
Ответ Тема 4. Опорно-двигательная система
36. Сформулируйте несколько вопросов, ответы на которые вы хотите получить при изучении этой темы.
37. Прочитайте §10. Представьте в виде схемы перечень функций опорно-двигательной системы.
-
Ответ:
38. Раскройте роль органических и неорганических веществ кости, дописав утверждения.
39. Впишите в текст о строении трубчатой кости пропущенные слова: суставный хрящ, надкостница, компактное вещество, костномозговая полость, губчатое вещество, красный костный мозг, жёлтый костный мозг.
-
Ответ: Трубчатая кость состоит из средней части – тела кости и двух головок, сочленяющих с другими костями. Тело кости и наружная часть головок покрыта надкостница, а суставные поверхности головок – суставным хрящом
 Внутри головок находится губчатое вещество, содержащее красный костный мозг, в котором образуются клетки крови. Тело кости состоит из компактного вещества внутри которого находится костномозговая полость. Она заполнена жёлтым костным мозгом, представляющим в основном жировую ткань.
Внутри головок находится губчатое вещество, содержащее красный костный мозг, в котором образуются клетки крови. Тело кости состоит из компактного вещества внутри которого находится костномозговая полость. Она заполнена жёлтым костным мозгом, представляющим в основном жировую ткань.
 Внутри головок находится губчатое вещество, содержащее красный костный мозг, в котором образуются клетки крови. Тело кости состоит из компактного вещества внутри которого находится костномозговая полость. Она заполнена жёлтым костным мозгом, представляющим в основном жировую ткань.
Внутри головок находится губчатое вещество, содержащее красный костный мозг, в котором образуются клетки крови. Тело кости состоит из компактного вещества внутри которого находится костномозговая полость. Она заполнена жёлтым костным мозгом, представляющим в основном жировую ткань.
40. Ответьте, почему, несмотря на то, что рост кости в толщину осуществляется непрерывно за счёт надкостницы, кость взрослого человека не становится массивнее.
41. Отчёт о лабораторной работе «Микроскопическое строение кости» (с. 69-70 учебника).
-
Ответ:
1.На микропрепарате представлен поперечный срез кости.
2.Рисунок микроскопического строения кости с обозначением деталей, увиденных в микроскоп.
3.Ответы на вопросы задания №4 лабораторной работы.
Ответ: Костные пластинки располагаются по направлению сил сжатия и растяжения.
Возникающих в связи с нагрузками на кость, это объясняет уникальную прочность кости.
 Возникающих в связи с нагрузками на кость, это объясняет уникальную прочность кости.
Возникающих в связи с нагрузками на кость, это объясняет уникальную прочность кости.
42. Прочитайте § 11 «Скелет человека. Осевой скелет». Заполните таблицу, впишите названия парных и непарных костей черепа, а также цифры, которыми эти кости обозначены на рисунке.
-
Ответ:
Кости мозгового черепа
№ на рисунке
Кости лицевого черепа
№ на рисунке
Парные
Теменная
Височная
2
8
Скуловые
Верхняя челюсть
5
6
Непарные
Лобная
Затылочная
3
1
Носовая кость
Нижняя челюсть
4
7
43. Какие функции опорно-двигательного аппарата осуществляются благодаря подвижности нижней челюсти?
Какие функции опорно-двигательного аппарата осуществляются благодаря подвижности нижней челюсти?
44. Рассмотрите рисунок, на котором показаны осевой скелет и таз шимпанзе и человека. Ответьте на следующие вопросы.
1) Почему у шимпанзе позвоночник имеет два изгиба, а у человека – четыре?
2) Почему затылочная кость шимпанзе имеет мощные гребни, а человека их нет?
3) Чем объяснить то, что разница в массе шейных и поясничных позвонков человека больше, чем у шимпанзе?
-
Ответ: Человек прямоходящий, голову и спину держит прямо по ходьбе – отсюда большая нагрузка на позвонки и их массивность.
4) Почему таз у человека имеет форму чаши, а шимпанзе – нет?
45. На рисунке изображены шейный, грудной и поясничный позвонки. Общие части позвонков обозначены цифрами, отличительные признаки частей позвонков цифры и буквы, которыми они отмечены на рисунке:
-
Ответ:
Тело позвонка – 5;
Дуга – 2;
Отверстие, образующее с другими позвонками позвоночный канал, — 3;
Остистый отросток – 1;
Поперечный отросток – 4;
Отверстия шейных позвонков, через которые проходят в голову кровеносные сосуды, — А;
Суставные площадки на теле грудных позвонков и поперечных отростков, сочленяющиеся с рёбрами,- Б;
Имеются ли эти признаки у поясничных позвонков?
46. Назовите показанные на рисунке кости.
Назовите показанные на рисунке кости.
-
Ответ:
1 – Череп;
2 – Шейные позвонки;
3 – Ключица;
4 – Лопатки;
5 – Грудина;
6 – Рёбра;
7 – Плечевая кость;
8 – Лучевая кость;
9 – Локтевая кость;
10 – Таз;
11 – Копчик;
12 – Головка бедренной кости;
47. Ответьте на вопросы.
1) Сколько пар рёбер соединяются с грудиной с помощью хрящей?
2) Сколько пар рёбер заканчивается свободно?
3) Какую функцию выполняют хрящевые диски между позвонками?
-
Ответ: Они придают позвоночнику подвижность, упругость, смягчают сотрясение при беге, ходьбе, прыжках.
48. Какие структуры образуют грудную полость человека? Вспомните, у какой группы организмов впервые в процессе эволюции появилась грудная полость такого строения. Какие органы расположены в грудной полости?
Какие органы расположены в грудной полости?
-
Ответ: Грудной отдел позвоночника, ребра и грудина. Впервые подобная грудная полость появилась у пресмыкающихся. В грудной полости находятся легкие и сердце.
49. Напишите названия указанных на рисунке частей сустава.
50. Сделайте подписи к рисункам.
-
Ответ:
I. Кости верхней конечности:
1 – Ключица
2 – Лопатка
3 – Плечевая кость
4 – Лучевая кость
5 – Локтевая кость
6 – Запястье
7 – Пястье
8 – Фаланги пальцев
II. Кости нижней конечности и крестец:
1 – Таз
2 – Крестец
3 – Бедренная кость
4 – Большая берцовая кость
5 – Малая берцовая кость
6 – Предплюсна
7 – Плюсна
8 – Фаланги пальцев
51. Допишите утверждения.
Допишите утверждения.
-
Ответ: Кисть от стопы отличается подвижными и длинными фалангами (кисть можно собрать в горсть!) и противопоставлением большого пальца всем остальным. Стопа представлена мощными костями таранной и пяточной, короткими пальцами, большой палец не противопоставлен всем остальным. Своды стопы позволяют человеку пружинить при ходьбе.
52. Повторите статью «Разновидности мышечной ткани» (§8). Нарисуйте клетку гладкой и клетку поперечнополосатой мышечной ткани.
Отметьте особенности поперечнополосатой мышечной ткани:
53. Рассмотрите рисунок 40 в учебнике. Найдите брюшко и сухожилия двуглавой мышцы. Прочитав статьи «Макроскопическое строение мышц» и «Движения в суставах» (§13), впишите определяемые ниже понятия.
-
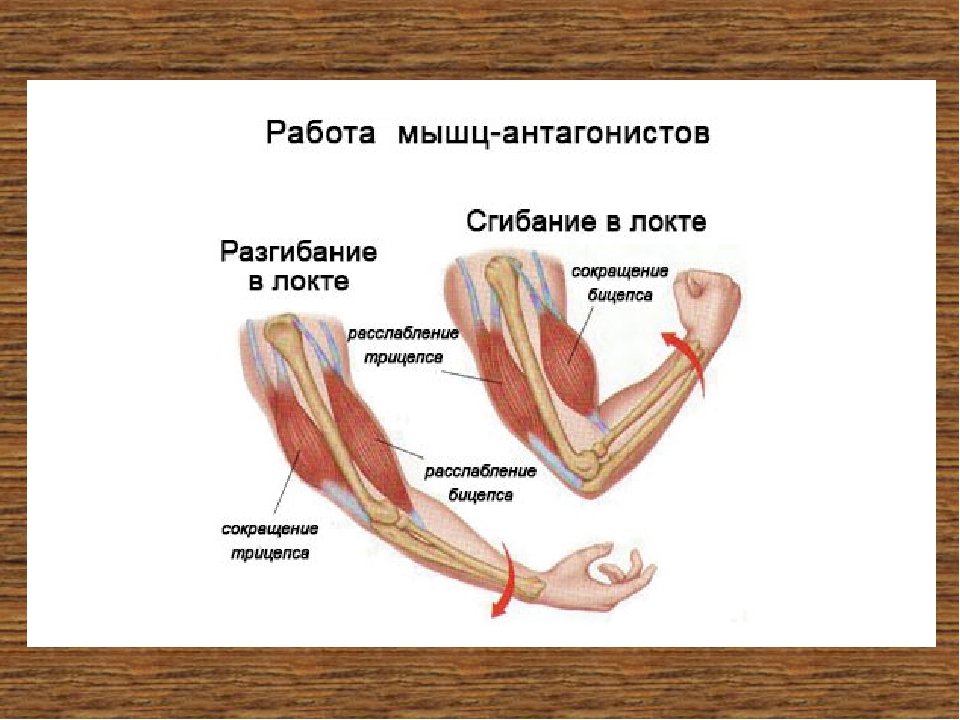
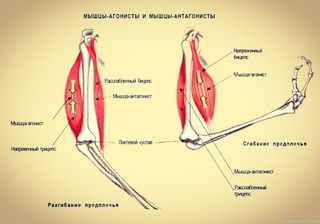
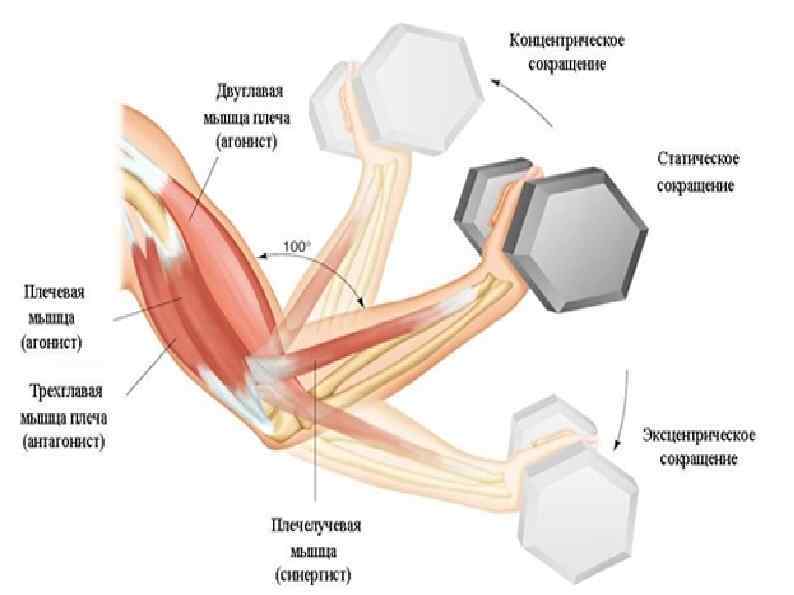
Ответ: Сухожилие, прикрепляющееся к участку кости, которая чаще всего остаётся неподвижной, называется
 Мышцы противоположного действия называются антагонистами, а мышцы действующие в одном направлении, — синергистами.
Мышцы противоположного действия называются антагонистами, а мышцы действующие в одном направлении, — синергистами.
54. Выполните лабораторную работу «Мышцы человеческого тела» (с. 86 – 89 учебника) и, изучив рисунки 41 и 42, заполните правый столбец таблицы.
-
Ответ:
Функции мышцы или мышечной группы
Название мышцы или мышечной группы
Мышцы, одним концом прикрепляющиеся к костям черепа, а другими к коже или только к коже.
Мимические.
Мышцы, приводящие в движение нижнюю челюсть.
Височные.
Мышца шеи и спины, откидывающая назад голову.
Трапециевидная.
Мышцы, наклоняющие голову вперёд, когда сокращаются вместе, и наклоняющие голову в стороны, когда сокращаются поодиночке.
Грудино-ключичнососцевидные.
Мышца спины, отводящая руки за спину.
Широчайшая мышца.
Мышцы груди, сводящие плечи и руки, согнутые в локте, вперёд.
Большая грудная.
Мышцы, обеспечивающие расширение и спадение грудной клетки.
Межреберные.
Мышцы, поднимающие туловище из положения лёжа в положение сидя при фиксированных ногах.
Брюшной пресс.
Мышцы, обеспечивающие прямохождение.
Глубокие и ягодичные.
Мышца руки, отводящая плечо в сторону
Дельтовидная.
Мышцы, сгибающие пальцы и кисть в кулак.
Мышцы предплечья.

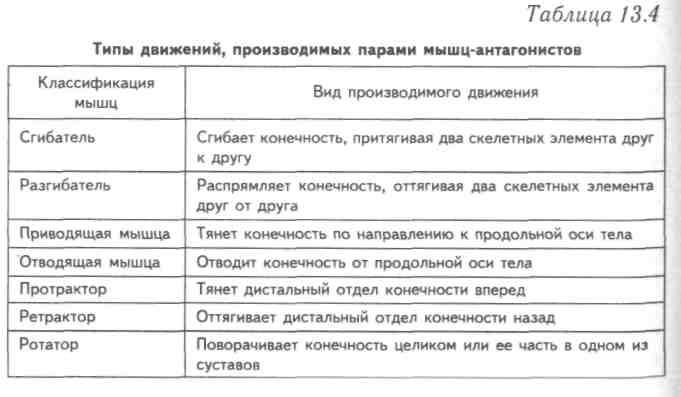
55. Приведите несколько примеров мышц, работающих как антагонисты и как синергисты.
-
Ответ:
Антагонисты
Синергисты
Бицепс — трицепс
Грудь — трицепс
56. На рисунке показан мотонейрон. Подпишите его части.
Укажите, сколькими мышечными волокнами управляет мотонейрон, показанный на рисунке.
57. Напишите, как надо организовать тренировку, чтобы был тренировочный эффект (синтез в мышечной ткани преобладал над распадом).
58. Укажите последствия гиподинамии.
-
Ответ: Изменения в мышцах: становятся дряблыми, теряют силу. Изменения в костях: уходят соли кальция. Изменения в крови: соли кальция соединяются с холестерином образуя бляшки.
59. Прочитайте статью «Регуляция работы мышц-антагонистов» (§14) и заполните таблицу, вписав в соответствующие столбцы слова: возбуждены, заторможены, сокращены, расслаблены.
-
Ответ:
Виды нервных
центров
и мышц
Состояние рук
Нервные центры сгибателей
Нервные центры разгибателей
Мышцы сгибатели
Мышцы разгибатели
Сгибание руки в локтевом суставе
Возбуждены
Заторможены
Сокращены
Расслаблены
Разгибание руки в локтевом суставе
Заторможены
Возбуждены
Расслаблены
Сокращены
Удержание груза в локтевом суставе
Возбуждены
Возбуждены
Сокращены
Сокращены
Рука опущена, нагрузка отсутствует
Заторможены
Заторможены
Расслаблены
Расслаблены
60. Впишите определяемые ниже понятия.
Впишите определяемые ниже понятия.
-
Ответ: Работа, связанная с перемещением тела или груза, называется динамическая. Работа, связанная с сохранением позы или удержанием груза, называется статическая.
61. Отчёт о лабораторной работе «Утомление при статической работе» (с. 94 учебника).
-
Ответ:
Фазы утомления
Признаки томления
Время
Начало опыта (отсутствие утомления)
Рука с грузом неподвижна
I фаза утомления
Рука опускается затем рывком поднимается на прежнее место.
II фаза утомления
Дрожание рук, потеря координации, пошатывание корпуса, покраснение лица, потоотделение.
III фаза утомления (предельное утомление)
Боль в мышцах, онемение, непроизвольное расслабление мышц.

62. Прочитайте §15 «Осанка. Предупреждение плоскостопия». Перечислите причины нарушения осанки.
-
Ответ: Неправильная посадка за столом, сутулиться (когда при ходьбе голова смотрит в землю). Привычка горбиться.
63. Назовите факторы, способствующие развитию плоскостопия.
64. Отчёт о лабораторной работе «Осанка и плоскостопие» (с. 98 учебника).
Выявление нарушений осанки.
1. Выявление сутулости.
-
Ответ:
Расстояние между левым и правым плечом
А/ Б
Со стороны груди (А)
Со стороны (Б)
55
50
1,1
Вывод: Нарушений нет.
2. Выявление нарушений в области поясничного изгиба.
-
Ответ: При положении тела спиной к стене, когда, пятки, голени, таз и лопатки касаются стены, между стеной и поясницей проходит (кулак или только ладонь) ладонь.
Вывод: Нарушений нет.
II. Выявление плоскостопия (работа выполняется дома).
-
Ответ: След в узкой своей части (заходит или не заходит) не заходит за линию, соединяющую центр пятки и центр третьего пальца (см. рис. 45 на с. 97 учебника).
Вывод: Нарушений нет.
65. Прочитайте § 16 «Первая помощь при ушибах, переломах костей и вывихах суставов», рассмотрите рисунки и заполните таблицу.
-
Ответ:
Тип травмы
Растяжение связок.
Перелом
Вывих
Меры первой помощи
Наложить тугую фиксирующую повязку и к пораженному месту прикладывают холод.
При открытых переломах – остановить кровотечение. Во всех случаях, кроме перелома костей грудной клетки, прикладывают шину. При травме позвоночника положить лицом вниз на ровный настил.
Обеспечить неподвижность.
Противопоказания
Тепло;
Движение;
Массаж.
Пытаться придать естественную форму, накладывая шину на грудную клетку.
Пытаться вправить кость.
66. При подозрении на перелом костей предплечья была наложена шина, а сам рука зафиксирована с помощью косынки.
Определите, какую ошибку допустили при оказании помощи пострадавшему. К каким последствиям она может привести?
-
Ответ: Шина не зафиксирована. Она может сместиться, в результате чего может сместиться сломанная кость. Это может привести к повреждению тканей, сосудов и др.
67. Оцените, что нового вы узнали при изучении этой темы. Предположите. Как могут пригодиться вам эти знания в повседневной жизни.
-
Ответ: При необходимости я могу, не навредив, оказать первую помощь, пока скорая помощь будет в пути.
68. Решите кроссворд №4.
-
Ответ:
По горизонтали:
1. Кость осевого скелета.
2. Кость плечевого пояса, лежащая среди мышц спины.
6. Мышцы однонаправленного действия.
7. Твёрдая основа тела, состоящая из костей, хрящей и связок.
11. Часть грудной клетки.
12. Болезнь позвоночника.
14. Часть кости, обеспечивающая её рост в толщину.
По вертикали:
1- Болезнь ног.
2- Элемент скелета.
4- Мышцы, действующие в противоположных направлениях.
5- Травма костей.
8- Кость плечевого пояса, сочленяющаяся с грудиной.
9- Часть мышцы, соединяющая её брюшко с костью.
10- Название пояса нижних конечностей.
13- Вид травмы.
Инструктивные карточки к лабораторным работам по анатомии физиологии и гигиене человека — Лабораторная работа
Инструктивные карточки
к лабораторным работам
по анатомии, физиологии и
гигиене человека
( по программе В.В. Пасечника)
Разработала учитель биологии
МОУ СОШ № 32
г. Череповца
Вологодской области
Череповца
Вологодской области
Григорьева Наталия Владимировна
Лабораторная работа №1
Рассматривание клеток и тканей в оптический микроскоп
Цель:
Оборудование: раствор йода, покровное и предметное стекло, микроскоп, готовый препарат эпителиальной ткани.
Охрана труда. При работе с микропрепаратами быть осторожным Аккуратно обращаться с микроскопом. При переводе объектива на большое увеличение осторожно работать с винтом, чтобы не раздавить микропрепарат.
Ход работы
1. Приготовьте временный препарат эпителиальной ткани. С помощью индивидуальной зубочистки соскоблите слизь с внутренней поверхности щеки. Полученный комочек слизи поместите на предметное стекло в каплю красящего раствора и накройте покровным стеклом.
2. Рассмотрите
препарат при большом увеличении
микроскопа. Обратите внимание на форму
клеток, контуры оболочки (мембрану),
цитоплазму, ядро, расположение клеток
относительно друг друга.
Рассмотрите
препарат при большом увеличении
микроскопа. Обратите внимание на форму
клеток, контуры оболочки (мембрану),
цитоплазму, ядро, расположение клеток
относительно друг друга.
3.Сравните приготовленный временный препарат с постоянным препаратом эпителиальной ткани.
4.Сделайте рисунок клеток эпителиальной ткани.
Вывод.
* Какие функции выполняет эпителиальная ткань, благодаря каким
особенностям строения?
Лабораторная работ № 3 Самонаблюдение мигательного рефлекса и условия его проявления Цель:
Охрана труда. Тем, кто носит очки, предлагаем на время опыта их снять.
Опыт проводить чистыми руками, использование карандашей и других
предметов для раздражения кожи
и век недопустимо.
Ход работы
1.Осторожно прикоснитесь рукой
а) к наружному углу глаза,
б) к внутреннему углу глаза,
в) к ресницам,
г) к бровям,
д) к коже век,
Отметьте те области, раздражение которых вызывает непроизвольное
мигание, знаком «+»
2.Результаты оформите в виде таблицы.
Область | Непроизвольное мигание |
3.Почему не наблюдается мигательный рефлекс при раздражении
некоторых областей?
4. Многократно прикасайтесь к внутреннему углу глаза со стороны носа, не задевая ресниц. Что происходит?
5. Составьте схему рефлекторной
дуги мигательного рефлекса.
Составьте схему рефлекторной
дуги мигательного рефлекса.
Укажите функцию каждого звена. Вывод:
Какое значение имеет мигательный рефлекс?
Это рефлекс условный или безусловный
* Лабораторная работ № 3 Самонаблюдение мигательного рефлекса и условия его проявления Цель:
Охрана труда. Тем, кто носит очки, предлагаем на время опыта их снять.
Опыт проводить чистыми руками, использование карандашей и других
предметов для раздражения кожи и век недопустимо.
Ход работы
1.Осторожно прикоснитесь рукой
а) к наружному углу глаза,
б) к внутреннему углу глаза,
в) к ресницам,
г) к бровям,
д) к коже век,
Отметьте те области, раздражение которых вызывает непроизвольное
мигание, знаком «+»
2. Результаты оформите в виде
таблицы.
Результаты оформите в виде
таблицы.
3.Почему не наблюдается мигательный рефлекс при раздражении
некоторых областей?
4. Многократно прикасайтесь к внутреннему углу глаза со стороны носа, не задевая ресниц. Что происходит?
5.Составьте схему рефлекторной дуги мигательного рефлекса.
Укажите функцию каждого звена. Вывод:
Какое значение имеет мигательный рефлекс?
Это рефлекс условный или безусловный?
Лабораторная работа № 4
Коленный рефлекс Цель:
Пояснение.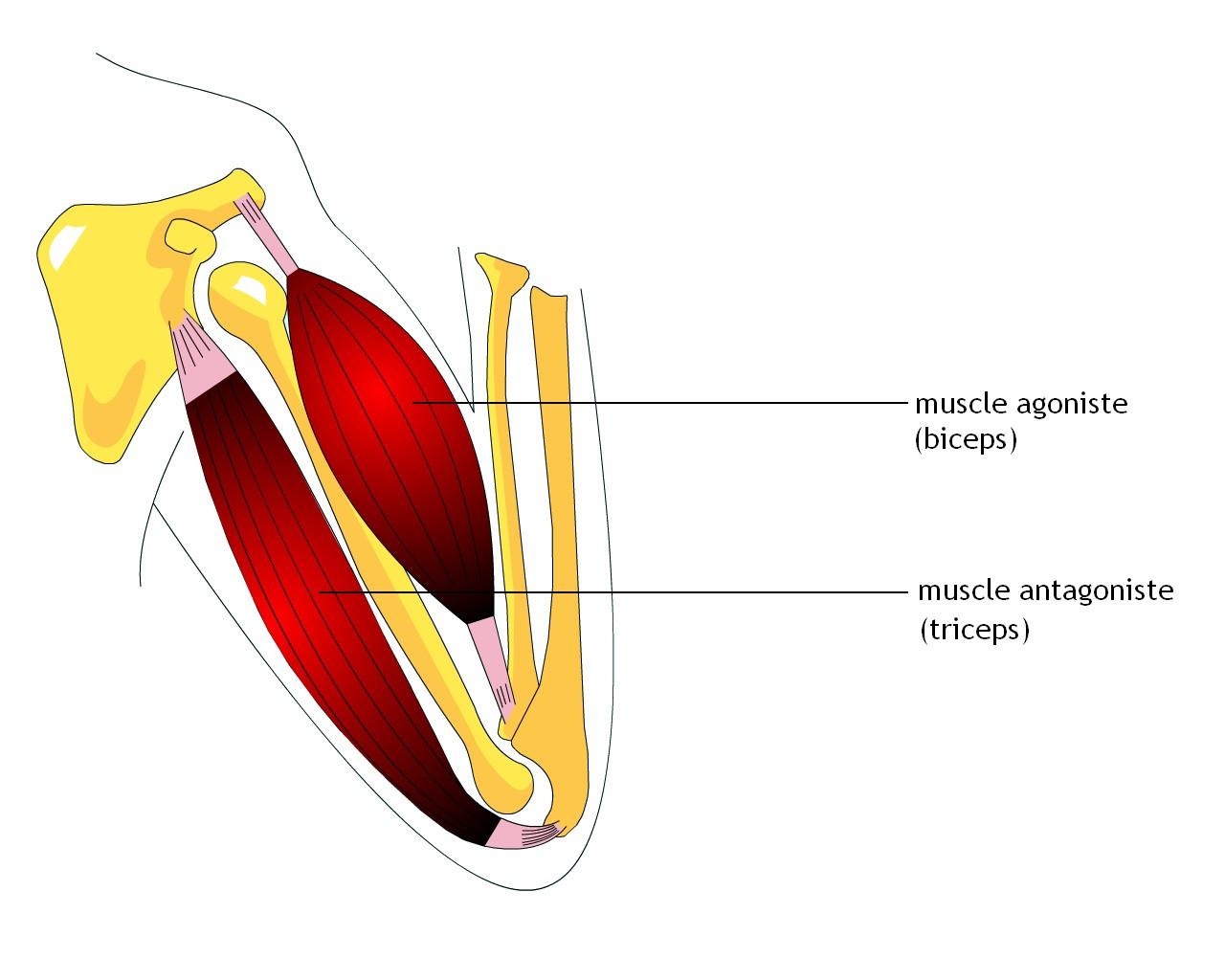 Коленный
рефлекс – это рефлекс, центр которого
находится в спинном мозге. Он проявляется
у всех здоровых людей. В условиях данного
опыта он
никакого биологического
значения не имеет. Его
используют врачи для определения
функций спинного мозга: при заболевании
они могут быть нарушены.
Коленный
рефлекс – это рефлекс, центр которого
находится в спинном мозге. Он проявляется
у всех здоровых людей. В условиях данного
опыта он
никакого биологического
значения не имеет. Его
используют врачи для определения
функций спинного мозга: при заболевании
они могут быть нарушены.
Ход работы
1.Посадите испытуемого на стул. Предложите ему перекинуть ногу на ногу,
затем ударьте ребром руки по сухожилию четырехглавой мышцы под коленной чашечкой.
2.Запишите свои наблюдения. 3.Составьте схему рефлекторной дуги коленного рефлекса.
Укажите функцию каждого звена. Вывод:
Вывод:
Лабораторная работа №5
Микроскопическое строение кости
Цель:
Ход работы
1.Рассмотрите микроскопическое строение кости. Ответьте на вопросы.
а) К какому типу ткани относится костная ткань?
б) Какие признаки характерны для данного типа ткани?
в) Какими свойствами обладает межклеточное вещество костной ткани?
2. Рассмотрите макроскопическое строение кости. Зарисуйте кость в разрезе, подпишите ее части: надкостницу, компактное вещество, губчатое вещество, красный костный мозг, желтый костный мозг.
Вывод. Какими свойствами обладают кости?
Какие особенности строения
обуславливают эти свойства.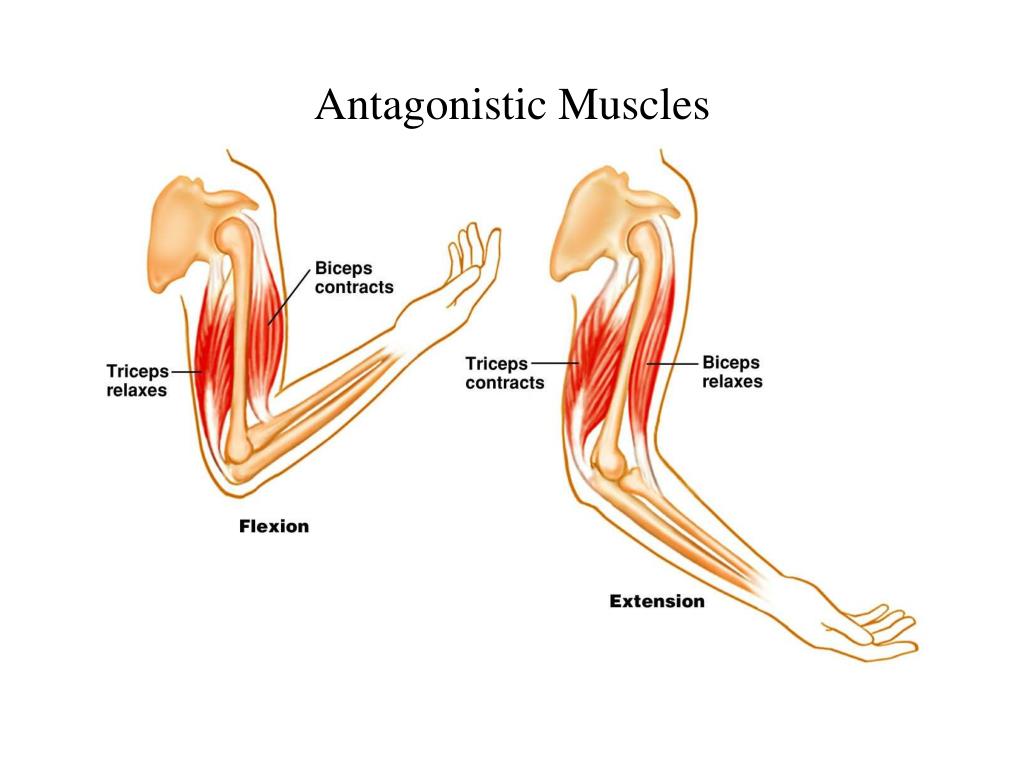
Лабораторная работа №6
Мышцы человеческого тела Цель:
Оборудование: учебник
Ход работы
1.Используя учебник и анатомическое описание (стр. учебника 65-68),
определите местоположение
мышечных групп и выполняемые ими
движения.
2.Результаты оформите в таблице.
Название мышцы | Местоположение | Функция |
* 3.Приведите несколько примеров мышц, работающих как
синергисты и как антагонисты
Антагонисты | Синергисты |
4.Вывод:
* Лабораторная работа №6
Мышцы человеческого тела Цель:
Оборудование: учебник
Ход
работы 1. Используя учебник и анатомическое
описание (стр. учебника 65-68),
Используя учебник и анатомическое
описание (стр. учебника 65-68),
определите местоположение мышечных групп и выполняемые ими
движения. 2.Результаты оформите в виде таблицы. * 3.Приведите несколько примеров мышц, работающих как
синергисты и как антагонисты
Антагонисты | Синергисты |
4.Вывод:
Лабораторная работа №7
Утомление при статической и динамической работе
Цель:
Оборудование: секундомер,
груз массой 1,5 и 3 кг.
Ход работы. Используя текст учебника (стр. 71),выясните различие между статической и динамической работой.
Опыт 1.Утомление при статической работе. 1.Испытуемый берет груз массой 1,5 кг, и держит его в руке, отведенной в сторону под прямым углом к туловищу. На уровне вытянутой руки
сделайте на доске отметку мелом и включите секундомер.
Наблюдайте, за какое время произойдет утомление мышц.
2.Испытуемый берет груз массой 3 кг, повторяет опыт.
Опыт 2 Утомление при динамической работе
1. Испытуемый поднимает тот же груз(1,5 кг,3кг) до сделанной метки и
опускает его. Наблюдайте, за
какое время произойдет утомление.
Наблюдайте, за
какое время произойдет утомление.
2. Результаты оформите в таблице.
Работа | Нагрузка 1,5 кг | Нагрузка 3 кг |
Статическая | ||
Динамическая |
Вывод:
Как нагрузка влияет на развитие утомления мышц?
Какая работа более утомительна?
* Почему при стирке белья спина устает больше, чем руки?
*Как лучше нести груз: без отдыха попеременно правой и левой рукой,
или одной правой, а потом, отдохнув минуту, груз снова нести в этой же руке?
* Лабораторная работа №7
Утомление при статической и динамической работе
Цель:
Оборудование: секундомер,
груз массой 1,5 и 3 кг.
Ход работы. Используя текст учебника (стр. 71),выясните различие между статической и динамической работой.
Опыт 1.Утомление при статической работе. 1.Испытуемый берет груз массой 1,5 кг и держит его в руке, отведенной в сторону под прямым углом к туловищу. На уровне вытянутой руки сделайте на доске отметку мелом и включите секундомер.
Наблюдайте, за какое время произойдет утомление мышц.
2.Испытуемый берет груз массой 3 кг, повторяет опыт.
Опыт 2 Утомление при динамической работе.
1. Испытуемый поднимает тот же груз(1,5 кг, 3 кг) до сделанной метки и
опускает его. Наблюдайте, за
какое время произойдет утомление
Наблюдайте, за
какое время произойдет утомление
2. Результаты оформите в таблице.
Вывод:
*Как лучше нести груз: без отдыха попеременно правой и левой рукой,
или одной правой, а потом, отдохнув минуту, груз снова нести в этой же руке?
* Почему при стирке белья спина устает больше, чем руки?
Лабораторная работа №8
Выявление нарушения осанки Цель:
Оборудование: сантиметровая лента
Ход работы.
Опыт№1
1.Для выявления сутулости сантиметровой лентой измерьте расстояние
между самыми отдаленными точками левого и правого плеча со стороны
А) груди,
Б) спины.
2.Первый результат разделите на второй.
3.Оформите в виде таблицы.
Расстояние между левым и правым плечом со стороны | А/Б | |
груди (А) | спины (Б) | |
4.Оцените результаты.
Если получается число, близкое к единице или больше, значит,
нарушений нет. Получение числа меньше единицы говорит о нарушении
осанки.
Опыт№2.
Встаньте спиной к стене так, чтобы пятки, голени, таз и лопатки
касались стены. Попробуйте между стеной и поясницей просунуть кулак.
Если он проходит — нарушение осанки есть. Если проходит только ладонь-
осанка нормальная.
Вывод: Есть ли у вас нарушение осанки?
Если есть, предположите, каковы причины?
Лабораторная работа №9
Выявление плоскостопия Цель:
Оборудование: таз с водой, лист бумаги, простой карандаш.
Ход работы.
Мокрой ногой встаньте на лист бумаги. Контуры следа обведите простым карандашом. Найдите центр пятки и центр третьего пальца.
Соедините две найденные точки
прямой линией. Оцените свои
результаты. Если в узкой
части след не заходит за
линию — плоскостопия нет
(стр.74,рис39).
Вывод: Есть ли у вас плоскостопие?
Если есть, предположите, каковы его причины?
Предложите 3-4 упражнения, предупреждающих плоскостопие.
Лабораторная работа №10
Роль плечевого пояса в движении руки
Цель:
Ход работы.
1. В положении сидя найдите нижний угол левой лопатки. Отведите левую руку в сторону до горизонтального уровня. Происходит ли смещение лопатки?
2. Продолжите движение руки до верхнего вертикального положения. Происходит ли смещение лопатки?
3. Положите руку на левую ключицу и повторите движение. В каком случае происходит движение ключицы?
Вывод: Какова роль плечевого пояса в движении руки?
Лабораторная работа №11
Рассматривание клеток крови человека и лягушки под микроскопом Цель:
Оборудование: готовые
микропрепараты крови человека и лягушки,
микроскоп.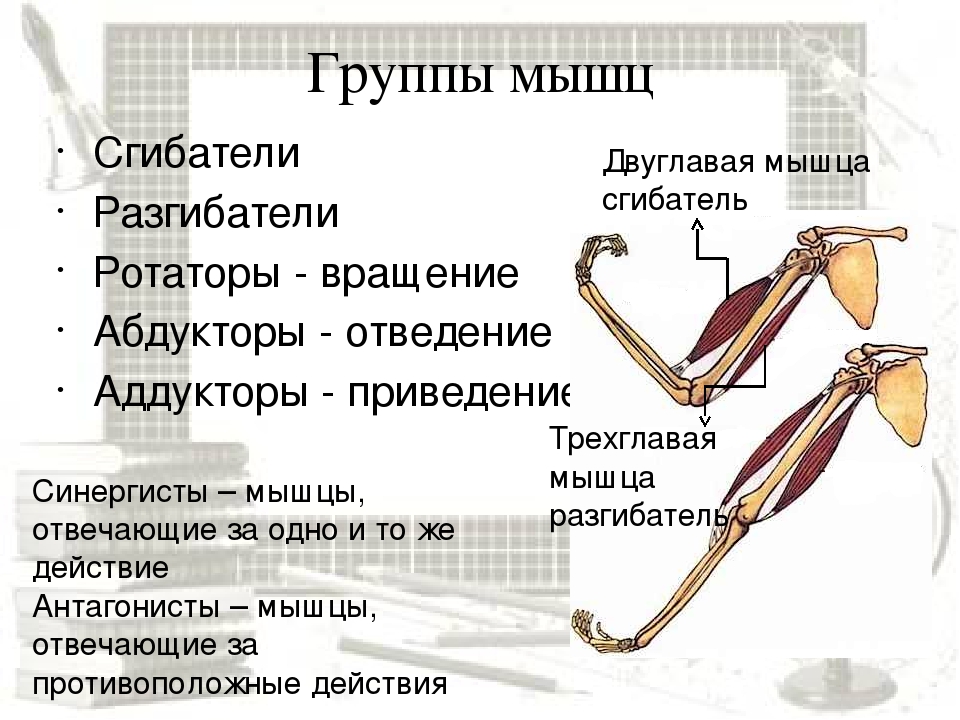 Охрана
труда. При
работе с микропрепаратами быть осторожным
Аккуратно обращаться с микроскопом.
При переводе объектива на большое
увеличение осторожно работать с винтом,
чтобы не раздавить микропрепарат.
Охрана
труда. При
работе с микропрепаратами быть осторожным
Аккуратно обращаться с микроскопом.
При переводе объектива на большое
увеличение осторожно работать с винтом,
чтобы не раздавить микропрепарат.
Ход работы
1.Рассмотреть и зарисовать эритроциты крови человека и лягушки (вид сбоку и спереди).
2.Сравнить эритроциты человека и лягушки: цвет, размеры, наличие ядра, форма, количество (приблизительно) в поле зрения микроскопа.
3.Результаты оформите в таблице.
Признаки | Эритроциты лягушки | Эритроциты человека |
Вывод.
Каковы черты сходства и различия
в строении эритроцитов человека и
лягушки?
* Объясните связь строения эритроцитов
человека и лягушки с выполняемой ими
функцией. * Чья кровь- человека или лягушки —
переносит больше кислорода. Почему?
* В каком направлении шла
эволюция эритроцитов позвоночных
животных?
* Чья кровь- человека или лягушки —
переносит больше кислорода. Почему?
* В каком направлении шла
эволюция эритроцитов позвоночных
животных?
* Лабораторная работа №11
Рассматривание клеток крови человека и лягушки под микроскопом Цель: Оборудование: готовые микропрепараты крови человека и лягушки, микроскоп. Охрана труда. При работе с микропрепаратами быть осторожным Аккуратно обращаться с микроскопом. При переводе объектива на большое увеличение осторожно работать с винтом, чтобы не раздавить микропрепарат.
Ход работы
1. Рассмотреть
и зарисовать эритроциты крови человека
и лягушки (вид сбоку и спереди).
Рассмотреть
и зарисовать эритроциты крови человека
и лягушки (вид сбоку и спереди).
2.Сравнить эритроциты человека и лягушки: цвет, размеры, наличие ядра, форма, количество (приблизительно) в поле зрения микроскопа.
3.Результаты оформите в таблице.
Вывод.
Каковы черты сходства и различия в строении эритроцитов человека и лягушки? * Объясните связь строения эритроцитов человека и лягушки с выполняемой ими функцией. * Чья кровь- человека или лягушки — переносит больше кислорода. Почему? * В каком направлении шла эволюция эритроцитов позвоночных животных?
Лабораторная работа №12
Функции венозных клапанов Цель:
Пояснение. Если
рука опущена, венозные клапаны не дают
крови стечь вниз. Клапаны раскрываются
лишь после того, как в ниже лежащих
сегментах накопится
достаточное количество крови, чтобы
открыть венозный клапан
и пропустить кровь вверх, в следующий
сегмент. Поэтому вены, по
которым кровь движется
против силы тяжести,
всегда набухшие.
Если
рука опущена, венозные клапаны не дают
крови стечь вниз. Клапаны раскрываются
лишь после того, как в ниже лежащих
сегментах накопится
достаточное количество крови, чтобы
открыть венозный клапан
и пропустить кровь вверх, в следующий
сегмент. Поэтому вены, по
которым кровь движется
против силы тяжести,
всегда набухшие.
Ход работы. 1.Поднимите одну руку вверх, а вторую опустите вниз. Спустя минуту
положите обе руки на стол. Наблюдения запишите в тетрадь.
Вывод.
Почему поднятая рука побледнела, а опущенная – покраснела?
В какой руке венозные клапаны были закрыты?
Лабораторная работа №13
Изменения в тканях при перетяжках, затрудняющих
кровообращение. Цель:
Оборудование: аптечное
резиновое кольцо или нитки. Пояснение. Перетяжка конечности затрудняет отток
крови по венам и лимфы по лимфатическим
сосудам. Расширение кровеносных капилляров
и вен приводит к покраснению,
а затем к посинению части органа,
изолированной перетяжкой. В дальнейшем
эта часть органа становится белой
из-за выхода плазмы крови в межклеточное
пространство. Тканевая жидкость
накапливается, сдавливая клетки. Орган
становится плотным на
ощупь. Начинается кислородное голодание
тканей, которое ощущается как
«ползание мурашек», покалывание. Работа
рецепторов нарушается.
Цель:
Оборудование: аптечное
резиновое кольцо или нитки. Пояснение. Перетяжка конечности затрудняет отток
крови по венам и лимфы по лимфатическим
сосудам. Расширение кровеносных капилляров
и вен приводит к покраснению,
а затем к посинению части органа,
изолированной перетяжкой. В дальнейшем
эта часть органа становится белой
из-за выхода плазмы крови в межклеточное
пространство. Тканевая жидкость
накапливается, сдавливая клетки. Орган
становится плотным на
ощупь. Начинается кислородное голодание
тканей, которое ощущается как
«ползание мурашек», покалывание. Работа
рецепторов нарушается.
Ход работы. 1.Накрутите на палец
резиновое кольцо или перетяните палец
ниткой. Обратите внимание
на изменение цвета и формы пальца.
1.Накрутите на палец
резиновое кольцо или перетяните палец
ниткой. Обратите внимание
на изменение цвета и формы пальца.
Наблюдения запишите в тетрадь. 2.Дотроньтесь перетянутым пальцем до какого-нибудь предмета. Наблюдения запишите в тетрадь. 3. Снимите перетяжку и помассируйте палец по направлению к сердцу. Что достигается этим приемом? Вывод.
Почему при перетяжке изменялся цвет пальца?
Почему ткани пальца уплотнены?
Как проявляются признаки кислородной недостаточности? Почему нарушена чувствительность?
*Почему вредно туго затягиваться ремнем, носить тесную одежду?
Лабораторная работа №14
Определение скорости
кровотока в капиллярах ногтевого ложа
Цель:
Оборудование: линейка,
секундомер или часы с секундной стрелкой.
