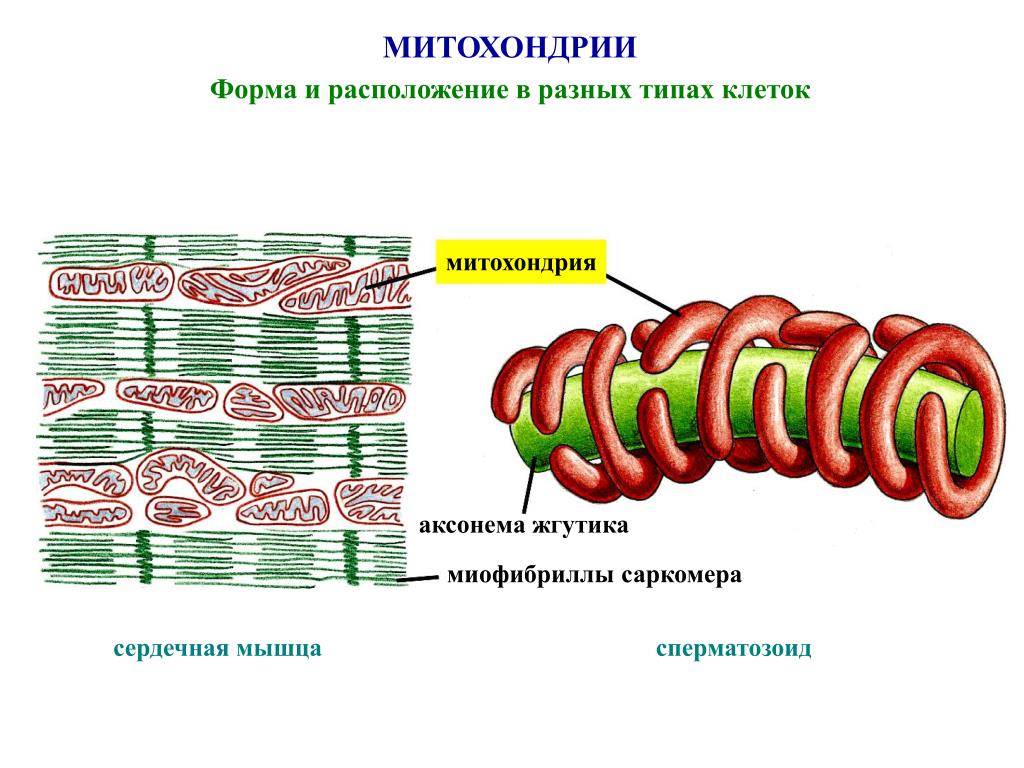
Методы гиперплазии миофибрилл в мышечных волокнах
В. Н. Селуянов, В. А. Рыбаков, М. П. Шестаков
Глава 4. Методы управления адаптационными процессами
Для управления адаптационными процессами в определенных клетках органов тела человека необходимо знать устройство органа, механизм его функционирования, факторы, обеспечивающие целевое направление адаптационных процессов. Модель организма спортсмена построена в предыдущих главах. На основе этой модели может быть реализовано теоретическое мышление в виде умозрительного или компьютерного (математического) моделирования. В ходе имитационного моделирования находятся различные варианты принятия управленческих решений, из которых в дальнейшем выбирается наиболее подходящая к данным условиям технология спортивной подготовки. Технология управления адаптационными процессами, реализуемая с помощью физических упражнений, характеризуется следующими параметрами: интенсивность сокращения мышц (ИС), средняя интенсивность упражнения (ИУ), продолжительность (П), интервал отдыха (ИО), количество повторений упражнения (КП), интервал отдыха до следующей тренировки (ИОТ).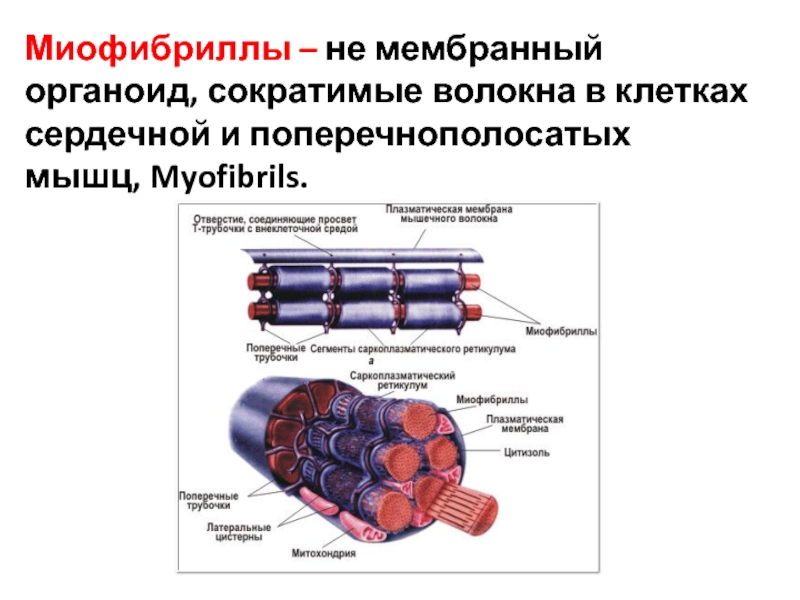
4.1. Методы гиперплазии миофибрилл в мышечных волокнах
Цель силовой подготовки — увеличить число миофибрилл в мышечных волокнах. Силовое воздействие человека на окружающую среду есть следствие функционирования мышц. Мышца состоит из мышечных волокон — клеток. Для увеличения силы тяги МВ необходимо добиться гиперплазии (увеличения) миофибрилл. Этот процесс возникает при ускорении синтеза и при прежних темпах распада белка. Исследования последних лет позволили выявить четыре основных фактора, определяющих ускоренный синтез белка в клетке:
- 1. Запас аминокислот в клетке. (Аминокислоты в клетке накапливаются после потребления пищи богатой белками.)
