
Метаболизм | справочник Пестициды.ru
Cхема метаболических процессовCхема метаболических процессов
Процессы метаболизма
Метаболизм включает две группы жизненно важных процессов – катаболизм (энергетический обмен) и анаболизм (биосинтез, или пластический обмен).[3]
- Катаболизм – это совокупность процессов расщепления питательных веществ, которые происходят в основном за счет реакций окисления. В результате выделяется энергия. Основными формами катаболизма у микроорганизмов являются брожение и дыхание. При брожении происходит неполный распад сложных органических веществ с выделением небольшого количества энергии и накоплении богатых энергией конечных продуктов. При дыхании (аэробном) обычно осуществляется полное окисление соединений с выходом большого количества энергии.

- Анаболизм объединяет процессы синтеза молекул из более простых веществ, которые присутствуют в окружающей среде. Реакции анаболизма связаны с потреблением свободной энергии, которая вырабатывается в процессах дыхания, брожения. Для протекания пластического обмена необходимо поступление в организм питательных веществ, на основе которых при участии выделенной в ходе катаболизма энергии обновляются структурные компоненты клеток, происходит рост и развитие.[3]

Катаболизм и анаболизм протекают параллельно, многие их реакции и промежуточные продукты являются общими. Тем не менее, на протяжении разных периодов существования интенсивность пластического и энергетического обмена неодинакова. Так, у насекомых в период размножения, линьки, во время ранних фаз развития (яйцо, личинка) синтетические процессы преобладают над процессами распада. В тоже время, определенные дегенеративные изменения в организме (старение, заболевания) способны приводить к преобладанию интенсивности катаболизма над анаболизмом, что порой угрожает гибелью живому объекту.
Превращение сульфооксида в сульфон
Использовано изображение:[2]
Метаболизм пестицидов
Метаболизм пестицидов – превращения пестицидов под влиянием продуктов жизнедеятельности различных живых организмов – бактерий, грибов, высших растений и животных.[4]
В результате биотрансформации токсичных веществ в большинстве случаев образуются менее токсичные продукты (метаболиты), более растворимые и легко выводимые из организма. В некоторых случаях токсичность метаболитов оказывается выше, чем попавших в организм веществ. Обмен промышленных ядов возможен за счет реакций окисления, восстановления, гидролитического расщепления, метилирования, ацилирования и др.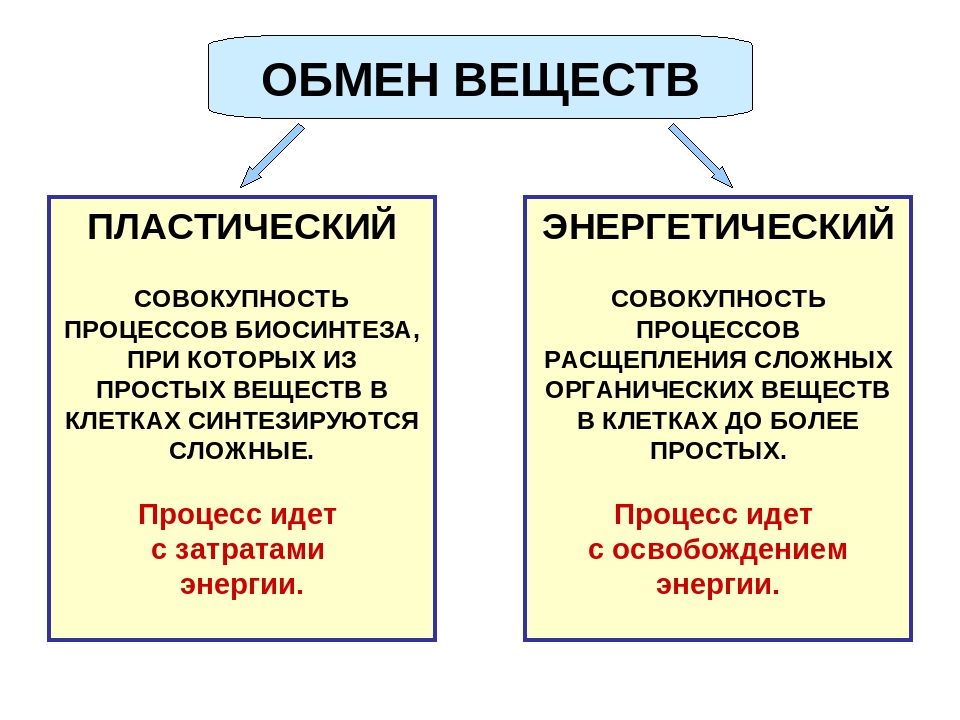
В метаболизме пестицидов большое значение имеют реакции окисления атома серы в молекулах некоторых веществ, что характерно, например, для инсектицидов из группы производных карбаминовой и фосфорной кислот. Окисление серы у этих соединений происходит независимо от структуры остальной части молекулы, при этом вначале образуется соответствующий сульфооксид, а затем сульфон: (фото) Продукты окисления не отличаются по токсичности от исходного вещества, но они значительно более стойки к гидролизу.
Окисление тионофосфатовОкисление тионофосфатов
А — тионофосфат, В – фосфат, 1 и 2- свободные радикалы, 3 — кислотный остаток
Использовано изображение:[2]
Реакции метаболизма, происходящие в растениях, обусловливают длительное инсектицидное действие для ряда эфиров фосфорных кислот с тиоэфирным радикалом. Окисление тионофосфатов в различных организмах рассматривается как активирующая ступень в процессах метаболизма этих веществ.[2](фото)
Окисление тионофосфатов в различных организмах рассматривается как активирующая ступень в процессах метаболизма этих веществ.[2](фото)
Токсичность продукта реакции для млекопитающих и насекомых увеличивается в десятки и сотни раз по сравнению с исходным веществом. Однако эти токсичные метаболиты легко гидролизуются и поэтому сохраняются в биологических средах непродолжительное время.
Близкие статьи
Ссылки:
Все статьи о токсикологии в разделе: Основы токсикологии
Статья составлена с использованием следующих материалов:
Литературные источники:
1.Голдовская Л.Ф. Химия окружающей среды. М.: Мир; БИНОМ. Лаборатория знаний, 2007. – 295 с
2.Груздев Г.С. Химическая защита растений. Под редакцией Г.С. Груздева — 3-е изд., перераб. и доп. — М.: Агропромиздат, 1987. — 415 с.: ил.
3.
Липунов И.Н., Первова И.Г.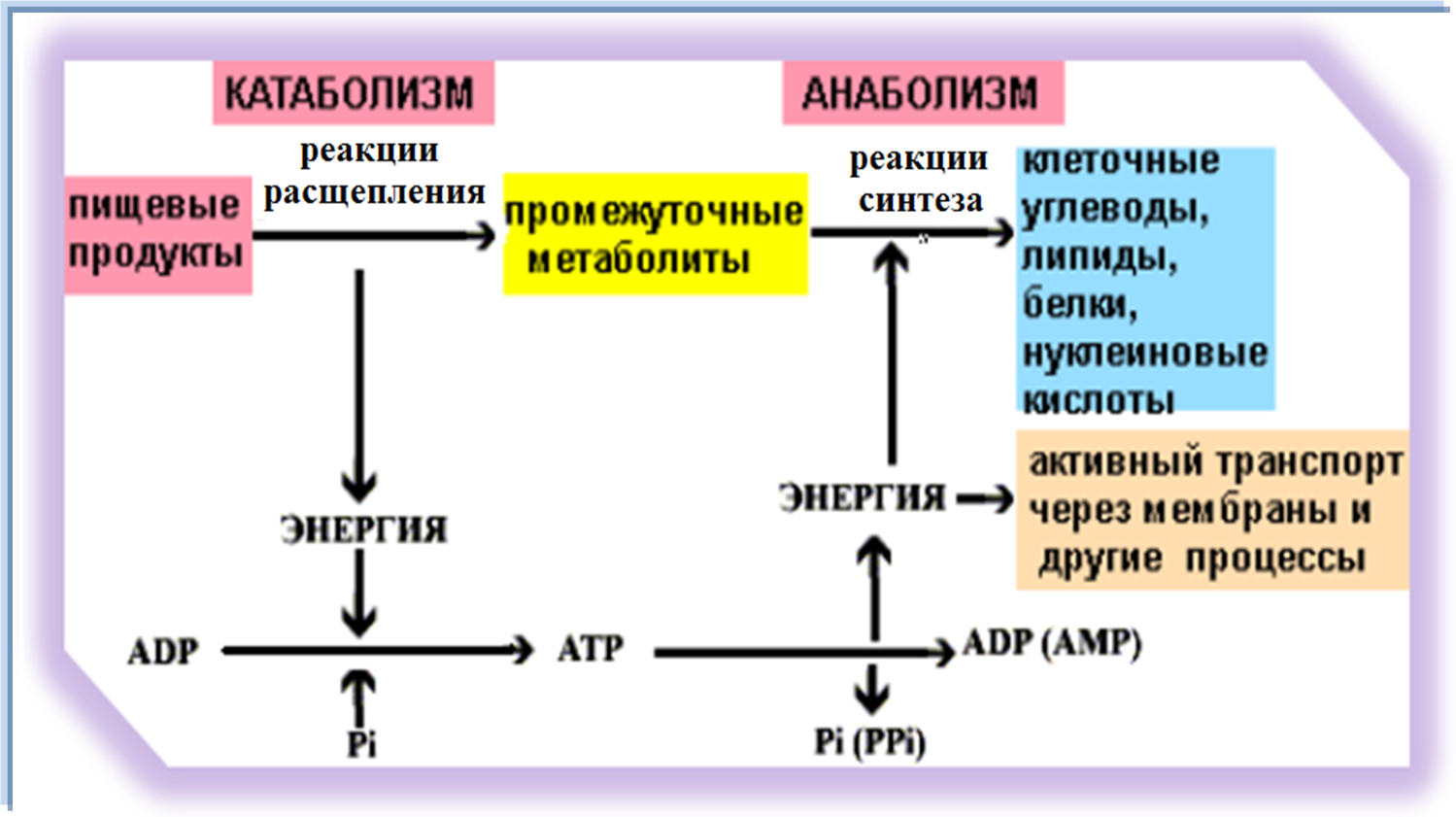 Основы микробиологии и биотехнологии: курс лекций. – Екатеринбург: Урал. гос. лесотехн. Университет, 2008. – 231 с
Основы микробиологии и биотехнологии: курс лекций. – Екатеринбург: Урал. гос. лесотехн. Университет, 2008. – 231 с
Мельников Н.Н., Новожилов К.В., Белан С.Р., Пылова Т.Н. Справочник по пестицидам — М.: Химия, 1985. — 352 с.
Свернуть Список всех источниковМетаболическая кардиомиопатия | Руководство по кардиологии
(Е.Г. Несукай)Определение
Метаболическая кардиомиопатия (ранее ее определяли как дистрофию миокарда, миокардиодистрофию) — невоспалительное поражение миокарда различной этиологии, в основе которого лежит нарушение обмена веществ, процесса образования энергии и/или нарушение ее превращения в механическую работу, приводящее к дистрофии миокарда и недостаточности сократительной и других функций сердца.
Этиология
Метаболическая кардиомиопатия развивается в результате воздействия патогенных факторов при различных заболеваниях и состояниях (схема 8.1).
Среди физических факторов могут рассматриваться радиация, вибрация, перегревание, переохлаждение, гиперинсоляция.
К химическим факторам относятся лекарственные средства, токсическое воздействие бытовых и промышленных ядов.
Патогенез
В возникновении и развитии метаболических поражений миокарда при разных заболеваниях существенное значение имеет нарушение иннервации, транспорта и утилизации энергии в кардиомиоцитах, то есть их энергообеспечение.
Напряжение регулирующих систем, функции миокарда и метаболических процессов в кардиомиоцитах ограничивает резервные возможности сердца. Длительная гиперфункция сама по себе, а особенно в неблагоприятных условиях на фоне основного заболевания, может привести к возникновению энергетического дефицита и нарушению приспособительных изменений в миокарде.
Механизмы снижения продукции энергии в поврежденном сердце включают снижение плотности капилляров, увеличение межкапиллярного расстояния, а также больший диаметр гипертрофированных кардиомиоцитов, что ухудшает диффузию кислорода и обусловливает возникновение гипоксии миокарда. Один из механизмов связан также с нарушением функции митохондрий, которое вызвано редуцированным синтезом окислительных энзимов вследствие нарушения пролиферативной реакции, которая частично опосредована экспрессией рецепторов PPARα, играющих ключевую роль в биогенезе митохондрий.
В прогрессировании метаболической кардиомиопатии ведущую роль играет усиление реакций свободнорадикального перекисного окисления липидов клеточных мембран. Повреждая мембраны, гидроперекиси и свободные радикалы снижают активность липидозависимых ферментативных реакций (к которым относятся основные жизненно важные ферменты ионного транспорта и дыхательной цепи митохондрий), изменяют мембранорецепторные системы клетки с развитием медиаторного дисбаланса, активируют протеолитические и лизосомальные ферменты.
Патологическая анатомия
Метаболические поражения миокарда охватывают все стадии нарушения обмена сердечной мышцы — от функциональных расстройств до грубых структурных изменений. Морфологические изменения происходят внутри клеток миокарда и не сопровождаются увеличением их количества. Наиболее чувствительны к патогенным воздействиям митохондрии и эндоплазматическая сеть. Для дегенеративных изменений миокарда характерно мозаичное нарушение структуры кардиомиоцитов: в одной и той же клетке среди набухших митохондрий с частично или полностью разрушенными внутренними перегородками могут быть митохондрии с нормальным строением.
Как правило, устранение патогенной причины приводит к постепенной нормализации ультраструктур кардиомиоцита, что обусловлено внутриклеточными регенераторными процессами. Поврежденные миофибриллы восстанавливаются в результате активной деятельности рибосом: постепенно устраняется внутриклеточный отек, появляются зерна гликогена, уменьшается количество жировых включений. При длительном и интенсивном воздействии повреждающих факторов на миокард дистрофические изменения могут приводить к глубоким морфологическим изменениям, заканчивающимся развитием миокардиофиброза.
При длительном и интенсивном воздействии повреждающих факторов на миокард дистрофические изменения могут приводить к глубоким морфологическим изменениям, заканчивающимся развитием миокардиофиброза.
Гибель части миокарда восполняется увеличением массы специфических структур в неповрежденных клетках, происходит гиперплазия митохондрий, саркоплазматического ретикулума, рибосом. В результате развивается гипертрофия миокарда, представляющая собой компенсаторную регенераторно-гиперпластическую реакцию, характерную для миокарда. Биохимические процессы чаще нарушаются в ЛЖ.
Клиническая картина
Клинические проявления многообразны и не являются специфичными. Начальные стадии могут протекать бессимптомно, со временем снижение сократительной способности миокарда может привести к тяжелой СН.
Нередко на фоне проявлений основного заболевания отмечают кардиалгию (чаще в области верхушки сердца (92%), реже за грудиной (15%)), расширение границ сердца, приглушенность тонов, небольшой систолический шум на верхушке сердца, нарушения ритма (в основном экстрасистолическую аритмию).
Диагностика
ЭКГ является ведущим методом в распознавании дистрофических изменений в миокарде, которые касаются в основном процесса реполяризации и проявляются чаще всего изменениями конечной части желудочкового комплекса: отмечается депрессия сегмента ST, которая имеет восходящий характер к положительному зубцу Т. Зубец Т также может быть деформированным, низкоамплитудным, сглаженным или отрицательным.
Также может определяться снижение вольтажа комплекса QRS, особенно выраженное при ожирении и микседеме, при тиреотоксикозе амплитуда зубцов чаще увеличена. В некоторых случаях могут возникать замедление внутрипредсердной проводимости, увеличение интервала Q–T, нарушения внутрижелудочковой проводимости. Из нарушений ритма наиболее часто отмечают синусовую тахикардию и экстрасистолическую аритмию.
При формулировке диагноза следует прежде всего указать основное заболевание или этиологический фактор, характер течения кардиомиопатии и основные клинические проявления (наличие нарушений ритма и проводимости, стадию СН).
В дифференциальной диагностике метаболической кардиомиопатии могут иметь значение нагрузочные и медикаментозные пробы, в случае необходимости — проведение коронарографии.
Лечение
Независимо от повреждающего фактора принципиальными для метаболической кардиомиопатии могут быть следующие положения:
- нарушения метаболизма миокарда при своевременном лечении обратимы;
- выраженная СН развивается сравнительно редко, в основном в конечной стадии заболевания, но возникшая СН резистентна к сердечным гликозидам и успех терапии целиком зависит от степени восстановления нарушенного обмена веществ в миокарде.
Помощь больным следует начинать с устранения причины развития дистрофии миокарда. Немаловажное значение имеет отказ от курения и злоупотребления алкоголем, исключение физического и психоэмоционального перенапряжения.
Наряду с лечением основного заболевания необходимо восстановление адекватного энергетического обмена. На первый план выступает применение комплекса лекарственных средств, направленных на улучшение транспорта кислорода в ткани и его утилизации.
На обмен веществ в клетке могут оказывать влияние две группы лекарственных средств: регуляторы экстрацеллюлярной природы (гормональные препараты, блокаторы и стимуляторы центральной и периферической нервной системы) и регуляторы метаболизма интрацеллюлярной природы (ферменты и антиферменты, витамины, кофакторы, разнообразные метаболиты), оказывающие действие на различные пути обмена веществ.
При нарушении процессов окислительного фосфорилирования применяют комплекс витаминов, включающий витамины В1, В2, пантотеновую и липоевую кислоты. Витамины группы В влияют на белковый, липидный, углеводно-энергетический обмен, синтез аминокислот, нуклеотидов.
Среди препаратов с антиоксидантными свойствами широко применяют токоферола ацетат, его сочетание с витамином РР (никотиновой кислотой) способствует улучшению энергетического обеспечения сократительной функции миокарда. Активным антиоксидантом, который участвует в окислительно-восстановительных процессах, является витамин С.
Большое значение для нормализации метаболизма миокарда имеет достаточное поступление в организм незаменимых аминокислот; в том числе метионина, лейцина, аланина, валина, лизина, трионина, триптофана, являющихся пластическим материалом для синтеза белка, ферментов, коферментов. Для улучшения их усвоения рекомендуется назначать их в комплексе с анаболическими стероидами (метандиенон, нандролон).
При прогрессировании дистрофического процесса показано применение внутрь калия хлорида, калия и магния аспарагината для устранения закономерного дефицита внутриклеточного калия, нарушения баланса кальция и магния, что приводит к восстановлению регуляции возбудимости и проводимости миокарда, его автоматизма и сократимости.
Для активации синтеза белков и нуклеиновых кислот применяют соли оротовой кислоты (оротат калия/магния).
Проводимая терапия должна быть направлена на повышение генерации энергии и повышение устойчивости миокарда к гипоксии. В последнее время большое внимание уделяют роли серотонинергической системы в регуляции стрессорной реакции. Специфической особенностью никотинамида является его способность стимулировать процессы аэробного окисления и обмен гликогена, тем самым повышая устойчивость кардиомиоцитов к гипоксии.
Прямое цито- и мембранопротекторное действие на кардиомиоциты в условиях гипоксических состояний оказывает триметазидин.
Продолжительность интенсивной метаболической терапии на ранних стадиях у больных с преимущественно функциональными нарушениями составляет 2–3 нед. При прогрессировании дистрофии миокарда и выявлении органического поражения сердца курс терапии следует повторять несколько раз в год.
ПОРАЖЕНИЯ СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ ПРИ ЭНДОКРИННЫХ НАРУШЕНИЯХ
Сердечно-сосудистая система часто вовлекается в патологический процесс при заболеваниях желез внутренней секреции. Функциональные изменения сердца могут превалировать в клинической картине, и пациент с эндокринным заболеванием становится фактически «кардиальным» больным. Поражение сердца при эндокринных заболеваниях в основном обусловлено обменными нарушениями, вызванными недостатком или избытком того или иного гормона в организме.
ПОРАЖЕНИЕ СЕРДЦА ПРИ САХАРНОМ ДИАБЕТЕ
Термин «диабетическая кардиомиопатия» впервые предложен в 1954 г. для обозначения кардиальных изменений, предшествующих ИБС.
Патогенез
Патогенез метаболической кардиомиопатии при сахарном диабете многофакторный, поражение сердечно-сосудистой системы обусловлено сложными обменными нарушениями, возникающими в связи с абсолютной или относительной недостаточностью инсулина и нарушением толерантности к глюкозе.
Патогенез миокардиальных нарушений включает несколько основных механизмов: повреждение кардиомиоцитов, микроциркуляторные и нейровегетативные нарушения. Первый механизм связан с нарушением метаболизма кардиомиоцитов, снижением эффективности энергетических, пластических процессов и изменением ионного метаболизма, в результате чего снижаются компенсаторные возможности сердечно-сосудистой системы, нарушается сократительная функция миокарда, уменьшается толерантность к физическим нагрузкам. Второй механизм основывается на микроциркуляторных нарушениях в мелких артериях миокарда как локального проявления генерализованной микроангиопатии. Третий механизм включает поражение вегетативной нервной системы в результате формирования нейровегетодистрофии.
Кардиомиопатия, не обусловленная нарушением коронарного кровообращения, возникает у больных молодого возраста с ювенильным сахарным диабетом, для которых нехарактерно развитие выраженного атеросклероза, или у пациентов старшего возраста без сопутствующей ИБС.
Инсулин оказывает на сердце прямое действие, которое заключается в увеличении поступления и стимуляции окисления глюкозы и лактата, увеличении образования гликогена в миокарде. Непрямой эффект инсулина состоит в снижении содержания жирных кислот в плазме крови, уменьшении их поступления в сердце.
Дефицит инсулина вызывает нарушение утилизации тканями глюкозы и усиливает расщепление липидов и белков, также приводит к выраженным изменениям состава внутренней среды организма — гипергликемии, гиперкетонемии, гиперлипидемии с накоплением в крови жирных кислот, диспротеинемии, метаболическому ацидозу, оксидантный стресс вызывает апоптоз миоцитов. Эти нарушения являются определяющими факторами изменения структуры и функции миокарда.
Патогенез и морфогенез диабетического поражения сердца обусловлены не только влиянием гиперинсулинемии на эндотелий сосудов, энергетические и метаболические процессы в миокарде, но и непосредственно связаны с токсико-метаболическим повреждением кардиомиоцитов.
Есть мнение, что причиной разрушения структур кардиомиоцитов, нарушения структуры сарколеммы и ее дериватов, изменения ионного равновесия и снижения активности актомиозинового комплекса кардиомиоцитов является прямая глюкозотоксичность.
В патогенезе кардиомиопатии важную роль играет тканевая гипоксия. Большое значение в развитии гипоксии имеет нарушение транспорта кислорода кровью, функции дыхательных ферментов под влиянием выраженного ацидоза. При сахарном диабете потребность тканей, в том числе миокарда, в кислороде повышена.
Важным фактором развития миокардиодистрофии является нарушение нейроэндокринной регуляции сердца, связанное с преобладанием эффектов контринсулярных гормонов. Доказано, что у пациентов происходит повышение продукции адренокортикотропного и соматотропного гормонов, а также глюкокортикоидов, катехоламинов и глюкагона, это приводит к инициации целой группы метаболических и ультраструктурных процессов, вызывающих развитие метаболической кардиомиопатии.
Патогенез увеличения жесткости миокарда связан с нарушением транспорта кальция, электромеханическим дисбалансом, сопровождающимся асинхронностью расслабления и механическими факторами.
Патологическая анатомия
Характерен фиброз миокарда, связанный с нарушением внутриклеточного метаболизма оксида азота и кальция, а также с пролиферативными процессами, обусловленными действием инсулина и ИФР. Морфологической основой дистрофии миокарда при сахарном диабете является микроангиопатия, характеризующаяся инфильтрацией тучными клетками и фибриноидным набуханием стенок мелких сосудов. При морфологическом исследовании выявляют развитие апоптозной дегенерации, потерю синаптических пузырьков, появление больших вакуолей в цитоплазме клеток симпатических ганглиев. При гистохимическом исследовании в стенках сосудов определяются отложения гликопротеинов. На ультраструктурном уровне определяется утолщение базальной мембраны сосудистой стенки. Важное значение придают дезорганизации мышечных волокон гипертрофированного миокарда.
Клиническая картина и диагностика
Больные с ювенильным сахарным диабетом изредка отмечают колющую боль в области сердца. Возникновение тахикардии покоя связано с поражением блуждающего нерва и относительным преобладанием тонуса симпатического отдела вегетативной нервной системы. Тахикардия сопровождается неэффективными сокращениями миокарда, что приводит к истощению энергоресурсов и в конечном счете к снижению сократительной функции миокарда и развитию СН.
Размеры сердца в пределах нормы. Некоторое приглушение тонов сердца и систолический шум на верхушке чаще отмечают у болеющих сахарным диабетом более 5 лет. В дальнейшем гипергликемия и инсулинорезистентность ассоциируются с увеличением массы ЛЖ и появлением симптомов СН.
На ЭКГ отмечаются синусовая тахикардия или брадикардия, желудочковая экстрасистолическая аритмия, нарушения процессов реполяризации: смещение сегмента ST, изменение амплитуды, инверсия, уплощение, сглаженность или двухфазность зубца Т, нарушение внутрижелудочковой проводимости.
При эхоКГ-исследовании наиболее ранним признаком поражения миокарда при сахарном диабете является нарушение диастолической функции, которое отмечают у 27–69% бессимптомных больных.
При анализе крови уровень гликемии в плазме крови натощак >7,0 ммоль/л.
Лечение
Одной из основных задач лечения больных диабетической кардиомиопатией является профилактика дальнейшего прогрессирования поражения миокарда и развития СН. Важным является борьба с факторами риска: курением, ожирением, малоподвижным образом жизни, несбалансированным питанием. Рекомендации по оптимизации образа жизни должны содержать обоснование соответствующей низкокалорийной диеты для уменьшения массы тела, отказ от курения, регулярные физические нагрузки.
Важной задачей является нормализация обмена веществ, что включает достижение целевых уровней глюкозы, аглюкозурии, нормализации уровня гликированного гемоглобина. Регулярные физические нагрузки позволяют снизить резистентность к инсулину, повысить толерантность к глюкозе, способствуют утилизации глюкозы крови и свободных жирных кислот в мышцах, оказывают благоприятное влияние на функционирование сердечно-сосудистой системы.
Фармакотерапия сахарного диабета II типа направлена на усиление секреции инсулина, снижение инсулинорезистентности и представлена препаратами с различными механизмами действия: бигуаниды, производные сульфонилмочевины, глитазоны, глиниды, ингибиторы α-глюкозидазы, инсулин. Применение метформина позволяет улучшить контроль глюкозы крови у больных сахарным диабетом и способствует снижению общей смертности на 36%.
Для восстановления метаболических нарушений в миокарде назначают препараты α-липоевой кислоты, которая активирует ферменты митохондрий, увеличивает окисление глюкозы, замедляет глюконеогенез и кетогенез, как антиоксидант защищает клетки от повреждающего действия свободных радикалов. Также применяют препараты, способствующие коррекции нарушений обмена в миокарде: триметазидин, триметилгидразиния пропионат.
ТИРЕОТОКСИЧЕСКАЯ БОЛЕЗНЬ СЕРДЦА
Патогенез
Нарушение функции сердечно-сосудистой системы — появление «тиреотоксического сердца» является частым осложнением тиреотоксикоза. Изменения сердечно-сосудистой системы при тиреотоксикозе («тиреотоксическое сердце») обусловлены воздействием избыточного количества тиреоидных гормонов (L-тироксина и 3,5,3-трийод-L-тиронина) на обменные процессы в миокарде, гемодинамику и симпатическую нервную систему. Одним из важных эффектов тиреоидных гормонов является разобщение окислительного фосфорилирования, что приводит к снижению в сердечной мышце содержания АТФ и креатинфосфата. В результате происходит угнетение анаболических процессов: снижается синтез и усиливается распад гликогена и белка, снижается содержание калия в эритроцитах и других клетках. Потребление кислорода миокардом увеличивается, однако эффективность его утилизации в процессе биологического окисления снижается. При избытке тироксина нарушается проницаемость митохондриальных мембран.
Под влиянием тиреоидных гормонов происходит усиление сократительной функции миокарда, вероятно, вследствие активизации стимулирующего влияния на сердце и прямого действия тироксина на сердечную мышцу. Вследствие нарушений энергетических процессов и изменения калий-натриевого насоса происходит ускорение спонтанной деполяризации в клетках синусного узла, что обусловливает более частое образование в нем импульсов. Избыток тиреоидных гормонов изменяет симпатические и парасимпатические влияния на миокард. При высокой степени тиреотоксикоза в результате резкого снижения эффективности биологического окисления, преобладания распада белка над его синтезом снижается уровень энергетических ресурсов и пластических процессов, что приводит в конечном итоге к угнетению сократительной функции миокарда.
Гемодинамика
В основе гиперфункции сердца при тиреотоксикозе лежит повышение сократительной способности миокарда, что обусловлено как повышением активности симпатической нервной системы, так и непосредственным действием тиреоидных гормонов на миокард. При тиреотоксикозе происходят резкие изменения гемодинамики: увеличивается МОК (в основном за счет повышения ЧСС), скорость кровотока и ОЦК. Периферическое сосудистое сопротивление в большом круге кровообращения снижается, а в малом повышается. В результате повышается пульсовое давление. Сердце испытывает диастолическую перегрузку, а правые отделы сердца еще и систолическую перегрузку, увеличенная работа сердца происходит в крайне неблагоприятном для него режиме: вследствие изменений гемодинамики ЛЖ работает в условиях постоянной изотонической гиперфункции, а правый — в условиях смешанного типа гиперфункции (нагрузка объемом и сопротивлением), однако при этом отсутствуют условия для развития компенсаторной гипертрофии миокарда (усилен распад и снижен синтез белка, уменьшено количество АТФ и креатинфосфата). Все это достаточно быстро приводит к развитию СН.
Патологическая анатомия
Гистологические изменения миокарда при тиреотоксикозе характеризуются воспалением и дегенерацией вплоть до развития очагов некроза и фиброза. Гистологические изменения в миокарде непостоянны и неспецифичны. Факторы, обусловливающие поражение сердечно-сосудистой системы у больных с диффузным токсическим зобом, вначале вызывают дистрофические изменения, а в дальнейшем дегенеративно-склеротические. При тяжелом течении заболевания возникают дегенеративные изменения в митохондриях и их распад.
Клиническая картина и диагностика
Больные нередко жалуются на боль в области сердца, часто ноющего, колющего, нередко стенокардитического характера, а также на сердцебиение, которое возникает в состоянии покоя, но при физических нагрузках неадекватно усиливается. Больные отмечают повышенную возбудимость, потливость, мышечную слабость, тремор рук, похудение. Существенным симптомом является постоянная синусовая тахикардия, выраженность которой соответствует тяжести токсического зоба. У 10–20% больных диагностируется тахисистолическая форма фибрилляции предсердий. Характерно повышение САД, что обусловлено увеличением сердечного выброса. Одышка отмечается как при нагрузках, так и в покое. СН, в основном правожелудочковую, отмечают в 15–25% случаев. Признаки левожелудочковой недостаточности обычно выражены меньше, поскольку очень быстро возникает слабость ПЖ.
При осмотре отмечается прекардиальная пульсация и пульсация артерий. Аускультативно определяется повышение звучности сердечных тонов, особенно первого, почти всегда выслушивается систолический шум на верхушке сердца и ЛА.
На ЭКГ, кроме синусовой тахикардии или фибрилляции предсердий, отмечается повышение амплитуды зубца Р, иногда изменения комплекса QRS, снижение сегмента ST и вольтажа зубца Т.
При эхоКГ-исследовании на ранней стадии заболевания выявляют умеренную гипертрофию — утолщение задней стенки, межжелудочковой перегородки и увеличение сократительной функции ЛЖ. В дальнейшем развивается дилатация полостей сердца, увеличивается масса миокарда, уменьшается систолический и минутный объем крови, снижается сократительная функция миокарда.
