. IGF-1, по-видимому, играет важную роль в регуляции этого процесса, поскольку было показано, что недостаток IGF-1 приводит к чрезмерному накоплению IMCT и, возможно, к нарушению регенеративного потенциала мышц.
В совокупности эти результаты указывают на то, что элиминация синергистов вызывает увеличение содержания IMCT и, в частности, утолщение эндомизиальных структур в перегруженных мышцах. Эти адаптации могут служить для модуляции несократительных структур мышц в соответствии с повышенными функциональными требованиями. IGF-1, по-видимому, играет важную роль в регуляции этого процесса, поскольку было показано, что недостаток IGF-1 приводит к чрезмерному накоплению IMCT и, возможно, к нарушению регенеративного потенциала мышц.
Одно из первых исследований по тестированию и сравнению различных форм упражнений с отягощениями у мужчин было проведено Brown et al.(1999), которые сообщили, что после одного приступа концентрических сокращений маркеры распада коллагена (гидроксипролин и сывороточный коллаген I типа) не увеличивались. Напротив, эксцентрические сокращения мышц повышают уровень коллагена в сыворотке более чем на 40% в течение 9 дней после тренировки, что указывает на то, что эксцентрические сокращения могут быть более эффективными в содействии разрушению коллагена. Эти результаты были подтверждены в двух более поздних исследованиях с использованием высокоинтенсивных эксцентрических упражнений, в которых были обнаружены повышенная обработка проколлагена и содержание коллагена IV типа, а также более высокая активность MMP и TIMP (Crameri et al., 2004; Макки и др., 2004). Интересно, что Crameri et al. (2004, 2007) также сообщили об увеличении тенасцина С, гликопротеина, присутствующего в ВКМ, который, как предполагается, направляет миграцию клеток после повреждения, независимо от того, было ли повреждение мышц вызвано произвольным или электрическим повреждением мышц. Временная активация тенасцина С и других гликопротеинов внеклеточного матрикса (например, фибронектина и гиалуроновой кислоты) обычно называют «транзиторной матрицей», появление которой считается важным первым шагом для успешного восстановления мышц, поскольку оно дает важные сигналы, управляющие регенеративный потенциал мышечных стволовых клеток (Calve et al., 2010; Тирни и др., 2016).
Эти результаты были подтверждены в двух более поздних исследованиях с использованием высокоинтенсивных эксцентрических упражнений, в которых были обнаружены повышенная обработка проколлагена и содержание коллагена IV типа, а также более высокая активность MMP и TIMP (Crameri et al., 2004; Макки и др., 2004). Интересно, что Crameri et al. (2004, 2007) также сообщили об увеличении тенасцина С, гликопротеина, присутствующего в ВКМ, который, как предполагается, направляет миграцию клеток после повреждения, независимо от того, было ли повреждение мышц вызвано произвольным или электрическим повреждением мышц. Временная активация тенасцина С и других гликопротеинов внеклеточного матрикса (например, фибронектина и гиалуроновой кислоты) обычно называют «транзиторной матрицей», появление которой считается важным первым шагом для успешного восстановления мышц, поскольку оно дает важные сигналы, управляющие регенеративный потенциал мышечных стволовых клеток (Calve et al., 2010; Тирни и др., 2016). Сообщается, что высвобождение гликопротеинов ECM сопровождается повышением активности MMP-9 у молодых и снижением активности MMP-9 и MMP-15 у пожилых людей (Wessner et al., 2019). Эти данные свидетельствуют о том, что острая тренировка с отягощениями вызывает катаболический ответ в молодых мышцах, но этот эффект может ослабевать в пожилом возрасте. Было обнаружено, что последующая анаболическая реакция, характеризующаяся активацией структурных коллагенов (I, III, IV) и ламинина, происходит со значительной задержкой, что позволяет предположить, что восстановление мышц после острого приступа повреждающих (удлиняющих) сокращений следует за двухфазный характер (Mackey et al., 2011; Хильдахл и др., 2015). Интересно, что недавнее исследование Sorensen et al. (2018) обнаружили, что появление переходной матрицы у физически активных пожилых людей было менее выраженным, чем у молодых. Это наблюдение подтверждает идею о том, что нарушение регуляции сигналов ВКМ может быть ответственным за повышенное отложение ВКМ и снижение активности стволовых клеток, обычно наблюдаемое в старых мышцах (Grounds, 1998).
Сообщается, что высвобождение гликопротеинов ECM сопровождается повышением активности MMP-9 у молодых и снижением активности MMP-9 и MMP-15 у пожилых людей (Wessner et al., 2019). Эти данные свидетельствуют о том, что острая тренировка с отягощениями вызывает катаболический ответ в молодых мышцах, но этот эффект может ослабевать в пожилом возрасте. Было обнаружено, что последующая анаболическая реакция, характеризующаяся активацией структурных коллагенов (I, III, IV) и ламинина, происходит со значительной задержкой, что позволяет предположить, что восстановление мышц после острого приступа повреждающих (удлиняющих) сокращений следует за двухфазный характер (Mackey et al., 2011; Хильдахл и др., 2015). Интересно, что недавнее исследование Sorensen et al. (2018) обнаружили, что появление переходной матрицы у физически активных пожилых людей было менее выраженным, чем у молодых. Это наблюдение подтверждает идею о том, что нарушение регуляции сигналов ВКМ может быть ответственным за повышенное отложение ВКМ и снижение активности стволовых клеток, обычно наблюдаемое в старых мышцах (Grounds, 1998).
Heinemeier et al. опубликовали одно из первых исследований, посвященных прямому сравнению различных форм мышечных сокращений с точки зрения их способности к острому ремоделированию внеклеточного матрикса.(2007). Эти авторы провели исследование на грызунах и обнаружили, что активность генов, связанных с биосинтезом коллагена (например, коллагенов I и III), а также факторов роста (например, TGF-β1) повышалась после всех форм физических упражнений, но наиболее заметно при этом. после эксцентрической тренировки. Было также высказано предположение, что у людей скорость фракционного синтеза белка коллагена выше после острого приступа эксцентрической тренировки по сравнению с концентрической тренировкой (Holm et al., 2017), хотя это мнение не бесспорно (Moore et al., 2005). В совокупности эти результаты свидетельствуют о том, что особенно эксцентрические упражнения являются мощным стимулом, который вызывает микротравмы и обновление клеток IMCT, причем последнее, как предполагается, представляет собой попытку организма предотвратить повторное повреждение мышц (Mackey et al. , 2011; Hyldahl et al. , 2015; Такаги и др., 2016). На самом деле, снижение активности ММР после длительной тренировки, состоящей из электрически вызванных изометрических сокращений у крыс, может отражать успешное усиление ВКМ (Ogasawara et al., 2014), тогда как длительное повышение активности ММР и ТИМП в подошвенной мышце мышей после хирургического удаления икроножной и камбаловидной мышцы может свидетельствовать о продолжающемся ремоделировании ВКМ (Mendias et al., 2017).
, 2011; Hyldahl et al. , 2015; Такаги и др., 2016). На самом деле, снижение активности ММР после длительной тренировки, состоящей из электрически вызванных изометрических сокращений у крыс, может отражать успешное усиление ВКМ (Ogasawara et al., 2014), тогда как длительное повышение активности ММР и ТИМП в подошвенной мышце мышей после хирургического удаления икроножной и камбаловидной мышцы может свидетельствовать о продолжающемся ремоделировании ВКМ (Mendias et al., 2017).
Помимо режима сокращения, внеклеточный матрикс скелетных мышц также может быть чувствителен к интенсивности упражнений. Кармели и др. (2005) проверили влияние бега на беговой дорожке как с высокой, так и с низкой интенсивностью на крысах и обнаружили, что ММР-2 (один из ферментов, ответственных за расщепление коллагена IV, в основном присутствующий в базальной мембране мышц) увеличивался после высокоинтенсивных упражнений. Только. Напротив, у людей одно исследование Holm et al. (2010) сравнили эффекты упражнений на одностороннее разгибание колена, выполняемых с низкой или высокой (16% или 70% от индивидуального одноповторного максимума соответственно) интенсивностью, с количеством повторений, скорректированным в соответствии с вмешательствами для общей нагрузки. поднят.В этом исследовании скорость фракционного синтеза коллагена была равномерно увеличена после обоих вмешательств.
поднят.В этом исследовании скорость фракционного синтеза коллагена была равномерно увеличена после обоих вмешательств.
Что касается адаптации ВКМ к продолжительным тренировкам с отягощениями, то существуют только данные исследований на животных. де Соуза Нето и др. (2018) сообщили, что 12 недель тренировок с отягощениями, состоящих из подъемов по лестнице с прогрессивными дополнительными нагрузками, эквивалентными 65–100% максимальной грузоподъемности каждого человека, повышали активность MMP-2 в подошвенных сгибателях мышц старых крыс, в то время как подавляли MMP-2. 2 и ММП-9 в кровообращении.Вывод авторов о том, что тренировки с отягощениями могут, таким образом, быть полезным инструментом для поддержания ремоделирования ВКМ в пожилом возрасте, недавно получил эмпирическое подтверждение в другом тренировочном исследовании на крысах, в котором использовался тот же тренировочный протокол, и было показано уменьшение отложения соединительной ткани у тренированных пожилых людей. мышцы (Guzzoni et al., 2018).
мышцы (Guzzoni et al., 2018).
Подводя итог, несколько исследований, изучающих острые эффекты физической активности как у грызунов, так и у людей, показали, что физические упражнения могут стимулировать как деградацию, так и синтез коллагена в скелетных мышцах.Репарация микротравм, вызванных физическими упражнениями, проходит по двухфазной схеме, при которой гликопротеины сначала создают переходную матрицу для управления катаболическими процессами, а анаболические процессы для укрепления структуры IMCT происходят со значительной задержкой. Потенциал упражнений для ремоделирования ВКМ, по-видимому, зависит от режима сокращения, при этом эксцентрические сокращения вызывают больший ответ, чем концентрические или изометрические мышечные действия. Доступно несколько исследований, проверяющих результаты упражнений различной интенсивности, и на данный момент результаты показывают, что расщепление белка (но не синтез) может сильнее провоцироваться более высокой интенсивностью.Неиспользование резко снижает активность ферментов, связанных с биосинтезом коллагена, хотя на уровне белков изменения происходят медленно.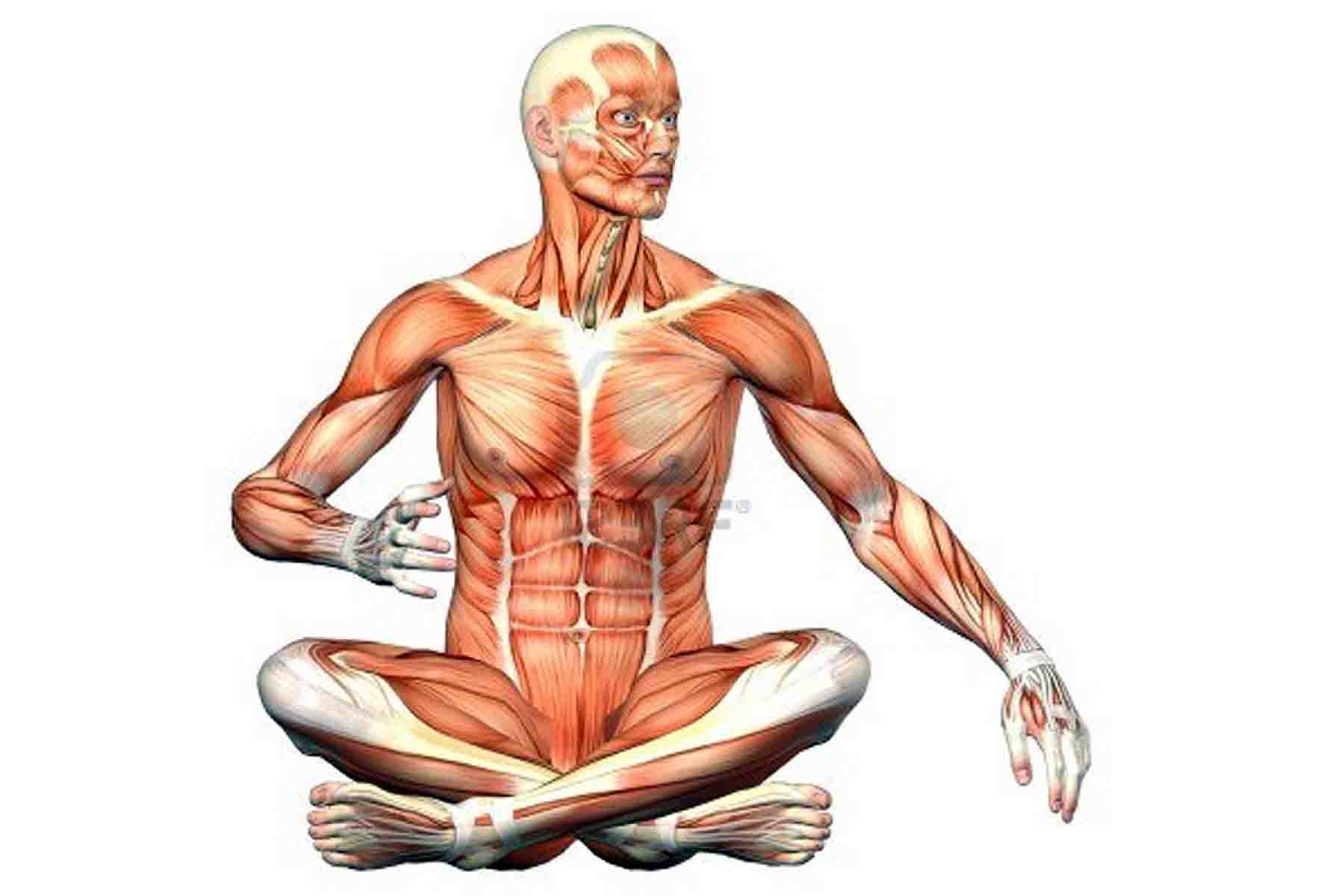 Поперечные сравнения с участием тренированных (в основном на выносливость) грызунов показывают, что хроническая физическая активность может привести к усиленному фенотипу IMCT. Единственные доступные на сегодняшний день длительные лонгитюдные исследования тренировок были проведены на грызунах и позволяют предположить, что длительные тренировки с отягощениями могут быть полезны для противодействия чрезмерному накоплению IMCT в пожилом возрасте.Физиологические и функциональные последствия ремоделирования IMCT, вызванного тренировкой, требуют дальнейшего изучения.
Поперечные сравнения с участием тренированных (в основном на выносливость) грызунов показывают, что хроническая физическая активность может привести к усиленному фенотипу IMCT. Единственные доступные на сегодняшний день длительные лонгитюдные исследования тренировок были проведены на грызунах и позволяют предположить, что длительные тренировки с отягощениями могут быть полезны для противодействия чрезмерному накоплению IMCT в пожилом возрасте.Физиологические и функциональные последствия ремоделирования IMCT, вызванного тренировкой, требуют дальнейшего изучения.
Заключение
Настоящий обзор направлен на то, чтобы дать обзор современного состояния знаний о ВКМ скелетных мышц, который играет важную, хотя часто недооцениваемую роль в поддержании мышечного гомеостаза, влияет на мышечную функцию и адаптацию и может быть ключом к лечению. мышечных и метаболических нарушений вследствие старения или болезней.
Будучи сложной сетью различных коллагенов, гликопротеинов, протеогликанов и эластина, внеклеточный матрикс включает в себя сократительные мышечные волокна и служит через интегрины и дистрофин-ассоциированный гликопротеиновый комплекс, соответственно, в качестве биохимического и механического интерфейса между мышечными клетками и их окружением. Сборке его коллагенового каркаса в основном способствуют факторы роста TGF-β и CTGF, которые регулируются различными протеогликанами, такими как декорин и бигликан. Кроме того, в регуляции ВКМ участвуют протеолитические ферменты (ММР), а также их ингибиторы (ТИМП).
Сборке его коллагенового каркаса в основном способствуют факторы роста TGF-β и CTGF, которые регулируются различными протеогликанами, такими как декорин и бигликан. Кроме того, в регуляции ВКМ участвуют протеолитические ферменты (ММР), а также их ингибиторы (ТИМП).
Функционально ВКМ служит средой для передачи сократительной силы, которая может служить не только для повышения эффективности мышечного сокращения, но и для защиты мышечных волокон от чрезмерного напряжения и облегчения заживления микротравм. В дополнение к своей функциональной роли ВКМ активно участвует в регуляции мышечного пула сателлитных клеток. Ниши ВКМ, расположенные между сарколеммой и базальной мембраной, защищают сателлитные клетки от включения в клеточный цикл и, таким образом, помогают поддерживать регенеративный потенциал мышц.Определенные компоненты ВКМ, такие как фибронектин, коллаген VI и различные протеогликаны, могут дополнительно способствовать делению стволовых клеток. И наоборот, было показано, что ламинин, гликозаминогликаны и другие протеогликаны способствуют дифференцировке сателлитных клеток и их слиянию в зрелые миофибриллы.
Научные данные дополнительно демонстрируют, что внеклеточный матрикс скелетных мышц представляет собой податливую ткань, которая может подвергаться процессам ремоделирования в результате старения, болезней, физических тренировок или неиспользования.В частности, старение обычно приводит к общему повышенному отложению коллагеновой ткани, изменениям в составе коллагена (сдвиг в сторону более высокого уровня коллагена I типа к типу III) и усилению неферментативного перекрестного связывания коллагена (посредством конечных продуктов гликирования). Эти изменения, которые, возможно, опосредованы снижением активности ММР, приводят к ригидности внеклеточного матрикса мышцы и могут нарушать ее функцию и регенеративный потенциал.