Ход работы 1.Измерьте длину ногтя большого пальца от корня до места, где кончается его розовая часть, и начинается прозрачный ноготь, который обычно срезается (см. стр. 117, рис.56.А). Запишите результат.
Ботулотоксин в косметологии принципы подбора доз препарата Диспорт
Принципы расчета дозировок препаратов ботулинического нейропротеина типа А остаются дискутабельными, специалисты пока не нашли единого решения
Все согласны с тем, что необходим строго индивидуальный подход. Но на чем он должен быть основан? Что нужно принимать во внимание? И почему так важно работать, сохраняя баланс между мышцами-антагонистами?
Ботулинотерапия с помощью препаратов ботулинического токсина типа А (БТ типа А) является распространенным методом лечения ряда патологических состояний, вызванных спазмом скелетных мышц и их гиперкинезами.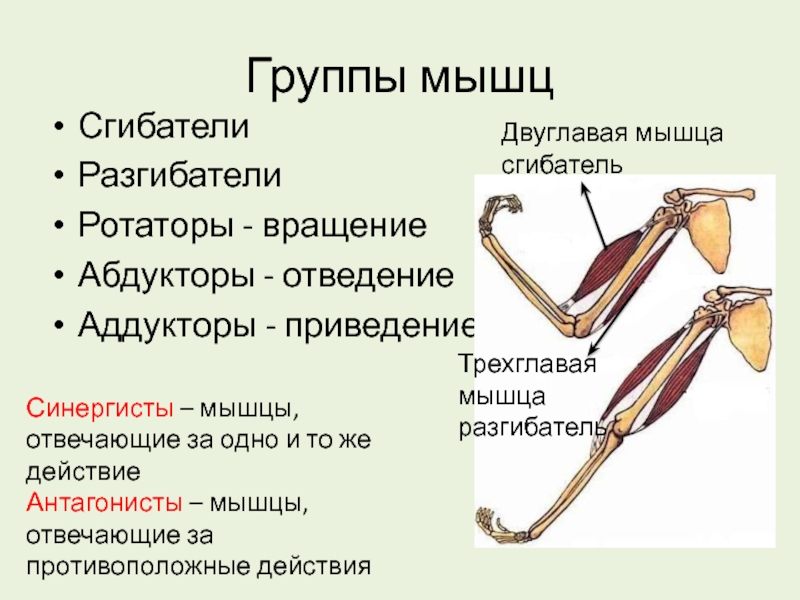 Ее применяют и для коррекции эстетических проблем, связанных с гиперактивностью мимических и жевательных мышц лица и шеи. Расчет дозировок для каждой мышцы у конкретного пациента производится с учетом множества составляющих и может представлять определенные трудности. Так, препараты БТ типа А разных производителей имеют специфические особенности, связанные с технологией производства и исключающие возможность унифицирования содержащихся в них активных единиц. Однако в связи со множеством факторов, сопряженных с особенностями мимики и архитектоники лица конкретного пациента, часто возникают трудности при расчете индивидуализированных дозировок ботулинического нейропротеина для получения управляемых клинических эффектов.
Ее применяют и для коррекции эстетических проблем, связанных с гиперактивностью мимических и жевательных мышц лица и шеи. Расчет дозировок для каждой мышцы у конкретного пациента производится с учетом множества составляющих и может представлять определенные трудности. Так, препараты БТ типа А разных производителей имеют специфические особенности, связанные с технологией производства и исключающие возможность унифицирования содержащихся в них активных единиц. Однако в связи со множеством факторов, сопряженных с особенностями мимики и архитектоники лица конкретного пациента, часто возникают трудности при расчете индивидуализированных дозировок ботулинического нейропротеина для получения управляемых клинических эффектов.
Индивидуализация подхода
Мимическая активность человека является индивидуальной, и ее коррекция требует такого же индивидуального подбора дозировок БТ типа А. Необходимость в изменении рекомендованной дозы также может возникать с течением времени у одного и того же пациента, поскольку ботулинотерапия способствует изменению его мимического паттерна (стереотипа мимической активности) и гармонизирует мимику в долгосрочной перспективе.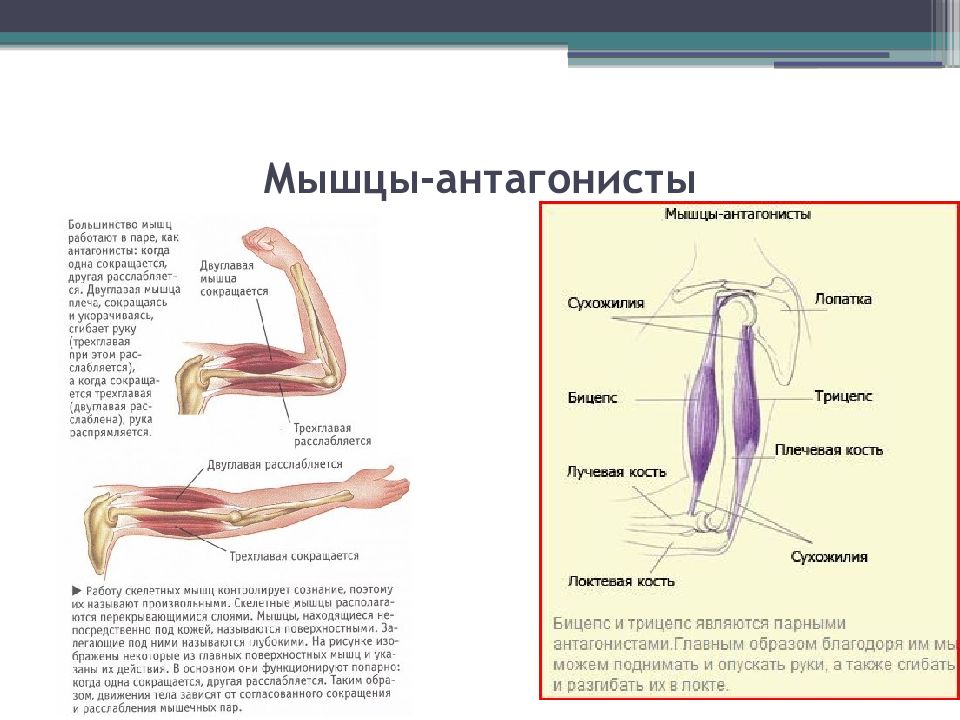 Потому и сформировалась общепринятая стратегия ботулинотерапии, нацеленная и на ближайший, и на отдаленный эстетический результат. Эта стратегия подразумевает также подбор и смену индивидуальных доз БТ типа А сообразно меняющемуся мимическому образу пациента.
Потому и сформировалась общепринятая стратегия ботулинотерапии, нацеленная и на ближайший, и на отдаленный эстетический результат. Эта стратегия подразумевает также подбор и смену индивидуальных доз БТ типа А сообразно меняющемуся мимическому образу пациента.
Отклонение в разумных пределах индивидуальной дозы БТ типа А от среднестатистической рекомендуемой – обычная клиническая практика. Эта коррекция должна быть обоснована: следует помнить, что при лечении антагонистических групп не всегда легко найти баланс, поскольку даже незначительный просчет в выборе дозировок может привести к нежелательной компенсаторной активации мимических мышц, неудовлетворительным клиническим результатам и даже нежелательным явлениям.
Еще одним весьма заметным препятствием при выборе адекватной дозы может стать региональный подход в коррекции мимических мышц лица без учета единой и взаимосвязанной двигательной активности всей группы мышц лица и шеи. Не существует мимических мышц, работающих в изоляции. Все они связаны с ближайшими и отдаленными «партнерами» синергическими или антагонистическими взаимоотношениями, которые могут быть и прямыми, и SMAS-опосредованными (SMAS – поверхностная мышечно-апоневротическая система). При любом типе взаимоотношений мимические мышцы образуют разнообразные кинематические цепи и функционируют как единая биомеханическая система. Недооценка роли этой системы при проведении ботулинотерапии и прогнозировании ее ближайшего и отдаленного клинического результата недопустима.
Все они связаны с ближайшими и отдаленными «партнерами» синергическими или антагонистическими взаимоотношениями, которые могут быть и прямыми, и SMAS-опосредованными (SMAS – поверхностная мышечно-апоневротическая система). При любом типе взаимоотношений мимические мышцы образуют разнообразные кинематические цепи и функционируют как единая биомеханическая система. Недооценка роли этой системы при проведении ботулинотерапии и прогнозировании ее ближайшего и отдаленного клинического результата недопустима.
Теория баланса
На первый план в современной ботулинотерапии вышла теория баланса (Е. З. Парасагашвили, Е. А. Разумовская). Эта ключевая стратегия имеет своей целью обеспечить гармоничную работу единой биомеханической системы всех взаимодействующих друг с другом мимических мышц, а также поддерживать в краткосрочной и долгосрочной перспективе динамическое равновесие этой системы как в покое, так и в движении.
Рекомендуемые производителями абсолютные значения доз ботулинического нейропротеина для отдельных зон лица являются эффективными лишь при изолированной работе с отдельными мышцами или областями лица, но они должны подвергаться коррекции при одновременной работе с несколькими зонами или по методу Full Face.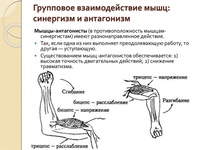 Потому рекомендуемые абсолютные значения доз не могут быть универсальными не только при применении препаратов БТ типа А от разных производителей и у разных пациентов, но и при применении одного и того же препарата в лечении одного и того же пациента. Таким образом, расчетные дозы ботулинического нейропротеина предполагают вариабельность в зависимости от решаемой эстетической задачи и при одновременной коррекции нескольких зон лица.
Потому рекомендуемые абсолютные значения доз не могут быть универсальными не только при применении препаратов БТ типа А от разных производителей и у разных пациентов, но и при применении одного и того же препарата в лечении одного и того же пациента. Таким образом, расчетные дозы ботулинического нейропротеина предполагают вариабельность в зависимости от решаемой эстетической задачи и при одновременной коррекции нескольких зон лица.
В настоящей статье представляется разумным дополнить рекомендации по применению абсолютных значений доз БТ типа А более универсальными относительными значениями. Возможность оперировать помимо абсолютных значений еще и относительными ни в коей мере не отрицает основополагающих принципов ботулинотерапии. Однако, как показывает личный клинический опыт автора, это может заметно улучшить прогноз ближайших и долгосрочных клинических результатов и управляемость ими.
Расчет относительных дозировок не только облегчает индивидуальный подбор доз, но и в соответствии с теорией баланса позволяет эффективно и деликатно сбалансировать единую биомеханическую систему лица с целью его гармонизации.
Прежде чем представить конкретный алгоритм расчета более универсальных относительных доз БТ типа А для получения управляемых клинических эффектов, рассмотрим, какие трудности этот метод позволяет преодолеть.
Для примера вспомним работу с управлением высотой расположения бровей.
Управление высотой расположения бровей
Бровями управляет в качестве леватора затылочно-лобная мышца, а точнее ее лобное брюшко – m. frontalis. Она тянет брови вверх и несколько в стороны, одновременно образуя горизонтальные морщины на лбу в местах наибольшего своего сокращения.
Анализ мы начнем с установления состава мышечной группы, в которой мышцы являются участниками одной и той же кинематической цепи. Известно, что только у части пациентов при коррекции глабеллы (с неактивной лобной мышцей и лишь на старте ботулинотерапии) возможно добиться нехирургического подъема бровей, избежав формирования морщин над бровями – процесса, сопутствующего их лифтингу.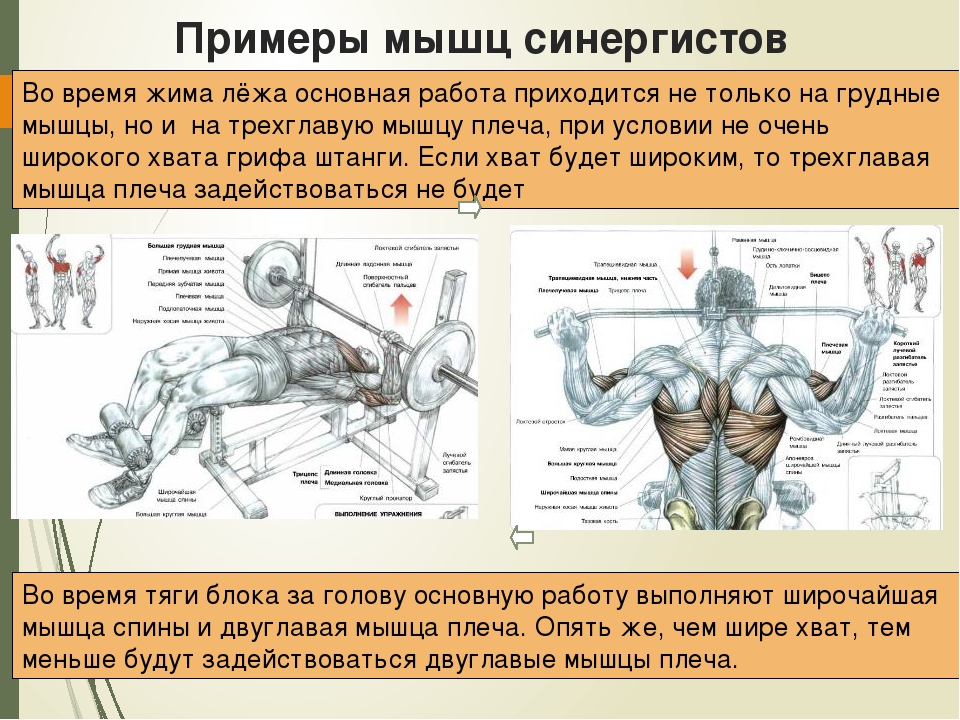 Но уже через 2–4 процедуры и у таких пациентов мы увидим компенсаторную активизацию m. frontalis с формированием горизонтальных морщин лба, особенно над бровями. Этот процесс, названный О. Р. Орловой активизацией «дремлющих мышц», – увы, неизбежная плата пациента за лифтинг бровей. Генез этих морщин – активизация оставшейся интактной лобной мышцы или ее латеральных порций. Эти мышечные волокна являются синергистами центральной порции m. frontalis и одновременно антагонистами (прямыми и непрямыми) глабеллярного комплекса. Таким образом, латеральные порции лобной мышцы из нормокинеза переходят в состояние гиперкинетической активности, компенсируя тем самым нарушенную функцию центральной порции лобной мышцы и глабеллы.
Но уже через 2–4 процедуры и у таких пациентов мы увидим компенсаторную активизацию m. frontalis с формированием горизонтальных морщин лба, особенно над бровями. Этот процесс, названный О. Р. Орловой активизацией «дремлющих мышц», – увы, неизбежная плата пациента за лифтинг бровей. Генез этих морщин – активизация оставшейся интактной лобной мышцы или ее латеральных порций. Эти мышечные волокна являются синергистами центральной порции m. frontalis и одновременно антагонистами (прямыми и непрямыми) глабеллярного комплекса. Таким образом, латеральные порции лобной мышцы из нормокинеза переходят в состояние гиперкинетической активности, компенсируя тем самым нарушенную функцию центральной порции лобной мышцы и глабеллы.
Латеральные порции лобной мышцы из нормокинеза переходят в состояние гиперкинетической активности, компенсируя тем самым нарушенную функцию центральной порции лобной мышцы и глабеллы.
Можно ли предупредить формирование подковообразных морщин над бровями, не опуская при этом сами брови? Ответ лежит, казалось бы, на поверхности: необходимо провести ботулинотерапию m. frontalis, включая все ее порции, и во избежание птоза бровей денервировать все ее антагонисты. Но тут и возникают трудности в расчете доз: как денервировать круговые мышцы глаз, находящиеся в антагонистических отношениях с m. frontalis в ее латеральных порциях? Когда и как инъецировать БТ типа А в мышцы-синергисты глабеллярного комплекса – леваторы крыльев носа и верхней губы и поперечное брюшко мышцы носа? Да и можно ли себе позволить энергичную денервацию круговых мышц глаз, которые граничат со слезными железами и леваторами верхних век? А если нет, то какую минимально эффективную дозу можно инъецировать безопасно?
frontalis, включая все ее порции, и во избежание птоза бровей денервировать все ее антагонисты. Но тут и возникают трудности в расчете доз: как денервировать круговые мышцы глаз, находящиеся в антагонистических отношениях с m. frontalis в ее латеральных порциях? Когда и как инъецировать БТ типа А в мышцы-синергисты глабеллярного комплекса – леваторы крыльев носа и верхней губы и поперечное брюшко мышцы носа? Да и можно ли себе позволить энергичную денервацию круговых мышц глаз, которые граничат со слезными железами и леваторами верхних век? А если нет, то какую минимально эффективную дозу можно инъецировать безопасно?
Иными словами, задача врача состоит в том, чтобы сохранить высоту бровей и разгладить морщины над ними с ограничением активности мышц-антагонистов и достижением баланса между ними. Идея применения относительных дозировок поможет нам этот баланс найти.
Управление леваторной и депрессорной порциями m. frontalis
Разберем еще один пример.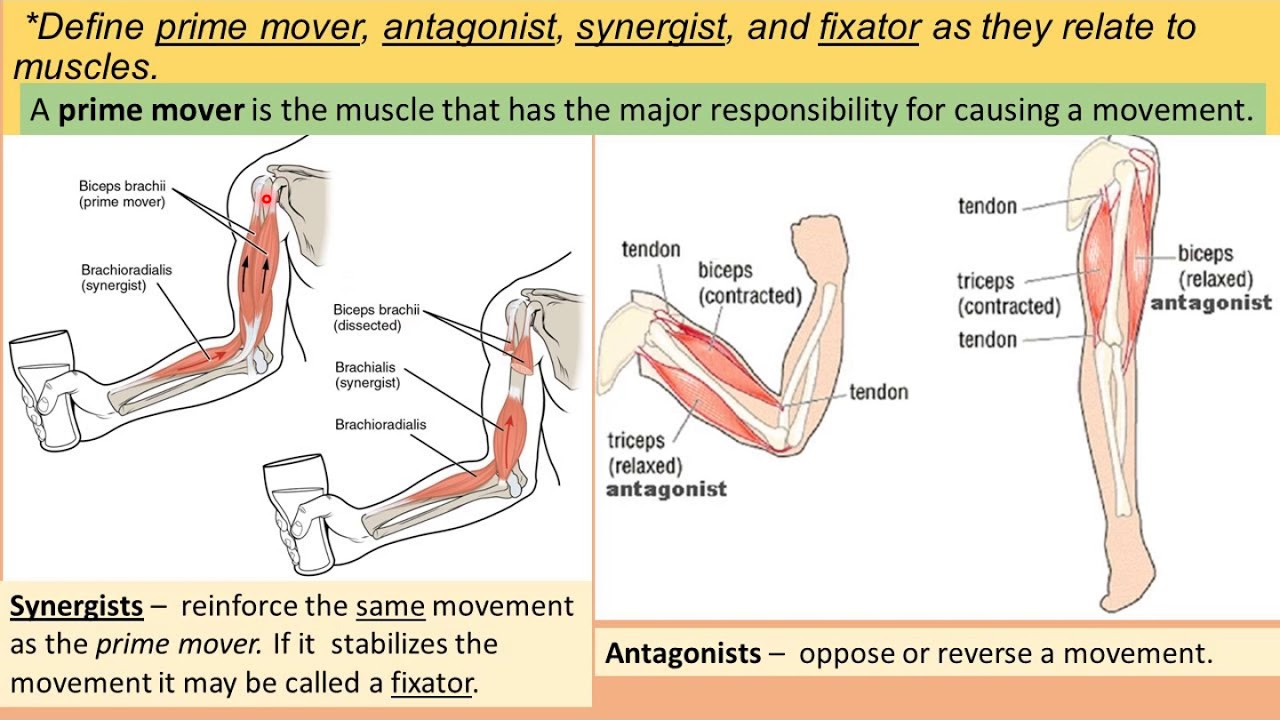 Иногда лобная мышца по срединной горизонтали лба делится на две порции (причем антагонистические!): нижнюю – леваторную и верхнюю – депрессорную. Этот морфовариант был описан доктором медицины Клодом Ле Луарном как иллюстрация к двухрядному расположению моторных пластин в волокнах лобной мышцы.
Иногда лобная мышца по срединной горизонтали лба делится на две порции (причем антагонистические!): нижнюю – леваторную и верхнюю – депрессорную. Этот морфовариант был описан доктором медицины Клодом Ле Луарном как иллюстрация к двухрядному расположению моторных пластин в волокнах лобной мышцы.
Лобная мышца разделена по срединной горизонтали лба на две антагонистические порции: нижнюю – леваторную и верхнюю – депрессорную.
Такое порционное деление лобной мышцы у пациента легко определяется по встречному сокращению леваторной и депрессорной порций при сморщивании лба. Оно отчетливо проявляется видимым смещением линии роста волос вниз, когда человек проявляет удивление. Позволю себе утверждать, что анатомо-функциональной основой такого морфоварианта лобного брюшка затылочно-лобной мышцы является наличие дополнительной опорной апоневротической фиксации депрессорной порции лобной мышцы. Эта опорная фиксация идет, скорее всего, вдоль средней горизонтали лба, где две антагонистические порции переплетаются друг с другом, и леваторная нижняя порция, вероятно, покрывает депрессорную верхнюю.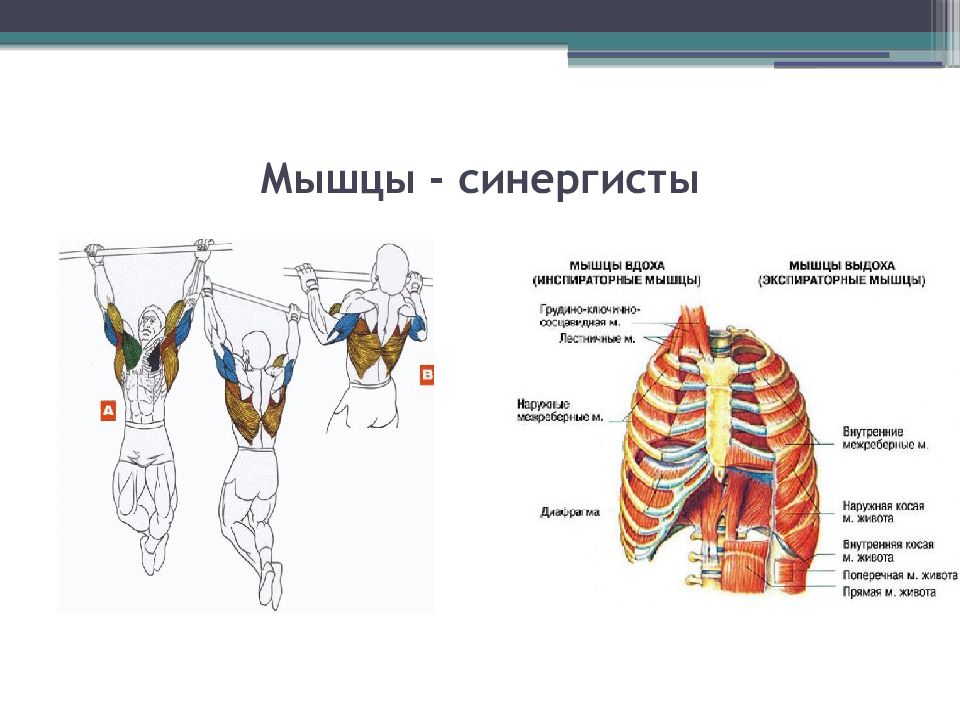
Предположение основано на универсальном анатомическом правиле: сокращение мышцы и направление этого сокращения определяется областью (точкой) опоры, а модуль вектора сокращения всегда направлен единственно в сторону этой опоры.
Отсутствие внимания к депрессорной порции m. frontalis и изолированная денервация ее леваторной порции при таком строении лба неизбежно приведут к птозу бровей. Но как распределить дозу с учетом этого необычного внутримышечного антагонизма в пределах одной и той же мышцы – m. frontalis? Как выбрать правильную пропорцию между двумя порциями одной и той же мышцы?
Золотое сечение, или золотая пропорция
Рассмотрим относительные величины, а именно относительные дозы БТ типа А, уместные при работе с единой биомеханической системой лица.
Метод расчета соотношения доз, адресуемых мимическим мышцам, в ботулинотерапии реализуется при работе с мышцами-антагонистами. Они являются моделью гармоничных отношений противоположностей, которые подпадают под соотношения чисел в знаменитой золотой пропорции, или золотом сечении. Древний Египет, Древняя Греция (школа Пифагора), средневековая Италия (Фибоначчи и Леонардо да Винчи) – неполный перечень стран и имен, где золотая пропорция применялась как стандарт в самых разных областях человеческой деятельности и бытия. К этой же пропорции спустя столетия пришел и калифорнийский пластический хирург Стивен Марквордт, изучая «идеальные» лица современников. Составив маску человеческого лица, он обнаружил, что черты его укладываются в два перевернутых пятиугольника, которые, в свою очередь, состоят из трех «золотых» треугольников. Внутри маски также встречаются пропорции, соответствующие золотому сечению.
Древний Египет, Древняя Греция (школа Пифагора), средневековая Италия (Фибоначчи и Леонардо да Винчи) – неполный перечень стран и имен, где золотая пропорция применялась как стандарт в самых разных областях человеческой деятельности и бытия. К этой же пропорции спустя столетия пришел и калифорнийский пластический хирург Стивен Марквордт, изучая «идеальные» лица современников. Составив маску человеческого лица, он обнаружил, что черты его укладываются в два перевернутых пятиугольника, которые, в свою очередь, состоят из трех «золотых» треугольников. Внутри маски также встречаются пропорции, соответствующие золотому сечению.
Золотая пропорция (золотое сечение) – это соотношение двух величин, равное соотношению их суммы к большей из данных величин. Приблизительная величина золотого сечения равна 1,6180339887. В процентном округленном значении это деление величины на 62 и 38% соответственно, что выражается формулой:
(a + b) / a = a / b = 1,6180339887 при условии, что а > b
Золотая пропорция, применяемая для расчета доз в ботулинотерапии, дает возможность дополнить абсолютные их значения относительными и эффективно оперировать соотношениями доз.
Работа с соотношениями доз в золотой пропорции парадоксальным образом гарантирует безопасность и эффективность ботулинотерапии мимических мышц-антагонистов, вместе с тем оставляя специалисту большую свободу в подборе индивидуальных абсолютных дозировок БТ типа А.
В подавляющем большинстве случаев, выбирая дозировки для мимических мышц, в качестве меньшего слагаемого мы выбираем мышцы-леваторы, чтобы сохранить вверенную им природой позитивную мимику. Но поскольку есть исключения (особенно для центральных или медиальных леваторов), то прежде всего необходимо определиться с целевой мышцей (или порцией мышцы, или группой мышц), подобрать необходимую для денервации дозу и далее строить пропорцию относительно нее с ее антагонистами.
Прежде чем перейти к рекомендациям по проведению расчета относительных доз БТ типа А для мышц, реализующих то или иное мимическое движение и являющихся таргетными, необходимо ввести несколько обязательных понятий.
Составив маску лица, С. Марквордт обнаружил, что черты его укладываются в два перевернутых пятиугольника, которые, в свою очередь, состоят из трех «золотых» треугольников. Внутри «маски красоты» встречаются пропорции, соответствующие золотому сечению.
Марквордт обнаружил, что черты его укладываются в два перевернутых пятиугольника, которые, в свою очередь, состоят из трех «золотых» треугольников. Внутри «маски красоты» встречаются пропорции, соответствующие золотому сечению.
Понятие кинематической цепи, давно применяемое в спортивной медицине. Кинематическая цепь – это любая произвольно взятая группа взаимодействующих друг с другом мышц или групп мышц. Она может состоять из пары и более мышц. Обязательным условием образования динамической структуры, называемой кинематической цепью, является наличие взаимодействия, прямого или опосредованного, между мышцами – участницами кинематической цепи. То есть состав кинематической цепи – это взаимодействующие друг с другом мышцы.
Одна и та же мышца при разной мимике может быть участницей разных кинематических цепей. Одна и та же мышца может иметь разные порции, участвующие в разных кинематических цепях. SMAS обеспечивает единство биомеханической системы лица в любых кинематических цепях и, соответственно, движениях, но самая важная роль принадлежит ей в кооперации работы удаленных друг от друга кинематических цепей.
Провести диагностику состава кинематических цепей с детальным анализом взаимоотношений мимических мышц, включенных в цепь.
Убедиться, что мышцы целевой области для коррекции БТ типа А находятся в антагонистических взаимоотношениях («леватор – депрессор», «депрессор – сфинктер» и «леватор – сфинктер»).
Определить таргетные мышцы и/или их отдельные порции в кинематических цепях, реализующие мимическую активность целевой области ботулинотерапии.
Проанализировать взаимоотношения целевых мышц и/или их порций в кинематических цепях.
Убедиться, что мышцы нецелевых областей (ими могут быть мышцы как соседних, так и удаленных областей лица и шеи) не влияют и/или не могут повлиять в дальнейшем (компенсаторно, исходя из антагонизма или синергии) на активность целевой области коррекции.
При расчете золотой пропорции для мышц-антагонистов, как правило, меньшей должна быть доза для леваторов, а большей – для депрессоров (есть исключения – например, подбородочная мышца, являющаяся леватором, имеет особенности содружественной активности с депрессорами).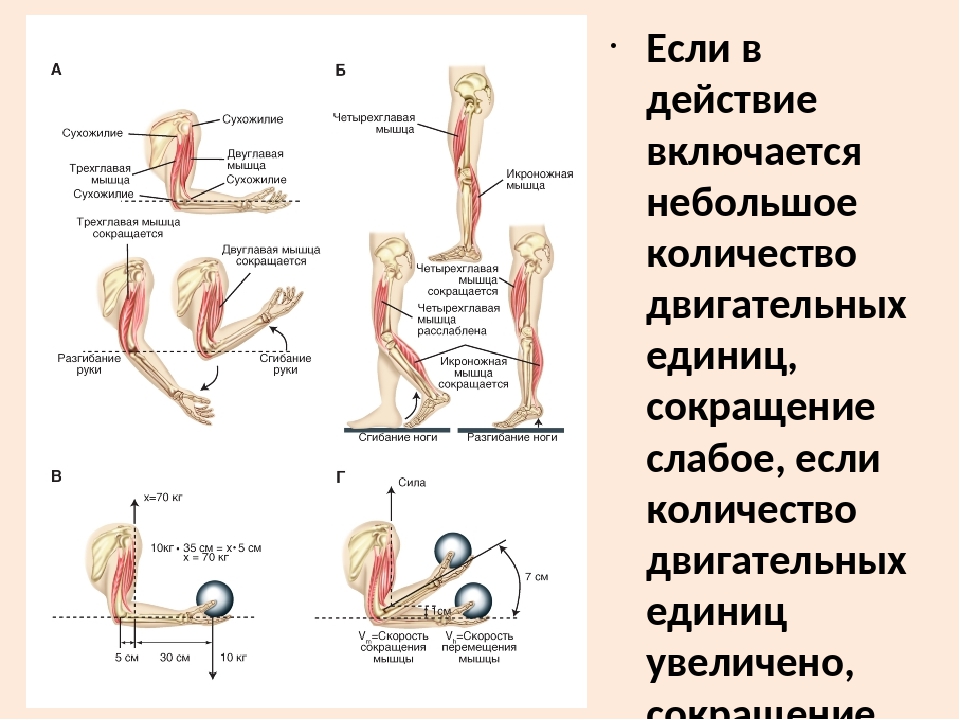 Слагаемые должны составить суммарно 100% подобранной для кинематической цепи дозы.
Слагаемые должны составить суммарно 100% подобранной для кинематической цепи дозы.
При любом изменении дозы для любой из мышц – участниц кинематической цепи (или порции мышцы-участницы, или группы мышц-участниц) необходимо в соответствии с золотым сечением пропорционально изменить дозы для всех мышц-антагонистов внутри общей с ними кинематической цепи, оставляя неизменным соотношение между дозами для мышц-антагонистов (одной мышцы, или ее порции, или группы мышц).
При увеличении дозы для мышцы-леватора пропорционально, в соответствии с золотым сечением, должна возрастать, как правило, доза для мышцы-депрессора. Это не означает, что дозу на леваторы можно увеличивать безнаказанно, надеясь на адекватную денервацию их антагонистов-депрессоров в кинематической цепи, поскольку закон меньшей дозы на мышцы-леваторы предполагает в первую очередь малые абсолютные дозы для них. Иначе какие мышцы станут осуществлять леваторные функции?
Золотая пропорция (золотое сечение) – это соотношение двух величин, равное соотношению их суммы к большей из данных величин.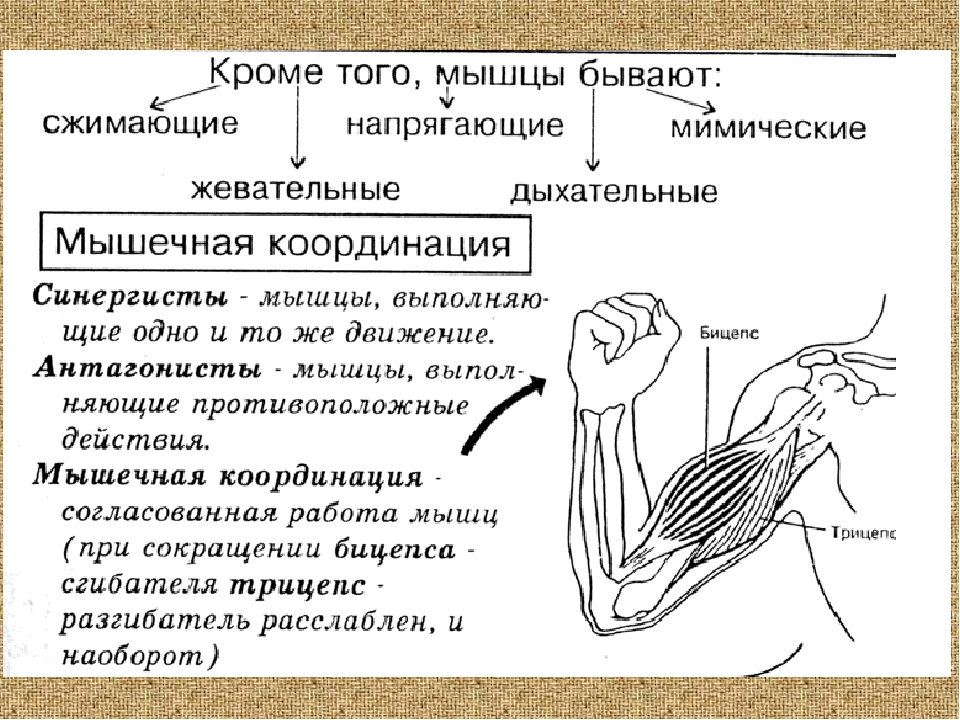 Приблизительная величина золотого сечения равна 1,6180339887.
Приблизительная величина золотого сечения равна 1,6180339887.
Практическое применение золотой пропорции
Рассмотрим усредненные практические примеры, которые демонстрируют, что в клинической практике мы во многих случаях эмпирически соблюдали правила золотой пропорции.
Работа с верхней третью лица, препарат «Диспорт»:
– при V-образном инъецировании лба: в леватор – лобную мышцу, – исключая ее латеральные порции – 24 ЕД препарата «Диспорт»; в депрессоры – мышцы глабеллярной области – в сумме 40 ЕД препарата «Диспорт».
24 + 40 = 64; 64 : 40 = 40 : 24 = 1,6
– при дополнительном инъецировании компенсаторных дугообразных морщин над каудальными отрезками бровей: в леватор – латеральные порции лобной мышцы – по 3–4 ЕД; всего на латеральные области лба – 8 ЕД препарата «Диспорт»; в депрессоры – верхнелатеральные участки круговых мышц глаз (каудальные участки бровей) – по 6–7 ЕД; всего под оба хвостика бровей – 12 ЕД препарата «Диспорт».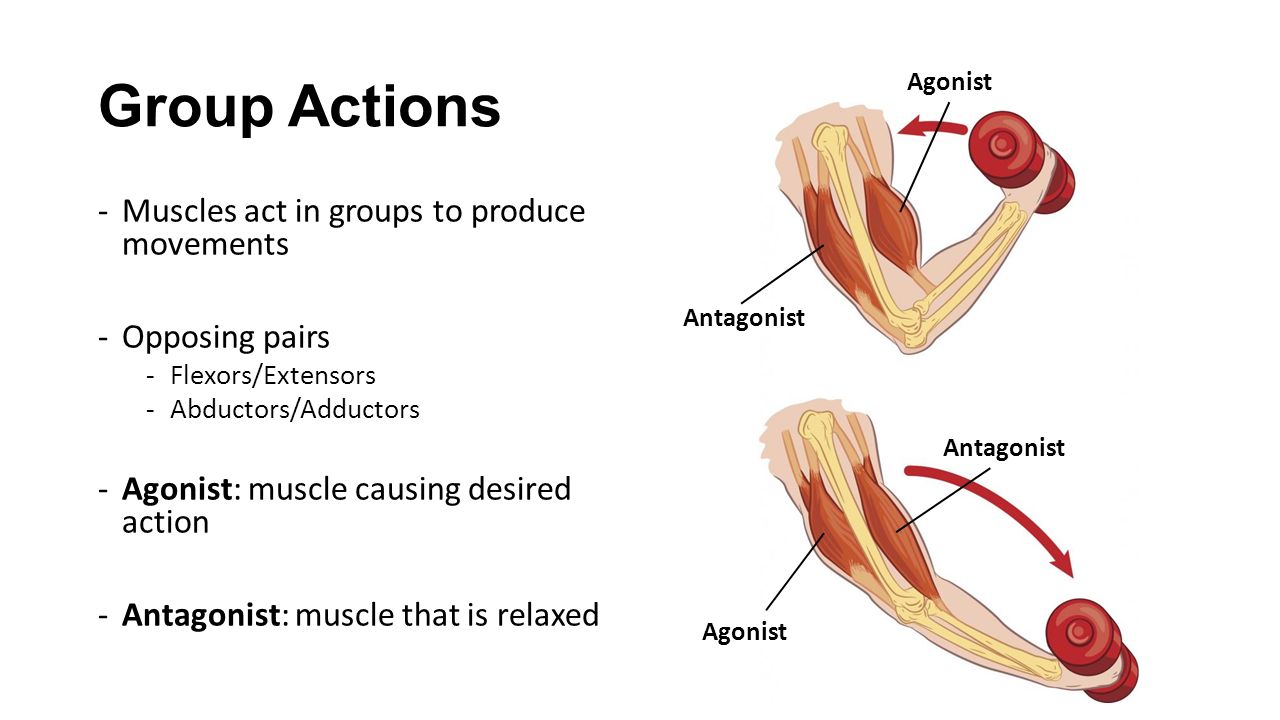
8 + 13 = 21; 21 : 13 = 13 : 8 = 1,6
Суммарно примеры 1–2:
32 + 53 = 85; 85 : 53 = 53 : 32 = 1,6
При увеличении дозы препарата на леваторы пропорционально должна возрастать доза на депрессоры. При снижении дозы препарата на леваторы допустимо пропорциональное уменьшение дозы на депрессоры.
Применение золотой пропорции в ботулинотерапии может быть особенно полезным для безопасного подбора соотношений доз для мимических мышц-антагонистов у пациентов со сложными индивидуальными особенностями мимики и архитектоники лица.
Данная стратегия не ограничена особенностями разных токсинов БТ типа А разных производителей, поскольку речь идет не об абсолютных дозировках, а об их соотношении.
ЗаключениеДозирование ботулинического нейропротеина типа А с целью его лечебного воздействия на любую кинематическую цепь мимических мышц-антагонистов представляется целесообразным производить, согласуясь с формулой золотого сечения (золотой пропорции): (a + b) / a = a / b = 1,6180339887 при условии, что а > b
При эстетической коррекции расчет абсолютных значений доз ботулинического нейропротеина типа А на мимические мышцы производится в зависимости от решаемой врачом задачи, индивидуальных особенностей мимики и архитектоники лица пациента.
Распределение абсолютных значений доз БТ типа А на мимические мышцы необходимо производить в золотой пропорции с соблюдением соотношения между дозами БТ типа А, предназначенными для мышц-антагонистов в одной кинематической цепи.
Использование принципов золотого сечения позволяет достичь динамического баланса между мимическими мышцами, гармонизировать единую биомеханическую систему лица в покое и движении и избежать нежелательных явлений в эстетической ботулинотерапии.
Елена Парсагашвили, дерматовенеролог, косметолог, физиотерапевт, ведущий специалист по инвазивным методикам, гл. врач «Эстима-клиник», Санкт-Петербург
Лабораторные работы 8 класс Пасечник
4) Сделайте выводы. Как особенности строения тканей связаны с выполненными функциями?Лабораторная работа № 2
Самонаблюдение мигательного рефлекса и условия его проявления и торможения. Коленный рефлекс.
Цель : Изучить мигательный и коленный рефлекс , условия их проявления и торможения .
Ход работы
1.Осторожно прикоснитесь рукой
а) к наружному углу глаза,б) к внутреннему углу глаза, в) к ресницам, г) к бровям, д) к коже век,
Отметьте те области, раздражение которых вызывает непроизвольное мигание, знаком «+»
2.Результаты оформите в виде таблицы.
3.Почему не наблюдается мигательный рефлекс при раздражении некоторых областей?4. Многократно прикасайтесь к внутреннему углу глаза со стороны носа, не задевая ресниц. Что происходит?
5. Проверьте возможность с помощью волевого усилия затормозить мигательный рефлекс. Объясните почему это удалось?
6. Посадите испытуемого на стул. Предложите ему перекинуть ногу на ногу, затем ударьте ребром руки по сухожилию четырехглавой мышцы под коленной чашечкой.
5.Составьте схему рефлекторной дуги мигательного и коленного рефлексов.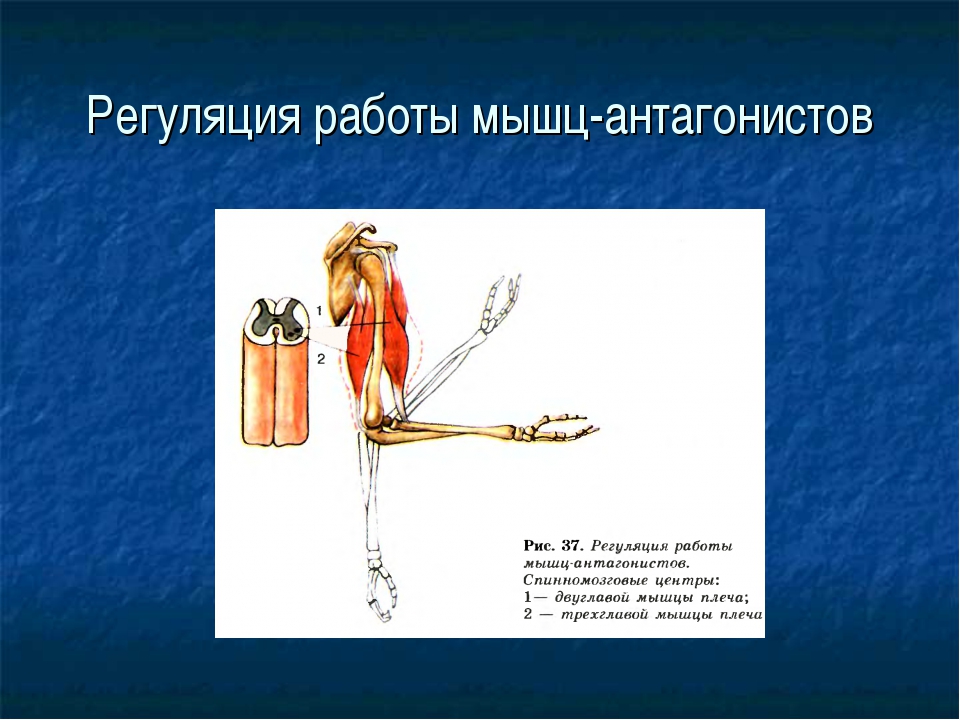 .
.
6. Сделайте вывод о значении мигательного и коленного рефлексов.
Лабораторная работа №3
Микроскопическое строение кости.
Цель : изучить особенности микроскопического строения кости
Оборудование: микроскоп, постоянный препарат «Костная ткань».
Ход работы
1. Рассмотрите при малом увеличении микроскопа костную ткань.
2. Найдите канальцы, по которым проходили сосуды и нервы. На поперечном срезе они имеют вид прозрачного кружка или овала.