В сыворотке крови определяется повышение уровней общего и свободного тироксина, трийодтиронина, снижение уровня тиреотропного гормона.
Лечение
Проводится по трем направлениям: нормализация функции щитовидной железы (достижение эутиреоидного состояния), устранение недостаточности кровообращения и восстановление синусового ритма (при фибрилляции предсердий).
Компенсация тиреотоксикоза достигается применением антитиреоидных препаратов или проведением хирургической операции или радиойодтерапии.
Для уменьшения синусовой тахикардии нецелесообразно применять сердечные гликозиды, широко назначают блокаторы β-адренорецепторов. При тахисистолической форме фибрилляции предсердий проводят комбинированное лечение антиаритмическими средствами (пропафенон) и блокаторами β-адренорецепторов, добиваясь восстановления синусового ритма или перевода фибрилляции предсердий в нормосистолическую форму.
Лечение СН не имеет специфических особенностей и обязательно должно проводиться на фоне антитиреоидной терапии. Следует учитывать, что чувствительность миокарда к гликозидам наперстянки может быть повышена.
КЛИМАКТЕРИЧЕСКАЯ (ДИСГОРМОНАЛЬНАЯ) КАРДИОМИОПАТИЯ
Эпидемиология
Изменение демографической структуры общества привело к увеличению в популяции доли женщин старшей возрастной группы (в настоящее время в мире около 500 млн женщин старше 50 лет, то есть в менопаузе).
О существовании связи между расстройством деятельности сердца и изменением функции женских половых органов известно давно. Заболевание может развиваться вследствие дефицита эстрогенов не только в климактерический период, но и у женщин молодого возраста с различными гинекологическими заболеваниями (миома матки, эндометриоз и др.), при посткастрационном и предменструальном синдромах. Климактерическая кардиомиопатия диагностируется иногда и у мужчин (климакс отмечают у 10–20% лиц мужского пола).
Патогенез
Менопауза, не являясь собственно заболеванием, приводит к нарушению эндокринного равновесия в организме и способствует развитию сердечно-сосудистых заболеваний.
В патогенезе обменных нарушений основное значение имеет нарушение активности эстрогенов, в норме благоприятно влияющих на белковый и электролитный обмен в миокарде и регулирующих симпатические влияния на сердце. При патологическом климаксе в миокарде происходят метаболические нарушения, приводящие к дистрофическим изменениям, в большинстве случаев носящим обратимый характер и лишь в некоторых случаях заканчивающимся развитием миокардиофиброза (кардиосклероза) (схема 8.2). Увеличение количества абдоминального жира и развитие абдоминального ожирения связано как с физиологическими изменениями, так и с изменениями образа жизни. Среди причин абдоминального ожирения после менопаузы можно выделить изменение баланса энергии — снижение скорости обменных процессов наряду с повышением аппетита и увеличением поступления энергии с пищей на фоне повышения тонуса симпатической нервной системы, усиления глюкокортикоидной стимуляции и падения уровня гормона роста. В основе патогенеза климактерической АГ лежит гипоэстрогения, которая сопровождается повышением возбудимости гипоталамо-гипофизарных структур, нарушением центральной и периферической регуляции сосудистого тонуса. Одним из механизмов является отсутствие в период менопаузы депрессорного эффекта фолликулярного гормона.
Клиническая картина
Наиболее распространенными являются жалобы на продолжительную, почти постоянную боль в области сердца разнообразного характера, локализующуюся слева от грудины, в области верхушки. Боль не провоцируется физическим напряжением. Кардиалгия не прекращается после приема нитроглицерина. Характерно сердцебиение при нормальном пульсе, не связанное с физической нагрузкой, нередко появляется в покое.
Больные часто жалуются на ощущение неудовлетворенности вдохом, невозможность вдохнуть полной грудью, которое не связано с физическими нагрузками и часто возникает в покое.
Типичны нарушения функции вегетативной нервной системы: гиперемия или побледнение кожи, потливость, приливы крови, сердцебиение, онемение конечностей, озноб, нарушение ритма дыхания, полиурия, головокружение, нарушение терморегуляции.
Большое количество жалоб обусловлено изменениями психического состояния: эмоциональная лабильность, раздражительность, плаксивость, повышенная возбудимость, нередко подавленное настроение, страхи, ухудшение памяти. Усугубление симптомов связано с нагрузками, особенно эмоциональными.
При патологическом климаксе нередко возникает симптоматическая АГ. Впоследствии, после исчезновения приливов крови и других проявлений климактерического синдрома, невротическое состояние может стать причиной развития гипертонической болезни.
У большинства мужчин с климактерической кардиомиопатией отмечают те или иные симптомы патологического климакса со стороны мочеполовой системы: отсутствие или снижение (редко повышение) либидо, снижение потенции. Больные часто жалуются на расстройства мочеиспускания, что обычно связано с доброкачественной гиперплазией предстательной железы.
Вазомоторный синдром проявляется в виде приливов крови, то есть внезапно возникающего ощущения жара в верхней половине туловища, коже лица, шеи, которое сменяется последовательно гиперемией и потоотделением. Наряду с приливами крови в отдельных областях тела периодически появляются парестезии: ощущение онемения, покалывания, ползания мурашек.
Климактерическая кардиомиопатия может возникнуть остро или развиваться постепенно. Характерно несоответствие между интенсивностью и длительностью болевого синдрома и удовлетворительным состоянием кровообращения.
При объективном обследовании характерно несоответствие между обилием жалоб и отсутствием клинических признаков коронарной или СН.
Диагностика
На ЭКГ самыми частыми изменениями являются снижение сегмента ST и/или инверсия зубца Т, которые в основном регистрируют в правых и средних грудных отведениях (V1–4). Зубец Т может длительное время быть отрицательным, затем положительным, а через несколько дней вновь отрицательным без какой-либо связи с клинической картиной болезни, на фоне удовлетворительного состояния больного. Изменения на ЭКГ не соответствуют клиническим проявлениям, физические нагрузки практически не влияют на конфигурацию зубцов. Часто возникают синусовая аритмия, предсердная и желудочковая экстрасистолия, пароксизмальная суправентрикулярная тахикардия. Изредка регистрируют нарушения предсердно-желудочковой и внутрижелудочковой проводимости.
На ранних стадиях климактерическая кардиомиопатия протекает чаще изолированно и характеризуется типичной клинической картиной заболевания. В более поздние периоды клиническая картина зависит от присоединения ИБС, воспалительных процессов в миокарде и других болезней, что несомненно отягощает течение кардиомиопатии и ухудшает прогноз.
Лечение
Должно быть направлено на устранение всех симптомов заболевания. Важное значение имеет модификация образа жизни, включающая повышение физической активности и соблюдение диеты с ограничением потребления насыщенных жиров и увеличением в рационе доли моно- и полиненасыщенных жиров и грубой клетчатки. Для нормализации деятельности нервной системы обычно назначают седативные препараты, транквилизаторы, иногда антидепрессанты.
Для лечения АГ в постменопаузе наиболее целесообразно назначение ингибиторов АПФ и диуретиков, которые должны быть нейтральными в отношении показателей углеводного и липидного обмена. Женщинам в постменопаузе должны назначаться только высокоселективные блокаторы β-адренорецепторов новой генерации, не оказывающие негативного воздействия на липидный и углеводный обмен.
Назначение заместительной гормонотерапии является патогенетически обоснованным в лечении больных с климактерической кардиомиопатией. Применяют препараты, содержащие эстрогены и гестагены. Половые гормоны подавляют повышенную активность гипоталамо-гипофизарных структур мозга и опосредованно влияют на сердце, нормализуя влияние вегетативной нервной системы. Не исключено, что половые гормоны ослабляют повышенную активность САС и тем самым нормализуют метаболические процессы в миокарде. Эстрогены оказывают непосредственное сосудорасширяющее действие на коронарные сосуды, а также нормализуют электролитный и белковый обмен в миокарде. Дозы и общая продолжительность лечения зависят от исходного гормонального фона и уровня эстрогенов, лечение следует проводить под наблюдением эндокринолога. Необходимо отметить, что климактерическая кардиомиопатия является самоизлечивающимся заболеванием, при котором гормоны оказывают лишь вспомогательное заместительное действие, гормональную терапию следует назначать на длительный срок. Лечение гормонами устраняет тягостные проявления климактерического синдрома и после окончания возрастной перестройки эндокринной системы заболевание исчезает.
Прогноз
Как правило, благоприятный. Снижение трудоспособности в большинстве случаев носит временный характер. Полное выключение больных из привычной трудовой обстановки, как правило, играет отрицательную роль, приводит к излишней концентрации внимания на тягостных ощущениях со стороны сердца.
ПОРАЖЕНИЯ СЕРДЦА ПРИ НАРУШЕНИИ ОБМЕНА ВЕЩЕСТВ
Нарушения обмена веществ в организме всегда отражаются на течении метаболических процессов в миокарде, нередко вызывая нарушение его функции и структуры. При различных заболеваниях первоначально могут нарушаться один или несколько путей метаболизма, что в дальнейшем обязательно отражается на энергообеспечении сердечной мышцы. При некоторых нарушениях обмена в межуточной ткани миокарда и в коронарных сосудах откладываются патологические продукты нарушенного метаболизма белков, углеводов, минералов или накапливаются избыточные компоненты нормального обмена. К таким заболеваниям относят амилоидоз, гликогеноз, гемохроматоз и др.
НАРУШЕНИЯ БЕЛКОВОГО ОБМЕНА. АМИЛОИДОЗ
Определение
Амилоидоз — системное заболевание неустановленной этиологии, характеризующееся внеклеточным отложением в органах и тканях (главным образом в медии артерий, периваскулярной соединительной и нервной ткани, в ретикулоэндотелиальной системе, а также миокарде, почках, печени, коже) особого белка β-фибриллярной структуры — амилоида.
Этиология и патогенез
Амилоидоз является следствием нарушения белкового обмена и может быть приобретенным или наследственным. Наследственный амилоидоз является аутосомным доминантным заболеванием. Ряд авторов связывают развитие заболевания с изменением свойств белков тканей вследствие аутоиммунных процессов под влиянием комплекса антиген — антитело. Диспротеинемия с накоплением в плазме крови грубодисперсных фракций белка и аномальных белков (парапротеинов) ведет к выходу последних из сосудов ткани с образованием амилоидных субстанций.
В последние годы стала возможной более точная биохимическая идентификация белков, входящих в состав амилоидных фибрилл, на основании чего выделены типы амилоида, определена связь отдельных типов с клиническими формами амилоидоза, изучены белки-предшественники, предположительно участвующие в синтезе белков.
Выделяют четыре типа амилоидоза: первичный (системный), вторичный, семейный (наследственный) и сенильный (старческий).
Наиболее распространен первичный тип (85%) с преимущественным поражением сердца, при котором амилоид образован легкими цепями молекул k и λ иммуноглобулина (AL-тип), часто ассоциирован с миеломной болезнью, более часто отмечают у мужчин и редко в возрасте младше 30 лет.
Вторичный амилоидоз возникает в результате образования неиммуноглобулиновых белков, миофибриллы содержат амилоидный протеин А, не относящийся к иммуноглобулинам (АА-тип), что часто происходит при хронических воспалительных заболеваниях — ревматоидном артрите, туберкулезе, болезни Крона и при семейной средиземноморской лихорадке.
Семейный или наследственный амилоидоз чаще всего является следствием образования мутантного белка преальбумина (транстиретина). Установлен аутосомно-доминантный тип наследования. Выявлены гены, ответственные за синтез этих белков, и идентифицирован характер генных мутаций.
Сенильный кардиальный амилоидоз, также известный как амилоид SSA, возникает вследствие образования патологического транстиретина у лиц старшего возраста. Выделяют две формы связанного с возрастом амилоидоза — амилоидоз предсердий, который охватывает только предсердия, и старческий аортальный амилоидоз, ограниченный аортой.
Патологическая анатомия
Миокард при амилоидозе сердца очень плотный на ощупь, утолщенный, мало поддается растяжению. Объем полостей сердца существенно не изменен или незначительно увеличен. Амилоид откладывается в разных отделах сердца, преимущественно в миокарде предсердий и желудочков, эндокарде, в клапанах, перикарде, нередко в синусном и AV-узлах, а также в мелких артериальных и венозных сосудах, включая vasa vasorum коронарных артерий, суживая их просвет вплоть до полной обтурации. В результате мышечные волокна сердца оказываются «замурованными» в массах амилоида, что приводит к атрофии сократительного миокарда.
Клиническая картина
Амилоидное поражение сердца не имеет специфических симптомов, развивается постепенно и может длительное время протекать бессимптомно, даже при выявлении отложений амилоида в миокарде при биопсии. Следует обратить внимание, что во время появления симптомов существует весьма значительная инфильтрация сердца амилоидом. У некоторых пациентов возникает боль в области сердца, иногда носящая стенокардический характер как следствие накопления депозитов амилоида в коронарных артериях.
В 10–15% случаев отмечается ортостатическая гипотензия, иногда с симптомами синкопальных состояний.
При аускультации на фоне глухих тонов сердца можно выслушать систолический шум митральной регургитации, при развитии СН — протодиастолический ритм галопа.
Часто определяют различные нарушения ритма, которые нередко могут быть причиной внезапной смерти. У некоторых больных отмечается выраженная брадикардия.
СН выявляют у 45–56% больных. Вначале доминирует правожелудочковая СН с повышенным давлением в яремных венах, гепатомегалией, периферическими отеками, асцитом. Затем возникает систолическая дисфункция и застойная СН.
Диагностика
Изменения на ЭКГ неспецифичны, наиболее типично наличие брадикардии, снижение амплитуды зубцов. Иногда наличие патологического зубца Q и отсутствие зубца R в отведениях V1–3 симулируют ИМ. Накопление депозитов амилоида в проводящей системе могут обусловливать различные расстройства образования импульса и проведения — возможны различные нарушения проводимости, включая полную блокаду сердца: часто выявляются предсердные и желудочковые нарушения ритма (синдром слабости синусного узла, фибрилляция предсердий (у 30% больных), желудочковая экстрасистолическая аритмия).
Двухмерная эхоКГ и допплерография являются основными методами неинвазивной диагностики. При обследовании выявляют нормальные или уменьшенные размеры полости ЛЖ со значительным утолщением миокарда и характерным нарушением его структуры с диффузным гранулярным блеском (рис. 8.1а, б). Отмечается также утолщение межпредсердной перегородки и створок клапанов, увеличение предсердий, наличие небольшого или умеренного перикардиального выпота. Нарушение диастолической функции ЛЖ и ПЖ происходит по рестриктивному типу нарушения их наполнения. В тяжелых случаях выявляются признаки различной степени нарушения систолической функции обоих желудочков.
При рентгеноскопии отмечают уменьшение пульсации контура сердца, размеры сердца увеличены (кардиомегалия) и обычно не соответствуют степени тяжести застойной СН.
К достижениям последних лет относится введение в клиническую практику метода сцинтиграфии с меченным 123I сывороточным Р-компонентом (SАР) для оценки распределения амилоида в организме. Р-компонент содержится в небольшом количестве (5–10%) в амилоиде всех типов; радиоактивный SАР, введенный больному амилоидозом, специфически связывается с амилоидными депозитами и может быть визуализирован и количественно оценен на серии сцинтиграмм. Метод особенно полезен для контроля за динамикой тканевых отложений амилоида в процессе лечения.
Для диагностики также используют сцинтиграфию с изотопом технеция 99mТс-пирофосфатом, способным связываться с амилоидом многих типов, однако эта проба оказывается положительной только при значительных отложениях амилоида в сердце, которые можно определить и с помощью эхоКГ.
МРТ используется для идентификации утолщения миокарда и небольшого размера полости ЛЖ при амилоидозе, что сопоставимо с данными эхоКГ.
Диагноз «амилоидоз» должен быть подтвержден эндомиокардиальной биопсией. При изучении биоптатов тканей важно не только выявить амилоид, но и провести иммуногистохимическое исследование для идентификации его типа.
Диагноз «амилоидоз сердца» чаще устанавливают при аутопсии, поскольку при жизни в ряде случаев не выявляют объективных причин, которыми можно было бы объяснить возникновение патологических признаков.
Лечение
Терапия при первичном амилоидозе включает клеточную антиплазменную терапию, которая останавливает продукцию легких цепей, а также применение алкилирующих средств (мелфалан) и преднизолона. Благоприятный эффект химиотерапии показан в двух рандомизированных испытаниях. Перспективна трансплантация стволовых клеток с органной ремиссией в 50% случаев. Другим подходом к лечению амилоидоза сердца может быть применение талидомида с дексаметазоном. Недавно показана эффективность леналидомида.
Для лечения пациентов с нарушениями ритма сердца назначают антиаритмические препараты. При явлениях полной поперечной блокады и слабости синусного узла эффективна имплантация искусственного водителя ритма. Кардиостимуляторы применяют для лечения пациентов с тяжелыми клинически выраженными нарушениями проводимости.
СН часто рефрактерна к медикаментозной терапии. Для уменьшения недостаточности кровообращения основными препаратами являются диуретики, которые применяют с осторожностью в низких дозах, и вазодилататоры — ингибиторы АПФ или блокаторы рецепторов ангиотензина II, хотя они плохо переносятся и могут вызвать значительную артериальную гипотензию или ортостатические симптомы, особенно у пациентов с амилоид-индуцированной дисфункцией автономной нервной системы. Не рекомендуется применять дигоксин из-за его токсичности и опасности развития аритмий, однако при тщательном ЭКГ-мониторировании его можно применять для контроля ритма у пациентов с фибрилляцией предсердий.
Блокаторы кальциевых каналов неэффективны в лечении при амилоидозе сердца. Больные могут быть гиперчувствительны к негативным инотропным эффектам блокаторов кальциевых каналов, их применение может привести к нарастанию симптомов декомпенсации.
Блокаторы β-адренорецепторов могут спровоцировать угрожающие жизни нарушения проводимости.
При резком снижении сократительной способности предсердий, свидетельствующем о массивной инфильтрации, даже при синусовом ритме показано применение антиагрегантов или антикоагулянтов, что обусловлено повышенным риском тромбообразования.
Трансплантация сердца обычно не проводится, поскольку возникают рецидивы амилоидоза в аллотрансплантате, а также неуклонное прогрессирование его в других органах, что сокращает продолжительность жизни больных.
Прогноз
Течение амилоидоза прогрессирующее, прогноз неблагоприятный, хотя зависит от формы, сроков диагностики и степени вовлечения жизненно важных органов. Каждый из четырех основных типов амилоидной болезни имеет различные степени вовлечения сердца, клинические симптомы и прогноз. Выживаемость больных со старческим амилоидозом намного выше, чем с первичным амилоидозом, — в среднем соответственно 60,0 и 5,5 мес со времени установления диагноза. Летальный исход (приблизительно через 1,5–2,5 года после появления первых признаков поражения сердца) обычно наступает вследствие нарушений ритма и проводимости, а также внесердечных осложнений (легочной или системной эмболии). У больных с вовлечением проводящей системы нередко возникает внезапная смерть. Самая низкая выживаемость отмечена у больных с рефрактерной к терапии застойной СН (в среднем 6 мес), особенностью которой является преимущественно правосердечный или тотальный ее тип с резким набуханием шейных вен и значительным повышением венозного давления, застойным увеличением печени и полостными отеками (гидроторакс, гидроперикард, асцит).
НАРУШЕНИЯ ПРЕИМУЩЕСТВЕННО ЛИПИДНОГО ОБМЕНА
ГОЛОДАНИЕ И КАХЕКСИЯ
Патогенез
Голодание, длительное неполноценное питание, кахексия приводят к нарушениям деятельности сердца, которые сопровождаются уменьшением массы миокарда, обычно пропорционально меньшим, чем уменьшение массы тела вследствие атрофии мышечных волокон, дегенеративным изменениям в миокарде и к СН.
Патологическая анатомия
При голодании микроскопически отмечают вакуолизацию миофибрилл, особенно вокруг ядер, изменения хроматина ядер и митохондрий. В далеко зашедших случаях выявляют бурую атрофию и дистрофические изменения миокарда.
Клиническая картина и диагностика
Основными проявлениями нарушения функции сердечно-сосудистой системы при голодании являются синусовая брадикардия, уменьшение МОК, снижение венозного давления и АД (преимущественно систолического), что нередко сопровождается головокружением, а при быстром перемещении из горизонтального положения в вертикальное — обмороком. Часто возникают отеки, обусловленные гипопротеинемией и увеличением ОЦК (но не СН).
На ЭКГ отмечаются отклонение оси сердца вправо, синусовая брадикардия, низкий вольтаж зубцов, иногда изменения зубца Т и комплекса QRS, которые, по-видимому, обусловлены нарушением обмена энергии и электролитов в миокарде.
Лечение заключается в восстановлении полноценного питания.
ОЖИРЕНИЕ
Эпидемиология
Эпидемиологические исследования свидетельствуют, что ожирение связано с сердечно-сосудистыми заболеваниями и преждевременной смертностью. Ожирение само по себе приводит к комплексному и прогностически неблагоприятному поражению сердца. Вероятность развития ГЛЖ у лиц с нормальной массой тела составляет 5,5%, а у лиц с ожирением — 30%. По данным Фремингемского исследования установлено наличие высокодостоверной связи между ИМТ, полостными размерами и толщиной стенок ЛЖ.
По современным представлениям ожирение представляет собой независимый фактор риска развития СН, являясь ее причиной у 11% мужчин и 14% женщин в США. По данным Фремингемского исследования увеличение ИМТ на каждый 1 кг/м2 повышает риск развития СН на 5% у мужчин и на 7% у женщин независимо от других факторов риска.
Этиология
Ожирение может быть самостоятельным заболеванием, возникающим вследствие избыточного потребления пищи с высокой калорийностью, или синдромом, сопровождающим различные заболевания, и развиваться вследствие ряда нейроэндокринных, социальных, поведенческих и генетических факторов. Генетические факторы играют важную роль в развитии ожирения. Результаты исследований свидетельствуют, что существует редко идентифицируемая группа генов, вызывающих значительное ожирение, однако чаще выявляют гены «восприимчивости», которые детерминируют склонность к ожирению и регулируют распределение жировой массы в организме, скорость обменных процессов и их реакцию на физическую активность и диету, контролируют пищевые привычки. Идентифицировано более 41 сайта в геноме, которые, возможно, связаны с развитием ожирения в популяции.
Патогенез
При ожирении происходит постепенное увеличение размеров клеток жировой ткани, ведущее к изменению их свойств. Гормонально-метаболические сдвиги, характерные для ожирения, могут напрямую воздействовать на структуру и массу миокарда. У пациентов с ожирением адипоциты жировой ткани высвобождают большое количество биологически активных субстанций, участвующих в регуляции сосудистого тонуса: ангиотензин II, интерлейкины, простагландины, эстрогены, ИФР, ФНО-α, ингибитор активатора плазминогена-1, лептин и другие, что повышает риск развития сердечно-сосудистых осложнений, при этом снижается уровень адипонектина, специфического циркулирующего белка жировой ткани, который вовлечен в регулирование метаболизма липидов и глюкозы (схема 8.3). Синтезируемый в жировой ткани лептин, важный маркер энергетического баланса, стимулирует гиперсимпатикотонию, способствует повышению уровня АКТГ, кортизола и альдостерона.
Ведущее значение в развитии различных форм ожирения имеют изменения функционирования гипоталамо-гипофизарной системы. Эндоканнабиноидная система, представленная в мозге (гипоталамусе) и периферически в жировой ткани (адипоцитах), печени, скелетных мышцах и пищеварительном тракте, посредством каннабиноидных рецепторов 1-го типа (СВ1) участвует в центральной и периферической регуляции энергетического баланса, а также метаболизме глюкозы и липидов, играет роль в контроле потребления пищи и массы тела. Гиперактивация этой системы ассоциирована с мотивацией к увеличенному потреблению пищи и ожирением и приводит к нарушению механизмов обратной связи, которые поддерживают устойчивый гомеостаз.
Присоединение АГ при ожирении происходит примерно у 60% больных, механизмы ее формирования связывают с развитием гормонально-метаболических отклонений, вызванных накоплением жировой ткани. Ключевую роль среди них играет развитие инсулинорезистентности и компенсаторной гиперинсулинемии, которая, усиливая задержку натрия почками, способствует дальнейшему росту ОЦК. Гипертензивное действие может оказывать и лептин, стимулирующий симпатическую нервную систему. Ожирение, АГ, дислипидемию и гипергликемию, в основе которых лежит инсулинорезистентность, объединяют в понятие «метаболический синдром».
При высокой степени ожирения нельзя исключить определенную роль гипоксии в изменении нейроэндокринной регуляции кровообращения и в развитии дистрофии миокарда. Включение гипоксического фактора в патогенез дистрофических поражений сердца может стать существенным механизмом не только их возникновения, но и развития СН.
Гемодинамика
Сердце у больных ожирением испытывает перегрузку объемом. ОЦК и объем плазмы крови увеличиваются пропорционально степени увеличения массы тела, что приводит к увеличению наполнения ЛЖ и ударного объема, дилатации и росту массы ЛЖ. Считается, что нарастание сердечного выброса при ожирении физиологически связано с удовлетворением метаболических потребностей возросшей тканевой массы тела. Развитие сердечно-сосудистых осложнений при ожирении связано с истощением компенсаторных механизмов миокарда, обусловленным увеличением величины ОЦК, которая формируется пропорционально объему сосудистой сети периферических тканей. Нарастающее содержание жировой ткани в организме десинхронизирует физиологические взаимосвязи между сердцем и кровотоком периферических метаболически активных тканей.
Сердечный выброс в состоянии покоя у больных с тяжелой степенью ожирения достигает 10 л/мин, причем на обеспечение кровотока в жировой ткани используется от ⅓ до ½ этого объема. Увеличенный объем крови в свою очередь увеличивает венозный возврат в ПЖ и ЛЖ, вызывая их дилатацию, увеличивая напряжение стенки. Это приводит к ГЛЖ, которая сопровождается снижением диастолической податливости камеры, приводя к повышению давления наполнения ЛЖ и его расширению.
Увеличение толщины миокарда снижает чрезмерное напряжение его волокон, что позволяет сохранить нормальную сократительную способность ЛЖ, одновременно создает предпосылки для диастолической дисфункции, в основе которой лежит относительное уменьшение количества капилляров на единицу объема мышечной ткани и ухудшение условий диффузии кислорода в гипертрофированных мышечных волокнах. По мере прогрессирования дилатации ЛЖ увеличение напряжения стенки приводит к систолической дисфункции.
Патологическая анатомия
При ожирении отмечают увеличенное отложение жировой ткани под эпикардом обоих желудочков и в поверхностных слоях миокарда, что со временем приводит к атрофии мышечных волокон, замещению их жировой тканью (cor adiposum). Миокард на разрезе имеет желтоватый оттенок. Выявляют наличие диффузной мышечной гипертрофии, которая является наиболее характерным проявлением ожирения со стороны сердечно-сосудистой системы.
Клиническая картина
У взрослого ожирение устанавливают при ИМТ >30,0 кг/м2. Клинически выраженные расстройства кровообращения развиваются у больных с ИМТ >40,0 кг/м2.
Жалобы на боль в сердце ноющего, колющего характера, сердцебиение и перебои в работе сердца при физических нагрузках. По мере накопления избыточной массы тела постепенно развивается прогрессирующая одышка при нагрузках, возникает ортопноэ и пароксизмальная ночная одышка, появляются отеки нижних конечностей, возможно увеличение живота в объеме.
Во многих проспективных исследованиях установлено, что увеличение массы тела приводит к повышению АД. У больных с ожирением высок риск присоединения ИБС, течение которой особенно агрессивно и тяжело.
Сердце принимает «поперечное» положение из-за высокого стояния диафрагмы, смещаясь влево и несколько кзади. Аускультативно определяется выраженная глухость тонов. Пульс имеет склонность к учащению.
При крайних степенях ожирения иногда отмечают клинический синдром, проявляющийся сочетанием сонливости, альвеолярной гиповентиляции и легочной гипертензии с гипертрофией ПЖ — синдром Пиквика.
Диагностика
На ЭКГ обычно синусовая тахикардия, отклонение электрической оси сердца влево, снижение сегмента ST в I–II и V5–6 отведениях, уплощенный и отрицательный зубец Т. У некоторых больных регистрируется низкоамплитудный зубец РІІІ и глубокий QІІІ. Отмечаются признаки ГЛЖ.
При эхоКГ-исследовании выявляют гипертрофию и дилатацию ЛЖ, увеличение левого предсердия, диаметра восходящей аорты. С помощью допплеровской эхоКГ выявляют признаки диастолической дисфункции, может определяться аортальная регургитация. В последующем происходит нарушение и систолической функции. Возможно расслоение листков перикарда за счет отложения жира. Проведение эхоКГ-исследования часто затруднено из-за большой толщины грудной клетки, сужения межреберных промежутков, смещения сердца кзади.
При изучении гемодинамических показателей у всех пациентов выявлено увеличение ОЦК, что сопровождается нарастанием ригидности миокарда ЛЖ, ростом давления его наполнения и УОК. С увеличением степени ожирения повышается конечное диастолическое давление в ПЖ, среднее давление в ЛА, давление заклинивания в легочных капиллярах и конечное диастолическое давление в ЛЖ. Эти изменения вызывают расширение полостей левого предсердия, ПЖ и правого предсердия. Давление крови в ПЖ, как правило, также повышено.
Рентгенологическая картина всегда изменена вследствие высокого стояния диафрагмы и скопления жира в области верхушки сердца, что создает картину его кажущегося увеличения. Пульсация вялая, тонус сердца понижен.
Лечение
Начальные дистрофические изменения миокарда при ожирении являются в значительной мере обратимыми при нормализации массы тела. Первоочередным этапом лечения является коррекция пищевых привычек и повышение физической активности. Специфические рекомендации включают 30 мин физической активности по крайней мере 5 раз в неделю, уменьшение калорийности пищи в среднем до 1500 ккал/сут, снижение потребления жиров до 30–35% дневной энергетической ценности (с оговоркой 10% для мононенасыщенных жирных кислот, например оливковое масло), отказ от трансгенных жиров, увеличение потребления продуктов, содержащих волокна, до 30 г/сут и отказ от жидких моно-и дисахаридов.
Для уменьшения массы тела применяют медикаментозные и хирургические методы лечения ожирения. Назначают ингибиторы липаз (средства периферического действия) и анорексигенные средства (центрального действия).
Лечение сердечно-сосудистых расстройств у больных с ожирением зависит от характера поражения сердца. Для лечения АГ наиболее целесообразно назначение ингибиторов АПФ и диуретиков, которые должны быть нейтральными в отношении показателей углеводного и липидного обмена. Должны назначаться только высокоселективные блокаторы β-адренорецепторов новой генерации, не оказывающие негативного воздействия на липидный и углеводный обмен.
При наличии признаков СН лечение проводят в соответствии с современными рекомендациями.
АЛКОГОЛЬНАЯ КАРДИОМИОПАТИЯ
Эпидемиология
Одна из форм алкогольного поражения сердца, отмечается у 50% лиц, на протяжении длительного времени злоупотребляющих алкоголем.
Алкогольную кардиомиопатию выявляют приблизительно у ⅓ всех больных с неишемической кардиомиопатией, 40–50% больных умирают в течение 3–6 лет.
Этиология
Этиологическим фактором является этанол и/или его метаболиты. Развитие алкогольной кардиомиопатии могут обусловить стрессовые состояния, недостаточность питания (дефицит белков, витаминов), наследственная предрасположенность, вирусная инфекция на фоне снижения иммунитета, изменения исходного состояния миокарда. Не всегда отмечается отчетливый параллелизм между количеством употребляемого этанола, длительностью интоксикации и выраженностью поражения сердца.