Ремоделирование внеклеточного матрикса также может быть связано с метаболическими нарушениями, такими как диабет.Было обнаружено, что чрезмерное потребление пищи приводит к увеличению экспрессии генов, связанных с ВКМ (коллагены I, III, IV, V, SPARC, интегрин). В свою очередь, такое ремоделирование может нарушать передачу сигналов интегрина, тем самым снижая чувствительность к инсулину. Другими компонентами ECM, потенциально представляющими мишени для резистентности к инсулину, являются гиалуронан, комплекс дистрофина-дистрогликана, а также MMP9.
В свою очередь, такое ремоделирование может нарушать передачу сигналов интегрина, тем самым снижая чувствительность к инсулину. Другими компонентами ECM, потенциально представляющими мишени для резистентности к инсулину, являются гиалуронан, комплекс дистрофина-дистрогликана, а также MMP9.
Наконец, ремоделирование ВКМ может быть вызвано физическими упражнениями. В то время как реальных исследований тренировок немного, есть данные, свидетельствующие о том, что упражнения могут резко способствовать как увеличению синтеза коллагена (коллагены I, III, TGF-β1), так и деградации (MMP2, MMP9).Поперечные исследования на людях и лонгитудинальные исследования на грызунах также предполагают, что такой повышенный обмен коллагена может привести к усилению коллагеновых структур у постоянно тренирующихся субъектов и предотвратить чрезмерное отложение коллагена (то есть фиброз) в мышцах пожилых людей. Исследования, изучающие последствия длительного неиспользования, показали противоречивые результаты.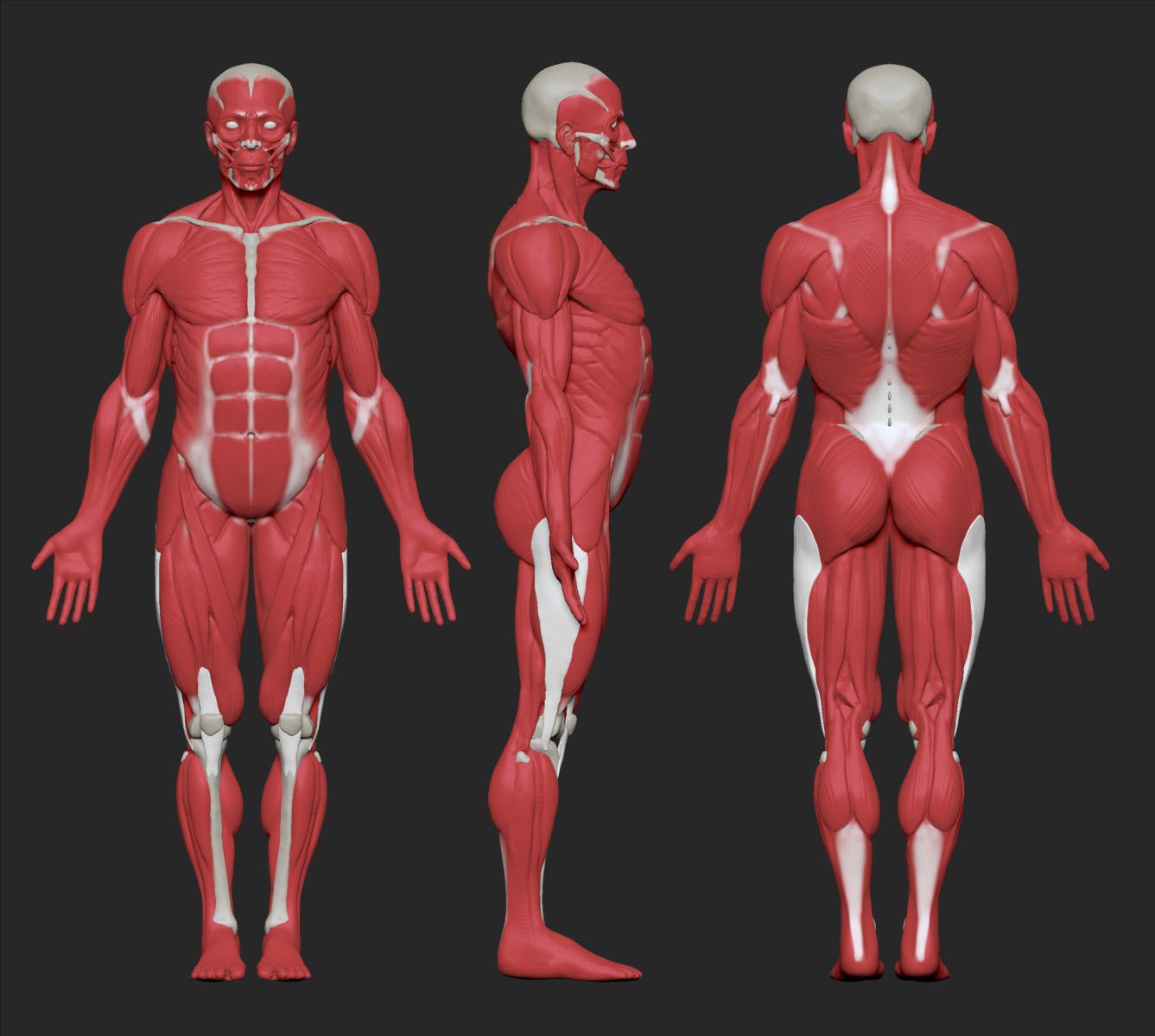 В то время как ранние исследования сообщали о снижении активности гидроксилазы и содержании гидроксипролина после кратковременной иммобилизации, более поздние работы обнаружили повышенное содержание коллагена I после 21 дня разгрузки задних конечностей у крыс, но никаких изменений после 60 дней постельного режима у людей.Необходимы дальнейшие исследования и, в частности, исследования в области обучения людей, чтобы изучить влияние различных методов обучения на структуру и состав ВКМ.
В то время как ранние исследования сообщали о снижении активности гидроксилазы и содержании гидроксипролина после кратковременной иммобилизации, более поздние работы обнаружили повышенное содержание коллагена I после 21 дня разгрузки задних конечностей у крыс, но никаких изменений после 60 дней постельного режима у людей.Необходимы дальнейшие исследования и, в частности, исследования в области обучения людей, чтобы изучить влияние различных методов обучения на структуру и состав ВКМ.
Вклад авторов
RC внес свой вклад в исследование литературы и подготовил рукопись. MG и BW внесли свой вклад в исследование литературы и отредактировали рукопись. Все авторы одобрили окончательную версию рукописи и согласились нести ответственность за все аспекты работы. Все лица, обозначенные как авторы, имеют право на авторство, и все те, кто имеет право на авторство, перечислены.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Австрийский научный фонд (FWF) за финансовую поддержку этого исследования: KLI 738-B27.
Каталожные номера
Аагард, П., Андерсен, Дж. Л., Дюре-Поульсен, П., Лефферс, А.-М., Вагнер, А., Magnusson, S.P., et al. (2001). Механизм увеличения сократительной способности перистых мышц человека в ответ на силовые тренировки: изменения в строении мышц. J. Physiol. 534, 613–623. doi: 10.1111/j.1469-7793.2001.t01-1-00613.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Адамс Г.Р., Хазер Б.М., Болдуин К.М. и Дадли Г.А. (1993). Состав тяжелой цепи миозина скелетных мышц и тренировка с отягощениями. J. Appl. Физиол. 74, 911–915. doi: 10.1152/jappl.1993.74.2.911
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Adstrum, S., Hedley, G., Schleip, R., Stecco, C., and Yucesoy, C.A. (2017). Определение фасциальной системы. Дж. Бодью. Мов.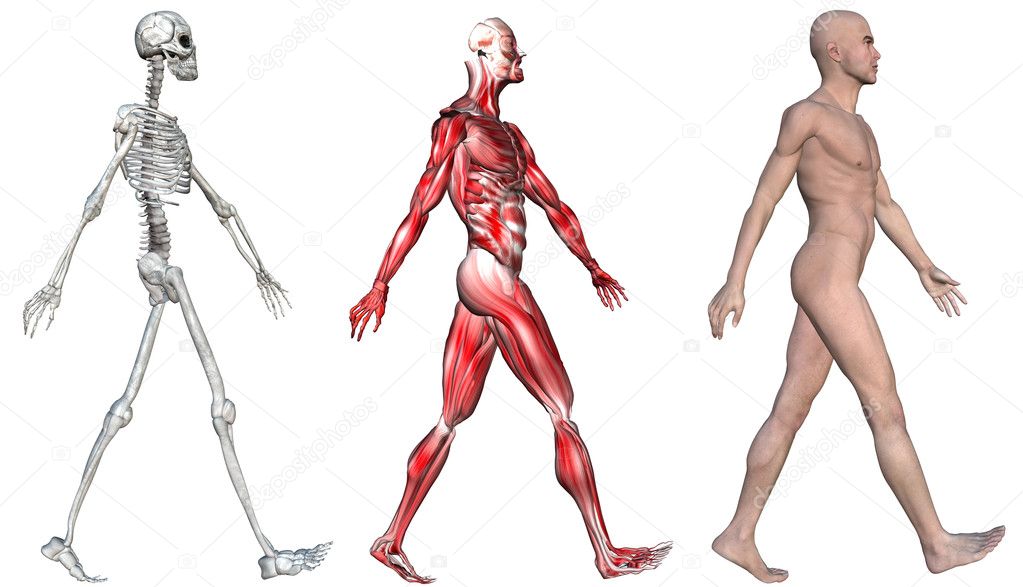 тер. 21, 173–177. doi: 10.1016/j.jbmt.2016.11.003
тер. 21, 173–177. doi: 10.1016/j.jbmt.2016.11.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Алмада, А. Э., и Вейджерс, А. Дж. (2016). Молекулярная схема судьбы стволовых клеток при регенерации, старении и заболеваниях скелетных мышц. Нац. Преподобный Мол. Клеточная биол. 17, 267–279. doi: 10.1038/nrm.2016.7
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Американский колледж спортивной медицины. (2009). Позиционный стенд Американского колледжа спортивной медицины. Модели прогрессии в тренировках с отягощениями для здоровых взрослых. Мед. науч. Спортивное упражнение. 41, 687–708. doi: 10.1249/mss.0b013e3181
0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Азизи Э., Делорье А.Р., Холт, Северная Каролина, и Итон, CE (2017). Сопротивление радиальному расширению ограничивает мышечное напряжение и работу. Биомех. Модель. механобиол.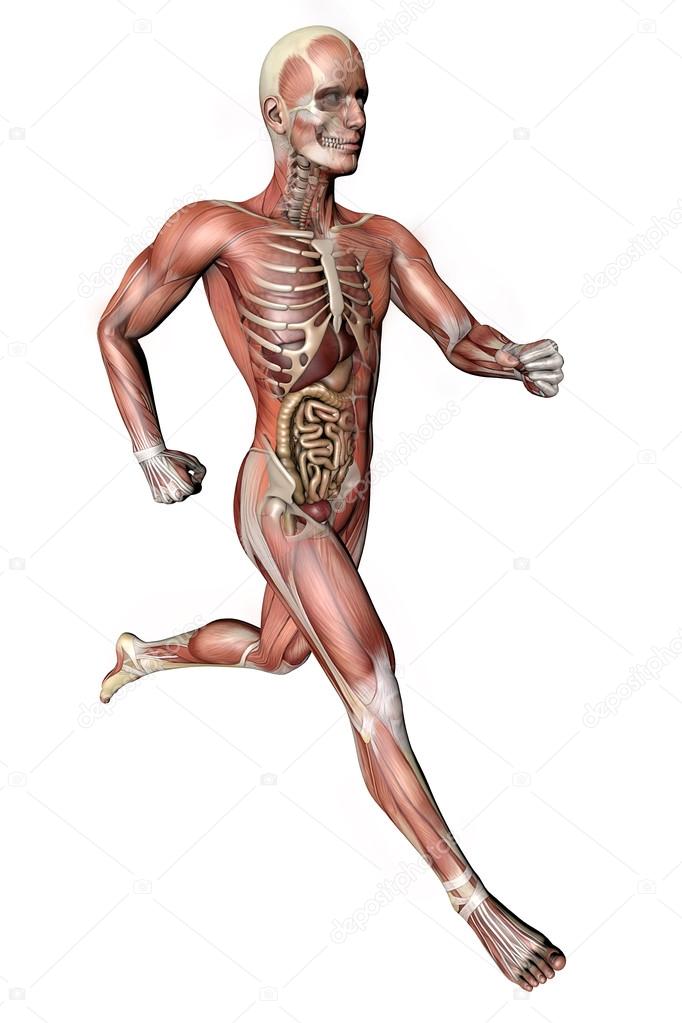 16, 1633–1643. doi: 10.1007/s10237-017-0909-3
16, 1633–1643. doi: 10.1007/s10237-017-0909-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Багдади М.Б., Кастель Д., Мачадо Л., Фукада С., Бирк Д.Е., Реле Ф. и др. (2018). Реципрокная передача сигналов с помощью Notch-Collagen V-CALCR удерживает мышечные стволовые клетки в их нише. Природа 557, 714–718. дои: 10.1038/с41586-018-0144-9
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Барретт, Б. (1962). Длина и способ окончания отдельных мышечных волокон в портняжной и задней бедренной мышцах человека. Акта Анат. 48, 242–257. дои: 10.1159/000141843
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bentzinger, C.F., Wang, Y.X., Dumont, N.A., and Rudnicki, M.A. (2013a). Клеточная динамика в нише сателлитных клеток мышц. EMBO Реп. 14, 1062–1072. doi: 10.1038/embor.2013.182
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bentzinger, C. F., Wang, Y.X., von Maltzahn, J., Soleimani, V.D., Yin, H., and Rudnicki, M.A. (2013b). Фибронектин регулирует передачу сигналов Wnt7a и расширение сателлитных клеток. Стволовая клетка 12, 75–87. doi: 10.1016/j.stem.2012.09.015
F., Wang, Y.X., von Maltzahn, J., Soleimani, V.D., Yin, H., and Rudnicki, M.A. (2013b). Фибронектин регулирует передачу сигналов Wnt7a и расширение сателлитных клеток. Стволовая клетка 12, 75–87. doi: 10.1016/j.stem.2012.09.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Берриа Р., Ван Л., Richardson, D.K., Finlayson, J., Belfort, R., Pratipanawatr, T., et al. (2006). Повышенное содержание коллагена в резистентных к инсулину скелетных мышцах. утра. Дж. Физиол. Эндокринол. Метаб. 290, Е560–Е565. doi: 10.1152/ajpendo.00202.2005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Блох, Р. Дж., и Гонсалес-Серратос, Х. (2003). Боковая передача силы через костамер в скелетных мышцах. Упр. Спортивная наука. Ред. 31, 73–78. дои: 10.1097/00003677-200304000-00004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боонен, К.Дж. М., Розария-Чак, К. Ю. , Баайенс, Ф. П. Т., ван дер Шафт, Д. В. Дж., и Пост, М. Дж. (2009). Основные экологические сигналы из ниши сателлитных клеток: оптимизация пролиферации и дифференцировки. утра. Дж. Физиол. Клеточная физиол. 296, C1338–C1345. doi: 10.1152/ajpcell.00015.2009