3. Найдите костные клетки, которые находятся между кольцами и имеют вид черных паучков. Они выделяют пластинки костного вещества, которые потом пропитываются минеральными солями.
4. Подумайте, почему компактное вещество состоит из многочисленных трубочек с прочными стенками. Как это способствует прочности кости при наименьшем расходе материала и массы костного вещества.
5. Сделайте вывод об особенностях микроскопического строения кости.
Сделайте вывод об особенностях микроскопического строения кости.
Лабораторная работа №4
Мышцы человеческого тела
Цель : изучить мышцы человеческого тела .
Оборудование: учебник
Ход работы
1.Используя учебник и анатомическое описание (стр. учебника 65-68), определите местоположение мышечных групп и выполняемые ими движения.
2.Результаты оформите в таблице.
3.Приведите несколько примеров мышц, работающих как синергисты и как антагонисты 4.Сделайте выводы о значении мышц.Лабораторная работа №5
Самонаблюдения работы основных мышц , роль плечевого пояса в движении руки.
Цель: пронаблюдать работы основных мышц , роль плечевого пояса в движении руки
Ход работы.
1. В положении сидя найдите нижний угол левой лопатки. Отведите левую руку в сторону до горизонтального уровня. Происходит ли смещение лопатки?
Отведите левую руку в сторону до горизонтального уровня. Происходит ли смещение лопатки?
2. Продолжите движение руки до верхнего вертикального положения. Происходит ли смещение лопатки?
3. Положите руку на левую ключицу и повторите движение. В каком случае происходит движение ключицы?
4. Сделайте вывод: Какова роль плечевого пояса в движении руки?
Лабораторная работа №6
Утомление при статической и динамической работе.
Цель : наблюдение признаков утомления при статической и динамической работе.
Оборудование: секундомер, груз массой 4-5 кг.
Ход работы
1. Испытуемый становится лицом к классу, вытягивает руку в сторону горизонтально. Мелом на доске отмечается тот уровень, на котором находится рука. Включите секундомер.
2. Наблюдайте, за какое время произойдет утомление мышц.
3. Результаты оформите в таблице.
Признаки утомления
Время
Отсутствие утомления
Рука с грузом неподвижна
Первая фаза утомления
Рука опускается, затем рывком поднимается на прежнее место
Вторая фаза утомления
Дрожание рук, пошатывание корпуса, покраснение лица, потоотделение
Предельное утомление
Рука с грузом опускается; опыт прекращается
4. Сделайте выводы о различие между динамической и статической работой?
Лабораторная работа №7
Выявление нарушения осанки
Цель : выявить нарушения осанки.
Оборудование : сантиметровая лента
Ход работы.
1. Для выявления сутулости( круглой спины) сантиметровой лентой измерьте расстояние между самыми отдаленными точками левого и правого плеча , отступя на 3-5 см от плечевого сустава, со стороны груди и со стороны спины. Первый результат разделите на второй. Если получиться число . близкое к единице или больше ее, значит нарушений нет. Получение числа меньше единицы говорит о нарушении осанки.
Первый результат разделите на второй. Если получиться число . близкое к единице или больше ее, значит нарушений нет. Получение числа меньше единицы говорит о нарушении осанки.
2. Встаньте спиной к стене так, чтобы пятки, голени, таз и лопатки касались стены. Попробуйте между стеной и поясницей просунуть кулак. Если он проходит — нарушение осанки есть. Если проходит только ладонь-осанка нормальная.
3. Сделайте вывод: Есть ли у вас нарушение осанки? Если есть, предположите, каковы причины?
Лабораторная работа №8
Выявление плоскостопия (работа выполняется дома).
Цель : выявить наличие или отсутствие плоскостопия
Оборудование: таз с водой, лист бумаги, простой карандаш.
Ход работы.
1. Мокрой ногой встаньте на лист бумаги. Контуры следа обведите простым карандашом.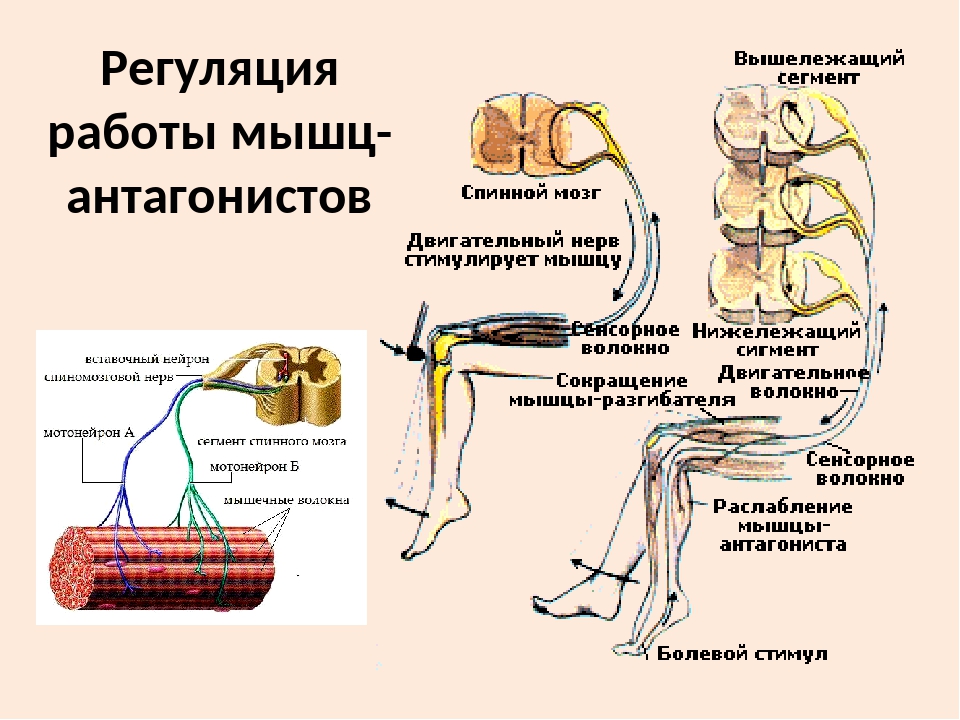 Найдите центр пятки и центр третьего пальца.
Найдите центр пятки и центр третьего пальца.
2. Соедините две найденные точки прямой линией. Оцените свои результаты. Если в узкой части след не заходит за линию — плоскостопия нет (стр.74,рис39).
3. Сделайте вывод: Есть ли у вас плоскостопие? Если есть, предположите, каковы его причины? Предложите упражнения, предупреждающие плоскостопие.
Лабораторная работа №9
Рассматривание клеток крови человека и лягушки под микроскопом
Цель: рассмотреть клетки крови человека и лягушки под микроскопом и выяснить их функции в зависимости от строения , количества.
Оборудование: готовые микропрепараты крови человека и лягушки, микроскоп.
Ход работы
1.Рассмотреть и зарисовать эритроциты крови человека и лягушки (вид сбоку и спереди).
2.Сравнить эритроциты человека и лягушки: цвет, размеры, наличие ядра, форма, количество (приблизительно) в поле зрения микроскопа.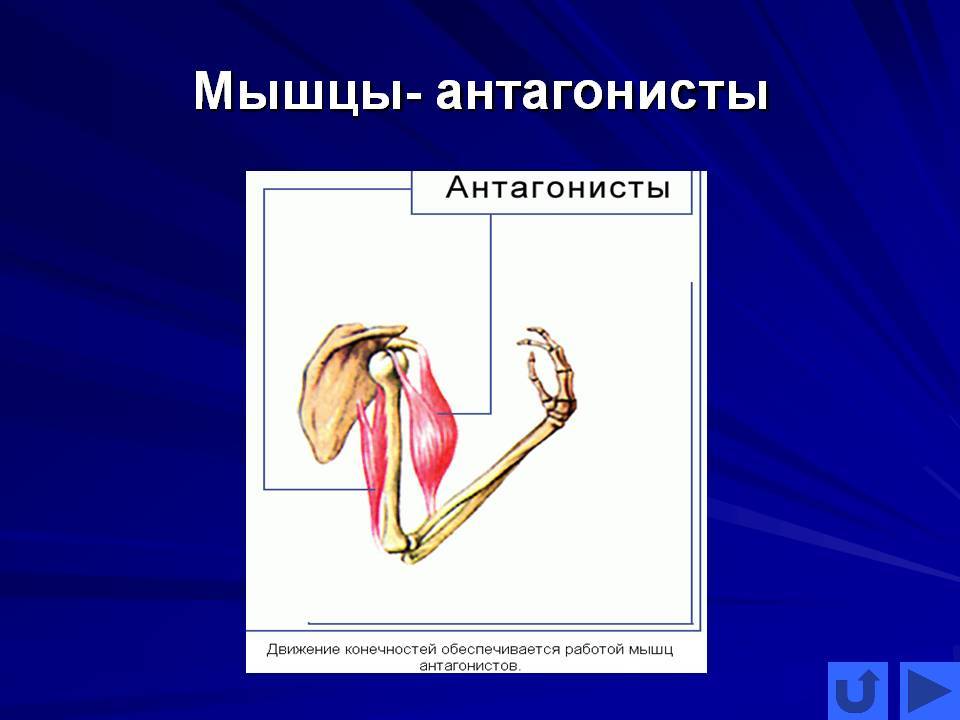
3.Результаты оформите в таблице.
ПризнакиЭритроциты лягушки
Эритроциты человека
Цвет
Размеры
Наличие ядра
Форма
Количество в поле зрения микроскопа
4. Сформулируйте вывод В чем отличие эритроцитов человека от эритроцитов лягушки?
Вывод: Эритроциты человека, в отличие от эритроцитов лягушки, не имеют ядра и приобрели двояковогнутую форму. Двояковогнутая форма эритроцита человека увеличивает поверхность клетки, а место ядра в них заполняется гемоглобином, поэтому каждый эритроцит человека может захватывать больше кислорода, чем эритроциты лягушки.
Эритроциты человека меньше по размерам, чем эритроциты лягушки, поэтому в крови человека в единице объема количество эритроцитов больше (в 1 мм 3 5 млн.), чем в крови у лягушки. Исходя из особенностей строения эритроцитов и большого их количества в крови человека, следует, что кровь человека содержит больше кислорода, чем кровь лягушки.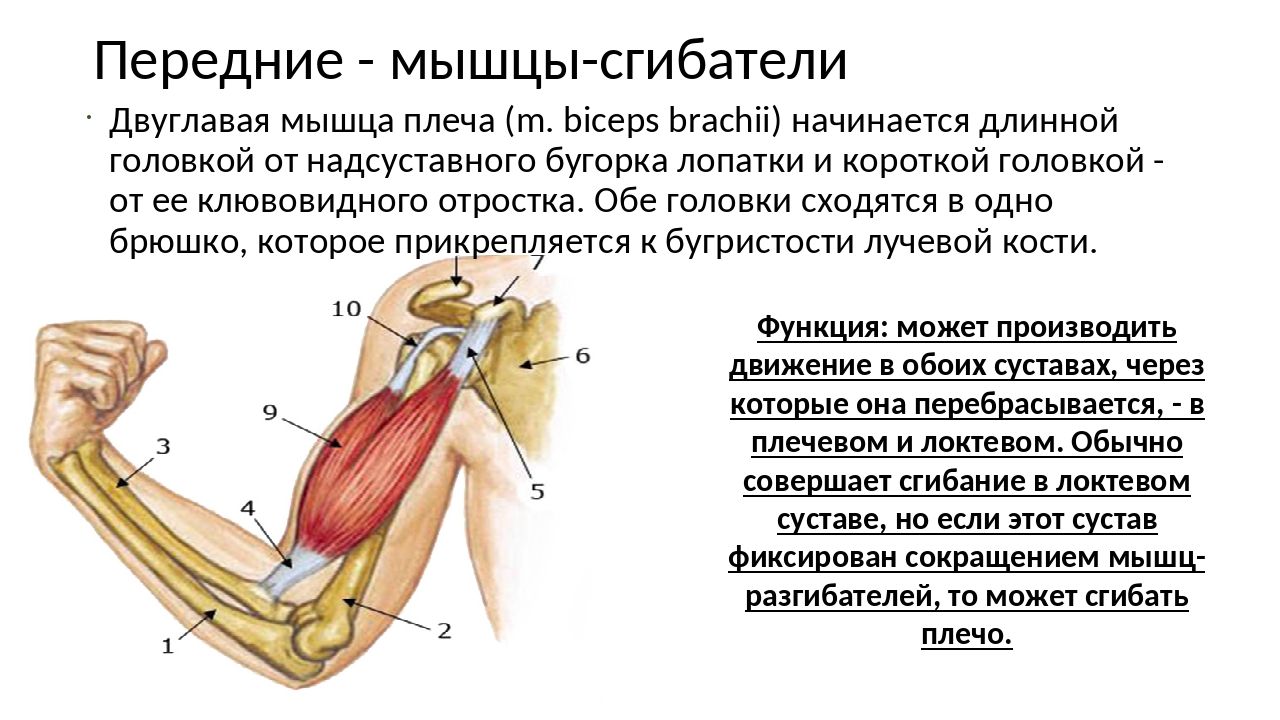 Дыхательная функция крови человека значительно эффективнее, чем у земноводных животных
Дыхательная функция крови человека значительно эффективнее, чем у земноводных животных
Лабораторная работа №10
Положение венозных клапанов в опущенной и поднятой руке.
Цель : изучить роль клапанов в опущенной и поднятой руке.
Пояснение. Если рука опущена, венозные клапаны не дают крови стечь вниз. Клапаны раскрываются лишь после того, как в ниже лежащих сегментах накопится достаточное количество крови, чтобы открыть венозный клапан и пропустить кровь вверх, в следующий сегмент. Поэтому вены, по которым кровь движется против силы тяжести, всегда набухшие.
Ход работы.
1.Поднимите одну руку вверх, а вторую опустите вниз. Спустя минуту положите обе руки на стол. Наблюдения запишите в тетрадь.
2.Сделайте выводы:
— Почему поднятая рука побледнела, а опущенная – покраснела? В какой руке венозные клапаны были закрыты?
Лабораторная работа №11
Изменения в тканях при перетяжках, затрудняющих кровообращение.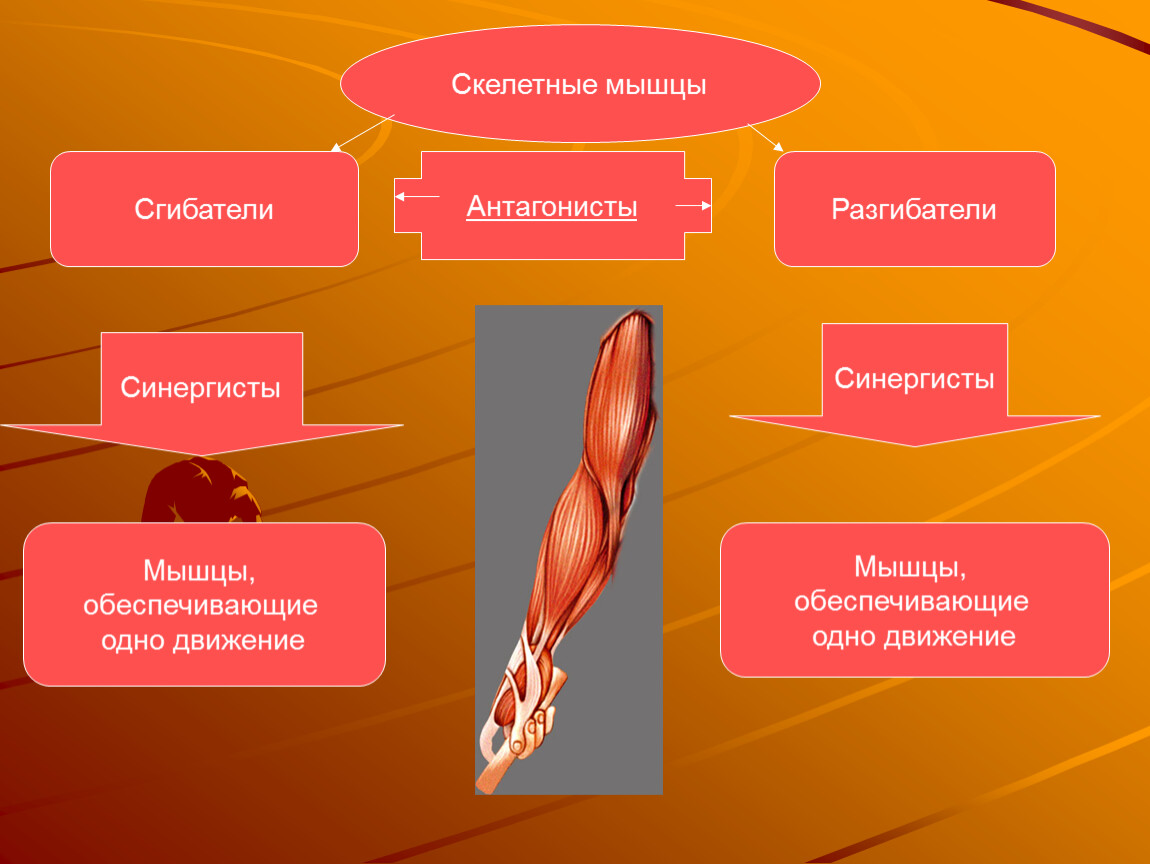
Цель : выяснить изменения в тканях при перетяжках
Оборудование: аптечное резиновое кольцо или нитки.
Пояснение. Перетяжка конечности затрудняет отток крови по венам и лимфы по лимфатическим сосудам. Расширение кровеносных капилляров и вен приводит к покраснению, а затем к посинению части органа, изолированной перетяжкой. В дальнейшем эта часть органа становится белой из-за выхода плазмы крови в межклеточное пространство. Тканевая жидкость накапливается, сдавливая клетки. Орган становится плотным на ощупь. Начинается кислородное голодание тканей, которое ощущается как «ползание мурашек», покалывание. Работа рецепторов нарушается.
Ход работы.
1. Накрутите на палец резиновое кольцо или перетяните палец ниткой. Обратите внимание на изменение цвета и формы пальца.
2. Дотроньтесь перетянутым пальцем до какого-нибудь предмета. Палец кажется каким-то ватным.
3. Снимите перетяжку и помассируйте палец по направлению к сердцу. Что достигается этим приемом?
4. Сделайте выводы:
— Почему при перетяжке изменялся цвет пальца?
— Почему ткани пальца уплотнены?
— Как проявляются признаки кислородной недостаточности?
— Почему вредно туго затягиваться ремнем, носить тесную одежду?
Лабораторная работа №12
Определение скорости кровотока в капиллярах ногтевого ложа.
Цель: определить скорость кровотока в капиллярах ногтевого ложа.
Оборудование: линейка, секундомер или часы с секундной стрелкой.
Ход работы
1. Измерьте длину ногтя большого пальца от корня до места, где кончается его розовая часть, и начинается прозрачный ноготь, который обычно срезается (стр. 117, рис.56.А). Запишите результат.
117, рис.56.А). Запишите результат.
2. Нажмите указательным пальцем на ноготь так, чтобы он побелел. При этом кровь будет вытеснена из сосудов ногтевого ложа. Уберите указательный палец. Через некоторое время ноготь начинает краснеть. Повторите опыт, зафиксируйте время до полного покраснения пальца. Это время, за которое кровь проделала свой путь.
3. Рассчитайте скорость кровотока в капиллярах ногтевого ложа по формуле
V=S/t , где
S- длина пути, которую пройдет кровь от корня ногтя до его вершины,
t- время, которое ей для этого потребуется.
4. Сравните скорость тока крови в крупных артериях, венах, капиллярах.
Оценка результатов: у большинства людей около 1-0.5 см/с.это в 50-100 раз меньше чем в аорте и в 25- 50 раз меньше чем в полых венах.
5. Сделайте вывод:
— Какое значение имеет медленное течение крови в капиллярах?
Лабораторная работа № 13
Опыты, выясняющие природу пульса
Цель: доказать, что пульс связан с колебаниями стенок артерий, а не с толчками, возникающими при движении крови.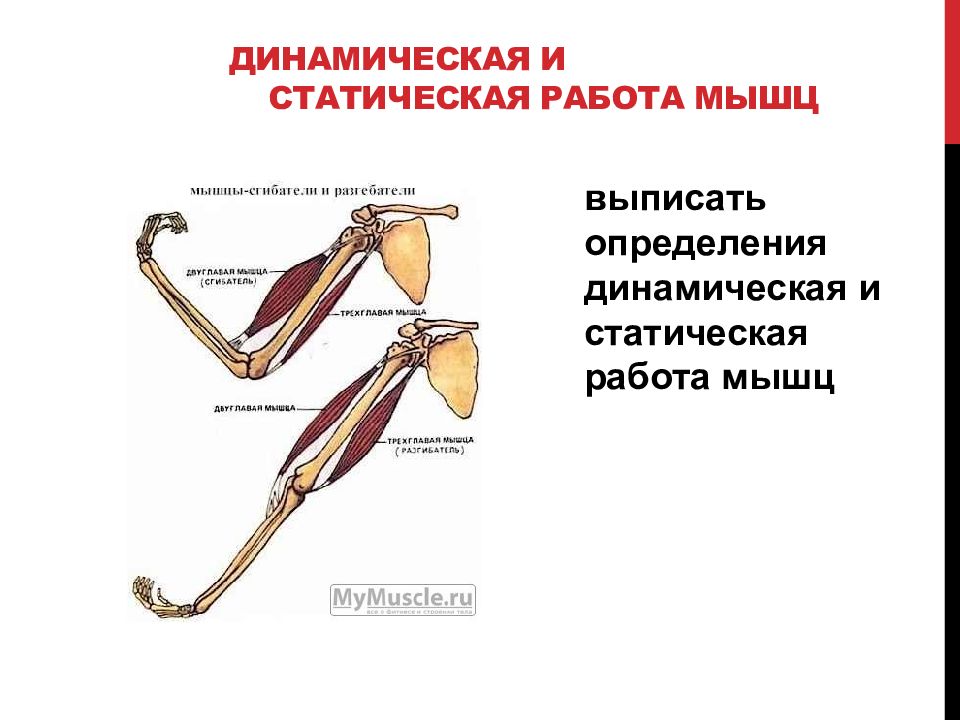
Пояснение: Чтобы доказать, что пульс связан с колебаниями стенок артерий, а не с толчками, возникающими при движении крови, надо на каком-то участке артерии остановить движение крови, но так, чтобы стенки артерий продолжали колебаться.
Ход работы.
1.Найдите пульс на лучевой артерии. Наметьте точку a , ближайшую к большому пальцу исследуемой руки, и точку b, наиболее удаленную от большого пальца. Кровь течет то точки b к точке a ( рис 56,Б, стр. 117).
2. Зажмите артерию в точке a. Движение крови на участке ba будет остановлено. Однако стенка артерии в точке b, будет продолжать колебаться, и пульс в этой точке будет прощупываться.
3. Теперь зажмите артерию в точке b. Вы не только остановили кровь, но и прекратили распространение пульсовой волны, которая не сможет пройти через участок b. В этом случае, в точке а пульс ощущаться не будет.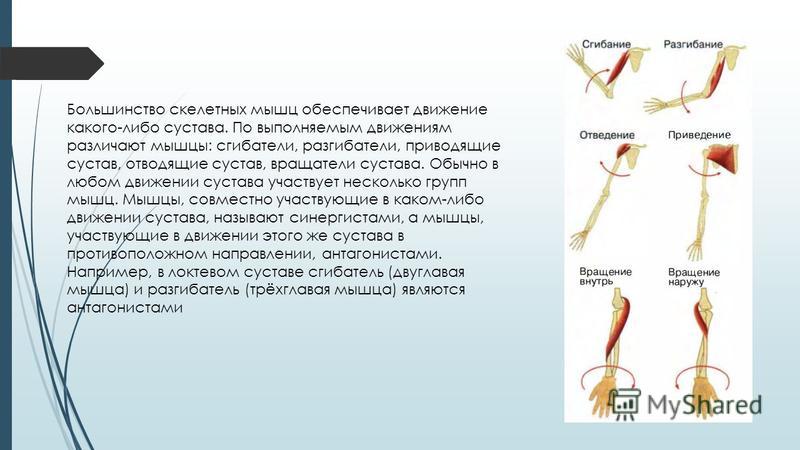
Оценка результатов. Пульсовая волна передается по стенке артерий и не зависит от наличия или отсутствия кровотока. Пульс прощупывается выше места, где артерия перетянута, а ниже этого места отсутствует и кровоток , и пульс, потому что, прижимая стенки артерий друг к другу мы не только останавливаем кровь, но и останавливаем колебание стенок артерий.
Лабораторная работа №14
Функциональная проба :реакция сердечно-сосудистой системы на дозированную нагрузку.
Цель: выяснить реакцию сердечно-сосудистой системы на дозированную нагрузку.
Оборудование: секундомер или часы с секундной стрелкой .
Ход работы.
1. Найдите пульс. Обычно принято определять пульс на лучевой артерии, у основания большого пальца, для чего 2, 3и 4-й пальцы накладывают несколько выше лучезапястного сустава, нащупывают артерию и прижимают ее к кости. 2.Подсчитайте число ударов в спокойном состоянии за 10 секунд. Этот показатель умножьте на шесть, и вы получите ваш пульс за 1 минуту. 3.Сделайте 10 приседаний в быстром темпе, снова подсчитайте число ударов за 10 секунд. Этот показатель умножьте на шесть, и вы получите пульс за 1 минуту. Определите показания за 1 минуту.
2.Подсчитайте число ударов в спокойном состоянии за 10 секунд. Этот показатель умножьте на шесть, и вы получите ваш пульс за 1 минуту. 3.Сделайте 10 приседаний в быстром темпе, снова подсчитайте число ударов за 10 секунд. Этот показатель умножьте на шесть, и вы получите пульс за 1 минуту. Определите показания за 1 минуту.
4.Свои результаты оформите в виде таблицы.
Число ударов за 1 мин.в состоянии покоя
Число ударов за 1 мин.
после 10 приседаний
5.Оцените свои результаты.
Пояснение (Частота пульса в возрасте 13-18 лет в норме составляет 60-90 ударов в минуту. Результаты хорошие, если частота пульса после приседаний повысилась на 1/3 или меньше от результатов покоя; если наполовину — результаты средние, если больше, чем наполовину — результаты неудовлетворительные. Частота пульса после приседаний нормализуется спустя 2-3 минуты).
6. Сделайте вывод:
— работа собственного сердца в состоянии покоя и при нагрузке. — почему после физической нагрузки частота пульса увеличилась?
— почему после физической нагрузки частота пульса увеличилась?
Лабораторная работа №15
Измерение обхвата грудной клетки в состоянии вдоха и выдоха Оборудование: сантиметровая лента.
Ход работы
1.Испытуемому предлагают приподнять руки и накладывают измерительную ленту так, чтобы на спине она касалась углов лопаток. Во время измерения руки должны быть опущены.
2.Измерьте обхват грудной клетки на вдохе. Испытуемый должен глубоко вдохнуть. Мышцы напрягать нельзя, плечи не поднимать. Запишите результат.
3. Измерьте обхват грудной клетки на выдохе. Испытуемый должен сделать глубокий выдох. Плечи не опускать, не сутулиться. Запишите результат.
4.Оцените свои результаты.
5.Сделайте вывод.
Пояснение: (В норме разница обхвата грудной клетки в состоянии глубокого вдоха и в состоянии глубокого выдоха равна 6-9 см).
Лабораторная работа № 15
Функциональные пробы с задержкой дыхания на фазе вдоха и выдоха.
Оборудование: секундомер.
Ход работы
Пояснение (Проба проводится в положении сидя).
1. Сделайте глубокий вдох и глубокий выдох.
2. После этого сделайте почти максимальный вдох и задержите дыхание. Начните отсчет.
3. Выключите секундомер, когда дыхание возобновится непроизвольно, и запишите результат.
4. Отдохните 5-7 минут.
5. Сделайте не очень глубокий выдох, задержите дыхание и тут же включите секундомер.
6. Выключите секундомер при непроизвольном восстановлении дыхания и запишите результат.
7. Оцените свои результаты.
Пояснение: (Результат задержки дыхания на вдохе считается удовлетворительным, если человек смог задержать дыхание на 16-55 сек. Более низкие результаты следует считать плохими, более высокие – хорошими.
Результат задержки дыхания на выдохе считается удовлетворительным, если он не ниже 12-13 сек).
Лабораторная работа №16
Действие ферментов слюны на крахмал
Цель: показать, что ферменты слюны способны расщеплять крахмал.
Оборудование: кусочек накрахмаленного бинта, вата, спички (ватные палочки), блюдце, вода, йод(5%).
Пояснение. Крахмал с йодом дает интенсивное синее окрашивание.
Ход работы.
1.Приготовьте реактив на крахмал — йодную воду. В блюдце налейте воду и добавьте несколько капель йода до получения жидкости цвета крепко заваренного чая.
2.Намотайте на спичку вату (можно взять ватную палочку), смочите ее слюной, а затем этой ватой со слюной напишите букву на накрахмаленном бинте.
3.Расправленный бинт зажмите в руках и подержите 1-2 минуты.
4.Опустите бинт в йодную воду, тщательно расправив его.
5.Наблюдайте, как окрасился бинт. Свои наблюдения запишите в тетрадь.
6. Сделайте вывод.
Объясните результаты опыта.
— Могла ли получиться синяя буква на белом фоне при проведении опыта?
— Будет ли слюна расщеплять крахмал, если ее прокипятить?
Лабораторная работа №17
Установление зависимости между нагрузкой и уровнем
энергетического обмена по результатам функциональной пробы с
задержкой дыхания до и после нагрузки.
Оборудование: секундомер или часы с секундной стрелкой.
Ход работы
1.В положении сидя задержите дыхание при вдохе на максимальный срок. Включите секундомер (предварительное глубокое дыхание перед опытом не допускается!).
2.Выключите секундомер в момент восстановления дыхания. Запишите результат (А). Отдохните 5 мин.
3.Встаньте и сделайте 20 приседаний за 30 с.
4.Вдохните, быстро задержите дыхание и включите секундомер, не дожидаясь, пока дыхание успокоится, сядьте на стул.
5.Выключите секундомер при восстановлении дыхания. Запишите результат(B).
6.Спустя минуту повторите первую пробу. Результат запишите(C).
7.Вычислите процентное отношение B/A X 100% и C/A X 100%
8.Оцените свои результаты (таблица стр.197 учебника).
9. Сделайте вывод.
10.Ответьте на вопросы.
— Почему происходит непроизвольное восстановление дыхания? Почему при задержке дыхания в крови накапливается углекислый газ?
— Как углекислый газ воздействует на дыхательный центр?- Почему после работы удается задержать дыхание на меньшее время, чем в состоянии покоя?
Лабораторная работа №18
Составление пищевых рационов в зависимости от энергозатрат.
Оборудование: калькулятор
Пояснения. При составлении пищевого рациона человека следует придерживаться следующих правил:
1)калорийность пищевого рациона должна соответствовать суточному расходу энергии;
2) необходимо учитывать оптимальное для лиц, занимающихся данным видам труда (а для детей — возраста), количество белков, жиров и углеводов;
3) наилучший режим питания предполагает четырехразовый прием пищи (первый завтрак должен составлять 10-15%, второй завтрак — 15-35%, обед-40 — 50% и ужин 15-20% от общей калорийности)
4) продукты, богатые белком (мясо, рыба, яйцо), рациональнее использовать для завтрака и обеда. На ужин следует оставлять молочно-растительные блюда;
На ужин следует оставлять молочно-растительные блюда;
5) в пищевом рационе около 30% должны составлять белки и жиры животного происхождения.
Потребность детей в калориях в зависимости от возраста
Потребление энергии в день( калорийность рациона), ккал
7-10
11-13
мальчики
девочки
14-17
юноши
девушки
2300
2700
2450
2900
2600
Калорийность пищевых продуктов.
ХЛЕБНЫЕ ИЗДЕЛИЯ, КРУПЫ, СЛАСТИХлеб: ржаной
пшеничный
батон
сухари
печенье
пирожные
пирожки печеные
макаронные изделия
Крупы: овсяная
гречневая, манная, рис
Сахар
Шоколад
Халва
Пастила, зефир
Варенье
МЯСО, РЫБА, ЯЙЦА
Мясо
Птица
Ветчина
Колбаса вареная
Сосиски
Судак, треска
Окунь
Икра
Яйцо 1 шт.
ЖИРОВЫЕ ПРОДУКТЫ
Масло: сливочное
растительное
Маргарин
МОЛОЧНЫЕ ПРОДУКТЫ
Молоко, простокваша, кефир
Сметана
Творог: жирный
нежирный
Сырки творожные сладкие
Сыры: твердые
плавленые
Молоко сгущенное с сахаром
ОВОЩИ, ФРУКТЫ ГРИБЫ, ЯГОДЫ, ОРЕХИ
Огурцы
Картофель
Морковь
Свекла
Капуста
Лук репчатый
Грибы
Ягоды (вишня, земляника, малина, крыжовник, смородина и др.)
Виноград
Бананы
Арбуз
Абрикосы, апельсины, лимоны
НАПИТКИ
Сладкие фруктовые воды
Хлебный квас
Минеральная вода
200
230
250
360
420
430
220
340
380
330
400
600
500
350
280
150
200
370
250
200
65
100
250
75
780
900
750
60
280
230
80
330
350
240
320
20
90
40
50
25
50
30
50
70
100
40
50
60
30
0
Ход работы
1. Составьте суточный пищевой рацион для вашей возрастной группы, используя таблицу калорийности пищевых продуктов.
Составьте суточный пищевой рацион для вашей возрастной группы, используя таблицу калорийности пищевых продуктов.
2. Результаты расчетов занесите в таблицу
Масса,г
Содержание
белков
Содержание
углеводов
Содержание
жиров
Калорийность
1-й завтрак
2-й завтрак
Обед Ужин
3. Сделайте выводы.
Лабораторная работа №19
Пальценосовая проба и особенности движений, связанных с функцией мозжечка и среднего мозга.
Ход работы
1). Закройте глаза. Вытяните вперед руку с выставленным указательным
пальцем. Коснитесь указательным пальцем кончика носа. Проделайте
Коснитесь указательным пальцем кончика носа. Проделайте
то же самое с левой рукой.
Пояснение. (Чтобы совершить это движение и попасть в цель необходимо рассчитать траекторию, определить последовательность
и время сокращения определенных мышечных групп, осуществляющих движение по заданной траектории. В этом движении участвуют 33
мышцы, каждая из которых должна в определенное время включиться и выйти из работы).
2). Сделайте вывод:
— Какая функция мозжечка была выявлена в этом эксперименте?
Лабораторная работа №20
Штриховое раздражение кожи – тест, определяющий
изменения тонуса симпатической и парасимпатической
автономной нервной системы при раздражении.
Пояснение. (Известно, что симпатические нервы сужают кровеносные сосуды кожи, а парасимпатические нервы их расширяют. )
)
Ход работы.
1. Ногтем проведите по коже. Почему вначале появляется белая полоска, а спустя некоторое время — красная? Объясните, почему через некоторое время эта полоска исчезает и никаких следов от раздражения не остается.
2. Сделайте вывод.
Лабораторная работа №21
Опыты, выявляющие иллюзии, связанные с
бинокулярным зрением.
Оборудование: трубка, свернутая из листа бумаги.
Ход работы.
Один конец трубки приставьте к правому глазу. Ко второму концу трубки приставьте левую руку так, чтобы трубка лежала между большим и указательным пальцами. Оба глаза открыты и должны смотреть вдаль. Если изображения, полученные в правом и левом глазах, попадут на соответствующие участки коры больших полушарий, возникает иллюзия- «дырка в ладони».
Лабораторная работа №22
Выработка навыков зеркального письма как пример разрушения
старого и образования нового динамического стереотипа.
Ход работы
1. Измерьте, сколько секунд потребуется, чтобы написать скорописью какое-либо слово, например «Психология». С правой стороны проставьте затраченное время.
2. Предложите испытуемому написать то же слово зеркальным шрифтом: справа налево. Писать надо так, чтобы все элементы букв были повернуты в противоположную сторону. Сделайте 10 попыток, около каждой из них с правой стороны проставьте время в секундах.
3. Постройте график. На оси X отложите порядковый номер попытки, на оси У — время, которое испытуемый потратил на написание очередного слова.
4. Посчитайте, сколько разрывов между буквами было при написании слова обычным способом, сколько разрывов стало при первой и последующих попытках написания слова справа налево.
5.Отметьте, в каких случаях возникают эмоциональные реакции: смех, жестикуляции, попытка бросить работу и др. 6. Назовите число букв, в которых встречаются элементы, написанные старым способом.
7. Проанализируйте полученный график. Встречаются ли моменты, где навык перестает вырабатываться, когда его результаты становятся хуже? Это случается всегда, когда образовавшаяся система связей исчерпывает себя и начинается новый поиск. Это случается несколько раз помимо нашего сознания и прекращается после того, как результаты станут стабильными, а динамический стереотип выработанным.
Сделайте выводы:
Какие факты говорят, что при разрушении динамического стереотипа происходит распад общей деятельности на отдельные элементы, например слово, написанное ранее одним росчерком, выписывается теперь по буквам?
Делаются ли при формировании нового динамического стереотипа попытки соединять буквы без дополнительной инструкции? Нужны ли эти инструкции для овладения приемами рационального письма?
В чем выражалась «борьба» между стереотипами — вновь создаваемым и старым, хорошо закрепленным? Об этом можно судить по наличию элементов букв, написанных по-старому.

Лабораторная работа №23
Измерение числа колебаний образа усеченной пирамиды при непроизвольном,
произвольном внимании и при активной работе с объектом.
Оборудование: секундомер или часы с секундной стрелкой.
Пояснения. Попытайтесь представить усеченную пирамиду ( рис 119, стр. 293), обращенную усеченным концом к вам и от вас. Когда оба образа будут сформированы, они станут сменять друг друга: пирамида будет казаться то обращенной к вам, то от вас. По числу колебаний этих образов можно судить об устойчивости внимания. Обычно измеряют число колебаний внимания в минуту. Для экономии времени можно измерить число колебаний за 30 с. И результат удвоить.
Ход работы
Опыт №1.
Определение устойчивости непроизвольного внимания
Смотрите на рисунок, не отрываясь от него в течение 30 с.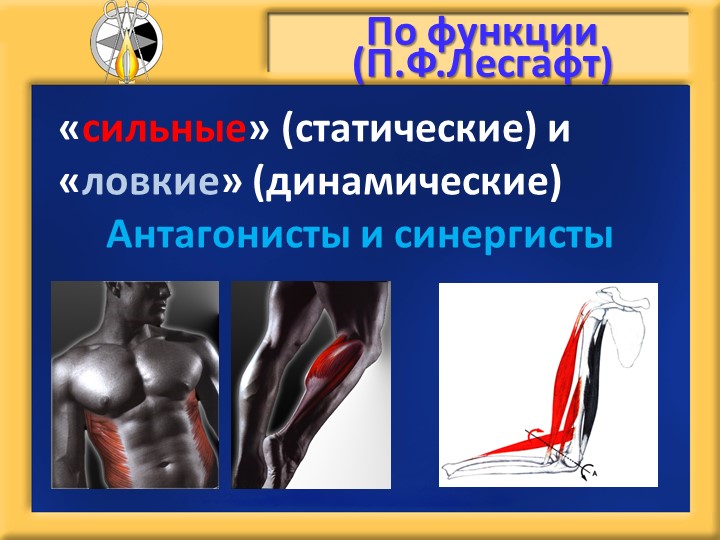 При каждом изменении образа делайте штрих в тетради. Число колебаний внимания за 30 с удвойте. Оба значения занесите в соответствующие графы таблицы.
При каждом изменении образа делайте штрих в тетради. Число колебаний внимания за 30 с удвойте. Оба значения занесите в соответствующие графы таблицы.
Опыт №2.
Удержание образа произвольным вниманием.
Повторите опыт, соблюдая ту же методику проведения, но постарайтесь как можно дольше удерживать тот образ, который сложился. Если же он все же изменился, надо удерживать новый образ как можно дольше. Подсчитайте число колебаний. Результаты занесите в таблицу.
Опыт №3
Определение устойчивости внимания при активной работе
с объектом.
Представьте себе, что рисунок изображает комнату. Маленький квадрат — ее задняя стенка. Подумайте, как расставить мебель: диван, кровать, телевизор, приемник и пр. Выполняйте эту работу в течение тех же 30 с. Не забывайте при каждом изменении образа делать штрих, причем каждый раз возвращайтесь к исходному образу и продолжайте «обставлять» комнату.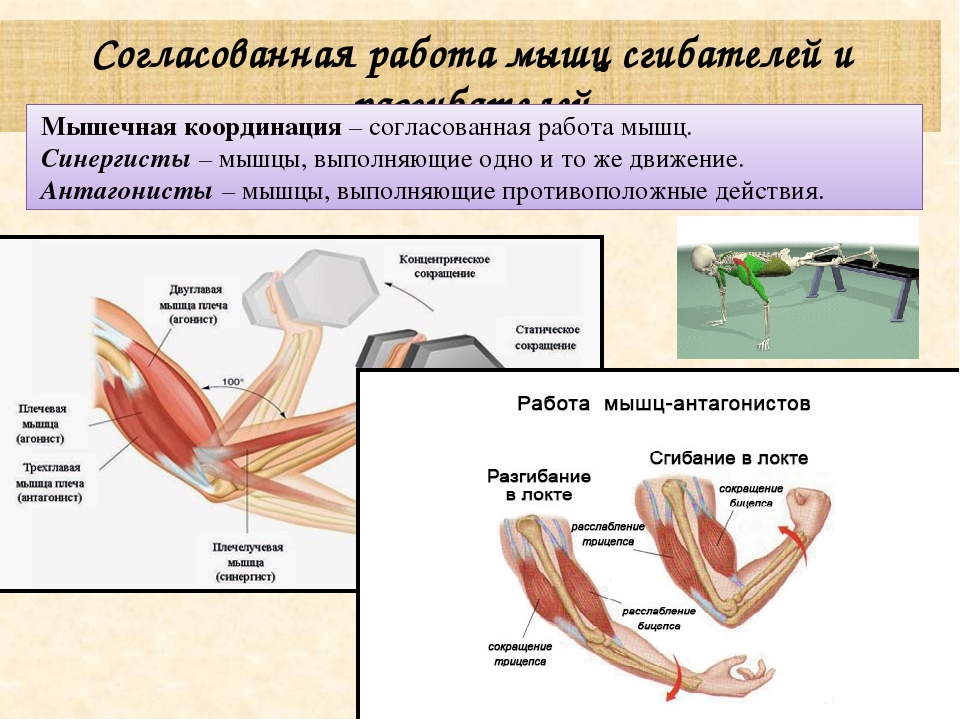 «Расставлять» мебель надо мысленно, не отрываясь от рисунка. Полученные результаты занесите в таблицу в соответствующие графы.
«Расставлять» мебель надо мысленно, не отрываясь от рисунка. Полученные результаты занесите в таблицу в соответствующие графы.
Произвольное внимание (с установкой сохранять
создавшийся образ)
Произвольное внимание при активной работе с объектом
Сделайте выводы: При каких условиях наблюдается наибольшее число колебаний внимания?
ВГУ имени П. М. Машерова, 2013. – 50 с.
19
(мышцы: трехглавая мышца голени, подошвенная, задняя большеберцовая,
длинный сгибатель большого пальца, длинный сгибатель пальцев, длинная ма-
лоберцовая, короткая малоберцовая), разгибание в коленном (мышцы: прямая
мышца бедра, латеральная широкая мышца бедра, медиальная широкая мышца
бедра, промежуточная широкая мышца бедра) и тазобедренном (мышцы: большая
ягодичная, двуглавая мышца бедра, полусухожильная, полуперепончатая, боль-
шая приводящая) суставах, разгибание позвоночного столба (мышцы: трапецие-
видная, верхняя и нижняя задние зубчатые мышцы, ременная мышца головы и
шеи, мышца, выпрямляющая позвоночник, поперечно-остистая мышца) и одно-
временный взмах руками кверху (мышцы: передняя часть дельтовидной, большая
грудная, клювовидно-плечевая, двуглавая мышца плеча, трехглавая мышца плеча,
локтевая), что способствует повышению положения ОЦТ тела. По законам балли-
По законам балли-
стики полет будет наиболее длинным, если прямая, показывающая направление
отталкивания, располагается под углом 45° к горизонтальной плоскости.