2. Повышенная концентрация анаболических гормонов в крови как результат психического напряжения (Holloszy et al.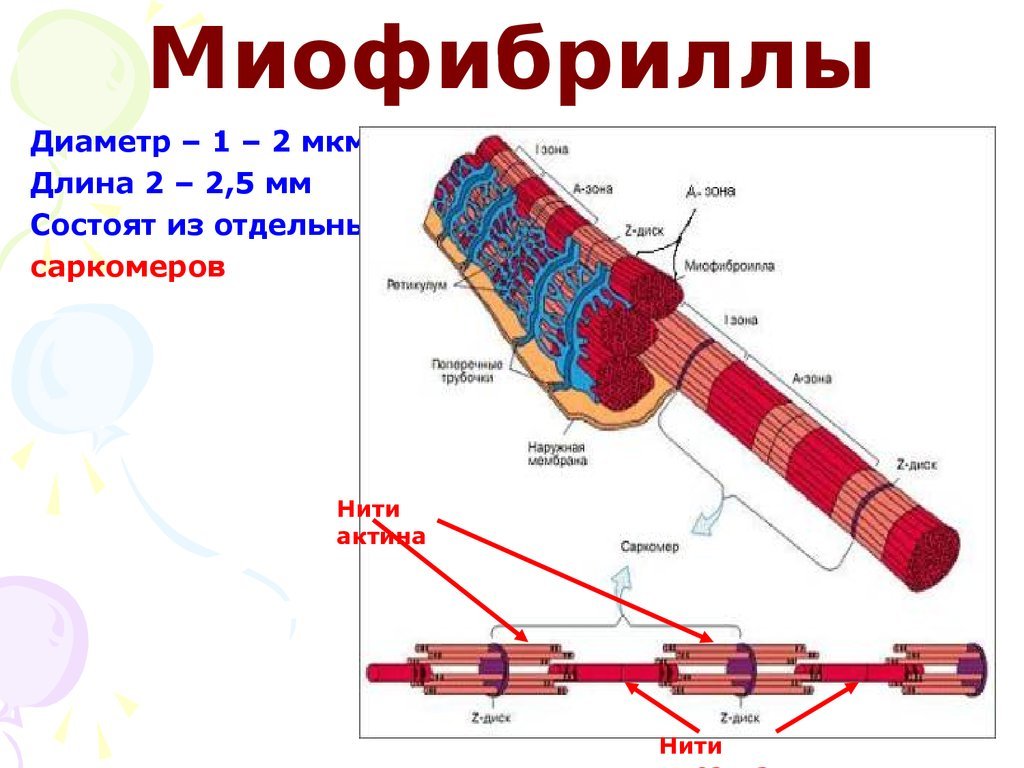 , 1971; Schants, 1986).
, 1971; Schants, 1986).
3. Повышенная концентрация «свободного» креатина в МВ (Walker, 1979).
4. Повышенная концентрация ионов водорода (Панин Л. Е., 1983).
Второй, третий и четвертый факторы прямо связаны с содержанием тренировочных упражнений.
Механизм синтеза органелл в клетке, в частности, миофибрилл, можно описать следующим образом.
В ходе выполнения упражнения энергия АТФ тратится на образование актин миозиновых соединений, выполнение механической работы. Ресинтез АТФ идет благодаря запасам КрФ. Появление свободного Кр активизирует деятельность всех метаболических путей, связанных с образованием АТФ (гликолиз в цитоплазме, аэробное окисление в различных митохондриях, например, миофибриллярных, а также в находящихся в ядрышке и на мембранах СПР). В быстрых мышечных волокнах (БМВ) преобладает мышечная лактат-дегидрогеназа (М ЛДГ), поэтому пируват, образующийся в ходе анаэробного гликолиза, в основном трансформируется в лактат. В ходе такого процесса в клетке накапливаются ионы Н.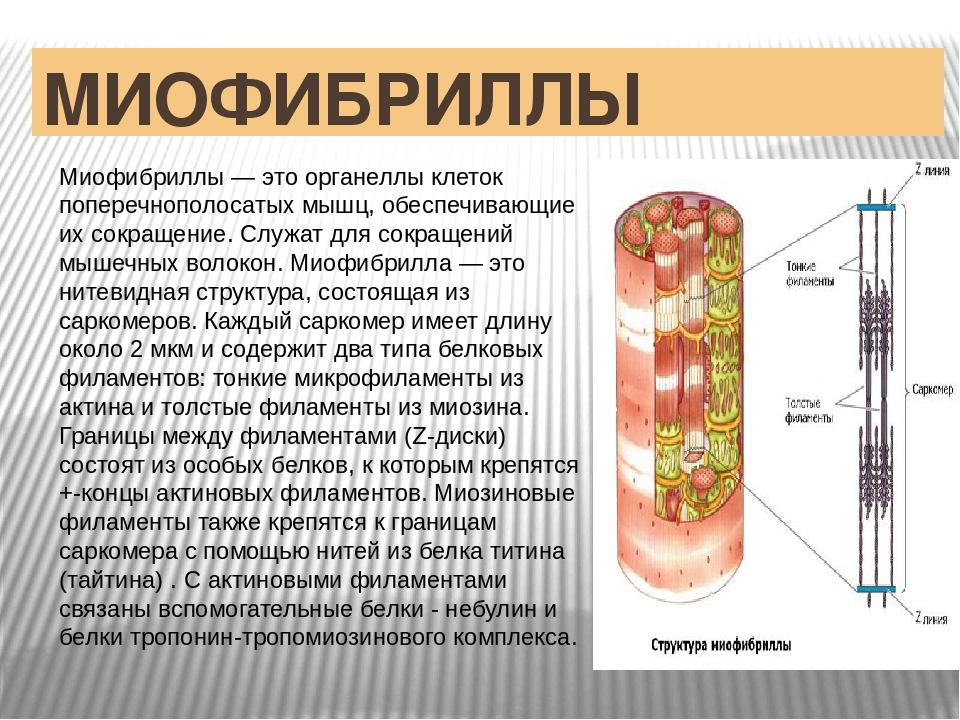
Наряду с важной ролью в определении сократительных свойств в регуляции энергетического метаболизма, накопление свободного креатина в саркоплазматическом пространстве служит мощным эндогенным стимулом, возбуждающим белковый синтез в скелетных мышцах (Walker J., 1979; Волков Н. И. с соав., 1983). Показано, что между содержанием сократительных белков и содержанием креатина имеется строгое соответствие. Свободный креатин, видимо, влияет на синтез и РНК, т. е. на транскрипцию в ядрышках МВ, либо активирует деятельность ядерных митохондрий, которые начинают в большей мере вырабатывать АТФ, которая используется для транскрипции ДНК (Walker, 1979).
Предполагается, что повышение концентрации ионов водорода вызывает лабилизацию мембран (увеличение размеров пор в мембранах, это ведет к облегчению проникновения гормонов в клетку), активизирует действие ферментов, облегчает доступ гормонов к наследственной информации, к молекулам ДНК (Панин Л. Е., 1983). В ответ на одновременное повышение концентрации Кр и Н интенсивнее образуются РНК. Срок жизни и РНК короток, несколько секунд в ходе выполнения силового упражнения плюс пять минут в паузе отдыха (Виру А. А., 1981). Затем молекулы и РНК разрушаются.