, Баайенс, Ф. П. Т., ван дер Шафт, Д. В. Дж., и Пост, М. Дж. (2009). Основные экологические сигналы из ниши сателлитных клеток: оптимизация пролиферации и дифференцировки. утра. Дж. Физиол. Клеточная физиол. 296, C1338–C1345. doi: 10.1152/ajpcell.00015.2009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боппарт, доктор медицины, и Махмассани, З.С. (2019). Передача сигналов интегрином: связь механической стимуляции с гипертрофией скелетных мышц. утра. Дж. Физиол. Клеточная физиол. 317, C629–C641. doi: 10.1152/ajpcell.00009.2019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Брак А.С., Конбой И.М., Конбой М.Дж., Шен Дж. и Рандо Т.А. (2008). Временное переключение с Notch на передачу сигналов Wnt в мышечных стволовых клетках необходимо для нормального миогенеза взрослых. Стволовая клетка 2, 50–59. doi: 10.1016/j.stem.2007.10.006
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Брак, А. С., Конбой М.Дж., Рой С., Ли М., Куо С.Дж., Келлер С. и соавт. (2007). Увеличение передачи сигналов Wnt во время старения изменяет судьбу мышечных стволовых клеток и увеличивает фиброз. Наука 317, 807–810. doi: 10.1126/наука.1144090
С., Конбой М.Дж., Рой С., Ли М., Куо С.Дж., Келлер С. и соавт. (2007). Увеличение передачи сигналов Wnt во время старения изменяет судьбу мышечных стволовых клеток и увеличивает фиброз. Наука 317, 807–810. doi: 10.1126/наука.1144090
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Брандан, Э., Кабельо-Верруджо, К., и Виал, К. (2008). Новые механизмы регуляции протеогликанов декорина и бигликана при формировании мышц и мышечной дистрофии. Матрикс Биол. 27, 700–708. doi: 10.1016/j.matbio.2008.07.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Brandt, P.W., Lopez, E., Reuben, J.P., and Grundfest, H. (1967). Взаимосвязь между плотностью упаковки миофиламентов и длиной саркомера в поперечнополосатых мышцах лягушки. J. Cell Biol. 33, 255–263. doi: 10.1083/jcb.33.2.255
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Браун С., Дэй С. и Доннелли А. (1999).Косвенные доказательства повреждения скелетных мышц человека и разрушения коллагена после эксцентрических мышечных движений. J. Sports Sci. 17, 397–402. дои: 10.1080/026404199365911
(1999).Косвенные доказательства повреждения скелетных мышц человека и разрушения коллагена после эксцентрических мышечных движений. J. Sports Sci. 17, 397–402. дои: 10.1080/026404199365911
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бушби, К.М.Д., Коллинз, Дж., и Хикс, Д. (2014). «Коллагеновые миопатии типа VI», в Progress in Heritable Soft Connective Diseases , ed. Дж. Халпер (Дордрехт: Springer), 185–199. дои: 10.1007/978-94-007-7893-1_12
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кальве, С., Одельберг, С.Дж., и Саймон, Х.-Г. (2010). Переходный внеклеточный матрикс определяет поведение клеток во время регенерации мышц. Дев. биол. 344, 259–271. doi: 10.1016/j.ydbio.2010.05.007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Calvo, A.C., Moreno, L., Moreno, L., Toivonen, J.M., Manzano, R., Molina, N., et al. (2020). Коллаген типа XIX: многообещающий биомаркер базальных мембран. Нейронная регенерация. Рез. 15, 988–995. дои: 10.4103/1673-5374.270299
Нейронная регенерация. Рез. 15, 988–995. дои: 10.4103/1673-5374.270299
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кармели Э., Моас М., Леннон С. и Пауэрс С.К. (2005). Упражнения высокой интенсивности увеличивают экспрессию матриксных металлопротеиназ в быстрых скелетных мышечных волокнах: упражнения и ММР в быстрых скелетных волокнах. Экспл. Физиол. 90, 613–619. doi: 10.1113/expphysiol.2004.029462
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шарве, Б., Гиро, А., Malbouyres, M., Zwolanek, D., Guillon, E., Bretaud, S., et al. (2013). Нокдаун гена col22a1 у рыбок данио вызывает мышечную дистрофию за счет нарушения мышечно-сухожильного соединения. Дев. Камб. англ. 140, 4602–4613. doi: 10.1242/dev.096024
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шике, М., Бирк, Д.Е., Беннеманн, К.Г., и Кох, М. (2014). Коллаген XII: защита целостности костей и мышц за счет организации коллагеновых фибрилл.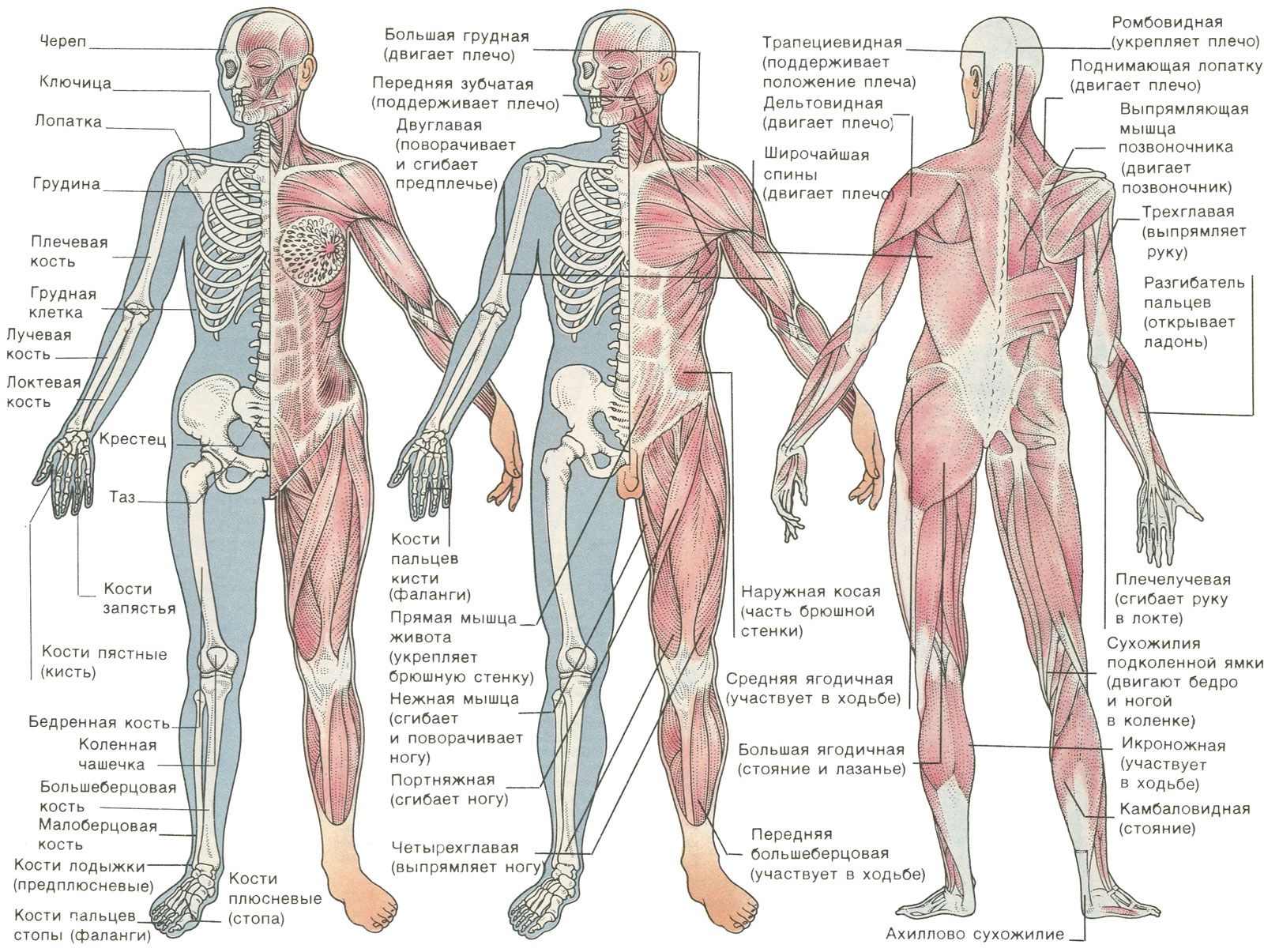 Междунар. Дж.Биохим. Клеточная биол. 53, 51–54. doi: 10.1016/j.biocel.2014.04.020
Междунар. Дж.Биохим. Клеточная биол. 53, 51–54. doi: 10.1016/j.biocel.2014.04.020
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кристенсен, С., и Пурслоу, П. П. (2016). Роль матриксных металлопротеиназ в развитии мышечной и жировой ткани и качестве мяса: обзор. Наука о мясе. 119, 138–146. doi: 10.1016/j.meatsci.2016.04.025
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Коркоран, М. Л., Хьюитт, Р. Э., Кляйнер, Д.E. Jr. и Stetler-Stevenson, WG (1996). ММП-2: экспрессия, активация и ингибирование. Ферментный белок 49, 7–19. дои: 10.1159/000468613
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Корнелисон, Д. Д., Филла, М. С., Стэнли, Х. М., Рапрегер, А. С., и Олвин, Б. Б. (2001). Синдекан-3 и синдекан-4 специфически маркируют сателлитные клетки скелетных мышц и участвуют в поддержании сателлитных клеток и регенерации мышц. Дев. биол.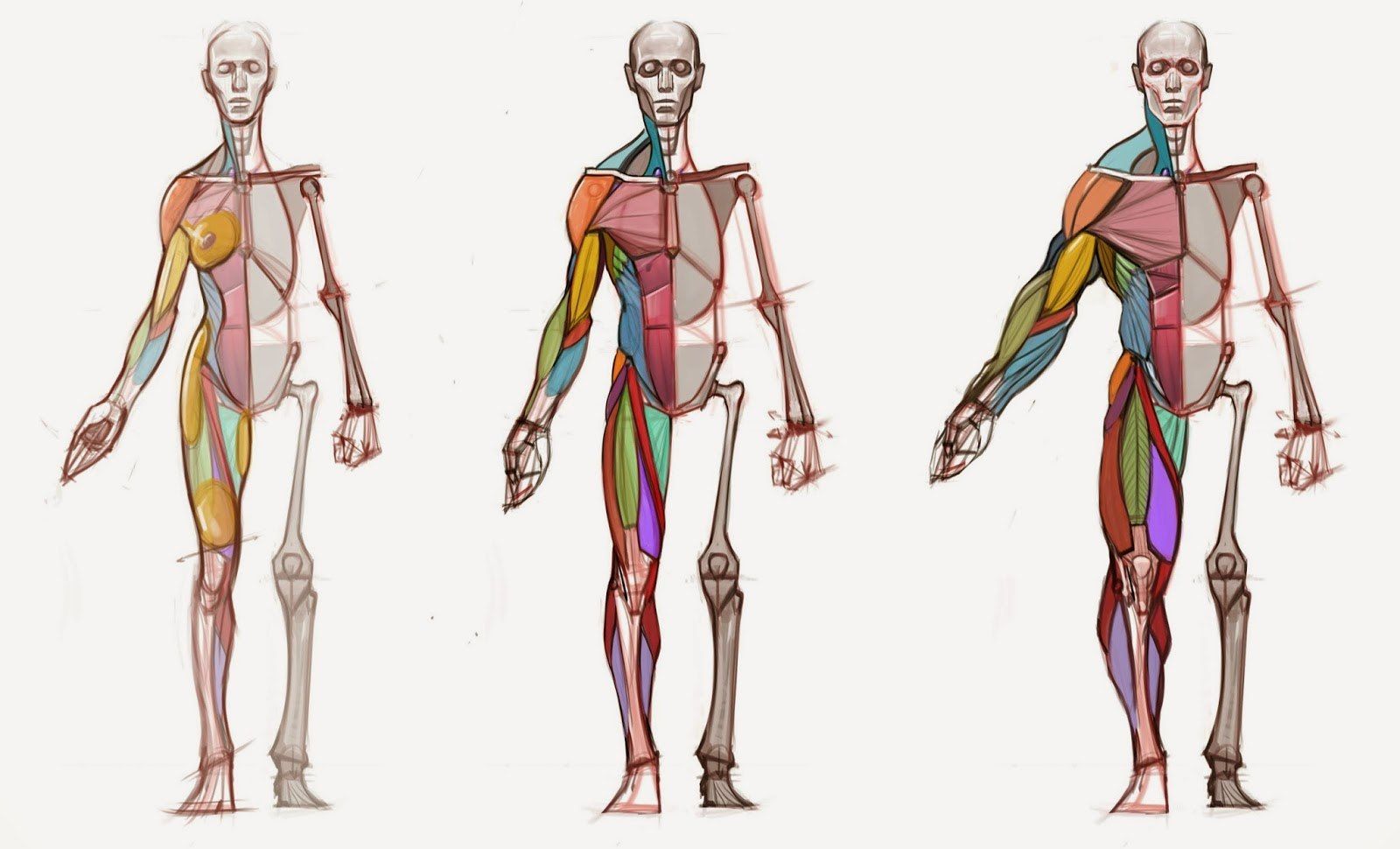 239, 79–94.doi: 10.1006/dbio.2001.0416
239, 79–94.doi: 10.1006/dbio.2001.0416
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cosgrove, B.D., Gilbert, P.M., Porpiglia, E., Mourkioti, F., Lee, S.P., Corbel, S.Y., et al. (2014). Омоложение популяции мышечных стволовых клеток восстанавливает силу поврежденных состарившихся мышц. Нац. Мед. 20, 255–264. doi: 10.1038/nm.3464
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Крамери, Р. М., Аагард, П., Квортруп, К., Лангберг, Х., Олесен, Дж., и Кьяер, М. (2007). Повреждение миофибрилл в скелетных мышцах человека: эффекты электрической стимуляции против произвольных сокращений: равномерная против гетерогенная активация мышечных волокон. J. Physiol. 583, 365–380. doi: 10.1113/jphysiol.2007.128827
Полнотекстовая перекрестная ссылка | Академия Google
Crameri, R.M., Langberg, H., Teisner, B., Magnusson, P., Schrøder, H.D., Olesen, J.L., et al. (2004). Усиленный процессинг проколлагена в скелетных мышцах после однократной эксцентрической нагрузки у людей. Матрикс Биол. 23, 259–264. doi: 10.1016/j.matbio.2004.05.009
(2004). Усиленный процессинг проколлагена в скелетных мышцах после однократной эксцентрической нагрузки у людей. Матрикс Биол. 23, 259–264. doi: 10.1016/j.matbio.2004.05.009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Цуй Н., Ху М. и Халил Р. А. (2017). Биохимические и биологические свойства матриксных металлопротеиназ. Прог. Мол. биол. Перевод науч. 147, 1–73. doi: 10.1016/bs.pmbts.2017.02.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
де Соуза Нето, И.В., Дуриган, Дж.Л.К., Гуццони, В., Тибана, Р.A., Prestes, J., de Araujo, H.S.S., et al. (2018). Влияние силовых тренировок на активность матричных металлопротеиназ в скелетных мышцах и кровообращение при старении. Перед. Физиол. 9:190. doi: 10.3389/fphys.2018.00190
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дитрих, А.В., Боттер, А., Виейра, Т.М., Пеолссон, А., Петцке, Ф., Дэйви, П. , и соавт. (2017). Пространственные вариации и несоответствие между оценками начала мышечной активации по данным ЭМГ и УЗИ. науч. Респ. 7:42011. дои: 10.1038/srep42011
, и соавт. (2017). Пространственные вариации и несоответствие между оценками начала мышечной активации по данным ЭМГ и УЗИ. науч. Респ. 7:42011. дои: 10.1038/srep42011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дитрих С., Абу-Ребье Ф., Броманн Х., Бладт Ф., Зонненберг-Ритмахер Э., Ямаай Т. и др. (1999). Роль SF/HGF и c-Met в развитии скелетных мышц. Дев. Камб. англ. 126, 1621–1629.