Чтобы в момент отталкивания в наибольшей мере использовать все мыш-
цы-сгибатели пальцев, обычно держат стопы в положении с несколько обращен-
ными внутрь носками. Работа мышц в фазе отталкивания характеризуется резко-
стью и силой. В течение короткого времени мышцы сокращаются до максимума,
в результате чего тело, подброшенное в воздух, проходит некоторое расстояние.
Главными мышцами, работающими при отталкивании, являются: в области сто-
пы – все мышцы ее подошвенной поверхности; в области голеностопного суста-
ва – задняя и латеральная группы мышц голени; в области коленного сустава –
бедренные головки четырехглавой мышцы бедра; в области тазобедренного сус-
тава – мышцы, расположенные на его задней поверхности; на туловище – мыш-
цы-разгибатели позвоночного столба, а также мышцы, поднимающие пояс верх-
ней конечности; в области верхней конечности – мышцы-сгибатели плеча, а
также мышцы-разгибатели предплечья.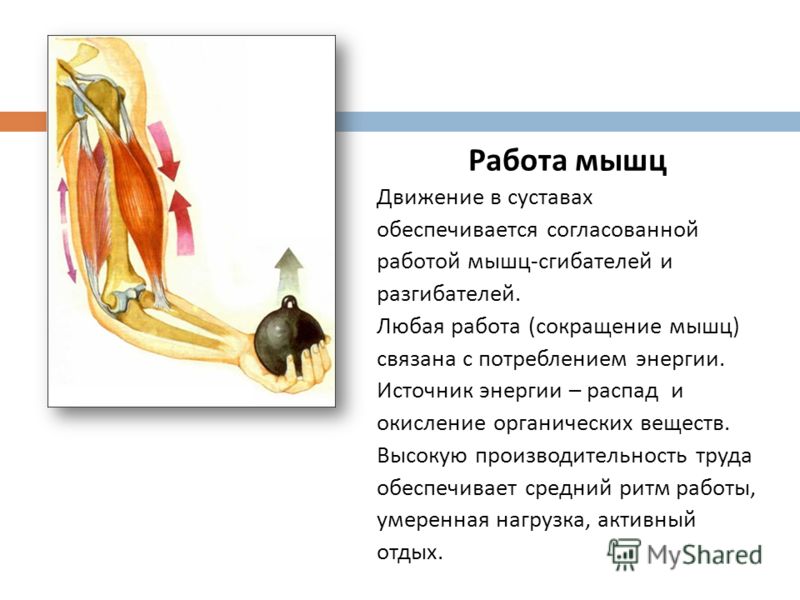 Большинство из них, в частности мыш-
Большинство из них, в частности мыш-
цы, расположенные на нижних конечностях и на туловище, находится в сокра-
щенном состоянии и в подготовительном периоде. Но в это время они выполня-
ют уступающую работу, а в момент отталкивания – преодолевающую.
После того как в суставах произошло почти полное разгибание, движения
в них, как уже говорилось, затормаживаются благодаря кратковременному со-
кращению мышц-антагонистов, превращающих все тело как бы в одно монолит-
ное целое, что способствует передаче сил толчка на ОЦТ тела и сохранению
равновесия во время полета. Роль мышц-антагонистов заключается также в том,
что они препятствуют переразгибанию звеньев нижней конечности в суставах,
предохраняя их от повреждения.
В ф а з а п о л е т а вначале тело наклонено вперед, постепенно выпрям-
ляется и затем отклоняется назад, ноги выносятся вперед, происходит сгибание в
коленном (мышцы: двуглавая мышца бедра, полусухожильная, полуперепонча-
тая, портняжная, тонкая, подколенная, икроножная, подошвенная) и тазобедрен-
ном (мышцы: портняжная, мышца-напрягатель широкой фасции, гребенчатая,
прямая мышца бедра) суставах, разгибание стопы (мышцы: передняя больше-
берцовая, длинный разгибатель пальцев, длинный разгибатель большого пальца)
и туловища (мышцы: трапециевидная, верхняя и нижняя задние зубчатые мыш-
цы, ременная мышца головы и шеи, мышца, выпрямляющая позвоночник, попе-
Репозиторий ВГУ
ДВИЖЕНИЕ и СКЕЛЕТНЫЕ МЫШЦЫ
ДВИЖЕНИЕ и СКЕЛЕТНЫЕ МЫШЦЫДВИЖЕНИЕ и СКЕЛЕТНЫЕ МЫШЦЫ
I Движение
А.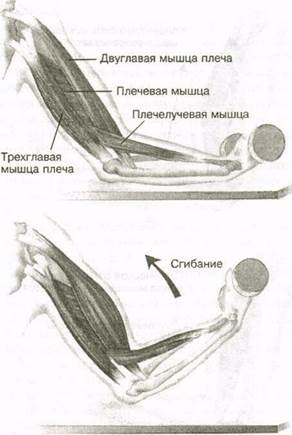 Требования
Требования
B. Происхождение и вставка
C. Мышца — мышечные взаимодействия: координация и контроль
1. противоположные пары
а.первичный двигатель (агонист)
б. антагонист
в. синергисты
1. фиксаторы
D. Взаимодействие мышц и костей: работа
1. рычажные системы
а. рычаг = кость
б. точка опоры = сустав
в.усилие (сила) = мышца
д. нагрузка
2. рычаги первого класса
а. точка опоры между усилием (силой) и нагрузкой (качели)
3. рычаги второго класса
а. нагрузка между усилие и точка опоры (тачка)
б.сила, меньше скорость и диапазон движения
4. рычаги третьего класса
а. усилие между нагрузкой и точкой опоры
б. быстрый, большой движения с небольшим усилием
в. чаще всего встречается в организме
Когда вы закончите этот модуль, вы должны уметь:
— перечислите, что необходим для обеспечения движения костей
— определять начало и место прикрепления мышцы
— идентифицировать начало и место прикрепления мышцы при наличии двух точек прикрепления
— определите мышцы могут играть разные роли: агонист, антагонист, синергист
— идентифицировать роль, которую выполняет мышца при определенном движении
— перечислить компоненты рычажной системы в целом и их аналоги в кузове
— Нарисовать Рычажная система 1-го, 2-го и 3-го класса
— сравнивать преимущества и различия трех рычажных систем
— идентифицировать пример каждого класса рычажных систем в кузове
II Мышечные возможности
А. Общие для всех типов мышц
Общие для всех типов мышц
B. Различия между типами мышц
III Структура скелетных мышц
А. Орган
1. скелет мышечная ткань
а. клетка: сарколемма, исчерченность, миофибриллы, миофиламенты
2. соединительная ткань
а.эндомизий (вокруг волокна)
б. перимизий (вокруг пучка)
в. эпимизий (вокруг всей мышцы)
д. глубокая фасция
е. сухожилие
3. нервная ткань
а. нервно-мышечное соединение
Б.Структура и расположение пучка
1. эффекты: движение и сила
2. типы
а. параллель
б. сходящийся
в. пеннатный (одно-, двух-, мульти-)
д. круговой (сфинктеры)
Когда вы закончите этот модуль, вы должны уметь:
— идентифицировать скелетные, сердечные и гладкие мышцы
1) по фотографии (или слайду!)
2) по конструктивным характеристикам
3) по функциональным характеристикам
— Нарисовать мышечное волокно и маркировка его основных частей
— указать причина полосатого внешнего вида и причина ядер на периферии волокна
— идентифицировать и опишите организацию мышцы от волокон до пучков до целого мышцы, включая соединительнотканные оболочки
— объяснять как мышца может тянуть кость, если она никогда не вступает в прямой контакт с вставка
— определять нервно-мышечное соединение и его расположение на мышечном волокне
— распознавать и опишите параллельное, конвергентное и перистое расположение мышц пучки
— обсуждать почему параллельное расположение обеспечивает большее движение
— обсуждать почему конвергентное расположение обеспечивает меньшую мощность и разнообразие движений
— обсуждать почему перистое расположение обеспечивает меньше движения и больше силы
— описывать сфинктеры и их роль в организме
IV Брюшная стенка
А. Маркировка поверхности
Маркировка поверхности
Б. Мышцы
1. прямая мышца живота
а. сухожильный перекрестки
2. наружная косая
3. внутренняя косая
4. поперечная мышца живота
C. Апоневрозы и белая линия
V Мышцы верхних конечностей
А.Наплечник
1. стабилизировать сустав (ротатор манжета)
а. все происхождение на лопатке
б. все вставки на плечевой кости
1. подлопаточная
2. подостная
3. малая круглая
4. надостная мышца
2. лопатка болотная
а. все исходники по оси скелет
б. все вставки на лопатку
1. trapezius (3 области)
2. поднимающая лопатку
3. ромбы
4. малая грудная мышца
5. передняя зубчатая мышца
3. переместить рычаг
а. все вставки на плечевой кости
1. дельты (3 зоны)
2. большая грудная
3. широчайшая мышца спины
широчайшая мышца спины
4. большая круглая
5. коракобрахиалис
Б. Рука
1. движение (в локте) предплечье
а. передний купе; сгибатели; мышечно-кожный нерв
1. двуглавая мышца плеча
2.брахиалис
3. (клювовидно-плечевой)
б. задний купе; разгибатели; лучевой нерв
1. трехглавая мышца плеча
C. Локтевая ямка
1. конструкции
а. плечевой артерия —-> лучевая и локтевая артерии
б.срединный нерв
в. двуглавый апоневроз
д. медиана локтевая вена
D. Предплечье
1. движение запястья и рука
а. передний купе; сгибатели; срединный нерв для большинства
б. плечелучевая; боковая сторона наружного отсека; сгибатель; лучевой нерв
в.задний купе; разгибатели; лучевой нерв
VI Мышцы нижних конечностей
А. Бедро
1. пошевелить бедром
а. происхождение на таз
б. вставки на
бедренная кость
вставки на
бедренная кость
1. подвздошно-поясничная мышца — сгибатель
2. большая ягодичная мышца
3. средняя ягодичная мышца
4. малая ягодичная мышца
2. отсеки
а. передний отсек — разгибатели голени (голени)
б. медиальный отсек — приводящие мышцы бедра
в. задний купе – поперечные два стыка; разгибатель бедра, согнутая нога
Б.Нога
1. передний отделение (расширение пальцев стопы, тыльное сгибание стопы)
2. боковое отделение (подошвенное сгибание, разворот стопы)
3. задний отсек (сгибатели)
В конце этого модуля вы должны быть в состоянии:
— определить некоторые маркировка на передней поверхности тела
— обсудить структурное расположение мышц живота и их роль в организме
— определить сухожилие, связка, апоневроз
— идентифицировать на схему и обсудите мышцы «вращательной манжеты плеча» (начало, прикрепление, индивидуальные действия, групповое действие),
— идентифицировать на нарисуйте схему и обсудите мышцы, прикрепляющие лопатку
— идентифицировать на схему и обсудите мышцы, приводящие в движение руку
— идентифицировать на схему и обсудите мышцы, приводящие в движение предплечье (в локтевом суставе)
— сравнить мышцы переднего отдела руки и заднего отдела руки
— Опишите структура локтевой ямки и структуры в ее пределах
— сравните функция мышц переднего и заднего отделов предплечья
— указать общее начало и место прикрепления мышц бедра
— обозначьте три отдела мышц бедра и сформулируйте общую функцию мышцы в каждом отделе
— обозначить три отделы голени и состояние общей функции мышц в каждом отделе
Вернуться к анатомические контуры
Вернуться к анатомия
Вернуться к домашняя страница
Последнее обновление: 05.
 01.22
01.22Совместное сокращение использует двойной контроль мышц-агонистов и антагонистов для улучшения двигательной активности
Производительность улучшается при совместном сокращении во время постуральной задачи робот Kinarm Exoskeleton
20 (Kinarm, Кингстон, Онтарио, Канада).Каждое испытание они начинали с перемещения пальца к центральной мишени. Импульс крутящего момента возмущения сгибает или разгибает локоть, смещая кончик пальца от цели. Участники успешно выполнили задание, если они возвращали кончик пальца к цели менее чем за 500 мс и удерживали его в этой области более 1000 мс. Задача выполнялась с применением фоновых крутящих моментов 1-5 Нм в локтевом суставе для предварительной активации либо разгибателей, либо сгибателей локтя (рис. 1а). Предварительно активированные мышцы считаются здесь агонистами.Участники также выполняли задание без фоновой нагрузки. В этом случае испытуемый начинал каждое испытание с совместного сокращения мышц-сгибателей и разгибателей локтевого сустава до уровней активности, соответствующих фоновым крутящим моментам 1, 3 или 5 Нм, используя визуальную обратную связь (рис. 1b). Рисунок 1:
1b). Рисунок 1: (a) Задача постурального возмущения с фоновой нагрузкой, которая предварительно активирует разгибатели или сгибатели локтевого сустава. Участники должны удерживать кончик пальца в неподвижной мишени, сопротивляясь фоновой нагрузке. Применяется импульс крутящего момента возмущения, и участники должны вернуть кончик пальца к цели как можно быстрее.(b) Задача постурального возмущения с целенаправленным совместным сокращением сгибателей и разгибателей локтевого сустава. Участники должны совместно сокращаться до желаемого уровня мышечной активности, используя визуальную обратную связь, удерживая кончик пальца в неподвижной цели. Применяется импульс крутящего момента возмущения, и участники должны вернуть кончик пальца к цели как можно быстрее. (c) Задача отслеживания цели. Участники должны отслеживать движущуюся цель кончиком пальца. Применяется импульс крутящего момента возмущения, и участники должны вернуть кончик пальца к движущейся цели как можно быстрее.Пробы выполняются с активным совместным сокращением сгибателей и разгибателей локтевого сустава и без него.
Мы обнаружили явное улучшение показателей участников, когда они сокращались до постурального нарушения по сравнению с тем, когда они противостояли фоновым нагрузкам (рис. 2а). Состояние нагрузки и уровень мышечной активности оказали значительное влияние на успешность выполнения задачи без значимого взаимодействия (F(условие) = 25, p = 2,1e-6; F(уровень) = 4,2, p = 0,028; F(условие x уровень). ) = 2.3, р = 0,071). Успешность задачи составила 56% при самом низком уровне совместного сокращения и увеличилась до 71% при самом высоком уровне (p = 0,021, парный t-критерий между участниками). Напротив, успешность задачи составляла всего ~35%, когда группа мышц, предварительно активированная фоновыми нагрузками (группа мышц-агонистов), растягивалась или сокращалась в результате возмущения. Было небольшое увеличение успеха до 45% для агонистов в длительных испытаниях при самом высоком уровне фоновой нагрузки. Повышение производительности при совместном выполнении по сравнению с противодействием фоновым нагрузкам (дополнительно от 23% до 39% от общего числа успешно завершенных испытаний) было статистически значимым (p = 0. 013 для совместного сокращения по сравнению с агонистами, растянутыми при низкой мышечной активности, p < 0,01 для совместного сокращения по сравнению со всеми остальными, парные t-тесты между участниками).
013 для совместного сокращения по сравнению с агонистами, растянутыми при низкой мышечной активности, p < 0,01 для совместного сокращения по сравнению со всеми остальными, парные t-тесты между участниками).
(a) Уровень успешности выполнения задачи постурального возмущения у всех испытуемых для трех экспериментальных условий нагрузки: растяжение агониста (треугольник), сокращение агониста (квадрат) и совместное сокращение (круг). (b) Относительные изменения в успехе задачи постурального возмущения с совместным сокращением по сравнению с предварительной активацией одной группы мышц.Красные точки обозначают отдельные предметы. Эквивалентная нагрузка указывает на уровень фоновой мышечной активности от сопротивления фоновым нагрузкам или совместного сокращения.
Одиннадцать из двенадцати участников улучшили производительность на всех трех уровнях мышечной активности при совместном сокращении по сравнению с тем, когда агонисты растягивались или укорачивались (рис. 2b). Было достигнуто относительное повышение производительности до 350 % при среднем улучшении производительности примерно на 100 %.
Было достигнуто относительное повышение производительности до 350 % при среднем улучшении производительности примерно на 100 %.
Характер корректирующих реакций при совместном сокращении участников явно отличался от того, когда предварительно активировалась только одна группа мышц (рис. 3).Уровень фоновой мышечной активности оказал значительное влияние на пиковое смещение с дополнительным эффектом взаимодействия от состояния нагрузки (F(условие) = 2,8, p = 0,083; F(уровень) = 155, p = 1,1e-13; F(условие x уровень). ) = 20, р = 2,5е-9). Условия нагрузки, уровень активности и их взаимодействие оказывали значительное влияние на время возврата после возмущения (F(условие) = 26, p = 1,7e-6; F(level) = 28, p = 9,3e-7; F(уровень) = 28, p = 9,3e-7; состояние х уровень) = 3,1, p = 0,026).
Рисунок 3: Изменение угла локтевого сустава после механических возмущений для трех экспериментальных условий нагрузки: (а) растяжение агониста, (б) сокращение агониста и (в) совместное сокращение. Уровень фоновой мышечной активности обозначен цветом линии, а начало возмущения — 0 мс (серая рамка). Показаны групповые результаты (среднее значение и стандартная ошибка).
Уровень фоновой мышечной активности обозначен цветом линии, а начало возмущения — 0 мс (серая рамка). Показаны групповые результаты (среднее значение и стандартная ошибка).
Подробный анализ изменений в движении локтевого сустава выявляет несколько критических различий в движении конечностей после возмущений. Во-первых, наблюдались существенные различия в характере движения локтевого сустава даже при самом низком уровне нагрузки независимо от того, были ли агонисты растянуты, укорочены или все мышцы сокращались одновременно (синие линии на рис. 3).Максимальное смещение было сходным, когда предварительно активировалась только одна группа мышц, тогда как при их совместном сокращении оно было на 14% (2,0 градуса) меньше, чем при растяжении агонистов (p = 0,21, парный t-критерий между участниками) и 23 % (3,5 градуса) меньше, чем при укорочении агонистов (p = 0,0027, парный t-критерий между участниками). Когда агонисты были растянуты, наблюдалось существенное превышение, которое не происходит, когда агонисты укорачиваются или когда одновременно сокращаются группы мышц-сгибателей и разгибателей. Хотя локоть возвращался к целевому углу примерно в одно и то же время для последних двух условий, была большая вариабельность, когда агонисты были укорочены возмущением (стандартное отклонение 1,3 градуса), чем при совместном сокращении (стандартное отклонение 0,5 градуса). вероятно, приводит к большему успеху в испытаниях на совместное сокращение (p = 1,2e-4, F-тест для двух образцов с использованием угла локтя через 500 мс после возмущения).
Хотя локоть возвращался к целевому углу примерно в одно и то же время для последних двух условий, была большая вариабельность, когда агонисты были укорочены возмущением (стандартное отклонение 1,3 градуса), чем при совместном сокращении (стандартное отклонение 0,5 градуса). вероятно, приводит к большему успеху в испытаниях на совместное сокращение (p = 1,2e-4, F-тест для двух образцов с использованием угла локтя через 500 мс после возмущения).
Второй важной кинематической особенностью было небольшое изменение в начальном движении локтя после возмущения для различных фоновых нагрузок или уровней совместного сокращения (вставка на рис. 3).Самая большая разница наблюдалась, когда агонисты были укорочены, что приводило к уменьшению подвижности конечности на 21% через 50 мс. Однако разница в движении во время совместного сокращения составила всего 5%. Это говорит о том, что внутренние свойства мышц влияют на начальное движение конечности, но эффект в этом коротком временном масштабе ограничен.
В-третьих, увеличение фоновой нагрузки привело к явному снижению пикового смещения. Пиковое смещение уменьшилось на 30–45 % от самого низкого до самого высокого уровня фоновой активности в зависимости от состояния нагрузки (p < 0.01, парные t-тесты между участниками). При самых высоких уровнях нагрузки максимальное смещение было одинаковым, независимо от того, были ли агонисты растянуты, укорочены ли агонисты или одновременно сокращались обе группы мышц.
В-четвертых, наиболее критическая разница в условиях нагрузки была связана с наличием или отсутствием перерегулирования в конце корректирующей реакции. Перерегулирование уменьшалось по мере увеличения фоновой нагрузки при растяжении агонистов, тогда как оно увеличивалось при укорочении агонистов.Увеличение совместного сокращения привело к несколько более быстрому возврату к целевому углу (уменьшение времени возврата на 200 мс между низким и высоким уровнями мышечной активности, p = 0,019), при сохранении минимального перерегулирования.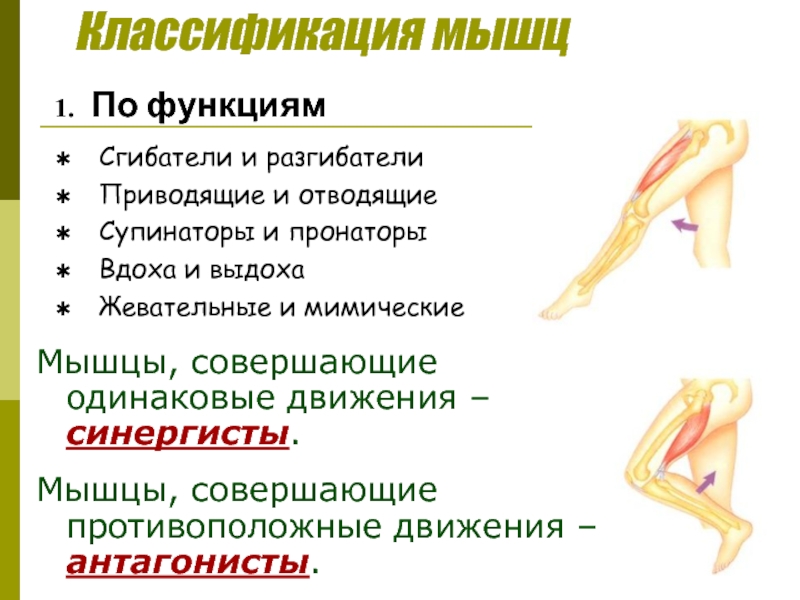 Таким образом, совместное сокращение приводило к простой стереотипной схеме двигательной коррекции, тогда как предварительная активация только одной группы мышц приводила к усложнению двигательной коррекции в зависимости от уровня фоновой мышечной активности.
Таким образом, совместное сокращение приводило к простой стереотипной схеме двигательной коррекции, тогда как предварительная активация только одной группы мышц приводила к усложнению двигательной коррекции в зависимости от уровня фоновой мышечной активности.
Критическое различие между совместным сокращением и другими условиями нагрузки заключалось в характере мышечной активности во время двигательной коррекции (рис. 4).Когда мышцы-агонисты растягивались, они реагировали всплеском активности в течение долгой латентной эпохи. По мере увеличения фоновой активности агонистов также происходил более ранний всплеск активности в течение короткой латентной эпохи. Величина этого всплеска увеличивалась с уровнем фоновой мышечной активности, обычно называемой масштабированием усиления 21–25 . Важно отметить, что при всех уровнях фоновой активности двигательная реакция в мышцах-антагонистах практически отсутствовала.
Рисунок 4: Активность латеральной головки трицепса и бицепса после механических воздействий, вызывающих сгибание в локтевом суставе.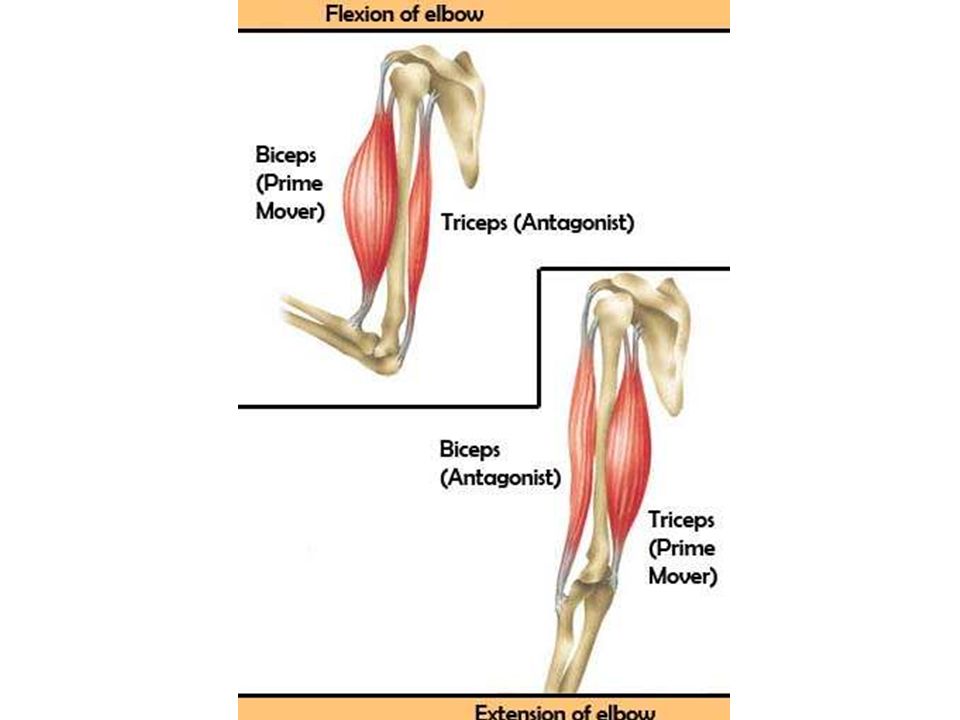 (а) Растянутый трицепс, предварительно активированный фоновой нагрузкой (растяжка-агонист). (b) Укороченный бицепс, предварительно активированный фоновой нагрузкой (укорочение агониста). (c) Мышцы сокращаются совместно. Увеличение активности растянутого трицепса отложено в положительном направлении, а увеличение активности укороченного бицепса — в отрицательном. Уровень фоновой мышечной активности обозначен цветом линии, а начало возмущения приходится на 0 мс. Показаны групповые результаты (среднее значение и стандартная ошибка). См. Дополнительный материал для получения дополнительных показателей мышечной активности.
(а) Растянутый трицепс, предварительно активированный фоновой нагрузкой (растяжка-агонист). (b) Укороченный бицепс, предварительно активированный фоновой нагрузкой (укорочение агониста). (c) Мышцы сокращаются совместно. Увеличение активности растянутого трицепса отложено в положительном направлении, а увеличение активности укороченного бицепса — в отрицательном. Уровень фоновой мышечной активности обозначен цветом линии, а начало возмущения приходится на 0 мс. Показаны групповые результаты (среднее значение и стандартная ошибка). См. Дополнительный материал для получения дополнительных показателей мышечной активности.
Когда мышцы-агонисты были укорочены, они угнетались и на короткое время снижались до минимальной активности для всех уровней фоновой нагрузки. Активность снова увеличилась со 120 мс до 350 мс после возмущения. Это увеличение коррелировало с уровнем фоновой активности агонистов. При всех уровнях фоновой агонистической активности мышцы-антагонисты проявляли минимальную реакцию на возмущение.
Наконец, при совместном сокращении корректирующая реакция задействовала как растянутые, так и укороченные мышцы.Реакция растянутых мышц была качественно такой же, как и при растянутом состоянии агонистов. Укороченные мышцы реагировали торможением в течение долгой латентной эпохи, которая масштабировалась с фоновой активностью. Было внезапное увеличение активности после торможения, которое произошло между 90 мс и 170 мс после возмущения. Этот всплеск масштабировался с фоновой активностью и был инициирован до смены направления руки, что, вероятно, предотвратило перерегулирование при возвращении к цели 26 .Кроме того, по-видимому, наблюдается небольшой всплеск активности в укороченных мышцах в период короткой латентности, что указывает на то, что мышцы могут рефлекторно сокращаться вместе после возмущения 27 . Однако это рефлекторное совместное сокращение присутствовало не у всех субъектов или мышц.
Модель для изучения вклада импеданса и контроля с обратной связью
Мы разработали модель, которая отражает два важных аспекта совместного сокращения: повышенный механический импеданс и контроль обратной связи растянутых и укороченных мышц (двойной контроль агонист-антагонист). Модель состоит из жесткого звена с шарнирным соединением на одном конце, которое представляет собой предплечье, кисть и локоть. Два торсионных привода действуют как разгибатели и сгибатели локтевого сустава. Эти исполнительные механизмы действуют в противоположных направлениях, могут генерировать только тяговые моменты и управляются приближением первого порядка динамики активации 28 . Приводы управляются оптимальным контроллером с обратной связью, предназначенным для возврата звена в исходное положение после импульсного возмущения. Совместное сокращение добавляется к модели двумя способами.Во-первых, жесткость и демпфирование в штифтовом суставе увеличиваются при более высоких уровнях совместного сокращения в зависимости от свойств силы-длины и силы-скорости мышц 28,29 . Во-вторых, базовая активность исполнительных механизмов увеличивается, что позволяет увеличивать, а также уменьшать до нуля активность для противодействия возмущению.
Модель состоит из жесткого звена с шарнирным соединением на одном конце, которое представляет собой предплечье, кисть и локоть. Два торсионных привода действуют как разгибатели и сгибатели локтевого сустава. Эти исполнительные механизмы действуют в противоположных направлениях, могут генерировать только тяговые моменты и управляются приближением первого порядка динамики активации 28 . Приводы управляются оптимальным контроллером с обратной связью, предназначенным для возврата звена в исходное положение после импульсного возмущения. Совместное сокращение добавляется к модели двумя способами.Во-первых, жесткость и демпфирование в штифтовом суставе увеличиваются при более высоких уровнях совместного сокращения в зависимости от свойств силы-длины и силы-скорости мышц 28,29 . Во-вторых, базовая активность исполнительных механизмов увеличивается, что позволяет увеличивать, а также уменьшать до нуля активность для противодействия возмущению.
Мы хотели изучить индивидуальный вклад повышенного механического импеданса и повышенной фоновой активности, связанных с ко-сокращением.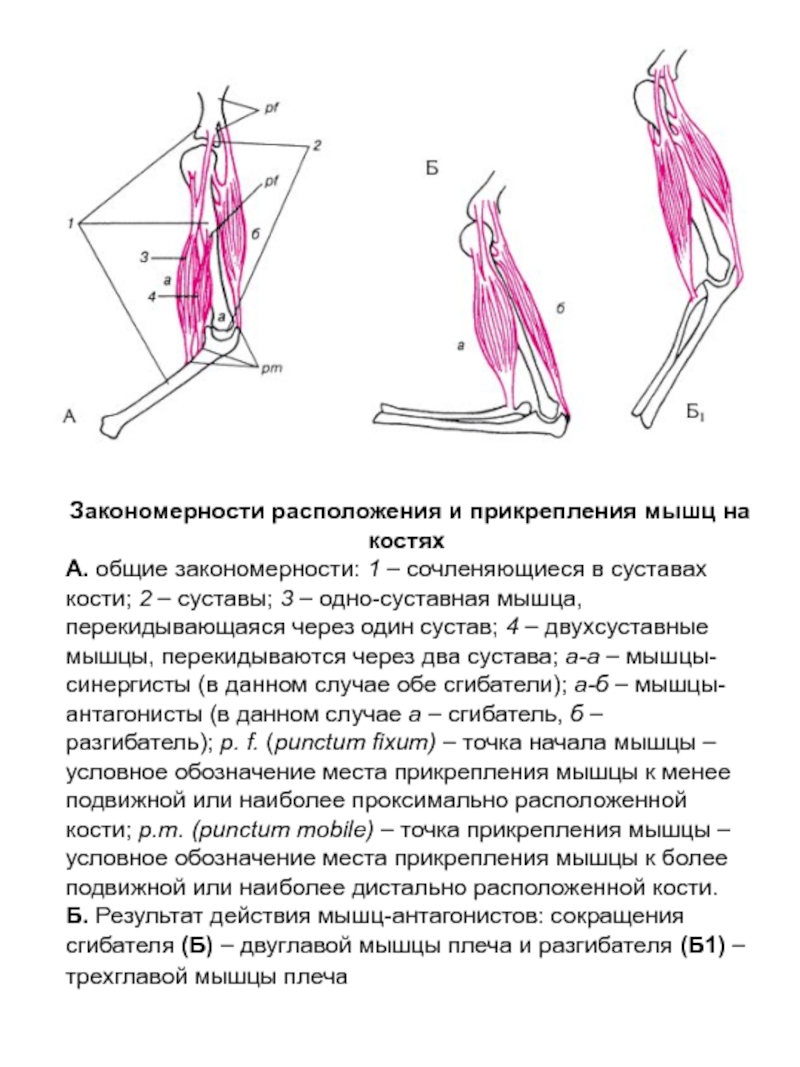 Влияние импеданса без обратной связи было зафиксировано пассивным моделированием (рис. 5а).Увеличение импеданса уменьшило максимальное смещение, перерегулирование при возврате к цели и время возврата системы. Моделирование с контролем с обратной связью и базовой активностью мышц-агонистов и антагонистов при фиксированном мышечном импедансе выявило эффект стратегии двойного контроля агонист-антагонист (рис. 5b). Двойное управление значительно уменьшило максимальное смещение системы и устранило превышение цели даже без увеличения импеданса. Важно отметить, что хорошие результаты наблюдались даже при самом низком уровне совместного сокращения.Наконец, когда импеданс был увеличен с базовой активностью (рис. 5c), производительность немного улучшилась, что обеспечило немного более быстрое время возврата.
Влияние импеданса без обратной связи было зафиксировано пассивным моделированием (рис. 5а).Увеличение импеданса уменьшило максимальное смещение, перерегулирование при возврате к цели и время возврата системы. Моделирование с контролем с обратной связью и базовой активностью мышц-агонистов и антагонистов при фиксированном мышечном импедансе выявило эффект стратегии двойного контроля агонист-антагонист (рис. 5b). Двойное управление значительно уменьшило максимальное смещение системы и устранило превышение цели даже без увеличения импеданса. Важно отметить, что хорошие результаты наблюдались даже при самом низком уровне совместного сокращения.Наконец, когда импеданс был увеличен с базовой активностью (рис. 5c), производительность немного улучшилась, что обеспечило немного более быстрое время возврата.
Результаты моделирования (изменение угла локтевого сгиба после механических возмущений), демонстрирующие влияние внутреннего импеданса мышц и контроля с обратной связью. (а) Пассивное моделирование, в котором импеданс увеличивается при совместном сокращении, но нет обратной связи. Обратите внимание, что траектория отклика на самом низком уровне импеданса выходит за пределы осей, чтобы поддерживать постоянный масштаб на графиках.(b) Моделирование, в котором присутствует контроль с обратной связью и базовая активность увеличивается при одновременном сокращении, но импеданс остается на самом низком уровне. (c) Моделирование как с управлением с обратной связью, так и с увеличением импеданса при совместном сокращении. Цвет линии представляет собой эквивалентный уровень мышечной активности, используемый для установки базовой активности и/или импеданса. Показаны среднее значение и стандартное отклонение 20 симуляций.
Обратите внимание, что траектория отклика на самом низком уровне импеданса выходит за пределы осей, чтобы поддерживать постоянный масштаб на графиках.(b) Моделирование, в котором присутствует контроль с обратной связью и базовая активность увеличивается при одновременном сокращении, но импеданс остается на самом низком уровне. (c) Моделирование как с управлением с обратной связью, так и с увеличением импеданса при совместном сокращении. Цвет линии представляет собой эквивалентный уровень мышечной активности, используемый для установки базовой активности и/или импеданса. Показаны среднее значение и стандартное отклонение 20 симуляций.
Мы хотели проверить преимущества наличия двух активных групп мышц для контроля по сравнению с тем, когда была активна только одна группа мышц.В каждом случае контроллер был оптимизирован, предполагая, что оба исполнительных механизма вносят свой вклад в реакцию обратной связи. Моделирование, в котором только растянутый (рис. 6ad) или укороченный (рис. 6be) актуатор может реагировать на возмущение, и моделирование, в котором оба могут реагировать (рис. 6cf), представляют собой растянутые агонисты, укороченные агонисты и экспериментальные условия совместного сжатия соответственно. Во всех трех случаях импеданс увеличивался в соответствии с уровнем фоновой активности. Результаты этих симуляций имеют много общих ключевых характеристик с экспериментальными результатами.
6be) актуатор может реагировать на возмущение, и моделирование, в котором оба могут реагировать (рис. 6cf), представляют собой растянутые агонисты, укороченные агонисты и экспериментальные условия совместного сжатия соответственно. Во всех трех случаях импеданс увеличивался в соответствии с уровнем фоновой активности. Результаты этих симуляций имеют много общих ключевых характеристик с экспериментальными результатами.
Изменение угла колена после механических возмущений при моделировании, где (а) реагирует только растянутый привод, (б) реагирует только укороченный привод и (в) реагируют оба привода. Соответствующие действия привода показаны на (d-f). Увеличение активации растянутого исполнительного механизма отображается в положительном направлении, а увеличение активации укороченного исполнительного механизма — в отрицательном направлении. Цвет линии представляет собой эквивалентный уровень мышечной активности, используемый для установки базовой активности и импеданса.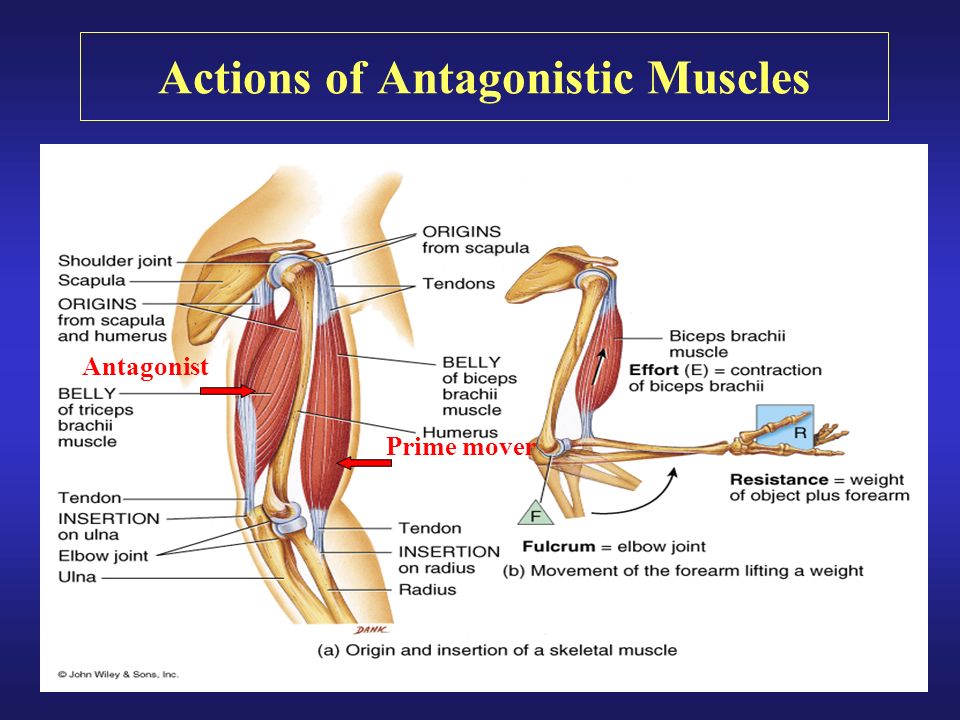 Показаны среднее значение и стандартное отклонение 20 симуляций.
Показаны среднее значение и стандартное отклонение 20 симуляций.
Наиболее важным результатом является то, что управление обоими исполнительными механизмами (двойное управление агонистом-антагонистом) устранило перерегулирование и колебания, которые возникали, когда был активен только один исполнительный механизм. Кроме того, увеличение фоновой активности уменьшило максимальное смещение во всех условиях и уменьшило перерегулирование при растяжении единственного активного исполнительного механизма.
Паттерны актуаторной активности в симуляциях были качественно аналогичны паттернам мышечной активности, использованным участниками (рис. 6d-f).При предварительной активации растянутый исполнительный механизм ответил всплеском активности, за которым последовало подавление. На самом низком базовом уровне это подавление было обрезано из-за ограничения, согласно которому привод мог создавать крутящий момент только в одном направлении. Когда укороченный привод был предварительно активирован, он реагировал ингибированием, которое обрезалось при нулевой активности для более низких исходных уровней. Когда оба исполнительных механизма были предварительно активированы, их реакции следовали тем же схемам, что и при индивидуальной предварительной активации.Однако с обоими активными клиппирование было менее распространенным и, в основном, наблюдалось лишь кратковременно в укороченном актуаторе на самом низком базовом уровне. В то время как модель фиксирует многие особенности двигательной коррекции человека, следует отметить, что, в отличие от эксперимента, величина взрыва растянутого привода не увеличивалась с базовой активацией. Кроме того, перерегулирование не увеличивалось с базовой активацией, когда реагировал только укороченный привод.
Когда оба исполнительных механизма были предварительно активированы, их реакции следовали тем же схемам, что и при индивидуальной предварительной активации.Однако с обоими активными клиппирование было менее распространенным и, в основном, наблюдалось лишь кратковременно в укороченном актуаторе на самом низком базовом уровне. В то время как модель фиксирует многие особенности двигательной коррекции человека, следует отметить, что, в отличие от эксперимента, величина взрыва растянутого привода не увеличивалась с базовой активацией. Кроме того, перерегулирование не увеличивалось с базовой активацией, когда реагировал только укороченный привод.
Производительность улучшается при совместном сокращении во время задачи отслеживания
Мы использовали задачу отслеживания возмущения, чтобы проверить, улучшает ли совместное сокращение производительность во время движения.В этом задании участники начинали каждое испытание, перемещая кончик пальца к неподвижной мишени (рис. 1с). Мишень начинала двигаться с постоянной скоростью по траектории, требующей лишь сгибания или разгибания локтя. Как только конец траектории был достигнут, цель изменила направление и по той же траектории вернулась к исходной точке. В случайный момент времени, когда цель находилась вблизи центра траектории, импульс крутящего момента возмущения (5 Нм, 50 мс) сгибал или разгибал локоть, смещая кончик пальца от цели.Это создало четыре условия возмущения: движение разгибания с возмущением сгибания, движение разгибания с возмущением разгибания, движение сгибания с возмущением разгибания и движение сгибания с возмущением сгибания. Участникам было предложено выполнить задание с активным антагонистическим совместным сокращением мышц, охватывающих локоть, и без него.
Как только конец траектории был достигнут, цель изменила направление и по той же траектории вернулась к исходной точке. В случайный момент времени, когда цель находилась вблизи центра траектории, импульс крутящего момента возмущения (5 Нм, 50 мс) сгибал или разгибал локоть, смещая кончик пальца от цели.Это создало четыре условия возмущения: движение разгибания с возмущением сгибания, движение разгибания с возмущением разгибания, движение сгибания с возмущением разгибания и движение сгибания с возмущением сгибания. Участникам было предложено выполнить задание с активным антагонистическим совместным сокращением мышц, охватывающих локоть, и без него.
Мы обнаружили явное улучшение показателей участников, когда они заключили совместный контракт (рис. 7). Совместное сокращение уменьшило отклонения от желаемого движения локтя, вызванные возмущением, независимо от направления движения и возмущения, и устранило перерегулирование в конце корректирующей реакции.Было небольшое уменьшение (12%) в начальном (50 мс) движении локтя после возмущения при совместном сокращении.
Углы в локтях во время повторяющихся движений слежения наружу и назад при механических возмущениях образцом субъекта (а) без и (б) с активным совместным сокращением. (c) Отклонение угла локтя от целевой траектории после механических возмущений без (синий) и с (красный) активным совместным сокращением. Начало возмущения при 0 мс (серый прямоугольник). Показаны групповые результаты (среднее значение и стандартная ошибка).(d) Максимальное расстояние от кончика пальца до цели и (e) время, чтобы вернуться к цели после механических возмущений без (синий) и с (красный) активного совместного сокращения. Показаны групповые результаты (среднее значение и стандартное отклонение).
Сосокращение, состояние возмущения и их взаимодействие оказали существенное влияние на максимальное смещение кончика пальца (F(сосокращение) = 287, p = 3,9e-8; F(условие) = 24, p = 8,1 e-8; F(совместное сокращение x условие) = 21, p 2.6e-7). На время возврата значительно влияло только совместное сокращение (F(совместное сокращение) = 81, p = 8. 4д-6; F(состояние) = 0,53, р = 0,66; F(совместное сокращение x условие) = 0,17, p = 0,92). Максимальное смещение и время возврата уменьшились на 50–57 % при совместном сокращении в зависимости от условий возмущения (p < 0,001 для всех условий, парные t-тесты между участниками).