Е., 1983). В ответ на одновременное повышение концентрации Кр и Н интенсивнее образуются РНК. Срок жизни и РНК короток, несколько секунд в ходе выполнения силового упражнения плюс пять минут в паузе отдыха (Виру А. А., 1981). Затем молекулы и РНК разрушаются.
Теоретический анализ показывает, что при выполнении силового упражнения до отказа, например 10 приседаний со штангой с темпом одно приседание за 3–5 с, упражнение длится до 50 с. В мышцах в это время идет циклический процесс: опускание и подъем со штангой 1–2 с выполняется за счет запасов АТФ; за 2–3 с паузы, когда мышцы становятся малоактивными (нагрузка распространяется вдоль позвоночного столба и костей ног), идет ресинтез АТФ из запасов КрФ, а КрФ ресинтезируется за счет аэробных процессов в ОМВ и анаэробного гликолиза в ГМВ. В связи с тем, что мощность аэробных и гликолитических процессов значительно ниже скорости расхода АТФ, запасы КрФ постепенно исчерпываются, продолжение упражнения заданной мощности становится невозможным наступает отказ.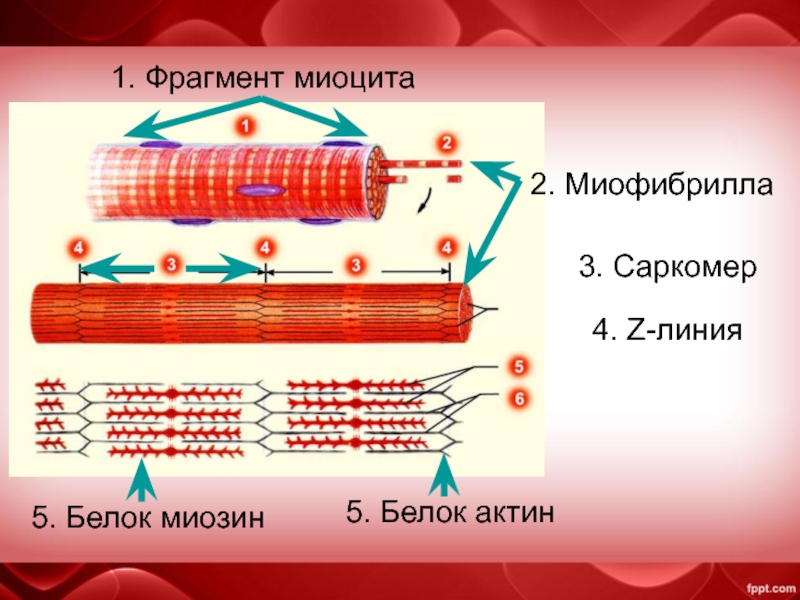
 В частности, в исследовании А. Salminen e. a. (1984) на крысах было показано, что интенсивный (гликолитический) бег вызывает некротические изменения и 4–5-кратное увеличение активности лизосомальных ферментов. Совместное действие ионов водорода и свободного Кр приводит к активизации синтеза РНК. Известно, что Кр присутствует в мышечном волокне в ходе упражнения и в течение 30–60 с после него, пока идет ресинтез КрФ. Поэтому можно считать, что за один подход к снаряду спортсмен набирает около одной минуты чистого времени, когда в его мышцах происходит образование и РНК. При повторении подходов количество накопленной и РНК будет расти, но одновременно с повышением концентрации ионов Н; поэтому возникает противоречие, то есть можно разрушить больше, чем потом будет синтезировано. Избежать этого можно при проведении подходов с большими интервалами отдыха или тренировках несколько раз в день с небольшим числом подходов в каждой тренировке.
В частности, в исследовании А. Salminen e. a. (1984) на крысах было показано, что интенсивный (гликолитический) бег вызывает некротические изменения и 4–5-кратное увеличение активности лизосомальных ферментов. Совместное действие ионов водорода и свободного Кр приводит к активизации синтеза РНК. Известно, что Кр присутствует в мышечном волокне в ходе упражнения и в течение 30–60 с после него, пока идет ресинтез КрФ. Поэтому можно считать, что за один подход к снаряду спортсмен набирает около одной минуты чистого времени, когда в его мышцах происходит образование и РНК. При повторении подходов количество накопленной и РНК будет расти, но одновременно с повышением концентрации ионов Н; поэтому возникает противоречие, то есть можно разрушить больше, чем потом будет синтезировано. Избежать этого можно при проведении подходов с большими интервалами отдыха или тренировках несколько раз в день с небольшим числом подходов в каждой тренировке.Вопрос об интервале отдыха между днями силовой тренировки связан со скоростью реализации и РНК в органеллы клетки, в частности, в миофибриллы.
Логика происходящего при выполнении силовой тренировки представляется в основном корректной, однако доказать ее истинность может лишь эксперимент. Проведение эксперимента требует затрат времени, привлечения испытуемых и др., а если логика окажется где-то порочной, то придется вновь проводить эксперимент. Понятно, что такой подход возможен, но малоэффективен. Более продуктивен подход с применением модели организма человека и имитационным моделированием физиологических функций и структурных, адаптационных перестроек в системах и органах. На ЭВМ возможно в короткое время систематически изучать процессы адаптации и проверять корректность планирования физической подготовки. Эксперимент же можно проводить уже после того, как будет ясно, что грубых ошибок в планировании не допущено.
Проведение эксперимента требует затрат времени, привлечения испытуемых и др., а если логика окажется где-то порочной, то придется вновь проводить эксперимент. Понятно, что такой подход возможен, но малоэффективен. Более продуктивен подход с применением модели организма человека и имитационным моделированием физиологических функций и структурных, адаптационных перестроек в системах и органах. На ЭВМ возможно в короткое время систематически изучать процессы адаптации и проверять корректность планирования физической подготовки. Эксперимент же можно проводить уже после того, как будет ясно, что грубых ошибок в планировании не допущено.
Из описания механизма должно быть ясно, что ОМВ и БГВ должны тренироваться в ходе выполнения разных упражнений, разными методиками.