Реферат PubMed | Академия Google
Duance, V.C., Restall, D.J., Beard, H., Bourne, F.J., and Bailey, A.Дж. (1977). Расположение трех типов коллагена в скелетных мышцах. ФЭБС Письмо. 79, 248–252. дои: 10.1016/0014-5793(77)80797-7
Полнотекстовая перекрестная ссылка | Академия Google
Эклунд, Л., Пиухола, Дж., Комулайнен, Дж., Сормунен, Р., Онгваррасопоне, К., Фасслер, Р., и соавт. (2001). Отсутствие коллагена типа XV вызывает скелетную миопатию и сердечно-сосудистые дефекты у мышей. Проц. Натл. акад. науч. США 98, 1194–1199. doi: 10.1073/pnas.031444798
науч. США 98, 1194–1199. doi: 10.1073/pnas.031444798
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Энглер, А.Дж., Гриффин, М.А., Сен, С., Беннеманн, К.Г., Суини, Х.Л., и Дишер, Д.Е. (2004). Myotubes оптимально дифференцируются на субстратах с тканеподобной жесткостью: патологические последствия для мягких или жестких микроокружений. J. Cell Biol. 166, 877–887. doi: 10.1083/jcb.200405004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Этьен, Дж., Лю, К., Скиннер, К.М., Конбой, М.Дж., и Конбой, И.М. (2020). Скелетная мышца как экспериментальная модель выбора для изучения старения и омоложения тканей. Скелет. Мышцы 10:4. doi: 10.1186/s13395-020-0222-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фишер, Дж., Стил, Дж., Брюс-Лоу, С., и Смит, Д. (2011). Рекомендации по тренировкам с отягощениями, основанные на фактических данных. Мед. Спорт. 15, 147–162. doi: 10.2478/v10036-011-0025-x
15, 147–162. doi: 10.2478/v10036-011-0025-x
Полнотекстовая перекрестная ссылка | Академия Google
Фолланд, Дж. П., и Уильямс, А. Г. (2007). Адаптация к силовым тренировкам: морфологический и неврологический вклад в увеличение силы. Спорт Мед. 37, 145–168. дои: 10.2165/00007256-200737020-00004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фрай, К.С., Кирби, Т.Дж., Космак, К., Маккарти, Дж.Дж., и Петерсон, К.А. (2017). Миогенные клетки-предшественники контролируют продукцию внеклеточного матрикса фибробластами во время гипертрофии скелетных мышц. Стволовая клетка 20, 56–69. doi: 10.1016/j.stem.2016.09.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гарбер, К.E., Blissmer, B., Deschenes, M.R., Franklin, B.A., Lamonte, M.J., Lee, I.-M., et al. (2011). Количество и качество упражнений для развития и поддержания кардиореспираторной, скелетно-мышечной и нейромоторной выносливости у практически здоровых взрослых: руководство по назначению упражнений.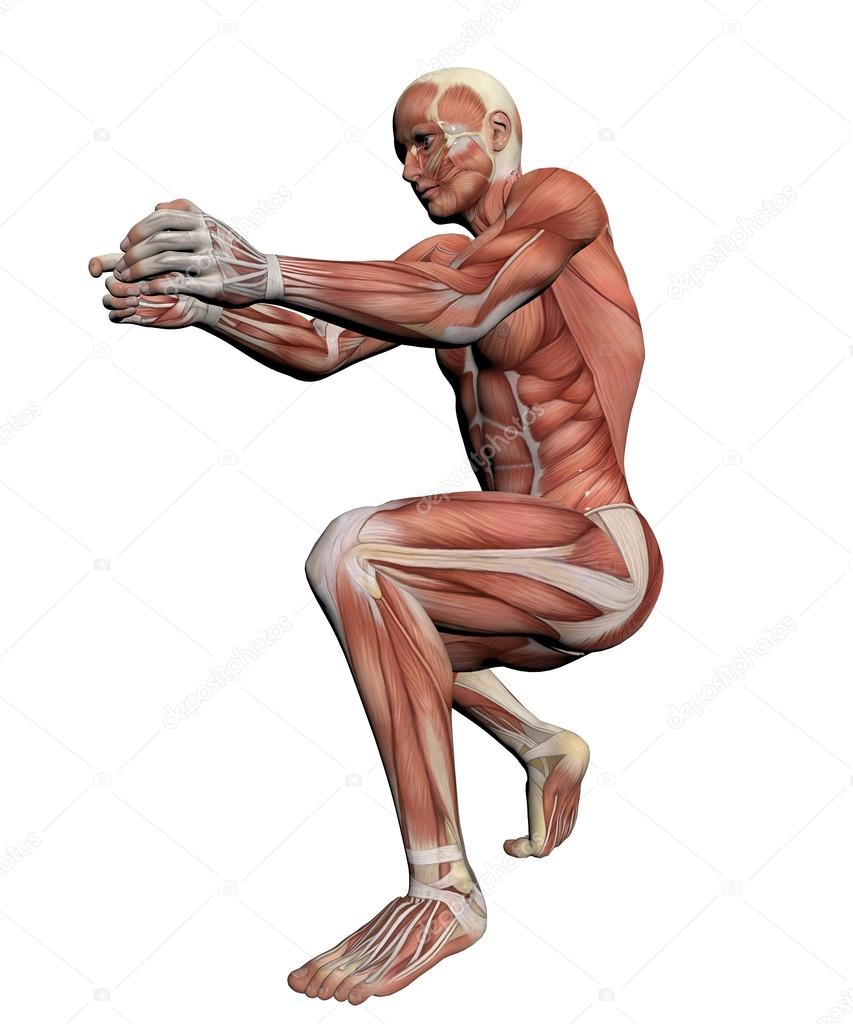 Мед. науч. Спортивное упражнение. 43, 1334–1359. doi: 10.1249/MSS.0b013e318213fefb
Мед. науч. Спортивное упражнение. 43, 1334–1359. doi: 10.1249/MSS.0b013e318213fefb
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гилберт, П. М., Хавенстрит, К. Л., Магнуссон, К.Э.Г., Сакко А., Леонарди Н.А., Крафт П. и соавт. (2010). Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре. Наука 329, 1078–1081. doi: 10.1126/science.11
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гиллис, А. Р., Чепмен, М. А., Бушонг, Э. А., Диринк, Т. Дж., Эллисман, М. Х., и Либер, Р. Л. (2017). Трехмерная реконструкция с высоким разрешением внеклеточного матрикса фиброзной скелетной мышцы: организация ECM фиброзной мышцы. J. Physiol. 595, 1159–1171. дои: 10.1113/JP273376
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Госселин, Л. Е., Адамс, К., Коттер, Т. А., Маккормик, Р. Дж., и Томас, Д. П. (1998). Влияние физических упражнений на пассивную жесткость локомоторных скелетных мышц: роль внеклеточного матрикса.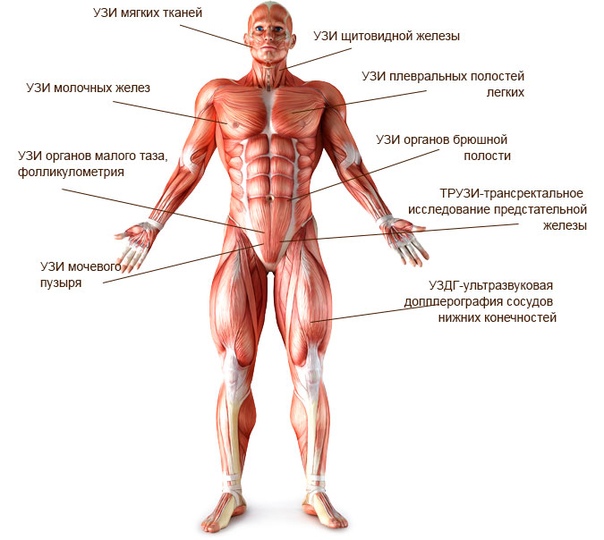 J. Appl. Физиол. 85, 1011–1016. doi: 10.1152/jappl.1998.85.3.1011
J. Appl. Физиол. 85, 1011–1016. doi: 10.1152/jappl.1998.85.3.1011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грефте, С., Vullinghs, S., Kuijpers-Jagtman, A.M., Torensma, R., and Von den Hoff, JW (2012). Матригель, но не коллаген I, поддерживает способность к дифференцировке мышечных клеток 90–196 in vitro 90–197 . Биомед. Матер. 7:055004. дои: 10.1088/1748-6041/7/5/055004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Grounds, MD (1998). Возрастные изменения реакции клеток скелетных мышц на нагрузку и регенерацию. Энн. Н. Я. акад.науч. 854, 78–91. doi: 10.1111/j.1749-6632.1998.tb09894.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Граундс, М. Д., Сорокин, Л., и Уайт, Дж. (2005). Сила на границе межклеточного матрикса и мышц. Скан. Дж. Мед. науч. Спорт 15, 381–391. doi: 10.1111/j.1600-0838.2005.00467.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гийон, Э.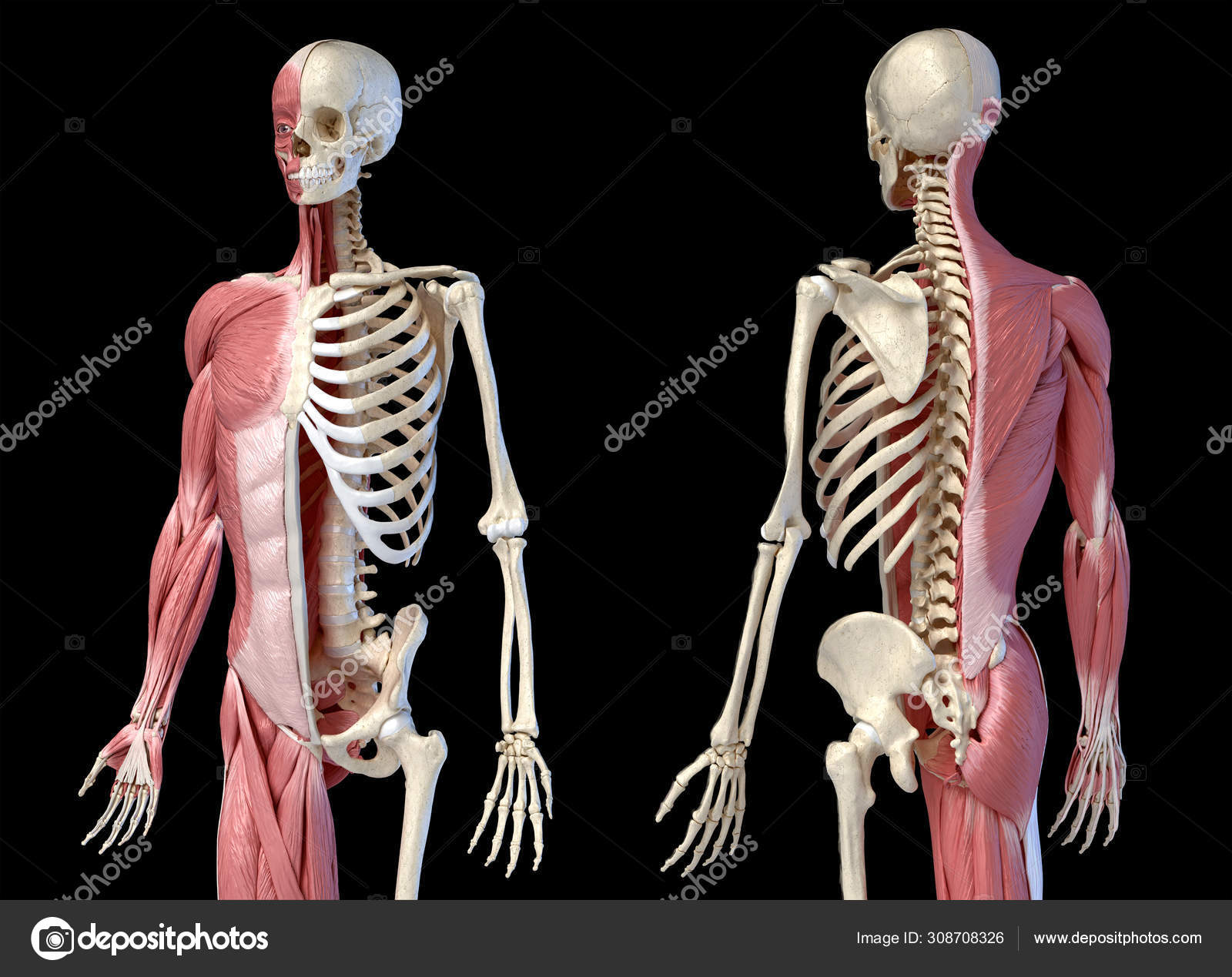 , Брето, С., и Руджеро, Ф. (2016). Предшественники медленных мышц откладывают отпечаток матрицы коллагена XV, чтобы направлять моторные аксоны. J. Neurosci. 36, 2663–2676. doi: 10.1523/JNEUROSCI.2847-15.2016
, Брето, С., и Руджеро, Ф. (2016). Предшественники медленных мышц откладывают отпечаток матрицы коллагена XV, чтобы направлять моторные аксоны. J. Neurosci. 36, 2663–2676. doi: 10.1523/JNEUROSCI.2847-15.2016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гутьеррес, Дж., и Брэндан, Э. (2010). Новый механизм секвестрации фактора роста фибробластов 2 глипиканом в липидных рафтах, обеспечивающий дифференцировку скелетных мышц. Мол. Клетка. биол. 30, 1634–1649. doi: 10.1128/mcb.01164-09
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гуццони, В., Рибейро, М.Б.Т., Лопес, Г.Н., де Кассиа Маркети, Р., де Андраде, Р.В., Селистре-де-Араужо, Х.С., и др. (2018). Влияние силовых тренировок на адаптацию внеклеточного матрикса скелетных мышц старых крыс. Перед. Физиол. 9:374. doi: 10.3389/fphys.2018.00374
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Халпер, Дж.![]() , и Кьяер, М. (2014). «Основные компоненты соединительной ткани и внеклеточного матрикса: эластин, фибриллин, фибулины, фибриноген, фибронектин, ламинин, тенасцины и тромбоспондины», в Progress in Heritable Soft Connective Tissue Diseases , ed.Дж. Халпер (Дордрехт: Springer), 31–47. дои: 10.1007/978-94-007-7893-1_3
, и Кьяер, М. (2014). «Основные компоненты соединительной ткани и внеклеточного матрикса: эластин, фибриллин, фибулины, фибриноген, фибронектин, ламинин, тенасцины и тромбоспондины», в Progress in Heritable Soft Connective Tissue Diseases , ed.Дж. Халпер (Дордрехт: Springer), 31–47. дои: 10.1007/978-94-007-7893-1_3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Харёнен Х., Зайнул З., Ту Х., Науменко Н., Сормунен Р., Мииналайнен И. и др. (2017). Коллаген XIII обеспечивает пре- и постсинаптическую целостность нервно-мышечного синапса. Гул. Мол. Жене. 26, 2076–2090. doi: 10.1093/hmg/ddx101
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хассон, П., ДеЛорье, А., Bennett, M., Grigorieva, E., Naiche, L.A., Papaioannou, V.E., et al. (2010). Tbx4 и tbx5, действующие в соединительной ткани, необходимы для формирования паттерна мышц и сухожилий конечностей. Дев. Ячейка 18, 148–156. doi: 10.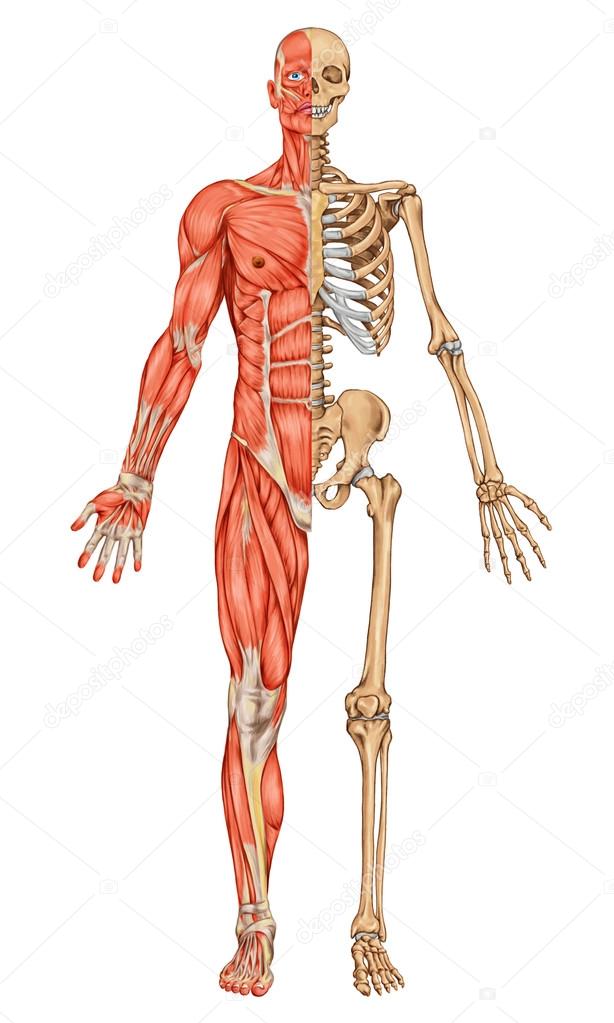 1016/j.devcel.2009.11.013
1016/j.devcel.2009.11.013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хаус, Дж. М., Кэрритерс, Дж. А., Кэрролл, К. С., Теш, П. А., и Траппе, Т. А. (2007a). Содержание белка в сократительной и соединительной ткани скелетных мышц человека: эффекты 35 и 90 дней имитации микрогравитации и противодействия физическим упражнениям. утра. Дж. Физиол. Регул. интегр. Комп. Физиол. 293, R1722–R1727. doi: 10.1152/ajpregu.00292.2007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хаус, Дж. М., Кэрритерс, Дж. А., Трапп, С. В., и Трапп, Т. А. (2007b). Коллаген, перекрестное связывание и конечные продукты гликирования в стареющих скелетных мышцах человека. J. Appl. Физиол. 103, 2068–2076. doi: 10.1152/japplphysiol.00670.2007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хейккинен, А., Харёнен, Х., Норман, О., и Пихлаяниеми, Т. (2019). Коллаген XIII и другие компоненты ECM в сборке и заболевании нервно-мышечного синапса. Анат. Рек. doi: 10.1002/ar.24092 [Epub перед печатью].
Анат. Рек. doi: 10.1002/ar.24092 [Epub перед печатью].
Полнотекстовая перекрестная ссылка | PubMed Резюме | Академия Google
Heinemeier, K.M., Olesen, J.L., Haddad, F., Langberg, H., Kjaer, M., Baldwin, K.M., et al. (2007). Экспрессия коллагена и родственных факторов роста в сухожилиях и скелетных мышцах крыс в ответ на определенные типы сокращения: экспрессия коллагена и TGF-β-1 в сухожилиях и мышцах, подвергающихся нагрузке. J. Physiol. 582, 1303–1316. doi: 10.1113/jphysiol.2007.127639
Полнотекстовая перекрестная ссылка | Академия Google
Хельясваара Р., Айкио М., Руотсалайнен Х. и Пихлаяниеми Т. (2017). Коллаген XVIII в тканевом гомеостазе и дисрегуляции — уроки, извлеченные из модельных организмов и пациентов-людей. Матрикс Биол. 57–58, 55–75. doi: 10.1016/j.matbio.2016.10.002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хиджиката, Т., Вакисака Х. и Ниида С. (1993). Функциональная комбинация сужающихся профилей и перекрывающихся структур в нерастягивающихся скелетных мышечных волокнах, оканчивающихся внутри пучка.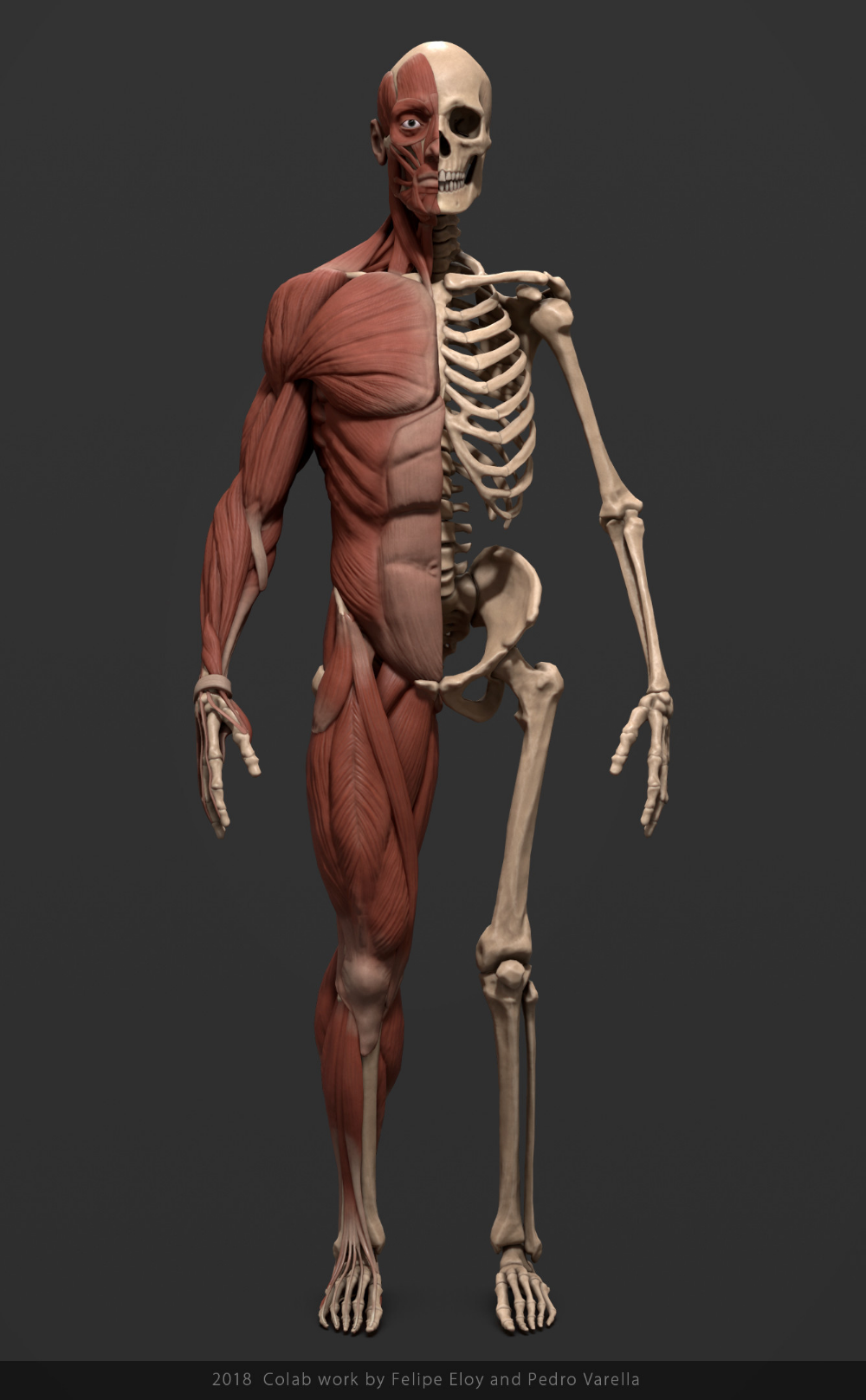 Анат. Рек. 236, 602–610. doi: 10.1002/ar.10403
Анат. Рек. 236, 602–610. doi: 10.1002/ar.10403
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хиндл, А.Г., Хорнинг, М., Меллиш, Дж.-А. Э. и Лоулер, Дж. М. (2009). Погружение в старость: мышечное старение у крупного долгоживущего млекопитающего, тюленя Уэдделла ( Leptonychotes weddellii ). Дж. Экспл. биол. 212, 790–796. doi: 10.1242/jeb.025387
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холм, Л., Рахбек, С.К., Фаруп, Дж., Вендельбо, М.Х., и Виссинг, К. (2017). Режим сокращения и прием сывороточного протеина влияют на скорость синтеза внутримышечной соединительной ткани: краткие отчеты. Мышечный нерв 55, 128–130. doi: 10.1002/mus.25398
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холм, Л., van Hall, G., Rose, A.J., Miller, B.F., Doessing, S., Richter, E.A., et al. (2010). Интенсивность сокращения и питание по-разному влияют на скорость синтеза коллагена и миофибриллярного белка в скелетных мышцах человека. утра. Дж. Физиол. Эндокринол. Метаб. 298, E257–E269. doi: 10.1152/ajpendo.00609.2009
утра. Дж. Физиол. Эндокринол. Метаб. 298, E257–E269. doi: 10.1152/ajpendo.00609.2009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хуанг Ю., Фан Ю., Саланова М., Ян X., Сунь Л. и Блоттнер Д. (2018). Влияние подошвенной вибрации на кость и глубокую фасцию в модели бездействия разгрузки задних конечностей крысы. Перед. Физиол. 9:616. doi: 10.3389/fphys.2018.00616
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Hughes, D.C., Marcotte, G.R., Marshall, A.G., West, D.W.D., Baehr, L.M., Wallace, M.A., et al. (2016). Возрастные различия дистрофина: влияние на белки передачи силы, целостность мембран и стабильность нервно-мышечных соединений. Дж. Геронтол. А. Биол. науч. Мед. науч. 72, 640–648. doi: 10.1093/gerona/glw109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хуйцзин, П.А. (1999). Мышца как композит, армированный коллагеновыми волокнами: обзор передачи силы в мышцах и конечностях в целом. Дж. Биомех. 32, 329–345. doi: 10.1016/s0021-9290(98)00186-9
Дж. Биомех. 32, 329–345. doi: 10.1016/s0021-9290(98)00186-9
Полнотекстовая перекрестная ссылка | Академия Google
Huijing, P.A., Baan, G.C., and Rebel, G.T. (1998). Немышечно-сухожильная передача силы в длинном разгибателе пальцев крысы. Дж. Экспл. биол. 201, 683–691.