Патогенез
Основной из метаболитов этанола, ацетальдегид, оказывает прямое повреждающее воздействие на клеточные и субклеточные мембраны кардиомиоцитов, связанное с их способностью растворять липиды и увеличивать текучесть биологических мембран. На определенном этапе интоксикации это может вызывать нарушение обмена веществ в миокарде и ингибирование основных путей утилизации энергии в клетках сердца, в результате угнетения функции дыхательной цепи митохондрий возникает гипоксия миокарда. Опосредованное воздействие происходит в результате влияния алкоголя на различные отделы нервной системы и функцию надпочечников.
Патологическая анатомия
Длительное употребление алкоголя вызывает жировую инфильтрацию миокарда, дегенеративные изменения в стенках коронарных артерий и нейронах, расположенных в сердце. При микроскопическом исследовании отмечают исчезновение поперечной исчерченности миофибрилл, пикноз ядер, интерстициальный отек, вакуольную и жировую дистрофию, иногда единичные или множественные очаги некроза, мелкие участки фиброза.
Клиническая картина
Как правило, больные упорно отрицают злоупотребление алкоголем.
Развернутую клиническую картину с явлениями СН, стойкими нарушениями ритма и проводимости, тромбоэмболическими осложнениями, кардиомегалией выявляют редко.
Первым клиническим проявлением наиболее часто бывают нарушения ритма без признаков застойной СН. Развитие заболевания имеет несколько стадий — от функциональных расстройств, нарушений ритма сердца преходящего характера до стойкой гипертрофии миокарда с последующим развитием СН.
К наиболее частым и типичным клиническим симптомам относят:
- возбуждение, тремор рук, суетливость, многословность;
- ощущение нехватки воздуха, кардиалгию, тахикардию;
- похолодение конечностей;
- ощущение жара во всем теле, гиперемию кожи лица, инъецированность склер;
- потливость;
- повышение АД.
Начальными признаками заболевания принято считать сердцебиение и одышку при физической нагрузке. На более поздних стадиях заболевания состояние пациентов постепенно ухудшается.
Диагностика
На ЭКГ характерными изменениями являются укорочение интервала Р–Q, удлинение интервала Q–T в сочетании с небольшой элевацией сегмента ST и заостренным высоким с широким основанием зубцом Т, синусовая аритмия, бради- или тахикардия. Нередко нарушения ритма (предсердная и желудочковая экстрасистолическая аритмия, фибрилляция предсердий) и проводимости (атриовентрикулярная и внутрижелудочковая блокады) возникают после длительного и/или однократного употребления большого количества алкоголя (синдром «праздничного» сердца).
О наличии алкогольной кардиомиопатии может свидетельствовать отсутствие определенной причины фибрилляции предсердий (тиреотоксикоз, ревматический порок сердца) у мужчин молодого возраста.
Диагностику затрудняет и отсутствие маркеров алкогольного поражения сердца.
Диагностировать алкогольную кардиомиопатию легче в том случае, если в анамнезе имеются указания на длительное употребление алкоголя и определяются клинические признаки кардиомегалии, аритмии или застойной СН при отсутствии других причин, способных привести к аналогичным нарушениям сердечной деятельности.
При эхоКГ-исследовании отмечается дилатация полости ЛЖ, снижение его сократительной способности, возможна диффузная гипоксия. При допплеровском исследовании могут выявляться признаки митральной регургитации.
Лечение
При лечении обязательно исключают употребление алкоголя. Полная абстиненция может остановить прогрессирование поражения сердца на ранних этапах (обычно в первые 2–6 мес).
На ранних стадиях без проявлений СН и при наличии кардиалгии, тахикардии, АГ и аритмии рекомендованы блокаторы β-адренорецепторов. При выраженной кардиомегалии следует назначать сердечные гликозиды, однако строго контролировать их прием в целях предупреждения кардиотоксического эффекта. В комплексное лечение включают мочегонные средства, витамины, анаболические гормоны, соли калия и магния.
Прогноз
При полном отказе от употребления спиртных напитков и под влиянием лечения размеры сердца у больных с алкогольной кардиомиопатией нередко уменьшаются. Восстановление основных функций миокарда и улучшение общего состояния наступают очень медленно, сроки относительного выздоровления исчисляются месяцами и годами.
ЛИТЕРАТУРА
- Александров А.А., Кухаренко С.С. (2006) Миокардиальные проблемы ожирения. Рос. кардиол. журн., 2: 11-17.
- Артемчук А.Ф. (2000) Клинические особенности и терапия сердечно-сосудистых нарушений при алкоголизме. Укр. кардіол. журн., 4: 68-71.
- Ефимов А.С., Соколова Л.К., Рыбченко Ю.Б. (2005) Сахарный диабет и сердце. Мистецтво лікування, 34: 44-49.
- Зубкова С.Т., Тронько Н.Д. (2006) Сердце при эндокринных заболеваниях. Библиотечка практикующего врача, Киев, 200 с.
- Коваленко В.Н., Несукай Е.Г. (2001) Некоронарогенные болезни сердца. Практ. руководство. Морион, Киев, 480 с.
- Моисеев В.С., Сумароков А.В. (2001) Болезни сердца. Универсум паблишинг, Москва, с. 369-378.
- Alpert M.A. (2001) Obesity cardiomyopathy: pathophysiology and evolution of the clinical syndrome. Amer. J. Med. Sci., 321: 225-236.
- Bartnik M., Van der Berghe G., Betteridge J. et al. (2007) Guidelines on diabetes, pre-diabetes and cardiovascular diseases. Eur. Heart J., 28: 88-136.
- Cooper L.T., Baughman K.L., Feldman A.M. (2007) The role of endomyocardial biopsy in the management of cardiovascular disease. Eur. Heart J., 28: 3077-3093.
- Cote M., Matias I., Lemieux I. et al. (2007) Circulating endocannabinoid levels, abdominal adiposity and related cardiometabolic risk factors in obese men. Int. J. Obes. (Lond), 31: 692-699.
- Di Marzo V., Matias I. (2005) Endocannabinoid control of food intake and energy balance. Nature Neuroscience, 8: 585-589.
- Falk R.H. (2005) Diagnosis and management of the cardiac amyloidoses. Circulation, 112: 2047-2060.
- Fauchier L. (2003) Alcoholic cardiomyopathy and ventricular arrhythmias. Chest., 123: 1320-1325.
- Galinier M., Pathak K., Roncalli J. et al. (2005) Obesity and cardiac failure. Arch. Mal. Coeur. Vaiss., 98: 39-45.
- Gertz M.A., Blood E., Vesole D.H. et al. (2004) A multicenter phase 2 trial of stem cell transplantation for immunoglobulin light-chain amyloidosis (E4A97): An Eastern Cooperative Oncology Group Study. Bone Marrow Transplant., 34: 149-154.
- Hemery Y., Broustet H., Guiraude O. et al. (2000) Alcohol and rhythm disorders. Ann. Cardiol. Angeiol., 49: 473-479.
- Huss J.M., Kelly D.P. (2005) Mitochondrial energy metabolism in heart failure: a question of balance. J. Clin. Invest., 115: 547-555.
- Ingwall J.S., Weiss R.G. (2004) Is the failing heart energy starved? On using chemical energy to support cardiac function. Circ. Res., 95: 135-145.
- Katz A.M. (2006) Physiology of the heart. 4th ed. Williams&Wilkins, Lippincot, 644 p.
- Kholova I., Niessen H.W. (2005) Amyloid in the cardiovascular system: a review. J. Clin. Pathol., 58: 125-133.
- Maceira A.M., Joshi J., Prasad S.K. et al. (2005) Cardiovascular magnetic resonance in cardiac amyloidosis. Circulation, 111: 186-193.
- Matias I., Gonthier M.P., Orlando P. et al. (2006) Regulation, function, and dysregulation of endocannabinoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia. J. Clin. Endocrinol. Metab., 91: 3171-3180.
- Miller S.R., SekijimaY., Kelly J.W. (2004) Native state stabilization by NSAIDs inhibits transthyretin amyloidogenesis from the most common familial disease variants. Lab. Invest., 84: 545-552.
- Murtagh B., Hammill S.C., Gertz M.A. et al. (2005) Electrocardiographic findings in primary systemic amyloidosis and biopsy-proven cardiac involvement. Amer. J. Cardiol., 95: 535-537.
- Poirier P., Giles T.D., Bray G.A. et al. (2006) Obesity and cardiovascular disease: pathophysiology, evaluation and effect of weight loss. Circulation,113: 898-918.
- Rajkumar S.V., Dispenzieri A., Kyle R.A. (2006) Monoclonal gammopathy of undetermined significance, Waldenstrom macroglobulinemia, AL amyloidosis, and related plasma cell disorders: Diagnosis and treatment. Mayo Clin. Proc., 81: 693-703.
- Rutter M.K., Parise H., Benjamin E.J. et al. (2003) Impact of glucose intolerance and insulin resistance on cardiac structure and function: sex-related differences in the Framingham Heart Study. Circulation, 107: 448-454.
- Topol E.J. (Ed.) (2007) Textbook of cardiovascular medicine. 3th ed. Williams& Wilkins, Lippincott, 1628 p.
- Torp-Pedersen C., Caterson I., Coutinho W. et al. (2007) Cardiovascular responses to weight management and sibutramine in high-risk subjects: an analysis from the SCOUT trial. Eur. Heart J., 28: 2915-2923.
- Trayhurn P., Wood I.S. (2004) Adipokines: inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr., 92: 347-355.
Метаболизм — это… Что такое Метаболизм?
Метаболи́зм (от греч. μεταβολή — «превращение, изменение»), или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты.
Обмен веществ происходит между клетками организма и межклеточной жидкостью, постоянство состава которой поддерживается кровообращением: за время прохождения крови в капиллярах через проницаемые стенки капилляров плазма крови 40 раз полностью обновляется с интерстициальной жидкостью. Серии химических реакций обмена веществ называют метаболическими путями, в них при участии ферментов одни биологически значимые молекулы последовательно превращаются в другие. Ферменты играют важную роль в метаболических процессах потому, что:
- действуют как биологические катализаторы и снижают энергию активации химической реакции;
- позволяют регулировать метаболические пути в ответ на изменения среды клетки или сигналы от других клеток.
Особенности метаболизма влияют на то, будет ли пригодна определенная молекула для использования организмом в качестве источника энергии. Так, например, некоторые прокариоты используют сероводород в качестве источника энергии, однако этот газ ядовит для животных.[1] Скорость обмена веществ также влияет на количество пищи, необходимой для организма.
Основные метаболические пути и их компоненты одинаковы для многих видов, что свидетельствует о единстве происхождения всех живых существ.[2] Например, некоторые карбоновые кислоты, являющиеся интермедиатами цикла трикарбоновых кислот присутствуют во всех организмах, начиная от бактерий и заканчивая многоклеточными организмами эукариот.[3] Сходства в обмене веществ, вероятно, связаны с высокой эффективностью метаболических путей, а также с их ранним появлением в истории эволюции.[4][5]
Биологические молекулы
Органические вещества, входящие в состав всех живых существ (животных, растений, грибов и микроорганизмов), представлены в основном аминокислотами, углеводами, липидами (часто называемые жирами) и нуклеиновыми кислотами. Так как эти молекулы имеют важное значение для жизни, метаболические реакции сосредоточены на создании этих молекул при строительстве клеток и тканей или разрушении их с целью использования в качестве источника энергии. Многие важные биохимические реакции объединяются вместе для синтеза ДНК и белков.
Аминокислоты и белки
Белки являются линейными биополимерами и состоят из остатков аминокислот, соединённых пептидными связями. Некоторые белки являются ферментами и катализируют химические реакции. Другие белки выполняют структурную или механическую функцию (например, образуют цитоскелет).[6] Белки также играют важную роль в передаче сигнала в клетках, иммунных реакциях, агрегации клеток, активном транспорте через мембраны и регуляции клеточного цикла.[7]
Липиды
Липиды входят в состав биологических мембран, например, плазматических мембран, являются компонентами коферментов и источниками энергии.[7] Липиды являются гидрофобными или амфифильными биологическими молекулами, растворимыми в органических растворителях таких, как бензол или хлороформ.[8]Жиры — большая группа соединений, в состав которых входят жирные кислоты и глицерин. Молекула трёхатомного спирта глицерина, образующая три сложные эфирные связи с тремя молекулами жирных кислот, называется триглицеридом.[9] Наряду с остатками жирных кислот, в состав сложных липидов может входить, например, сфингозин (сфинголипиды), гидрофильные группы фосфатов (в фосфолипидах). Стероиды, например холестерол, представляют собой ещё один большой класс липидов.[10]
Углеводы
Сахара могут существовать в кольцевой или линейной форме в виде альдегидов или кетонов, имеют несколько гидроксильных групп. Углеводы являются наиболее распространёнными биологическими молекулами. Углеводы выполняют следующие функции: хранение и транспортировка энергии (крахмал, гликоген), структурная (целлюлоза растений, хитин у животных).[7] Наиболее распространенными мономерами сахаров являются гексозы — глюкоза, фруктоза и галактоза. Моносахариды входят в состав более сложных линейных или разветвленных полисахаридов.[11]
Нуклеотиды
Полимерные молекулы ДНК и РНК представляют собой длинные неразветвленные цепочки нуклеотидов. Нуклеиновые кислоты выполняют функцию хранения и реализации генетической информации, которые осуществляются в ходе процессов репликации,транскрипции, трансляции, и биосинтеза белка.[7] Информация, закодированная в нуклеиновых кислотах, защищается от изменений системами репарации и мультиплицируется при помощи репликации ДНК.
Некоторые вирусы имеют РНК-содержащий геном. Например, вирус иммунодефицита человека использует обратную транскрипцию для создания матрицы ДНК из собственного РНК-содержащего генома.[12] Некоторые молекулы РНК обладают каталитическими свойствами (рибозимы) и входят в состав сплайсосом и рибосом.
Нуклеозиды — продукты присоединения азотистых оснований к сахару рибозе. Примерами азотистых оснований являются гетероциклические азотсодержащие соединения — производные пуринов и пиримидинов. Некоторые нуклеотиды также выступают в качестве коферментов в реакциях переноса функциональных групп.[13]
Коферменты
- Подробное рассмотрение темы: Коферменты
Метаболизм включает широкий спектр химических реакций, большинство из которых относятся к нескольким основным типам реакций переноса функциональных групп.[14] Для переноса функциональных групп между ферментами, катализирующими химические реакции, используются коферменты.[13] Каждый класс химических реакций переноса функциональных групп катализируется отдельными ферментами и их кофакторами.[15]
Аденозинтрифосфат (АТФ) — один из центральных коферментов, универсальный источник энергии клеток. Этот нуклеотид используется для передачи химической энергии, запасенной в макроэргических связях между различными химическими реакциями. В клетках существует небольшое количество АТФ, который постоянно регенерируется из ADP и AMP. Организм человека за сутки расходует массу АТФ, равную массе собственного тела.[15] АТР выступает в качестве связующего звена между катаболизмом и анаболизмом: при катаболических реакциях образуется АТФ, при анаболических — энергия потребляется. АТФ также выступает донором фосфатной группы в реакциях фосфорилирования.
Витамины — низкомолекулярные органические вещества, необходимые в небольших количествах, причём, например, у человека большинство витаминов не синтезируется, а получается с пищей или через микрофлору КТ. В организме человека большинство витаминов являются кофакторами ферментов. Большинство витаминов приобретают биологическую активность в измененном виде, например, все водорастворимые витамины в клетках фосфорилируются или соединяются с нуклеотидами.[16]Никотинамидадениндинуклеотид (NADH) является производным витамина B3 (ниацина), и представляет собой важный кофермент — акцептора водорода. Сотни различных ферментов дегидрогеназ отнимают электроны из молекул субстратов и переносят их на молекулы NAD+, восстанавливая его до NADH. Окисленная форма кофермента является субстратом для различных редуктаз в клетке.[17] NAD в клетке существует в двух связанных формах NADH и NADPH. NAD+/NADH больше важен для протекания катаболических реакций, а NADP+/NADPH чаще используется в анаболических реакциях.
Структура гемоглобина. Белковые субъединицы окрашены красным и синим, а железосодержащий гем — зелёным. Из PDB 1GZX.Минералы и кофакторы
Неорганические элементы играют важнейшую роль в обмене веществ. Около 99 % массы млекопитающего состоит из углерода, азота, кальция, натрия, магния, хлора, калия, водорода, фосфора, кислорода и серы.[18] Биологически значимые органические соединения (белки, жиры, углеводы и нуклеиновые кислоты) содержат большое количество углерода, водорода, кислорода, азота и фосфора.[18]
Многие неорганические соединения являются ионными электролитами. Наиболее важны для организма ионы натрия, калия, кальция, магния, хлоридов, фосфатов и гидрокарбонатов. Баланс этих ионов внутри клетки во внеклеточной среде определяет осмотическое давление и рН.[19] Концентрации ионов также играют важную роль для функционирования нервных и мышечных клеток. Потенциал действия в возбудимых тканях возникает при обмене ионами между внеклеточной жидкостью и цитоплазмой.[20] Электролиты входят и выходят из клетки через ионные каналы в плазматической мембране. Например, в ходе мышечного сокращения в плазматической мембране, цитоплазме и Т-трубочках перемещаются ионы кальция, натрия и калия.[21]
Переходные металлы в организме являются микроэлементами, наиболее распространены цинк и железо.[22][23] Эти металлы используются некоторыми белками (например, ферментами в качестве кофакторов) и имеют важное значение для регуляции активности ферментов и транспортных белков.[24] Кофакторы ферментов обычно прочно связаны со специфическим белком, однако могут модифицироваться в процессе катализа, при этом после окончания катализа всегда возвращаются к своему первоначальному состоянию (не расходуются). Металлы-микроэлементы усваиваются организмом при помощи специальных транспортных белков и не встречаются в организме в свободном состоянии, так как связаны со специфическими белками-переносчиками (например, ферритином или металлотионеинами).[25][26]
Катаболизм
Катаболизмом называют метаболические процессы, при которых расщепляются относительно крупные органические молекулы сахаров, жиров, аминокислот. В ходе катаболизма образуются более простые органические молекулы, необходимые для реакций анаболизма (биосинтеза). Часто, именно в ходе реакций катаболизма организм мобилизует энергию, переводя энергию химических связей органических молекул, полученных в процессе переваривания пищи, в доступные формы: в виде АТФ, восстановленных коферментов и трансмембранного электрохимического потенциала. Термин катаболизм не является синонимом «энергетического обмена»: у многих организмов (например, у фототрофов) основные процессы запасания энергии не связаны напрямую с расщеплением органических молекул. Классификация организмов по типу метаболизма может быть основана на источнике получения энергии и углерода, что отражено в таблице ниже. Органические молекулы используются в качестве источника энергии органотрофами, литотрофы используют неорганические субстраты, а фототрофы потребляют энергию солнечного света. Однако, все эти различные формы обмена веществ зависят от окислительно-восстановительных реакций, которые связаны с передачей электронов от восстановленных доноров молекул, таких как органические молекулы, вода, аммиак, сероводород, на акцепторные молекулы, такие как кислород, нитраты или сульфат.[27] У животных эти реакции сопряжены с расщеплением сложных органических молекул до более простых, таких как двуокись углерода и воду. В фотосинтезирующих организмах — растениях и цианобактериях — реакции переноса электрона не высвобождают энергию, но они используются как способ запасания энергии, поглощаемой из солнечного света.[28]
Катаболизм у животных может быть разделён на три основных этапа. Во-первых, крупные органические молекулы, такие как белки, полисахариды и липиды расщепляются до более мелких компонентов вне клеток. Далее эти небольшие молекулы попадают в клетки и превращается в ещё более мелкие молекулы, например, ацетил-КоА. В свою очередь, ацетильная группа кофермента А окисляется до воды и углекислого газа в цикле Кребса и дыхательной цепи, высвобождая при этом энергию, которая запасается в форме АТР.
Пищеварение
- Подробное рассмотрение темы: Пищеварение и Желудочно-кишечный тракт
Такие макромолекулы, как крахмал, целлюлоза или белки, должны расщепляться до более мелких единиц прежде, чем они могут быть использованы клетками. Несколько классов ферментов принимают участие в деградации: протеазы, которые расщепляют белки до пептидов и аминокислот, гликозидазы, которые расщепляют полисахариды до олиго- и моносахаридов.
Микроорганизмы выделяют гидролитические ферменты в пространство вокруг себя,[29][30] чем отличаются от животных, которые выделяют такие ферменты только из специализированных железистых клеток.[31] Аминокислоты и моносахариды, образующиеся в результате активности внеклеточных ферментов, затем поступают в клетки с помощью активного транспорта.[32][33]
Получение энергии
- Подробное рассмотрение темы: Клеточное дыхание, Брожение, Липолиз
В ходе катаболизма углеводов сложные сахара расщепляются до моносахаридов, которые усваиваются клетками.[34] Попав внутрь, сахара (например, глюкоза и фруктоза) в процессе гликолиза превращаются в пируват, при этом вырабатывается некоторое количество АТР.[35] Пировиноградная кислота (пируват) является промежуточным продуктом в нескольких метаболических путях. Основной путь метаболизма пирувата — превращаение в ацетил-КоА и далее поступление в цикл трикарбоновых кислот. При этом в цикле Кребса в форме АТР запасается часть энергии, а также восстанавливаются молекулы NADH и FAD. В процессе гликолиза и цикла трикарбоновых кислот образуется диоксид углерода, который является побочным продуктом жизнедеятельности. В анаэробных условиях в результате гликолиза из пирувата при участии фермента лактатдегидрогеназы образуется лактат, и происходит окисление NADH до NAD+, который повторно используется в реакциях гликолиза. Существует также альтернативный путь метаболизма моносахаридов — пентозофосфатный путь, в ходе реакций которого энергия запасается в форме восстановленного кофермента NADPH и образуются пентозы, например, рибоза, необходимая для синтеза нуклеиновых кислот.
Жиры на первом этапе катаболизма гидролизуются в свободные жирные кислоты и глицерин. Жирные кислоты расщепляются в процессе бета-окисления с образованием ацетил-КоА, который в свою очередь далее катаболизируется в цикле Кребса, либо идет на синтез новых жирных кислот. Жирные кислоты выделяют больше энергии, чем углеводы, так как жиры содержат удельно больше атомов водорода в своей структуре.
Аминокислоты либо используются для синтеза белков и других биомолекул, либо окисляются до мочевины, диоксида углерода и служат источником энергии.[36] Окислительный путь катаболизма аминокислот начинается с удаления аминогруппы ферментами трансаминазами. Аминогруппы утилизируются в цикле мочевины; аминокислоты, лишённые аминогрупп называют кетокислотами. Некоторые кетокислоты — промежуточные продукты цикла Кребса. Например, при дезаминировании глутамата образуется альфа-кетоглутаровая кислота.[37] Гликогенные аминокислоты также могут быть преобразованы в глюкозу в реакциях глюконеогенеза.[38]
Энергетические превращения
Окислительное фосфорилирование
- Подробное рассмотрение темы: Окислительное фосфорилирование, Хемиосмос и Митохондрия
При окислительном фосфорилировании электроны, удалённые из пищевых молекул в метаболических путях (например, в цикле Кребса), переносятся на кислород, а выделяющаяся энергия используется для синтеза АТР. У эукариот данный процесс осуществляется при участии ряда белков, закреплённых в мембранах митохондрий, называемые дыхательной цепью переноса электронов. У прокариот эти белки присутствуют во внутренней мембране клеточной стенки.[39] Белки цепи переноса электронов используют энергию, полученную при передаче электронов от восстановленных молекул (например NADH) на кислород, для перекачки протонов через мембрану.[40]
При перекачке протонов создаётся разница концентраций ионов водорода и возникает электрохимический градиент.[41] Эта сила возвращает протоны обратно в митохондрии через основание АТР-синтазы. Поток протонов заставляет вращаться кольцо из c-субъединиц фермента, в результате чего активный центр синтазы изменяет форму и фосфорилирует аденозиндифосфат, превращая его в АТР.[15]
Энергия из неорганических соединений
Хемолитотрофами называют прокариот, имеющих особый тип обмена веществ, при котором энергия образуется в результате окисления неорганических соединений. Хемолитотрофы могут окислять молекулярный водород,[42] соединения серы (например, сульфиды, сероводород и тиосульфат),[1]оксид железа(II)[43] или аммиак.[44] При этом энергия от окисления этих соединений образуется с помощью акцепторов электронов, таких как кислород или нитриты.[45] Процессы получения энергии из неорганических веществ играют важную роль в таких биогеохимических циклах, как ацетогенез, нитрификация и денитрификация.[46][47]
Энергия из солнечного света
Энергия солнечного света поглощается растениями, цианобактериями, пурпурными бактериями, зелёными серными бактериями и некоторыми простейшими. Этот процесс часто сочетается с превращением диоксида углерода в органические соединения, как часть процесса фотосинтеза (см. ниже). Системы захвата энергии и фиксации углерода у некоторых прокариот могут работать раздельно (например, у пурпурных и зелёных серных бактерий).[48][49]
У многих организмов поглощение солнечной энергии в принципе аналогично окислительному фосфорилированию, так как при этом энергия запасается в форме градиента концентрации протонов и движущая сила протонов приводит к синтезу АТР.[15] Электроны, необходимые для этой цепи переноса, поступают от светособирающих белков, называемых центрами фотосинтетических реакций (примером являются родопсины). В зависимости от вида фотосинтетических пигментов классифицируют два типа центров реакций; в настоящее время большинство фотосинтезирующих бактерий имеют только один тип, в то время как растения и цианобактерии два.[50]
У растений, водорослей и цианобактерий, фотосистема II использует энергию света для удаления электронов из воды, при этом молекулярный кислород выделяется как побочный продукт реакции. Электроны затем поступают в комплекс цитохрома b6f, который использует энергию для перекачки протонов через тилакоидную мембрану в хлоропластах.[7] Под действием электрохимического градиента протоны движутся обратно через мембрану и запускают АТР-синтазу. Электроны затем проходят через фотосистему I и могут быть использованы для окисления кофермента NADP+, для использования в цикле Кальвина или рециркуляции для образования дополнительных молекул АТР.[51]
Анаболизм
- Подробное рассмотрение темы: Анаболизм
Анаболизм — совокупность метаболических процессов биосинтеза сложных молекул с затратой энергии. Сложные молекулы, входящие в состав клеточных структур, синтезируются последовательно из более простых предшественников. Анаболизм включает три основных этапа, каждый из которых катализируется специализированным ферментом. На первом этапе синтезируются молекулы-предшественники, например, аминокислоты, моносахариды, терпеноиды и нуклеотиды. На втором этапе предшественники с затратой энергии АТР преобразуются в активированные формы. На третьем этапе активированные мономеры объединяются в более сложные молекулы, например, белки, полисахариды, липиды и нуклеиновые кислоты.
Не все живые организмы могут синтезировать все биологически активные молекулы. Автотрофы (например, растения) могут синтезировать сложные органические молекулы из таких простых неорганических низкомолекулярных веществ, как углекислый газ и вода. Гетеротрофам необходим источник более сложных веществ, таких как моносахариды и аминокислоты, для создания более сложных молекул. Организмы классифицируют по их основным источникам энергии: фотоавтотрофы и фотогетеротрофы получают энергию из солнечного света, в то время как хемоавтотрофы и хемогетеротрофы получают энергию из неорганических реакций окисления.