Гистология.mp3 — Мышечные ткани
Слушать (13 464 Кб):
Общая гистология — скелетная, сердечная и гладкая мышечные ткани
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям.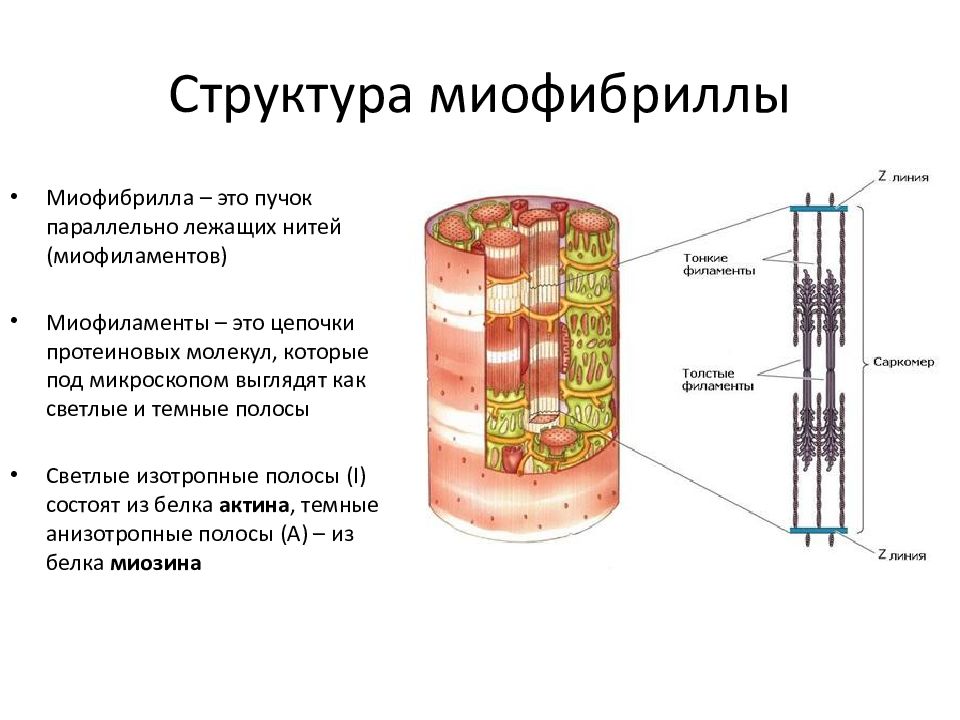 Они обеспечивают перемещения в пространстве всего организма в целом или его частей (пример – скелетная мускулатура) и движение органов внутри организма (пример – сердце, язык, кишечник).
Они обеспечивают перемещения в пространстве всего организма в целом или его частей (пример – скелетная мускулатура) и движение органов внутри организма (пример – сердце, язык, кишечник).
Свойством изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
Общая характеристика и классификация
Основные морфологические признаки элементов мышечных тканей — удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы — миофиламенты обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков — актина и миозина при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды.
Миоглобин — это белок-пигмент (наподобие гемоглобина), обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (и поступление кислорода при этом резко падает).
Запас источников энергии образуют гликоген и липиды.
Миоглобин — это белок-пигмент (наподобие гемоглобина), обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (и поступление кислорода при этом резко падает).
В основу классификации мышечных тканей положены два принципа — морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы: исчерченные мышечные ткани и гладкие мышечные ткани.
Поперечнополосатые (исчерченные) мышечные ткани. В цитоплазме их элементов миозиновые филаменты постоянно полимеризованы, образуют с актиновыми нитями постоянно существующие миофибриллы. Последние организованы в характерные комплексы — саркомеры. В соседних миофибриллах структурные субъединицы саркомеров расположены на одинаковом уровне и создают поперечную исчерченность.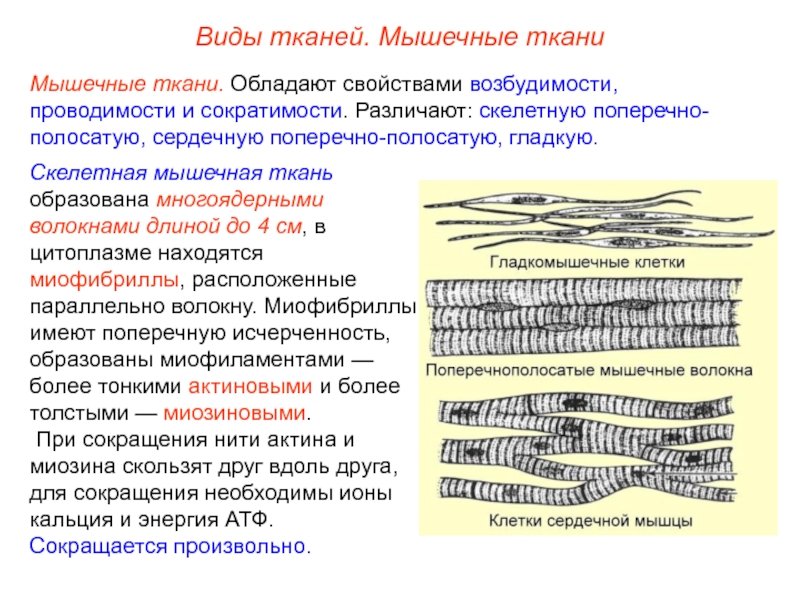 Исчерченные мышечные ткани сокращаются быстрее, чем гладкие.
Исчерченные мышечные ткани сокращаются быстрее, чем гладкие.
Гладкие (неисчерченные) мышечные ткани. Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине нитями.