Академия Google
Хилдаль, Р. Д., Нельсон, Б., Xin, L., Welling, T., Groscost, L., Hubal, M.J., et al. (2015). Ремоделирование внеклеточного матрикса и его вклад в защитную адаптацию после удлинения мышц человека. FASEB J. 29, 2894–2904. doi: 10.1096/fj.14-266668
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ивата Дж., Судзуки А., Пеликан Р. К., Хо Т.-В. и Чай Ю. (2013). Передача сигналов неканонического трансформирующего фактора роста β (TGFβ) в клетках краниального нервного гребня вызывает дефекты развития мышц языка. Дж. Биол. хим. 288, 29760–29770. doi: 10.1074/jbc.M113.493551
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Якобсен, Дж.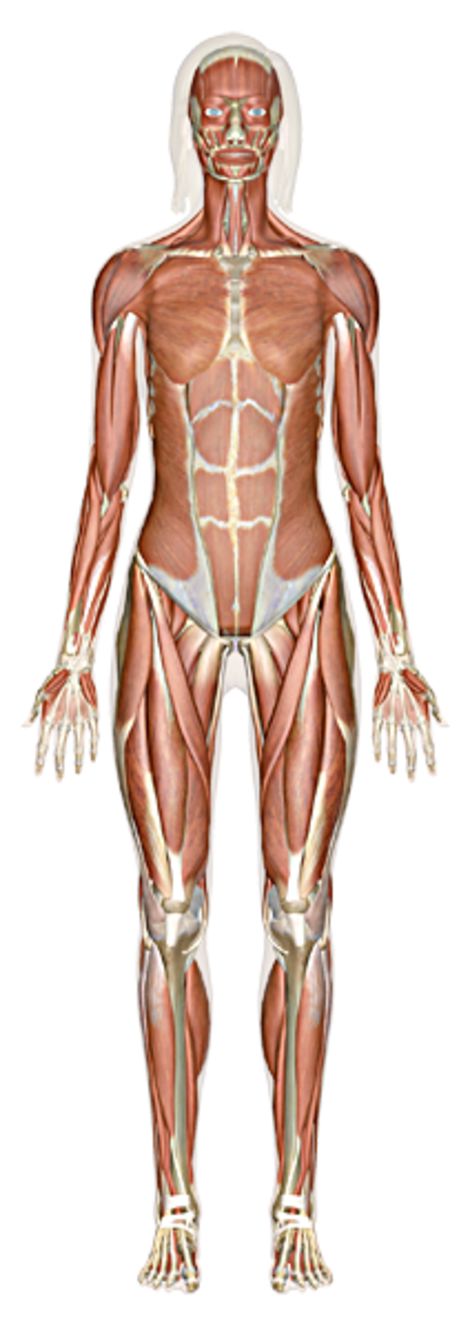 Р., Макки, А. Л., Кнудсен, А. Б., Кох, М., Кьер, М., и Крогсгаард, М. Р. (2017). Состав и адаптация мышечно-сухожильного соединения человека и соседних мышечных волокон к тренировкам с тяжелым сопротивлением. Скан. Дж. Мед. науч. Спорт 27, 1547–1559. doi: 10.1111/смс.12794
Р., Макки, А. Л., Кнудсен, А. Б., Кох, М., Кьер, М., и Крогсгаард, М. Р. (2017). Состав и адаптация мышечно-сухожильного соединения человека и соседних мышечных волокон к тренировкам с тяжелым сопротивлением. Скан. Дж. Мед. науч. Спорт 27, 1547–1559. doi: 10.1111/смс.12794
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Яначек, Й., Чебасек В., Кубинова Л., Рибарич С. и Эрцен И. (2009). Трехмерная визуализация и измерение капилляров, снабжающих метаболически разные типы волокон в длинном разгибателе пальцев крысы во время денервации и реиннервации. J. Histochem. Цитохим. 57, 437–447. doi: 10.1369/jhc.2008.953018
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ярвинен Т.А.Х., Йожа Л., Каннус П., Ярвинен Т.Л.Н. и Ярвинен М. (2002). Организация и распределение внутримышечной соединительной ткани в нормальных и иммобилизованных скелетных мышцах.Иммуногистохимическое, поляризационное и сканирующее электронное микроскопическое исследование. Дж. Мускул Рез. Селл Мотил. 23, 245–254. дои: 10.1023/a:10208336
Дж. Мускул Рез. Селл Мотил. 23, 245–254. дои: 10.1023/a:10208336
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Kang, L., Lantier, L., Kennedy, A., Bonner, J.S., Mayes, W.H., Bracy, D.P., et al. (2013). Гиалуронан накапливается при кормлении с высоким содержанием жиров и способствует резистентности к инсулину. Диабет 62, 1888–1896. дои: 10.2337/db12-1502
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кан, Л., Мэйс, У. Х., Джеймс, Ф. Д., Брейси, Д. П., и Вассерман, Д. Х. (2014). Матриксная металлопротеиназа 9 противостоит индуцированной диетой резистентности мышц к инсулину у мышей. Диабетология 57, 603–613. doi: 10.1007/s00125-013-3128-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кардон, Г., Харф, Б.Д., и Табин, С.Дж. (2003). Tcf4-позитивная мезодермальная популяция обеспечивает препаттерн для формирования паттерна мышц конечностей позвоночных. Дев. Ячейка 5, 937–944. дои: 10.1016/с1534-5807(03)00360-5
Дев. Ячейка 5, 937–944. дои: 10.1016/с1534-5807(03)00360-5
Полнотекстовая перекрестная ссылка | Академия Google
Карпакка Дж., Вяананен К., Орава С. и Такала Т. Е. (1990). Влияние предиммобилизационной тренировки и иммобилизации на синтез коллагена в скелетных мышцах крыс. Междунар. Дж. Спорт Мед. 11, 484–488. doi: 10.1055/s-2007-1024842
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Карпакка Дж., Виртанен П., Ваананен К., Орава С. и Такала Т. Е. (1991).Синтез коллагена в скелетных мышцах крыс при иммобилизации и ремобилизации. J. Appl. Физиол. 70, 1775–1780 гг. doi: 10.1152/jappl.1991.70.4.1775
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Khaleduzzaman, M., Sumiyoshi, H., Ueki, Y., Inoguchi, K., Ninomiya, Y. и Yoshioka, H. (1997). Структура гена коллагена типа XIX человека (COL19A1), которая предполагает, что он произошел от гена-предка семейства FACIT. Геномика 45, 304–312.doi: 10.1006/geno.1997.4921
Геномика 45, 304–312.doi: 10.1006/geno.1997.4921
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ким, Дж., Ван, З., Хеймсфилд, С.Б., Баумгартнер, Р.Н., и Галлахер, Д. (2002). Масса скелетных мышц всего тела: оценка новым методом двухэнергетической рентгеновской абсорбциометрии. утра. Дж. Клин. Нутр. 76, 378–383. doi: 10.1093/ajcn/76.2.378
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кнудсен, А. Б., Ларсен, М., Макки, А. Л., Хьорт, М., Hansen, K.K., Qvortrup, K., et al. (2015). Мышечно-сухожильное соединение человека: исследование ультраструктуры и трехмерного анализа. Скан. Дж. Мед. науч. Спорт 25, е116–е123. doi: 10.1111/смс.12221
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Koch, M., Schulze, J., Hansen, U., Ashwodt, T., Keene, D.R., Brunken, W.J., et al. (2004). Новый маркер тканевых соединений, коллаген XXII. Дж. Биол. хим. 279, 22514–22521. doi: 10.1074/jbc.M400536200
279, 22514–22521. doi: 10.1074/jbc.M400536200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кованен В., Суоминен Х., Ристели Дж. и Ристели Л. (1988). Коллаген IV типа и ламинин в медленных и быстрых скелетных мышцах крыс — влияние возраста и тренировок на выносливость в течение всей жизни. Сб. Относ. Рез. 8, 145–153. doi: 10.1016/s0174-173x(88)80026-8
Полнотекстовая перекрестная ссылка | Академия Google
Куо, Х.Дж., Маслен, К.Л., Кин, Д.Р., и Гланвилл, Р.В. (1997). Коллаген типа VI закрепляет базальные мембраны эндотелия, взаимодействуя с коллагеном типа IV. Дж. Биол. хим. 272, 26522–26529. doi: 10.1074/jbc.272.42.26522
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Леонг Д. П., Тео К. К., Рангараджан С., Лопес-Харамильо П., Авезум А., Орландини А. и др. (2015). Прогностическое значение силы хвата: результаты проспективного исследования городской и сельской эпидемиологии (PURE).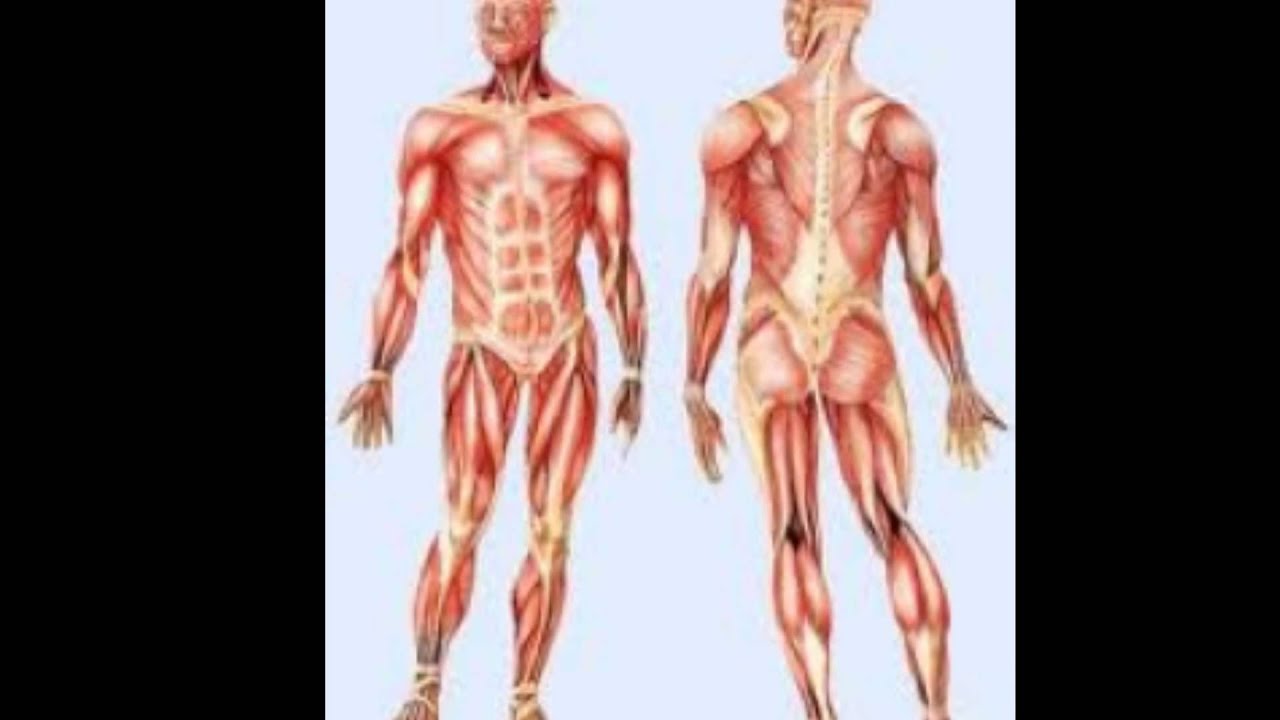 Ланцет Лонд. англ. 386, 266–273. doi: 10.1016/s0140-6736(14)62000-6
Ланцет Лонд. англ. 386, 266–273. doi: 10.1016/s0140-6736(14)62000-6
Полнотекстовая перекрестная ссылка | Академия Google
Ли, Дж.J., Wittert, G.A., Vincent, A., Atlantis, E., Shi, Z., Appleton, S.L., et al. (2016). Сила мышечного сжатия предсказывает возникновение диабета 2 типа: популяционное когортное исследование. Метаболизм 65, 883–892. doi: 10.1016/j.metabol.2016.03.011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Либер Р.Л. и Уорд С.Р. (2013). Клеточные механизмы фиброза тканей. 4. Структурно-функциональные последствия фиброза скелетных мышц. утра. Дж. Физиол.Клеточная физиол. 305, C241–C252. doi: 10.1152/ajpcell.00173.2013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лопес-Отин, К., Бласко, М.А., Партридж, Л., Серрано, М., и Кремер, Г. (2013). Признаки старения. Сотовый 153, 1194–1217. doi: 10.1016/j.cell.2013.05.039
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Maaß, T. , Bayley, C.P., Mörgelin, M., Lettmann, S., Bonaldo, P., Paulsson, M., et al. (2016).Неоднородность микрофибрилл коллагена VI: структурный анализ неколлагеновых областей. Дж. Биол. хим. 291, 5247–5258. doi: 10.1074/jbc.M115.705160
, Bayley, C.P., Mörgelin, M., Lettmann, S., Bonaldo, P., Paulsson, M., et al. (2016).Неоднородность микрофибрилл коллагена VI: структурный анализ неколлагеновых областей. Дж. Биол. хим. 291, 5247–5258. doi: 10.1074/jbc.M115.705160
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Mackey, A.L., Brandstetter, S., Schjerling, P., Bojsen-Moller, J., Qvortrup, K., Pedersen, M.M., et al. (2011). Последовательный ответ мертвогезии внеклеточного матрикса и регуляторов фиброза после повреждения мышц участвует в защите скелетных мышц человека от будущих повреждений. FASEB J. 25, 1943–1959. doi: 10.1096/fj.10-176487
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Mackey, A.L., Donnelly, A.E., Turpeenniemi-Hujanen, T., and Roper, H.P. (2004). Содержание коллагена в скелетных мышцах человека после высокоинтенсивных эксцентрических сокращений. J. Appl. Физиол. 97, 197–203. doi: 10.1152/japplphysiol.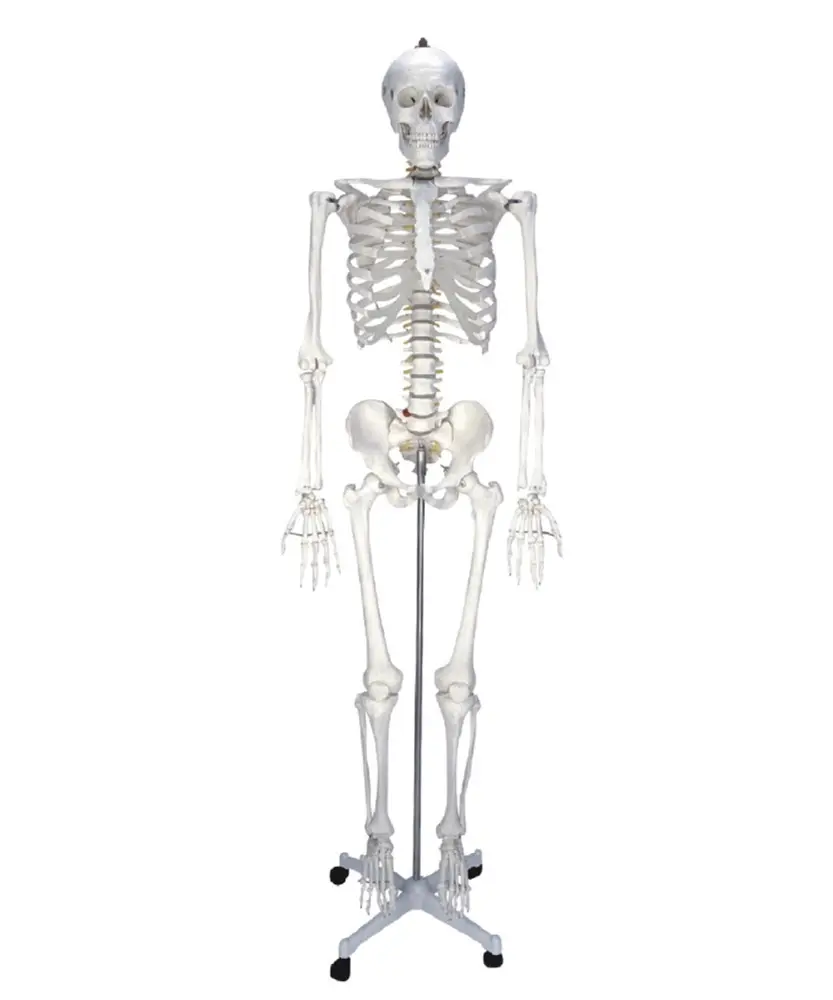 01174.2003
01174.2003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Махновский П.А., Згода В.Г., Боков Р. О., Шагимарданова Е. И., Газизова Г. Р., Гусев О. А. и др. (2020). Регуляция белков в скелетных мышцах человека: роль транскрипции. науч. Респ. 10:3514.