Связывание углерода
- Подробное рассмотрение темы: Фотосинтез и Хемосинтез
Фотосинтезом называют процесс биосинтеза сахаров из углекислого газа, при котором необходимая энергия поглощается из солнечного света. У растений, цианобактерий и водорослей, при кислородном фотосинтезе происходит фотолиз воды, при этом, как побочный продукт, выделяется кислород. Для преобразования CO2 в 3-фосфоглицерат используется энергия АТР и NADPH, запасенная в фотосистемах. Реакция связывания углерода осуществляется с помощью фермента рибулозобисфосфаткарбоксилазы и является частью цикла Кальвина.[52] У растений классифицируют три типа фотосинтеза — по пути трехуглеродых молекул, по пути четырехуглеродых молекул (С4), и CAM фотосинтез. Три типа фотосинтеза отличаются по пути связывания углекислого газа и его вхождения в цикл Кальвина; у C3 растений связывание CO2 происходит непосредственно в цикле Кальвина, а при С4 и CAM CO2 предварительно включается в состав других соединений. Разные формы фотосинтеза являются приспособлениями к к интенсивному потоку солнечных лучей и к сухим условиям.[53]
У фотосинтезирующих прокариот механизмы связывания углерода более разнообразны. Углекислый газ может быть фиксирован в цикле Кальвина, в обратном цикле Кребса,[54] или в реакциях карбоксилирования ацетил-КоА.[55][56] Прокариоты — хемоавтотрофы также связывают CO2 через цикл Кальвина, но для протекания реакции используют энергию из неорганических соединений.[57]
Углеводы и гликаны
- Подробное рассмотрение темы: Глюконеогенез и Гликозилирование
В процессе анаболизма сахаров простые органические кислоты могут быть преобразованы в моносахариды, например, в глюкозу, и затем использованы для синтеза полисахаридов, таких как крахмал. Образование глюкозы из соединений, как пируват, лактат, глицерин, 3-фосфоглицерат и аминокислот называют глюконеогенезом. В процессе глюконеогенеза пируват превращается глюкозо-6-фосфат через ряд промежуточных соединений, многие из которых образуются и при гликолизе.[35] Однако, глюконеогенез не просто является гликолизом в обратном направлении, так как несколько химических реакций катализируют специальные ферменты, что дает возможность независимо регулировать процессы образования и распада глюкозы.[58][59]
Многие организмы запасают питательные вещества в форме липидов и жиров, однако, позвоночные не имеют ферментов, катализирующих превращение ацетил-КоА (продукта метаболизма жирных кислот) в пируват (субстрат глюконеогенеза).[60] После длительного голодания позвоночные начинают синтезировать кетоновые тела из жирных кислот, которые могут заменять глюкозу в таких тканях, как головной мозг.[61] У растений и бактерий, данная метаболическая проблема решается использованием глиоксилатного цикла, который обходит этап декарбоксилирования в цикле лимонной кислоты и позволяет превращать ацетил-КоА в оксалоацетат, и далее использовать для синтеза глюкозы.[60][62]
Полисахариды выполняют структурные и метаболические функции, а также могут быть соединены с липидами (гликолипиды) и белками (гликопротеиды) при помощи ферментов олигосахаридтрансфераз.[63][64]
Жирные кислоты, изопреноиды и стероиды
- Подробное рассмотрение темы: Стероиды
Жирные кислоты образуются синтазами жирных кислот из ацетил-КоА. Углеродный скелет жирных кислот удлиняется в цикле реакций, в которых сначала присоединяется ацетильная группа, далее карбонильная группа восстанавливается до гидроксильной, затем происходит дегидратация и последующее восстановление. Ферменты биосинтеза жирных кислот классифицируют на две группы: у животных и грибов все реакции синтеза жирных кислот осуществляются одним многофункциональным белком I типа,[65] в пластидах растений и у бактерий каждый этап катализируют отдельные ферменты II типа.[66][67]
Терпены и терпеноиды являются представителями самого многочисленного класса растительных натуральных продуктов.[68] Представители данной группы веществ являются производными изопрена и образуются из активированных предшественников изопентилпирофосфата и диметилаллилпирофосфата, которые, в свою очередь, образуются в разных реакциях обмена веществ.[69] У животных и архей изопентилпирофосфат и диметилаллилпирофосфат синтезируются из ацетил-КоА в мевалонатном пути,[70] в то время как у растений и бактерий субстратами не-мевалонатного пути являются пируват и глицеральдегид-3-фосфат.[69][71] В реакциях биосинтеза стероидов молекулы изопрена объединяются и образуют сквалены, которые далее формируют циклические структуры с образованием ланостерола.[72] Ланостерол может быть преобразован в другие стероиды, например холестерин и эргостерин.[72][73]
Белки
- Подробное рассмотрение темы: Биосинтез белка
Организмы различаются по способности к синтезу 20 общих аминокислот. Большинство бактерий и растений могут синтезировать все 20, но млекопитающие способны синтезировать лишь 11 заменимых аминокислот.[7] Таким образом, в случае млекопитающих 9 незаменимых аминокислот должны быть получены из пищи. Все аминокислоты синтезируются из промежуточных продуктов гликолиза, цикла лимонной кислоты или пентозомонофосфатного пути. Перенос аминогрупп с аминокислот на альфа-кетокислоты называется трансаминированием. Донорами аминогрупп являются глутамат и глутамин. [74]
Аминокислоты, соединенными пептидными связями, образуют белки. Каждый белок имеет уникальную последовательность аминокислотных остатков (первичная структура белка). Подобно тому, как буквы алфавита могут комбинироваться с образованием почти бесконечных вариаций слов, аминокислоты могут связываться в той или иной последовательности и формировать разнообразные белки. Фермент Аминоацил-тРНК-синтетаза катализирует АТР-зависимое присоединение аминокислот к тРНК сложноэфирными связями, при этом образуются аминоацил-тРНК.[75] Аминоацил-тРНК являются субстратами для рибосом, которая объединяют аминокислоты в длинные полипептидные цепочки, используя матрицу мРНК.[76]
Нуклеотиды
- Подробное рассмотрение темы: Пурин, пиримидин
Нуклеотиды образуются из аминокислот, углекислого газа и муравьиной кислоты в цепи реакций, для протекания которых требуется большое количество энергии.[77][78] Именно поэтому большинство организмов имеют эффективные системы сохранения ранее синтезированных нуклеотидов и азотистых оснований.[77][79]Пурины синтезируются как нуклеозиды (в основном связанные с рибозой). Аденин и гуанин образуются из инозин-монофосфата, который синтезируется из глицина, глутамина и аспартата при участии метенил-тетрагидрофолата. Пиримидины синтезируются из оротата, который образуется из глутамина и аспартата.[80]
Ксенобиотики и окислительный метаболизм
- Подробное рассмотрение темы: Антиоксиданты
Все организмы постоянно подвергаются воздействию соединений, накопление которых может быть вредно для клеток. Такие потенциально опасные чужеродные соединения называются ксенобиотиками.[81] Ксенобиотики, например синтетические лекарства и яды природного происхождения, детоксифицируются специализированными ферментами. У человека такие ферменты представлены, например, цитохром-оксидазами,[82] глюкуронилтрансферазой,[83] и глутатион S-трансферазой.[84] Эта система ферментов действует в три этапа: на первой стадии ксенобиотики окисляются, затем происходит конъюгирование водорастворимых групп в молекулы, далее модифицированные водорастворимые ксенобиотики могут быть удалены из клеток и метаболизированы перед их экскрецией. Описанные реакции играют важную роль в разложении микробами загрязняющих веществ и биоремедиации загрязнённых земель и разливов нефти.[85] Многие подобные реакции протекают при участии многоклеточных организмов, однако, ввиду невероятного разнообразия, микроорганизмы справляются с гораздо более широким спектром ксенобиотиков, чем многоклеточные организмы, и способны даже разрушать стойкие органические загрязнители, например хлорорганические соединения.[86]
Связанной с этим проблемой для аэробных организмов является оксидативный стресс.[87] В процессе окислительного фосфорилирования и образования дисульфидных связей при укладке белка образуются активные формы кислорода, например пероксид водорода.[88] Эти повреждающие оксиданты удаляются антиоксидантами, например глутатионом и ферментами каталазой и пероксидазами.[89][90]
Термодинамика живых организмов
Живые организмы подчиняются началам термодинамики, которые описывают превращения тепла и работы. Второе начало термодинамики гласит, что в любой изолированной системе энтропия не уменьшается. Хотя невероятная сложность живых организмов очевидно противоречит этому закону, жизнь возможна, так как все организмы открытые системы, которые обмениваются веществом и энергией с окружающей средой. Таким образом живые системы не находятся в термодинамическом равновесии, но вместо этого выступает диссипативной системой, которая поддерживают своё состояние сложно организованности, вызывая большее увеличение энтропии окружающей средой.[91] В метаболизм клеток это достигается путём сочетания спонтанных процессов катаболизма с не спонтанных процессов анаболизма. В термодинамических условиях, метаболизм поддерживает порядок за счёт создания беспорядка.[92]
Регуляция и контроль
- Подробное рассмотрение темы: Гормоны, Передача сигнала в клетке
Гомеостазом называют постоянство внутренней среды организма. Так как внешняя среда, окружающая большинство организмов, постоянно меняется, для поддержания постоянных условий внутри клеток, реакции обмена веществ должны точно регулироваться.[93][94] Регуляция метаболизма позволяет организмам отвечать на сигналы и активно взаимодействовать с окружающей средой.[95] В случае фермента, регуляция заключается в повышении и снижении его активности в ответ на сигналы. С другой стороны, фермент оказывает некоторый контроль над метаболическим путем, который определяется как эффект от изменения активности фермента на данный метаболический путь.[96]
Влияние инсулина на поглощение глюкозы и обмен веществ. Инсулин связывается со своим рецептором (1), который в свою очередь запускает касакад реакций активации множества белков (2). К ним относятся: транслокация переносчика GLUT-4 к плазматической мембране и поступление глюкозы в клетку (3), синтез гликогена (4), гликолиз (5) и синтез жирных кислот (6).Выделяют несколько уровней регуляции метаболизма. В метаболическом пути происходит саморегуляция на уровне субстрата или продукта; например, уменьшение количества продукта может компенсированно увеличить поток субстрата реакции по данному пути.[97] Этот тип регулирования часто включает аллостерическое регулирование активности некоторых ферментов в метаболических путях.[98] Внешний контроль включает клетку многоклеточного организма, изменяющую свой метаболизм в ответ на сигналы от других клеток. Эти сигналы, как правило, в виде растворимых мессенджеров, например гормоны и факторы роста, определяются специфическими рецепторами на поверхности клеток.[99] Затем эти сигналы передаются внутрь клетки системой вторичных мессенджеров, которые зачастую связаны с фосфорилированием белков.[100]
Хорошо изученный пример внешнего контроля — регуляция метаболизма глюкозы инсулином.[101] Инсулин вырабатывается в ответ на повышение уровня глюкозы в крови. Гормон связывается с инсулиновым рецептором на поверхности клетки, затем активируется каскад протеинкиназ, которые обеспечивают поглощение молекул глюкозы клетками и преобразовать их в молекулы жирных кислот и гликогена.[102] Метаболизм гликогена контролируется активностью фосфорилазы (фермента, который расщепляет гликоген) и гликогенсинтазы (фермента, который образует его). Эти ферменты взаимосвязаны; фосфорилирование ингибируется гликогенсинтазой, но активируется фосфорилазой. Инсулин вызывает синтез гликогена путём активации белковых фосфатаз и уменьшает фосфорилирование этих ферментов.[103]
Эволюция
- Подробное рассмотрение темы: Филогенетика
Главные пути метаболизма, описанные выше, например, гликолиза и цикла Кребса, присутствуют у всех трёх доменах живых существ и обнаруживаются у последнего универсального общего предка.[3][104] Этот универсальный предок был прокариотом и, вероятно, метаногеном с аминокислотным, нуклеотидным, углеводным и липидным метаболизмом.[105][106] Сохранение этих древних метаболических путей в эволюции может быть результатом того, что эти реакции оптимальны для решения конкретных проблем с метаболизмом. Так, конечные продукты гликолиза и цикла Кребса образуются с высокой эффективностью и с минимальным количеством стадий.[4][5] Первые метаболические пути на основе ферментов могли быть частями пуринового метаболизма нуклеотидов с предыдущим метаболических путей были частью древнего мира РНК.[107]
Многие модели были предложены для описания механизмов, посредством которых новые метаболические пути эволюционировали. К ним относятся последовательное добавление новых ферментов на короткий предковый путь, дупликация, а затем дивергенция всех путей, а также набор уже существующих ферментов и их сборка в новый путь реакций.[108] Относительную важность этих механизмов неясна, однако геномные исследования показали, что ферменты в метаболическом пути, скорее всего, имеют общее происхождение, предполагая, что многие пути эволюционировали шаг за шагом с новыми функциями, созданными из уже существующих этапов пути.[109] Альтернативная модель основана на исследованиях, в которых прослеживается эволюция структуры белков в метаболических связях; предполагают, что ферменты собирались для выполнения схожих функций в различных метаболических путях[110] Эти процессы сборки привели к эволюционированию ферментативной мозаики.[111] Некоторые части обмена веществ возможно существовали в качестве «модулей», которые могли быть повторно использованы в различных путях для выполнения схожих функций.[112]
Эволюция также может приводить к потере метаболических функций. Например, у некоторых паразитов метаболические процессы, которые не важны для выживания, утрачены и готовые аминокислоты, нуклеотиды и углеводы получаются от хозяина.[113] Подобные упрощения метаболических возможностей наблюдают у эндосимбиотических организмов.[114]
Методы исследования
- Подробное рассмотрение темы: Протеомика, Метабономика
Классически, метаболизм изучается упрощённым подходом, который фокусируется на одном метаболическом пути. Особенно ценно использование меченых атомов на организменном, тканевом и клеточном уровнях, которые определяют пути от предшественников до конечных продуктов путём выявления радиоактивно меченых промежуточных продуктов.[115] Ферменты, которые катализируют эти химические реакции, могут затем быть выделены для исследования их кинетики и ответа на ингибиторы. Параллельный подход заключается в выявлении небольших молекул в клетки или ткани; полный набор этих молекул называется метаболом. В целом, эти исследования дают хорошее представление о структуре и функциях простых путей метаболизма, но недостаточны в применении к более сложных системам, например полной метаболизм клетки.[116]
Идея сложности метаболических сетей в клетках, которые содержат тысячи различных ферментов, отражена на изображении справа, показывающее взаимодействия только между 43 белками и 40 метаболитами, которые регулируются 45000 генов.[117] Тем не менее, сейчас можно использовать такие данные о геномах для воссоздания полной сети биохимических реакций и образовывать более целостные математические модели, которые могут объяснить и предсказать их поведение.[118] Эти модели особенно сильны, когда используются для интеграции данных о путях и метаболитах, полученных на основе классических методов, с данными по экспрессии генов из протеомных и ДНК-микрочиповых исследований.[119] С помощью этих методов, модель человеческого метаболизма в настоящее время создаётся, которая будет служить ориентиром для будущих исследований лекарств и биохимических исследований.[120] Эти модели в настоящее время используются в анализах сети, для классификации болезней человека по группам, которые различаются по общим белкам или метаболитам.[121][122]
Яркий пример бактериальных метаболических сетей — устройство галстук-бабочки[123][124][125], структура которой позволяет вводить широкий спектр питательных веществ и производить большое разнообразие продуктов и сложных макромолекул, используя сравнительно немного общих промежуточных веществ.
Основная технологическая основа этой информации — метаболическая инженерия. Здесь организмы, например дрожжи, растения или бактерии, генетически модифицируются, чтобы сделать их более эффективными в биотехнологии и помочь в производстве лекарств, например антибиотиков или промышленных химических веществ, таких как 1,3-пропандиола и шикимовой кислоты.[126] Эти генетические модификации обычно направлены на уменьшение количества энергии, используемой для производства продукции, повышения урожайности и снижения производственных отходов.[127]
История
Санторио взвешивает сам себя до и после принятия пищи, из Ars de statica medicina, впервые опубликованной в 1614 годуИстория изучения метаболизма охватывает несколько столетий. Исследования начинались с изучения организмов животных, в современной биохимии изучают отдельные метаболические реакции. Понятие обмена веществ впервые встречается в работах Ибн аль-Нафиса (1213—1288), который писал, что «тело и его части находятся в постоянном состоянии распада и питания, так что оно неизбежно претерпевает постоянные изменения».[128] Первые контролируемые эксперименты по метаболизму у человека были опубликованы Санторио Санторио в 1614 году в книге итал. Ars de statica medicina.[129] Он рассказал, как он сам взвесил себя до и после приёма пищи, сна, работы, секса, натощак, после питья и выделения мочи. Он обнаружил, что большая часть пищи, которую он принял, была утрачена в результате процесса, названного «незаметным испарением».
В ранних исследованиях механизмы метаболических реакций не были обнаружены и считалось, что живой тканью управляет живая сила.[130] В XIX веке при исследовании ферментации сахара спирта дрожжами Луи Пастер сделал вывод, что брожение катализируется веществами из дрожжевых клеток, которые он назвал ферментами. Пастер писал, что «алкогольное брожение — действие, связанное с жизнью и организуется дрожжевыми клетками, не связано со смертью или разложением клеток».[131] Это открытие, вместе с публикацией Фридриха Вёлера в 1828 году о химическом синтезе мочевины,[132] доказали, что органические соединения и химические реакции, обнаруженные в клетках, не имеют различий в принципе, как и любые другие разделы химии.
Открытие ферментов в начале XX века Эдуардом Бухнером разделило изучение метаболических реакций от изучения клеток и дало начало развитию биохимии как науки.[133] Одним из успешных биохимиков начала двадцатого века был Ханс Адольф Кребс, который внёс огромный вклад в изучение метаболизма.[134] Кребс описал цикл мочевины и позднее, работая вместе с Хансом Корнбергом, цикл лимонной кислоты и глиоксилатный цикл.[135][62] В современных биохимических исследованиях широко используют новые методы, такие как хроматография, рентгеноструктурный анализ, ЯМР-спектроскопия, электронная микроскопия и метод классической молекулярной динамики. Эти методы позволяют открывать и подробно изучать множество молекул и метаболических путей в клетках.
См. также
Примечания
- ↑ 1 2 Friedrich C (1998). «Physiology and genetics of sulfur-oxidizing bacteria». Adv Microb Physiol 39: 235–89. DOI:10.1016/S0065-2911(08)60018-1. PMID 9328649.
- ↑ Pace NR (January 2001). «The universal nature of biochemistry». Proc. Natl. Acad. Sci. U.S.A. 98 (3): 805–8. DOI:10.1073/pnas.98.3.805. PMID 11158550.
- ↑ 1 2 Smith E, Morowitz H (2004). «Universality in intermediary metabolism». Proc Natl Acad Sci USA 101 (36): 13168–73. DOI:10.1073/pnas.0404922101. PMID 15340153.
- ↑ 1 2 Ebenhöh O, Heinrich R (2001). «Evolutionary optimization of metabolic pathways. Theoretical reconstruction of the stoichiometry of ATP and NADH producing systems». Bull Math Biol 63 (1): 21–55. DOI:10.1006/bulm.2000.0197. PMID 11146883.
- ↑ 1 2 Meléndez-Hevia E, Waddell T, Cascante M (1996). «The puzzle of the Krebs citric acid cycle: assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution». J Mol Evol 43 (3): 293–303. DOI:10.1007/BF02338838. PMID 8703096.
- ↑ Michie K, Löwe J (2006). «Dynamic filaments of the bacterial cytoskeleton». Annu Rev Biochem 75: 467–92. DOI:10.1146/annurev.biochem.75.103004.142452. PMID 16756499.
- ↑ 1 2 3 4 5 6 Nelson David L. Lehninger Principles of Biochemistry. — New York: W. H. Freeman and company, 2005. — P. 841. — ISBN 0-7167-4339-6
- ↑ Fahy E, Subramaniam S, Brown H, Glass C, Merrill A, Murphy R, Raetz C, Russell D, Seyama Y, Shaw W, Shimizu T, Spener F, van Meer G, VanNieuwenhze M, White S, Witztum J, Dennis E (2005). «A comprehensive classification system for lipids». J Lipid Res 46 (5): 839–61. DOI:10.1194/jlr.E400004-JLR200. PMID 15722563.
- ↑ Nomenclature of Lipids. IUPAC-IUB Commission on Biochemical Nomenclature (CBN). Архивировано из первоисточника 22 августа 2011. Проверено 8 марта 2007.
- ↑ Hegardt F (1999). «Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase: a control enzyme in ketogenesis». Biochem J 338 (Pt 3): 569–82. DOI:10.1042/0264-6021:3380569. PMID 10051425.
- ↑ Raman R, Raguram S, Venkataraman G, Paulson J, Sasisekharan R (2005). «Glycomics: an integrated systems approach to structure-function relationships of glycans». Nat Methods 2 (11): 817–24. DOI:10.1038/nmeth807. PMID 16278650.
- ↑ Sierra S, Kupfer B, Kaiser R (2005). «Basics of the virology of HIV-1 and its replication». J Clin Virol 34 (4): 233–44. DOI:10.1016/j.jcv.2005.09.004. PMID 16198625.
- ↑ 1 2 Wimmer M, Rose I (1978). «Mechanisms of enzyme-catalyzed group transfer reactions». Annu Rev Biochem 47: 1031–78. DOI:10.1146/annurev.bi.47.070178.005123. PMID 354490.
- ↑ Mitchell P (1979). «The Ninth Sir Hans Krebs Lecture. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems». Eur J Biochem 95 (1): 1–20. DOI:10.1111/j.1432-1033.1979.tb12934.x. PMID 378655.
- ↑ 1 2 3 4 Dimroth P, von Ballmoos C, Meier T (March 2006). «Catalytic and mechanical cycles in F-ATP synthases. Fourth in the Cycles Review Series». EMBO Rep 7 (3): 276–82. DOI:10.1038/sj.embor.7400646. PMID 16607397.
- ↑ Stanford School of Medicine Nutrition Courses. — SUMMIT, 2006.
- ↑ Pollak N, Dölle C, Ziegler M (2007). «The power to reduce: pyridine nucleotides—small molecules with a multitude of functions». Biochem J 402 (2): 205–18. DOI:10.1042/BJ20061638. PMID 17295611.
- ↑ 1 2 Heymsfield S, Waki M, Kehayias J, Lichtman S, Dilmanian F, Kamen Y, Wang J, Pierson R (1991). «Chemical and elemental analysis of humans in vivo using improved body composition models». Am J Physiol 261 (2 Pt 1): E190–8. PMID 1872381.
- ↑ Sychrová H (2004). «Yeast as a model organism to study transport and homeostasis of alkali metal cations» (PDF). Physiol Res 53 Suppl 1: S91–8. PMID 15119939.
- ↑ Levitan I (1988). «Modulation of ion channels in neurons and other cells». Annu Rev Neurosci 11: 119–36. DOI:10.1146/annurev.ne.11.030188.001003. PMID 2452594.
- ↑ Dulhunty A (2006). «Excitation-contraction coupling from the 1950s into the new millennium». Clin Exp Pharmacol Physiol 33 (9): 763–72. DOI:10.1111/j.1440-1681.2006.04441.x. PMID 16922804.
- ↑ Mahan D, Shields R (1998). «Macro- and micromineral composition of pigs from birth to 145 kilograms of body weight». J Anim Sci 76 (2): 506–12. PMID 9498359.
- ↑ Husted S, Mikkelsen B, Jensen J, Nielsen N (2004). «Elemental fingerprint analysis of barley (Hordeum vulgare) using inductively coupled plasma mass spectrometry, isotope-ratio mass spectrometry, and multivariate statistics». Anal Bioanal Chem 378 (1): 171–82. DOI:10.1007/s00216-003-2219-0. PMID 14551660.
- ↑ Finney L, O’Halloran T (2003). «Transition metal speciation in the cell: insights from the chemistry of metal ion receptors». Science 300 (5621): 931–6. DOI:10.1126/science.1085049. PMID 12738850.
- ↑ Cousins R, Liuzzi J, Lichten L (2006). «Mammalian zinc transport, trafficking, and signals». J Biol Chem 281 (34): 24085–9. DOI:10.1074/jbc.R600011200. PMID 16793761.
- ↑ Dunn L, Rahmanto Y, Richardson D (2007). «Iron uptake and metabolism in the new millennium». Trends Cell Biol 17 (2): 93–100. DOI:10.1016/j.tcb.2006.12.003. PMID 17194590.
- ↑ Nealson K, Conrad P (1999). «Life: past, present and future». Philos Trans R Soc Lond B Biol Sci 354 (1392): 1923–39. DOI:10.1098/rstb.1999.0532. PMID 10670014.
- ↑ Nelson N, Ben-Shem A (2004). «The complex architecture of oxygenic photosynthesis». Nat Rev Mol Cell Biol 5 (12): 971–82. DOI:10.1038/nrm1525. PMID 15573135.
- ↑ Häse C, Finkelstein R (December 1993). «Bacterial extracellular zinc-containing metalloproteases». Microbiol Rev 57 (4): 823–37. PMID 8302217.
- ↑ Gupta R, Gupta N, Rathi P (2004). «Bacterial lipases: an overview of production, purification and biochemical properties». Appl Microbiol Biotechnol 64 (6): 763–81. DOI:10.1007/s00253-004-1568-8. PMID 14966663.
- ↑ Hoyle T (1997). «The digestive system: linking theory and practice». Br J Nurs 6 (22): 1285–91. PMID 9470654.
- ↑ Souba W, Pacitti A (1992). «How amino acids get into cells: mechanisms, models, menus, and mediators». JPEN J Parenter Enteral Nutr 16 (6): 569–78. DOI:10.1177/0148607192016006569. PMID 1494216.
- ↑ Barrett M, Walmsley A, Gould G (1999). «Structure and function of facilitative sugar transporters». Curr Opin Cell Biol 11 (4): 496–502. DOI:10.1016/S0955-0674(99)80072-6. PMID 10449337.
- ↑ Bell G, Burant C, Takeda J, Gould G (1993). «Structure and function of mammalian facilitative sugar transporters». J Biol Chem 268 (26): 19161–4. PMID 8366068.
- ↑ 1 2 Bouché C, Serdy S, Kahn C, Goldfine A (2004). «The cellular fate of glucose and its relevance in type 2 diabetes». Endocr Rev 25 (5): 807–30. DOI:10.1210/er.2003-0026. PMID 15466941.
- ↑ Sakami W, Harrington H (1963). «Amino acid metabolism». Annu Rev Biochem 32: 355–98. DOI:10.1146/annurev.bi.32.070163.002035. PMID 14144484.
- ↑ Brosnan J (2000). «Glutamate, at the interface between amino acid and carbohydrate metabolism». J Nutr 130 (4S Suppl): 988S–90S. PMID 10736367.
- ↑ Young V, Ajami A (2001). «Glutamine: the emperor or his clothes?». J Nutr 131 (9 Suppl): 2449S–59S; discussion 2486S–7S. PMID 11533293.
- ↑ Hosler J, Ferguson-Miller S, Mills D (2006). «Energy transduction: proton transfer through the respiratory complexes». Annu Rev Biochem 75: 165–87. DOI:10.1146/annurev.biochem.75.062003.101730. PMID 16756489.
- ↑ Schultz B, Chan S (2001). «Structures and proton-pumping strategies of mitochondrial respiratory enzymes». Annu Rev Biophys Biomol Struct 30: 23–65. DOI:10.1146/annurev.biophys.30.1.23. PMID 11340051.
- ↑ Capaldi R, Aggeler R (2002). «Mechanism of the F(1)F(0)-type ATP synthase, a biological rotary motor». Trends Biochem Sci 27 (3): 154–60. DOI:10.1016/S0968-0004(01)02051-5. PMID 11893513.
- ↑ Friedrich B, Schwartz E (1993). «Molecular biology of hydrogen utilization in aerobic chemolithotrophs». Annu Rev Microbiol 47: 351–83. DOI:10.1146/annurev.mi.47.100193.002031. PMID 8257102.
- ↑ Weber K, Achenbach L, Coates J (2006). «Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction». Nat Rev Microbiol 4 (10): 752–64. DOI:10.1038/nrmicro1490. PMID 16980937.
- ↑ Jetten M, Strous M, van de Pas-Schoonen K, Schalk J, van Dongen U, van de Graaf A, Logemann S, Muyzer G, van Loosdrecht M, Kuenen J (1998). «The anaerobic oxidation of ammonium». FEMS Microbiol Rev 22 (5): 421–37. DOI:10.1111/j.1574-6976.1998.tb00379.x. PMID 9990725.
- ↑ Simon J (2002). «Enzymology and bioenergetics of respiratory nitrite ammonification». FEMS Microbiol Rev 26 (3): 285–309. DOI:10.1111/j.1574-6976.2002.tb00616.x. PMID 12165429.
- ↑ Conrad R (1996). «Soil microorganisms as controllers of atmospheric trace gases (H2, CO, CH4, OCS, N2O, and NO)». Microbiol Rev 60 (4): 609–40. PMID 8987358.
- ↑ Barea J, Pozo M, Azcón R, Azcón-Aguilar C (2005). «Microbial co-operation in the rhizosphere». J Exp Bot 56 (417): 1761–78. DOI:10.1093/jxb/eri197. PMID 15911555.
- ↑ van der Meer M, Schouten S, Bateson M, Nübel U, Wieland A, Kühl M, de Leeuw J, Sinninghe Damsté J, Ward D (July 2005). «Diel variations in carbon metabolism by green nonsulfur-like bacteria in alkaline siliceous hot spring microbial mats from Yellowstone National Park». Appl Environ Microbiol 71 (7): 3978–86. DOI:10.1128/AEM.71.7.3978-3986.2005. PMID 16000812.
- ↑ Tichi M, Tabita F (2001). «Interactive control of Rhodobacter capsulatus redox-balancing systems during phototrophic metabolism». J Bacteriol 183 (21): 6344–54. DOI:10.1128/JB.183.21.6344-6354.2001. PMID 11591679.
- ↑ Allen J, Williams J (1998). «Photosynthetic reaction centers». FEBS Lett 438 (1–2): 5–9. DOI:10.1016/S0014-5793(98)01245-9. PMID 9821949.
- ↑ Munekage Y, Hashimoto M, Miyake C, Tomizawa K, Endo T, Tasaka M, Shikanai T (2004). «Cyclic electron flow around photosystem I is essential for photosynthesis». Nature 429 (6991): 579–82. DOI:10.1038/nature02598. PMID 15175756.
- ↑ Miziorko H, Lorimer G (1983). «Ribulose-1,5-bisphosphate carboxylase-oxygenase». Annu Rev Biochem 52: 507–35. DOI:10.1146/annurev.bi.52.070183.002451. PMID 6351728.
- ↑ Dodd A, Borland A, Haslam R, Griffiths H, Maxwell K (2002). «Crassulacean acid metabolism: plastic, fantastic». J Exp Bot 53 (369): 569–80. DOI:10.1093/jexbot/53.369.569. PMID 11886877.
- ↑ Hügler M, Wirsen C, Fuchs G, Taylor C, Sievert S (May 2005). «Evidence for autotrophic CO2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria». J Bacteriol 187 (9): 3020–7. DOI:10.1128/JB.187.9.3020-3027.2005. PMID 15838028.
- ↑ Strauss G, Fuchs G (1993). «Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle». Eur J Biochem 215 (3): 633–43. DOI:10.1111/j.1432-1033.1993.tb18074.x. PMID 8354269.
- ↑ Wood H (1991). «Life with CO or CO2 and H2 as a source of carbon and energy». FASEB J 5 (2): 156–63. PMID 1900793.
- ↑ Shively J, van Keulen G, Meijer W (1998). «Something from almost nothing: carbon dioxide fixation in chemoautotrophs». Annu Rev Microbiol 52: 191–230. DOI:10.1146/annurev.micro.52.1.191. PMID 9891798.
- ↑ Boiteux A, Hess B (1981). «Design of glycolysis». Philos Trans R Soc Lond B Biol Sci 293 (1063): 5–22. DOI:10.1098/rstb.1981.0056. PMID 6115423.
- ↑ Pilkis S, el-Maghrabi M, Claus T (1990). «Fructose-2,6-bisphosphate in control of hepatic gluconeogenesis. From metabolites to molecular genetics». Diabetes Care 13 (6): 582–99. DOI:10.2337/diacare.13.6.582. PMID 2162755.
- ↑ 1 2 Ensign S (2006). «Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation». Mol Microbiol 61 (2): 274–6. DOI:10.1111/j.1365-2958.2006.05247.x. PMID 16856935.
- ↑ Finn P, Dice J (2006). «Proteolytic and lipolytic responses to starvation». Nutrition 22 (7–8): 830–44. DOI:10.1016/j.nut.2006.04.008. PMID 16815497.
- ↑ 1 2 Kornberg H, Krebs H (1957). «Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle». Nature 179 (4568): 988–91. DOI:10.1038/179988a0. PMID 13430766.
- ↑ Opdenakker G, Rudd P, Ponting C, Dwek R (1993). «Concepts and principles of glycobiology». FASEB J 7 (14): 1330–7. PMID 8224606.
- ↑ McConville M, Menon A (2000). «Recent developments in the cell biology and biochemistry of glycosylphosphatidylinositol lipids (review)». Mol Membr Biol 17 (1): 1–16. DOI:10.1080/096876800294443. PMID 10824734.
- ↑ Chirala S, Wakil S (2004). «Structure and function of animal fatty acid synthase». Lipids 39 (11): 1045–53. DOI:10.1007/s11745-004-1329-9. PMID 15726818.
- ↑ White S, Zheng J, Zhang Y (2005). «The structural biology of type II fatty acid biosynthesis». Annu Rev Biochem 74: 791–831. DOI:10.1146/annurev.biochem.74.082803.133524. PMID 15952903.
- ↑ Ohlrogge J, Jaworski J (1997). «Regulation of fatty acid synthesis». Annu Rev Plant Physiol Plant Mol Biol 48: 109–136. DOI:10.1146/annurev.arplant.48.1.109. PMID 15012259.
- ↑ Dubey V, Bhalla R, Luthra R (2003). «An overview of the non-mevalonate pathway for terpenoid biosynthesis in plants» (PDF). J Biosci 28 (5): 637–46. DOI:10.1007/BF02703339. PMID 14517367.
- ↑ 1 2 Kuzuyama T, Seto H (2003). «Diversity of the biosynthesis of the isoprene units». Nat Prod Rep 20 (2): 171–83. DOI:10.1039/b109860h. PMID 12735695.
- ↑ Grochowski L, Xu H, White R (May 2006). «Methanocaldococcus jannaschii uses a modified mevalonate pathway for biosynthesis of isopentenyl diphosphate». J Bacteriol 188 (9): 3192–8. DOI:10.1128/JB.188.9.3192-3198.2006. PMID 16621811.
- ↑ Lichtenthaler H (1999). «The 1-Ddeoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants». Annu Rev Plant Physiol Plant Mol Biol 50: 47–65. DOI:10.1146/annurev.arplant.50.1.47. PMID 15012203.
- ↑ 1 2 Schroepfer G (1981). «Sterol biosynthesis». Annu Rev Biochem 50: 585–621. DOI:10.1146/annurev.bi.50.070181.003101. PMID 7023367.
- ↑ Lees N, Skaggs B, Kirsch D, Bard M (1995). «Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review». Lipids 30 (3): 221–6. DOI:10.1007/BF02537824. PMID 7791529.
- ↑ Guyton Arthur C. Textbook of Medical Physiology. — Philadelphia: Elsevier, 2006. — P. 855–6. — ISBN 0-7216-0240-1
- ↑ Ibba M, Söll D (2001). «The renaissance of aminoacyl-tRNA synthesis». EMBO Rep 2 (5): 382–7. PMID 11375928.
- ↑ Lengyel P, Söll D (1969). «Mechanism of protein biosynthesis». Bacteriol Rev 33 (2): 264–301. PMID 4896351.
- ↑ 1 2 Rudolph F (1994). «The biochemistry and physiology of nucleotides». J Nutr 124 (1 Suppl): 124S–127S. PMID 8283301.
- ↑ Zrenner R, Stitt M, Sonnewald U, Boldt R (2006). «Pyrimidine and purine biosynthesis and degradation in plants». Annu Rev Plant Biol 57: 805–36. DOI:10.1146/annurev.arplant.57.032905.105421. PMID 16669783.
- ↑ Stasolla C, Katahira R, Thorpe T, Ashihara H (2003). «Purine and pyrimidine nucleotide metabolism in higher plants». J Plant Physiol 160 (11): 1271–95. DOI:10.1078/0176-1617-01169. PMID 14658380.