В соответствии с гистогенетическим принципом в зависимости от источников развития (т.е. эмбриональных зачатков) мышечные ткани подразделяются на 5 типов:
- мезенхимные (из десмального зачатка в составе мезенхимы)
- эпидермальные (из кожной эктодермы и из прехордальной пластинки)
- нейральные (из нервной трубки)
- целомические (из миоэпикардиальной пластинки висцерального листка спланхнотома)
- соматические (миотомные)
Первые три типа относятся к подгруппе гладких мышечных тканей, четвертый и пятый — к подгруппе поперечнополосатых.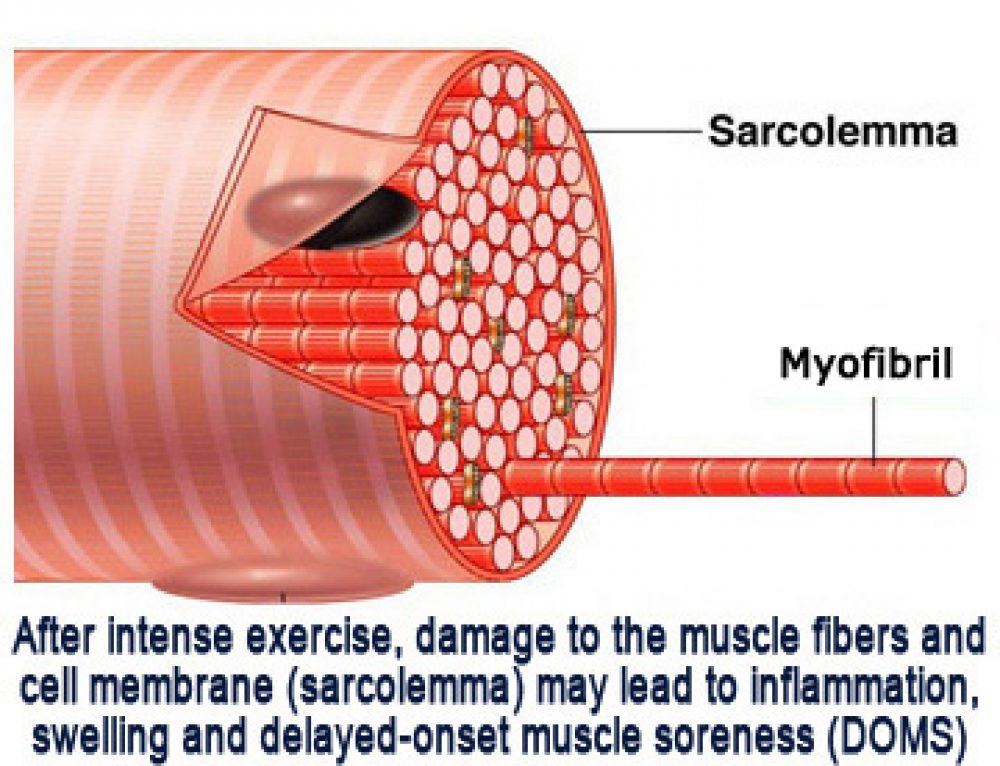
Поперечнополосатые мышечные ткани
Имеется две основные разновидности поперечнополосатых (исчерченных) тканей — скелетная мышечная ткань и сердечная мышечная ткань.
Скелетная мышечная ткань
Гистогенез
Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов — миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела.
В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами.
Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (или миосателлиты). Эти клетки располагаются на поверхности миосимпластов.
Строение
Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной.
Длина всего волокна может измеряться сантиметрами при толщине всего 50—100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
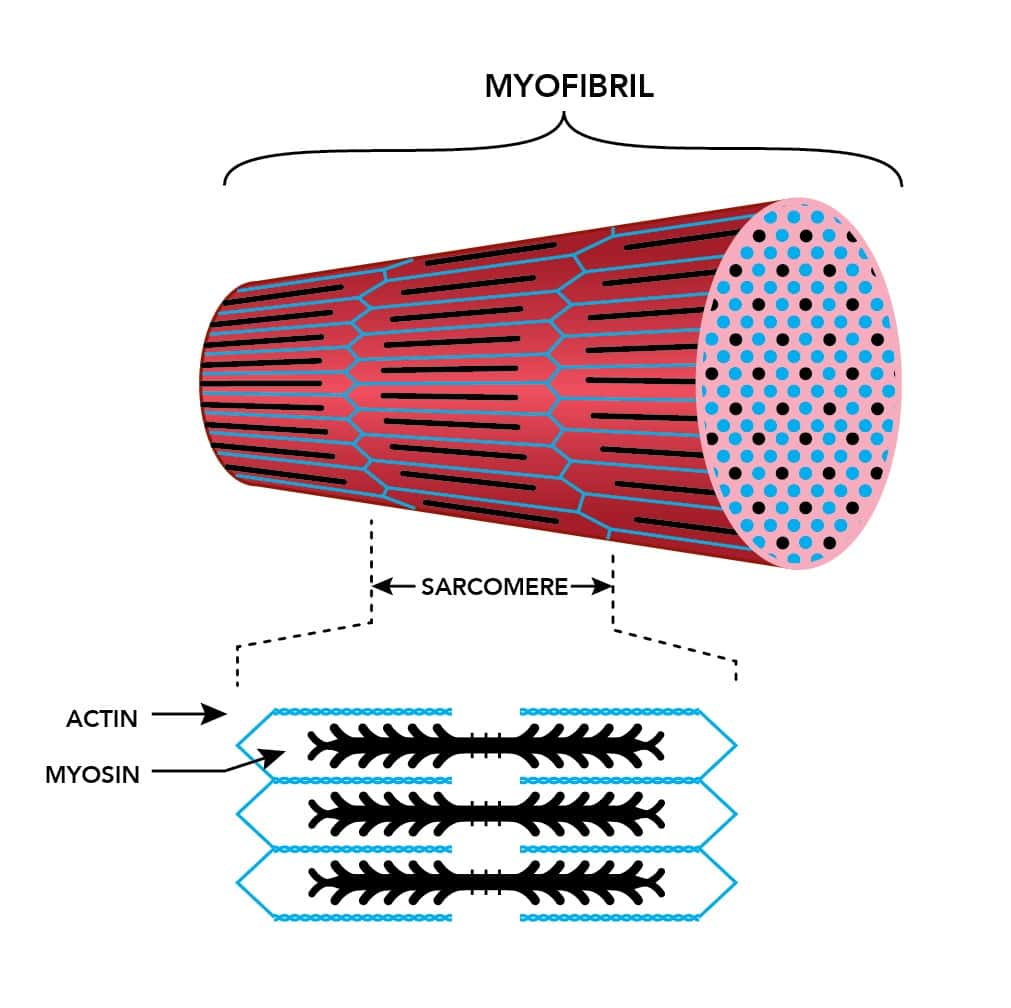
Саркомер — это структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски, имеющие неодинаковое лучепреломление (анизотропные A-диски и изотропные I-диски). Каждая миофибрилла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети, или саркоплазматического ретикулума. Соседние саркомеры имеют общую пограничную структуру — Z-линию (или телофрагму). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы тонких, актиновых, филаментов. От соседних Z-линий актиновые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию, или мезофрагму. В узлах этой М-линии закреплены концы толстых, миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию, или мезофрагму. В узлах этой М-линии закреплены концы толстых, миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Z-линии сближаются, саркомер укорачивается.
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в кортикальном слое цитоплазмы, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна.
Источником ионов кальция служат цистерны агранулярной эндоплазматической сети. Они вытянуты вдоль миофибрилл около каждого саркомера и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или (латеральные) L-цистерны.