Академия Google
Мартин, Г. Р., и Тимпл, Р. (1987). Ламинин и другие компоненты базальной мембраны. год. Преподобный Cell Biol. 3, 57–85. doi: 10.1146/annurev.cb.03.110187.000421
Полнотекстовая перекрестная ссылка | Академия Google
Макки, Т.Дж., Перлман, Г., Моррис М. и Комарова С.В. (2019). Состав внеклеточного матрикса соединительной ткани: систематический обзор и метаанализ. науч. Респ. 9:10542.
Академия Google
Мендиас, К.Л., Шварц, А.Дж., Грекин, Дж.А., Гумуцио, Дж.П., и Сагг, К.Б. (2017). Изменения сократительной способности мышечных волокон и продукции внеклеточного матрикса при гипертрофии скелетных мышц.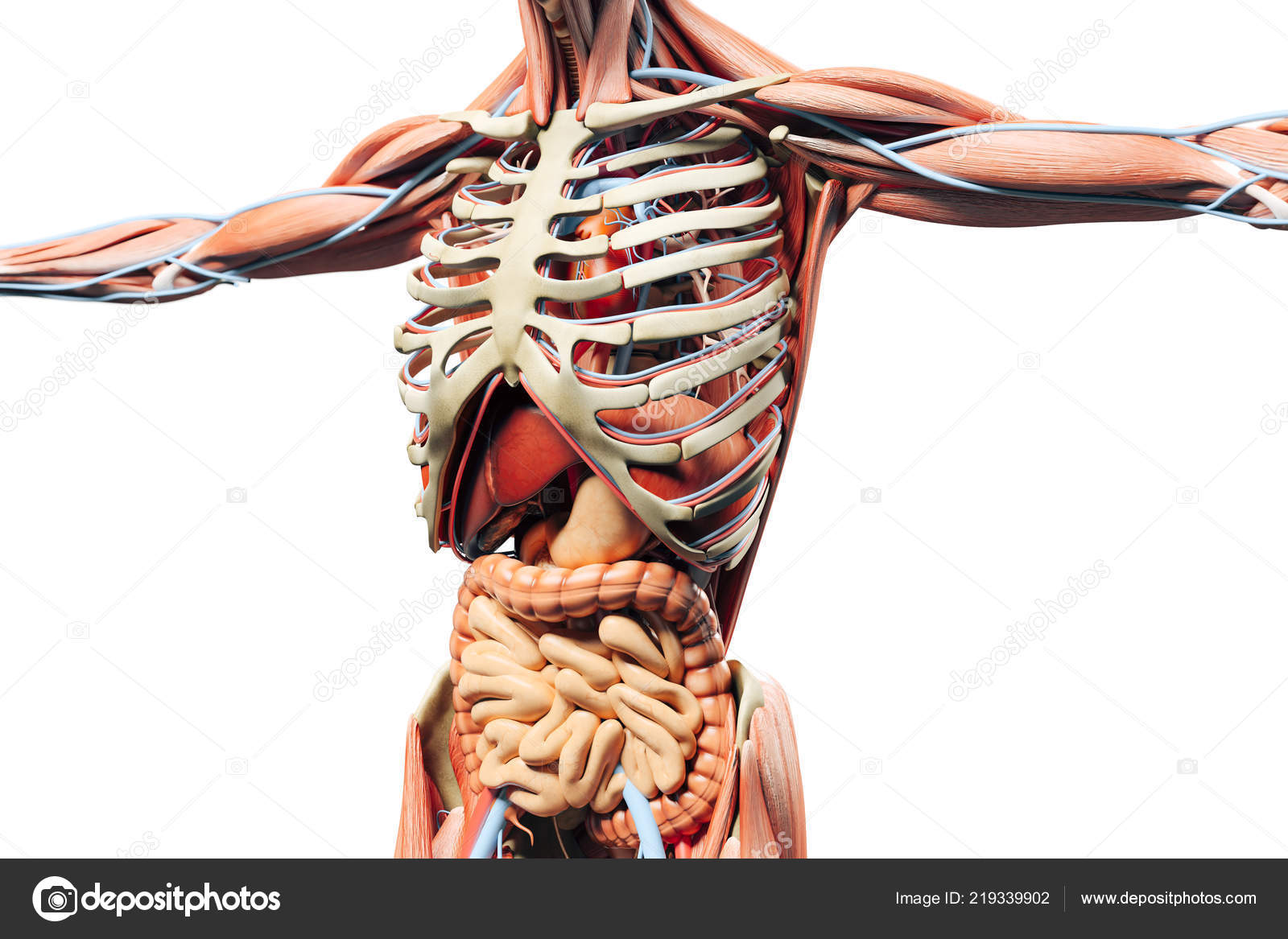 J. Appl. Физиол. 122, 571–579. doi: 10.1152/japplphysiol.00719.2016
J. Appl. Физиол. 122, 571–579. doi: 10.1152/japplphysiol.00719.2016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Монти, Р.Дж., Рой, Р.Р., Ходжсон, Дж.А., и Реджи Эдгертон, В. (1999). Передача сил в скелетных мышцах млекопитающих. Дж. Биомех. 32, 371–380. doi: 10.1016/s0021-9290(98)00189-4
Полнотекстовая перекрестная ссылка | Академия Google
Мур, Д. Р., Филлипс, С. М., Бабрай, Дж. А., Смит, К., и Ренни, М. Дж. (2005). Синтез миофибриллярных и коллагеновых белков в скелетных мышцах человека у молодых мужчин после максимальных укорачивающих и удлиняющих сокращений. утра. Дж. Физиол.Эндокринол. Метаб. 288, E1153–E1159. doi: 10.1152/ajpendo.00387.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Моралес, М. Г., Кабельо-Верруджо, К., Сантандер, К., Кабрера, Д., Гольдшмединг, Р., и Брандан, Э. (2011). Сверхэкспрессия CTGF/CCN-2 может непосредственно индуцировать признаки дистрофии скелетных мышц.![]() Дж. Патол. 225, 490–501. doi: 10.1002/path.2952
Дж. Патол. 225, 490–501. doi: 10.1002/path.2952
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Малви, К., Харно Э., Кинан А. и Олендик К. (2005). Экспрессия скелетно-мышечного комплекса дистрофина-дистрогликана и комплекса синтрофина-синтазы оксида азота сильно нарушена у крыс Goto-Kakizaki с диабетом 2 типа. евро. Дж. Клеточная биология. 84, 867–883. doi: 10.1016/j.ejcb.2005.06.007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Наба, А., Клаузер, К.Р., Дин, Х., Уиттакер, К.А., Карр, С.А., и Хайнс, Р.О. (2016). Внеклеточный матрикс: инструменты и идеи для эпохи «омики». Матрикс Биол. 49, 10–24. doi: 10.1016/j.matbio.2015.06.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нассари, С., Дюпре, Д., и Фурнье-Тибо, К. (2017). Немиогенный вклад в развитие мышц и гомеостаз: роль соединительной ткани. Перед. Сотовый Дев. биол. 5:22. doi: 10.3389/fcell.2017.00022
doi: 10.3389/fcell.2017.00022
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нордес, А., Галло, Т., Кэтлин, С., Гевель, А., Корню, К., и Хуг, Ф. (2009). Электромеханическая задержка пересмотрена с использованием ультразвука с очень высокой частотой кадров. J. Appl. Физиол. 106, 1970–1975 гг. doi: 10.1152/japplphysiol.00221.2009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Новицки, Дж. Л., Такимото, Р., и Берк, А. С. (2003). Латеральная сомитическая граница: дорсо-вентральные аспекты передне-задней регионализации у эмбрионов птиц. Мех. Дев. 120, 227–240. doi: 10.1016/s0925-4773(02)00415-x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Огасавара, Р., Наказаво, К., Сато, К., Боппарт, М.Д., и Фудзита, С. (2014). Упражнения с отягощениями увеличивают активную экспрессию MMP и белка бета1-интегрина в скелетных мышцах. Физиол. Респ. 2:e12212. дои: 10.14814/phy2.12212
дои: 10.14814/phy2.12212
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ольгин, Х.К., и Олвин, Б.Б. (2004). Повышающая регуляция Pax-7 ингибирует миогенез и прогрессирование клеточного цикла в сателлитных клетках: потенциальный механизм самообновления. Дев. биол. 275, 375–388.doi: 10.1016/j.ydbio.2004.08.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Олссон Л., Фальк П., Лопес К., Кобб Дж. и Ханкен Дж. (2001). Клетки черепного нервного гребня вносят вклад в соединительную ткань в черепных мышцах бесхвостых амфибий Bombina orientalis . Дев. биол. 237, 354–367. doi: 10.1006/dbio.2001.0377
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пассерьё, Э., Россиньоль, Р., Летелье, Т., и Делаж, Дж. (2007). Физическая непрерывность перимизия от мышечных волокон до сухожилий: участие в передаче боковой силы в скелетных мышцах. Дж. Структура. биол. 159, 19–28. doi: 10.1016/j.jsb.2007.01.022
биол. 159, 19–28. doi: 10.1016/j.jsb.2007.01.022
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Паттисон, Дж. С., Фолк, Л. К., Мэдсен, Р. В., Чайлдс, Т. Е., и Бут, Ф. В. (2003). Транскрипционное профилирование выявляет обширное подавление экспрессии генов внеклеточного матрикса в саркопенической камбаловидной мышце крысы. Физиол. Геномика 15, 34–43. doi: 10.1152/physiolgenomics.00040.2003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Паван П.Г., Стекко А., Стерн Р. и Стекко К. (2014). Болезненные соединения: уплотнение против фиброза фасции. Курс. Боль Головная боль Респ. 18:441.
Академия Google
Пирс, Р. В., Шерц, П. Дж., Кэмпбелл, Дж. К., и Табин, С. Дж. (2007). Анализ клеточного происхождения зачатка куриной конечности. Дев. биол. 310, 388–400. doi: 10.1016/j.ydbio.2007.08.002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Питер, А. К., Ченг, Х., Росс, Р. С., Ноултон, К. У., и Чен, Дж. (2011). Костамера соединяет саркомеры с сарколеммой в поперечнополосатых мышцах. Прог. Педиатр. Кардиол. 31, 83–88. doi: 10.1016/j.ppedcard.2011.02.003
К., Ченг, Х., Росс, Р. С., Ноултон, К. У., и Чен, Дж. (2011). Костамера соединяет саркомеры с сарколеммой в поперечнополосатых мышцах. Прог. Педиатр. Кардиол. 31, 83–88. doi: 10.1016/j.ppedcard.2011.02.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Purslow, PP (2002). Структура и функциональное значение изменений соединительной ткани в мышцах. Комп. Биохим. Физиол. Мол. интегр. Физиол. 133, 947–966. doi: 10.1016/s1095-6433(02)00141-1
Полнотекстовая перекрестная ссылка | Академия Google
Рамасвами, К.С., Палмер, М.Л., ван дер Меулен, Дж.Х., Ренукс, А., Костроминова, Т.Ю., Мишель, Д.Е., и соавт. (2011). Боковая передача силы нарушена в скелетных мышцах дистрофических мышей и очень старых крыс: латеральная передача силы в скелетных мышцах мышей и крыс. J. Physiol. 589, 1195–1208.doi: 10.1113/jphysiol.2010.201921
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Раягири, С. С., Ранальди, Д., Рэйвен, А., Мохамад Ажар, Н.И.Ф., Лефевр, О., Заммит, П.С., и соавт. (2018). Ремоделирование базальной пластинки в нише стволовых клеток скелетных мышц опосредует самообновление стволовых клеток. Нац. коммун. 9:1075.
С., Ранальди, Д., Рэйвен, А., Мохамад Ажар, Н.И.Ф., Лефевр, О., Заммит, П.С., и соавт. (2018). Ремоделирование базальной пластинки в нише стволовых клеток скелетных мышц опосредует самообновление стволовых клеток. Нац. коммун. 9:1075.
Академия Google
Риз, С.П., Андервуд, С.Дж., и Вайс, Дж.А. (2013). Влияние декоринового протеогликана на фибриллогенез, ультраструктуру и механику гелей коллагена I типа. Матрикс Биол. 32, 414–423. doi: 10.1016/j.matbio.2013.04.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Роннинг, С. Б., Педерсен, М. Е., Андерсен, П. В., и Холлунг, К. (2013). Комбинация гликозаминогликанов и волокнистых белков улучшает пролиферацию клеток и раннюю дифференцировку первичных клеток скелетных мышц крупного рогатого скота. Дифференциация 86, 13–22. doi: 10.1016/j.diff.2013.06.006
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Роуленд, Л.А. , Бал, Н.К., и Периасами, М. (2015). Роль скелетно-мышечных термогенных механизмов в эндотермии позвоночных. биол. Преподобный Кэмб. Филос. соц. 90, 1279–1297. doi: 10.1111/brv.12157
, Бал, Н.К., и Периасами, М. (2015). Роль скелетно-мышечных термогенных механизмов в эндотермии позвоночных. биол. Преподобный Кэмб. Филос. соц. 90, 1279–1297. doi: 10.1111/brv.12157
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рубенштейн А.Б., Смит Г.Р., Рауэ У., Бег Г., Минчев К., Руф-Замойски Ф. и соавт. (2020). Профили транскрипции одиночных клеток в скелетных мышцах человека. науч. Респ. 10:229.
Академия Google
Сабателли, П., Gualandi, F., Gara, S.K., Grumati, P., Zamparelli, A., Martoni, E., et al. (2012). Экспрессия цепей α5 и α6 коллагена VI в мышцах человека и при мышечном фиброзе, связанном с мышечной дистрофией Дюшенна. Матрикс Биол. 31, 187–196. doi: 10.1016/j.matbio.2011.12.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Санес, младший (1982). Ламинин, фибронектин и коллаген в синаптических и экстрасинаптических участках базальной мембраны мышечных волокон. Дж.Клеточная биол. 93, 442–451. doi: 10.1083/jcb.93.2.442
Дж.Клеточная биол. 93, 442–451. doi: 10.1083/jcb.93.2.442
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Savolainen, J., Väänänen, K., Puranen, J., Takala, T.E., Komulainen, J., and Vihko, V. (1988). Синтез коллагена и протеолитическая активность в скелетных мышцах крыс: эффект гипсовой иммобилизации в удлиненном и укороченном положениях. Арх. физ. Мед. Реабилит. 69, 964–969.