- ↑ Smith J (1995). «Enzymes of nucleotide synthesis». Curr Opin Struct Biol 5 (6): 752–7. DOI:10.1016/0959-440X(95)80007-7. PMID 8749362.
- ↑ Testa B, Krämer S (2006). «The biochemistry of drug metabolism—an introduction: part 1. Principles and overview». Chem Biodivers 3 (10): 1053–101. DOI:10.1002/cbdv.200690111. PMID 17193224.
- ↑ Danielson P (2002). «The cytochrome P450 superfamily: biochemistry, evolution and drug metabolism in humans». Curr Drug Metab 3 (6): 561–97. DOI:10.2174/1389200023337054. PMID 12369887.
- ↑ King C, Rios G, Green M, Tephly T (2000). «UDP-glucuronosyltransferases». Curr Drug Metab 1 (2): 143–61. DOI:10.2174/1389200003339171. PMID 11465080.
- ↑ Sheehan D, Meade G, Foley V, Dowd C (November 2001). «Structure, function and evolution of glutathione transferases: implications for classification of non-mammalian members of an ancient enzyme superfamily». Biochem J 360 (Pt 1): 1–16. DOI:10.1042/0264-6021:3600001. PMID 11695986.
- ↑ Galvão T, Mohn W, de Lorenzo V (2005). «Exploring the microbial biodegradation and biotransformation gene pool». Trends Biotechnol 23 (10): 497–506. DOI:10.1016/j.tibtech.2005.08.002. PMID 16125262.
- ↑ Janssen D, Dinkla I, Poelarends G, Terpstra P (2005). «Bacterial degradation of xenobiotic compounds: evolution and distribution of novel enzyme activities». Environ Microbiol 7 (12): 1868–82. DOI:10.1111/j.1462-2920.2005.00966.x. PMID 16309386.
- ↑ Davies K (1995). «Oxidative stress: the paradox of aerobic life». Biochem Soc Symp 61: 1–31. PMID 8660387.
- ↑ Tu B, Weissman J (2004). «Oxidative protein folding in eukaryotes: mechanisms and consequences». J Cell Biol 164 (3): 341–6. DOI:10.1083/jcb.200311055. PMID 14757749.
- ↑ Sies H (1997). «Oxidative stress: oxidants and antioxidants» (PDF). Exp Physiol 82 (2): 291–5. PMID 9129943.
- ↑ Vertuani S, Angusti A, Manfredini S (2004). «The antioxidants and pro-antioxidants network: an overview». Curr Pharm Des 10 (14): 1677–94. DOI:10.2174/1381612043384655. PMID 15134565.
- ↑ von Stockar U, Liu J (1999). «Does microbial life always feed on negative entropy? Thermodynamic analysis of microbial growth». Biochim Biophys Acta 1412 (3): 191–211. DOI:10.1016/S0005-2728(99)00065-1. PMID 10482783.
- ↑ Demirel Y, Sandler S (2002). «Thermodynamics and bioenergetics». Biophys Chem 97 (2–3): 87–111. DOI:10.1016/S0301-4622(02)00069-8. PMID 12050002.
- ↑ Albert R (2005). «Scale-free networks in cell biology». J Cell Sci 118 (Pt 21): 4947–57. DOI:10.1242/jcs.02714. PMID 16254242.
- ↑ Brand M (1997). «Regulation analysis of energy metabolism». J Exp Biol 200 (Pt 2): 193–202. PMID 9050227.
- ↑ Soyer O, Salathé M, Bonhoeffer S (2006). «Signal transduction networks: topology, response and biochemical processes». J Theor Biol 238 (2): 416–25. DOI:10.1016/j.jtbi.2005.05.030. PMID 16045939.
- ↑ Westerhoff H, Groen A, Wanders R (1984). «Modern theories of metabolic control and their applications (review)». Biosci Rep 4 (1): 1–22. DOI:10.1007/BF01120819. PMID 6365197.
- ↑ Salter M, Knowles R, Pogson C (1994). «Metabolic control». Essays Biochem 28: 1–12. PMID 7925313.
- ↑ Fell D, Thomas S (1995). «Physiological control of metabolic flux: the requirement for multisite modulation». Biochem J 311 (Pt 1): 35–9. PMID 7575476.
- ↑ Hendrickson W (2005). «Transduction of biochemical signals across cell membranes». Q Rev Biophys 38 (4): 321–30. DOI:10.1017/S0033583506004136. PMID 16600054.
- ↑ Cohen P (2000). «The regulation of protein function by multisite phosphorylation—a 25 year update». Trends Biochem Sci 25 (12): 596–601. DOI:10.1016/S0968-0004(00)01712-6. PMID 11116185.
- ↑ Lienhard G, Slot J, James D, Mueckler M (1992). «How cells absorb glucose». Sci Am 266 (1): 86–91. DOI:10.1038/scientificamerican0192-86. PMID 1734513.
- ↑ Roach P (2002). «Glycogen and its metabolism». Curr Mol Med 2 (2): 101–20. DOI:10.2174/1566524024605761. PMID 11949930.
- ↑ Newgard C, Brady M, O’Doherty R, Saltiel A (2000). «Organizing glucose disposal: emerging roles of the glycogen targeting subunits of protein phosphatase-1» (PDF). Diabetes 49 (12): 1967–77. DOI:10.2337/diabetes.49.12.1967. PMID 11117996.
- ↑ Romano A, Conway T (1996). «Evolution of carbohydrate metabolic pathways». Res Microbiol 147 (6–7): 448–55. DOI:10.1016/0923-2508(96)83998-2. PMID 9084754.
- ↑ Koch A (1998). «How did bacteria come to be?». Adv Microb Physiol 40: 353–99. DOI:10.1016/S0065-2911(08)60135-6. PMID 9889982.
- ↑ Ouzounis C, Kyrpides N (1996). «The emergence of major cellular processes in evolution». FEBS Lett 390 (2): 119–23. DOI:10.1016/0014-5793(96)00631-X. PMID 8706840.
- ↑ Caetano-Anolles G, Kim HS, Mittenthal JE (2007). «The origin of modern metabolic networks inferred from phylogenomic analysis of protein architecture». Proc Natl Acad Sci USA 104 (22): 9358–63. DOI:10.1073/pnas.0701214104. PMID 17517598.
- ↑ Schmidt S, Sunyaev S, Bork P, Dandekar T (2003). «Metabolites: a helping hand for pathway evolution?». Trends Biochem Sci 28 (6): 336–41. DOI:10.1016/S0968-0004(03)00114-2. PMID 12826406.
- ↑ Light S, Kraulis P (2004). «Network analysis of metabolic enzyme evolution in Escherichia coli». BMC Bioinformatics 5: 15. DOI:10.1186/1471-2105-5-15. PMID 15113413. Alves R, Chaleil R, Sternberg M (2002). «Evolution of enzymes in metabolism: a network perspective». J Mol Biol 320 (4): 751–70. DOI:10.1016/S0022-2836(02)00546-6. PMID 12095253.
- ↑ Kim HS, Mittenthal JE, Caetano-Anolles G (2006). «MANET: tracing evolution of protein architecture in metabolic networks». BMC Bioinformatics 19 (7): 351. DOI:10.1186/1471-2105-7-351. PMID 16854231.
- ↑ Teichmann SA, Rison SC, Thornton JM, Riley M, Gough J, Chothia C (2001). «Small-molecule metabolsim: an enzyme mosaic». Trends Biotechnol 19 (12): 482–6. DOI:10.1016/S0167-7799(01)01813-3. PMID 11711174.
- ↑ Spirin V, Gelfand M, Mironov A, Mirny L (June 2006). «A metabolic network in the evolutionary context: multiscale structure and modularity». Proc Natl Acad Sci USA 103 (23): 8774–9. DOI:10.1073/pnas.0510258103. PMID 16731630.
- ↑ Lawrence J (2005). «Common themes in the genome strategies of pathogens». Curr Opin Genet Dev 15 (6): 584–8. DOI:10.1016/j.gde.2005.09.007. PMID 16188434. Wernegreen J (2005). «For better or worse: genomic consequences of intracellular mutualism and parasitism». Curr Opin Genet Dev 15 (6): 572–83. DOI:10.1016/j.gde.2005.09.013. PMID 16230003.
- ↑ Pál C, Papp B, Lercher M, Csermely P, Oliver S, Hurst L (2006). «Chance and necessity in the evolution of minimal metabolic networks». Nature 440 (7084): 667–70. DOI:10.1038/nature04568. PMID 16572170.
- ↑ Rennie M (1999). «An introduction to the use of tracers in nutrition and metabolism». Proc Nutr Soc 58 (4): 935–44. DOI:10.1017/S002966519900124X. PMID 10817161.
- ↑ Phair R (1997). «Development of kinetic models in the nonlinear world of molecular cell biology». Metabolism 46 (12): 1489–95. DOI:10.1016/S0026-0495(97)90154-2. PMID 9439549.
- ↑ Sterck L, Rombauts S, Vandepoele K, Rouzé P, Van de Peer Y (2007). «How many genes are there in plants (… and why are they there)?». Curr Opin Plant Biol 10 (2): 199–203. DOI:10.1016/j.pbi.2007.01.004. PMID 17289424.
- ↑ Borodina I, Nielsen J (2005). «From genomes to in silico cells via metabolic networks». Curr Opin Biotechnol 16 (3): 350–5. DOI:10.1016/j.copbio.2005.04.008. PMID 15961036.
- ↑ Gianchandani E, Brautigan D, Papin J (2006). «Systems analyses characterize integrated functions of biochemical networks». Trends Biochem Sci 31 (5): 284–91. DOI:10.1016/j.tibs.2006.03.007. PMID 16616498.
- ↑ Duarte NC, Becker SA, Jamshidi N, et al. (February 2007). «Global reconstruction of the human metabolic network based on genomic and bibliomic data». Proc. Natl. Acad. Sci. U.S.A. 104 (6): 1777–82. DOI:10.1073/pnas.0610772104. PMID 17267599.
- ↑ Goh KI, Cusick ME, Valle D, Childs B, Vidal M, Barabási AL (May 2007). «The human disease network». Proc. Natl. Acad. Sci. U.S.A. 104 (21): 8685–90. DOI:10.1073/pnas.0701361104. PMID 17502601.
- ↑ Lee DS, Park J, Kay KA, Christakis NA, Oltvai ZN, Barabási AL (July 2008). «The implications of human metabolic network topology for disease comorbidity». Proc. Natl. Acad. Sci. U.S.A. 105 (29): 9880–9885. DOI:10.1073/pnas.0802208105. PMID 18599447.
- ↑ Csete M, Doyle J (2004). «Bow ties, metabolism and disease». Trends Biotechnol. 22 (9): 446–50. DOI:10.1016/j.tibtech.2004.07.007. PMID 5249808.
- ↑ Ma HW, Zeng AP (2003). «The connectivity structure, giant strong component and centrality of metabolic networks». Bioinformatics 19 (11): 1423–30. DOI:10.1093/bioinformatics/btg177. PMID 12874056.
- ↑ Zhao J, Yu H, Luo JH, Cao ZW, Li YX (2006). «Hierarchical modularity of nested bow-ties in metabolic networks». BMC Bioinformatics 7: 386. DOI:10.1186/1471-2105-7-386. PMID 16916470.
- ↑ Thykaer J, Nielsen J (2003). «Metabolic engineering of beta-lactam production». Metab Eng 5 (1): 56–69. DOI:10.1016/S1096-7176(03)00003-X. PMID 12749845. González-Pajuelo M, Meynial-Salles I, Mendes F, Andrade J, Vasconcelos I, Soucaille P (2005). «Metabolic engineering of Clostridium acetobutylicum for the industrial production of 1,3-propanediol from glycerol». Metab Eng 7 (5–6): 329–36. DOI:10.1016/j.ymben.2005.06.001. PMID 16095939. Krämer M, Bongaerts J, Bovenberg R, Kremer S, Müller U, Orf S, Wubbolts M, Raeven L (2003). «Metabolic engineering for microbial production of shikimic acid». Metab Eng 5 (4): 277–83. DOI:10.1016/j.ymben.2003.09.001. PMID 14642355.
- ↑ Koffas M, Roberge C, Lee K, Stephanopoulos G (1999). «Metabolic engineering». Annu Rev Biomed Eng 1: 535–57. DOI:10.1146/annurev.bioeng.1.1.535. PMID 11701499.
- ↑ Dr. Abu Shadi Al-Roubi (1982), «Ibn Al-Nafis as a philosopher», Symposium on Ibn al Nafis, Second International Conference on Islamic Medicine: Islamic Medical Organization, Kuwait (cf. Ibnul-Nafees As a Philosopher, Encyclopedia of Islamic World [1]).
- ↑ Eknoyan G (1999). «Santorio Sanctorius (1561–1636) — founding father of metabolic balance studies». Am J Nephrol 19 (2): 226–33. DOI:10.1159/000013455. PMID 10213823.
- ↑ Williams, H. S. (1904) A History of Science: in Five Volumes. Volume IV: Modern Development of the Chemical and Biological Sciences Harper and Brothers (New York) Retrieved on 2007-03-26
- ↑ Dubos J. (1951). «Louis Pasteur: Free Lance of Science, Gollancz. Quoted in Manchester K. L. (1995) Louis Pasteur (1822–1895)—chance and the prepared mind». Trends Biotechnol 13 (12): 511–515. DOI:10.1016/S0167-7799(00)89014-9. PMID 8595136.
- ↑ Kinne-Saffran E, Kinne R (1999). «Vitalism and synthesis of urea. From Friedrich Wöhler to Hans A. Krebs». Am J Nephrol 19 (2): 290–4. DOI:10.1159/000013463. PMID 10213830.
- ↑ Eduard Buchner’s 1907 Nobel lecture at http://nobelprize.org Accessed 2007-03-20
- ↑ Kornberg H (2000). «Krebs and his trinity of cycles». Nat Rev Mol Cell Biol 1 (3): 225–8. DOI:10.1038/35043073. PMID 11252898.
- ↑ Krebs HA, Henseleit K (1932). «Untersuchungen über die Harnstoffbildung im tierkorper». Z. Physiol. Chem. 210: 33–66.Krebs H, Johnson W (April 1937). «Metabolism of ketonic acids in animal tissues». Biochem J 31 (4): 645–60. PMID 16746382.
Ссылки
Что такое и как работает метаболизм – правда и мифы?
Что такое и как работает метаболизм – правда и мифы?
Распространение неверных трактовок относительно понятия метаболизм и надуманные правила похудения приводят в заблуждение многих людей. Мы разоблачаем мифы об обмене веществ и объясняем суть этого процесса.
Правильное питание – основа здоровья, долголетия и красивой фигуры. Когда человек желает снизить вес, он старается всевозможными способами сжигать больше калорий, начинает изучать многочисленные статьи о похудении и пытается разобраться в таких понятиях, как метаболизм и обмен веществ. При этом очень важно отделять действительно полезную информацию от мифов.
Метаболизм и существующие факты
Сегодня чуть ли не ежедневно появляются новые мифы и неподтвержденные факты по поводу похудения, сжигания калорий, ведения правильного образа жизни. Понимание своего организма и изучение проверенной информации позволит вам принимать разумные решения для достижения желаемой физической формы. Необходимо уметь отделять истину от выдумки и отсеивать мифические факты.
Метаболизм – это процесс обмена веществ, нормальное функционирование которого важно для здоровья и помогает убрать лишние килограммы. Метаболизм способствует нормализации естественного баланса. В таком случае полезные компоненты усваиваются организмом.
Внимание! Нарушение этой функции приводит к увеличению массы тела и причиняет существенный дискомфорт.
Понятия из области фантастики
Метаболизм – основа жизни любого организма, поэтому важно понимание этого процесса. Некоторые существующие сегодня факты об обмене веществ недостоверны. Вот некоторые из них:
- Метаболизм – это просто преобразование полученных организмом калорий в энергетические запасы. На самом деле это более широкое понятие, включающее разделение поступающих органических соединений и синтез нужных организму веществ.
- Скорость метаболизма не подвергается корректировке. Это утверждение из мифической области, поскольку скорость этого процесса зависит от количества калорий, которые нужны для поддержания стабильной формы. Чтобы снизить массу тела, необходимо увеличить нагрузку на организм и потреблять столько же калорий в день, сколько он расходует.
- С возрастом метаболизм замедляется, снижается физическая активность, и организм требует больше покоя. Это так. Если не снизить количество калорий, то их избыток начнет накапливаться день за днем в виде лишних килограммов.
- Употребление холодной пищи позволит сжигать больше калорий, поскольку дополнительное их количество будет потрачено на ее разогрев. Лабораторные исследования позволили опровергнуть правдоподобность этого утверждения.
- Сокращение калорий ведет к снижению обменных реакций. Значительного их замедления при неизменных физических нагрузках и нахождении организма в длительном покое заметно не будет.
- Отказ от еды после 18:00 приводит к похудению, поскольку интенсивность метаболизма в этот период снижена. В данном случае вступает в силу правило: сколько потребил – столько потратил. Отказ от питания после 6 вечера предполагает сокращение потребления калорий за день.
Внимание! Если человек начнет отказываться от еды вечером, но будет переедать до назначенного времени, то норма калорий за день повысится. Сроки приема продуктов будут распределены неравномерно, поэтому риск перенасыщения и срыва диеты увеличится.
Это лишь некоторые мифы о метаболизме. Не попадайтесь на вымышленные факты, научитесь вытягивать то зерно истины, которое поможет добиться результата!
Отказ от ответсвенности
Обращаем ваше внимание, что вся информация, размещённая на сайте Prowellness предоставлена исключительно в ознакомительных целях и не является персональной программой, прямой рекомендацией к действию или врачебными советами. Не используйте данные материалы для диагностики, лечения или проведения любых медицинских манипуляций. Перед применением любой методики или употреблением любого продукта проконсультируйтесь с врачом. Данный сайт не является специализированным медицинским порталом и не заменяет профессиональной консультации специалиста. Владелец Сайта не несет никакой ответственности ни перед какой стороной, понесший косвенный или прямой ущерб в результате неправильного использования материалов, размещенных на данном ресурсе.
Метаболизм в масштабе всей жизни
Измерение расхода энергии на протяжении всей жизни человека позволяет выявить особенности различных фаз метаболизма.
Под метаболизмом понимается не только процесс получения энергии, но и то, как происходит переработка и преобразование питательных веществ в «удобную энергетическую валюту». В понятие метаболизма входят также синтез, модификация и обмен строительных блоков для всех клеточных функций; процесс обмена веществ и энергии выступает в качестве сенсора и регулятора активности жизнедеятельности клетки, когда отдельные части метаболических путей оказывают влияние на протекание клеточных реакций на различные раздражители. Значительное количество энергии, потребляемой каждый день, требуется для поддержания жизни; энергетические потребности для осуществления физической активности накладываются на чрезвычайно сложный механизм. Состояние метаболизма связано с огромным числом заболеваний и расстройств, в том числе тех, которые проявляются с возрастом [1–3]. В этом выпуске «Science» Pontzer с соавт. [4] проанализировали потребление энергии у более чем 6400 мужчин и женщин в возрасте от 8 дней до 95 лет из 29 стран мира и выявили различные метаболические фазы в ходе постнатального развития и старения.
В понимание энергозатрат на протяжении всей жизни должно входить разнообразие фенотипов человека с учетом пола, расы, состава тела с точки зрения содержания мышечной массы и подкожного жира, а также окружающей среды. Оценить энергозатраты можно с помощью непрямой калориметрии, которая определяет газообмен и теплопродукцию у человека в специальной изолированной камере, или с помощью метода дважды меченой воды (DLW) у людей с обычным образом жизни. Метод DLW основан на относительной скорости выведения изотопов кислорода и водорода из организма [5]. Использование метода DLW неуклонно возрастало тех пор, как велась разработка методов для массового применения среди людей [6]. Сопутствующие затраты на дозирование изотопов ограничивали большинство исследований относительно небольшими выборками, однако исследовательским сообществом были взяты обязательства по обмену данными и разработке интегративных методов, которые позволили бы проводить анализ данных на значительно больших когортах [7].
В исследовании Pontzer с соавт. энергозатраты организма были скорректированы с учетом массы без жировой ткани для учета различий в размерах тела при выявлении внутренних сдвигов состояния обмена веществ в процессе постнатального развития, полового созревания и старения. Авторы определили переходные моменты, которые отображают границы четырех отдельных фаз. Согласно их данным, дети и подростки образуют две различные группы по уровню обмена веществ. Как уже ранее отмечалось, дети — это не просто маленькие взрослые [8]. То, что среди молодых людей определяются различные группы по состоянию метаболизма, является крайне важным для рекомендаций по рациону питания и физической активности, не говоря уже о рекомендациях по дозированию фармацевтических препаратов для молодых людей. Оставшиеся две фазы включают взрослых людей и пожилых. Если скорректировать показатели энергозатрат, они остаются на примерно одном и том же уровне в возрасте от 20 до 60 лет, после чего наблюдается постепенное снижение расхода энергии (см. рисунок).
Рисунок 1 | Метаболизм в масштабе всей жизни
Считается, что данное снижение скорости обмена веществ после 60 лет отражает изменение уровня тканевого метаболизма, т. е. количества энергии, затрачиваемой на поддержание жизнедеятельности. Не может быть совпадением, что рост неинфекционной заболеваемости начинается в то же время [9]. Согласно этим результатам, при выборе моделей болезни необходимо тщательно учитывать период жизни человека. Это оказывается особенно важным для исследований этиологии возрастных заболеваний [10]. Не всегда возможно оказать воздействие на те же молекулярные пути и факторы у пожилого, что и у молодого растущего организма. Влияние на точки приложения лекарственного воздействия, например, может не привести к такому же отклику. И наоборот, неправильным является подход переложения условий жизнедеятельности пожилого организма на молодой. Суть многих метаболических процессов у пожилых и молодых в значительной мере различается.
Влияние размера тела на скорость метаболизма обсуждается и исследуется на протяжении десятилетий [11]. Общий расход энергии характеризуется половым диморфизмом: он более низкий у женщин, чем у мужчин; однако подсчет скорости обмена веществ без массы жировой ткани устраняет это различие. Для того, чтобы вычислительные модели соответствовали наблюдаемым данным, важно учитывать вклад физической активности и скорости метаболизма в тканях, ведь как первое, так и второе претерпевают изменения на протяжении всей жизни человека. Хотя это и не является основной темой работы, Pontzer с соавт. выявили существенные различия в составе тканей тела среди людей. Проблемы, возникающие вследствие этой гетерогенности, находят отражение во все возрастающем энтузиазме относительно прецизионной (индивидуализированной) медицины [12]. Совершенно очевидно, что один размер одежды и обуви не подходит всем. Путем корректировки массы без жировой ткани, это исследование устраняет некоторые из этих различий для выявления внутренне обусловленных изменений обмена веществ, соответствующих периоду жизни. Существуют значимые различия в характере и сроках проявления признаков старения с точки зрения заболеваемости [13]. Важно изучить, как предрасположенность к заболеваниям в среднем возрасте влияет на исходы этих патологий в пожилом возрасте, а также насколько тяжесть заболеваний у людей связана с возрастными изменениями их метаболизма, специфичного относительно конкретных тканей.
Причинные факторы возрастной склонности к развитию болезней, несомненно, кроются в четко фиксируемых изменениях клеточной биологии, физиологии тканей и системного гомеостаза. Благодаря исследованиям на лабораторных животных, метаболизм превратился в центральное звено процессов старения, и пришло понимание того, что замедление старения происходит в том числе и за счет ограничения калорийности рациона [14]. Предполагается, что различия в метаболизме влияют на получение энергии из нутриентов, являющихся основным материалом для синтеза различных веществ внутри клетки и работы путей сигнализации, а также на способность согласовывать работу клетки в соответствии с существующими условиями (как внешними, так и внутренними). Поэтому неудивительно, что в попытках определить фармакологические средства, способные положительно влиять на здоровье при старении, все подходящие вещества так или иначе оказываются теми, что способны влиять на обмен веществ [15]. Исследование Pontzer с соавт. позволяет по-новому взглянуть на метаболизм человека; исследование отличается беспрецедентным масштабом, но оно было бы невозможным без совместной работы специалистов из разных областей.
Лактат
Лактат – это продукт клеточного метаболизма, производная молочной кислоты. Может находиться в клетках в виде самой молочной кислоты либо в виде ее солей.
Синонимы русские
Молочная кислота, соли молочной кислоты.
Синонимы английские
Lactate, lactic acid.
Метод исследования
Кинетический колориметрический метод.
Единицы измерения
Ммоль/л (миллимоль на литр).
Какой биоматериал можно использовать для исследования?
Венозную кровь.
Как правильно подготовиться к исследованию?
- Не принимать пищу в течение 12 часов до исследования.
- Исключить физическое и эмоциональное перенапряжение в течение 30 минут до исследования.
- Не курить в течение 30 минут до исследования.
Общая информация об исследовании
В ходе анализа измеряется количество лактатов в крови. Они являются продуктом клеточного метаболизма и в зависимости от рН (кислотности) могут присутствовать в клетках в виде молочной кислоты или при нейтральной рН в форме солей молочной кислоты.
В норме концентрация лактатов в крови очень низкая. В мышцах, эритроцитах, клетках мозга и в других тканях она повышается при недостатке кислорода в клетке либо если первичный путь производства энергии в клетках нарушен.
Основные запасы клеточной энергии производятся в митохондриях, крошечных «энергетических станциях» внутри клеток организма. Митохондрии используют глюкозу и кислород для производства АТФ (аденозинтрифосфата), главного энергетического источника в организме. Это называется аэробным образованием энергии.
При падении уровня кислорода в клетке либо при нарушении нормального функционирования митохондрий организм переключается на менее эффективное производство энергии (анаэробное) путем расщепления глюкозы с образованием АТФ. Лактат является основным побочным продуктом этого анаэробного процесса. Молочная кислота может накапливаться в случае, если она производится быстрее, чем печень успевает ее утилизировать. Когда ее содержание в крови значительно повышается, наступает гиперлактатацидемия, которая может далее развиться в лактацидоз, если молочная кислота будет продолжать накапливаться. Организму часто удается компенсировать эффект лактацидоза, однако в тяжелых случаях нарушается кислотно-щелочной баланс, что сопровождается слабостью, учащенным дыханием, тошнотой, рвотой, потливостью и даже комой.
Причины повышения уровня лактатов подразделяются на две группы в соответствии с механизмом, который вызывает лактацидоз.
Лактацидоз А-типа наиболее распространен и ассоциирован с факторами, вызывающими недостаточный захват кислорода легкими либо замедленное кровообращение, что приводит к уменьшению снабжения тканей кислородом. Примеры лактацидоза А-типа:
Лактацидоз Б-типа не связан с поступлением кислорода к тканям, он является причиной повышенной потребности в кислороде из-за проблем обмена веществ. Примеры лактацидоза Б-типа:
- болезни печени,
- почечные заболевания,
- сахарный диабет,
- лейкемия,
- СПИД,
- болезни, связанные с сохранением гликогена (например, глюкозо-6-фосфатазная недостаточность),
- лекарства и токсины, такие как салицилаты, цианиды, метанол, метформин,
- различные наследственные митохондриальные и метаболические заболевания, являющиеся формами мышечной дистрофии и затрагивающие синтез АТФ,
- состояние при интенсивных физических нагрузках.
Для чего используется исследование?
- Для выявления лактацидоза, то есть высокого содержания лактатов.
- Чтобы определить гипоксию и лактацидоз и оценить их тяжесть, если есть факторы, понижающие снабжение кислородом клеток и тканей (лактацидоз чаще всего возникает именно из-за этого), например шок или застойная сердечная недостаточность.
- Для оценки кислотно-щелочного баланса и оксигенации (вместе с анализом на газы в крови).
- При диагностике болезней, которые способны привести к повышенному содержанию лактатов, а также при симптомах ацидоза, поскольку лактацидоз может вызываться факторами, не связанными с уровнем кислорода в тканях.
- Чтобы выяснить, не являются ли сопутствующие заболевания, например болезни печени или почек, причиной лактацидоза (вместе с другими исследованиями, такими как клинический анализ крови или мочи, некоторые биохимические тесты).
- Для обследования больных с подозрением на сепсис. Если уровень лактатов у них падает ниже нормы, лечение им назначается незамедлительно. При своевременной диагностике и безотлагательном лечении сепсиса шансы на успешное выздоровление увеличиваются во много раз.
- Для наблюдения за течением гипоксии и контроля за эффективностью ее лечения в случае обострения таких болезней, как сепсис, инфаркт и застойная сердечная недостаточность.
Когда назначается исследование?
- При симптомах недостатка кислорода (одышка, учащенное дыхание, бледность, потливость, тошнота, слабость в мышцах).
- При подозрении на сепсис, шок, инфаркт, сердечную недостаточность, почечную недостаточность или сахарный диабет.
- При острых головных болях, лихорадке, расстройстве и потере сознания, а также признаках менингита.
Что означают результаты?
Референсные значения: 0,5 — 2,2 ммоль/л.
Клиническое значение имеет лишь повышение концентрации лактата в крови.
- Высокая концентрация лактата указывает на болезнь (либо иные факторы), которая является причиной накопления лактатов в тканях. В целом чем выше уровень лактатов, тем острее протекает заболевание. Если накопление лактатов связано с гипоксией, то их повышение означает, что организм не способен ее компенсировать. В то же время сама по себе избыточная концентрация лактатов не является прямым указанием на диагноз, она лишь помогает подтвердить либо исключить возможные причины наблюдаемых симптомов.
- Если есть подозрение на состояние, ведущее к кислородной недостаточности, например на шок, полученный в результате травмы или сильной кровопотери, сепсис, инфаркт, застойную сердечную недостаточность, острые респираторные или легочные заболевания, отек легких, острую анемию, то повышенный уровень лактатов может быть признаком гипоксии и/или дисфункции органов.
- Иногда лактацидоз является осложнением болезней печени, почек, диабета, лейкемии, СПИДа, болезней, связанных с сохранением гликогена (например, глюкозо-6-фосфатазной недостаточностью), различных наследственных митохондриальных и метаболических заболеваний (форм мышечной дистрофии и тех, которые затрагивают синтез АТФ).
- Увеличивать концентрацию лактатов способны лекарства и токсины (салицилаты, цианиды, метанол, метформин) и интенсивные физические нагрузки.
- При симптомах менингита значительно повышенный уровень лактатов в цереброспинальной жидкости указывает на вероятность бактериального менингита, в то время как слегка повышенный – на его вирусную разновидность.
- Нормальный уровень лактатов свидетельствует о том, что у пациента нет лактацидоза, а также о достаточном снабжении кислородом на клеточном уровне.
- При лечении лактацидоза или гипоксии уменьшение концентрации лактатов со временем отражает реакцию организма на процесс лечения.
«Метаболический синдром» – проблема 21 века. 2017023.new
«Метаболический синдром»– проблема, которая набирает обороты во всем Мире. Подробнее о том, что такое метаболический синдром, как с ним бороться расскажет Красильникова Марина Олеговна, врач-гастроэнтеролог, гепатолог сети многопрофильных клиник ДИАЛАЙН, кандидат медицинский наук и куратор программы по ведению пациентов с метаболическим синдромом.— Что такое метаболический синдром?
— Метаболический синдром— одна из наиболее актуальных проблем современной медицины, связанная с ведением нездорового образа жизни. Метаболический синдром — это комплекс нарушений обмена веществ, при котором повышен риск развития сердечно-сосудистых заболеваний и диабета 2 типа. Снижение физической активности и высококалорийный характер питания являются главными причинами того, что заболеваемость метаболическим синдромом растет. На данный момент им страдает около 25% населения.
— Как определить его наличие?