С поверхности миосимпласта плазмолемма образует длинные трубочки, идущие поперечно в глубину клетки (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда клетка получает сигнал о начале сокращения, этот сигнал перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в цистернах саркоплазматического ретикулума и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет АТФ- АДФ-превращений. Роль АТФазы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
Когда клетка получает сигнал о начале сокращения, этот сигнал перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в цистернах саркоплазматического ретикулума и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет АТФ- АДФ-превращений. Роль АТФазы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма.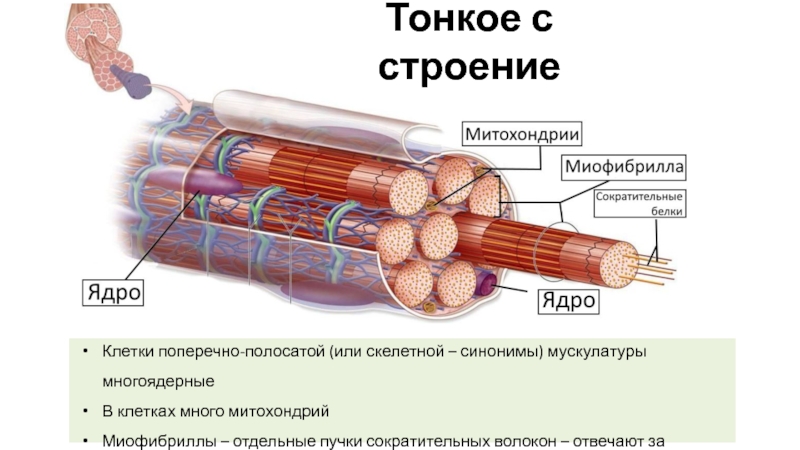 Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается из миоглобина и участвует в биохимических реакциях.
Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается из миоглобина и участвует в биохимических реакциях.
Миосателлитоциты — это малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосателлитоциты одноядерны, их ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов — гликолитических и окислительных.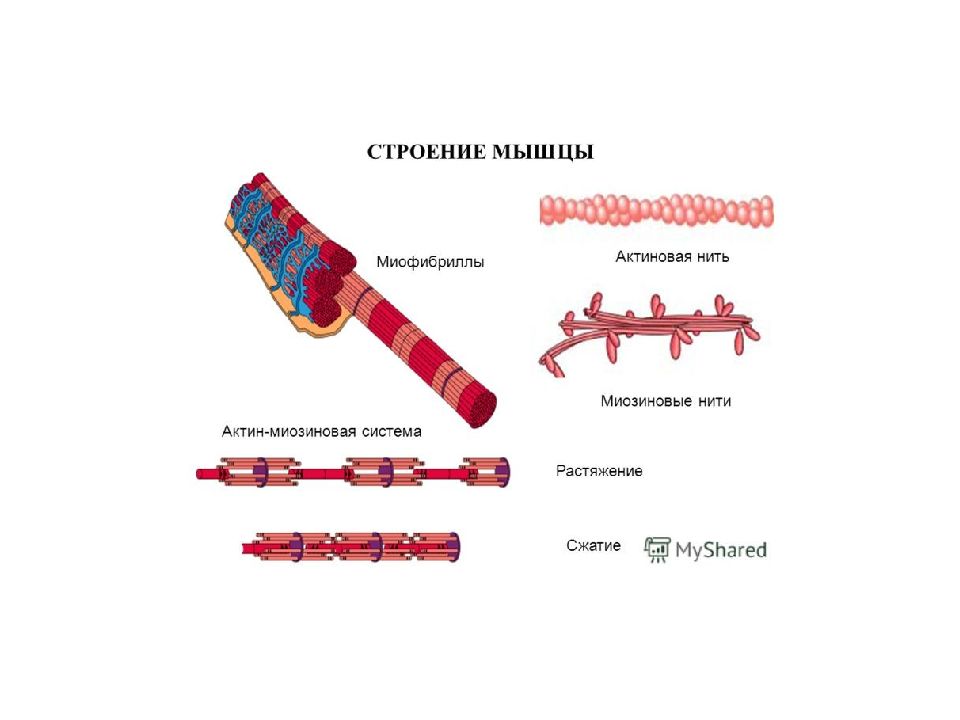
По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные. Наиболее заметно мышечные волокна различаются особенностями молекулярной организации миозина. Среди различных его изоформ существуют две основных — «быстрая» и «медленная». При постановке гистохимических реакций их различают по АТФазной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они более богаты гликогеном, в них меньше миоглобина, поэтому их называют также белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными.
Свойства мышечных волокон меняются при изменении нагрузок — спортивных, профессиональных, а также в экстремальных условиях (таких как невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
Регенерация скелетной мышечной ткани
Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами.
Восстановление любых тканей организма может осуществляется за счет двух механизмов: гипертрофии и гиперплазии.
Под гипертрофией подразумевают компенсаторное увеличение объема самого симпласта, в т.ч. за счет увеличения количества миофибрилл. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмолеммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку.
Под гиперплазией понимают пролиферацию миосателлитоцитов. Сохранившиеся рядом с повреждением миосателлитоциты делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмолеммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку.
Под гиперплазией понимают пролиферацию миосателлитоцитов. Сохранившиеся рядом с повреждением миосателлитоциты делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Скелетная мышца как орган
Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна.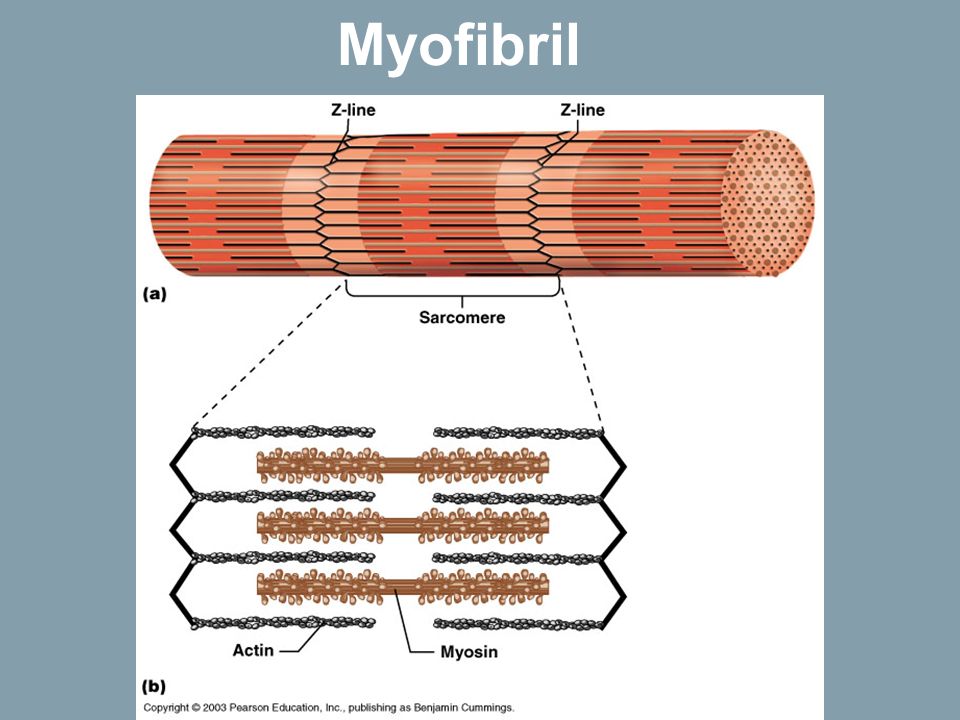 Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани.
Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани — эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Васкуляризация. Артерии вступают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви 5—6-го порядка образуют в перимизии артериолы. В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тканевых базофилов, принимающих участие в регуляции проницаемости сосудистой стенки.
В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тканевых базофилов, принимающих участие в регуляции проницаемости сосудистой стенки.
Иннервация. В мышцах выявлены миелинизированные эфферентные (двигательные), афферентные (чувствительные), а также немиелинизированные вегетативные нервные волокна. Отросток нервной клетки, приносящий импульс от моторного нейрона спинного мозга, ветвится в перимизии. Каждая его ветвь проникает сквозь базальную мембрану, и у поверхности симпласта на плазмолемме образует терминали, участвуя в организации так называемой моторной бляшки, или нервно-мышечного соединения. При поступлении нервного импульса из терминали выделяется ацетилхолин — медиатор, который вызывает возбуждающий потенциал действия, распространяющееся отсюда по плазмолемме миосимпласта.
Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется нервно-мышечной единицей. Характерно, что мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Группа же мышечных волокон, иннервируемых одним мотонейроном, называется нервно-мышечной единицей. Характерно, что мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Чувствительные нервные окончания располагаются не на рабочих мышечных волокнах, а связаны со специализированными мышечными волокнами в так называемых мышечных веретенах, которые расположены в перимизии. Волокна в таких чувствительных мышечных веретенах именуются интрафузальными волокнами, а обычные рабочие мышечные волокна – экстрафузальными.
Интрафузальные мышечные волокна веретен значительно тоньше рабочих. Существует два их вида — волокна с ядерной сумкой и волокна с ядерной цепочкой. Каждое мышечное волокно веретена спирально обвито терминалью чувствительного нервного волокна. В результате сокращения или расслабления рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена, соответственно изменяется тонус интрафузальных мышечных волокон.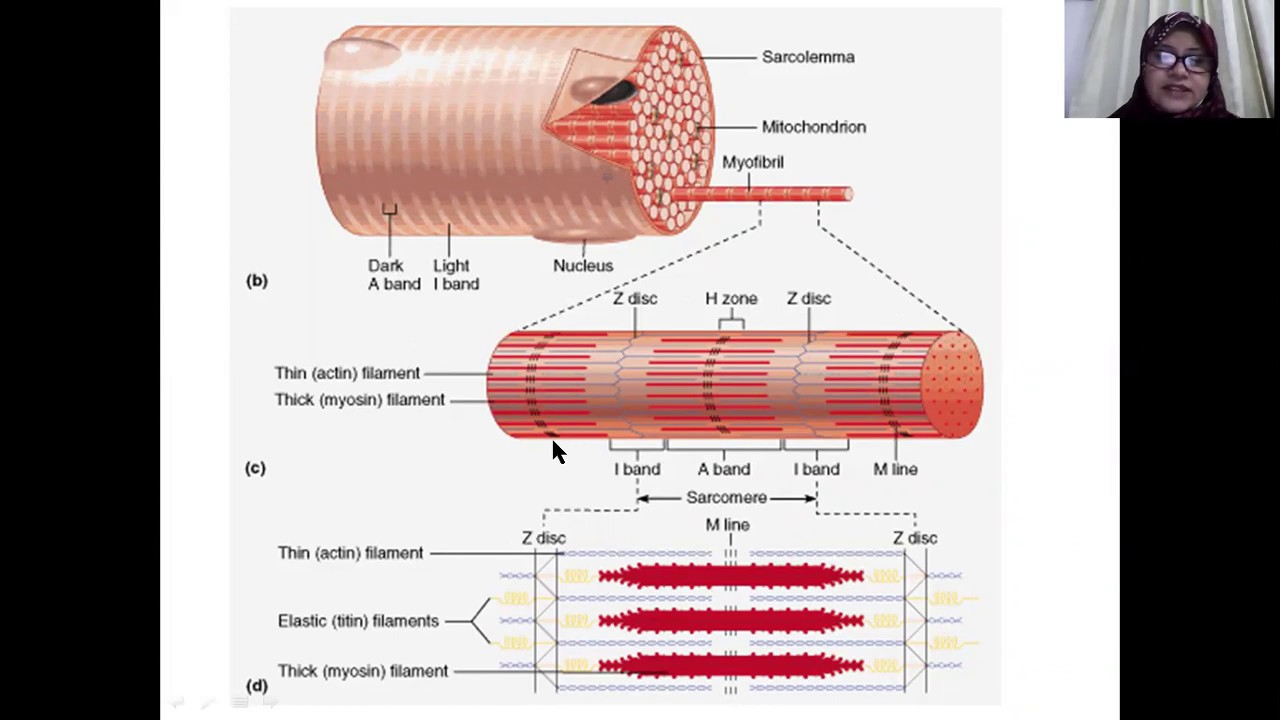 Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом.
Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом.
Сердечная мышечная ткань
Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани — симметричные участки висцерального листка спланхнотома в шейной части зародыша — так называемые миоэпикардиалъные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 3 вида кардиомиоцитов:
- рабочие, или типичные, или же сократительные, кардиомиоциты,
- атипичные кардиомиоциты (сюда входят пейсмекерные, проводящие и переходные кардиомиоциты, а также
- секреторные кардиомиоциты.