Реферат PubMed | Академия Google
Шенфельд, Б.Дж. (2010). Механизмы мышечной гипертрофии и их применение в тренировках с отягощениями. Дж. Сила конд. Рез. 24, 2857–2872. doi: 10.1519/JSC.0b013e3181e840f3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шенрок Б., Зандер В., Дерн С., Лимпер У., Малдер Э., Веракситш А. и др. (2018). Постельный режим, меры противодействия физическим нагрузкам и восстановительное воздействие на систему мышечного тонуса человека в состоянии покоя. Перед.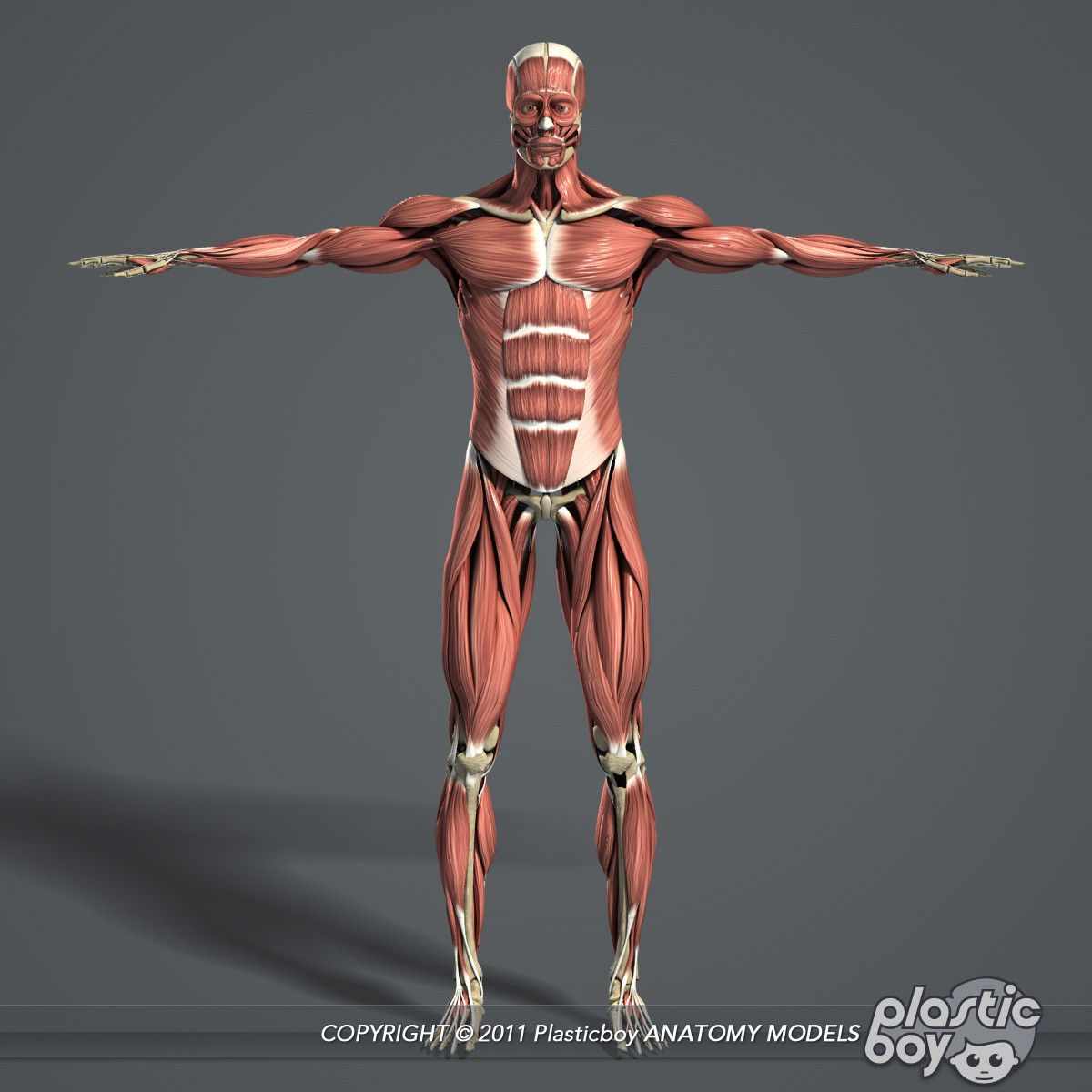 Физиол. 9:810. дои: 10.3389/ффиз.2018.00810
Физиол. 9:810. дои: 10.3389/ффиз.2018.00810
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Schönherr, E., Witsch-Prehm, P., Harrach, B., Robenek, H., Rauterberg, J., and Kresse, H. (1995). Взаимодействие бигликана с коллагеном I типа. Дж. Биол. хим. 270, 2776–2783. doi: 10.1074/jbc.270.6.2776
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шварц, Р.И. (2015). Коллаген I и фибробласты: высокая экспрессия белка требует новой парадигмы посттранскрипционной регуляции с обратной связью. Биохим. Биофиз. 3, 38–44. doi: 10.1016/j.bbrep.2015.07.007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сефтон, Э. М., и Кардон, Г. (2019). Связь развития мышц, врожденных дефектов и эволюции: важная роль мышечной соединительной ткани. Курс. Вверх. Дев. биол. 132, 137–176. doi: 10.1016/bs.ctdb.2018.12.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шарафи Б. и Блемкер С.С. (2011). Математическая модель передачи силы от внутрипучковых окончаний мышечных волокон. Дж. Биомех. 44, 2031–2039. doi: 10.1016/j.jbiomech.2011.04.038
и Блемкер С.С. (2011). Математическая модель передачи силы от внутрипучковых окончаний мышечных волокон. Дж. Биомех. 44, 2031–2039. doi: 10.1016/j.jbiomech.2011.04.038
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Соренсен, Дж. Р., Скаузен, К., Холланд, А., Уильямс, К., и Хилдал, Р. Д. (2018). Острая реакция внеклеточного матрикса, воспаления и МАРК на удлинение скелетных мышц человека пожилого возраста. Экспл. Геронтол. 106, 28–38.doi: 10.1016/j.exger.2018.02.013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Street, SF (1983). Боковая передача напряжения в миофибриллах лягушки: возможными передатчиками являются миофибриллярная сеть и поперечные цитоскелетные связи. Дж. Сотовый. Физиол. 114, 346–364. doi: 10.1002/jcp.1041140314
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сумиёси Х., Лауб Ф., Йошиока Х. и Рамирес Ф. (2001).Эмбриональная экспрессия коллагена типа XIX носит временный характер и ограничена мышечными клетками. Дев. Дин. 220, 155–162. doi: 10.1002/1097-0177(2000)9999:9999<::aid-dvdy1099>3.0.co;2-w
Дев. Дин. 220, 155–162. doi: 10.1002/1097-0177(2000)9999:9999<::aid-dvdy1099>3.0.co;2-w
Полнотекстовая перекрестная ссылка | Академия Google
Суоминен Х. и Хейккинен Э. (1975). Активность ферментов в мышечной и соединительной ткани M. Vastus lateralis у мужчин в возрасте от 33 до 70 лет, которые обычно тренируются и ведут малоподвижный образ жизни. евро. Дж. Заявл. Физиол. 34, 249–254. дои: 10.1007/bf00999938
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Суоминен Х., Хейккинен Э. и Паркатти Т. (1977). Влияние восьминедельной физической тренировки на мышцы и соединительную ткань M. Vastus lateralis у 69-летних мужчин и женщин. Дж. Геронтол. 32, 33–37. doi: 10.1093/geronj/32.1.33
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шварц, М.Е., Эберхарт, Дж., Паскуале, Э.Б., и Крулл, К.Э. (2001). Взаимодействия EphA4/ephrin-A5 при миграции клеток-предшественников мышц в передних конечностях птиц. Дев. Камб. англ. 128, 4669–4680.
Дев. Камб. англ. 128, 4669–4680.
Реферат PubMed | Академия Google
Такаги Р., Огасавара Р., Цутаки А., Накадзато К. и Исии Н. (2016). Региональная адаптация коллагена в скелетных мышцах к повторным приступам напряженных эксцентрических упражнений. Арка Пфлюгера. 468, 1565–1572. doi: 10.1007/s00424-016-1860-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Такала, Т.Э. и Виртанен П. (2000). Биохимический состав внеклеточного матрикса мышц: влияние нагрузки. Скан. Дж. Мед. науч. Спорт 10, 321–325. doi: 10.1034/j.1600-0838.2000.010006321.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Такала, Т.Е.С., Мюллюля, Р., Салминен, А., Анттинен, Х., и Вихко, В. (1983). Повышение активности пролил-4-гидроксилазы и галактозилгидроксилизилглюкозилтрансферазы, ферментов биосинтеза коллагена, в скелетных мышцах мышей, тренированных на выносливость. Арка Пфлюгера. 399, 271–274. дои: 10.1007/BF00652751
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Там, К.С., Чаудхури, Р., Хатчисон, А.Т., Самоча-Бонет, Д., и Хейлбронн, Л.К. (2017). Ремоделирование внеклеточного матрикса скелетных мышц после кратковременного переедания у здоровых людей. Метаболизм 67, 26–30. doi: 10.1016/j.metabol.2016.10.009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Там, К.С., Ковингтон, Дж. Д., Байпеи, С., Чукалова, Ю., Бурк, Д., Йоханнсен, Д. Л., и соавт. (2014). Прибавка в весе показывает резкое усиление ремоделирования внеклеточного матрикса скелетных мышц. Дж. Клин. Эндокринол. Метаб. 99, 1749–1757. doi: 10.1210/jc.2013-4381
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Tam, C.S., Power, J.E., Markovic, T.P., Yee, C., Morsch, M., McLennan, S.V., et al. (2015). Влияние питания с высоким содержанием жиров на физическую функцию и внеклеточный матрикс скелетных мышц. Нутр. Диабет 5:e187. doi: 10.1038/nutd.2015.39
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Thorsteinsdóttir, S., Deries, M., Cachaço, A.S., and Bajanca, F. (2011). Размер внеклеточного матрикса развития скелетных мышц. Дев. биол. 354, 191–207. doi: 10.1016/j.ydbio.2011.03.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тирни М.Т., Громова А., Сесильо Ф.Б., Сала Д., Спенле К., Orend, G., et al. (2016). Автономное ремоделирование внеклеточного матрикса контролирует прогрессивную адаптацию регенеративной способности мышечных стволовых клеток во время развития. Cell Rep. 14, 1940–1952. doi: 10.1016/j.celrep.2016.01.072
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уэдзуми А., Фукада С., Ямамото Н., Такэда С. и Цучида К. (2010). Мезенхимальные предшественники, отличные от сателлитных клеток, способствуют образованию эктопических жировых клеток в скелетных мышцах. Нац. Клеточная биол. 12, 143–152. doi: 10.1038/ncb2014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Улен, М., Оксволд, П., Фагерберг, Л., Лундберг, Э., Йонассон, К., Форсберг, М., и соавт. (2010). К основанному на знаниях атласу белков человека. Нац. Биотехнолог. 28, 1248–1250. doi: 10.1038/nbt1210-1248
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Урчуоло А., Кварта М., Морбидони В., Гаттаццо Ф., Молон С., Грумати П. и соавт. (2013). Коллаген VI регулирует самообновление сателлитных клеток и регенерацию мышц. Нац. коммун. 4:1964. дои: 10.1038/ncomms2964
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Vallecillo-García, P., Orgeur, M., Vom Hofe-Schneider, S., Stumm, J., Kappert, V., Ibrahim, D.M., et al. (2017). Odd skipped-related 1 идентифицирует популяцию эмбриональных фибро-адипогенных предшественников, регулирующих миогенез во время развития конечностей. Нац. коммун. 8:1218.
Академия Google
Вирджилио, К.М., Мартин, К.С., Пирс, С.М., и Блемкер, С.С. (2015). Многомасштабные модели скелетных мышц показывают сложные эффекты мышечной дистрофии на механику тканей и восприимчивость к повреждениям. Интерфейс Focus 5:20140080. doi: 10.1098/rsfs.2014.0080
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Виссе Р. и Нагасе Х. (2003). Матриксные металлопротеиназы и тканевые ингибиторы металлопротеиназ: структура, функции и биохимия. Обр. Рез. 92, 827–839. doi: 10.1161/01.RES.0000070112.80711.3D
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Wang, D., Eraslan, B., Wieland, T., Hallström, B., Hopf, T., Zolg, D.P., et al. (2019). Глубокий атлас изобилия протеомов и транскриптомов 29 здоровых тканей человека. Мол. Сист. биол. 15:e8503. doi: 10.15252/msb.20188503
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Весснер Б., Либенштайнер М., Нахбауэр, В., и Чапо, Р. (2019). Возрастная реакция внеклеточного матрикса скелетных мышц на острые упражнения с отягощениями: экспериментальное исследование. евро. Дж. Спортивные науки. 19, 354–364. дои: 10.1080/17461391.2018.1526974
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
White, J.P., Reecy, J.M., Washington, T.A., Sato, S., Le, M.E., Davis, J.M., et al. (2009). Индуцированное перегрузкой ремоделирование внеклеточного матрикса скелетных мышц и рост миофибрилл у мышей, лишенных IL-6. Acta Physiol. 197, 321–332. doi: 10.1111/j.1748-1716.2009.02029.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Wilschut, KJ, Haagsman, H.P., and Roelen, B.A.J. (2010). Компоненты внеклеточного матрикса определяют поведение стволовых клеток свиных мышц. Экспл. Сотовый рез. 316, 341–352. doi: 10.1016/j.yexcr.2009.10.014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вуд Л.К., Каюпов Э., Гумуцио Дж.П., Мендиас, К.Л., Клафлин, Д.Р., и Брукс, С.В. (2014). Внутренняя жесткость внеклеточного матрикса увеличивается с возрастом в скелетных мышцах мышей. J. Appl. Физиол. 117, 363–369. doi: 10.1152/japplphysiol.00256.2014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Yucesoy, C.A., Maas, H., Koopman, B.H.F.J.M., Grootenboer, H.J., и Huijing, P.A. (2006). Механизмы, вызывающие влияние положения мышц на различия проксимально-дистальной мышечной силы при внемышечной миофасциальной передаче силы. Мед. англ. физ. 28, 214–226. doi: 10.1016/j.medengphy.2005.06.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zammit, PS (2017). Функция миогенных регуляторных факторов Myf5, MyoD, Myogenin и MRF4 в скелетных мышцах, сателлитных клетках и регенеративном миогенезе. Семин. Сотовый Дев. биол. 72, 19–32. doi: 10.1016/j.semcdb.2017.11.011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжан, К.и Гао Ю. (2012). Анализ методом конечных элементов механики латеральной передачи силы в одиночном мышечном волокне. Дж. Биомех. 45, 2001–2006 гг. doi: 10.1016/j.jbiomech.2012.04.026
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zhu, J., Li, Y., Shen, W., Qiao, C., Ambrosio, F., Lavasani, M., et al. (2007). Взаимоотношения между трансформирующим фактором роста-β1, миостатином и декорином: значение для фиброза скелетных мышц. Дж. Биол. хим. 282, 25852–25863.doi: 10.1074/jbc.M704146200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zong, H., Bastie, C.C., Xu, J., Fassler, R., Campbell, K.P., Kurland, I.J., et al. (2009). Инсулинорезистентность у мышей с дефицитом бета1-рецептора интегрина, специфичного для поперечнополосатых мышц. Дж. Биол. хим. 284, 4679–4688. doi: 10.1074/jbc.M807408200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зюгель М., Маганарис С. Н., Вилке Дж., Юркат-Ротт К., Klingler, W., Wearing, S.C., et al. (2018). Исследования фасциальных тканей в спортивной медицине: от молекул к адаптации тканей, травмам и диагностике: консенсусное заявление. руб. Дж. Спорт Мед. 52:1497. дои: 10.1136/bjsports-2018-099308
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Скелетные мышцы – анатомия и физиология
OpenStaxCollege
Цели обучения
К концу этого раздела вы сможете:
- Опишите слои соединительной ткани, упаковывающие скелетные мышцы
- Объясните, как мышцы работают с сухожилиями для движения тела
- Определение областей скелетных мышечных волокон
- Описать муфту возбуждения-сокращения
Наиболее известной особенностью скелетных мышц является их способность сокращаться и вызывать движения.Скелетные мышцы действуют не только для создания движения, но и для его остановки, например, сопротивления гравитации для сохранения осанки. Небольшие постоянные корректировки скелетных мышц необходимы, чтобы удерживать тело в вертикальном положении или в равновесии в любом положении. Мышцы также предотвращают чрезмерное движение костей и суставов, поддерживая стабильность скелета и предотвращая повреждение или деформацию скелетной структуры. Суставы могут быть смещены или полностью вывихнуты из-за натяжения связанных с ними костей; мышцы работают, чтобы суставы оставались стабильными.Скелетные мышцы расположены по всему телу у отверстий внутренних путей и контролируют движение различных веществ. Эти мышцы позволяют контролировать такие функции, как глотание, мочеиспускание и дефекация. Скелетные мышцы также защищают внутренние органы (особенно органы брюшной полости и таза), выступая в качестве внешнего барьера или щита от внешних травм и поддерживая вес органов.
Скелетные мышцы способствуют поддержанию гомеостаза в организме, вырабатывая тепло.Для сокращения мышц требуется энергия, а при расщеплении АТФ выделяется тепло. Это тепло очень заметно во время физических упражнений, когда продолжительное движение мышц вызывает повышение температуры тела, а в случаях сильного холода, когда дрожь вызывает случайные сокращения скелетных мышц для выработки тепла.
Каждая скелетная мышца представляет собой орган, состоящий из различных интегрированных тканей. Эти ткани включают волокна скелетных мышц, кровеносные сосуды, нервные волокна и соединительную ткань. Каждая скелетная мышца имеет три слоя соединительной ткани (так называемые «мизии»), которые ее окружают и обеспечивают структуру мышцы в целом, а также разделяют мышечные волокна внутри мышцы ([ссылка]).Каждая мышца покрыта оболочкой из плотной соединительной ткани неправильной формы, называемой эпимизием, которая позволяет мышце сокращаться и двигаться, сохраняя при этом свою структурную целостность. Эпимизий также отделяет мышцу от других тканей и органов в этой области, позволяя мышце двигаться независимо.
Три слоя соединительной ткани
Пучки мышечных волокон, называемые пучками, покрыты перимизием. Мышечные волокна покрыты эндомизием.
Внутри каждой скелетной мышцы мышечные волокна организованы в отдельные пучки, каждый из которых называется пучком, при помощи среднего слоя соединительной ткани, называемого перимизием.Эта фасцикулярная организация обычна для мышц конечностей; это позволяет нервной системе запускать определенное движение мышцы, активируя подмножество мышечных волокон в пучке или пучке мышц. Внутри каждого пучка каждое мышечное волокно заключено в тонкий слой соединительной ткани из коллагена и ретикулярных волокон, называемый эндомизием. Эндомизий содержит внеклеточную жидкость и питательные вещества для поддержки мышечного волокна. Эти питательные вещества поступают через кровь в мышечную ткань.