— Предпосылками для появления метаболического синдрома являются малоподвижный образ жизни, избыток массы тела и ожирение, артериальная гипертензия, наличие ишемическая болезнь сердца, диабет 2 типа и некоторые другие заболевания. Важно вовремя диагностировать данное заболевание и начать лечение. Возможны следующие жалобы при метаболическом синдроме — повышенная утомляемость, апатия, одышка, повышенный аппетит, жажда, учащенное мочеиспускание, головная боль, ночная потливость, вспышки ярости, исчезающие после приема пищи, слабость, сонливость после мясной пищи. Основной признак наличия метаболического синдрома центральный (абдоминальный) тип ожирения—т.е. окружность талии более 80 см. у женщин и более 94 см. у мужчин, а также индекс массы тела более 25. Дополнительные критерии- определяет специалист: артериальное давление больше 140/90 мм рт. ст., повышение уровня ТТГ(тиреотропный гормон), изменение липидного спектра, гипогликемия натощак.
— Многие люди живут и не подозревают, что причиной ухудшения их состояния может быть метаболический синдром. Расскажите подробнее о том, что могут предложить в клиниках по диагностике и профилактике метаболического синдрома?
— В наших клиниках разработаны чек-апы – специальные комплексные программы обследования, которые позволят в короткий срок диагностировать наличие метаболического синдрома, своевременно назначить лечение и подобрать индивидуальную программу питания. Остановлюсь на чек-апе «метаболический синдром расширенный + ТТГ». Что он в себя включает: анализ ТТГ(тиреотропный гормон)- это гормон, определяющий функцию щитовидной железы и обмен веществ. Так же проводится ЭКГ и биоимпедансометрия -определение процентного соотношения воды, мышечной и жировой ткани для выявления метаболического синдрома.
— Как это происходит на практике и в какой срок?
— Пациент проходит лабораторные и инструментальные обследования, после чего следует консультация специалиста с назначением лечения и подбором программы питания. Срок прохождения данного чек-апа два дня.
Если вы обнаружили у себя признаки метаболического синдрома, не стоит затягивать, а лучше своевременно обратиться к специалисту.
Метаболизм: основные концепции и дизайн — Биохимия
Концепции конформации и динамики, разработанные в Части I, особенно те, которые касаются специфичности и каталитической силы ферментов, регуляции их каталитической активности и транспорта молекул и ионов через мембраны — позвольте нам теперь задать вопросы, фундаментальные для биохимии:
Эти процессы осуществляются высоко интегрированной сетью химических реакций, которые вместе известны как метаболизм .
Более тысячи химических реакций происходят даже в таком простом организме, как Escherichia coli . На первый взгляд множество реакций может показаться ошеломляющим. Однако более пристальное рассмотрение показывает, что метаболизм имеет согласованный дизайн , содержащий множество общих мотивов . Эти мотивы включают использование энергетической валюты и повторяющееся появление ограниченного числа активированных промежуточных продуктов. Фактически, группа из примерно 100 молекул играет центральную роль во всех формах жизни.Более того, хотя количество реакций в метаболизме велико, количество видов реакций невелико, и механизмы этих реакций обычно довольно просты. Метаболические пути также регулируются обычным образом. Цель этой главы — познакомить с некоторыми общими принципами и мотивами метаболизма, чтобы заложить основу для последующих более подробных исследований.
14.0.1. Клетки трансформируют различные типы энергии:
Живые организмы нуждаются в непрерывном поступлении свободной энергии для трех основных целей: (1) выполнение механической работы по сокращению мышц и других клеточных движениях, (2) активный транспорт молекул и ионов, и (3) синтез макромолекул и других биомолекул из простых предшественников.Свободная энергия, используемая в этих процессах, которые поддерживают организм в состоянии, далеком от равновесия, поступает из окружающей среды.
Первый закон термодинамики гласит, что энергия не может быть ни создана, ни уничтожена. Количество энергии во Вселенной постоянно. Тем не менее, энергия может быть преобразована из одной формы в другую.
Фотосинтезирующие организмы, или фототрофов , используют энергию солнечного света для преобразования простых молекул с низким энергопотреблением в более сложные молекулы, богатые энергией, которые служат топливом.Другими словами, фотосинтезирующие организмы преобразуют световую энергию в химическую энергию. Действительно, это преобразование, в конечном счете, является основным источником химической энергии для подавляющего большинства организмов, включая человека. Хемотрофы , в состав которых входят животные, получают химическую энергию в результате окисления пищевых продуктов, производимых фототрофами.
Химическая энергия, полученная при окислении углеродных соединений, может трансформироваться в неравномерное распределение ионов через мембрану, что приводит к ионному градиенту.Этот градиент, в свою очередь, является источником энергии, который можно использовать для перемещения молекул через мембраны, который может быть преобразован в другие типы химической энергии или может передавать информацию в форме нервных импульсов. Кроме того, химическая энергия может быть преобразована в механическую. Мы преобразуем химическую энергию топлива в структурные изменения сократительных белков, которые приводят к сокращению и движению мышц. Наконец, химическая энергия поддерживает реакции, приводящие к синтезу биомолекул.
В любой момент в клетке происходят тысячи преобразований энергии. Энергия извлекается из топлива и используется для обеспечения биосинтетических процессов. Эти преобразования обозначаются как метаболизм или промежуточный метаболизм .
Рисунок
Колибри обладают невероятной выносливостью. Например, крошечный колибри с рубиновым горлом может накапливать достаточно топлива, чтобы без отдыха перелететь через Мексиканский залив на расстояние около 500 миль.Это достижение возможно благодаря (подробнее …)
4.1 Энергия и метаболизм — Концепции биологии — 1-е канадское издание
Цели обучения
К концу этого раздела вы сможете:
- Объясните, что такое метаболические пути
- Изложите первый и второй законы термодинамики.
- Объясните разницу между кинетической и потенциальной энергией
- Опишите эндергонические и экзэргонические реакции.
- Обсудите, как ферменты действуют как молекулярные катализаторы
Посмотрите видео о гетеротрофах.
Ученые используют термин биоэнергетика для описания концепции потока энергии (рис. 4.2) через живые системы, такие как клетки. Клеточные процессы , такие как построение и разрушение сложных молекул , происходят посредством ступенчатых химических реакций . Некоторые из этих химических реакций являются спонтанными и высвобождают энергию, тогда как другие требуют энергии для протекания. Точно так же, как живые существа должны постоянно потреблять пищу для пополнения своих запасов энергии, клетки должны постоянно производить больше энергии, чтобы восполнить то, что используется многими химическими реакциями, требующими энергии, которые постоянно происходят.В совокупности, , все химические реакции , которые происходят внутри клеток, включая те, которые потребляют или генерируют энергию, называются метаболизмом клетки .
Рис. 4.2. В конечном счете, большинство форм жизни получают энергию от солнца. Растения используют фотосинтез для захвата солнечного света, а травоядные животные поедают растения для получения энергии. Плотоядные животные едят травоядных, и возможное разложение растительного и животного материала способствует пополнению запасов питательных веществ.Рассмотрим метаболизм сахара.Это классический пример одного из многих клеточных процессов, которые используют и производят энергию. Живые существа потребляют сахар в качестве основного источника энергии, потому что молекулы сахара имеют много энергии, хранящейся в их связях. По большей части фотосинтезирующие организмы, такие как растения, производят эти сахара. Во время фотосинтеза растения используют энергию (первоначально из солнечного света) для преобразования газообразного диоксида углерода (CO 2 ) в молекулы сахара (например, глюкозы: C 6 H 12 O 6 ).Они потребляют углекислый газ и выделяют кислород в качестве побочного продукта. Эта реакция резюмируется как:
6CO 2 + 6H 2 O + энергия ——-> C 6 H 12 O 6 + 6O 2
Поскольку этот процесс включает синтез молекулы, накапливающей энергию, для его выполнения требуется подача энергии. Во время световых реакций фотосинтеза энергия обеспечивается молекулой, называемой аденозинтрифосфатом (АТФ) , которая является основным источником энергии для всех клеток.Так же, как доллар используется в качестве валюты для покупки товаров, клетки используют молекулы АТФ в качестве энергетической валюты для выполнения немедленной работы. Напротив, молекулы-накопители энергии, такие как глюкоза, потребляются только для того, чтобы расщепиться для использования своей энергии. Реакцию, которая собирает энергию молекулы сахара в клетках, нуждающихся в кислороде для выживания, можно описать обратной реакцией на фотосинтез. В этой реакции расходуется кислород и выделяется углекислый газ в качестве побочного продукта. Реакция резюмируется как:
C 6 H 12 O 6 + 6O 2 ——> 6CO 2 + 6H 2 O + энергия
Обе эти реакции включают много этапов.
Процессы образования и расщепления молекул сахара иллюстрируют два примера метаболических путей. Метаболический путь — это серия химических реакций, в которых исходная молекула изменяется, шаг за шагом, через ряд промежуточных продуктов метаболизма, в конечном итоге давая конечный продукт. В примере метаболизма сахара первый метаболический путь синтезирует сахар из более мелких молекул, а другой путь расщепляет сахар на более мелкие молекулы. Эти два противоположных процесса — первый, требующий энергии, а второй — производящий энергию — называются анаболическими путями (строительные полимеры) и катаболическими путями (разрушение полимеров на их мономеры) соответственно.Следовательно, метаболизм состоит из синтеза (анаболизма) и деградации (катаболизма) (рис. 4.3).
Важно знать, что химические реакции метаболических путей не происходят сами по себе. Каждая стадия реакции ускоряется или катализируется белком, называемым ферментом. Ферменты важны для катализирования всех типов биологических реакций — как тех, которые требуют энергии, так и тех, которые выделяют энергию.
Рис. 4.3. Катаболические пути — это те пути, которые генерируют энергию, расщепляя более крупные молекулы.Анаболические пути — это те, которые требуют энергии для синтеза более крупных молекул. Оба типа путей необходимы для поддержания энергетического баланса клетки.Термодинамика относится к изучению энергии и передачи энергии с участием физической материи. Материя, относящаяся к конкретному случаю передачи энергии, называется системой, а все, что находится вне этой материи, называется окружающей средой. Например, при нагревании кастрюли с водой на плите система включает плиту, кастрюлю и воду.Энергия передается внутри системы (между плитой, кастрюлей и водой). Есть два типа систем: открытая и закрытая. В открытой системе можно обмениваться энергией с окружающей средой. Плита открыта, потому что тепло может отдаваться воздуху. Закрытая система не может обмениваться энергией со своим окружением.
Биологические организмы — это открытые системы. Между ними и их окружением происходит обмен энергией, поскольку они используют энергию солнца для фотосинтеза или потребляют молекулы, накапливающие энергию, и выделяют энергию в окружающую среду, выполняя работу и выделяя тепло.Как и все в физическом мире, энергия подчиняется физическим законам. Законы термодинамики управляют передачей энергии внутри и между всеми системами во Вселенной.
В общем, энергия определяется как способность выполнять работу или создавать какие-либо изменения. Энергия существует в разных формах. Например, электрическая энергия, световая энергия и тепловая энергия — это разные виды энергии. Чтобы понять, как энергия поступает в биологические системы и выходит из них, важно понимать два физических закона, управляющих энергией.
Первый закон термодинамики гласит, что общее количество энергии во Вселенной постоянно и сохраняется. Другими словами, во Вселенной всегда было и будет точно такое же количество энергии. Энергия существует во многих различных формах . Согласно первому закону термодинамики, энергия может передаваться с места на место или преобразовываться в разные формы, , но не может быть создана или уничтожена . Передачи и преобразования энергии происходят вокруг нас постоянно.Лампочки преобразуют электрическую энергию в световую и тепловую. Газовые плиты преобразуют химическую энергию природного газа в тепловую. Растения осуществляют одно из наиболее биологически полезных преобразований энергии на Земле: преобразование энергии солнечного света в химическую энергию, хранящуюся в органических молекулах (рис. 4.2). Некоторые примеры преобразования энергии показаны на рисунке 4.4.
Задача всех живых организмов состоит в том, чтобы получать энергию из окружающей среды в формах, которые они могут передавать или преобразовывать в полезную энергию для выполнения работы.Живые клетки эволюционировали, чтобы справиться с этой задачей. Химическая энергия, хранящаяся в органических молекулах, таких как сахара и жиры, передается и преобразуется посредством ряда клеточных химических реакций в энергию в молекулах АТФ. Энергия в молекулах АТФ легко доступна для работы. Примеры типов работы, которую должны выполнять клетки, включают построение сложных молекул, транспортировку материалов, обеспечение движения ресничек или жгутиков и сокращение мышечных волокон для создания движения.
Рисунок 4.4 Показаны некоторые примеры передачи и преобразования энергии из одной системы в другую и из одной формы в другую. Пища, которую мы потребляем, обеспечивает наши клетки энергией, необходимой для выполнения функций организма, так же как световая энергия дает растениям средства для создания необходимой им химической энергии. (кредит «мороженое»: модификация работы Д. Шэрон Прюитт; кредит «дети»: модификация работы Макса из Провиденса; кредитный «лист»: модификация работы Кори Занкера)Основные задачи получения живой клетки, преобразование и использование энергии для работы может показаться простым.Однако второй закон термодинамики объясняет, почему эти задачи сложнее, чем кажется. Всякая передача энергии и преобразование никогда не бывают полностью эффективными . При каждой передаче энергии некоторое количество энергии теряется в непригодной для использования форме. В большинстве случаев это форма тепловой энергии. Термодинамически тепловая энергия определяется как неработающая энергия, передаваемая от одной системы к другой. Например, когда включается электрическая лампочка, часть энергии, преобразуемой из электрической энергии в энергию света, теряется в виде тепловой энергии.Точно так же часть энергии теряется в виде тепловой энергии во время клеточных метаболических реакций.
Важным понятием физических систем является понятие порядка и беспорядка. Чем больше энергии теряется системой в свое окружение, тем менее упорядоченной и случайной является система. Ученые называют меру случайности или беспорядка в системе энтропией . Высокая энтропия означает высокий беспорядок и низкую энергию. Молекулы и химические реакции также имеют разную энтропию. Например, энтропия увеличивается, когда молекулы с высокой концентрацией в одном месте диффундируют и разлетаются.Второй закон термодинамики гласит, что энергия всегда будет теряться в виде тепла при передаче или преобразовании энергии.
Живые существа очень упорядочены, и для их поддержания в состоянии низкой энтропии требуется постоянный ввод энергии.
Когда объект находится в движении, с ним связана энергия. Подумайте о шаре для разрушения. Даже медленно движущийся шар-разрушитель может нанести большой урон другим объектам. Энергия, связанная с движущимися объектами, называется кинетической энергией (Рисунок 4.5). Ускоряющаяся пуля, идущий человек и быстрое движение молекул в воздухе (выделяющих тепло) — все они обладают кинетической энергией.
А что, если тот же самый неподвижный шар для разрушения поднять краном на два этажа над землей? Если подвешенный шар для разрушения неподвижен, связана ли с ним энергия? Ответ положительный. Энергия, которая требовалась для подъема разрушающего шара, не исчезла, но теперь сохраняется в разрушающем шаре в силу его положения и силы тяжести, действующей на него.Этот вид энергии называется потенциальной энергией (рис. 4.5). Если мяч упадет, потенциальная энергия будет преобразована в кинетическую энергию до тех пор, пока вся потенциальная энергия не будет исчерпана, когда мяч упадет на землю. Шары-крушители тоже качаются, как маятник; во время качания происходит постоянное изменение потенциальной энергии (самая высокая в верхней части качания) на кинетическую энергию (самая высокая в нижней части качания). Другие примеры потенциальной энергии включают энергию воды, удерживаемой за плотиной, или человека, который собирается прыгнуть с парашютом из самолета.
Рисунок 4.5 У негазированной воды есть потенциальная энергия; движущаяся вода, например, в водопаде или в быстро текущей реке, обладает кинетической энергией. (кредит «дамба»: модификация работы «Паскаля» / Flickr; кредит «водопад»: модификация работы Фрэнка Гуалтьери)Потенциальная энергия связана не только с расположением материи, но и со структурой материи. Даже пружина на земле имеет потенциальную энергию, если она сжата; то же самое происходит и с туго натянутой резинкой. На молекулярном уровне связи, которые удерживают атомы молекул вместе, существуют в определенной структуре, обладающей потенциальной энергией.Помните, что анаболические клеточные пути требуют энергии для синтеза сложных молекул из более простых, а катаболические пути высвобождают энергию при расщеплении сложных молекул. Тот факт, что энергия может выделяться при разрыве определенных химических связей, означает, что эти связи обладают потенциальной энергией. Фактически, в связях всех пищевых молекул, которые мы едим, хранится потенциальная энергия, которая в конечном итоге используется для использования. Это потому, что эти связи могут высвобождать энергию при разрыве.Тип потенциальной энергии, которая существует в химических связях и высвобождается при разрыве этих связей, называется химической энергией. Химическая энергия отвечает за обеспечение живых клеток энергией из пищи. Высвобождение энергии происходит при разрыве молекулярных связей в молекулах пищи.
Посмотрите видео о килокалориях.
Концепция в действии
Посетите сайт и выберите «Маятник» в меню «Работа и энергия», чтобы увидеть изменение кинетической и потенциальной энергии маятника в движении.
Узнав, что химические реакции высвобождают энергию при разрыве энергонакопительных связей, возникает следующий важный вопрос: как количественно и выражается энергия, связанная с этими химическими реакциями? Как можно сравнить энергию, выделяемую в результате одной реакции, с энергией другой реакции? Измерение свободной энергии используется для количественной оценки этой передачи энергии. Напомним, что согласно второму закону термодинамики, любая передача энергии связана с потерей некоторого количества энергии в непригодной для использования форме, такой как тепло.Свободная энергия, в частности, относится к энергии, связанной с химической реакцией, которая доступна после учета потерь. Другими словами, свободная энергия — это полезная энергия или энергия, доступная для выполнения работы.
Если энергия выделяется во время химической реакции, то изменение свободной энергии, обозначенное как ∆G (дельта G), будет отрицательным числом. Отрицательное изменение свободной энергии также означает, что продукты реакции имеют меньше свободной энергии, чем реагенты, потому что они выделяют некоторую свободную энергию во время реакции.Реакции, которые имеют отрицательное изменение свободной энергии и, следовательно, высвобождают свободную энергию, называются экзергоническими реакциями. Подумайте: ex эргономичный означает, что энергия ex в системе. Эти реакции также называются спонтанными реакциями, и их продукты имеют меньше накопленной энергии, чем реагенты. Необходимо провести важное различие между термином «спонтанный» и идеей немедленного протекания химической реакции. В отличие от повседневного использования этого термина, спонтанная реакция — это не реакция, которая возникает внезапно или быстро.Ржавчина железа — это пример спонтанной реакции, которая происходит медленно, мало-помалу, с течением времени.
Если химическая реакция поглощает энергию, а не высвобождает ее в балансе, то ∆G для этой реакции будет положительным значением. В этом случае у продуктов больше свободной энергии, чем у реагентов. Таким образом, продукты этих реакций можно рассматривать как молекулы, запасающие энергию. Эти химические реакции называются эндергоническими реакциями , а — несамопроизвольными .Эндергоническая реакция не будет происходить сама по себе без добавления свободной энергии.
Рисунок 4.6. Показаны некоторые примеры эндергонических процессов (требующих энергии) и экзэргонических процессов (тех, которые выделяют энергию). (кредит a: модификация работы Натали Мэйнор; кредит b: модификация работы Министерством сельского хозяйства США; кредит c: модификация работы Кори Занкера; кредит d: модификация работы Гарри Мальша)Посмотрите на каждый из представленных процессов и решите если он эндергонический или экзергонический.
Есть еще одна важная концепция, которую необходимо учитывать в отношении эндергонических и экзэргонических реакций. Экзергонические реакции требуют небольшого количества энергии для начала, прежде чем они смогут приступить к своим этапам высвобождения энергии. Эти реакции имеют чистое высвобождение энергии, но все же требуют некоторого ввода энергии вначале. Это небольшое количество энергии, необходимое для протекания всех химических реакций, называется энергией активации.
Концепция в действии
Посмотрите анимацию перехода от свободной энергии к переходному состоянию реакции.
Вещество, которое способствует протеканию химической реакции, называется катализатором, а молекулы, катализирующие биохимические реакции, называются ферментами. Большинство ферментов представляют собой белки и выполняют критическую задачу , снижая энергии активации химических реакций внутри клетки. Большинство реакций, критических для живой клетки, протекают слишком медленно при нормальной температуре, чтобы быть полезными для клетки. Без ферментов ускорить эти реакции жизнь не могла бы существовать.Ферменты делают это, связываясь с молекулами реагентов и удерживая их таким образом, чтобы облегчить процессы разрыва и образования химических связей. Важно помнить, что ферменты не изменяют, является ли реакция экзергонической (спонтанной) или эндергонической. Это потому, что они не изменяют свободную энергию реагентов или продуктов. Они только уменьшают энергию активации, необходимую для продолжения реакции (рис. 4.7). Кроме того, сам фермент не изменяется в результате реакции, которую он катализирует.После того, как одна реакция катализируется, фермент может участвовать в других реакциях.
Рис. 4.7. Ферменты снижают энергию активации реакции, но не изменяют свободную энергию реакции.Химические реагенты, с которыми связывается фермент, называются субстратами фермента. В зависимости от конкретной химической реакции может быть один или несколько субстратов. В некоторых реакциях один реагент-субстрат распадается на несколько продуктов. В других случаях два субстрата могут объединиться, чтобы создать одну большую молекулу.Два реагента также могут вступить в реакцию, и оба они станут модифицированными, но выходят из реакции в виде двух продуктов. Место внутри фермента, где связывается субстрат, называется активным сайтом фермента . Активный сайт — это место, где происходит «действие». Поскольку ферменты являются белками, в активном центре существует уникальная комбинация боковых цепей аминокислот. Каждая боковая цепь характеризуется разными свойствами. Они могут быть большими или маленькими, слабокислотными или основными, гидрофильными или гидрофобными, положительно или отрицательно заряженными или нейтральными.Уникальная комбинация боковых цепей создает очень специфическую химическую среду в активном центре. Эта специфическая среда подходит для связывания с одним конкретным химическим субстратом (или субстратами).
Активные сайты подвержены влиянию местной среды. Повышение температуры окружающей среды обычно увеличивает скорость реакции, катализируемой ферментами или иначе. Однако температуры за пределами оптимального диапазона снижают скорость, с которой фермент катализирует реакцию. Высокие температуры в конечном итоге вызывают денатурирование ферментов, необратимое изменение трехмерной формы и, следовательно, функции фермента.Ферменты также подходят для наилучшего функционирования в определенном диапазоне pH и концентрации соли, и, как и в случае с температурой, экстремальные значения pH и концентрации соли могут вызывать денатурирование ферментов.
В течение многих лет ученые считали, что связывание фермента с субстратом происходит простым «замком и ключом». Эта модель утверждает, что фермент и субстрат идеально сочетаются друг с другом за один мгновенный шаг. Однако текущие исследования поддерживают модель, называемую индуцированной подгонкой (рис. 4.8). Модель индуцированной подгонки расширяет модель блокировки и ключа, описывая более динамическое связывание между ферментом и субстратом.Когда фермент и субстрат объединяются, их взаимодействие вызывает небольшой сдвиг в структуре фермента, который формирует идеальную структуру связывания между ферментом и субстратом.
Концепция в действии
Просмотрите анимацию индуцированной посадки.
Когда фермент связывает свой субстрат, образуется комплекс фермент-субстрат. Этот комплекс снижает энергию активации реакции и способствует ее быстрому развитию одним из множества возможных способов. На базовом уровне ферменты способствуют химическим реакциям, в которых участвует более одного субстрата, объединяя субстраты вместе в оптимальной ориентации для реакции.Другой способ, которым ферменты способствуют реакции своих субстратов, — это создание оптимальной среды в активном центре для протекания реакции. Химические свойства, проистекающие из особого расположения R-групп аминокислот в активном центре, создают идеальную среду для реакции определенных субстратов фермента.
Комплекс фермент-субстрат может также снизить энергию активации за счет нарушения структуры связи, так что ее легче разорвать. Наконец, ферменты также могут снижать энергию активации, принимая участие в самой химической реакции.В этих случаях важно помнить, что фермент всегда возвращается в исходное состояние по завершении реакции. Одним из отличительных свойств ферментов является то, что они в конечном итоге остаются неизменными в результате катализируемых ими реакций. После того, как фермент катализирует реакцию, он высвобождает свой продукт (ы) и может катализировать новую реакцию.
Рис. 4.8. Модель индуцированной подгонки представляет собой корректировку модели «замок-и-ключ» и объясняет, как ферменты и субстраты претерпевают динамические модификации во время переходного состояния для увеличения сродства субстрата к активному сайту.Казалось бы, идеальным иметь сценарий, в котором все ферменты организма существуют в изобилии и оптимально функционируют во всех клеточных условиях, во всех клетках, во все времена. Однако множество механизмов гарантирует, что этого не произойдет. Клеточные потребности и условия постоянно меняются от клетки к клетке и со временем меняются внутри отдельных клеток. Необходимые ферменты клеток желудка отличаются от ферментов жировых клеток, клеток кожи, клеток крови и нервных клеток. Кроме того, клетка пищеварительного органа намного усерднее обрабатывает и расщепляет питательные вещества в течение времени, которое следует за едой, по сравнению со многими часами после еды.Поскольку эти клеточные потребности и условия меняются, должны меняться количества и функциональность различных ферментов.
Поскольку скорость биохимических реакций контролируется энергией активации, а ферменты ниже и определяют энергии активации химических реакций, относительные количества и функционирование различных ферментов в клетке в конечном итоге определяют, какие реакции будут протекать и с какой скоростью. Это определение строго контролируется в клетках. В определенных клеточных средах активность ферментов частично контролируется факторами окружающей среды, такими как pH, температура, концентрация соли и, в некоторых случаях, кофакторами или коферментами.
Ферменты также могут регулироваться способами, которые либо способствуют, либо снижают активность фермента. Есть много видов молекул, которые подавляют или стимулируют функцию ферментов, и различные механизмы, с помощью которых они это делают. В некоторых случаях ингибирования фермента молекула ингибитора достаточно похожа на субстрат, чтобы она могла связываться с активным сайтом и просто блокировать связывание субстрата. Когда это происходит, фермент ингибируется посредством конкурентного ингибирования , потому что молекула ингибитора конкурирует с субстратом за связывание с активным сайтом.
С другой стороны, при неконкурентном ингибировании молекула ингибитора связывается с ферментом в месте, отличном от активного сайта, называемом аллостерическим сайтом , но все же удается блокировать связывание субстрата с активным сайтом. Некоторые молекулы ингибитора связываются с ферментами в том месте, где их связывание вызывает конформационное изменение, которое снижает сродство фермента к его субстрату. Этот тип торможения называется аллостерическим торможением (рис. 4.9).Большинство аллостерически регулируемых ферментов состоят из более чем одного полипептида, что означает, что они имеют более одной белковой субъединицы. Когда аллостерический ингибитор связывается с областью фермента, все активные центры белковых субъединиц слегка изменяются, так что они связывают свои субстраты с меньшей эффективностью. Есть аллостерические активаторы, а также ингибиторы. Аллостерические активаторы связываются с участками фермента, удаленными от активного сайта, вызывая конформационные изменения, которые увеличивают сродство активного сайта (ов) фермента к его субстрату (ам) (Рисунок 4.9).
Рис. 4.9. Аллостерическое ингибирование работает, косвенно вызывая конформационные изменения активного сайта, так что субстрат больше не подходит. Напротив, при аллостерической активации молекула активатора изменяет форму активного сайта, чтобы обеспечить лучшее прилегание субстрата.Через призму коренных народов
Растения не могут убежать или спрятаться от своих хищников, и они разработали множество стратегий, чтобы отпугнуть тех, кто их съел. Подумайте о шипах, раздражителях и вторичных метаболитах: это соединения, которые напрямую не помогают растениям расти, а созданы специально для защиты от хищников.Вторичные метаболиты — наиболее распространенный способ отпугивания хищников. Некоторые примеры вторичных метаболитов — атропин, никотин, ТГК и кофеин. Люди обнаружили, что эти вторичные метаболиты являются богатым источником материалов для лекарств. Подсчитано, что 90% лекарств в современной аптеке имеют свои «корни» в этих вторичных метаболитах.
Лечение травами первых людей открыло миру эти вторичные метаболиты. Например, коренные народы издавна использовали кору ивовых кустарников и ольхи для приготовления чая, тонизирующего средства или припарок, чтобы уменьшить воспаление.Вы узнаете больше о воспалительной реакции иммунной системы в главе 11.
Рис. 4.10. Кора тихоокеанской ивы содержит соединение салицин.И ива, и кора ольхи содержат соединение салицин. У большинства из нас в аптечке есть это соединение в виде салициловой кислоты или аспирина. Доказано, что аспирин уменьшает боль и воспаление, а попав в наши клетки, салицин превращается в салициловую кислоту.
Так как это работает? Салицин или аспирин действуют как ингибитор фермента.В воспалительной реакции ключевыми в этом процессе являются два фермента, COX1 и COX2. Салицин или аспирин специфически модифицируют аминокислоту (серин) в активном центре этих двух родственных ферментов. Эта модификация активных центров не позволяет нормальному субстрату связываться и, таким образом, нарушается воспалительный процесс. Как вы читали в этой главе, это делает его конкурентным ингибитором ферментов.
Разработчик фармацевтических препаратов
Рис. 4.11 Задумывались ли вы, как создаются фармацевтические препараты? (кредит: Дебора Остин)Ферменты — ключевые компоненты метаболических путей.Понимание того, как работают ферменты и как их можно регулировать, — ключевые принципы, лежащие в основе разработки многих фармацевтических препаратов, представленных сегодня на рынке. Биологи, работающие в этой области, совместно с другими учеными разрабатывают лекарства (рис. 4.11).
Рассмотрим, к примеру, статины. Статины — это название одного класса лекарств, которые могут снижать уровень холестерина. Эти соединения являются ингибиторами фермента HMG-CoA редуктазы, который является ферментом, синтезирующим холестерин из липидов в организме.Ингибируя этот фермент, можно снизить уровень холестерина, синтезируемого в организме. Точно так же ацетаминофен, широко продаваемый под торговой маркой Tylenol, является ингибитором фермента циклооксигеназы. Хотя он используется для снятия лихорадки и воспаления (боли), его механизм действия до сих пор полностью не изучен.
Как обнаруживаются наркотики? Одна из самых больших проблем в открытии лекарств — это определение мишени для лекарства. Мишень лекарства — это молекула, которая буквально является мишенью лекарства.В случае статинов мишенью для лечения является HMG-CoA редуктаза. Цели лекарств определяются путем кропотливых лабораторных исследований. Одной идентификации цели недостаточно; ученым также необходимо знать, как мишень действует внутри клетки и какие реакции идут наперекосяк в случае болезни. Как только цель и путь определены, начинается фактический процесс разработки лекарств. На этом этапе химики и биологи работают вместе, чтобы разработать и синтезировать молекулы, которые могут блокировать или активировать определенную реакцию.Однако это только начало: если и когда прототип лекарства успешно выполняет свою функцию, он подвергается множеству тестов, от экспериментов in vitro до клинических испытаний, прежде чем он получит одобрение Управления по санитарному надзору за качеством пищевых продуктов и медикаментов США. магазин.
Многие ферменты не работают оптимально или даже не работают, если они не связаны с другими специфическими небелковыми вспомогательными молекулами. Они могут связываться либо временно посредством ионных или водородных связей, либо навсегда посредством более прочных ковалентных связей.Связывание с этими молекулами способствует оптимальной форме и функционированию соответствующих ферментов. Двумя примерами этих типов вспомогательных молекул являются кофакторы и коферменты. Кофакторы — это неорганические ионы, такие как ионы железа и магния. Коферменты — это органические вспомогательные молекулы, имеющие базовую атомную структуру, состоящую из углерода и водорода. Подобно ферментам, эти молекулы участвуют в реакциях, не меняясь сами, и в конечном итоге перерабатываются и используются повторно. Витамины являются источником коферментов.Некоторые витамины являются предшественниками коферментов, а другие действуют непосредственно как коферменты. Витамин C является прямым коферментом множества ферментов, которые участвуют в создании важной соединительной ткани — коллагена. Следовательно, функция фермента частично регулируется обилием различных кофакторов и коферментов, которые могут поступать с пищей организма или, в некоторых случаях, вырабатываться организмом.