4д-6; F(состояние) = 0,53, р = 0,66; F(совместное сокращение x условие) = 0,17, p = 0,92). Максимальное смещение и время возврата уменьшились на 50–57 % при совместном сокращении в зависимости от условий возмущения (p < 0,001 для всех условий, парные t-тесты между участниками).
При выполнении задачи без совместного сокращения реакция на возмущение почти полностью ограничивалась растянутой мышцей, что вызывало всплеск активности в эпоху долгой латентности (рис. 8, синие кривые).Укороченная мышца практически не давала ответа. Напротив, при совместном сокращении двигательная реакция включала как укороченные, так и растянутые мышцы (рис. 8, красные линии). Растянутая мышца вызывала более выраженную короткую и длительную латентную реакцию, чем в условиях отсутствия совместного сокращения. Кроме того, укороченная мышца вызывала торможение в течение периода длинной латентности, за которым следовал всплеск активности, начинающийся примерно через 100 мс после возмущения, который длился от 100 мс до 200 мс. Как и в статической задаче, этот всплеск произошел до поворота руки, что, вероятно, предотвратило промах при возвращении к цели.
Как и в статической задаче, этот всплеск произошел до поворота руки, что, вероятно, предотвратило промах при возвращении к цели.
Активность бицепса (верхний ряд) и трицепса (нижний ряд) во время задачи отслеживания цели с возмущениями сгибания (первый столбец) и разгибания (второй столбец). Групповые результаты (среднее значение и стандартная ошибка) показаны для испытаний без (синий) и с (красный) активным совместным сокращением. Начало возмущения при 0 мс.
Спинальный путь между синергистами может модулировать активность мышц-сгибателей локтевого сустава человека
Abstract
Было показано, что электрическая стимуляция плечелучевой ветви лучевого нерва ингибирует разряд произвольно активированных двигательных единиц в двуглавой мышце плеча во время слабых сокращений с помощью мышцы-сгибатели локтя.Целью настоящего исследования было охарактеризовать тормозной рефлекс путем сравнения его силы в короткой и длинной головках двуглавой мышцы плеча и изучения влияния положения предплечья на силу рефлекса. Для оценки влияния стимуляции лучевого нерва на разряд одиночных двигательных единиц в двуглавой мышце плеча у 15 человек использовали импульсную стимуляцию. Раздражение лучевого нерва удлиняло межспайковый интервал ( P < 0,001) двигательных единиц в длинном ( n = 31, 4.8 ± 5,6 мс) и короткие головки ( n = 26, 8,1 ± 12,3 мс) двуглавой мышцы плеча без разницы между двумя головками ( P = 0,11). Сила торможения варьировала в зависимости от положения предплечья для двигательных единиц обеих голов ( n = 18, P < 0,05). Величина торможения была наибольшей в пронации (7,9 ± 8,9 мс), средней в нейтральной (5,8 ± 7,1 мс) и наименьшей в супинации (2,8 ± 3,4 мс). Эти результаты показывают, что торможение, вызванное афферентной обратной связью от плечелучевой мышцы к низкопороговым двигательным единицам (средняя сила 3–5% MVC) в двуглавой мышце плеча, варьировалось в зависимости от положения предплечья, но было одинаковым для двух головок двуглавой мышцы плеча.
Для оценки влияния стимуляции лучевого нерва на разряд одиночных двигательных единиц в двуглавой мышце плеча у 15 человек использовали импульсную стимуляцию. Раздражение лучевого нерва удлиняло межспайковый интервал ( P < 0,001) двигательных единиц в длинном ( n = 31, 4.8 ± 5,6 мс) и короткие головки ( n = 26, 8,1 ± 12,3 мс) двуглавой мышцы плеча без разницы между двумя головками ( P = 0,11). Сила торможения варьировала в зависимости от положения предплечья для двигательных единиц обеих голов ( n = 18, P < 0,05). Величина торможения была наибольшей в пронации (7,9 ± 8,9 мс), средней в нейтральной (5,8 ± 7,1 мс) и наименьшей в супинации (2,8 ± 3,4 мс). Эти результаты показывают, что торможение, вызванное афферентной обратной связью от плечелучевой мышцы к низкопороговым двигательным единицам (средняя сила 3–5% MVC) в двуглавой мышце плеча, варьировалось в зависимости от положения предплечья, но было одинаковым для двух головок двуглавой мышцы плеча. Этот рефлекторный путь обеспечивает механизм регулирования активации двуглавой мышцы плеча при изменении положения предплечья и представляет собой спинальную основу для мышечной синергии у людей.
Этот рефлекторный путь обеспечивает механизм регулирования активации двуглавой мышцы плеча при изменении положения предплечья и представляет собой спинальную основу для мышечной синергии у людей.
Ключевые слова: Двигательная единица, Торможение, Двуглавая мышца плеча, Плечево-лучевая мышца, Синергия
Введение
Момент сгибания в локтевом суставе создается активацией ряда мышц, которые пересекают локтевой сустав и прикрепляются к лучевой или локтевой кости. Из-за этих мест прикрепления большинство мышц, которые вносят вклад в крутящий момент сгибателя локтя, также производят действия относительно других осей вращения.Вследствие такой анатомической организации некоторые мышцы являются синергистами для вращательного момента сгибателей локтевого сустава, но являются антагонистами для других действий. Одним из примеров такого расположения является двуглавая мышца плеча и плечелучевая мышца. Оба способствуют сгибанию локтевого сустава, но двуглавая мышца плеча также может генерировать супинационный крутящий момент, тогда как плечелучевая мышца может генерировать как супинационный, так и пронационный крутящий момент в зависимости от ориентации предплечья (Gielen and van Zuylen 1986; Buchanan et al.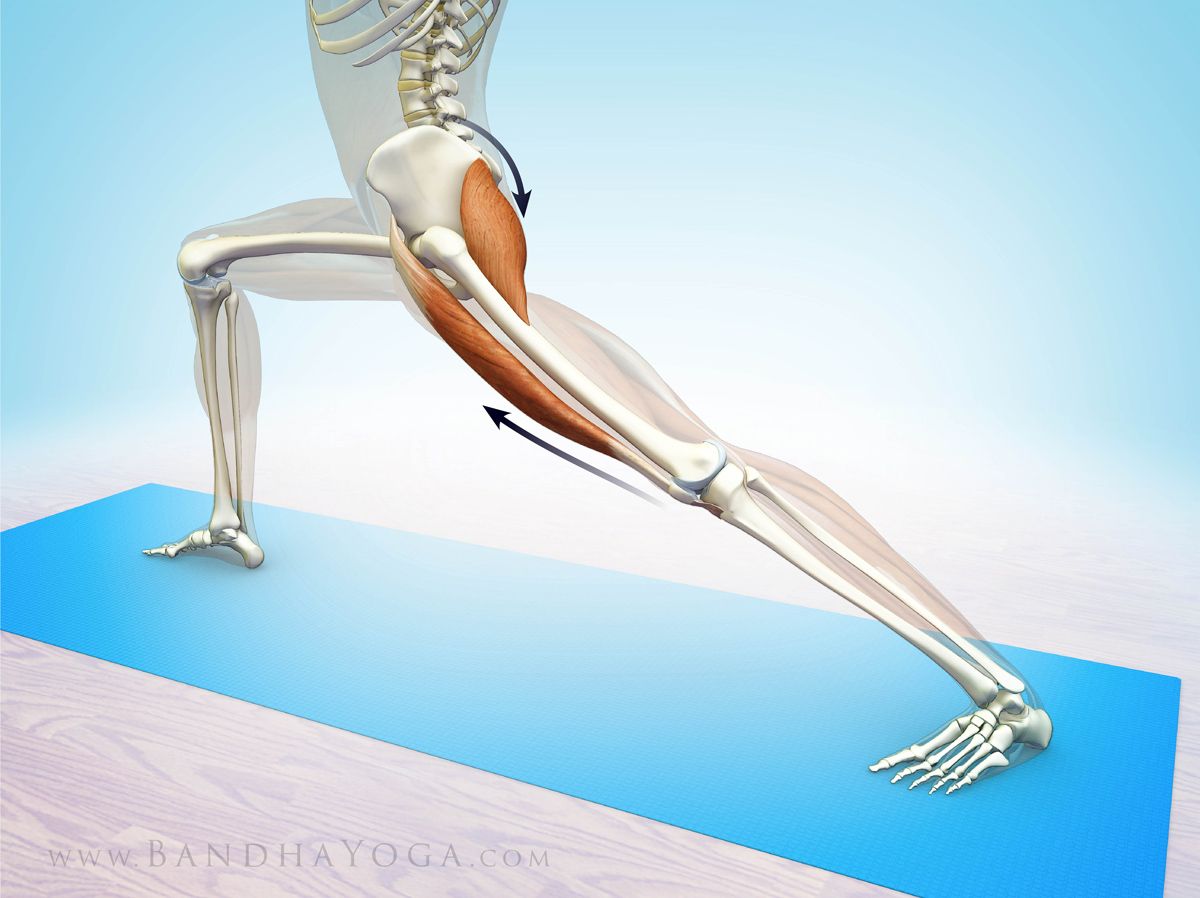 1989; Jamison and Caldwell 1993; Мюррей и др.1995 год; Чжан и др. 1998). Противоположные способности этих двух мышц к пронации и супинации изменяют количество активности, которую каждая из них генерирует во время упражнения на сгибание локтя, в соответствии с одновременными требованиями относительно длинной оси предплечья (Buchanan et al., 1989; Jamison and Caldwell, 1993). Например, ЭМГ-активность двуглавой мышцы плеча больше, когда задача включает одновременное сгибание и супинацию, по сравнению со сгибанием и пронацией (Barry and Carson 2004; Shemmell et al.2005).
1989; Jamison and Caldwell 1993; Мюррей и др.1995 год; Чжан и др. 1998). Противоположные способности этих двух мышц к пронации и супинации изменяют количество активности, которую каждая из них генерирует во время упражнения на сгибание локтя, в соответствии с одновременными требованиями относительно длинной оси предплечья (Buchanan et al., 1989; Jamison and Caldwell, 1993). Например, ЭМГ-активность двуглавой мышцы плеча больше, когда задача включает одновременное сгибание и супинацию, по сравнению со сгибанием и пронацией (Barry and Carson 2004; Shemmell et al.2005).
Также было показано, что относительная активация длинной и короткой головок двуглавой мышцы плеча зависит от вращающего момента пронации или супинации, сопровождающего сгибание локтя (Basmajian and Latif, 1957; Buchanan et al., 1986; Jamison and Caldwell, 1993). Эксперименты с записью отдельных двигательных единиц показали, что некоторые двигательные единицы в длинной головке двуглавой мышцы плеча и все двигательные единицы в короткой головке задействуются только тогда, когда задача требует как сгибательного, так и супинационного торка (ter Haar Romeny et al.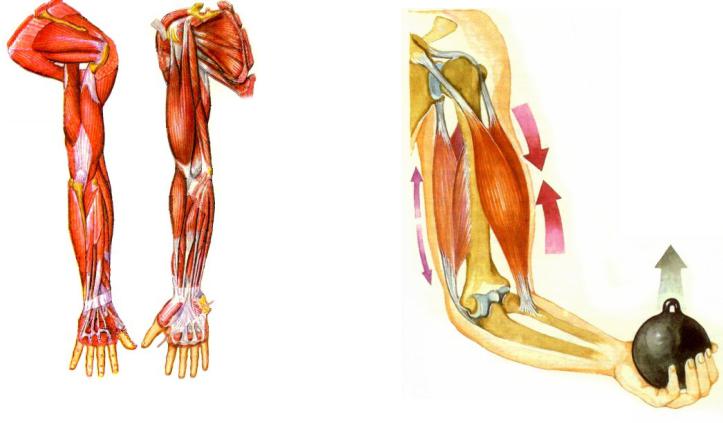 1982 год; тер Хаар Ромени и др. 1984 год; ван Зуйлен и др. 1988). Было высказано предположение, что спинномозговые рефлекторные пути (Windhorst et al. 1989), в частности пути торможения между мышцами-антагонистами (Jongen et al. 1989), могут лежать в основе дифференцированного рекрутирования двигательных единиц в популяции. Соответственно, неоднородная активация пула моторных нейронов двуглавой мышцы плеча может контролироваться таким механизмом (van Zuylen et al., 1988; Jongen et al., 1989). Также было высказано предположение, что двуглавая мышца плеча рефлекторно тормозится, когда предплечье находится в положении лежа (Basmajian and Latif, 1957), и что рекрутирование некоторых двигательных единиц двуглавой мышцы плеча может блокироваться тормозными путями, которые активны во время пронации (ter Haar Romeny et al.1984).
1982 год; тер Хаар Ромени и др. 1984 год; ван Зуйлен и др. 1988). Было высказано предположение, что спинномозговые рефлекторные пути (Windhorst et al. 1989), в частности пути торможения между мышцами-антагонистами (Jongen et al. 1989), могут лежать в основе дифференцированного рекрутирования двигательных единиц в популяции. Соответственно, неоднородная активация пула моторных нейронов двуглавой мышцы плеча может контролироваться таким механизмом (van Zuylen et al., 1988; Jongen et al., 1989). Также было высказано предположение, что двуглавая мышца плеча рефлекторно тормозится, когда предплечье находится в положении лежа (Basmajian and Latif, 1957), и что рекрутирование некоторых двигательных единиц двуглавой мышцы плеча может блокироваться тормозными путями, которые активны во время пронации (ter Haar Romeny et al.1984).
Найто и др. (1996) описал тормозной рефлекторный путь от афферентов плечелучевого нерва к двуглавой мышце плеча, который может способствовать профилю рекрутирования двигательных единиц в двуглавой мышце плеча. При случайной стимуляции лучевого нерва ниже двигательного порога они обнаружили, что разрядка 52% зарегистрированных одиночных двигательных единиц была задержана. 21 двигательная единица, изученная Naito et al. (1996) располагались в медиальной части двуглавой мышцы плеча, а предплечье располагалось в супинированном положении.Учитывая различные профили рекрутирования двигательных единиц в короткой и длинной головках двуглавой мышцы плеча и влияние положения предплечья на ЭМГ-активность двуглавой мышцы плеча, мы предположили, что сила тормозного рефлекса будет различаться между двумя головками и зависит от положения предплечья, поэтому что она максимальна, когда предплечье пронировано, а активация двуглавой мышцы плеча обычно снижена. Цель настоящего исследования состояла в том, чтобы охарактеризовать тормозной рефлекс путем сравнения его силы в короткой и длинной головках двуглавой мышцы плеча и изучения влияния положения предплечья на силу рефлекса.Некоторые из этих данных представлены в абстрактной форме (Pascoe et al. 2006; Riley et al. 2006).
2006; Riley et al. 2006).
Методы
Проведена серия экспериментов с 15 здоровыми взрослыми (13 мужчин, 2 женщины, 27,2 ± 4,8 года, диапазон 21–39 лет). Комитет по исследованиям человека Университета Колорадо в Боулдере одобрил процедуры, и эксперименты проводились в соответствии с Хельсинкской декларацией. Все испытуемые дали письменное информированное согласие до участия в исследовании.
Экспериментальная установка
Испытуемых усаживали вертикально на стул с левым плечом вертикально и слегка отведенным от туловища, а локоть опирался на опору. Локтевой сустав был согнут до 1,57 рад, предплечье горизонтально. Кисть и предплечье были зафиксированы с помощью модифицированного ортеза запястье-кисть (Orthomerica; Ньюпорт-Бич, Калифорния, США), а сила, оказываемая мышцами-сгибателями локтевого сустава, была измерена с помощью датчика силы-момента (JR-3, диапазон 900 Н, 89,4 N/V; JR-3, Вудленд, Калифорния, США).Датчик крепился к ортезу на уровне запястья. Субъектам было предложено напрячь мышцы-сгибатели локтя, чтобы подтянуть ортез вверх.
Записи отдельных двигательных единиц
Потенциалы отдельных двигательных единиц регистрировали с длинной и короткой головок двуглавой мышцы плеча с использованием проволоки из нержавеющей стали (диаметр 50 мкм, California Fine Wire, Гровер-Бич, Калифорния, США), изолированной формваром. и склеены на записывающих наконечниках. Изоляция отсутствовала только на записывающем наконечнике каждого провода, а в каждый электрод входило по два-три провода.Провода вводили в мышечное брюшко с помощью иглы для подкожных инъекций 27 или 30 калибра, которую удаляли после того, как провода были на месте. Регулировка глубины электрода и использование альтернативных биполярных конфигураций проводов улучшили качество сигнала двигательной единицы. Референтный электрод помещали на кожу над латеральным надмыщелком. Записи отдельных двигательных единиц усиливались (1000–5000 раз) и подвергались полосовой фильтрации в диапазоне от 0,3 до 8,5 кГц (Coulbourn Instruments, Аллентаун, Пенсильвания, США). Сигнал двигательной единицы регистрировали на частоте 20 кГц с помощью Power 1401 (CED, Кембридж, Великобритания), сохраняли на компьютере, а потенциалы отдельной двигательной единицы определяли в режиме онлайн и в автономном режиме с использованием программного обеспечения Spike2 (v.5.16, КЭД).
Сигнал двигательной единицы регистрировали на частоте 20 кГц с помощью Power 1401 (CED, Кембридж, Великобритания), сохраняли на компьютере, а потенциалы отдельной двигательной единицы определяли в режиме онлайн и в автономном режиме с использованием программного обеспечения Spike2 (v.5.16, КЭД).
Запись интерференционной ЭМГ
Интерференционную электромиограмму (ЭМГ) регистрировали с помощью биполярных поверхностных электродов (Ag–AgCl, диаметром 4 или 8 мм, межэлектродное расстояние 20 мм), расположенных над длинной и короткой головками двуглавой и трехглавой мышц плеча, brachioradialis и лучевой разгибатель запястья. Интерференционная ЭМГ была получена от плечевой мышцы с использованием внутримышечных биполярных электродов, изготовленных из проволоки из нержавеющей стали с формваровой изоляцией диаметром 75 мкм с удаленной изоляцией на 1 мм на записывающем наконечнике.Два отдельных провода были введены в плечевую мышцу на расстоянии примерно 20 мм друг от друга с помощью игл для подкожных инъекций 30 калибра и 2,54 см. Референтные электроды располагали на латеральном надмыщелке плечевой кости, на головке лучевой кости и на акромиальном отростке лопатки. Сигналы ЭМГ усиливали (в 100–10 000 раз; серия S, Coulbourn Instruments) и подвергали полосовой фильтрации от 13 Гц до 1 кГц. Интерференционные ЭМГ регистрировались на частоте 2 или 4 кГц и сохранялись, как описано для внутримышечных сигналов.
Референтные электроды располагали на латеральном надмыщелке плечевой кости, на головке лучевой кости и на акромиальном отростке лопатки. Сигналы ЭМГ усиливали (в 100–10 000 раз; серия S, Coulbourn Instruments) и подвергали полосовой фильтрации от 13 Гц до 1 кГц. Интерференционные ЭМГ регистрировались на частоте 2 или 4 кГц и сохранялись, как описано для внутримышечных сигналов.
Стимуляция нерва
Стимулятор Grass S88 (Grass Technologies, West Warwick, RI, США) использовали последовательно с блоком изоляции SIU5 и блоком постоянного тока CCU1 для подачи импульса 0.Прямоугольный импульс длительностью 5 мс на плечелучевую ветвь лучевого нерва при 0,9 × моторном пороге (МТ). Катодную стимуляцию проводили с латеральной стороны плеча, над плечевой костью и на 3–4 см выше латерального надмыщелка. После определения подходящего места стимуляции с помощью биполярного металлического электрода-зонда клейкие электроды (электроды для стимуляции периферических нервов NDM, 10-миллиметровый диск Ag-AgCl с гелевой контактной поверхностью 2,4 × 1,9 см, Conmed, Ютика, Нью-Йорк, США) были помещены в вызывают отчетливую М-волну плечелучевой мышцы при низкой интенсивности стимула без какой-либо активации лучевого разгибателя запястья при интенсивности стимула не менее 2 х МТ для брахиорадиальной мышцы (Naito et al. 1996). Хотя стимулирующие электроды меньшего размера позволяли бы проводить более очаговую стимуляцию, электроды большего размера обеспечивали более последовательную активацию афферентных волокон при исследовании различных положений предплечья и движении поверхностного электрода относительно нижележащего нерва.
1996). Хотя стимулирующие электроды меньшего размера позволяли бы проводить более очаговую стимуляцию, электроды большего размера обеспечивали более последовательную активацию афферентных волокон при исследовании различных положений предплечья и движении поверхностного электрода относительно нижележащего нерва.
Поскольку плечевая мышца может получать некоторую иннервацию от лучевого нерва в дополнение к своей первичной иннервации от мышечно-кожного нерва (Mahakkanukrauh and Somsarp 2002; Vicente et al.2005 г.; Блэкберн и др. 2007) был проведен отдельный эксперимент для оценки происхождения афферентов, ответственных за торможение от стимуляции лучевого нерва. Ветвь лучевого нерва к плечевой мышце расположена относительно близко к той, которая иннервирует плечелучевую мышцу, и рядом с электродами, которые использовались для вызова ответа. Чтобы оценить возможное участие афферентов, исходящих из плечевой мышцы, стимуляция была оптимизирована для получения М-волны либо в плечевой, либо в плечелучевой мышце, при этом гарантируя отсутствие активации лучевого разгибателя запястья при интенсивности до 2. 0 × МТ. Для одной и той же двигательной единицы двуглавой мышцы плеча были зарегистрированы ответы примерно на 100 стимулов на стимуляцию, сначала оптимизированную для одной из двух мышц, а затем оптимизированную для другой мышцы. Моторный порог как для брахиалиса, так и для брахиорадиалиса был определен для каждого места стимуляции, чтобы определить эффективную интенсивность стимуляции для нецелевой ветви нерва, когда целевая ветвь нерва была активирована при 0,9 × MT.
0 × МТ. Для одной и той же двигательной единицы двуглавой мышцы плеча были зарегистрированы ответы примерно на 100 стимулов на стимуляцию, сначала оптимизированную для одной из двух мышц, а затем оптимизированную для другой мышцы. Моторный порог как для брахиалиса, так и для брахиорадиалиса был определен для каждого места стимуляции, чтобы определить эффективную интенсивность стимуляции для нецелевой ветви нерва, когда целевая ветвь нерва была активирована при 0,9 × MT.
Стимуляция сухожилий
Короткие механические импульсы (3 импульса при 200 Гц, 15 мс) применяли к дистальному отделу сухожилия или брюшку плечелучевой мышцы с помощью механического вибратора (вибратор LDS V203 и усилитель мощности PA25E, Ling Dynamic Systems, Herts, UK) .На вибратор насаживали алюминиевый зонд диаметром 8 мм для взаимодействия с кожей. Силу реакции на кожу измеряли с помощью датчика силы MLP-10 (Transducer Techniques, Temecula, CA) и поддерживали на фоновом уровне 1–2 Н. Расположение и сила механических постукиваний, вызывающих брахиорадиальные Т-рефлексы, были определяется путем усреднения ответа ЭМГ на 24 механических импульса (~ 0,5 Гц), поскольку субъект поддерживал небольшое фоновое сокращение. Интенсивность во время стимуляции, запускаемой спайком, была установлена примерно на 0.в 5–0,8 раза выше порога моторной реакции, а стимул доставлялся с задержкой 25 мс после разряда каждой триггерной двигательной единицы. Тщательное позиционирование вибратора требовалось, чтобы избежать моносинаптического возбуждения 1а двуглавой мышцы плеча путем активации мышечных веретен в лучевом разгибателе запястья (Cavallari and Katz 1989). Для некоторых испытуемых было необходимо расположить вибратор над плечелучевой мышцей живота, а не над дистальным сухожилием.
Интенсивность во время стимуляции, запускаемой спайком, была установлена примерно на 0.в 5–0,8 раза выше порога моторной реакции, а стимул доставлялся с задержкой 25 мс после разряда каждой триггерной двигательной единицы. Тщательное позиционирование вибратора требовалось, чтобы избежать моносинаптического возбуждения 1а двуглавой мышцы плеча путем активации мышечных веретен в лучевом разгибателе запястья (Cavallari and Katz 1989). Для некоторых испытуемых было необходимо расположить вибратор над плечелучевой мышцей живота, а не над дистальным сухожилием.
Максимальные произвольные сокращения
Максимальные произвольные сокращения (МВС) выполняли мышцами-сгибателями локтевого сустава с предплечьем в нейтральном положении.Участникам было предложено увеличить крутящий момент во время изометрического сокращения до максимума примерно через 3 с, а затем приложить максимально возможный крутящий момент в течение дополнительных 3 с. После нескольких тренировочных попыток МПК выполняли 3 раза с последовательными попытками, разделенными 2–3-минутным отдыхом. Максимальный крутящий момент, достигнутый в трех попытках, принимали за крутящий момент MVC.
Максимальный крутящий момент, достигнутый в трех попытках, принимали за крутящий момент MVC.
Протокол
Силу рефлекса оценивали путем регистрации разряда отдельных двигательных единиц из двуглавой мышцы плеча с помощью электрода из тонкой проволоки, который вставляли либо в длинную, либо в короткую головку двуглавой мышцы плеча.Субъект производил низкие силы сгибания до тех пор, пока не была изолирована одна двигательная единица, которую можно было четко различить с помощью дискриминатора порогового окна (серия S, Coulbourn Instruments). Затем импульсная стимуляция (Stephens et al., 1976; Fournier et al., 1986) проводилась каждые 2–3 с, поскольку испытуемые поддерживали постоянную скорость разряда ~ 11 импульсов в секунду (pps). Каждый из 100 стимулов давался с установленной задержкой после выбранного разряда.
Помимо пороговой дискриминации, с помощью программного обеспечения Spike2 (CED) было проведено сопоставление онлайн-шаблона потенциала действия двигательной единицы, чтобы убедиться, что на протяжении всего эксперимента отслеживалась одна и та же отдельная двигательная единица. Субъекту была предоставлена звуковая обратная связь о разрядке двигательных единиц и визуальная обратная связь о силе сгибания локтя и скорости разрядки двигательных единиц, чтобы помочь в поддержании постоянной скорости разрядки. Для измерения стабильности фоновой скорости разряда контрольные импульсы перемежались между последовательными стимулами через случайный интервал не менее 1 с после триггера стимуляции и за 0,7 с до следующего триггера стимуляции.
Субъекту была предоставлена звуковая обратная связь о разрядке двигательных единиц и визуальная обратная связь о силе сгибания локтя и скорости разрядки двигательных единиц, чтобы помочь в поддержании постоянной скорости разрядки. Для измерения стабильности фоновой скорости разряда контрольные импульсы перемежались между последовательными стимулами через случайный интервал не менее 1 с после триггера стимуляции и за 0,7 с до следующего триггера стимуляции.
Найто и др. (1996) не сообщили о задержке стимуляции, используемой в их исследовании, поэтому в первом эксперименте проверялось влияние трех задержек стимуляции на одну и ту же единственную двигательную единицу.Результаты () свидетельствовали об отсутствии разницы в удлинении межспайкового интервала при задержках 30, 40 или 50 мс, поэтому во всех последующих экспериментах использовалась задержка 30 мс. Задержка в 30 мс минимизировала загрязнение последующей записи двигательной единицы артефактом стимула и максимизировала влияние стимуляции на распределение всего интервала между спайками.
a Схематическое представление времени торможения, вызванного при 3 различных задержках (30, 40 и 50 мс после триггерной разрядки двигательной единицы в момент времени 0 мс), и гистограмма нормально распределенных интервалов между спайками для разрядки двигательной единицы примерно в 11 секунд. стр. с.Эти смоделированные данные показывают типичную изменчивость интервалов между спайками для «регулярно» разряжающейся двигательной единицы со средним интервалом между спайками 90 мс (11,1 pps) и коэффициентом вариации для интервала между спайками 18% (Matthews 1996; Moritz et al. ., 2005; Barry et al., 2007), который был аналогичен коэффициенту вариации для межспайкового интервала для экспериментальных данных. Ожидаемое появление тормозного залпа через 22 мс после стимуляции указано правым краем горизонтальной линии рядом с каждой числовой меткой, а левым краем указано время, в которое была доставлена стимуляция. b Средние данные по разнице интервалов между спайками для 16 двигательных единиц, протестированных с 3 задержками стимула.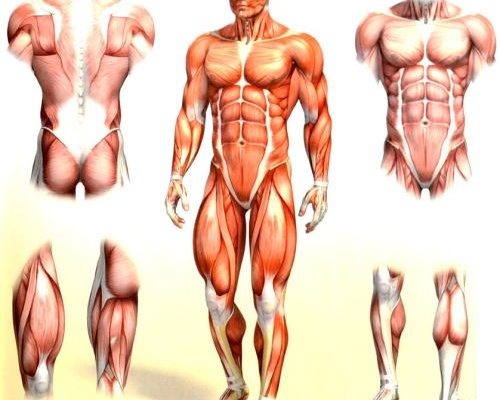 Этот показатель представляет собой разницу между интервалом между пиками, включающими стимул (ISI-0), и тремя интервалами между пиками, которые предшествовали стимуляции (ISI-pre), для стимула и контрольных условий. Положительные значения разности ISI означают ингибирование разряда двигательной единицы, которое было одинаковым для всех трех задержек ( P = 0,59). Несколько меньшая разница ISI для задержки в 50 мс, по-видимому, возникла из-за того, что самые ранние разряды двигательных единиц произошли бы до прихода тормозного залпа на двигательный нейрон.Разница ISI для контрольных условий была близка к 0 мс, что указывало на то, что целевая скорость фонового разряда поддерживалась постоянно на протяжении каждого испытания. Раздражители подавались на плечелучевую ветвь лучевого нерва при нейтральном положении предплечья
Этот показатель представляет собой разницу между интервалом между пиками, включающими стимул (ISI-0), и тремя интервалами между пиками, которые предшествовали стимуляции (ISI-pre), для стимула и контрольных условий. Положительные значения разности ISI означают ингибирование разряда двигательной единицы, которое было одинаковым для всех трех задержек ( P = 0,59). Несколько меньшая разница ISI для задержки в 50 мс, по-видимому, возникла из-за того, что самые ранние разряды двигательных единиц произошли бы до прихода тормозного залпа на двигательный нейрон.Разница ISI для контрольных условий была близка к 0 мс, что указывало на то, что целевая скорость фонового разряда поддерживалась постоянно на протяжении каждого испытания. Раздражители подавались на плечелучевую ветвь лучевого нерва при нейтральном положении предплечья
Исследовано также возможное влияние кожных афферентов на рефлекторное торможение. Электрические стимулы подавались либо на сторону плеча, прилегающую к месту стимуляции лучевого нерва, либо на кожу, покрывающую костные выступы локтевого сустава.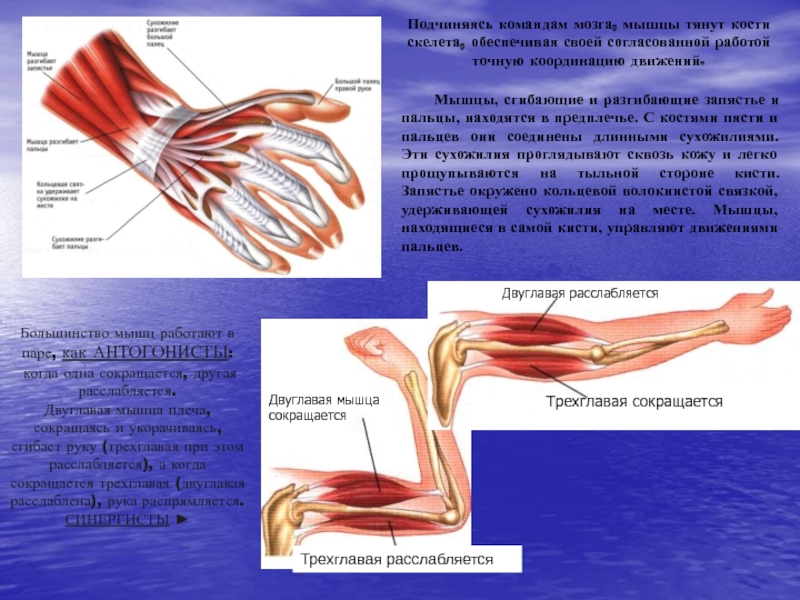 Эти места были выбраны потому, что испытуемые обычно испытывали только локальные ощущения при стимуляции лучевого нерва, а не парестезию, которая иррадиировала вниз по руке (Burke et al., 1992). Интенсивность кожных ощущений была примерно в 2 раза больше порога восприятия, чтобы напоминать либо подаваемый ток, либо воспринимаемую интенсивность, связанную со стимуляцией лучевого нерва при 0,9 × MT. Реакцию одной и той же двигательной единицы в двуглавой мышце плеча регистрировали на 100 стимулов, доставленных отдельно к лучевому нерву и участкам кожи, или на случайную комбинацию стимулов из двух мест.
Эти места были выбраны потому, что испытуемые обычно испытывали только локальные ощущения при стимуляции лучевого нерва, а не парестезию, которая иррадиировала вниз по руке (Burke et al., 1992). Интенсивность кожных ощущений была примерно в 2 раза больше порога восприятия, чтобы напоминать либо подаваемый ток, либо воспринимаемую интенсивность, связанную со стимуляцией лучевого нерва при 0,9 × MT. Реакцию одной и той же двигательной единицы в двуглавой мышце плеча регистрировали на 100 стимулов, доставленных отдельно к лучевому нерву и участкам кожи, или на случайную комбинацию стимулов из двух мест.
Для оценки электрического порога ответа оценивали силу рефлекторного торможения в одной и той же двигательной единице при восьми интенсивностях стимуляции (0,5, 0,6, 0,7, 0,75, 0,8, 0,9, 1,0 и 1,1 × МТ), приложенных к плечелучевая ветвь лучевого нерва. В качестве дополнительного теста на участие 1а-афферентов от плечелучевой мышцы сравнивали ответы одной и той же отдельной двигательной единицы на стимуляцию плечелучевой ветви лучевого нерва и механические постукивания по плечелучевой мышце.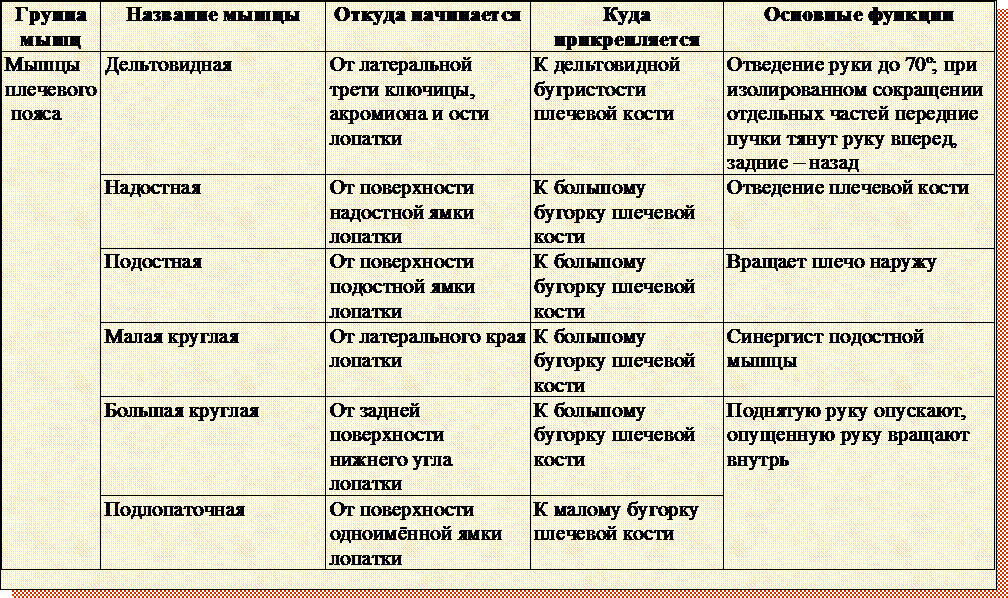 Оба эти эксперимента и эксперименты по стимуляции кожи проводились с предплечьем в нейтральном положении. Чтобы сравнить силу рефлекса между длинной и короткой головками, выборка двигательных единиц, зарегистрированных с обеих головок, была расширена, и стимуляция применялась к плечелучевой ветви лучевого нерва с предплечьем в нейтральном положении. В подмножестве экспериментов образцы двигательных единиц были отобраны с обеих голов в одном и том же экспериментальном сеансе, чтобы обеспечить согласованность стимуляции нерва.
Оба эти эксперимента и эксперименты по стимуляции кожи проводились с предплечьем в нейтральном положении. Чтобы сравнить силу рефлекса между длинной и короткой головками, выборка двигательных единиц, зарегистрированных с обеих головок, была расширена, и стимуляция применялась к плечелучевой ветви лучевого нерва с предплечьем в нейтральном положении. В подмножестве экспериментов образцы двигательных единиц были отобраны с обеих голов в одном и том же экспериментальном сеансе, чтобы обеспечить согласованность стимуляции нерва.
Чтобы изучить влияние положения предплечья на силу рефлекса, одна и та же отдельная двигательная единица в любой головке двуглавой мышцы плеча отслеживалась, когда предплечье удерживалось в трех разных положениях: пронация (~0,79 рад), супинация (~0,79 рад). рад) или нейтральный (~0 рад). Реакции примерно на 100 стимулов регистрировались для каждой позы, но испытания проводились только тогда, когда амплитуда, ширина и форма потенциала действия двигательной единицы были достаточно постоянными в разных положениях, чтобы указать, что запись была от одной и той же двигательной единицы. Рука минимально перемещалась между различными положениями, чтобы не нарушить запись отдельной двигательной единицы. В качестве показателя согласованности стимула измерялись М-волны брахиорадиальной мышцы до или после каждой последовательности в разных положениях предплечья.
Рука минимально перемещалась между различными положениями, чтобы не нарушить запись отдельной двигательной единицы. В качестве показателя согласованности стимула измерялись М-волны брахиорадиальной мышцы до или после каждой последовательности в разных положениях предплечья.
Анализ данных
Средняя сила, средняя скорость разряда и коэффициент вариации (CV) для интервала между спайками (ISI) рассчитывались в зависимости от времени подачи стимулов. Сопоставление шаблона с программным обеспечением Spike2 повторялось в автономном режиме для различения потенциалов действия отдельных двигательных единиц.Точность этого различения была подтверждена визуальным осмотром каждого выделенного потенциала действия и просмотром распределения интервалов между спайками. Офлайн-анализ также подтвердил, что в период между стимуляцией или контрольным событием и предшествующим триггерным разрядом двигательной единицы не происходило разрядов двигательных единиц. Гистограммы времени после стимула (PSTH) были построены с интервалами 0,5 мс для периода 100 мс после доставки стимула и для контрольных условий, когда стимул не был предоставлен.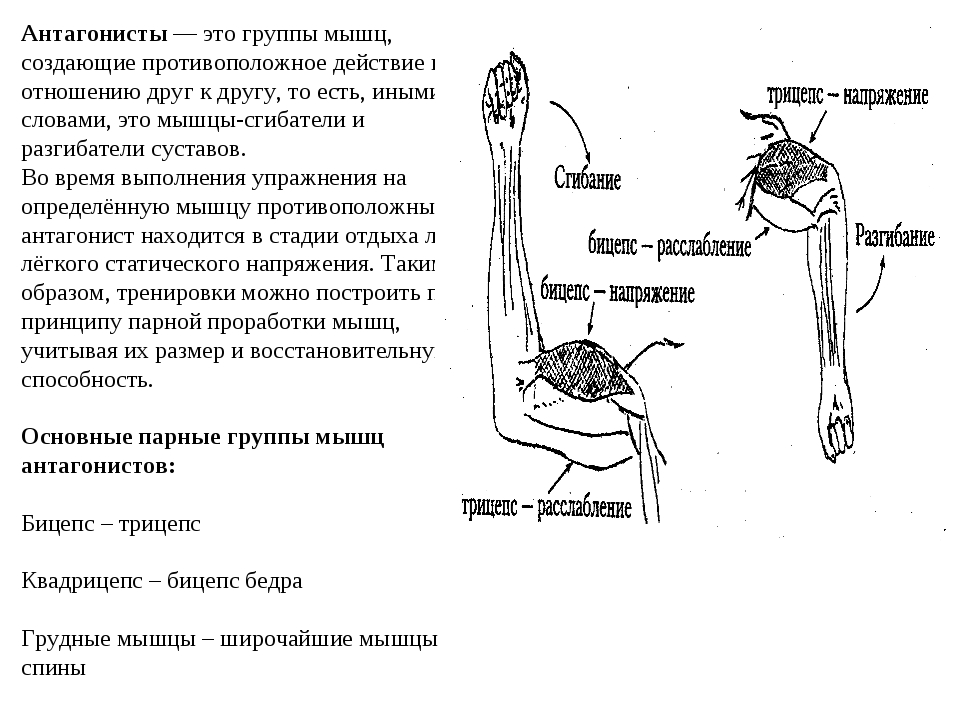 Совокупные суммы рассчитывали по гистограммам стимуляции и контроля (Ellaway, 1978).
Совокупные суммы рассчитывали по гистограммам стимуляции и контроля (Ellaway, 1978).
Степень торможения в образце двигательной единицы была количественно определена путем измерения удлинения интервала между спайками, вызванного стимуляцией лучевого нерва (Datta and Stephens 1981; Nafati et al. 2005). Три значения ISI до стимуляции усреднялись (ISI-до) и измерялось значение ISI, которое включало стимуляцию (ISI-0). Разницу между двумя значениями (мс) использовали для обозначения силы торможения рефлекса.Положительное значение любого индекса соответствовало торможению, тогда как отрицательное значение указывало на облегчение. Эти интервалы были извлечены для каждого триггера стимуляции и контроля, и были рассчитаны средние значения для каждой двигательной единицы.
Селективность стимуляции лучевого нерва измеряли путем усреднения ЭМГ от плечелучевой мышцы и лучевого разгибателя запястья после стимуляции на двигательном пороге или выше для обеих мышц. Пиковую амплитуду плечелучевой М-волны в ответ на стимуляцию в 1-2 х MT регистрировали до и в конце каждой экспериментальной сессии или во время каждого изменения положения предплечья, чтобы убедиться, что условия стимуляции не изменились.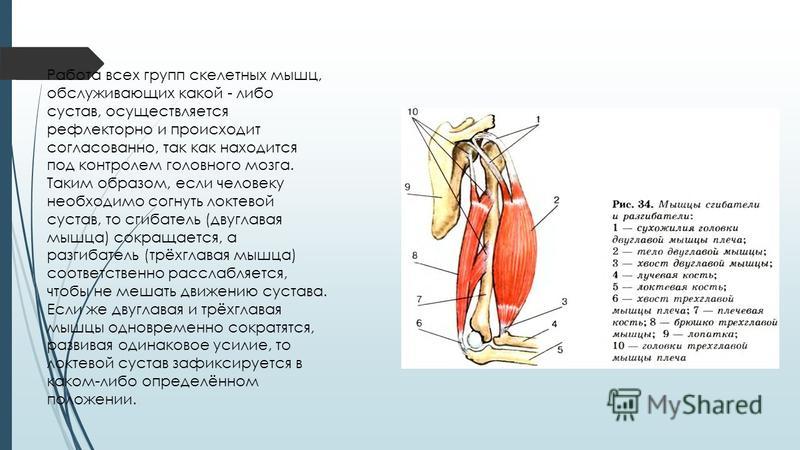 Форма волны также усреднялась во время испытаний двигательных единиц, чтобы гарантировать отсутствие двигательной реакции на стимуляцию во время сокращений с малой силой.
Форма волны также усреднялась во время испытаний двигательных единиц, чтобы гарантировать отсутствие двигательной реакции на стимуляцию во время сокращений с малой силой.