В скелетных мышцах, которые работают с сухожилиями, чтобы тянуть кости, коллаген в трех тканевых слоях (мизия) переплетается с коллагеном сухожилия. На другом конце сухожилие срастается с надкостницей, покрывающей кость. Напряжение, создаваемое сокращением мышечных волокон, затем передается через мизии на сухожилия, а затем на надкостницу, натягивая кость для движения скелета. В других местах мизия может сливаться с широким, похожим на сухожилие листком, называемым апоневрозом, или с фасцией, соединительной тканью между кожей и костями.Широкий слой соединительной ткани в нижней части спины, в который сливаются широчайшие мышцы спины («широчайшие»), является примером апоневроза.
Каждая скелетная мышца также богато снабжена кровеносными сосудами для питания, доставки кислорода и удаления отходов. Кроме того, каждое мышечное волокно в скелетной мышце снабжается аксонной ветвью соматического моторного нейрона, который сигнализирует волокну о сокращении. В отличие от сердечных и гладких мышц, единственный способ функционального сокращения скелетных мышц — передача сигналов от нервной системы.
Поскольку клетки скелетных мышц длинные и цилиндрические, их обычно называют мышечными волокнами. Скелетные мышечные волокна могут быть довольно большими для клеток человека, с диаметром до 100 92 147 μ 90 148 мкм и длиной до 30 см (11,8 дюйма) в портняжной мышце бедра. Во время раннего развития эмбриональные миобласты, каждый со своим собственным ядром, сливаются с сотнями других миобластов, образуя многоядерные скелетные мышечные волокна. Множественные ядра означают множество копий генов, позволяющих производить большое количество белков и ферментов, необходимых для сокращения мышц.
Некоторые другие термины, связанные с мышечными волокнами, восходят к греческому sarco , что означает «плоть». Плазматическая мембрана мышечных волокон называется сарколеммой, цитоплазма – саркоплазмой, а специализированный гладкий эндоплазматический ретикулум, который хранит, высвобождает и извлекает ионы кальция (Са ++), называется саркоплазматическим ретикулумом (СР). ([ссылка на сайт]). Как будет вскоре описано, функциональной единицей скелетных мышечных волокон является саркомер, высокоорганизованная структура сократительных миофиламентов актина (тонкая нить) и миозина (толстая нить) вместе с другими поддерживающими белками.
Мышечное волокно
Скелетные мышечные волокна окружены плазматической мембраной, называемой сарколеммой, которая содержит саркоплазму, цитоплазму мышечных клеток. Мышечное волокно состоит из множества фибрилл, которые придают клетке поперечно-полосатый вид.
Исчерченность скелетных мышечных волокон обусловлена расположением миофиламентов актина и миозина в последовательном порядке от одного конца мышечного волокна к другому. Каждый пакет этих микрофиламентов и их регуляторных белков, тропонина и тропомиозина (наряду с другими белками) называется саркомером.
Посмотрите это видео, чтобы узнать больше о макро- и микроструктурах скелетных мышц. а) Как называются «точки соединения» между саркомерами? б) Как называются «субъединицы» внутри миофибрилл, которые проходят по всей длине волокон скелетных мышц? в) Что такое «двойная нить жемчуга», описанная в видео? г) Что придает скелетным мышечным волокнам поперечно-полосатый вид?
Саркомер является функциональной единицей мышечного волокна. Сам саркомер связан внутри миофибриллы, которая проходит по всей длине мышечного волокна и прикрепляется к сарколемме на его конце.При сокращении миофибрилл сокращается вся мышечная клетка. Поскольку диаметр миофибрилл составляет приблизительно 1,2 мкм мкм, внутри одного мышечного волокна можно найти от сотен до тысяч (каждый с тысячами саркомеров). Каждый саркомер имеет длину примерно 2 мкм м, имеет трехмерное цилиндрическое расположение и граничит со структурами, называемыми Z-дисками (также называемыми Z-линиями, поскольку изображения двумерные), к которым присоединяются актиновые миофиламенты. закреплено ([ссылка]).Поскольку актин и его тропонин-тропомиозиновый комплекс (выступающие из Z-дисков к центру саркомера) образуют более тонкие нити, чем миозин, его называют тонкой нитью саркомера. Точно так же, поскольку нити миозина и их многочисленные головки (выступающие из центра саркомера по направлению к Z-дискам, но не до конца) имеют большую массу и толще, их называют толстыми нитями саркомера.
Саркомер
Саркомер, область от одной Z-линии до следующей Z-линии, является функциональной единицей скелетного мышечного волокна.
Другой специализацией скелетных мышц является место, где окончание моторного нейрона встречается с мышечным волокном, называемое нервно-мышечным соединением (НМС). Именно здесь мышечное волокно в первую очередь отвечает на сигналы двигательного нейрона. Каждое мышечное волокно в каждой скелетной мышце иннервируется моторным нейроном в НМС. Сигналы возбуждения от нейрона — единственный способ функционально активировать сокращение волокна.
Каждое волокно скелетной мышцы иннервируется моторным нейроном в НМС.Посмотрите это видео, чтобы узнать больше о том, что происходит в NMJ. а) Каково определение двигательной единицы? б) Каковы структурные и функциональные различия между крупной двигательной единицей и мелкой двигательной единицей? в) Можете ли вы привести пример каждого из них? (г) Почему нейротрансмиттер ацетилхолин расщепляется после связывания с его рецептором?
Все живые клетки имеют мембранные потенциалы или электрические градиенты на своих мембранах. Внутренняя часть мембраны обычно составляет от -60 до -90 мВ относительно внешней.Это называется мембранным потенциалом клетки. Нейроны и мышечные клетки могут использовать свои мембранные потенциалы для генерации электрических сигналов. Они делают это, контролируя движение заряженных частиц, называемых ионами, через свои мембраны, создавая электрические токи. Это достигается открытием и закрытием специализированных белков в мембране, называемых ионными каналами. Хотя токи, генерируемые ионами, движущимися через белки этих каналов, очень малы, они составляют основу как передачи нервных сигналов, так и сокращения мышц.
И нейроны, и клетки скелетных мышц электрически возбудимы, что означает, что они способны генерировать потенциалы действия. Потенциал действия — это особый тип электрического сигнала, который может распространяться вдоль клеточной мембраны в виде волны. Это позволяет передавать сигнал быстро и точно на большие расстояния.
Хотя термин «сопряжение возбуждения-сокращения» сбивает с толку или пугает некоторых студентов, он сводится к следующему: для того, чтобы скелетное мышечное волокно сокращалось, его мембрана должна быть сначала «возбуждена» — другими словами, она должна быть стимулирована для запуска потенциала действия. .Потенциал действия мышечного волокна, который волной проносится вдоль сарколеммы, «сцеплен» с собственно сокращением за счет высвобождения ионов кальция (Са ++) из СР. После высвобождения Са ++ взаимодействует с экранирующими белками, заставляя их отойти в сторону, чтобы сайты связывания актина были доступны для прикрепления миозиновыми головками. Затем миозин тянет актиновые филаменты к центру, укорачивая мышечное волокно.
В скелетных мышцах эта последовательность начинается с сигналов сомато-двигательного отдела нервной системы.Другими словами, стадия «возбуждения» в скелетных мышцах всегда запускается сигналом от нервной системы ([ссылка]).
Торцевая пластина двигателя и иннервация
В СНС окончание аксона высвобождает АХ. Моторная концевая пластинка представляет собой место расположения АХ-рецепторов в сарколемме мышечного волокна. Когда молекулы АХ высвобождаются, они диффундируют через крошечное пространство, называемое синаптической щелью, и связываются с рецепторами.
Мотонейроны, которые сообщают скелетным мышечным волокнам о сокращении, происходят из спинного мозга, при этом меньшее количество расположено в стволе головного мозга для активации скелетных мышц лица, головы и шеи.Эти нейроны имеют длинные отростки, называемые аксонами, которые специализируются на передаче потенциалов действия на большие расстояния — в данном случае от спинного мозга до самих мышц (которые могут находиться на расстоянии до трех футов). Аксоны нескольких нейронов связываются вместе, образуя нервы, как провода, связанные вместе в кабеле.
Передача сигналов начинается, когда потенциал действия нейрона проходит по аксону двигательного нейрона, а затем по отдельным ветвям и заканчивается в НМС. В НМС окончание аксона высвобождает химический мессенджер или нейротрансмиттер, называемый ацетилхолином (АХ).Молекулы АХ диффундируют через крошечное пространство, называемое синаптической щелью, и связываются с рецепторами АХ, расположенными внутри моторной концевой пластинки сарколеммы на другой стороне синапса. Как только АХ связывается, канал в рецепторе АХ открывается, и положительно заряженные ионы могут проникать в мышечное волокно, вызывая его деполяризацию, а это означает, что мембранный потенциал мышечного волокна становится менее отрицательным (ближе к нулю)
.Когда мембрана деполяризуется, открывается другой набор ионных каналов, называемых потенциалзависимыми натриевыми каналами.Ионы натрия проникают в мышечное волокно, и потенциал действия быстро распространяется (или «выстреливает») по всей мембране, инициируя сопряжение возбуждения и сокращения.
В мире возбудимых мембран все происходит очень быстро (только подумайте, как быстро вы можете щелкнуть пальцами, как только решите это сделать). Сразу после деполяризации мембраны она реполяризуется, восстанавливая отрицательный мембранный потенциал. Между тем, АХ в синаптической щели расщепляется ферментом ацетилхолинэстеразой (АХЭ), так что АХ не может повторно связываться с рецептором и снова открывать свой канал, что может вызвать нежелательное длительное возбуждение и сокращение мышц.
Распространение потенциала действия вдоль сарколеммы является частью возбуждения сопряжения возбуждения-сокращения. Напомним, что это возбуждение фактически запускает высвобождение ионов кальция (Са ++) из его запасов в СР клетки. Чтобы потенциал действия достиг мембраны СР, в сарколемме имеются периодические впячивания, называемые Т-трубочками («Т» означает «поперечные»). Вы помните, что диаметр мышечного волокна может достигать 100 мкм м, поэтому эти Т-трубочки обеспечивают возможность сближения мембраны с СР в саркоплазме.Расположение Т-трубочки с мембранами СР по обе стороны называется триадой ([ссылка]). Триада окружает цилиндрическую структуру, называемую миофибриллой, которая содержит актин и миозин.
Т-образная трубка
Узкие Т-трубочки позволяют проводить электрические импульсы. SR функционирует, чтобы регулировать внутриклеточные уровни кальция. Две терминальные цистерны (где увеличенный СР соединяется с Т-трубочкой) и одна Т-трубочка составляют триаду — «тройку» мембран, с мембранами СР с двух сторон и Т-трубочкой, зажатой между ними.
Т-трубочки несут потенциал действия внутрь клетки, что запускает открытие кальциевых каналов в мембране соседнего СР, в результате чего Са ++ диффундирует из СР в саркоплазму. Именно прибытие Ca ++ в саркоплазму инициирует сокращение мышечного волокна его сократительными единицами, или саркомерами.
Скелетные мышцы содержат соединительную ткань, кровеносные сосуды и нервы.Различают три слоя соединительной ткани: эпимизий, перимизий и эндомизий. Скелетные мышечные волокна организованы в группы, называемые пучками. Кровеносные сосуды и нервы входят в соединительную ткань и разветвляются в клетке. Мышцы прикрепляются к костям непосредственно или через сухожилия или апоневрозы. Скелетные мышцы поддерживают осанку, стабилизируют кости и суставы, контролируют внутренние движения и вырабатывают тепло.
Волокна скелетных мышц представляют собой длинные многоядерные клетки. Мембрана клетки — сарколемма; цитоплазма клетки – саркоплазма.Саркоплазматический ретикулум (СР) представляет собой форму эндоплазматического ретикулума. Мышечные волокна состоят из миофибрилл. Исчерченность создается организацией актина и миозина, что приводит к полосатости миофибрилл.
Посмотрите это видео, чтобы узнать больше о макро- и микроструктурах скелетных мышц. а) Как называются «точки соединения» между саркомерами? б) Как называются «субъединицы» внутри миофибрилл, которые проходят по всей длине волокон скелетных мышц? в) Что такое «двойная нить жемчуга», описанная в видео? г) Что придает скелетным мышечным волокнам поперечно-полосатый вид?
(а) Z-линии.(б) Саркомеры. (в) Это расположение актиновых и миозиновых филаментов в саркомере. (г) чередующиеся нити актиновых и миозиновых филаментов.
Каждое волокно скелетной мышцы иннервируется моторным нейроном в НМС. Посмотрите это видео, чтобы узнать больше о том, что происходит в нервно-мышечном соединении. а) Каково определение двигательной единицы? б) Каковы структурные и функциональные различия между крупной двигательной единицей и мелкой двигательной единицей? Можете ли вы привести пример каждого? в) Почему нейротрансмиттер ацетилхолин расщепляется после связывания с его рецептором?
(а) Количество волокон скелетных мышц, иннервируемых одним мотонейроном.(b) Крупная двигательная единица имеет один нейрон, снабжающий многие волокна скелетных мышц для крупных движений, например височная мышца, где 1000 волокон снабжаются одним нейроном. Небольшой двигатель имеет один нейрон, который снабжает несколько волокон скелетных мышц для очень тонких движений, как экстраокулярные мышцы глаза, где шесть волокон снабжаются одним нейроном. (c) Чтобы избежать продления мышечного сокращения.
Правильный порядок организации мышечной ткани от наименьшей до наибольшей ________.
- пучок, филамент, мышечное волокно, миофибрилла
- нить, миофибрилла, мышечное волокно, пучок
- мышечное волокно, пучок, филамент, миофибрилла
- миофибриллы, мышечные волокна, нити, пучки
Деполяризация сарколеммы означает ________.
- внутренняя часть мембраны стала менее отрицательной по мере накопления ионов натрия
- внешняя сторона мембраны стала менее отрицательной по мере накопления ионов натрия
- внутренняя часть мембраны стала более отрицательной из-за накопления ионов натрия
- сарколемма полностью потеряла электрический заряд
Что произойдет со скелетными мышцами, если разрушить эпимизиум?
Мышцы теряли свою целостность при сильных движениях, что приводило к повреждению мышц.
Опишите, как сухожилия облегчают движения тела.
Когда мышца сокращается, сила движения передается через сухожилие, которое натягивает кость, вызывая скелетное движение.
Каковы пять основных функций скелетных мышц?
Обеспечивает движение скелета, поддерживает осанку и положение тела, поддерживает мягкие ткани, окружает отверстия пищеварительного, мочевого и других путей и поддерживает температуру тела.
Каковы противоположные роли потенциалзависимых натриевых каналов и потенциалзависимых калиевых каналов?
Открытие потенциалзависимых натриевых каналов с последующим притоком Na + передает потенциал действия после достаточной деполяризации мембраны.Отсроченное открытие калиевых каналов позволяет К + выйти из клетки, чтобы реполяризовать мембрану.
Глоссарий
- ацетилхолин (АХ)
- нейротрансмиттер, который связывается с концевой пластинкой двигателя, вызывая деполяризацию
- актин
- белок, составляющий большую часть тонких миофиламентов в саркомерном мышечном волокне
- потенциал действия
- изменение напряжения клеточной мембраны в ответ на раздражитель, приводящее к передаче электрического сигнала; уникальный для нейронов и мышечных волокон
- апоневроз
- широкий, похожий на сухожилие слой соединительной ткани, который прикрепляет скелетную мышцу к другой скелетной мышце или к кости
- деполяризовать
- для уменьшения разницы потенциалов между внутренней и внешней частью клеточной плазматической мембраны (сарколемма мышечного волокна), делая внутреннюю часть менее отрицательной, чем в состоянии покоя
- эндомизий
- рыхлая и хорошо гидратированная соединительная ткань, покрывающая каждое мышечное волокно в скелетной мышце
- эпимизий
- наружный слой соединительной ткани вокруг скелетной мышцы
- Муфта возбуждения-сокращения
- последовательность событий от передачи сигналов двигательного нейрона волокну скелетной мышцы до сокращения саркомеров волокна
- выпуск
- пучок мышечных волокон внутри скелетной мышцы
- торцевая пластина двигателя
- сарколемма мышечного волокна в нервно-мышечном соединении с рецепторами нейротрансмиттера ацетилхолина
- миофибриллы
- длинная цилиндрическая органелла, идущая параллельно мышечному волокну и содержащая саркомеры
- миозин
- белок, который составляет большую часть толстого цилиндрического миофиламента в мышечном волокне саркомера
- нервно-мышечное соединение (НМС)
- синапс между окончанием аксона моторного нейрона и участком мембраны мышечного волокна с рецепторами ацетилхолина, высвобождаемого окончанием
- нейротрансмиттер
- сигнальное химическое вещество, высвобождаемое нервными окончаниями, которое связывается и активирует рецепторы на клетках-мишенях
- перимизий
- соединительная ткань, которая связывает волокна скелетных мышц в пучки внутри скелетной мышцы
- саркомер
- продольно повторяющаяся функциональная единица скелетной мышцы, в которой участвуют все сократительные и ассоциированные белки
- сарколемма
- плазматическая мембрана скелетно-мышечного волокна
- саркоплазма
- цитоплазма мышечной клетки
- саркоплазматический ретикулум (SR)
- специализированный гладкий эндоплазматический ретикулум, который хранит, высвобождает и извлекает Ca ++
- синаптическая щель
- пространство между окончанием нерва (аксона) и двигательной концевой пластинкой
- Т-образная трубка
- проекция сарколеммы внутрь клетки
- толстая нить
- толстые нити миозина и их многочисленные головки, выступающие из центра саркомера по направлению к Z-дискам, но не полностью к ним
- тонкая нить
- тонкие нити актина и его тропонин-тропомиозиновый комплекс, выступающие из Z-дисков к центру саркомера
- триада
- группа из одной Т-трубочки и двух терминальных цистерн
- тропонин
- регуляторный белок, который связывается с актином, тропомиозином и кальцием
- тропомиозин
- регуляторный белок, покрывающий сайты связывания миозина, чтобы предотвратить связывание актина с миозином
- потенциалзависимые натриевые каналы
- мембранные белки, которые открывают натриевые каналы в ответ на достаточное изменение напряжения и инициируют и передают потенциал действия, когда Na + входит через канал