Рис. 4.12. Витамины являются важными коферментами или предшественниками коферментов и необходимы для правильного функционирования ферментов.Мультивитаминные капсулы обычно содержат смеси всех витаминов в разном процентном соотношении.Ингибирование обратной связи в метаболических путях
Молекулы могут регулировать функцию ферментов разными способами. Однако остается главный вопрос: что это за молекулы и откуда они берутся? Как вы уже знаете, некоторые из них являются кофакторами и коферментами. Какие другие молекулы в клетке обеспечивают ферментативную регуляцию, такую как аллостерическая модуляция, а также конкурентное и неконкурентное ингибирование? Возможно, наиболее подходящими источниками регуляторных молекул для ферментативного клеточного метаболизма являются продукты самих клеточных метаболических реакций.Наиболее эффективным и элегантным образом клетки эволюционировали, чтобы использовать продукты собственных реакций для подавления активности ферментов с помощью обратной связи. Подавление обратной связи предполагает использование продукта реакции для регулирования его собственного дальнейшего производства (рис. 4.12). Клетка реагирует на обилие продуктов замедлением производства во время анаболических или катаболических реакций. Такие продукты реакции могут ингибировать ферменты, которые катализируют их производство с помощью механизмов, описанных выше.
Рисунок 4.13 Метаболические пути — это серия реакций, катализируемых множеством ферментов. Ингибирование обратной связи, когда конечный продукт пути ингибирует вышестоящий процесс, является важным регуляторным механизмом в клетках.Производство как аминокислот, так и нуклеотидов контролируется посредством ингибирования с обратной связью. Кроме того, АТФ является аллостерическим регулятором некоторых ферментов, участвующих в катаболическом распаде сахара, процессе, который создает АТФ. Таким образом, когда АТФ в избытке, клетка может предотвратить производство АТФ.С другой стороны, АДФ служит положительным аллостерическим регулятором (аллостерическим активатором) для некоторых из тех же ферментов, которые ингибируются АТФ. Таким образом, когда относительные уровни АДФ высоки по сравнению с АТФ, клетка начинает производить больше АТФ за счет катаболизма сахара.
Клетки выполняют жизненные функции посредством различных химических реакций. Метаболизм клетки — это комбинация химических реакций, которые происходят в ней. Катаболические реакции расщепляют сложные химические вещества на более простые и связаны с выделением энергии.Анаболические процессы создают сложные молекулы из более простых и требуют энергии.
При изучении энергии термин «система» относится к веществу и окружающей среде, участвующим в передаче энергии. Энтропия — это мера беспорядка системы. Физические законы, описывающие передачу энергии, являются законами термодинамики. Первый закон гласит, что общее количество энергии во Вселенной постоянно. Второй закон термодинамики гласит, что каждая передача энергии включает некоторую потерю энергии в непригодной для использования форме, такой как тепловая энергия.Энергия бывает разных форм: кинетической, потенциальной и свободной. Изменение свободной энергии реакции может быть отрицательным (высвобождает энергию, экзергоническое) или положительным (потребляет энергию, эндергоническое). Все реакции требуют начального ввода энергии, называемой энергией активации.
Ферменты — это химические катализаторы, которые ускоряют химические реакции за счет снижения их энергии активации. Ферменты имеют активный центр с уникальной химической средой, которая соответствует определенным химическим реагентам для этого фермента, называемым субстратами.Считается, что ферменты и субстраты связываются в соответствии с моделью индуцированной подгонки. Действие ферментов регулируется для сохранения ресурсов и оптимального реагирования на окружающую среду.
Глоссарий
энергия активации: количество начальной энергии, необходимой для протекания реакции
активный сайт: специфическая область на ферменте, где субстрат связывается
аллостерическое ингибирование: механизм ингибирования действия фермента, при котором регуляторная молекула связывается со вторым сайтом (не активным сайтом) и инициирует изменение конформации в активном сайте, предотвращая связывание с субстратом
анаболический: описывает путь, который требует ввода чистой энергии для синтеза сложных молекул из более простых
биоэнергетика: концепция потока энергии через живые системы
катаболический: описывает путь, по которому сложные молекулы распадаются на более простые, выделяя энергию в качестве дополнительного продукта реакции.
конкурентное ингибирование: общий механизм регуляции активности фермента, при котором молекула, отличная от субстрата фермента, способна связывать активный сайт и предотвращать связывание самого субстрата, тем самым подавляя общую скорость реакции фермента
endergonic: описывает химическую реакцию, в результате которой образуются продукты, которые хранят больше химической потенциальной энергии, чем реагенты.
фермент: молекула, катализирующая биохимическую реакцию
exergonic: описывает химическую реакцию, в результате которой образуются продукты с меньшей химической потенциальной энергией, чем у реагентов, плюс высвобождение свободной энергии
ингибирование обратной связи: механизм регулирования активности фермента, в котором продукт реакции или конечный продукт ряда последовательных реакций ингибирует фермент на более ранней стадии в серии реакций
тепловая энергия: энергия, передаваемая из одной системы в другую, которая не работает
кинетическая энергия: тип энергии, связанной с движущимися объектами
метаболизм: все химические реакции, происходящие внутри клеток, включая те, которые используют энергию, и те, которые выделяют энергию
неконкурентное ингибирование: общий механизм регуляции активности фермента, при котором регуляторная молекула связывается с сайтом, отличным от активного сайта, и предотвращает связывание активного сайта с субстратом; таким образом, молекула ингибитора не конкурирует с субстратом за активный центр; аллостерическое торможение — это форма неконкурентного торможения
потенциальная энергия: тип энергии, который относится к потенциалу совершать работу
субстрат: молекула, на которую действует фермент
термодинамика: наука о взаимосвязи тепла, энергии и работы
4.1 Энергия и обмен веществ — Основные понятия биологии
Цели обучения
К концу этого раздела вы сможете:- Объясните, что такое метаболические пути
- Изложите первый и второй законы термодинамики.
- Объясните разницу между кинетической и потенциальной энергией
- Опишите эндергонические и экзэргонические реакции.
- Обсудите, как ферменты действуют как молекулярные катализаторы
Ученые используют термин биоэнергетика для описания концепции потока энергии (Рисунок 4.2) через живые системы, например клетки. Клеточные процессы, такие как построение и разрушение сложных молекул, происходят через ступенчатые химические реакции. Некоторые из этих химических реакций являются спонтанными и высвобождают энергию, тогда как другие требуют энергии для протекания. Точно так же, как живые существа должны постоянно потреблять пищу для пополнения своих запасов энергии, клетки должны постоянно получать больше энергии для пополнения энергии, используемой многими химическими реакциями, требующими энергии, которые постоянно происходят.Вместе все химические реакции, происходящие внутри клеток, в том числе те, которые потребляют или генерируют энергию, называются метаболизмом клетки.
Фигура 4.2 В конечном счете, большинство форм жизни получают энергию от солнца. Растения используют фотосинтез для захвата солнечного света, а травоядные животные поедают растения для получения энергии. Плотоядные животные едят травоядных, и возможное разложение растительного и животного материала способствует пополнению запасов питательных веществ.
Метаболические пути
Рассмотрим метаболизм сахара.Это классический пример одного из многих клеточных процессов, которые используют и производят энергию. Живые существа потребляют сахар в качестве основного источника энергии, потому что молекулы сахара имеют много энергии, хранящейся в их связях. По большей части фотосинтезирующие организмы, такие как растения, производят эти сахара. Во время фотосинтеза растения используют энергию (первоначально из солнечного света) для преобразования газообразного диоксида углерода (CO 2 ) в молекулы сахара (например, глюкозы: C 6 H 12 O 6 ).Они потребляют углекислый газ и выделяют кислород в качестве побочного продукта. Эта реакция резюмируется как:
6CO2 + 6h3O -> C6h22O6 + 6O26CO2 + 6h3O -> C6h22O6 + 6O2 размер 12 {C rSub {размер 8 {6}} H rSub {размер 8 {12}} O rSub {размер 8 {2}}} {}Поскольку этот процесс включает синтез молекулы, накапливающей энергию, для его выполнения требуется подача энергии. Во время световых реакций фотосинтеза энергия обеспечивается молекулой, называемой аденозинтрифосфатом (АТФ), которая является основной энергетической валютой всех клеток.Так же, как доллар используется в качестве валюты для покупки товаров, клетки используют молекулы АТФ в качестве энергетической валюты для выполнения немедленной работы. Напротив, молекулы-накопители энергии, такие как глюкоза, потребляются только для того, чтобы расщепиться для использования своей энергии. Реакцию, которая собирает энергию молекулы сахара в клетках, нуждающихся в кислороде для выживания, можно описать обратной реакцией на фотосинтез. В этой реакции расходуется кислород и выделяется углекислый газ в качестве побочного продукта. Реакция резюмируется как:
C6h22O6 + 6O2 -> 6h3O + 6CO2C6h22O6 + 6O2 -> 6h3O + 6CO2 размер 12 {C rSub {размер 8 {6}} H rSub {размер 8 {12}} или rSub {размер 8 {2}}} {}Обе эти реакции включают много этапов.
Процессы образования и расщепления молекул сахара иллюстрируют два примера метаболических путей. Метаболический путь — это серия химических реакций, в которых исходная молекула изменяется, шаг за шагом, через ряд промежуточных продуктов метаболизма, в конечном итоге давая конечный продукт. В примере метаболизма сахара первый метаболический путь синтезирует сахар из более мелких молекул, а другой путь расщепляет сахар на более мелкие молекулы. Эти два противоположных процесса — первый, требующий энергии, а второй — производящий энергию — называются анаболическими путями (строительство полимеров) и катаболическими путями (разрушение полимеров на их мономеры) соответственно.Следовательно, метаболизм состоит из синтеза (анаболизма) и деградации (катаболизма) (рис. 4.3).
Важно знать, что химические реакции метаболических путей не происходят сами по себе. Каждая стадия реакции ускоряется или катализируется белком, называемым ферментом. Ферменты важны для катализирования всех типов биологических реакций — как тех, которые требуют энергии, так и тех, которые выделяют энергию.
Фигура 4.3 Катаболические пути — это те пути, которые генерируют энергию, расщепляя более крупные молекулы.Анаболические пути — это те, которые требуют энергии для синтеза более крупных молекул. Оба типа путей необходимы для поддержания энергетического баланса клетки.
Энергия
Термодинамика относится к изучению энергии и передачи энергии с участием физической материи. Материя, относящаяся к конкретному случаю передачи энергии, называется системой, а все, что находится вне этой материи, называется окружающей средой. Например, при нагревании кастрюли с водой на плите система включает плиту, кастрюлю и воду.Энергия передается внутри системы (между плитой, кастрюлей и водой). Есть два типа систем: открытая и закрытая. В открытой системе можно обмениваться энергией с окружающей средой. Плита открыта, потому что тепло может отдаваться воздуху. Закрытая система не может обмениваться энергией со своим окружением.
Биологические организмы — это открытые системы. Между ними и их окружением происходит обмен энергией, поскольку они используют энергию солнца для фотосинтеза или потребляют молекулы, накапливающие энергию, и выделяют энергию в окружающую среду, выполняя работу и выделяя тепло.Как и все в физическом мире, энергия подчиняется физическим законам. Законы термодинамики управляют передачей энергии внутри и между всеми системами во Вселенной.
В общем, энергия определяется как способность выполнять работу или создавать какие-либо изменения. Энергия существует в разных формах. Например, электрическая энергия, световая энергия и тепловая энергия — это разные виды энергии. Чтобы понять, как энергия поступает в биологические системы и выходит из них, важно понимать два физических закона, управляющих энергией.
Термодинамика
Первый закон термодинамики гласит, что общее количество энергии во Вселенной постоянно и сохраняется. Другими словами, во Вселенной всегда было и будет точно такое же количество энергии. Энергия существует во многих различных формах. Согласно первому закону термодинамики, энергия может передаваться с места на место или преобразовываться в различные формы, но не может быть создана или уничтожена. Передачи и преобразования энергии происходят вокруг нас постоянно.Лампочки преобразуют электрическую энергию в световую и тепловую. Газовые плиты преобразуют химическую энергию природного газа в тепловую. Растения осуществляют одно из наиболее биологически полезных преобразований энергии на Земле: преобразование энергии солнечного света в химическую энергию, хранящуюся в органических молекулах (рис. 4.2). Некоторые примеры преобразования энергии показаны на рисунке 4.4.
Задача всех живых организмов состоит в том, чтобы получать энергию из окружающей среды в формах, которые они могут передавать или преобразовывать в полезную энергию для выполнения работы.Живые клетки эволюционировали, чтобы справиться с этой задачей. Химическая энергия, хранящаяся в органических молекулах, таких как сахара и жиры, передается и преобразуется посредством ряда клеточных химических реакций в энергию в молекулах АТФ. Энергия в молекулах АТФ легко доступна для работы. Примеры типов работы, которую должны выполнять клетки, включают построение сложных молекул, транспортировку материалов, обеспечение движения ресничек или жгутиков и сокращение мышечных волокон для создания движения.
Фигура 4.4 Показаны некоторые примеры передачи и преобразования энергии из одной системы в другую и из одной формы в другую. Пища, которую мы потребляем, обеспечивает наши клетки энергией, необходимой для выполнения функций организма, так же как световая энергия дает растениям средства для создания необходимой им химической энергии. (кредит «мороженое»: модификация работы Д. Шэрон Прюитт; кредит «дети»: модификация работы Макса из Providence; кредит «лист»: модификация работы Кори Занкера)
Основные задачи живой клетки по получению, преобразованию и использованию энергии для работы могут показаться простыми.Однако второй закон термодинамики объясняет, почему эти задачи сложнее, чем кажется. Все передачи и преобразования энергии никогда не бывают полностью эффективными. При каждой передаче энергии некоторое количество энергии теряется в непригодной для использования форме. В большинстве случаев это форма тепловой энергии. Термодинамически тепловая энергия определяется как неработающая энергия, передаваемая от одной системы к другой. Например, когда включается электрическая лампочка, часть энергии, преобразуемой из электрической энергии в энергию света, теряется в виде тепловой энергии.Точно так же часть энергии теряется в виде тепловой энергии во время клеточных метаболических реакций.
Важным понятием физических систем является понятие порядка и беспорядка. Чем больше энергии теряется системой в свое окружение, тем менее упорядоченной и случайной является система. Ученые называют меру случайности или беспорядка в системе энтропией. Высокая энтропия означает высокий беспорядок и низкую энергию. Молекулы и химические реакции также имеют разную энтропию. Например, энтропия увеличивается, когда молекулы с высокой концентрацией в одном месте диффундируют и разлетаются.Второй закон термодинамики гласит, что энергия всегда будет теряться в виде тепла при передаче или преобразовании энергии.
Живые существа очень упорядочены, и для их поддержания в состоянии низкой энтропии требуется постоянный ввод энергии.
Потенциальная и кинетическая энергия
Когда объект находится в движении, с ним связана энергия. Подумайте о шаре для разрушения. Даже медленно движущийся шар-разрушитель может нанести большой урон другим объектам. Энергия, связанная с движущимися объектами, называется кинетической энергией (Рисунок 4.5). Ускоряющаяся пуля, идущий человек и быстрое движение молекул в воздухе (выделяющих тепло) — все они обладают кинетической энергией.
А что, если тот же самый неподвижный шар для разрушения поднять краном на два этажа над землей? Если подвешенный шар для разрушения неподвижен, связана ли с ним энергия? Ответ положительный. Энергия, которая требовалась для подъема разрушающего шара, не исчезла, но теперь сохраняется в разрушающем шаре в силу его положения и силы тяжести, действующей на него.Этот вид энергии называется потенциальной энергией (рис. 4.5). Если мяч упадет, потенциальная энергия будет преобразована в кинетическую энергию до тех пор, пока вся потенциальная энергия не будет исчерпана, когда мяч упадет на землю. Шары-крушители тоже качаются, как маятник; во время качания происходит постоянное изменение потенциальной энергии (самая высокая в верхней части качания) на кинетическую энергию (самая высокая в нижней части качания). Другие примеры потенциальной энергии включают энергию воды, удерживаемой за плотиной, или человека, который собирается прыгнуть с парашютом из самолета.
Фигура 4.5 У негазированной воды есть потенциальная энергия; движущаяся вода, например, в водопаде или в быстро текущей реке, обладает кинетической энергией. (кредит «Плотина»: модификация работы «Pascal» / Flickr; кредит «водопад»: модификация работы Фрэнка Гуалтьери)
Потенциальная энергия связана не только с расположением материи, но и со структурой материи. Даже пружина на земле имеет потенциальную энергию, если она сжата; то же самое происходит и с туго натянутой резинкой.На молекулярном уровне связи, которые удерживают атомы молекул вместе, существуют в определенной структуре, обладающей потенциальной энергией. Помните, что анаболические клеточные пути требуют энергии для синтеза сложных молекул из более простых, а катаболические пути высвобождают энергию при расщеплении сложных молекул. Тот факт, что энергия может выделяться при разрыве определенных химических связей, означает, что эти связи обладают потенциальной энергией. Фактически, в связях всех пищевых молекул, которые мы едим, хранится потенциальная энергия, которая в конечном итоге используется для использования.Это потому, что эти связи могут высвобождать энергию при разрыве. Тип потенциальной энергии, которая существует в химических связях и высвобождается при разрыве этих связей, называется химической энергией. Химическая энергия отвечает за обеспечение живых клеток энергией из пищи. Высвобождение энергии происходит при разрыве молекулярных связей в молекулах пищи.
Ссылка на обучение
Концепция в действии
Посетите сайт и выберите «Маятник» в меню «Работа и энергия», чтобы увидеть изменение кинетической и потенциальной энергии маятника в движении.
Свободная энергия и энергия активации
Узнав, что химические реакции высвобождают энергию при разрыве энергонакопительных связей, возникает следующий важный вопрос: как количественно и выражается энергия, связанная с этими химическими реакциями? Как можно сравнить энергию, выделяемую в результате одной реакции, с энергией другой реакции? Измерение свободной энергии используется для количественной оценки этой передачи энергии. Напомним, что согласно второму закону термодинамики, любая передача энергии связана с потерей некоторого количества энергии в непригодной для использования форме, такой как тепло.Свободная энергия, в частности, относится к энергии, связанной с химической реакцией, которая доступна после учета потерь. Другими словами, свободная энергия — это полезная энергия или энергия, доступная для выполнения работы.
Если энергия выделяется во время химической реакции, то изменение свободной энергии, обозначенное как ∆G (дельта G), будет отрицательным числом. Отрицательное изменение свободной энергии также означает, что продукты реакции имеют меньше свободной энергии, чем реагенты, потому что они выделяют некоторую свободную энергию во время реакции.Реакции, которые имеют отрицательное изменение свободной энергии и, следовательно, высвобождают свободную энергию, называются экзергоническими реакциями. Подумайте: ex эргономичный означает, что энергия ex идет в систему. Эти реакции также называются спонтанными реакциями, и их продукты имеют меньше накопленной энергии, чем реагенты. Необходимо провести важное различие между термином «спонтанный» и идеей немедленного протекания химической реакции. В отличие от повседневного использования этого термина, спонтанная реакция — это не реакция, которая возникает внезапно или быстро.Ржавчина железа — это пример спонтанной реакции, которая происходит медленно, мало-помалу, с течением времени.
Если химическая реакция поглощает энергию, а не высвобождает ее в балансе, то ∆G для этой реакции будет положительным значением. В этом случае у продуктов больше свободной энергии, чем у реагентов. Таким образом, продукты этих реакций можно рассматривать как молекулы, запасающие энергию. Эти химические реакции называются эндергоническими реакциями, и они не являются спонтанными. Эндергоническая реакция не будет происходить сама по себе без добавления свободной энергии.
Визуальное соединение
Визуальное соединение
Фигура 4.6 Показаны некоторые примеры эндергонических процессов (требующих энергии) и экзэргонических процессов (тех, которые высвобождают энергию). (кредит а: модификация работы Натали Мэйнор; кредит b: модификация работы Министерством сельского хозяйства США; кредит c: модификация работы Кори Занкера; кредит d: модификация работы Гарри Мальша)
Посмотрите на каждый из показанных процессов и решите, является ли он эндергоническим или экзергоническим.
Есть еще одна важная концепция, которую необходимо учитывать в отношении эндергонических и экзэргонических реакций. Экзергонические реакции требуют небольшого количества энергии для начала, прежде чем они смогут приступить к своим этапам высвобождения энергии. Эти реакции имеют чистое высвобождение энергии, но все же требуют некоторого ввода энергии вначале. Это небольшое количество энергии, необходимое для протекания всех химических реакций, называется энергией активации.
Ссылка на обучение
Концепция в действии
Посмотрите анимацию перехода от свободной энергии к переходному состоянию реакции.
Ферменты
Вещество, которое способствует протеканию химической реакции, называется катализатором, а молекулы, катализирующие биохимические реакции, называются ферментами. Большинство ферментов являются белками и выполняют важную задачу по снижению энергии активации химических реакций внутри клетки. Большинство реакций, критических для живой клетки, протекают слишком медленно при нормальной температуре, чтобы быть полезными для клетки. Без ферментов, ускоряющих эти реакции, жизнь не могла бы существовать.Ферменты делают это, связываясь с молекулами реагентов и удерживая их таким образом, чтобы облегчить процессы разрыва и образования химических связей. Важно помнить, что ферменты не изменяют, является ли реакция экзергонической (спонтанной) или эндергонической. Это потому, что они не изменяют свободную энергию реагентов или продуктов. Они только уменьшают энергию активации, необходимую для продолжения реакции (рис. 4.7). Кроме того, сам фермент не изменяется в результате реакции, которую он катализирует.После того, как одна реакция катализируется, фермент может участвовать в других реакциях.
Фигура 4,7 Ферменты снижают энергию активации реакции, но не изменяют свободную энергию реакции.
Химические реагенты, с которыми связывается фермент, называются субстратами фермента. В зависимости от конкретной химической реакции может быть один или несколько субстратов. В некоторых реакциях один реагент-субстрат распадается на несколько продуктов.В других случаях два субстрата могут объединиться, чтобы создать одну большую молекулу. Два реагента также могут вступить в реакцию, и оба они станут модифицированными, но выходят из реакции в виде двух продуктов. Место внутри фермента, где связывается субстрат, называется активным центром фермента. Активный сайт — это место, где происходит «действие». Поскольку ферменты являются белками, в активном центре существует уникальная комбинация боковых цепей аминокислот. Каждая боковая цепь характеризуется разными свойствами. Они могут быть большими или маленькими, слабокислотными или основными, гидрофильными или гидрофобными, положительно или отрицательно заряженными или нейтральными.Уникальная комбинация боковых цепей создает очень специфическую химическую среду в активном центре. Эта специфическая среда подходит для связывания с одним конкретным химическим субстратом (или субстратами).
Активные сайты подвержены влиянию местной среды. Повышение температуры окружающей среды обычно увеличивает скорость реакции, катализируемой ферментами или иначе. Однако температуры за пределами оптимального диапазона снижают скорость, с которой фермент катализирует реакцию. Высокие температуры в конечном итоге вызывают денатурирование ферментов, необратимое изменение трехмерной формы и, следовательно, функции фермента.Ферменты также подходят для наилучшего функционирования в определенном диапазоне pH и концентрации соли, и, как и в случае с температурой, экстремальные значения pH и концентрации соли могут вызывать денатурирование ферментов.
В течение многих лет ученые считали, что связывание фермента с субстратом происходит простым «замком и ключом». Эта модель утверждает, что фермент и субстрат идеально сочетаются друг с другом за один мгновенный шаг. Однако текущие исследования поддерживают модель, называемую индуцированной подгонкой (рис. 4.8). Модель индуцированной подгонки расширяет модель блокировки и ключа, описывая более динамическое связывание между ферментом и субстратом.Когда фермент и субстрат объединяются, их взаимодействие вызывает небольшой сдвиг в структуре фермента, который формирует идеальную структуру связывания между ферментом и субстратом.
Ссылка на обучение
Концепция в действии
Просмотрите анимацию индуцированной посадки.
Когда фермент связывает свой субстрат, образуется комплекс фермент-субстрат. Этот комплекс снижает энергию активации реакции и способствует ее быстрому развитию одним из множества возможных способов.На базовом уровне ферменты способствуют химическим реакциям, в которых участвует более одного субстрата, объединяя субстраты вместе в оптимальной ориентации для реакции. Другой способ, которым ферменты способствуют реакции своих субстратов, — это создание оптимальной среды в активном центре для протекания реакции. Химические свойства, проистекающие из особого расположения R-групп аминокислот в активном центре, создают идеальную среду для реакции определенных субстратов фермента.
Комплекс фермент-субстрат может также снизить энергию активации за счет нарушения структуры связи, так что ее легче разорвать. Наконец, ферменты также могут снижать энергию активации, принимая участие в самой химической реакции. В этих случаях важно помнить, что фермент всегда возвращается в исходное состояние по завершении реакции. Одним из отличительных свойств ферментов является то, что они в конечном итоге остаются неизменными в результате катализируемых ими реакций. После того, как фермент катализирует реакцию, он высвобождает свой продукт (ы) и может катализировать новую реакцию.
Фигура 4.8 Модель индуцированной подгонки представляет собой корректировку модели «замок-и-ключ» и объясняет, как ферменты и субстраты претерпевают динамические модификации во время переходного состояния, чтобы увеличить сродство субстрата к активному центру.
Казалось бы, идеальным иметь сценарий, в котором все ферменты организма существуют в изобилии и оптимально функционируют во всех клеточных условиях, во всех клетках, во все времена. Однако множество механизмов гарантирует, что этого не произойдет.Клеточные потребности и условия постоянно меняются от клетки к клетке и со временем меняются внутри отдельных клеток. Необходимые ферменты клеток желудка отличаются от ферментов жировых клеток, клеток кожи, клеток крови и нервных клеток. Кроме того, клетка пищеварительного органа намного усерднее обрабатывает и расщепляет питательные вещества в течение времени, которое следует за едой, по сравнению со многими часами после еды. Поскольку эти клеточные потребности и условия меняются, должны меняться количества и функциональность различных ферментов.
Поскольку скорость биохимических реакций контролируется энергией активации, а ферменты ниже и определяют энергию активации химических реакций, относительные количества и функционирование множества ферментов в клетке в конечном итоге определяют, какие реакции будут протекать и с какой скоростью. Это определение строго контролируется в клетках. В определенных клеточных средах активность ферментов частично контролируется факторами окружающей среды, такими как pH, температура, концентрация соли и, в некоторых случаях, кофакторами или коферментами.
Ферменты также можно регулировать способами, которые либо способствуют, либо снижают активность ферментов. Есть много видов молекул, которые подавляют или стимулируют функцию ферментов, и различные механизмы, с помощью которых они это делают. В некоторых случаях ингибирования фермента молекула ингибитора достаточно похожа на субстрат, чтобы она могла связываться с активным сайтом и просто блокировать связывание субстрата. Когда это происходит, фермент ингибируется за счет конкурентного ингибирования, потому что молекула ингибитора конкурирует с субстратом за связывание с активным сайтом.
С другой стороны, при неконкурентном ингибировании молекула ингибитора связывается с ферментом в месте, отличном от активного сайта, называемом аллостерическим сайтом, но все же удается предотвратить связывание субстрата с активным сайтом. Некоторые молекулы ингибитора связываются с ферментами в том месте, где их связывание вызывает конформационное изменение, которое снижает активность фермента, поскольку больше не катализирует превращение субстрата в продукт. Этот тип торможения называется аллостерическим торможением (Рисунок 4.9). Большинство аллостерически регулируемых ферментов состоят из более чем одного полипептида, что означает, что они имеют более одной белковой субъединицы. Когда аллостерический ингибитор связывается с областью фермента, все активные центры белковых субъединиц слегка изменяются, так что они связывают свои субстраты с меньшей эффективностью. Есть аллостерические активаторы, а также ингибиторы. Аллостерические активаторы связываются с участками фермента, удаленными от активного сайта, вызывая конформационные изменения, которые увеличивают сродство активного сайта (ов) фермента к его субстрату (ам) (Рисунок 4.9).
Фигура 4.9 Аллостерическое ингибирование работает, косвенно вызывая конформационные изменения активного сайта, так что субстрат больше не подходит. Напротив, при аллостерической активации молекула активатора изменяет форму активного сайта, чтобы обеспечить лучшее прилегание субстрата.
Карьера Связь
Карьера в действии
Разработчик фармацевтических препаратов
Фигура 4.10 Вы когда-нибудь задумывались, как создаются фармацевтические препараты? (кредит: Дебора Остин)
Ферменты — ключевые компоненты метаболических путей.Понимание того, как работают ферменты и как их можно регулировать, — ключевые принципы, лежащие в основе разработки многих фармацевтических препаратов, представленных сегодня на рынке. Биологи, работающие в этой области, совместно с другими учеными разрабатывают лекарства (рис. 4.10).
Рассмотрим, к примеру, статины. Статины — это название одного класса препаратов, которые могут снижать уровень холестерина. Эти соединения являются ингибиторами фермента HMG-CoA редуктазы, который является ферментом, синтезирующим холестерин из липидов в организме.Ингибируя этот фермент, можно снизить уровень холестерина, синтезируемого в организме. Точно так же ацетаминофен, широко продаваемый под торговой маркой Tylenol, является ингибитором фермента циклооксигеназы. Хотя он используется для снятия лихорадки и воспаления (боли), его механизм действия до сих пор полностью не изучен.
Как обнаруживаются наркотики? Одна из самых больших проблем в открытии лекарств — это определение мишени для лекарства. Мишень лекарства — это молекула, которая буквально является мишенью лекарства.В случае статинов мишенью для лечения является HMG-CoA редуктаза. Цели лекарств определяются путем кропотливых лабораторных исследований. Одной идентификации цели недостаточно; ученым также необходимо знать, как мишень действует внутри клетки и какие реакции идут наперекосяк в случае болезни. Как только цель и путь определены, начинается фактический процесс разработки лекарств. На этом этапе химики и биологи работают вместе, чтобы разработать и синтезировать молекулы, которые могут блокировать или активировать определенную реакцию.Однако это только начало: если и когда прототип лекарства успешно выполняет свою функцию, он подвергается множеству тестов, от экспериментов in vitro до клинических испытаний, прежде чем он получит одобрение Управления по санитарному надзору за качеством пищевых продуктов и медикаментов США. магазин.
Многие ферменты не работают оптимально или даже не работают, если они не связаны с другими специфическими небелковыми вспомогательными молекулами. Они могут связываться либо временно посредством ионных или водородных связей, либо навсегда посредством более прочных ковалентных связей.Связывание с этими молекулами способствует оптимальной форме и функционированию соответствующих ферментов. Двумя примерами этих типов вспомогательных молекул являются кофакторы и коферменты. Кофакторы — это неорганические ионы, такие как ионы железа и магния. Коферменты — это органические вспомогательные молекулы, имеющие базовую атомную структуру, состоящую из углерода и водорода. Подобно ферментам, эти молекулы участвуют в реакциях, не меняясь сами, и в конечном итоге перерабатываются и используются повторно. Витамины являются источником коферментов.Некоторые витамины являются предшественниками коферментов, а другие действуют непосредственно как коферменты. Витамин C является прямым коферментом множества ферментов, которые участвуют в создании важной соединительной ткани — коллагена. Следовательно, функция фермента частично регулируется обилием различных кофакторов и коферментов, которые могут поступать с пищей организма или, в некоторых случаях, вырабатываться организмом.