Статистический анализ
Характеристики скорости разряда, сила и ЭМГ для каждой мышцы сравнивались с дисперсионным анализом повторных измерений для каждого испытания стимуляции. Пиковые амплитуды М-волн сравнивали между положениями предплечий с помощью дисперсионного анализа с повторными измерениями. Статистика хи-квадрат использовалась для изучения влияния стимуляции на ПСТГ для триггеров стимуляции и контроля.ПСТГ сравнивали в течение 25–95 мс после стимуляции. Наблюдаемые и ожидаемые подсчеты для расчета статистики хи-квадрат были взяты из групп из 10 бинов PSTH, чтобы обеспечить среднее количество бинов ≥5, чтобы удовлетворить предположения теста хи-квадрат. ANOVA с повторными измерениями использовали для сопоставления средних интервалов между спайками, зарегистрированных до (ISI-pre) и во время (ISI-0) разрядов двигательных единиц, выбранных для генерации триггеров (стимуляция и контроль), и для оценки разницы между двумя головками.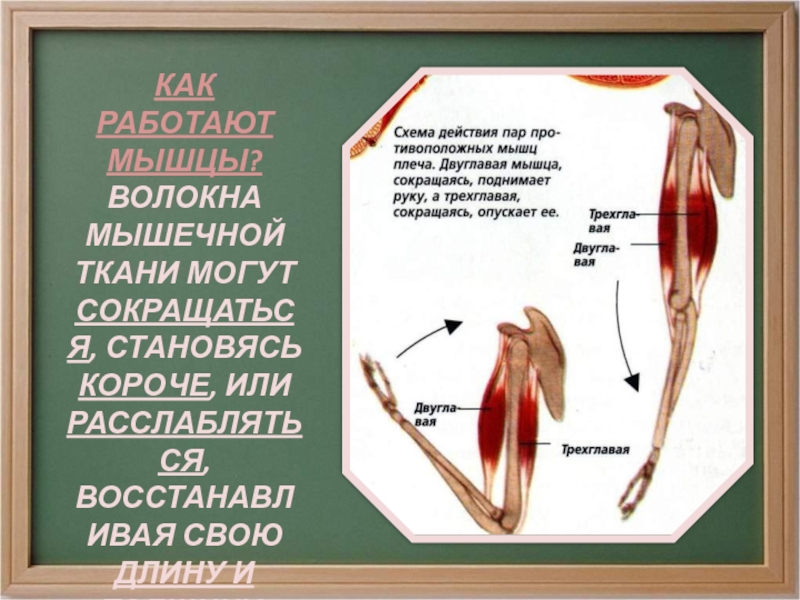 двуглавой мышцы плеча и различные положения предплечий.Все двигательные единицы были включены в анализ средних интервалов между спайками. Альфа-уровень P <0,05 использовали для выявления существенных различий. Все статистические анализы были выполнены в SPSS (Чикаго, Иллинойс). Данные представлены в тексте и таблицах как среднее ± стандартное отклонение (SD).
двуглавой мышцы плеча и различные положения предплечий.Все двигательные единицы были включены в анализ средних интервалов между спайками. Альфа-уровень P <0,05 использовали для выявления существенных различий. Все статистические анализы были выполнены в SPSS (Чикаго, Иллинойс). Данные представлены в тексте и таблицах как среднее ± стандартное отклонение (SD).
Результаты
Удлинение интервалов между спайками было одинаковым ( P = 0,59; ) для всех трех задержек стимула (30, 40 и 50 мс) в 16 двигательных единицах; по 8 от короткой и длинной головок двуглавой мышцы плеча (3 предмета).Значительное ингибирование наблюдалось у 14 из 16 ПСТГ при задержке 30 мс и у 13 из 16 ПСТГ при задержке 40 и 50 мс. Пиковые отрицательные значения для кумулятивных сумм составили -28,3 ± 19,3 импульса/стимул для задержки 30 мс, -29,8 ± 17,6 импульса/стимул для задержки 40 мс и -22,0 ± 8,5 импульса/стимул для 50-мс задержки. задержка ( P = 0,13). Латентность торможения по отношению к разряду триггерной двигательной единицы составила 64,2 ± 8,8 мс для задержки 30 мс (34,2 мс после стимуляции) [69].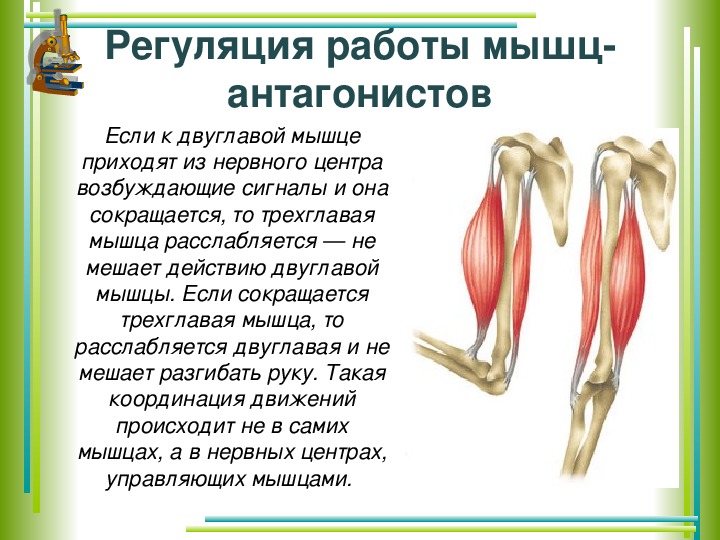 0 ± 6,1 мс для задержки 40 мс (29,0 мс после стимуляции) и 72,1 ± 8,1 мс для задержки 50 мс (22,1 мс после стимуляции) ( P <0,05). Продолжительность впадины в кумулятивной сумме не различалась ( P = 0,96) для 3 задержек стимуляции (30 мс: 19,3 ± 13,0 мс; 40 мс: 18,0 ± 11,7 мс; 50 мс: 18,3 ± 20,9 мс).
0 ± 6,1 мс для задержки 40 мс (29,0 мс после стимуляции) и 72,1 ± 8,1 мс для задержки 50 мс (22,1 мс после стимуляции) ( P <0,05). Продолжительность впадины в кумулятивной сумме не различалась ( P = 0,96) для 3 задержек стимуляции (30 мс: 19,3 ± 13,0 мс; 40 мс: 18,0 ± 11,7 мс; 50 мс: 18,3 ± 20,9 мс).
Динамика входного рефлекса во времени была дополнительно исследована у 66 двигательных единиц у 13 испытуемых путем оценки влияния стимуляции лучевого нерва (задержка 30 мс) на несколько интервалов между спайками после запуска выбранной двигательной единицы ().Наблюдался значительный эффект стимуляции ( P <0,001), который варьировался в интервалах между спайками ( P <0,01). Независимые тесты 90 555 t 90 556 показали значительные различия при каждом ISI после стимуляции (90 555 P <0,05), при этом наиболее стойкий эффект наблюдался во время ISI-0 и ISI + 1. После начального удлинения интервала между спайками (ISI-0) продолжительность следующего межспайкового интервала (ИСИ + 1) была короче, но затем снова увеличивалась для второго межспайкового интервала после стимуляции (ИСИ + 2). К четвертому межспайковому интервалу после стимуляции (ISI + 4) продолжительность межспайкового интервала была близка к контрольным уровням.
К четвертому межспайковому интервалу после стимуляции (ISI + 4) продолжительность межспайкового интервала была близка к контрольным уровням.
Влияние стимуляции плечелучевой ветви лучевого нерва на последовательные интервалы между спайками. Стимуляцию проводили во время ISI-0. Данные показаны для 66 двигательных единиц, зарегистрированных с предплечьем в нейтральном положении. Столбики ошибок указывают 95% доверительные интервалы
Ответы были получены для девяти двигательных единиц (шесть субъектов) при стимуляции, оптимизированной для плечелучевой ветви лучевого нерва в одной последовательности и для плечевой ветви в другой последовательности () .Две нервные ветви можно было стимулировать только с ограниченной избирательностью. Стимуляция плечелучевой ветви при 0,9×МТ соответствовала интенсивности 0,78±0,15×МТ для плечевой ветви лучевого нерва. Точно так же стимуляция ветви брахиалиса при 0,9 × MT была связана с интенсивностью 0,69 ± 0,13 × MT для ветви brachioradialis. Стимуляция в каждом месте вызывала значительное торможение разряда двигательных единиц ( P <0,001), и хотя процентное изменение ISI было больше при стимуляции ветви плечелучевого нерва (6.1%), чем для ветви брахиалиса (2,2%), разница не была значимой для двух местоположений ( P = 0,197).
Стимуляция в каждом месте вызывала значительное торможение разряда двигательных единиц ( P <0,001), и хотя процентное изменение ISI было больше при стимуляции ветви плечелучевого нерва (6.1%), чем для ветви брахиалиса (2,2%), разница не была значимой для двух местоположений ( P = 0,197).
Таблица 1
Таблица 1
Относительный стимул интенсивностей для брахиорадиалиса и брахилиса, когда стимуляция была оптимизирована для других мышц
| Mother Unit | Brachioradialis 0,9 MT | Brachialis 0,9 MT | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Доля плечелучевой МТ | ||||||||||||||||||||
| 1 | 0. 95 95 | 0,65 | ||||||||||||||||||
| 2 | 0,54 | 0,88 | ||||||||||||||||||
| 3 | 0,90 | 0,72 | ||||||||||||||||||
| 4 | 0,78 | 0,74 | ||||||||||||||||||
| 5 | 0,57 | 0,82 | ||||||||||||||||||
| 6 | 0.86 | 0.56 | 0.56 | |||||||||||||||||
| 7 | 7 | 0. 86 86 | 0.64 | |||||||||||||||||
| 8 | 0.72 | 0.46 | ||||||||||||||||||
| 9 | 0.87 | 0.73 | 0,73 | |||||||||||||||||
| Среднее (SD) | 0,78 (0,15) | 9078 (0,15) | 0,69 (0,13) |
Дополнительные измерения подтвердили специфичность стимуляции, используемую для вызова рефлекторного ингибирования разряда моторного блока Biceps. Влияние стимуляции плечелучевой ветви лучевого нерва сравнивали с электростимуляцией кожи на 14 двигательных единиц (семь испытуемых).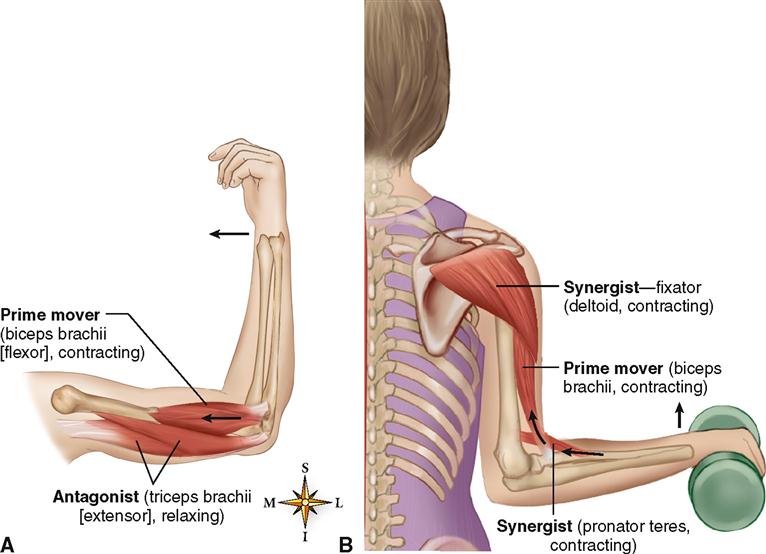 Значительное торможение наблюдалось только после стимуляции плечелучевой ветви лучевого нерва ( P < 0.05). Интервал между спайками удлинялся на 3,68 ± 3,31 мс при стимуляции лучевого нерва по сравнению с 0,52 ± 2,01 мс при кожной стимуляции (12). Сила рефлекторного торможения в одной и той же двигательной единице ( n = 4) достоверно увеличивалась ( P < 0,05) при увеличении интенсивности раздражения от 0,5 до 1,1×МТ (). Интервал между спайками значительно удлинялся при механической активации афферентов плечелучевой мышцы ( P < 0,05) и при электрической стимуляции плечелучевой ветви лучевого нерва ( P < 0.05).05) для шести одиночных двигательных единиц от шести испытуемых ().
Значительное торможение наблюдалось только после стимуляции плечелучевой ветви лучевого нерва ( P < 0.05). Интервал между спайками удлинялся на 3,68 ± 3,31 мс при стимуляции лучевого нерва по сравнению с 0,52 ± 2,01 мс при кожной стимуляции (12). Сила рефлекторного торможения в одной и той же двигательной единице ( n = 4) достоверно увеличивалась ( P < 0,05) при увеличении интенсивности раздражения от 0,5 до 1,1×МТ (). Интервал между спайками значительно удлинялся при механической активации афферентов плечелучевой мышцы ( P < 0,05) и при электрической стимуляции плечелучевой ветви лучевого нерва ( P < 0.05).05) для шести одиночных двигательных единиц от шести испытуемых ().
Данные для 14 одиночных двигательных единиц, показывающие разницу ISI, вызванную стимуляцией плечелучевой ветви лучевого нерва ( b ) и комбинированным эффектом двух разных мест кожной стимуляции ( a ). Эти данные были получены с предплечьем в нейтральном положении.
Средняя реакция 4 двигательных единиц двуглавой мышцы плеча на повышение интенсивности стимуляции плечелучевой ветви лучевого нерва.Было произведено 87 ± 12 стимулов при каждой из 8 интенсивностей стимула с предплечьем в нейтральном положении. Не было получено ответов от одной двигательной единицы на стимулы в 0,75 и 1,1 × MT. одинаковая двигательная единица. a Образцы PSTH и кумулятивные суммы от одного субъекта обнаруживают ранее описанный возбуждающий эффект стимуляции мышечных веретен в лучевом разгибателе запястья, который контрастирует с тормозящим эффектом, вызванным механическими постукиваниями по плечелучевой мышце. b Средние данные для 6 одиночных двигательных единиц в ответ на случайно смешанную электрическую стимуляцию плечелучевой ветви лучевого нерва и механические постукивания по сухожилию или брюшку плечелучевой мышцы. Эти данные были получены с предплечьем в нейтральном положении
Силу торможения двигательных единиц длинной и короткой головок двуглавой мышцы плеча сравнивали со спайк-триггерной стимуляцией для 57 двигательных единиц (31 длинная головка, 26 короткая головка). из 11 предметов.В среднем ответы на 99 ± 22 стимула регистрировались в течение 271 ± 64 с. Субъекты прилагали среднюю силу 3,6 ± 2,2% силы MVC, чтобы поддерживать постоянную скорость разряда двигательных единиц 11,2 ± 0,8 импульсов в секунду (11,2 ± 0,8 импульсов в секунду). Не было различий в продолжительности испытаний, количестве стимулов или характеристиках разряда ( P > 0,3) между длинной и короткой головками, за исключением немного более высокой средней силы для короткой головки ( P <0,05). Анализ хи-квадрат ПСТГ выявил значительное торможение для 40 из 57 гистограмм без различий между длинной (22/31, 71%) и короткой (18/26, 69%) головками двуглавой мышцы плеча.Также не было статистически значимой разницы ( P = 0,20) пикового отрицательного значения кумулятивной суммы для двигательных единиц с длинной (-18,4 ± 13,2 имп./стимул) и короткой головкой (-24,4 ± 19,7 имп./стимул). двуглавой мышцы плеча. Таблица 2 31)
из 11 предметов.В среднем ответы на 99 ± 22 стимула регистрировались в течение 271 ± 64 с. Субъекты прилагали среднюю силу 3,6 ± 2,2% силы MVC, чтобы поддерживать постоянную скорость разряда двигательных единиц 11,2 ± 0,8 импульсов в секунду (11,2 ± 0,8 импульсов в секунду). Не было различий в продолжительности испытаний, количестве стимулов или характеристиках разряда ( P > 0,3) между длинной и короткой головками, за исключением немного более высокой средней силы для короткой головки ( P <0,05). Анализ хи-квадрат ПСТГ выявил значительное торможение для 40 из 57 гистограмм без различий между длинной (22/31, 71%) и короткой (18/26, 69%) головками двуглавой мышцы плеча.Также не было статистически значимой разницы ( P = 0,20) пикового отрицательного значения кумулятивной суммы для двигательных единиц с длинной (-18,4 ± 13,2 имп./стимул) и короткой головкой (-24,4 ± 19,7 имп./стимул). двуглавой мышцы плеча. Таблица 2 31)
 0 ± 2.0
0 ± 2.0 Значительный интервал между спайками × триггерный эффект ( P <0,001) указывал на то, что интервалы между спайками удлинялись за счет стимуляции плечелучевой ветви лучевого нерва. ISI во время стимуляции (ISI-0) был значительно длиннее, чем среднее значение трех предыдущих ISI (ISI-pre) как для длинного (94,3 ± 6,9 и 89,5 ± 5,4 мс соответственно) и короткие головки (98,9 ± 14,5 и 90,8 ± 5,7 мс соответственно). Не было существенной разницы ( P = 0,11) в продолжительности ISI-0 между 31 двигательной единицей длинной головки и 26 двигательными единицами короткой головки двуглавой мышцы плеча. В 13 случаях были отобраны двигательные единицы как из длинной ( n = 13), так и из короткой головки ( n = 13) двуглавой мышцы плеча в ходе одного и того же сеанса эксперимента, что позволило провести сравнение с обычным местом стимуляции.Эти данные также продемонстрировали значительный эффект стимуляции ( P < 0,05) без существенной разницы между длинной и короткой головками двуглавой мышцы плеча (разница ISI: 4,9 ± 5,2 и 9,8 ± 15,9 мс соответственно; P = 0,22). ). Существовала значительная вариабельность влияния стимуляции на разряд отдельных двигательных единиц как между субъектами, так и внутри них, особенно в отношении короткой головки двуглавой мышцы плеча (10).
Разница ISI для стимула и условий контроля для каждой из 31 двигательной единицы длинной головки ( a ) и 26 двигательных единиц короткой головки ( b ) двуглавой мышцы плеча.Разница между исходными ISI и ISI, которая включала стимуляцию, была значимой для обеих головок ( P < 0,001) без различий между головками ( P = 0,11). Данные получали, когда предплечье находилось в нейтральном положении, а стимулы подавались на плечелучевую ветвь лучевого нерва. Две двигательные единицы короткой головки двуглавой мышцы плеча ( b ), которые демонстрировали наибольшее торможение, были зарегистрированы у одного и того же субъекта, а двигательные единицы, зарегистрированные в длинной головке этого субъекта, также демонстрировали одно из самых сильных торможений в выборке
Данные получали, когда предплечье находилось в нейтральном положении, а стимулы подавались на плечелучевую ветвь лучевого нерва. Две двигательные единицы короткой головки двуглавой мышцы плеча ( b ), которые демонстрировали наибольшее торможение, были зарегистрированы у одного и того же субъекта, а двигательные единицы, зарегистрированные в длинной головке этого субъекта, также демонстрировали одно из самых сильных торможений в выборке
Для Во второй части задачи ответ на стимуляцию плечелучевой ветви лучевого нерва регистрировали для 18 одиночных двигательных единиц (9 длинных головок, 9 коротких головок) в 3 различных положениях предплечий у 8 испытуемых.Субъекты прилагали небольшие усилия (4,0 ± 1,8% силы MVC) к мышцам-сгибателям локтя и поддерживали среднюю скорость разряда 11,4 ± 0,8 импульсов в секунду для активной двигательной единицы (1). Не было никаких существенных различий в продолжительности испытания, количестве стимулов, силе сгибания локтя или характеристиках разряда ( P > 0,14) для трех положений предплечья.
Таблица 3
Сила и разряда Характеристики для 18 моторных единиц, записанные в каждом из трех позиций предплечья
| Пронал | Пронал | Supined | Supined | |
|---|---|---|---|---|
| Пробный продолжительностью (ы) | 234 ± 58 | 275 ± 87 | 233 ± 62 | |
| Количество стимулов | 90 ± 24 | 94 ± 23 | 94 ± 23 | 88 ± 26 | 88 ± 26
| Средняя сила (% MVC) | 4. 1 ± 1,5 1 ± 1,5 | 3,7 | 3.7 ± 1,9 | 4.3 ± 1,9 |
| Средний интервал Interspike (MS) | 88,3 ± 5.2 | 89,1 ± 5,8 | 86,7-7-61 | |
| Коэффициент вариации для ISI (%) | 15,5 ± 5,2 | 17,1 ± 6,3 | 14,9 ± 4,4 |
Хи-квадрат статистики PSTH показал значительное ингибирование в 12/18 гистограммах, когда предплечье было положении и 11/18 при супинации; пиковые отрицательные значения для кумулятивных сумм были -21.9 ± 18,6 импульсов/стимул в пронации, -23,1 ± 20,0 импульсов/стимул в нейтральном положении и -17,4 ± 13,7 импульсов/стимул в супинации. Примеры PSTH, кумулятивных сумм и связанных с ними потенциалов действия двигательных единиц показаны для трех положений предплечья. Удлинение интервалов между спайками после стимуляции лучевого нерва было выражено во всех трех положениях руки, с наибольшим тормозным эффектом в пронации (7,9 ± 8,2 мс), промежуточным в нейтральном (5,8 ± 7,1 мс) и наименьшим в супинации (2.8 ± 3,4 мс; Р < 0,05; ). Между отдельными двигательными единицами наблюдалась значительная вариабельность, и эта вариабельность была меньше в положении супинации, чем в нейтральном и пронированном положениях (2). Не было различий в реакции на стимуляцию в трех положениях между двигательными единицами короткой ( n = 9) и длинной ( n = 9) головок двуглавой мышцы плеча ( n = 0,75).
Примеры PSTH, кумулятивных сумм и связанных с ними потенциалов действия двигательных единиц показаны для трех положений предплечья. Удлинение интервалов между спайками после стимуляции лучевого нерва было выражено во всех трех положениях руки, с наибольшим тормозным эффектом в пронации (7,9 ± 8,2 мс), промежуточным в нейтральном (5,8 ± 7,1 мс) и наименьшим в супинации (2.8 ± 3,4 мс; Р < 0,05; ). Между отдельными двигательными единицами наблюдалась значительная вариабельность, и эта вариабельность была меньше в положении супинации, чем в нейтральном и пронированном положениях (2). Не было различий в реакции на стимуляцию в трех положениях между двигательными единицами короткой ( n = 9) и длинной ( n = 9) головок двуглавой мышцы плеча ( n = 0,75).
Гистограммы времени после стимула и наложенные потенциалы действия двигательных единиц для одной и той же отдельной двигательной единицы, записанные с предплечьем в пронированном, нейтральном и супинированном положениях.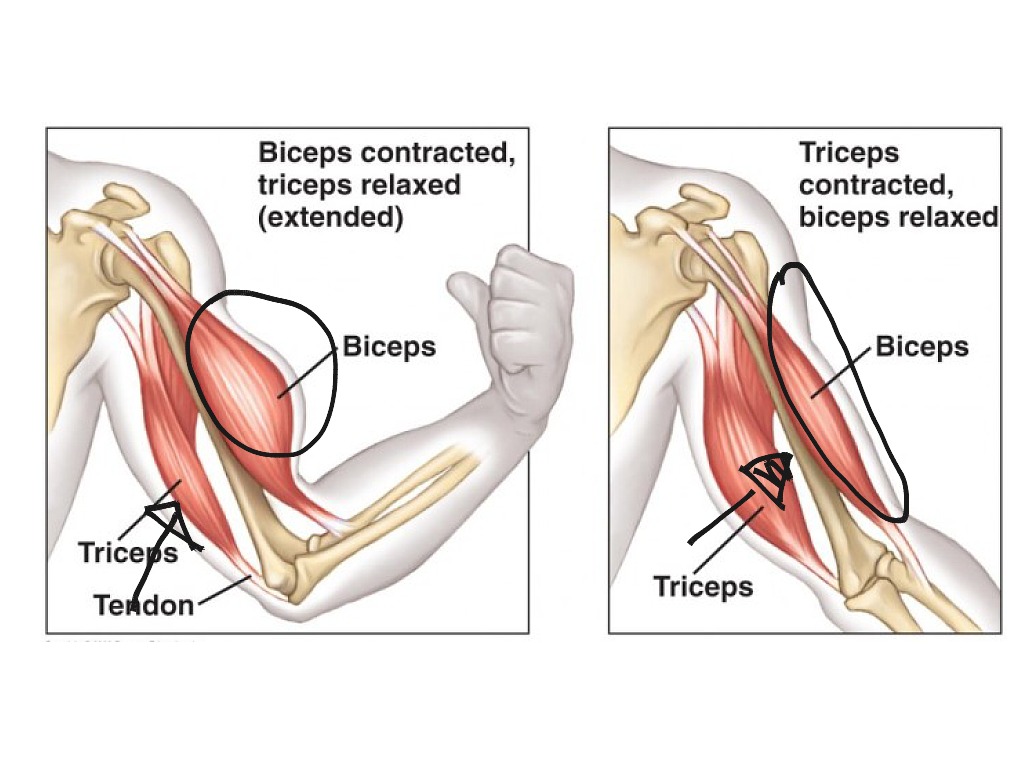 Гистограммы постстимульного времени строили для условий стимуляции и контроля, а к разнице между гистограммами стимуляции и контроля применяли процедуру кумулятивной суммы; отрицательное отклонение означает ингибирующий эффект стимуляции, которая продлевает время до следующей разрядки потенциала действия двигательным нейроном. Стимуляция проводилась в момент времени 0 мс, что соответствовало задержке в 30 мс после разряда предшествующей двигательной единицы ISI-pre) для условий стимуляции и контроля, усредненных для всех двигательных единиц ( a ) и для каждой двигательной единицы ( b ), когда предплечье находилось в пронированном, нейтральном и супинированном положениях.Сила торможения значительно модулируется положением предплечья ( P <0,05). В качестве показателя постоянства стимула показаны размахи амплитуд ответов М-волны ( c ), выраженные как доля контрольной М-волны, для каждой двигательной единицы, когда предплечье находилось в пронированном положении.
Гистограммы постстимульного времени строили для условий стимуляции и контроля, а к разнице между гистограммами стимуляции и контроля применяли процедуру кумулятивной суммы; отрицательное отклонение означает ингибирующий эффект стимуляции, которая продлевает время до следующей разрядки потенциала действия двигательным нейроном. Стимуляция проводилась в момент времени 0 мс, что соответствовало задержке в 30 мс после разряда предшествующей двигательной единицы ISI-pre) для условий стимуляции и контроля, усредненных для всех двигательных единиц ( a ) и для каждой двигательной единицы ( b ), когда предплечье находилось в пронированном, нейтральном и супинированном положениях.Сила торможения значительно модулируется положением предплечья ( P <0,05). В качестве показателя постоянства стимула показаны размахи амплитуд ответов М-волны ( c ), выраженные как доля контрольной М-волны, для каждой двигательной единицы, когда предплечье находилось в пронированном положении. , нейтральное и супинированное положения ( P = 0,9)
, нейтральное и супинированное положения ( P = 0,9)
В качестве показателя постоянства стимула амплитуда плечелучевой М-волны не изменилась () в трех положениях (выражается как доля контрольного М-зубца: пронация 0.94 ± 0,15; нейтральный 0,95 ± 0,14; супинация 1,05 ± 0,19; P = 0,9). Кроме того, фоновые уровни мышечной активности количественно определялись для каждого из трех положений руки путем усреднения среднеквадратичного значения амплитуды ЭМГ за несколько 0,5-секундных интервалов, начиная за 1 с до каждой стимуляции или контрольного события. Для нормализованных данных ЭМГ имелась значительная задача × мышечное взаимодействие ( P <0,05). Аналогичные уровни нормализованной ЭМГ были зарегистрированы для плечелучевой мышцы (1.41 ± 0,94, 1,42 ± 0,86, 1,40 ± 0,90% для пронации, нейтральной и супинации соответственно), плечевой (8,44 ± 11,20, 7,93 ± 10,38, 7,97 ± 10,41%) и трехглавой мышцы плеча (3,49 ± 2,89, 3,41 ± 2,86). , 3,45 ± 3,00%) в трех положениях рук, хотя активность брахиалиса была выше, чем у всех других мышц в трех задачах. Амплитуда ЭМГ была ниже, когда предплечье находилось в пронированном положении для длинной головки (3,08 ± 2,10, 5,29 ± 5,85, 5,05 ± 3,79% MVC для трех положений соответственно) и короткой головки (2.01 ± 1,53, 2,55 ± 1,90%, 2,54 ± 2,07% МОК) двуглавой мышцы плеча.
Амплитуда ЭМГ была ниже, когда предплечье находилось в пронированном положении для длинной головки (3,08 ± 2,10, 5,29 ± 5,85, 5,05 ± 3,79% MVC для трех положений соответственно) и короткой головки (2.01 ± 1,53, 2,55 ± 1,90%, 2,54 ± 2,07% МОК) двуглавой мышцы плеча.
Обсуждение
Основными результатами этого исследования были наличие рефлекторного торможения в обеих головках двуглавой мышцы плеча без различий в уровне торможения разряда двигательных единиц между двумя головками и модуляция силы рефлекторного торможения с помощью изменение положения предплечья. Эти результаты подтверждают первоначальные выводы Naito et al. (1996), что спинномозговой рефлекторный путь от плечелучевой мышцы, мышцы-синергиста для сгибания локтя, может замедлить текущую разрядку произвольно активированной двигательной единицы в двуглавой мышце плеча.Результаты по влиянию положения предплечья согласуются с гипотезой, но сходство силы рефлекса в двух головках двуглавой мышцы плеча контрастирует с ожидаемыми результатами.
Идентификация источника ингибирования
Ингибирование было определено количественно путем изучения влияния стимуляции на среднюю продолжительность интервала между спайками (Datta and Stephens 1981; Nafati et al. 2005). Гистограммы времени после стимула также были рассчитаны для сравнения с предыдущими данными (Naito et al.(1996). Анализ среднего интервала между спайками показал прогрессивное усиление торможения по мере того, как положение предплечья менялось от супинации к пронации. Надежность анализа среднего интервала между спайками была подтверждена анализом контрольного интервала между спайками путем также оценки средних интервалов между спайками до и после контрольных триггеров, чередующихся со стимулирующими триггерами по всей серии спайков.
Латентность ингибирования в текущем исследовании была аналогична той, о которой сообщили Naito et al.(1996). При стимуляции плечелучевой ветви лучевого нерва с задержкой 50 мс после разряда триггерной двигательной единицы снижение кумулятивной суммы происходило через 22 мс после стимуляции. Кажущаяся латентность торможения была больше, когда стимуляция проводилась через 30 и 40 мс, но разрешение для точной идентификации начала торможения снижается, когда происходило относительно небольшое количество разрядов в более короткие интервалы между спайками под влиянием более ранней стимуляции. Средняя продолжительность впадины в PSTH 18,5 мс была больше, чем 8–15 мс, о которых сообщают Naito et al.(1996), но их интенсивность стимуляции составляла 1,0–1,3 × MT по сравнению с 0,9 × MT в текущем исследовании. На расхождение в продолжительности впадин PSTH могло повлиять меньшее количество триггеров, которые использовались в настоящем исследовании, но существуют ограничения в точности измерения продолжительности тормозного синаптического потенциала от PSTH (Turker and Powers 2003, 2005). .
Кажущаяся латентность торможения была больше, когда стимуляция проводилась через 30 и 40 мс, но разрешение для точной идентификации начала торможения снижается, когда происходило относительно небольшое количество разрядов в более короткие интервалы между спайками под влиянием более ранней стимуляции. Средняя продолжительность впадины в PSTH 18,5 мс была больше, чем 8–15 мс, о которых сообщают Naito et al.(1996), но их интенсивность стимуляции составляла 1,0–1,3 × MT по сравнению с 0,9 × MT в текущем исследовании. На расхождение в продолжительности впадин PSTH могло повлиять меньшее количество триггеров, которые использовались в настоящем исследовании, но существуют ограничения в точности измерения продолжительности тормозного синаптического потенциала от PSTH (Turker and Powers 2003, 2005). .
Текущее исследование предполагает, что ингибирование длится дольше, чем продолжительность, оцененная по впадинам PSTH. Удлинение интервала между спайками не отличалось при задержке стимула в 30, 40 или 50 мс, несмотря на доказательства того, что короткие (~ 20 мс) тормозные постсинаптические потенциалы (ТПСП) вызывают большее увеличение интервала между спайками, когда ТПСП происходит позже. в интервале между шипами (Turker and Powers 1999).Анализ нескольких интервалов между спайками после стимуляции (Miles et al., 1989; Mattei et al., 2003) показал длительное торможение в течение 150–300 мс после стимуляции (1). Однако эти данные следует интерпретировать с осторожностью из-за влияния удлинения предшествующего интервала между спайками (например, ISI-0) на последующие разряды (т. е. от ISI + 1 до ISI + 4) независимо от какого-либо продолжающегося ингибирования (Turker and Powers 1999).
в интервале между шипами (Turker and Powers 1999).Анализ нескольких интервалов между спайками после стимуляции (Miles et al., 1989; Mattei et al., 2003) показал длительное торможение в течение 150–300 мс после стимуляции (1). Однако эти данные следует интерпретировать с осторожностью из-за влияния удлинения предшествующего интервала между спайками (например, ISI-0) на последующие разряды (т. е. от ISI + 1 до ISI + 4) независимо от какого-либо продолжающегося ингибирования (Turker and Powers 1999).
Найто и др. (1996) оценили центральную задержку торможения как 0.на 7–1,2 мс позже, чем моносинаптическая Ia гомонимная фасилитация, что указывает на то, что в этом пути задействовано более одного синапса. Поскольку в рефлексе участвуют мышцы, которые не являются строгими антагонистами, он может представлять собой форму невзаимного торможения I типа, включающую Ia и Ib афференты и интернейроны (Naito et al., 1996; Pierrot-Deseilligny and Burke, 2005). Однако торможение длится дольше, чем типичная продолжительность (≤10 мс) невзаимного торможения I типа (Pierrot-Deseilligny and Burke 2005). Продолжительное торможение может иметь пресинаптическое происхождение (Hultborn et al., 1987; Pierrot-Deseilligny and Burke, 2005) и включать нарушение разрядки мотонейронов из-за кратковременного снижения Ia-афферентного входа в пул мотонейронов (Priori et al. др. 1998). Такое действие может накладываться на невзаимное торможение типа I, что позволяет предположить, что стимуляция лучевого нерва влияет на двигательные нейроны двуглавой мышцы плеча в спинном мозге через пре- и постсинаптические механизмы.
Продолжительное торможение может иметь пресинаптическое происхождение (Hultborn et al., 1987; Pierrot-Deseilligny and Burke, 2005) и включать нарушение разрядки мотонейронов из-за кратковременного снижения Ia-афферентного входа в пул мотонейронов (Priori et al. др. 1998). Такое действие может накладываться на невзаимное торможение типа I, что позволяет предположить, что стимуляция лучевого нерва влияет на двигательные нейроны двуглавой мышцы плеча в спинном мозге через пре- и постсинаптические механизмы.
Текущее исследование предоставило ограниченные данные о мышцах, от которых произошли афферентные залпы. Двигательные единицы, которые тормозились при электрической стимуляции плечелучевой ветви лучевого нерва, аналогичным образом подавлялись механическими импульсами, доставляемыми к сухожилию или брюшку плечелучевой мышцы. Это контрастирует с возбуждающим эффектом механической активации афферентов от соседнего лучевого разгибателя запястья (Cavallari and Katz, 1989) и подтверждает роль 1а-афферентов от плечелучевой мышцы в торможении, вызванном электрической стимуляцией лучевого нерва. Низкий электрический порог торможения (~0,5 × МТ) согласовывался с участием афферентов 1-й группы. Текущие результаты также отвергают роль кожных афферентов в торможении; на разряд двигательных единиц в двуглавой мышце плеча не влияла стимуляция кожи в двух разных местах на плече.
Низкий электрический порог торможения (~0,5 × МТ) согласовывался с участием афферентов 1-й группы. Текущие результаты также отвергают роль кожных афферентов в торможении; на разряд двигательных единиц в двуглавой мышце плеча не влияла стимуляция кожи в двух разных местах на плече.
Не удалось с достаточной избирательностью стимулировать ветви лучевого нерва к плечевой и плечелучевой мышцам поверхностными электродами. Сила торможения при стимуляции брахиалиса (2.2%), как правило, меньше, чем при стимуляции плечелучевой мышцы (6,1%), но оптимизация стимуляции ветви плечевой мышцы по-прежнему составляла ~0,7 × MT для ветви плечелучевой мышцы. Поскольку интенсивность 0,7 × MT была достаточной, чтобы задержать разряд двигательной единицы двуглавой мышцы плеча (1), это может объяснить наблюдаемое торможение при стимуляции ветви плечевого нерва. Однако исключить вовлечение афферентов от плечевой мышцы не удалось.
Торможение в двух головках двуглавой мышцы плеча
Характеристики рекрутирования и разрядки двигательных единиц в двух головках двуглавой мышцы плеча могут различаться (ter Haar Romeny et al. 1984 год; ван Зуйлен и др. 1988 год; Херрманн и Фландерс, 1998). Например, некоторые двигательные единицы в медиальной части длинной головки двуглавой мышцы плеча имели более низкие пороги рекрутирования, когда супинационные крутящие моменты выполнялись одновременно с крутящим моментом локтевых сгибателей, тогда как двигательные единицы в латеральной части могли рекрутироваться только при приложении крутящего момента сгибателей. сам по себе (ter Haar Romeny et al., 1984). Напротив, двигательные единицы в короткой головке двуглавой мышцы плеча могли быть задействованы только тогда, когда задача включала комбинацию вращательных моментов сгибания и супинации (van Zuylen et al.1988). Однако в этих исследованиях было зарегистрировано несколько двигательных единиц, а более свежие данные указывают на то, что двигательные единицы с различными паттернами рекрутирования непрерывно распределяются по двуглавой мышце плеча, а не группируются в отдельные мышечные области (Herrmann and Flanders, 1998).
1984 год; ван Зуйлен и др. 1988 год; Херрманн и Фландерс, 1998). Например, некоторые двигательные единицы в медиальной части длинной головки двуглавой мышцы плеча имели более низкие пороги рекрутирования, когда супинационные крутящие моменты выполнялись одновременно с крутящим моментом локтевых сгибателей, тогда как двигательные единицы в латеральной части могли рекрутироваться только при приложении крутящего момента сгибателей. сам по себе (ter Haar Romeny et al., 1984). Напротив, двигательные единицы в короткой головке двуглавой мышцы плеча могли быть задействованы только тогда, когда задача включала комбинацию вращательных моментов сгибания и супинации (van Zuylen et al.1988). Однако в этих исследованиях было зарегистрировано несколько двигательных единиц, а более свежие данные указывают на то, что двигательные единицы с различными паттернами рекрутирования непрерывно распределяются по двуглавой мышце плеча, а не группируются в отдельные мышечные области (Herrmann and Flanders, 1998).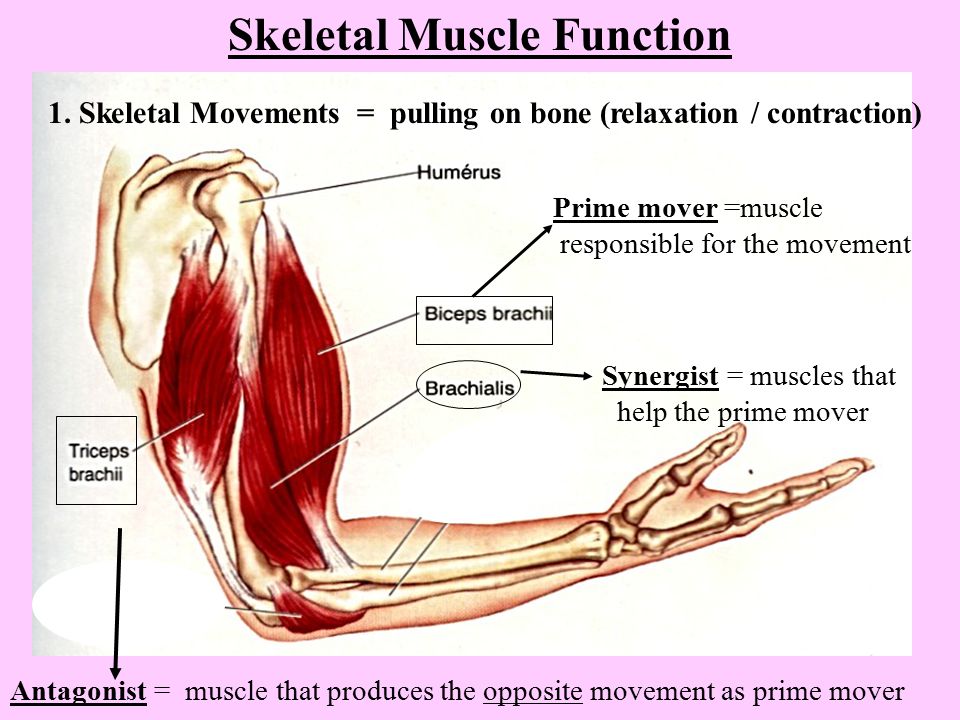 Тем не менее, паттерны активации моторных единиц длинной и короткой головок двуглавой мышцы плеча различаются (Herrmann and Flanders, 1998).
Тем не менее, паттерны активации моторных единиц длинной и короткой головок двуглавой мышцы плеча различаются (Herrmann and Flanders, 1998).
В соответствии с возможными функциональными различиями между двигательными единицами в короткой и длинной головках двуглавой мышцы плеча исследование трупа показало, что дистальное прикрепление сухожилия имело две отдельные части в 10 из 17 образцов и было встречно-пальцевым в других 7 образцах (Eames et al. др.2007). Сухожилие короткой головки прикрепляется дистально к бугристости лучевой кости, что обеспечивает более выгодное положение для создания сгибательных моментов. Сухожилие длинной головки, прикрепляемое проксимально к лучевому бугорку и, судя по его происхождению в супрагленоидном бугорке, имеет более сильное влияние на супинационный крутящий момент (Athwal et al. 2007; Eames et al. 2007).
Было высказано предположение, что пути спинальных рефлекторных путей ответственны за различие в активации популяций двигательных единиц в двух головках двуглавой мышцы плеча (Jongen et al. 1989). Однако результаты, полученные в настоящем исследовании, позволяют предположить, что рефлекторное торможение от плечелучевой мышцы к двуглавой мышце плеча не является одним из путей, которые способствуют избирательной активации двигательных единиц от двух головок двуглавой мышцы плеча, по крайней мере, не для низкопороговых двигательных единиц. . Тер Хаар Ромени и др. (1984) обнаружили, что вариации в активации наблюдались в двигательных единицах с порогом рекрутирования до 45% силы MVC в длинной головке двуглавой мышцы плеча, и, возможно, результаты настоящего исследования ограничены только зарегистрированными низкими значениями. пороговые двигательные единицы.Кроме того, в текущем эксперименте основное внимание уделялось активности двигательных единиц, когда задачей было сгибание локтевого сустава, а действия по оси пронация-супинация не измерялись.
1989). Однако результаты, полученные в настоящем исследовании, позволяют предположить, что рефлекторное торможение от плечелучевой мышцы к двуглавой мышце плеча не является одним из путей, которые способствуют избирательной активации двигательных единиц от двух головок двуглавой мышцы плеча, по крайней мере, не для низкопороговых двигательных единиц. . Тер Хаар Ромени и др. (1984) обнаружили, что вариации в активации наблюдались в двигательных единицах с порогом рекрутирования до 45% силы MVC в длинной головке двуглавой мышцы плеча, и, возможно, результаты настоящего исследования ограничены только зарегистрированными низкими значениями. пороговые двигательные единицы.Кроме того, в текущем эксперименте основное внимание уделялось активности двигательных единиц, когда задачей было сгибание локтевого сустава, а действия по оси пронация-супинация не измерялись.
В качестве альтернативы тормозные рефлекторные пути от других мышц плеча могут отвечать за контроль дифференциальных профилей рекрутирования двигательных единиц в двух головках двуглавой мышцы плеча (Cavallari and Katz, 1989; Jongen et al. , 1989; Naito, 2004). Реципрокное торможение было продемонстрировано между двуглавой и трехглавой мышцами плеча (Katz et al.1991), а коактивация этих мышц во время упражнений на сгибание в локтевом суставе приводит к различным паттернам активации мышц внутри и между двумя головками двуглавой мышцы плеча (Jongen et al. 1989).
, 1989; Naito, 2004). Реципрокное торможение было продемонстрировано между двуглавой и трехглавой мышцами плеча (Katz et al.1991), а коактивация этих мышц во время упражнений на сгибание в локтевом суставе приводит к различным паттернам активации мышц внутри и между двумя головками двуглавой мышцы плеча (Jongen et al. 1989).
Положение предплечья
Двуглавая мышца плеча и плечелучевая мышца действуют как синергисты при сгибании локтя, но могут быть синергистами или антагонистами при приложении крутящего момента вокруг оси пронации-супинации предплечья (Gielen and van Zuylen 1986; Buchanan et al. 1989; Джеймисон и Колдуэлл 1993; Мюррей и др.1995 год; Чжан и др. 1998). Двуглавая мышца плеча всегда имеет момент супинации независимо от положения предплечья, тогда как плечелучевая мышца имеет момент пронации, когда предплечье супинировано, и момент супинации, когда предплечье пронировано (Zhang et al., 1998). Активность ЭМГ двуглавой мышцы плеча больше, когда задача включает одновременные моменты сгибания и супинации по сравнению с моментами сгибания и пронации, но ЭМГ плечелучевой мышцы остается относительно постоянной при изменении положения предплечья (Buchanan et al. 1989). Было показано, что двигательные единицы, расположенные медиально в длинной головке двуглавой мышцы плеча, которые участвуют в моментах сгибания и супинации, умолкают даже при минимальной пронации (ter Haar Romeny et al., 1984). Было высказано предположение, что разрядка этих двигательных единиц «запирается» пронацией, что может объяснить сниженную мышечную активность, наблюдаемую в двуглавой мышце плеча при создании пронационных крутящих моментов или когда предплечье находится в пронированном положении (Buchanan et al., 1986; van Zuylen). и другие.1988 год; Бьюкенен и др. 1989 год; Йонген и др. 1989 год; Джеймисон и Колдуэлл, 1993).
1989). Было показано, что двигательные единицы, расположенные медиально в длинной головке двуглавой мышцы плеча, которые участвуют в моментах сгибания и супинации, умолкают даже при минимальной пронации (ter Haar Romeny et al., 1984). Было высказано предположение, что разрядка этих двигательных единиц «запирается» пронацией, что может объяснить сниженную мышечную активность, наблюдаемую в двуглавой мышце плеча при создании пронационных крутящих моментов или когда предплечье находится в пронированном положении (Buchanan et al., 1986; van Zuylen). и другие.1988 год; Бьюкенен и др. 1989 год; Йонген и др. 1989 год; Джеймисон и Колдуэлл, 1993).
В соответствии с этими наблюдениями результаты текущего исследования включали наибольшую общую силу торможения двуглавой мышцы плеча, когда предплечье находилось в пронации, в сочетании с наименьшей активностью ЭМГ. Это открытие устанавливает основу спинального рефлекса для сниженной активации двуглавой мышцы плеча, когда предплечье пронировано, и согласуется с сообщениями о модуляции H-рефлекса лучевого сгибателя запястья между пронированным и супинированным положениями предплечья (Baldissera et al. 2000). Поскольку рефлекторное торможение двуглавой мышцы плеча было наименьшим при супинации предплечья, кажется вероятным, что уменьшенное торможение обеспечивает большую активацию двуглавой мышцы плеча в этом положении предплечья. Кроме того, Naito et al. (1996) сообщили о торможении только 11 из 21 двигательной единицы двуглавой мышцы плеча, и это могло быть результатом супинированного положения предплечья, использованного в этом исследовании. Хотя задача в текущем исследовании требовала, чтобы испытуемые производили небольшое усилие сгибания, аналогичные паттерны активности ЭМГ между мышцами-сгибателями локтя наблюдались во время более сильных сокращений в направлениях сгибания и пронации или супинации (Jamison and Caldwell 1993).
2000). Поскольку рефлекторное торможение двуглавой мышцы плеча было наименьшим при супинации предплечья, кажется вероятным, что уменьшенное торможение обеспечивает большую активацию двуглавой мышцы плеча в этом положении предплечья. Кроме того, Naito et al. (1996) сообщили о торможении только 11 из 21 двигательной единицы двуглавой мышцы плеча, и это могло быть результатом супинированного положения предплечья, использованного в этом исследовании. Хотя задача в текущем исследовании требовала, чтобы испытуемые производили небольшое усилие сгибания, аналогичные паттерны активности ЭМГ между мышцами-сгибателями локтя наблюдались во время более сильных сокращений в направлениях сгибания и пронации или супинации (Jamison and Caldwell 1993).
Хотя модуляция этого тормозного рефлекса точно соответствовала типичному паттерну активации мышц-сгибателей локтевого сустава, парадоксально, что тормозная обратная связь от плечелучевой мышцы к двуглавой мышце плеча была фактически наибольшей, когда предплечье было пронировано, и обе мышцы вносили вклад в супинационный крутящий момент, и был наименьшим в малом сегменте диапазона движений, когда предплечье было супинировано, а двуглавая и плечелучевая мышцы действовали как антагонисты относительно оси пронация-супинация (Zhang et al. 1998). Однако понимание функционального значения этого пути требует информации о противоположном пути торможения от двуглавой мышцы плеча к плечелучевой (Naito 2004) и оценки рефлекторных действий, когда задача включает в себя действия по оси пронация-супинация в дополнение к крутящий момент в направлении сгибания.
1998). Однако понимание функционального значения этого пути требует информации о противоположном пути торможения от двуглавой мышцы плеча к плечелучевой (Naito 2004) и оценки рефлекторных действий, когда задача включает в себя действия по оси пронация-супинация в дополнение к крутящий момент в направлении сгибания.
Таким образом, не было обнаружено различий в силе торможения, наблюдаемого при разрядке отдельных двигательных единиц, расположенных в двух головках двуглавой мышцы плеча, хотя степень торможения варьировала в зависимости от положения предплечья.Поскольку сила торможения зависела от положения предплечья, результаты предполагают, что этот рефлекс играет роль в модуляции активации двуглавой мышцы плеча. Торможение было наибольшим, когда предплечье находилось в пронированном положении, а активация двуглавой мышцы плеча была снижена, но в этом положении обе мышцы могут способствовать супинации.
Коактивация агонистов и антагонистов повышает кортикомоторную возбудимость мышц голеностопного сустава
Спинальные пути, лежащие в основе реципрокных сгибательно-разгибательных сокращений, хорошо охарактеризованы, но степень влияния активации мышц-антагонистов на кортикально вызванные моторно-вызванные потенциалы (МВП) остается неясной. Большинство исследований с использованием транскраниальной магнитной стимуляции (ТМС) вызывали МВП для оценки возбудимости корково-спинномозгового пути, фокусируясь на мышцах верхних конечностей. Из-за различий в функциональном и нервном контроле между мышцами нижних и верхних конечностей необходимо оценить методологические факторы, влияющие на МВП, вызванные ТМС, особенно в мускулатуре нижних конечностей. Влияет ли и в какой степени активация нецелевых мышц, таких как мышцы-антагонисты, на МВП, вызванные ТМС, плохо изучено, и такие пробелы в наших знаниях могут ограничивать строгость и воспроизводимость исследований ТМС.Здесь мы оценили влияние состояния активации мышцы-антагониста на MEP, вызванные ТМС, полученные от целевой (агонистической) мышцы лодыжки как для передней большеберцовой (TA), так и для камбаловидной мышцы. Четырнадцать здоровых участников (11 женщин, возраст: годы) завершили одну экспериментальную сессию; в анализ были включены данные 12 человек. ТМС доставляли в 4 условиях: отдых, активация ТА, активация камбаловидной мышцы и коактивация ТА и камбаловидной мышцы.
Большинство исследований с использованием транскраниальной магнитной стимуляции (ТМС) вызывали МВП для оценки возбудимости корково-спинномозгового пути, фокусируясь на мышцах верхних конечностей. Из-за различий в функциональном и нервном контроле между мышцами нижних и верхних конечностей необходимо оценить методологические факторы, влияющие на МВП, вызванные ТМС, особенно в мускулатуре нижних конечностей. Влияет ли и в какой степени активация нецелевых мышц, таких как мышцы-антагонисты, на МВП, вызванные ТМС, плохо изучено, и такие пробелы в наших знаниях могут ограничивать строгость и воспроизводимость исследований ТМС.Здесь мы оценили влияние состояния активации мышцы-антагониста на MEP, вызванные ТМС, полученные от целевой (агонистической) мышцы лодыжки как для передней большеберцовой (TA), так и для камбаловидной мышцы. Четырнадцать здоровых участников (11 женщин, возраст: годы) завершили одну экспериментальную сессию; в анализ были включены данные 12 человек. ТМС доставляли в 4 условиях: отдых, активация ТА, активация камбаловидной мышцы и коактивация ТА и камбаловидной мышцы. Были проведены три попарных сравнения амплитуды ВМО и коэффициента вариабельности (CV): отдых против коактивации, отдых против активации антагониста и активация агониста против коактивации.Мы продемонстрировали, что коактивация агониста-антагониста увеличивала амплитуду ВМО и уменьшала CV ВОС как для TA, так и для камбаловидной мышцы. Наши результаты обеспечивают методологические соображения для будущих исследований ТМС и прокладывают путь для будущих исследований зависимой от коактивации модуляции кортикомоторной возбудимости в патологических когортах, таких как инсульт или травма спинного мозга.
Были проведены три попарных сравнения амплитуды ВМО и коэффициента вариабельности (CV): отдых против коактивации, отдых против активации антагониста и активация агониста против коактивации.Мы продемонстрировали, что коактивация агониста-антагониста увеличивала амплитуду ВМО и уменьшала CV ВОС как для TA, так и для камбаловидной мышцы. Наши результаты обеспечивают методологические соображения для будущих исследований ТМС и прокладывают путь для будущих исследований зависимой от коактивации модуляции кортикомоторной возбудимости в патологических когортах, таких как инсульт или травма спинного мозга.
1. Введение
Транскраниальная магнитная стимуляция (ТМС) — это метод неинвазивной стимуляции головного мозга, который можно использовать для оценки нисходящей кортикомоторной активности мышц голеностопного сустава при выполнении как статических [1], так и динамических задач [2].Амплитуда моторных вызванных потенциалов (МВП), возникающих в ответ на ТМС, отражает кумулятивную возбудимость корково-спинномозгового пути [3, 4]. Из-за различий в анатомии и физиологии нервных цепей управления, контролирующих мышцы конечностей, возникает необходимость оценки методологических факторов, влияющих на МВП, вызванные ТМС, именно в мускулатуре нижних конечностей [5, 6]. Ключевым препятствием для использования вызванных ТМС амплитуд МВП в качестве меры возбудимости первичной моторной коры (М1) является их чувствительность к одновременным изменениям возбудимости подкорковых и сегментарных спинальных цепей [7, 8].Кроме того, МВП, вызванные ТМС, могут модулировать за счет активации целевых мышц ноги, контралатеральной по отношению к стимулируемому полушарию [9–12], а также активации гомологичных мышц в ноге, ипсилатеральной по отношению к стимулируемому полушарию [13]. Недавняя работа нашей и других лабораторий показала, что постуральное состояние также влияет на вызванную ТМС амплитуду ВМО мышц ног [14–16]. Однако вопрос о том, влияет ли активация нецелевых мышц, таких как мышцы-антагонисты, на выход МВП, вызванный ТМС, и в какой степени, плохо изучен, и такие пробелы в наших знаниях могут ограничивать строгость и воспроизводимость исследований ТМС, сосредоточенных на нижних конечностях.
Нервный контроль мышц-агонистов и мышц-антагонистов (например, тыльных сгибателей голеностопного сустава и подошвенных сгибателей) организован реципрокно, так что активация агониста сопровождается одновременным торможением мышцы-антагониста. При активации передней большеберцовой мышцы (ПП) угнетаются Н-рефлексы с мышцы-антагониста (камбаловидной мышцы) [17]. По сравнению с состоянием покоя, во время активации ТА мышц наблюдается усиление дисинаптического спинального реципрокного торможения от ТА к мотонейронам камбаловидной мышцы, опосредованное интернейронами Ia [17].Интересно, что в отличие от этого реципрокного ингибирования мотонейронов-антагонистов в спинальных сегментарных цепях, было показано, что ТМС-вызванные МВП из камбаловидной мышцы облегчаются во время [12] и до [7] произвольного тыльного сгибания. Таким образом, при усилении реципрокного торможения мотонейрона камбаловидной мышцы и угнетении Н-рефлексов камбаловидной мышцы при активации ТА корково-вызванные ВВП камбаловидной мышцы могут проявлять облегчение при активации ТА [7]. Герцен и др. предполагают, что во время выполнения функциональных задач, требующих быстрой модуляции направления движения, произвольному сокращению голеностопного сустава предшествует стимуляция антагонистов, вероятно, опосредованная подкорковыми моторными программами.В то время как спинальные дисинаптические пути, лежащие в основе реципрокных сокращений сгибания-разгибания голеностопного сустава, были широко охарактеризованы, степень, в которой изменения возбудимости коры или нисходящего движения, фиксируемые амплитудой MEP, зависят от активации мышц-антагонистов, остается неясной.
Эксперименты как на нечеловеческих приматах [18, 19], так и на людях [20, 21] свидетельствуют о специализированном нисходящем контроле коактивации агонистов-антагонистов по сравнению с изолированными движениями сгибания и разгибания.Во время коактивации спинальное реципрокное торможение между антагонистическими мышцами голеностопного сустава угнетается, вероятно, за счет нисходящей супрессии интернейронов Ia. Во время коактивации агонист-антагонист усиление специфического для коактивации нисходящего влечения модулирует сегментарную цепь позвоночника и может влиять на амплитуду MEP, индуцированную ТМС. Коактивация мышц-агонистов и мышц-антагонистов является важным компонентом нормального моторного контроля. Коактивация является ключевой стратегией прямой связи, используемой для регулирования жесткости сустава [22] и увеличения импеданса сустава в ответ на внешние возмущения [23] для задач, требующих высокой точности [24] или при нарушении стабильности сустава [25, 26].Задания, требующие специфических для коактивации двигательных программ, например стояние на неустойчивой платформе, сопровождаются снижением реципрокного торможения [27] и одновременным снижением амплитуды рефлекса [25, 28]. Кроме того, повышенная коактивация агонистов и антагонистов во время функциональных задач является распространенным нарушением моторного контроля, наблюдаемым у людей с нейропатологией, такой как инсульт [29, 30]. Таким образом, использование ТМС для изучения изменений кортикомоторной возбудимости во время задачи коактивации голеностопного сустава дает возможность получить представление о нейромоторном контроле мышц нижних конечностей.
Относительно немногочисленные данные явно характеризуют, как коактивность мышц-антагонистов влияет на MEP, вызванные ТМС. Например, Geertsen et al. продемонстрировали, что произвольное сокращение в голеностопном суставе сопровождается предшествующим облегчением антагонистов, вероятно, опосредованным подкорковыми цепями [7]. Модуляция возбудимости тормозных интернейронов 1а, альфа-мотонейронов и подкорковых участков нисходящего торможения может заметно влиять на то, как вызванный ТМС нисходящий залп активирует конечный общий путь (нижний пул мотонейронов), а также на размер МЭП, вызванных от мишени. мышцы во время ТМС.Исследования ТМС обычно регистрируют и отслеживают фоновую активацию и МВП только от целевой мышцы. Тем не менее, активация мышц-синергистов и антагонистов может модулировать возбудимость супраспинальных и спинномозговых сегментарных цепей и влиять на вызванные ТМС МЭП-ответы целевой мышцы. Понимание того, как состояние активации мышцы-антагониста влияет на амплитуду ВМО, вызванную ТМС, может дать информацию для интерпретации и дизайна исследований ТМС. Таким образом, целью данного исследования было оценить влияние состояния активации мышцы-антагониста на вызванные ТМС MEP, полученные от целевой (агонистической) мышцы лодыжки как для ТА, так и для камбаловидной мышцы.
2. Методы
Процедуры исследования были одобрены Институциональным наблюдательным советом Университета Эмори. Все участники предоставили информированное письменное согласие в соответствии с Хельсинкской декларацией.
2.1. Участники
В это исследование были включены девятнадцать молодых людей с неврологическими нарушениями, не имевших в анамнезе ортопедических или неврологических заболеваний. Критерии исключения включали предшествующие или текущие неврологические расстройства и противопоказания к ТМС, включая судороги в анамнезе, металлические имплантаты в голове, недавнее сотрясение мозга, использование препаратов, действующих на центральную нервную систему, беременность или эпизод обморока или потери сознания за последние 12 месяцев. [31].Пять из 19 участников не завершили протокол исследования или были исключены из-за противопоказаний к ТМС, включая случай обморока [32], прием лекарств, предшествующее сотрясение мозга и головокружение/дискомфорт во время ТМС. Остальные 14 участников (11 женщин, возраст: годы) завершили протокол исследования.
2.2. Дизайн исследования
Представленные данные были собраны во время одного сеанса в рамках более крупного исследования, изучающего влияние мышечной активации, осанки и ходьбы на возбудимость коры нижних конечностей.
2.2.1. Процедуры электромиографии
После стандартных процедур подготовки кожи для минимизации сопротивления кожи к коже над животом прикрепляли биполярные, круглые и самоклеящиеся поверхностные датчики ЭМГ (диаметр 11 мм, межэлектродное расстояние 11 мм, BIOPAC Systems Inc., США). правой ТА и камбаловидной мышцы, с общим датчиком земли, прикрепленным над правой латеральной лодыжкой [33]. Размещение датчика ЭМГ было подтверждено проверкой сигнала ЭМГ во время произвольного сокращения мышц.Данные ЭМГ были отобраны на частоте 1000 Гц, отфильтрованы полосой пропускания от 10 Гц до 500 Гц и амплифицированы на 2000 (программное обеспечение AcqKnowledge, BIOPAC Systems Inc.). Чтобы обеспечить постоянное расположение датчиков на протяжении всего сеанса и свести к минимуму артефакты движения ЭМГ, датчики и провода ЭМГ были надежно закреплены на ноге (Sensi-Wrap, Dynarex Inc.).
2.2.2. Активность ЭМГ во время максимальных произвольных сокращений (МВС)
Данные ЭМГ были собраны, когда участник выполнял 3 максимальных произвольных сокращения (МВС) продолжительностью 5 с правой камбаловидной и ТА мышц.Для камбаловидной MVC участников просили встать на правую ногу и подошвенно согнуть лодыжку, отрывая пятку от земли. Для TA MVC участник максимально согнул лодыжку, преодолевая сопротивление, в положении сидя. 3-секундное окно во время пикового сокращения использовалось для расчета средней среднеквадратичной амплитуды ЭМГ, а максимальное значение из 3 сокращений было определено как ЭМГ MVC. MVC EMG использовался для установки низкого целевого уровня фоновой активации (10% MVC), который участники будут поддерживать во время оценок TMS.
2.2.3. Процедуры TMS
Одиночные импульсы TMS подавались с использованием специальной катушки в форме восьмерки «крыло летучей мыши», которая имеет немного другой угол и форму, чем катушка с двойным конусом (диаметр 70 мм, Magstim Company Ltd., Великобритания). подключен к монофазному стимулятору (Magstim 200 2 ). Стереотаксическая нейронавигация (Brainsight, Rogue Research Inc., Канада) использовалась для отслеживания трехмерного положения и ориентации катушки, при этом голова участника была зарегистрирована в стандартном шаблоне мозга (MNI 152) на протяжении всего эксперимента.Во время ТМС катушку удерживали тангенциально к коже головы, а ручку катушки — параллельно межполушарной щели, чтобы индуцировать заднепередний ток в коре головного мозга. Горячая точка для правой ТА-мышцы в контралатеральной (левой) первичной моторной коре (М1) определялась как оптимальное положение катушки, вызывающее максимальные МЭП-ответы от правой ТА-мышцы при самой низкой интенсивности стимулятора [34]. Для каждого участника положение катушки и ориентация в точке доступа были сохранены и использовались в качестве эталона на протяжении всего сеанса.С помощью компьютеризированного адаптивного алгоритма [35] определяли моторный порог покоя (rMT) с амплитудным критерием ВМО 80 мк В [34]. Критериальная амплитуда МЭП для rMT была увеличена с 50 мк В [34] до 80 мк В, чтобы гарантировать получение различимых МЭП в состоянии покоя как от ТА, так и от камбаловидной мышцы. Для оставшегося экспериментального протокола интенсивность ТМС поддерживалась на одном и том же надпороговом уровне (120 или 130% от rMT) для всех условий тестирования.
2.2.4. Биологическая обратная связь ЭМГ для поддержания согласованности фоновой активации ЭМГ
Необработанные сигналы ЭМГ от правого предплечья и камбаловидной мышцы отправлялись в виде аналоговых выходов с аппаратного обеспечения BIOPAC на плату сбора данных (USB-6343 X-Series, National Instruments) и вводились в пользовательскую ЭМГ. программа биологической обратной связи (LabVIEW, National Instruments) для расчета среднеквадратичной (RMS) ЭМГ для TA и камбаловидной мышцы и для визуального отображения RMS EMG для участника в режиме реального времени. Когда условия тестирования требовали, чтобы была активирована одна мышца (например,g., активирована камбаловидная мышца), участнику была предоставлена биологическая обратная связь, чтобы помочь поддерживать активацию конкретной мышцы в пределах целевого окна ЭМГ (10% MVC, ). Когда условие тестирования требовало, чтобы обе мышцы были активированы (например, коактивация), биологическая обратная связь обеспечивалась для обеих мышц одновременно. Биологическая обратная связь ЭМГ гарантировала, что участники поддерживали желаемые фоновые уровни активации ЭМГ для каждого состояния и что величина фоновой ЭМГ для мышцы (например, ТА) была постоянной в разных условиях тестирования (например,г., между активацией ТА и коактивацией).
2.2.5. Сбор данных при различных условиях тестирования ТМС
Специально написанная программа (AcqKnowledge, BIOPAC Systems Inc.) использовалась для доставки 10-15 импульсов ТМС с частотой 0,2 Гц. Сбор данных был организован таким образом, что 50 мс данных были собраны до и 450 мс после импульса ТМС, с общим временем сбора 500 мс для каждого импульса ТМС (рис. 1). Участники удобно сидели на стуле с опорой на спину, шею без опоры, колено и бедро сгибались под углом 90°, а лодыжка — под углом 0° тыльного сгибания.Затем выравнивание ног участников было отмечено лентой на напольном коврике, чтобы обеспечить постоянное положение ног во время сеанса. Груз на лодыжку помещали на спину для стабилизации. МВП, вызванные ТМС, были собраны в течение 4 условий тестирования в случайном порядке (рис. 1): (1) Отдых : камбаловидная мышца и камбаловидная мышца находятся в состоянии покоя(2) TA активирована : TA активирована, а камбаловидная мышца находится в состоянии покоя(3) камбаловидная мышца активирована : камбаловидная мышца активирована, а TA находится в состоянии покоя (4) Коактивация : были активированы как TA, так и камбаловидная мышца
Для всех условий тестирования ТМС доставлялась катушкой над горячей точкой правой TA (определено в начале сеанса), и MEP были одновременно регистрируется с ТА и камбаловидной мышцы.В этом исследовании мышца-агонист была мышцей-мишенью во время ТМС (т. е. мышцей, от которой ТМС-вызванные ВОС оценивались как зависимая переменная). При оценке вызванных ТМС МВП от ТА мышцы агонист относится к ТА, а антагонист к камбаловидной мышце. При оценке МВП, вызванных ТМС, для камбаловидной мышцы агонист относится к камбаловидной мышце, а антагонист к ТА.
Во время активации TA или камбаловидной мышцы участникам была предоставлена визуальная биологическая обратная связь о текущей ЭМГ для мышцы, которую необходимо было активировать.Участникам было предложено поднять ногу вверх (тыльное сгибание) или вниз (подошвенное сгибание), чтобы увеличить текущую ЭМГ до целевого значения. Во время состояния коактивации участникам показывали текущую ЭМГ как для TA, так и для камбаловидной мышцы на экране биологической обратной связи EMG, и их просили попытаться активировать каждую мышцу до целевого значения. При необходимости им давали словесные инструкции, такие как «постарайтесь напрячь лодыжку, отталкиваясь вверх от подушечки для стопы», чтобы помочь в выполнении задания на коактивацию.Данные этих 4 состояний использовались для проведения 3 сравнений для каждой мышцы (TA и камбаловидной мышцы) и для проверки следующих 3 гипотез, перечисленных ниже. Мы не сравнивали отдых с активацией агонистов, потому что это было хорошо изучено ранее. 3 сравнения следующие: Сравнение (1) Отдых по сравнению с коактивацией : как для TA, так и для камбаловидной мышцы MEP, мы предположили, что по сравнению с состоянием покоя вызванные TMS MEP будут усиливаться во время коактивации. Сравнение (2) Отдых по сравнению с активацией антагониста : как для TA, так и для камбаловидной мышцы, мы предположили, что по сравнению с состоянием покоя вызванные ТМС MEP будут облегчаться во время состояния активации антагонистаСравнение (3) Активация агониста по сравнению с коактивацией : как для TA, так и для MEP камбаловидной мышцы при сходных фоновых уровнях активации агониста ( мишень) мышцы, мы предположили, что MEP, вызванные ТМС, будут больше во время коактивации агониста-антагониста по сравнению с активацией только агониста
2.3. Анализ данных
Предварительная стимуляция ЭМГ фоновой среднеквадратичной амплитуды как для TA, так и для камбаловидной мышцы была рассчитана для окна 50 мс до проведения ТМС (рис. 1). Исходные данные ЭМГ RMS были проанализированы исследователями для каждого сравнения; испытания, в которых ЭМГ субъекта не соответствовала тестируемому состоянию (оцениваемому с помощью парных тестов между состояниями в испытаниях MEP отдельных субъектов), были удалены, чтобы гарантировать, что сравнения между различными условиями тестирования не были искажены различиями в фоновой активности ЭМГ.Размах амплитуды ТА и камбаловидной мышцы вычисляли для каждой попытки как разницу между максимумами и минимумами в пределах окна МВП. Для каждого испытуемого в качестве меры корково-спинномозговой возбудимости во время каждого условия тестирования использовались средние амплитуды от пика до пика для 10-15 испытаний MEP. Для каждого состояния вариабельность амплитуды МВП от испытания к испытанию индексировали путем расчета коэффициента вариации (CV) (стандартное отклонение амплитуд МВП, деленное на ).
2.4. Статистический анализ
Первичными зависимыми переменными были амплитуды ВМО для ТА и камбаловидной мышцы. Поскольку вариации фоновой преактивации ЭМГ могут влиять на МВП, вызванные ТМС, данные среднеквадратичного значения фоновой ЭМГ проверялись для отдельных испытаний и участников, а также оценивались как вторичная зависимая переменная. Дополнительные вторичные переменные включали CV амплитуды MEP от испытания к испытанию. Мы оценили предположение о нормальном распределении, используя критерий Колмогорова-Смирнова, который был выполнен для наших зависимых измерений, за исключением амплитуды TA MEP для парных состояний и среднеквадратичного значения фоновой ЭМГ в состоянии покоя.
ANOVA с повторными измерениями использовали для сравнения первичных переменных (амплитуда TA и MEP камбаловидной мышцы) среди 4 тестовых условий (отдых, включение агониста, включение антагониста и коактивация). Кроме того, для каждой мышцы были проведены заранее определенные попарные сравнения, чтобы сделать 3 конкретных сравнения: отдых против коактивации, отдых против активации антагониста и активация агониста против коактивации. Повторные измерения одностороннего ANOVA с апостериорными парными тестами также проводились для вторичных переменных (среднеквадратичное значение ЭМГ фона, CV амплитуды MEP).
Кроме того, для систематической оценки данных амплитуды МВП у отдельных субъектов для каждого парного сравнения рассчитывались баллы изменений как разница в амплитуде МВП для двух условий сравнения (например, амплитуда МВП во время коактивации минус амплитуда МВП в состоянии покоя). Оценки изменения MEP были нанесены на график для каждого участника и использованы для оценки межиндивидуальной вариабельности оценок изменений и для оценки того, включает ли 95% доверительный интервал (ДИ) оценки изменений ноль [36].Аналогичным образом, для проверки данных фоновой ЭМГ были рассчитаны и нанесены на график среднеквадратичные значения фоновой ЭМГ баллы изменений для каждого сравнения. Показатели изменения фоновой ЭМГ использовались для подтверждения того, что фоновая ЭМГ соответствовала интересующему сравнению, и для исключения выбросов, то есть любого участника с показателями изменения, которые превышали стандартное отклонение среднего значения для этого сравнения в 2,5 раза.
Для сравнения величины модуляции вызванных ТМС МВП, индуцированных изменением условий тестирования, для каждой мышцы (TA и камбаловидной мышцы) был выполнен односторонний ANOVA с повторными измерениями с попарными сравнениями post hoc для сравнения показателей изменения МВП среди мышц. 3 исследования сравнения.Три показателя изменения были (коактивация минус отдых), (активация антагониста минус отдых) и (коактивация минус активация агониста). Для статистического анализа использовалась версия 21 SPSS. Альфа-уровень был установлен на уровне 0,05.
3. Результаты
Двое из 14 участников, завершивших исследование, были исключены из всех сравнений из-за того, что их фоновая активация превышала указанное целевое значение.
3.1. Общие эффекты состояния активации мышцы-антагониста
Повторные измерения ANOVA выявили значительное основное влияние условий тестирования как на амплитуды TA (;), так и на амплитуды MEP камбаловидной мышцы (;).ANOVA, сравнивающий коэффициент вариации между испытаниями (CV) амплитуды MEP при 4 состояниях, показал основное влияние состояния на TA (;) и камбаловидную мышцу (;). Повторные измерения ANOVA выявили значительное основное влияние условий тестирования как на TA (; ), так и на среднеквадратичные амплитуды фоновой ЭМГ (; ) камбаловидной мышцы. Результаты апостериорных парных сравнений перечислены для каждого из сравнений ниже.
3.2. Сравнение покоя и коактивации агонистов-антагонистов
3.2.1. TA MEP Amplitudes
Амплитуда MEP для TA была значительно больше () во время коактивации () по сравнению с отдыхом () (рис. 2 (a), (i)). Индивидуальные оценки изменений участников показали, что у всех участников наблюдалось увеличение амплитуды MEP во время коактивации (оценка положительных изменений) по сравнению с состоянием покоя, а 95% ДИ оценки изменений не включал ноль (рис. 2 (a), (ii)). Отмечалось значительно большее () значение CV амплитуд МВП в покое () по сравнению с коактивацией () (рис. 3(b)).
3.2.2. Амплитуда камбаловидной мышцы
Амплитуда камбаловидной мышцы была значительно больше () во время коактивации () по сравнению с состоянием покоя () (рис. 4 (a), (i)). Индивидуальные оценки изменения субъекта показали, что у всех участников наблюдалось увеличение амплитуды MEP во время коактивации по сравнению с состоянием покоя, а 95% ДИ оценки изменения не включал ноль (рис. 4 (a), (ii)). Отмечалось значительно большее () значение CV амплитуд камбаловидной мышцы в покое () по сравнению с коактивацией () (рис. 3(d)).
3.2.3. Фоновая электромиография (ЭМГ) Активация
Среднеквадратичная амплитуда (RMS) фоновой ЭМГ для ТА была значительно ниже () во время отдыха () по сравнению с коактивацией () (рис. 2(a), (iii) и 4(a). , (iii)). Точно так же среднеквадратичное значение фоновой ЭМГ для камбаловидной мышцы было значительно ниже () во время отдыха () по сравнению с коактивацией () (рис. 2 (а), (iii) и 4 (а), (iii)).
3.3. Сравнение покоя и активации антагониста
3.3.1. Амплитуды TA MEP
Амплитуды TA MEP были значительно выше () во время активации антагониста (камбаловидной мышцы) () по сравнению с состоянием покоя () (рис. 2(b), (i)). Индивидуальные оценки изменений для TA MEP показали, что у всех испытуемых, кроме одного, наблюдалось увеличение амплитуды MEP (показатель положительного изменения) во время активации антагониста по сравнению с состоянием покоя, при этом 95% ДИ не включал ноль (рис. 2(b), (ii)). Отмечалось значительно большее () значение CV амплитуд МВП в покое в сравнении с активацией антагониста (рис. 3(b)).
3.3.2. Амплитуды МВП камбаловидной мышцы
Амплитуды МВП камбаловидной мышцы были значительно выше () во время активации антагониста (ТА акт.) () по сравнению с состоянием покоя () (рис. 4(b), (i)). Индивидуальные оценки изменения амплитуды камбаловидной мышцы показали, что у большинства испытуемых наблюдалось увеличение амплитуды МВП во время активации антагониста по сравнению с состоянием покоя (оценка положительных изменений) с 95% ДИ, не включая ноль (рис. 4(b), (ii)). Во время отдыха наблюдался значительно больший CV между испытаниями по сравнению с активацией антагониста (рис. 3(d)).
3.3.3. Фоновая ЭМГ
Наблюдалась значительная разница () в фоновой ЭМГ ТА между активацией покоя () и антагониста (камбаловидная мышца) (рис. 2(b), (iii)). Наблюдалась значительная разница () в фоновой ЭМГ камбаловидной мышцы между отдыхом () и активацией антагониста (камбаловидной мышцы) () (рис. 2(b), (iii)).
Наблюдалась значительная разница () в фоновой ЭМГ ТА между отдыхом () и активацией антагониста (ТА) () (рис. 4(b), (iii)). Наблюдалась значительная разница () в фоновой ЭМГ камбаловидной мышцы между отдыхом () и активацией антагониста (ТА) () (рис. 4(b), (iii)).
3.4. Сравнение активации агонистов и коактивации
3.4.1. Амплитуды TA MEP
Амплитуды TA MEP были значительно выше () во время коактивации () по сравнению с активацией агонистом () (рис. 2(c), (i)). Показатели индивидуальных изменений показали высокую межсубъектную изменчивость; однако у большинства испытуемых наблюдалось увеличение амплитуды ВМО во время коактивации по сравнению с активацией агонистом, а 95% ДИ не включал ноль (рис. 2(c), (ii)). Амплитуда CV TA MEP не показала существенной разницы () во время активации агониста по сравнению с коактивацией (рис. 3(b)).
3.4.2. Амплитуда камбаловидной мышцы
Амплитуда камбаловидной мышцы была значительно больше () во время коактивации () по сравнению с активацией агонистом () (рис. 4 (c), (i)). Показатели индивидуальных изменений показали высокую межсубъектную вариабельность, при этом у большинства субъектов наблюдалось увеличение амплитуды камбаловидной мышцы при коактивации по сравнению с активацией агонистом, а 95% ДИ не включал ноль (рис. 4 (c), (ii)). CV камбаловидной мышцы были значительно выше () во время активации агонистом по сравнению с коактивацией (рис. 3(d)).
3.4.3. Фоновая ЭМГ
Не было различий () в фоновой ЭМГ ТА между активацией агониста (ТА) () и коактивацией () (рис. 2(c), (iii)). Существовала значительная разница () в фоновой ЭМГ камбаловидной мышцы между активацией агонистом (ТА) () и коактивацией () (рис. 2(c), (iii)).
Существовала значительная разница () в фоновой ЭМГ TA между активацией агониста (камбаловидная мышца) () и коактивацией () (рис. 4 (c), (iii)). Не было различий () в фоновой ЭМГ камбаловидной мышцы между активацией агониста (камбаловидной мышцы) () и коактивацией () (рис. 4 (с), (iii)).
3.5. Сравнение модуляции амплитуды МВП с различной активацией антагониста
Односторонний дисперсионный анализ с повторными измерениями, оценивающий влияние 3 типов сравнения на изменение амплитуды МВП, показал основное влияние типа сравнения как для ТА (; ), так и для камбаловидной мышцы (; ) Показатели изменения MEP (рис. 3). Попарные апостериорные сравнения для ТА мышцы показали, что наибольшее изменение амплитуды ВМО наблюдалось при переходе от покоя к коактивации. Показатели изменения TA MEP для коактивации-отдыха были значительно выше, чем показатели изменения для антагонист-отдых () и коактивация-агонист () (рис. 3(а)).Аналогичным образом, апостериорные сравнения показали, что для камбаловидных МВП показатели изменения МВП для коактивации-отдыха были значительно выше, чем для антагониста-покоя (), но не отличались от показателей изменения коактивации-агониста (). Наблюдалась значительная разница между антагонистом-покоем и коактивацией-агонистом для камбаловидной мышцы () (рис. 3(с)).
4. Обсуждение
Наши результаты показали, что коактивация мышц агонистов и антагонистов значительно усиливает MEP, вызванные ТМС, регистрируемые в мышцах голеностопного сустава.Как для ТА, так и для камбаловидной мышцы, по сравнению с состоянием, когда обе мышцы находились в состоянии покоя, коактивация агониста-антагониста приводила к большей амплитуде ВМО. После сопоставления фоновой ЭМГ-активации мышцы-агониста между состояниями амплитуда ВМО была значительно выше во время коактивации по сравнению с активацией агониста. Наши результаты показывают, что во время задачи произвольной коактивации специфический для коактивации нисходящий драйв и модуляция реципрокного торможения позволяют нисходящим залпам, индуцированным ТМС, вызывать более крупные MEP.Мы также продемонстрировали, что по сравнению с отдыхом активация мышцы-антагониста или участие в коактивации агонист-антагонист приводит к меньшей вариабельности амплитуды МВП от попытки к попытке (CV). Таким образом, коактивация агонистов и антагонистов, а также активация антагонистов могут быть возможными стратегиями для увеличения вероятности получения устойчивых и измеримых МВП от мышц нижних конечностей, особенно у лиц с неврологическими нарушениями, которые демонстрируют повышенный двигательный порог. Кроме того, наши результаты подчеркивают важность мониторинга фоновой активации мышц-антагонистов во время экспериментов с ТМС.
Увеличение размера МВП, вызванного ТМС, было документально подтверждено параллельным увеличением произвольной фоновой активации мишени или агониста как в мышцах верхних конечностей [37], так и в мышцах нижних конечностей [1]. Тем не менее, существует мало данных о том, влияет ли коактивация антагонистов на амплитуды МВП. Здесь мы провели прямое сравнение влияния коактивности антагонистов на MEP-агонисты. Уникальным аспектом нашего подхода было то, что мы сравнивали коактивацию агонистов и антагонистов с условиями покоя, а также с условиями активации только агонистов.Наши результаты показали, что по сравнению с отдыхом, что неудивительно, амплитуда MEP целевой мышцы увеличилась в условиях коактивации. Что еще более важно и интересно, коактивация также индуцировала значительное увеличение амплитуды MEP по сравнению с состоянием активации агониста, несмотря на незначительную разницу между состояниями в уровне фоновой активации агониста или целевой мышцы. Таким образом, взятые вместе, оба этих сравнения подтверждают наш вывод о том, что коактивация агониста и антагониста увеличивает амплитуду MEP, вызванную ТМС, в мышце-мишени или мышце-агонисте.
Активнозависимая модуляция возбудимости спинного мозга ранее изучалась при тонических [21, 38] и динамических задачах [17], а также при ходьбе [2]. Несмотря на эту характеристику, в нескольких исследованиях ТМС использовалась для явной оценки корково-спинальной возбудимости пары мышц-антагонистов/агонистов во время задачи на волевую коактивацию. На спинно-сегментарном уровне коактивация регулируется различными спинальными механизмами, включая реципрокное торможение [39] и пресинаптическое торможение афферентов Ia [40].Однако на корковом уровне нисходящая регуляция передачи внутри спинальных тормозных интернейронов Ia играет роль в обеспечении коактивации мышц-антагонистов [21, 41]. Потенциально, некоторые корково-спинномозговые нейроны могут быть коактивационно-специфичными [18], однако степень, в которой коактивационно-зависимое увеличение вызванных ТМС МВП отражает активацию возбудимости в коактивационно-специфических нисходящих цепях, еще предстоит определить. Хотя наши методы и результаты не могут проанализировать вклад специфического механизма нервного контроля в коактивацию мышц голеностопного сустава, они предоставляют уникальные доказательства повышенной корково-спинальной возбудимости, которая сопровождает коактивацию агонистов-антагонистов.
Предыдущие предположения о более сильном нисходящем входе в пулы ТА-мотонейронов [42] по сравнению с камбаловидной мышцей [43] приводят к вопросу о том, различаются ли активация антагониста или зависимые от коактивации влияния на амплитуду ВМО между камбаловидной и ТА-мышцами. Герцен и др. ранее сообщалось об облегчении МВП камбаловидной мышцы во время сокращений антагонистов до и во время начала движения [7]. Интересно, что здесь мы наблюдали реципрокный эффект увеличения MEP во время коактивации, независимо от того, функционировали ли TA или камбаловидная мышца как агонист или мышца-мишень во время TMS.Во время функциональных задач, связанных с голеностопным суставом, нисходящий контроль, характерный для коактивации, может подавлять передачу при реципрокном торможении [44, 45], чтобы обеспечить коактивацию пулов агонистов и антагонистов мотонейронов. Разъединение нисходящего входа к тормозным интернейронам Ia и нисходящего импульса к мотонейронам может облегчить задачи коактивации и, по-видимому, проявляется как для TA, так и для камбаловидной мышцы.
Другие данные свидетельствуют о дифференциальной активации коры во время коактивации по сравнению с задачами активации только агонистов.Кроме того, активация мышц голеностопного сустава, как преднамеренная, так и в ответ на залп, индуцированный ТМС, не обязательно может активировать изолированные пулы мотонейронов. Визуализирующие исследования выявляют четкий и более широкий паттерн церебральной активации M1 во время коактивации по сравнению с активацией только агонистов или только антагонистов после сопоставления уровней ЭМГ при изолированном тыльном/подошвенном сгибании [46]. Однако, в то время как повышенная активация может указывать на повышенное использование подмножества корковых сетей, специфичных для коактивации, потребность коры, связанная со сложностью задачи, может альтернативно объяснять интенсивность корковой активации.Активация коры связана с потребностью [47] и типом двигательной задачи [46, 48]. Используемая здесь задача произвольной коактивации может быть достаточно сложной, чтобы потребовать больших когнитивных усилий и активации коры и, возможно, отдельных нисходящих цепей. Анализ влияния когнитивных усилий во время коактивации или квалифицированных двигательных задач на МВП, вызванные ТМС, может стать интересным исследованием в будущем. Текущие данные, к сожалению, не позволяют определить, было ли увеличение амплитуды МВП во время коактивации по сравнению с состоянием покоя вызвано различиями в концентрации внимания или сложностью двигательных задач.
Дальнейшая характеристика эффекта коактивации между гетеронимными парами мышц (например, камбаловидной и четырехглавой) может дать дополнительное представление об обобщаемости текущих результатов [49]. Систематическая количественная оценка зависимой от коактивации модуляции МВП, вызванных ТМС, в нескольких мышцах нижних конечностей может помочь определить специфический корковый контроль мышц-синергистов и мышц-антагонистов. Кроме того, записи во время динамических задач, таких как линейные и удерживающие сокращения, и во время других постуральных условий, таких как стояние, могут прояснить специфику задачи текущих наблюдений.Дальнейшее изучение зависимой от коактивации модуляции МВП может быть особенно уместным во время ходьбы, когда фазоспецифическая модуляция корково-спинномозговой возбудимости [50] и реципрокное торможение [2] подтверждают участие специфичной для походки моторной коры в настройке коактивации мышц.