Ингибирование обратной связи в метаболических путях
Молекулы могут регулировать функцию ферментов разными способами.Однако остается главный вопрос: что это за молекулы и откуда они берутся? Как вы уже знаете, некоторые из них являются кофакторами и коферментами. Какие другие молекулы в клетке обеспечивают ферментативную регуляцию, такую как аллостерическая модуляция, а также конкурентное и неконкурентное ингибирование? Возможно, наиболее подходящими источниками регуляторных молекул для ферментативного клеточного метаболизма являются продукты самих клеточных метаболических реакций. Наиболее эффективным и элегантным образом клетки эволюционировали, чтобы использовать продукты собственных реакций для подавления активности ферментов с помощью обратной связи.Подавление обратной связи предполагает использование продукта реакции для регулирования его собственного дальнейшего производства (рис. 4.11). Клетка реагирует на обилие продуктов замедлением производства во время анаболических или катаболических реакций. Такие продукты реакции могут ингибировать ферменты, которые катализируют их производство с помощью механизмов, описанных выше.
Фигура 4.11 Метаболические пути — это серия реакций, катализируемых множеством ферментов. Ингибирование обратной связи, когда конечный продукт пути ингибирует вышестоящий процесс, является важным регуляторным механизмом в клетках.
Производство как аминокислот, так и нуклеотидов контролируется посредством ингибирования с обратной связью. Кроме того, АТФ является аллостерическим регулятором некоторых ферментов, участвующих в катаболическом распаде сахара, процессе, который создает АТФ. Таким образом, когда АТФ в избытке, клетка может предотвратить производство АТФ. С другой стороны, АДФ служит положительным аллостерическим регулятором (аллостерическим активатором) для некоторых из тех же ферментов, которые ингибируются АТФ. Таким образом, когда относительные уровни АДФ высоки по сравнению с АТФ, клетка начинает производить больше АТФ за счет катаболизма сахара.
понятий метаболизма | SpringerLink
ChapterПервый онлайн:
Abstract
Хотя группа виталистов первоначально использовала термин «органический» для соединений, производимых только организмами, позже он был использован для соединений углерода. Wohler (1928) обнаружил, что мочевина, которая раньше считалась производимой только в живых существах, также может быть произведена в лаборатории из аммиака и бикарбоната. В 1897 году немецкие химики Эдуард Бюхнер и Ганс Бюхнер продемонстрировали, что ферментация может осуществляться с помощью бесклеточного экстракта дрожжей .Эти наблюдения привели к развитию науки биохимии. В начале двадцатого века, благодаря открытию различных метаболических путей, в биохимии преобладала органическая химия, за которой следовали энзимология и биоэнергетика . Некоторые из аналитических методов, которые сделали возможным изучение биохимии, включали выделение органелл, высокоэффективную жидкостную хроматографию, электрофорез, использование радиоактивных индикаторов, методы трансформации растений с использованием Agrobacterium tumefaciens, подавление гена , прямую генетику, обратную генетику, масс-спектрометрию, и микроматрица ДНК, среди прочего.С помощью вычислительных технологий теперь можно полностью понять взаимосвязь метаболических путей.
Ключевые слова
Аллостерическая регуляция Биоэнергетика Кофактор Сопряженные реакции Свободная энергия Гиббса Холофермент Метаболическое каналирование Метаболическая избыточность Константа Михаэлиса-Ментен Редокс-потенциалЭто предварительный просмотр содержимого подписки,
войдите в систему, чтобы проверить доступ.
Вопросы с множественным выбором
- 1.
Биоэнергетика относится к:
- (a)
Обмен энергией между клеткой и окружающей средой
- (b)
Наука, которая занимается преобразованием энергии внутри клетки
- (в)
Выделение энергии при химической реакции
- (г)
- (a)
- 2.
Живая клетка является открытой системой, потому что:
- (a)
Она не обменивается энергией или веществом с окружающей средой.
- (b)
Он может обмениваться энергией и веществом с окружающей средой.
- (c)
Он может обмениваться энергией с окружающей средой, но не с веществом.
- (d)
Он может обмениваться веществом, но не энергией с окружающей средой.
- (a)
- 3.
Согласно второму закону термодинамики, произойдет спонтанная реакция:
- (a)
Когда менее сложные молекулы будут преобразованы в более сложные.
- (б)
Когда происходит поглощение энергии из окружающей среды.
- (c)
Молекулы с более высокой энтропией превращаются в молекулы с меньшей энтропией.
- (d)
Молекулы с меньшей энтропией превращаются в молекулы с высокой энтропией.
- (a)
- 4.
ΔG клеточной реакции будет отрицательным, если:
- (a)
Продукты реакции имеют меньшую энтропию, чем реагенты.
- (б)
Продукты реакции обладают большей энтропией, чем реагенты.
- (c)
Реакция не является спонтанной.
- (d)
Для протекания реакции требуется подвод энергии.
- (a)
- 5.
Какое из следующих утверждений верно?
- (a)
В живой клетке константа равновесия поддерживается равной 0.
- (b)
Свободная энергия — это полная энергия, присутствующая в молекуле.
- (c)
Свободная энергия — это энергия, изотермически доступная для выполнения работы.
- (d)
ΔG ° ‘ определяется как изменение свободной энергии во время реакции, которая не находится в равновесии.
- (a)
- 6.
Высокоэнергетическая связь (~) АТФ означает:
- (a)
Для образования этой связи требуется энергия.
- (б)
Гидролиз этой связи высвобождает энергию.
- (c)
Продукты гидролиза обладают меньшей энергией, чем сама молекула.
- (г)
Продукты гидролиза обладают большей энергией, чем сама молекула.
- (a)
- 7.
АТФ — это молекула с высокой энергией, потому что:
- (a)
Это нуклеозидтрифосфат.
- (б)
АТФ более резонансно стабилизирован, чем продукты его гидролиза.
- (c)
АТФ присутствует в клетке в виде комплекса Mg-ATP.
- (г)
Продукты его гидролиза стабилизируются за счет резонанса.
- (a)
- 8.
В окислительно-восстановительной реакции движутся электроны:
- (a)
От соединений с более положительным окислительно-восстановительным потенциалом к соединениям с менее положительным окислительно-восстановительным потенциалом
- (b)
От соединений с меньшим положительным окислительно-восстановительным потенциалом до более положительного окислительно-восстановительного потенциала
- (c)
От соединений с меньшим отрицательным окислительно-восстановительным потенциалом до соединений с более отрицательным окислительно-восстановительным потенциалом
- (d)
- (a)
- 9.
В молекуле фермента, состоящей из белковой и небелковой структуры, белковая часть известна как:
- 10.
Небелковая структура, ковалентно присоединенная к белковой части молекулы фермента, называется:
- 11.
Константа Михаэлиса (K
m) фермента:
- (a)
Концентрация субстрата, при которой фермент полностью насыщен
- (b)
Концентрация субстрата при где V max составляет половину максимального значения
- (c)
Концентрация фермента при V max
- (d)
Концентрация фермента, при которой V max составляет половину максимальной
- (a)
Предлагаемая дополнительная литература
Jones RL, Ougham H, Thomas H, Waaland S (2013) Молекулярная жизнь растений.Wiley-Blackwell, Chichester, pp 42–70
Google ScholarNelson DL, Cox MM (2017) Принципы биохимии Ленингера, 7-е изд. W.H. Freeman, New York, pp 495–525
Google ScholarVoet DJ, Voet JG, Charlotte WP (2008) Принципы биохимии, 3-е изд. Wiley, Hoboken, pp. 448–484
Google Scholar
Информация об авторских правах
© Springer Nature Singapore Pte Ltd. 2018
Авторы и аффилированные лица
- 1.Кафедра ботаникиКирори Мал Колледж, Университет Дели Нью-ДелиИндия
Границы | Проблемы городского метаболизма: устойчивость и благополучие в городах
Концепция городского метаболизма восходит к девятнадцатому веку (Wolman, 1965; Odum, 1996), поскольку города стали двигателем экономического развития и благосостояния людей. Термин городской метаболизм, как подчеркнули Сеспедес Рестрепо и Моралес-Пинсон (2018), «представляет собой концепцию, в которой город использует биологическое понятие, относящееся к внутренним процессам, посредством которых живые организмы поддерживают непрерывный обмен вещества и энергии с окружающей средой. для обеспечения работы, роста и воспроизводства.«Города — это сложные экосистемы, полностью естественные, даже если мы иногда думаем, что созданные человеком структуры не являются природой. Понять города намного легче, если мы осознаем, что они представляют собой естественные экосистемы, подчиняющиеся тем же законам (принцип максимальной мощности Лотки, эволюция Дарвина, естественный отбор), что и все другие части и виды. Хотя города покрывают <2% поверхности Земли, они потребляют около 78% энергии, доступной на планете, к которой необходимо добавить количество материалов и продуктов (продукты питания, строительные минералы, металлы и т. Д.)), которые косвенно также требуют потребления энергии. Эта цифра может быть объяснена с экономической и социальной точек зрения, поскольку города предлагают жителям новые возможности для бизнеса, образования, здравоохранения, безопасности, социальной жизни. Поддержка этой деятельности (и связанных с ней выгод) требует огромных потоков ресурсов, что чаще всего приводит к экологическому стрессу как на местном, так и на глобальном уровне. Таким образом, оценка метаболизма ресурсов городской системы, хотя и очень трудная из-за ее сложности, становится все более важной не только в отношении взаимосвязи с окружающей средой как источника или поглотителя, но и в отношении внутренней динамики обмена ресурсами между компонентами города. секторы, а также конкуренция между городами за ограниченные ресурсы.За последние несколько лет наблюдается рост интереса к городам и их использованию энергии и материалов - отчасти из-за проблем, связанных с изменением климата, а отчасти из-за социальных проблем, обусловленных социальными и организационными проблемами, порожденными огромными миграционными потоками из сельских районов в другие. городские районы. Города играют центральную роль в сокращении выбросов CO 2 и борьбе с изменением климата, одним из наиболее серьезных вызовов, с которыми в настоящее время сталкивается наше общество (Zhang et al., 2018).Изменение климата — не единственная проблема, определяемая городами и затрагивающая их: заброшенность сельских районов, потребность в воде, отводимой от естественных путей, концентрация городских отходов и потребность в соответствующих способах их удаления, потребность в транспорте. и связанные с этим ограничения по пространству, энергии и загрязнению воздуха, огромное количество ресурсов для поддержки городского образа жизни (образование, медицинские услуги, цепочка поставок продуктов питания и промышленных товаров и т. д.) не только требуют энергии и вызывают загрязнение, но и являются источником загрязнения. потенциальный источник новой рыночной динамики, реорганизации рабочей силы, конфликтов, новых инструментов управления и дорожных карт, спроса на общественные услуги и, наконец, упадка прежних культурных и социальных моделей и их замены новыми (или ничем, что еще хуже).Мониторинг притока и оттока ресурсов и понимание того, как они соотносятся с населением, доступностью ресурсов и пропускной способностью окружающей среды, имеют решающее значение для осознанной и заинтересованной политики устойчивого развития городов и управления. Реорганизация городских структур, связывающая биофизические аспекты с социальными аспектами благополучия сообществ, является приоритетной задачей при разработке политики во всем мире. Чтобы влиять на метаболизм в городах, нам необходимо лучше понимать отношения между обществами, окружающей средой, массой и потоками энергии (производство и потребление), не недооценивая фактор роста населения.Благополучие, а не богатство или изобилие, будет ключевым словом для будущего городов, а именно осознанных и сбалансированных отношений городского населения с окружающей средой, ресурсами, другими видами, чтобы способствовать справедливому и равноправному доступу к социальным услугам. , улучшение культуры, экономики и здоровья не обязательно связано с увеличением потребления ресурсов на душу населения.
Распределение человеческого населения по всему миру ясно показывает, что все больше и больше людей переезжают из сельской местности в города, что ведет к росту проблем (и осведомленности о) доступности и использования энергии и ресурсов, городскому развитию и целостности окружающей среды (Cui, 2018) .Согласно отчетам Организации Объединенных Наций, к 2030 году будут существовать 43 мегаполиса (т.е. города с населением более 10 миллионов человек), большинство из которых в развивающихся странах (United Nations, 2018b). Ожидается, что наибольшая часть мирового населения будет проживать в городах или мегаполисах, и ожидаемое значительное изменение стиля жизни, цепочек поставок, моделей управления, общественной организации, производства отходов, образовательных инструментов и структур и многого другого в общей экологической, экономической и социальной динамике, трансформируются в острую необходимость во всесторонних исследованиях устойчивости и благополучия городов.
Городские системы и города представляют собой сложные образования со своим собственным метаболизмом, чрезвычайно зависимыми от различных материальных и энергетических ресурсов извне (Currie et al., 2017). Связь воды и энергии в городской системе с учетом моделей потребления воды и энергии в зданиях, жилых секторах и производственных секторах может дать важную информацию для устойчивого городского планирования (Fan et al., 2019). Метаболизм ресурсов влияет и зависит от того, как городские сообщества самоорганизуются и принимают решения, потребляют и сохраняют, деградируют и перерабатывают ресурсы, совместно используют общественные услуги, проектируют и используют инфраструктуры.Тем не менее, общественный аспект и управление метаболизмом ресурсов все еще плохо изучены. Вторая важная тема — это то, как городские системы соотносятся с окружающими сельскими районами и дикой природой, уважают их, помогают им развиваться и сохранять свои модели целостности (Wei et al., 2015). Наконец, много было написано о замедлении роста активов, населения и потребления, но это очень сильно ограничивалось количественным замедлением, без изучения все еще возможных моделей качественного роста с точки зрения качества работы, безопасности, взаимодействия с сообществом, совместного использования. , счастье, справедливый доступ к ресурсам на всех уровнях и совместные процессы.Это, вероятно, самый сложный, но также и самый важный аспект устойчивости и стабильности городов. Интегрированное городское управление посредством надлежащего управления и экологической оценки преимуществ и компромиссов в отношении планирования развития, пространственного планирования инфраструктуры, а также соответствующего вспомогательного вмешательства становится фундаментальным (Fan et al., 2019). Вместо того, чтобы просто заниматься городским развитием и сохранением экологии, градостроители всегда должны помнить об аспектах устойчивости, связанных с качеством и типологией метаболизма (Cui, 2018).Проблема не в том, чтобы максимально увеличить или обеспечить огромное энергоснабжение или максимально возможное количество и разнообразие продуктов питания или максимально быстрый транспорт, а в том, чтобы оптимизировать на всех уровнях доступ всего городского населения к услугам, товарам, образованию, отдыху, здоровью. услуг, качества окружающей среды в достаточной степени для обеспечения качества жизни, стабильности, справедливости и уважения.
Благополучие в городах, также называемое пригодностью для жизни в городе, представляет способность города обеспечивать адекватные условия для процветания и хорошего качества жизни граждан (del Mar Martínez-Bravo et al., 2019). Доступность ресурсов и их справедливое распределение / распределение, индивидуальное восприятие, различные организации и условия сообщества (например, уровень образования, общественные услуги, наличие свободного времени, готовность зеленых насаждений, среди прочего), экологическая осведомленность, социальное и экономическое развитие это взаимосвязанные вопросы, которые сильно влияют на метаболизм города. Устойчивость и благополучие городских территорий были предметом обсуждения как в научном мире (Wei et al., 2015; del Mar Martínez-Bravo et al., 2019), а также политические дебаты (United Nations, 2018a) из-за решающей связи между ростом, эксплуатацией природных ресурсов (использование невозобновляемых источников энергии и материалов) и последствиями для состояния среда.
Городские системы широко исследовались с помощью нескольких различных экономических и биофизических подходов. Показатели эффективности для городов, регионов и стран были разработаны на основе хорошо известных методов оценки, иногда интегрированных в специальный инструментарий: воплощенная энергия, анализ материальных потоков (MFA), анализ жизненного цикла (LCA), выбросы CO 2 , затраты / выгода и экономическая отдача (Yong et al., 2012). Эти индикаторы по отдельности или в сочетании не обязательно обеспечивают полностью адекватную характеристику экологической целостности городской системы и использования ресурсов, поскольку они не были предназначены для оценки целостных систем, замкнутых циклов и характеристик обратной связи, которые являются ключевыми характеристиками устойчивого городского развития. экономика (Yong et al., 2013). Чаще всего в этих подходах не учитываются качество и характеристики потока, а также сложность взаимодействия между природной средой и социально-экономическими системами (Huang et al., 2006). Поэтому очень важно обсудить и глубже понять сложность будущего развития и управления городами, выходя далеко за рамки линейного и одномерного подхода простого измерения городского населения, потребления энергии или ВВП. Связь городского метаболизма со стратегиями политики необходима для измерения и изменения показателей устойчивости городов, развития междисциплинарной практики городского метаболизма (Dijst et al., 2018). На социальное поведение и индивидуальные предпочтения сильно влияет конкуренция за доступные ресурсы, пространство, возможности.Невозможно иметь образованное следующее поколение без вложений в образование; невозможно наслаждаться здоровой пищей, не инвестируя ресурсы в более чистое производство; Невозможно иметь стабильное и счастливое сообщество без обеспечения равного доступа к основным ресурсам. Как следствие, надлежащее и справедливое управление ресурсами, вероятно, повлияет (и будет затронуто) образованием, демократией, стабильностью, чувством принадлежности, сбалансированным взаимодействием с окружающими территориями и населением.
Одной из наиболее сложных задач ближайшего будущего является разработка новых экологических стратегий и альтернатив ископаемым источникам энергии с целью удовлетворения потребностей и ожиданий нынешнего и будущих поколений на устойчивой основе.Во времена сокращения ресурсной базы более круговые и гибкие модели предоставления ресурсов в сочетании с изменениями образа жизни могут стать основой менее ресурсоемкой городской жизни (Zucaro et al., 2014). Поскольку городская территория представляет собой сложную систему, связанную с различными производственно-потребительскими цепочками поставок, исследование экологических, экономических и социальных аспектов с использованием общесистемной основы анализа для обеспечения устойчивости является ключевым вопросом для защиты благосостояния и разработки адекватных политических решений. .Даже если энергоэффективность и внедрение возобновляемых источников энергии, а также схемы рециркуляции материалов (круговая экономика), кажется, развиваются быстро и эффективно, многое еще предстоит сделать. Общество представляет собой сложную сеть производственной и потребительской деятельности, связанных друг с другом взаимодействующими цепочками поставок и торговли. Необязательно, что повышение энергоэффективности в одной точке сети (например, за счет рециркуляции или обмена ресурсами или реализации схем циркулярной экономики) приводит к общему повышению эффективности на уровне всей системы; Аналогичным образом, уменьшение одного вида воздействия (истощение энергии, истощение водных ресурсов, потребность в земле, выбросы и т. д.)) не обязательно приводит к общему снижению воздействия системы. Следовательно, исследования городского метаболизма должны срочно подключиться к разработке решений по устойчивости для социально-экологической динамики, описывающих и понимающих взаимозависимости и динамику городов и экосистем (John et al., 2019).
Обсуждение, диалог, совместное управление и соответствующие процессы решения проблем, основанные на подходящей количественной и качественной информации, являются важными аспектами для разработки соответствующего процесса принятия решений для моделей городского метаболизма, которые восстанавливаются как по замыслу, так и по замыслу, чтобы для достижения целей устойчивости и благополучия.
Устойчивое развитие — это то, что произойдет в городах или вряд ли произойдет. Переход от риторики устойчивости к реализации сообщества и благополучия — это то, что не произойдет само по себе и не может быть решено с помощью линейных и детерминированных подходов. Подобно экосистемам, городам требуется оптимизация использования ресурсов во всех секторах и на всех уровнях, взаимосвязь между компонентами и процессами, сбалансированное сотрудничество между городскими и сельскими районами, модели восстановления и рециркуляции остатков и отходов, импульсные стратегии в соответствии с колеблющейся доступностью ресурсов, образовательные программы, ориентированные на более глубокое понимание сложных экологических ограничений, стратегии качественного роста вместо стремления к неограниченному и вряд ли устойчивому количественному росту.
Взносы авторов
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Сеспедес Рестрепо, Дж. Д. и Моралес-Пинсон, Т. (2018).Городской метаболизм и устойчивость: прецеденты, генезис и перспективы исследования. Ресурс. Консерв. Рецикл . 131, 216–224. DOI: 10.1016 / j.resconrec.2017.12.023
CrossRef Полный текст | Google Scholar
Цуй, X. (2018). Как города могут поддерживать устойчивость: библиометрический анализ городского метаболизма. Ecol. Инд . 93, 704–717. DOI: 10.1016 / j.ecolind.2018.05.056
CrossRef Полный текст | Google Scholar
Карри, П. К., Мусанго, Дж.К., и Мэй, Н. Д. (2017). Городской метаболизм: обзор со ссылкой на Кейптаун. Города 70, 91–110. DOI: 10.1016 / j.cities.2017.06.005
CrossRef Полный текст | Google Scholar
дель Мар Мартинес-Браво, М., Мартинес-дель-Рио, Х. и Антолин-Лопес, Р. (2019). Компромиссы между устойчивостью городов, загрязнением и пригодностью для жизни в европейских городах. J. Clean. Прод . 224, 651–660. DOI: 10.1016 / j.jclepro.2019.03.110
CrossRef Полный текст | Google Scholar
Дийст, М., Worrell, E., Böcker, L., Brunner, P., Davoudi, S., Geertman, S., et al. (2018). Изучение городского метаболизма — к междисциплинарной перспективе. Ресурс. Консерв. Рецикл . 132, 190–203. DOI: 10.1016 / j.resconrec.2017.09.014
CrossRef Полный текст | Google Scholar
Фань, Дж. Л., Конг, Л. С., Ван, Х. и Чжан, X. (2019). Обзор взаимосвязи воды и энергии с точки зрения городского метаболизма. Ecol. Модель . 392, 128–136. DOI: 10.1016 / j.ecolmodel.2018.11.019
CrossRef Полный текст | Google Scholar
Хуанг, С. Л., Ли, К. Л., и Чен, К. В. (2006). Социально-экономический метаболизм в Тайване: неожиданный синтез против анализа материального потока. Ресурс. Консерв. Рецикл . 48, 166–196. DOI: 10.1016 / j.resconrec.2006.01.005
CrossRef Полный текст | Google Scholar
Джон Б., Людериц К., Ланг Д. Дж. И фон Верден Х. (2019). На пути к устойчивому метаболизму в городах. от понимания системы к трансформации системы. Ecol. Экон . 157, 402–414. DOI: 10.1016 / j.ecolecon.2018.12.007
CrossRef Полный текст | Google Scholar
Одум, Х. Т. (1996). Экологический учет: принятие решений по чрезвычайным ситуациям и окружающей среде . Нью-Йорк, штат Нью-Йорк: Wiley-Indersciene.
Google Scholar
Организация Объединенных Наций (2018b). Мировые перспективы урбанизации: редакция 2018 г. .
Вэй, Х., Шэнхуэй, К., Яримэ, М., Хашимото, С., и Манаги, С. (2015).Улучшение городского метаболизма исследования для устойчивой городской трансформации. Environ. Technol. Иннов . 4, 62–72. DOI: 10.1016 / j.eti.2015.04.004
CrossRef Полный текст | Google Scholar
Юн, Г., Фу, Дж., Саркис, Дж., И Сюэ, Б. (2012). На пути к национальной системе показателей замкнутой экономики в Китае: оценка и критический анализ. J. Clean. Прод . 23, 216. doi: 10.1016 / j.jclepro.2011.07.005
CrossRef Полный текст | Google Scholar
Йонг, Г., Саркис, Дж., Улгиати, С., и Чжан, П. (2013). Измерение круговой экономики Китая. Наука 339, 1526–1527. DOI: 10.1126 / science.1227059
CrossRef Полный текст | Google Scholar
Чжан Ю., Вуа К. и Фатх Б. Д. (2018). Обзор пространственного анализа городского углеродного метаболизма Ecol . Модель . 371, 18–24. DOI: 10.1016 / j.ecolmodel.2018.01.005
CrossRef Полный текст
Зукаро, А., Рипа, М., Меллино, С., Асьоне, М., и Ульгиати, С. (2014). Показатели использования городских ресурсов и экологической результативности. Приложение декомпозиционного анализа. Ecol. Инд . 47, 16–25. DOI: 10.1016 / j.ecolind.2014.04.022
CrossRef Полный текст | Google Scholar
Метаболизм: определение и обзор — видео и стенограмма урока
Катаболизм
Представьте, что вы едите кусок пиццы пепперони. В этом куске у вас есть несколько ключевых питательных веществ, необходимых для жизни.У вас есть белка , которые содержат аминокислоты . У вас есть крахмала , которые содержат сахар для энергии. У вас также есть жира , которые можно использовать для производства энергии и гормонов. Но для того, чтобы получить необходимые компоненты (например, аминокислоты и сахар), у вас должен быть способ расщепить эти материалы.
Катаболизм — это процесс разрушения материалов в организме. Когда мы перевариваем пищу, мы расщепляем (или катаболизируем) белки, жиры и крахмалы, чтобы получить полезные компоненты.Это делается путем добавления воды к химическим связям в этих веществах, что представляет собой процесс, известный как гидролиз . Пищевые продукты, которые не перевариваются или не могут быть расщеплены, не могут подвергаться гидролизу в организме.
Анаболизм
Для того, чтобы наши тела функционировали, мы должны иметь возможность производить необходимые нам материалы из тех, которые мы перевариваем. Катаболизируя нашу пищу, мы остаемся с основными компонентами, которые нам нужны для жизни. Например, если у нас есть поврежденные ткани, мы должны производить белки для их лечения.Следовательно, аминокислоты, которые мы получаем с пищей, могут использоваться для восстановления белков. Сахар, полученный путем переваривания крахмалов, может использоваться для производства более крупных сахаров для хранения. Жиры можно воспроизводить для хранения энергии. Все эти материалы производятся в результате анаболизма.
Анаболизм — это процесс производства более крупных веществ из более мелких. Это в некотором смысле противоположность катаболизму в том смысле, что он достигается за счет удаления воды для создания связей. Этот процесс известен как дегидратационный синтез .Посредством синтеза обезвоживания мы можем производить белки, жиры и другие необходимые материалы.
Метаболизм непродовольственных товаров также является важным процессом, необходимым для выживания. Хотя потребление пищи необходимо для жизни, употребление многих лекарств и ядов может нанести вред здоровью организма. Следовательно, способность расщеплять эти материалы и выводить их из организма также является частью метаболизма.
Например, печень отвечает за метаболизм и детоксикацию алкоголя, который может быть ядовитым для организма.Точно так же почки , отвечают за удаление этих материалов через образование мочи. Эта форма метаболизма так же важна, как и процессы, которые используются для усвоения питательных веществ.
Несколько физиологических факторов могут влиять на скорость и эффективность метаболизма в нашем организме. Во-первых, количество и тип потребляемой пищи влияют на скорость метаболизма. Например, жирная пища дольше переваривается в организме. Кроме того, большие порции перевариваются намного медленнее, чем маленькие.
Возраст и пол могут влиять на скорость метаболизма в организме. Мужчины разрушают и восстанавливают материалы быстрее, чем женщины, а молодые люди, как правило, усваивают материалы быстрее, чем люди старшего возраста. Дополнительные факторы включают температуру, наследственность и заболевания, которые могут увеличивать или уменьшать скорость метаболизма в организме.
Резюме
Метаболизм — это сумма всех химических реакций в организме. Катаболизм — это процесс разрушения материалов с целью извлечения необходимых компонентов. Анаболизм , с другой стороны, представляет собой процесс создания более крупных материалов из более мелких компонентов. Метаболизм важен для удаления ядовитых материалов из организма, а также для сохранения здоровых материалов. Наконец, несколько факторов, включая температуру, наследственность, возраст и пол, могут повлиять на скорость метаболизма в организме.
Метаболизм | Очерки биохимии
Фенилкетонурия (ФКУ) и дефицит ацил-КоА-дегидрогеназы со средней длиной цепи (MCADD) — два наиболее часто наследуемых метаболических нарушения, которым страдает примерно 1 из 10000 новорожденных в США.К.
ФКУ — это аминокислотное заболевание, вызванное дефицитом фермента фенилаланингидроксилазы, вызывающим ферментативный блок. Это приводит к снижению метаболизма аминокислоты фенилаланина, вызывая повышенное накопление в крови и головном мозге. Если не лечить новорожденных, это может вызвать задержку развития или повреждение головного мозга. Лечение начинается рано с диеты с низким содержанием белка, дополненной смесью аминокислот с удаленным фенилаланином.Однако небольшая часть людей с диагнозом ФКУ не отвечает на этот предложенный вид лечения. Эти люди обычно имеют дефекты синтеза дигидроптеридинредуктазы или биоптерина, вызывающие нарушение функции фенилаланингидроксилазы. Эти люди также обычно имеют дефекты тирозингидроксилазы, что может привести к дефициту нейротрансмиттеров. Затем этим пациентам требуются дополнительные добавки с нейротрансмиттерами, а также диета с низким содержанием фенилаланина.
MCADD — это пожизненное состояние, которое возникает из-за мутации ацил-CoA дегидрогеназы со средней длиной цепи (MCAD) β-окисления жирных кислот. Эта мутация нарушает расщепление жирных кислот со средней длиной цепи в ацетил-КоА. Потеря или недостаточность MCAD снижает окисление жирных ацил-CoA, которые содержат более шести атомов углерода, поскольку первая стадия дегидрирования β-окисления не может происходить. Используя тандемную масс-спектрометрию, можно увидеть, что профиль жирных кислот крови в MCADD показывает накопление C6, C8 и C10: 1.MCADD является основной причиной гипокетотической гипогликемии и может вызывать нарушение функции печени с метаболическим ацидозом, гипераммониемией и внезапной смертью. MCADD особенно опасен во время голодания, когда организм использует запасы гликогена, а свободные жирные кислоты высвобождаются из жировой ткани для получения энергии. Сниженная способность метаболизировать средние жирные кислоты значительно снижает доступность субстратов для кетогенеза, синтеза АТФ и цикла TCA при низкой энергии. Накопление промежуточных продуктов жирных кислот подавляет глюконеогенез, усугубляя гипогликемию.Это накопление также может способствовать сердечно-сосудистым и неврологическим осложнениям, обнаруживаемым в этих условиях. Лечение пациентов с MCADD включает потребление напитков с высоким содержанием сахара и избегание длительных периодов голодания.
Последнее наследственное нарушение обмена веществ, которое мы обсуждаем, встречается гораздо реже и встречается у 1 из 100 000 или 1 50000 новорожденных. Болезнь мочи кленового сиропа (MSUD) возникает из-за дефицита или снижения функции комплекса дегидрогеназы α-кетокислоты с разветвленной цепью (BCKAD).Это приводит к накоплению аминокислот с разветвленной цепью (BCAA), таких как лейцин, изолейцин и валин, в крови и моче. Название заболевания происходит от запаха мочи кленового сиропа из-за избытка BCAA. BCAA потребляются в рационе, богатом белком, в таких продуктах, как мясо, рыба, яйца и молоко. Обычно избыточные аминокислоты расщепляются через аминотрансферазы с разветвленной цепью (BCAT) на α-кетокислоты в митохондриях. На второй стадии катаболизма комплекс BCKAD инициирует окислительное декарбоксилирование α-кетокислот, что приводит к образованию ацетоацетата, ацетил-КоА и сукцинил-КоА.Нормальное функционирование катаболизма аминокислот необходимо для синтеза белка, передачи клеточных сигналов и метаболизма глюкозы. BCKAD состоит из четырех субъединиц. Мутации в каталитических компонентах BCKAD снижают его активность и, следовательно, увеличивают уровни BCAA, проявляясь как MSUD и вызывая дисфункцию иммунной системы, скелетных мышц и центральной нервной системы. По мере накопления токсичных метаболитов, таких как молочная кислота и аммиак, функция иммунных клеток подавляется, вызывая нарушение их регуляции.Скелетные мышцы поражены, как показали исследования, в которых обнаружено уменьшение диаметра мышечных волокон и поражения миофибрилл у крыс MSUD, однако его механизм полностью не изучен. Нарушение регуляции нервной системы, в частности, поражение головного мозга, было связано с накоплением токсичных метаболитов. Однако исследования показали, что образование азот-активных форм у пациентов с MSUD может вызывать морфологические изменения в клетках глиомы C6. Кроме того, у пациентов с MSUD обнаруживаются маркеры окислительного повреждения белков, ДНК и липидов, возможно, в результате продукции свободных радикалов.
.