С методологической точки зрения повышение кортикомоторной возбудимости агониста путем модулирования активации антагониста может быть полезным при оценке кортикомоторной возбудимости мышц нижних конечностей с помощью ТМС.Например, получение МВП, вызванных ТМС, в паретичной ноге особенно сложно у лиц с повышенным моторным порогом после инсульта. Использование направленной коактивации в качестве условия тестирования может устранить проблемы, связанные со сниженной кортикомоторной возбудимостью в паретичной конечности у людей с инсультом, вероятно, путем включения специализированной схемы коактивации. Наши нынешние результаты, если они наблюдаются у людей после инсульта, предполагают, что поддержание низкого уровня коактивации мышц-антагонистов может потенциально снизить пороги активации и увеличить вероятность возникновения ответов MEP нижних конечностей.Кроме того, учитывая растущую важность выявления наличия или отсутствия ВОС в качестве прогностического инструмента в нейрореабилитации у людей, перенесших инсульт [51], альтернативные условия тестирования, такие как задание на коактивацию, могут быть полезными. В качестве другого примера, в клиническом сценарии, когда у человека, перенесшего инсульт, наблюдается большая слабость в TA, чем в камбаловидной мышце, и получение MEP из TA (целевой мышцы) невозможно, наши результаты показывают, что активация камбаловидной мышцы (антагонист) может облегчить измерение MEP. из парализованной ТА мышцы.Это особенно важно для ТМС-исследований у лиц с парезом мышц тыльного сгибателя или подошвенного сгибателя, где коактивация менее пораженной мышцы-антагониста может в достаточной степени увеличить MEPs агониста паретичной мышцы [48, 52]. Однако МВП, вызванные с использованием модифицированных условий тестирования (таких как коактивация или активация антагониста), могут предлагать различные механистические представления или интерпретации по сравнению с МВП, вызванными в состоянии покоя. Аналогичным образом, во время ТМС нижних конечностей также было показано, что активация мышц контралатеральной конечности усиливает вызванные ТМС MEP от целевых мышц лодыжки [13].Кроме того, более низкая CV амплитуд MEP, продемонстрированная в наших результатах в условиях коактивации, обеспечивает еще одно методологическое преимущество использования условия коактивации в будущих исследованиях.
Наши результаты также показали, что по сравнению с отдыхом состояние активации антагониста приводило к большей амплитуде ВМО как для камбаловидной, так и для TA мышц. Однако это открытие было ограничено нашим наблюдением, что при указании активировать только мышцу-антагонист участники исследования также непреднамеренно увеличивали фоновую активность агониста.Таким образом, во время состояния активации антагониста присутствовала небольшая, но значительная степень активации агониста. Из-за этого методологического ограничения мы не смогли продемонстрировать условия тестирования, которые представляли собой «чистые» условия активации только антагониста. Эта низкоуровневая активация агониста, которая сопровождала команду задания на активацию антагониста, может быть связана с методологическими факторами, например, мы обеспечивали биологическую обратную связь ЭМГ только для мышцы, которую нужно активировать, и мы проверяли фоновую ЭМГ неактивированной мышцы во время анализа данных post hoc и не во время эксперимента.С другой стороны, в отличие от мышц кисти или пальцев, мышцы нижних конечностей могут иметь большую склонность к коактивации, а не к индивидуации. Несмотря на причину, это ограничение повлияло на наши сравнения, включающие условия активации антагониста, а не условия коактивации.
Наше исследование имеет несколько ограничений. Требуемые здесь целевые 10% MVC, вероятно, не приближаются к верхнему пределу прогрессивного рекрутирования двигательных единиц, когда вероятность повышенного стимульного ответа на ТМС резко снижается [53, 54].В то время как в мышцах верхних конечностей наблюдается линейное увеличение ответов МВП до плато около 50% MVC [37], в камбаловидной мышце не наблюдалось снижения ответа МВП в широком диапазоне уровней сокращения [1]. К сожалению, в этих исследованиях не сообщалось об уровнях коактивации антагонистов. Настоящие результаты не позволяют определить, как более высокие уровни добровольной коактивации антагонистов влияют на кривую ответа на стимул. Тем не менее, текущее исследование предоставляет предварительные доказательства того, что корково-спинальная возбудимость мышцы-агониста облегчается даже при маломощных совместных сокращениях мышц голеностопного сустава.Хотя мы протестировали размещение датчика ЭМГ и проверили перекрестные помехи, перекрестные помехи могут повлиять на результаты исследования. Точно так же изменение интенсивности стимуляции ТМС может повлиять на размер вызванных ответов для обеих мышц. Интенсивность стимула ТМС была дозирована до моторного порога ТА в состоянии покоя, а не камбаловидной мышцы. Надпороговая интенсивность была выбрана для достижения согласованных ответов MEP как от TA, так и от камбаловидной мышцы для каждого состояния. Хотя горячие точки ТА и камбаловидной мышцы, вероятно, перекрываются, способность избирательно активировать моторные репрезентации ТА и камбаловидной мышцы затруднена из-за их близкого расположения в межполушарной щели и ограниченной очаговости магнитного поля, индуцированного ТМС [6].Учитывая распространение электрического поля, индуцированного ТМС, на мышцы, не являющиеся мишенями, диссоциирующие эффекты, вызванные генерализованной стимуляцией коры нижних конечностей, затруднены. Наконец, в текущем исследовании использовалась ТМС для оценки изменений общей возбудимости кортикоспинального тракта при модуляции состояния активации мышц-антагонистов; включение показателей возбудимости позвоночника (например, реципрокного торможения TA-Soleus) в будущие исследования позволит получить дополнительное представление о месте и механизме наших результатов.Более того, поскольку активация мышц-агонистов не была эквивалентна в условиях активации и коактивации антагонистов, эти два состояния не сравнивались в нашем исследовании.
5. Выводы
В настоящем исследовании сделан важный шаг к выяснению влияния состояния активации мышц-антагонистов во время ТМС и представлены методологические соображения для будущей оценки ТМС мышц нижних конечностей. Мы продемонстрировали, что активация антагониста, а также коактивация агонист-антагонист увеличивают амплитуду ВМП, вызванную ТМС, при этом уменьшая вариабельность ВМП от испытания к испытанию как для TA, так и для камбаловидной мышцы.Наши результаты указывают на необходимость изучения влияния коактивационной модуляции кортикомоторной возбудимости в патологических когортах, таких как инсульт или травма спинного мозга. Будущие исследования ТМС в сочетании с измерениями спинальных рефлексов могут дополнительно пролить свет на нейронный контроль коактивации.
Доступность данных
Нейрофизиологические данные, использованные для поддержки результатов этого исследования, можно получить у соответствующего автора по запросу.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Благодарности
Эта работа была поддержана грантами Национального института здравоохранения (NIH) K01HD079584 (Kesar), K12HD055931 (Borich), 5R24HD050821-11 (Borich), U10NS086607 (Wolf) и U01NS091951 (Wolf) и Китайской Номер гранта стипендиального совета 201706165058 (Сюй). Мы хотели бы поблагодарить студентов-физиотерапевтов Хейли Шор Макдональд, Лию Энн Алмквист, Сумире Сато, Даниэля Шеффера, Эмили Та и Сару Шарбо за их помощь в сборе данных.
Мышцы-агонисты и мышцы-антагонисты: определение и упражнения
Пары мышц в нашем теле состоят из мышц-агонистов и мышц-антагонистов, которые контролируют диапазон их движения, а также эффективность их функционирования. Но что представляет собой каждая из этих мышц и как они работают вместе, создавая движение?
Подробный отчет OriGym исследует пары мышц-агонистов и антагонистов, как они формируют растяжку и тягу, и как вы можете нацелить их на свои тренировки, чтобы максимизировать свой прогресс.
Мы также рассмотрим советы и рекомендации экспертов о том, как обеспечить максимальное использование потенциала этих мышц, и действия, которые необходимо предпринять, чтобы смягчить любые потенциальные травмы, от которых могут пострадать ваши мышцы-агонисты и мышцы-антагонисты.
Содержание :
В качестве альтернативы, если вы уже знакомы с тем, как работают мышцы, но хотите поднять эти знания на новый уровень, то, возможно, вашим следующим шагом может стать карьера в сфере персональных тренировок.
Престижный диплом персонального тренера от OriGym поможет вам пройти путь от отсутствия предварительного обучения до полностью квалифицированного физиотерапевта всего за четыре недели, с экспертным руководством, доступным 7 дней в неделю, множеством ресурсов и материалов под рукой, бесплатными повторными экзаменами и гарантированное собеседование после выпуска.
Задайте вопрос сегодня или загрузите наш БЕСПЛАТНЫЙ проспект, чтобы узнать больше о том, кто мы такие, что мы предлагаем и как это может быть идеальным для вас.
Загрузите БЕСПЛАТНУЮ 16-недельную домашнюю программу силовых тренировок
Написано профессиональными тренерами S & C
Что такое мышца-агонист?
Сначала давайте рассмотрим определение мышцы-агониста. Это напряженная или «напряженная» мышца во время действия, она действует как основной двигатель во время любого действия.Это абсолютно важный компонент любого действия, и он работает вместе с мышцей-антагонистом, создавая сокращения и растяжения.
В любой паре мышца-агонист сокращается, а мышца-антагонист расслабляется, обеспечивая свободное движение суставов и мышц. Давайте воспользуемся повседневным примером пар мышц-агонистов и мышц-антагонистов, чтобы полностью понять определение мышцы-антагониста и ее аналога — бицепса и трицепса .
Когда мы сгибаем руку (например, сгибая бицепс), бицепс сокращается, делая его мышцей-агонистом, а трицепс расслабляется и, следовательно, мышца-антагонист в этом сценарии.
Затем, когда мы возвращаем руку в естественное положение, наш бицепс расслабляется (мышца-антагонист), а трицепс сокращается и называется мышцей-агонистом.
Эта логика применима ко многим движениям, которые мы выполняем, и является абсолютно неотъемлемой частью выполнения любого упражнения, а также повседневных задач, таких как подъем по лестнице или доставание чего-либо из шкафа.
Мы можем укрепить эти мышцы-агонисты и антагонисты с помощью простых упражнений на растяжку трицепса, а также следуя правильной технике при выполнении упражнений, в которых эти мышцы используются в полной мере.А как насчет определения мышцы-антагониста?
Что такое мышца-антагонист?
Наряду с мышцами-агонистами мышцы-антагонисты функционируют как часть пары, которая работает в тандеме, позволяя суставам и конечностям выполнять более сложные движения. Эти мышцы движутся в направлении, противоположном мышцам-агонистам, и компенсируют силу, которую эти мышцы оказывают, чтобы мы не повреждали наши хрупкие суставы.
Важно отметить, что мышца может двигаться только в одном направлении.Мышца-антагонист в паре растягивается или удлиняется, в то время как мышца-агонист сокращается, что, в свою очередь, создает движение, которое мы ищем.
Это, пожалуй, основное определение мышц-антагонистов и то, что отличает их от мышц-агонистов. Более конкретным примером этого могут быть две мышцы, которые мы находим в наших плечах, которые мы использовали в нашем предыдущем примере с мышцами-агонистами — бицепс и трицепс .
Когда мы расслабляем руку, двуглавая мышца является мышцей-антагонистом, поскольку она расслаблена, а трицепс сокращается, и, следовательно, является мышцей-агонистом.Когда мы напрягаем руки, происходит обратное: бицепс становится мышцей-агонистом, а трицепс расслабляется и становится мышцей-антагонистом.
Мы рассмотрели эти движения более подробно в нашем всестороннем обзоре лучших упражнений на бицепс для массы и силы, но эти принципы применимы даже к самым простым движениям.
Как мышцы-агонисты и мышцы-антагонисты работают вместе?
Теперь, когда мы полностью понимаем определения мышц-агонистов и мышц-антагонистов и какую функцию выполняет каждый из них, важно изучить, как они работают вместе, чтобы создать движения, которые мы ожидаем, и как вы можете максимально использовать их при тренировках. .
По сути, с каждой парой мышц-агонистов и мышц-антагонистов одна мышца будет сокращаться (мышца-агонист ), а другая расслабляться (мышца-антагонист ) во время каждого движения.
Одновременное функционирование этих мышц абсолютно необходимо, поскольку это предотвращает повреждение суставов и костей, а также позволяет мышцам успешно вернуться в исходное положение. Если одна мышца превосходит другую, мы рискуем перенапрячься или не сможем эффективно выполнять действия, к которым стремимся.
Перенапряжение (или перетренированность) часто связано с более интенсивными видами деятельности, такими как бег по пересеченной местности. Ознакомьтесь с дополнительной информацией в нашем всестороннем исследовании бега по пересеченной местности, а также о том, как смягчить последствия перетренированности.
Проще говоря, мышца-антагонист обеспечивает необходимое сопротивление движению, которое совершает мышца-агонист , позволяя использовать нужное количество силы. Это включает в себя простые задачи, о которых мы даже не задумываемся, например, способность стоять прямо или держать руки в естественном положении.
Также важно отметить, что существует два основных типа этих движений — изометрические (действие, при котором не происходит никакого движения, например, отталкивание неподвижной поверхности или объекта) и изотоническое (действие, при котором происходит движение). места, такие как толкание или вытягивание предмета) сокращения.
Итак, например, если вы задаетесь вопросом, «что такое мышца-агонист в отжимании», вам сначала нужно принять во внимание, что это изометрическое сокращение , то есть никакого движения не происходит.Это означает, что меньшее давление оказывается на мышцу-агонист , которая здесь является бицепсом , и, следовательно, больше давления на мышцу-антагонист (в данном случае на трицепс ).
Несмотря на то, что наше тело не нуждается в специальной подготовке к какому-либо из этих различных типов сокращений, очень важно знать о них, чтобы максимально увеличить мышечную массу, которую вы испытываете.
Стань персональным тренером с OriGym!
- Квалифицируйтесь и начните зарабатывать всего за 2 недели
- Учитесь полный рабочий день, неполный рабочий день или онлайн
- REPS & CIMSPA Accredited
Всего за 999 фунтов стерлингов
Узнать большеТеперь давайте сосредоточимся на более практических примерах пар мышц-агонистов и антагонистов, а также на некоторых основных функциях, которые выполняют эти пары мышц, и на том, где вы можете столкнуться с ними в своих упражнениях.
Бицепс и трицепс
Возможно, это один из наиболее легко узнаваемых примеров мышц-антагонистов и агонистов, бицепс и трицепс — две самые большие мышцы плеча. Мы используем эти мышцы каждый божий день, и в основном, не осознавая этого, поскольку они используются во многих повседневных задачах.
Как мы уже говорили в предыдущих разделах о мышцах-агонистах и мышцах-антагонистах, бицепсы и трицепсы функционируют как мышцы-агонисты, так и мышцы-антагонисты.
Напомним, когда мы сгибаем (или сгибаем) руку, бицепс действует как мышца-агонист во время ее сокращения, тогда как трицепс является мышцей-антагонистом , когда он расслабляется. Когда мы разгибаем или расслабляем руку, эти роли меняются местами: бицепс становится мышцей-антагонистом , а трицепс — мышцей-агонистом .
В ваших упражнениях это движение мышц-антагонистов и агонистов наиболее узнаваемо как часть сгибания рук с гантелями или штангой, но оно также имеет место и в других распространенных упражнениях, таких как становая тяга и жим от плеч.
Если вы планируете силовую тренировку рук, мы советуем выполнять одни из лучших упражнений на растяжку и движения бицепса — они помогут расширить диапазон движений, а также облегчат некоторые более сложные движения.
Подколенные сухожилия и квадрицепсы (квадрицепсы)
Две основные мышцы верхней части ног, это один из ярких примеров пар мышц-агонистов и мышц-антагонистов. Подобно паре бицепсов и трицепсов, которые мы только что рассмотрели, их часто упускают из виду, когда речь идет об их вкладе в повседневные задачи.
Квадрицепсы и подколенные сухожилия контролируют разгибание и сжатие коленей, которые являются неотъемлемой частью некоторых из самых простых вещей, которые мы можем делать, таких как ходьба или сидение.
Когда наши ноги расслаблены (например, когда мы находимся в более естественном положении стоя), квадрицепсы функционируют как мышца-агонист , т. это расслаблено.
Однако, когда нога согнута (например, когда вы приседаете или сидите на корточках), эти роли меняются — подколенное сухожилие теперь является мышцей-агонистом , а четырехглавая мышца в этом сценарии — мышцами-антагонистами .
Хотя мы коснулись некоторых из более простых действий, которые требуют этих пар мышц-агонистов и антагонистов, они также являются фундаментальной частью некоторых из самых основных упражнений и одинаково важны для правильной формы и осанки.
Чтобы узнать больше о том, насколько важно правильно тренироваться, ознакомьтесь с полным отчетом OriGym о преимуществах хорошей осанки.
Например, вы будете использовать эту конкретную группу мышц-агонистов и мышц-антагонистов для приседаний и определенных подъемов, особенно тех, где вам нужно слегка согнуть колени, прежде чем согнуть бедра.
Большие ягодичные мышцы и бедра
Хотя мы часто используем ягодицы и бедра, не осознавая этого полностью, они имеют решающее значение для поддержания правильной формы во время упражнений, а также помогают нам сохранять равновесие и стабильность в повседневной жизни.
Эти специфические мышцы-агонисты и антагонисты помогают двигать бедрами и функционируют аналогично другим парам мышц-агонистов и антагонистов, которые мы рассматривали до сих пор.
По сути, когда вы толкаете бедра вперед, вы напрягаете ягодичные мышцы (что делает их мышцами-агонистами) и расслабляете бедра (что делает их мышцами-антагонистами).Когда вы вернетесь в более нейтральное положение, вы расслабите ягодичные мышцы, которые затем являются мышцами-антагонистами, и начнете сокращать ягодичные мышцы, мышцы-агонисты.
Этот принцип часто лежит в основе ходьбы, и как более интенсивные упражнения (например, ходьба или бег в гору) могут значительно улучшить здоровье сердечно-сосудистой системы, а также помочь привести в тонус ключевые области вокруг ягодиц и бедер.
Вы также будете использовать это сокращение и расслабление этих мышц-агонистов и мышц-антагонистов во время становой тяги и рывковых движений, особенно если вы сосредоточены на поднятии более тяжелых весов.
Другие примеры пар мышц-агонистов и мышц-антагонистов
Хотя мы подробно рассмотрели несколько наиболее распространенных движений мышц-антагонистов и мышц-агонистов, есть еще несколько, которые не играют столь важной роли, но это не менее важно, особенно если вы хотите максимизировать прибыль и тренироваться до максимально возможного уровня.
Если вы хотите полностью тренировать пары мышц-агонистов и мышц-антагонистов, вам также необходимо обратить внимание на:
- Дельтовидные мышцы (плечо) и широчайшие мышцы спины (верхняя часть спины)
- Большая грудная мышца (грудь) и трапециевидная мышца (верхняя часть спины)
- Передняя большеберцовая и икроножная (передняя и задняя мышцы голени)
- Брюшной отдел (живот) и мышца, выпрямляющая позвоночник (нижняя часть спины)
В запястье также есть более мелкие и незначительные образцы мышц-антагонистов и мышц-агонистов , шея и лодыжки, которые помогают с гибкостью и могут играть ключевую роль в захвате, движении и устойчивости, особенно во время упражнений.
—
Вам нравится эта статья? Вот еще 3, которые вам могут понравиться:
Упражнения, в которых используются пары мышц-антагонистов и мышц-агонистов
Теперь, когда мы полностью изучили, какие пары мышц-агонистов и мышц-антагонистов, а также примеры мышц-антагонистов и мышц-агонистов, одинаково важно взглянуть на как они могут играть ключевую роль в ваших тренировках.
Понимание различных мышц и того, как каждая из них может оказать огромное влияние, имеет решающее значение для создания устойчивого и эффективного режима.Давайте рассмотрим несколько ключевых примеров.
Сгибание рук на бицепс
Один из ключевых примеров, в котором используются мышцы-агонисты и мышцы-антагонисты, сгибание рук на бицепс является основным элементом многих упражнений, и не зря. Его легкая адаптируемость в сочетании с широким спектром потенциального оборудования для этого упражнения делает его идеальным вариантом для тех, кто хочет разнообразить свои тренировки.
Как мы упоминали ранее, сгибание рук на бицепс состоит из двух основных движений — подъема и последующего расслабления.Во время подъема бицепс становится мышцей-агонистом , напрягаясь и сокращаясь, а трицепс становится мышцей-антагонистом , расслабляясь при подъеме.
Затем, когда вы опускаете руку, все меняется на противоположное: бицепс становится мышцей-агонистом , сокращаясь при опускании веса, а трицепс становится антагонистом , расслабляясь при опускании веса.
Добавляя вес к смеси, вы не только увеличиваете нагрузку на напряженную мышцу ( агонист ), но также увеличиваете количество мышц, которые антагонисту необходимо растянуть, чтобы компенсировать нагрузку на агонист .
В связи с этим мы настоятельно рекомендуем медленно увеличивать вес, особенно потому, что это может иметь огромное влияние и потенциально вызвать проблемы или травмы, если вы увеличите его слишком быстро.
Полное исследование OriGym сгибания рук на бицепс с кабелем подробно исследует это разнообразное упражнение, а также предоставляет примеры того, как вы можете настроить его в соответствии со своими потребностями.
Становая тяга
Еще одна основная программа упражнений для тех, кто хочет нарастить мышечную массу и усердно тренироваться. Становая тяга использует несколько различных пар мышц-агонистов и мышц-антагонистов для создания более сложных движений.
В становой тяге вам нужно, чтобы ваши руки были в выпрямленном положении, что означает, что ваш бицепс находится в расслабленном положении, мышца-антагонист , а трицепс функционирует как мышца-агонист .
Становая тяга начинается со слегка согнутых коленей (чтобы избежать «блокировки» коленей), что требует использования квадрицепсов и подколенных сухожилий. В исходном положении ваши подколенные сухожилия будут мышцами-агонистами , сокращенными и напряженными, а ваши квадрицепсы займут место мышц-антагонистов , то есть они будут более расслабленными.
По мере того, как вы переходите в положение стоя, эти мышцы меняются ролями: квадрицепсы становятся сокращенными, мышцами-агонистами , а подколенные сухожилия находятся в более расслабленном положении, мышцами-антагонистами .
Узнайте больше о становой тяге в нашем полном руководстве и узнайте о правильной осанке, а также о том, как добиться максимальной эффективности.
В становой тяге также используются другие пары мышц-агонистов и антагонистов, и они имеют первостепенное значение при выборе правильной техники становой тяги, а также для обеспечения безопасного выполнения этих упражнений.
Вам также нужно будет задействовать сгибатели бедра, выталкивая их наружу, чтобы добиться дополнительной подъемной силы на перекладине. Это включает в себя выталкивание бедер (это означает, что они находятся в положении мышц-агонистов ) и расслабление ягодичных мышц (здесь они будут мышцами-антагонистами ), прежде чем вернуться в более естественное положение.
Наконец, ваши запястья, хотя они и являются второстепенными мышцами-агонистами и антагонистами, абсолютно необходимы для надежного удержания грифа. Обязательно изучите различные варианты хвата, чтобы найти вариант, который подходит именно вам, тем более, что это может сильно повлиять на эффективность ваших упражнений.
Сердечно-сосудистые упражнения
Физическая активность является неотъемлемой частью здорового образа жизни, и один из самых простых способов сделать это — воспользоваться преимуществами кардио- и аэробных упражнений. Это также один из лучших способов одновременной активации ключевых пар мышц-агонистов и мышц-антагонистов, что позволяет вам работать с этими группами мышц без необходимости нацеливаться на них конкретно.
Как и следовало ожидать, когда мы идем (или бежим), основными мышцами, которые мы будем использовать, являются мышцы ног, в основном квадрицепсы, подколенные сухожилия, икры и ягодицы.Однако мы также будем двигать руками и плечами, что требует некоторых движений мышц-антагонистов и агонистов.
Давайте сначала сосредоточимся на ногах. Когда мы сгибаем колено, чтобы шагнуть вперед, мы в первую очередь задействуем наши подколенные сухожилия (сокращенные, или 90 569, агонисты 90 570) и четырехглавые мышцы (расслабленные, или 90 569, антагонисты 90 570).
Затем они меняются ролями, когда мы ставим ногу обратно, при этом квадрицепсы теперь сокращаются ( мышца-агонист ), а подколенные сухожилия расслабляются ( мышца-антагонист ).Это представляет собой наш основной шаг и происходит без нашего участия, особенно на беговой дорожке.
Узнайте больше о преимуществах бега на беговой дорожке из подробного отчета OriGym.
Наши плечевые мышцы также являются движущей силой, когда мы выполняем кардиоупражнения, и помогают толкать тело вперед. По мере того, как мы шагаем вперед, мы также двигаем вперед и плечи, а это означает, что большая грудная мышца (грудная мышца) берет на себя роль 90 569 агониста 90 570, а 90 569 антагониста 90 570 – трапециевидной (верхней части спины).
Как мы видели с предыдущими парами мышц-агонистов и мышц-антагонистов, эти роли меняются местами, когда мы возвращаемся в естественное положение: трапециевидная мышца теперь является мышцей-агонистом , а большая грудная мышца — мышцей-антагонистом .
В наших лодыжках также наблюдается незначительная активность, когда речь идет о движении мышц-антагонистов и агонистов. Это в основном для обеспечения хорошего баланса, сохранения осанки и обеспечения того, чтобы мы могли постоянно двигаться в одном и том же темпе.
Часто задаваемые вопросы
Хотя мы уже коснулись всех ключевых аспектов пар мышц-агонистов и мышц-антагонистов, а также определений мышц-агонистов и мышц-антагонистов, по-прежнему важно развеять любые заблуждения и ответить на некоторые вопросы. вопросы, которые могут возникнуть.
Почему мышцы работают парами?
В этой статье мы в основном сосредоточились на ключевых примерах мышц-агонистов и мышц-антагонистов, но не менее важно исследовать первопричину того, почему мышцы должны работать в паре, и что может произойти, если одна мышца ослабнет или не работает. не работает должным образом.
Как мы упоминали ранее, каждая пара мышц состоит из мышц-агонистов и мышц-антагонистов , которые чередуются по мере того, как мы совершаем движения и действия. Мышцы обычно располагаются парами по одной очень конкретной, очень важной причине.
Когда одна мышца сокращается (это мышца-агонист ) и прилагает необходимую силу для завершения действия, которое вы хотите выполнить, противоположная мышца (мышца-антагонист ) создает силу в противоположном направлении, чтобы сбалансируйте усилия, которые вы прилагаете.
Без этой противодействующей силы вы серьезно рискуете повредить мышцы, суставы и общее состояние скелета, особенно при таких движениях, как сгибание рук на бицепс или становая тяга, которые часто могут сочетаться со значительным весом в дополнение к мышечной нагрузке.
Несмотря на многочисленные преимущества становой тяги, жизненно важно понимать основы и то, как необходимые движения мышц-антагонистов и агонистов формируют основу всех упражнений, которые вы выполняете.
Этот принцип применим ко всем парам мышц-агонистов и мышц-антагонистов по всему телу, включая меньшие и менее заметные пары, которые мы находим в ключевых суставах (например, в наших запястьях и лодыжках).
Стань персональным тренером с OriGym!
- Квалифицируйтесь и начните зарабатывать всего за 2 недели
- Учитесь полный рабочий день, неполный рабочий день или онлайн
- REPS & CIMSPA Accredited
Всего за 999 фунтов стерлингов
Узнать больше
Это вполне понятный вопрос, тем более, что агонист мышечных движений в основном отвечает за приложение силы, необходимой для выполнения действия или упражнения.
Хотя это верно для всех примеров мышц-агонистов, они просто не будут функционировать правильно и эффективно без своего аналога, мышцы-антагониста . Давайте посмотрим на пример этого.
Когда мы сгибаем колено, чтобы сделать шаг вперед, независимо от темпа, в котором мы движемся, мы будем использовать две основные мышцы — подколенное сухожилие и четырехглавую мышцу.
В этом сценарии подколенное сухожилие является мышцей-агонистом (в том смысле, что она сокращается и прилагает необходимую силу для движения колена), а четырехглавая мышца является мышцей-антагонистом (они расслаблены и обеспечивают противовес сила, которую прикладывает мышца-агонист).
Когда мы снова разгибаем ногу, эти роли меняются: мышца-агонист теперь является четырехглавой мышцей, а мышца-антагонист — подколенным сухожилием. Каждое мышечное движение требует противодействующей силы, чтобы гарантировать, что мы не перенапрягаемся и что мы можем вернуться в более естественное положение после того, как закончим движение мышц-агонистов.
Мышцы-антагонисты также абсолютно необходимы уже по одной этой причине — они позволяют нам вернуть наше тело в более комфортное, естественное состояние.Однако они также помогают защитить кости от повреждений, а наши хрупкие сухожилия, суставы и связки — от разрывов или болезненных травм.
Полезно ли тренировать мышцы-антагонисты?
Когда мы стремимся достичь вершины своей личной физической формы, естественно, что мы хотим использовать возможности для активации и укрепления групп мышц, о которых мы, возможно, даже не думали. То же самое относится к определению и тонусу наших мышц-антагонистов.
Тем не менее, может быть трудно понять, как именно мы воздействуем на эти области, особенно потому, что они в основном используются для расслабления наших мышц-агонистов или тех, которые принимают на себя основную нагрузку.
Самый простой ответ на этот вопрос заключается в том, что вы, вероятно, уже тренируете мышцы-антагонисты, не осознавая этого, тем более что они часто являются частью каждого упражнения, особенно тех, о которых мы упоминали ранее.
Например, несмотря на то, что мышца-агонист в положении приседания — это подколенное сухожилие, вы все равно будете активировать и напрягать четырехглавую мышцу, что позволит вам укрепить свою мышцу-антагонист без сознательного воздействия на эту область.
Вариации приседаний могут даже обеспечить более последовательный способ нацеливания на эти забытые области — узнайте больше в нашем полном руководстве по приседаниям и их преимуществам.
Важно отметить, что правильная разминка и растяжка жизненно важны для обеспечения безопасной тренировки, особенно когда вы стремитесь проработать мышцы, на которых вы, возможно, раньше не обращали внимания или которые слабее. чем другие области вашего тела.
Растяжка ног (особенно с использованием растяжки подколенного сухожилия) и рук перед сердечно-сосудистыми упражнениями или выполнение ключевых растяжек бицепса и трицепса перед поднятием тяжестей может не только помочь вам максимизировать ваши результаты, но также смягчить некоторые из наиболее распространенных травмы, которые вы можете получить.
Перед уходом!
Целью этой статьи было дать вам полное определение мышцы-антагониста, а также того, как мышцы-агонисты и мышцы-антагонисты работают в парах, создавая ожидаемое нами движение.
Искали ли вы вдохновение для своих тренировок или искали примеры мышц-агонистов и антагонистов, чтобы улучшить свою тренировку, наше руководство предоставит вам все, что вам нужно знать, и как вы можете использовать эти уникальные пары мышц.
Но если вы уже знакомы с тем, как использовать пары мышц-агонистов и мышц-антагонистов, и ищете способ поделиться этим опытом, то, возможно, карьера в фитнесе может стать вашим призванием.
Диплом индивидуального обучения OriGym, аккредитованный CIMSPA, предлагает полный пакет услуг: экспертное руководство доступно 7 дней в неделю, специально созданная платформа онлайн-обучения, неограниченное руководство по профориентации и гарантированное собеседование после окончания обучения.
Заинтересованы? Нажмите здесь, чтобы загрузить наш БЕСПЛАТНЫЙ всеобъемлющий проспект и узнать больше о том, что мы предлагаем, и о том, как это может быть идеальным для вас.
Загрузите БЕСПЛАТНУЮ 16-недельную домашнюю программу силовых тренировок
Написано профессиональными тренерами S & C
11.1 Взаимодействие скелетных мышц, их расположение пучков и их рычажные системы — анатомия и физиология
Цели обучения
К концу этого раздела вы сможете:
- Сравните и сопоставьте мышцы-агонисты и мышцы-антагонисты
- Опишите, как пучки расположены в скелетной мышце
- Объясните основные события сокращения скелетных мышц внутри мышцы при создании силы
Для движения скелета напряжение, создаваемое сокращением волокон в большинстве скелетных мышц, передается на сухожилия.Сухожилия представляют собой прочные полосы плотной, правильной соединительной ткани, которые соединяют мышцы с костями. Благодаря связи с костями эта мышечная ткань называется скелетной мышцей.
Взаимодействие скелетных мышц в организме
Чтобы натянуть кость, то есть изменить угол в ее синовиальном суставе, который существенно перемещает скелет, скелетная мышца также должна быть прикреплена к неподвижной части скелета. Подвижный конец мышцы, прикрепляющийся к вытягиваемой кости, называется местом прикрепления мышцы, а конец мышцы, прикрепленный к неподвижной (стабилизированной) кости, называется началом.Во время сгибания предплечья — сгибания локтя — плечелучевая мышца помогает плечевой мышце.
Несмотря на то, что в действие может быть вовлечено несколько мышц, основная задействованная мышца называется первичным двигателем или агонистом. Чтобы поднять чашку, мышца, называемая двуглавой мышцей плеча, на самом деле является основным двигателем; однако, поскольку ему может помогать брахиалис, брахиалис называется синергистом в этом действии (рис. 11.2). Синергист также может быть фиксатором, стабилизирующим кость, являющуюся придатком источника первичного двигателя.
Рис. 11.2 Первичные двигатели и синергисты Двуглавая мышца плеча сгибает предплечье. Плечелучевая мышца в предплечье и плечевая мышца, расположенная глубоко в бицепсе в верхней части руки, являются синергистами, помогающими этому движению.
Мышца с действием, противоположным первичному двигателю, называется антагонистом. Антагонисты играют две важные роли в функционировании мышц: (1) они поддерживают положение тела или конечностей, например, вытягивают руку или стоят прямо; и (2) они контролируют быстрое движение, как в бою с тенью, без нанесения удара или возможности контролировать движение конечности.
Например, чтобы разогнуть колено, активируется группа из четырех мышц, называемых четырехглавой мышцей бедра, в передней части бедра (их можно назвать агонистами разгибания колена). Однако, чтобы согнуть коленный сустав, активируется противоположный или антагонистический набор мышц, называемый подколенными сухожилиями.
Как видите, эти термины также могут быть перевернуты для противодействующего действия. Если рассматривать первое действие как сгибание колена, подколенные сухожилия будут называться агонистами, а четырехглавые мышцы бедра — антагонистами.См. Таблицу 11.1 для списка некоторых агонистов и антагонистов.
Пары скелетных мышц-агонистов и антагонистов
| Агонисты | Антагонист | Механизм |
|---|---|---|
| Двуглавая мышца плеча: в переднем отделе плеча | Трехглавая мышца плеча: в заднем отделе плеча | Двуглавая мышца плеча сгибает предплечье, тогда как трехглавая мышца плеча разгибает его. |
| Подколенные сухожилия: группа из трех мышц в задней части бедра | Четырехглавая мышца бедра: группа из четырех мышц в передней части бедра | Подколенные сухожилия сгибают ногу, а четырехглавая мышца бедра разгибает ее. |
| Поверхностный сгибатель пальцев и глубокий сгибатель пальцев: в переднем отделе предплечья | Разгибатель пальцев: в заднем отделе предплечья | Поверхностный сгибатель пальцев и глубокий сгибатель пальцев сгибают пальцы и кисть в запястье, тогда как разгибатель пальцев разгибает пальцы и кисть в запястье. |
Таблица 11.1
Существуют также скелетные мышцы, которые не тянутся к скелету при движениях.Например, есть мышцы, отвечающие за выражение лица. Прикрепление и начало лицевых мышц находятся в коже, так что определенные отдельные мышцы сокращаются, чтобы вызвать улыбку или хмурый взгляд, произнесение звуков или слов и поднятие бровей. На языке также есть скелетные мышцы, а также наружный мочевой и анальный сфинктеры, которые обеспечивают произвольную регуляцию мочеиспускания и дефекации соответственно. Кроме того, диафрагма сокращается и расслабляется, чтобы изменить объем плевральных полостей, но не перемещает скелет для этого.
Связь на каждый день
Упражнения и растяжка
При выполнении упражнений важно сначала разогреть мышцы. Растяжка растягивает мышечные волокна, что также приводит к увеличению притока крови к работающим мышцам. Без надлежащей разминки вы можете либо повредить некоторые мышечные волокна, либо потянуть сухожилие. Растянутое сухожилие, независимо от его местоположения, вызывает боль, отек и снижение функции; если это от умеренной до тяжелой, травма может обездвижить вас на длительный период.
Вспомните обсуждение мышц, пересекающих суставы для создания движения. Большинство суставов, которые вы используете во время упражнений, являются синовиальными суставами, у которых есть синовиальная жидкость в суставном пространстве между двумя костями. Упражнения и растяжка также могут оказывать благотворное влияние на синовиальные суставы. Синовиальная жидкость представляет собой тонкую, но вязкую пленку консистенции яичного белка. Когда вы впервые встаете и начинаете двигаться, ваши суставы скованны по ряду причин. После надлежащей растяжки и разогрева синовиальная жидкость может стать менее вязкой, что способствует улучшению функции сустава.
Модели организации пучков
Скелетная мышца заключена в соединительнотканный каркас на трех уровнях. Каждое мышечное волокно (клетка) покрыто эндомизием, а вся мышца покрыта эпимизием. Когда группа мышечных волокон «связана» как единое целое внутри целой мышцы дополнительным покрытием из соединительной ткани, называемым перимизием, такая связанная группа мышечных волокон называется пучком. Расположение пучков перимизией коррелирует с силой, создаваемой мышцей; это также влияет на диапазон движения мышцы.По характеру расположения пучков скелетные мышцы можно классифицировать несколькими способами. Ниже приведены наиболее распространенные схемы пучков.
Параллельные мышцы имеют пучки, расположенные в том же направлении, что и длинная ось мышцы (рис. 11.3). Большинство скелетных мышц в организме имеют такой тип организации. Некоторые параллельные мышцы представляют собой плоские листы, которые расширяются на концах, образуя широкие прикрепления. Другие параллельные мышцы округлые с сухожилиями на одном или обоих концах.Мышцы, которые кажутся пухлыми, имеют большую массу ткани, расположенную в середине мышцы, между местом прикрепления и началом, которая известна как центральное тело. Более распространенное название этой мышцы – живот. Когда мышца сокращается, сократительные волокна укорачивают ее до еще большей выпуклости. Например, растяните, а затем согните двуглавую мышцу плеча; большой, средний отдел — это живот (рис. 11.4). Когда параллельная мышца имеет центральное большое брюшко, имеющее веретенообразную форму, что означает, что оно сужается по мере того, как оно простирается к своему началу и месту прикрепления, его иногда называют веретенообразным.
Рис. 11.3 Форма мышц и расположение волокон Скелетные мышцы тела обычно имеют семь различных форм.
Рис. 11.4 Сокращение двуглавой мышцы плеча Большая масса в центре мышцы называется животом. Сухожилия выходят из обоих концов живота и соединяют мышцы с костями, позволяя скелету двигаться. Сухожилия бицепса соединяются с плечом и предплечьем. (кредит: Виктория Гарсия)
Круговые мышцы также называют сфинктерами (см. рисунок 11.3). Когда они расслабляются, концентрически расположенные пучки мышечных волокон сфинктеров увеличивают размер отверстия, а когда они сокращаются, размер отверстия сжимается до точки закрытия. Круговая мышца рта представляет собой круговую мышцу, огибающую рот. Когда он сокращается, ротовое отверстие становится меньше, как при сморщивании губ для свиста. Другим примером является круговая мышца глаза, одна из которых окружает каждый глаз. Рассмотрим, например, названия двух круговых мышц (orbicularis oris и oribicularis oculi), где часть первого названия обеих мышц одинакова.Первая часть orbicularis, orb (orb = «круговой»), является ссылкой на круглую или круглую структуру; это также может навести на мысль об орбите, такой как путь Луны вокруг Земли. Слово oris (oris = «оральный») относится к ротовой полости или рту. Слово oculi (окуляр = «глаз») относится к глазу.
По всему телу есть и другие мышцы, названные по их форме или расположению. Дельтовидная мышца представляет собой крупную мышцу треугольной формы, покрывающую плечо. Она названа так потому, что греческая буква дельта выглядит как треугольник.Прямая мышца живота (rector = «прямая») — это прямая мышца передней стенки живота, а прямая мышца бедра — прямая мышца передней поверхности бедра.
Когда мышца имеет обширное расширение на значительной площади, но затем пучки приходят к одной общей точке прикрепления, мышца называется конвергентной. Точкой прикрепления сходящейся мышцы может быть сухожилие, апоневроз (плоское, широкое сухожилие) или шов (очень тонкое сухожилие).Большая грудная мышца, большая грудная мышца, является примером сходящейся мышцы, потому что она сходится на большом бугорке плечевой кости через сухожилие. Височная мышца черепа — другая.
Перистые мышцы (penna = «перья») переходят в сухожилие, которое проходит через центральную область мышцы по всей ее длине, что-то вроде пера пера с мышцей, расположенной подобно перьям. Из-за такой конструкции мышечные волокна перистых мышц могут тянуться только под углом, и в результате сокращающиеся перистые мышцы не перемещают свои сухожилия очень далеко.Однако, поскольку перистая мышца обычно может удерживать внутри себя больше мышечных волокон, она может создавать относительно большее напряжение для своего размера. Существует три подтипа перистых мышц.
В одноперистой мышце пучки расположены на одной стороне сухожилия. Разгибатель пальцев предплечья является примером одноперистой мышцы. Двуперистая мышца имеет пучки с обеих сторон сухожилия. В некоторых перистых мышцах мышечные волокна обвивают сухожилие, иногда образуя при этом отдельные пучки.Такое расположение называется мультипеннатным. Типичным примером является дельтовидная мышца плеча, которая покрывает плечо, но имеет единственное сухожилие, которое прикрепляется к дельтовидной бугристости плечевой кости.
Из-за пучков часть многоперистой мышцы, такая как дельтовидная, может стимулироваться нервной системой для изменения направления тяги. Например, когда дельтовидная мышца сокращается, рука отводится (отходит от средней линии в сагиттальной плоскости), но когда стимулируется только передний пучок, рука отводится и сгибается (двигается вперед в плечевом суставе).
Рычажная система взаимодействия мышц и костей
Скелетные мышцы не работают сами по себе. Мышцы располагаются парами в зависимости от их функций. Для мышц, прикрепленных к костям скелета, соединение определяет силу, скорость и диапазон движения. Эти характеристики зависят друг от друга и могут объяснить общую организацию мышечной и скелетной систем.
Скелет и мышцы действуют вместе, чтобы двигать тело. Вы когда-нибудь выбивали гвоздь из дерева тыльной стороной молотка? Рукоятка действует как рычаг, а головка молотка действует как точка опоры, фиксированная точка, к которой прикладывается сила, когда вы оттягиваете или нажимаете на рукоятку.Усилие, прилагаемое к этой системе, представляет собой вытягивание или нажатие на ручку для удаления гвоздя, который является нагрузкой, или «сопротивлением» движению ручки в системе. Наша опорно-двигательная система работает аналогичным образом: кости являются жесткими рычагами, а суставные окончания костей, заключенные в синовиальные суставы, действуют как точки опоры. Нагрузкой может быть поднимаемый объект или любое сопротивление движению (ваша голова представляет собой груз, когда вы ее поднимаете), а усилие или приложенная сила исходит от сокращения скелетных мышц.
Уровень 3 (70) Физические упражнения и фитнес: плечевой сустав
Плечевой сустав представляет собой шаровидный сустав, при этом головка плечевой кости входит в очень неглубокую полость на лопатке, называемую суставной ямкой. Плечевой сустав является наиболее подвижным суставом в организме, но также и одним из самых нестабильных, поскольку неглубокая впадина дает слабую поддержку головке плечевой кости. Стабильность должна обеспечиваться связками и мышцами, и в общей сложности девять мышц пересекают каждый плечевой сустав, чтобы прикрепиться к плечевой кости.Все эти мышцы берут начало на плечевом поясе.
Нажмите, чтобы увеличить
Только три из девяти мышц действуют как первичные двигатели движений рук – большая грудная мышца , широчайшая мышца спины и дельтовидная мышца . Остальные шесть используются как синергисты или фиксаторы.
Четыре из девяти мышц, надостная мышца , подостная мышца , малая круглая мышца и подлопаточная мышца , вместе известны как вращательная манжета плеча мышц.Их основная функция заключается в укреплении капсулы плечевого сустава для предотвращения вывиха плечевой кости. Однако они также действуют как синергисты при угловых или вращательных движениях руки.
Последние две мышцы, большая круглая мышца и клювовидно-плечевая мышца , пересекают плечевой сустав, но не укрепляют его.
Возможные движения в плечевом суставе включают сгибание, разгибание, горизонтальное сгибание, горизонтальное разгибание, отведение, приведение, медиальное вращение, латеральное вращение и круговое движение.
Мышцы, берущие начало в передней (передней) части плечевого сустава, имеют тенденцию сгибать руку (большая грудная, клювовидно-плечевая и передние волокна дельтовидной мышцы). Большая грудная мышца — это мышца, которая действует как первичный двигатель при сгибании плеча. Двуглавая мышца плеча помогает этому движению.
Мышцы, берущие начало в задней части плечевого сустава, разгибают руку.