Рабочие (сократительные) кардиомиоциты образуют свои цепочки.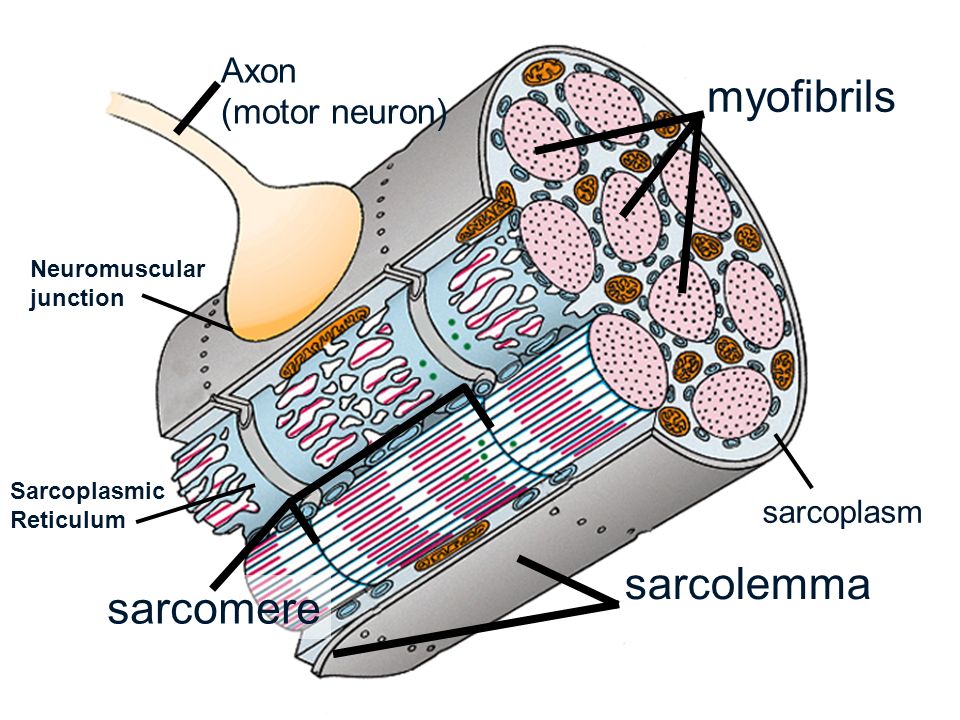 Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы.
Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу.
Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления.
Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности.
Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим.
Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим.
Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы.
Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу.
Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления.
Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности.
Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим.
Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим.
Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают гормон — натрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах.
Сократительные кардиомиоциты имеют удлиненную (100—150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски.
Кардиомиоциты могут ветвиться и образуют трехмерную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Миофибриллы слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены, контактируют с мембранами гладкой эндоплазматической (т.е. саркоплазматической) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта.
Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта.
Кардиомиоциты соединяются друг с другом своими торцевыми концами. Здесь образуются так называемые вставочные диски: эти участки выглядят как тонкие пластинки при увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (или щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани.
Гладкие мышечные ткани
По происхождению различают три группы гладких (или неисчерченных) мышечных тканей — мезенхимные, эпидермальные и нейральные.
Мышечная ткань мезенхимного происхождения
Гистогенез. Стволовые клетки и клетки-предшественники гладкой мышечной ткани, будучи уже детерминированными, мигрируют к местам закладки органов. Дифференцируясь, они синтезируют компоненты матрикса и коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Структурно-функциональной единицей гладкой, или неисчерченной, мышечной ткани является гладко-мышечная клетка, или гладкий миоцит — это веретеновидная клетка длиной 20—500 мкм, шириной 5—8 мкм. Ядро клетки палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены в цитоплазме около полюсов ядра. Аппарат Гольджи и гранулярная эндо плазматическая сеть развиты слабо, что свидетельствует о малой активности синтетических функций. Рибосомы в большинстве своем расположены свободно.
Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно, точнее косо-продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца.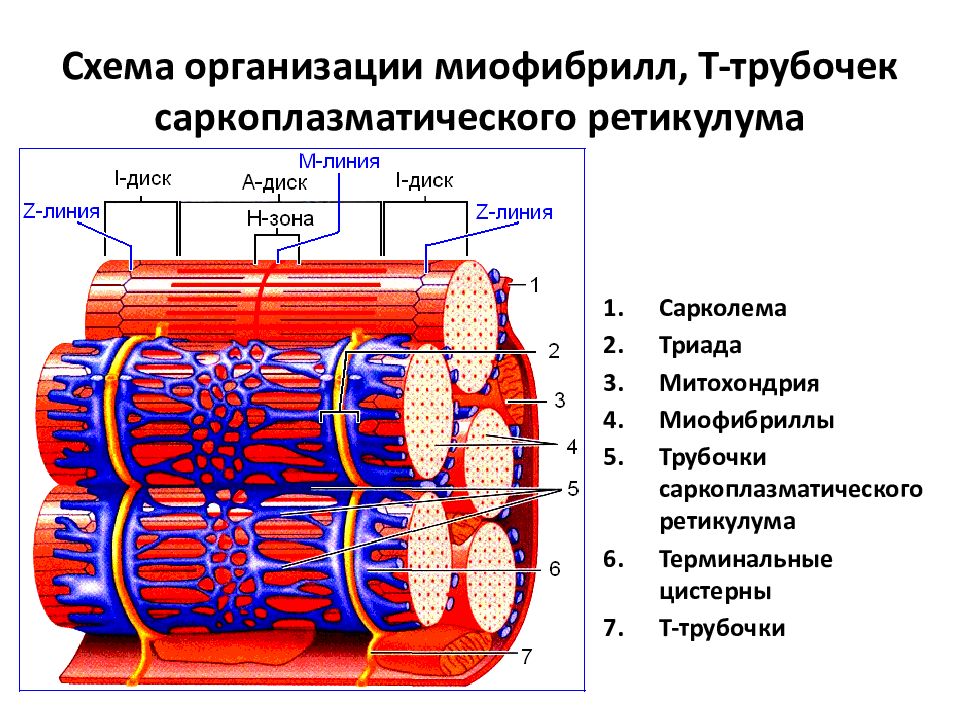
Миозиновые филаменты находятся в деполимеризованном состоянии. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует впячивания — кавеолы, в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков (здесь из пузырьков освобождается кальций). Это влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и «миофибриллы» распадаются. Таким образом, актино-миозиновые комплексы существуют в гладких миоцитах только в период сокращения.
Гладкие миоциты располагаются без заметных межклеточных пространств и разделены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Ретикулярные волокна проникают в щели на концах миоцитов, закрепляются там и передают усилие сокращения клетки всему их объединению.
На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Ретикулярные волокна проникают в щели на концах миоцитов, закрепляются там и передают усилие сокращения клетки всему их объединению.
Регенерация. Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается (рабочая гипертрофия клеток). Не исключена, однако, и пролиферация клеток (т.е. гиперплазия).
В составе органов миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани.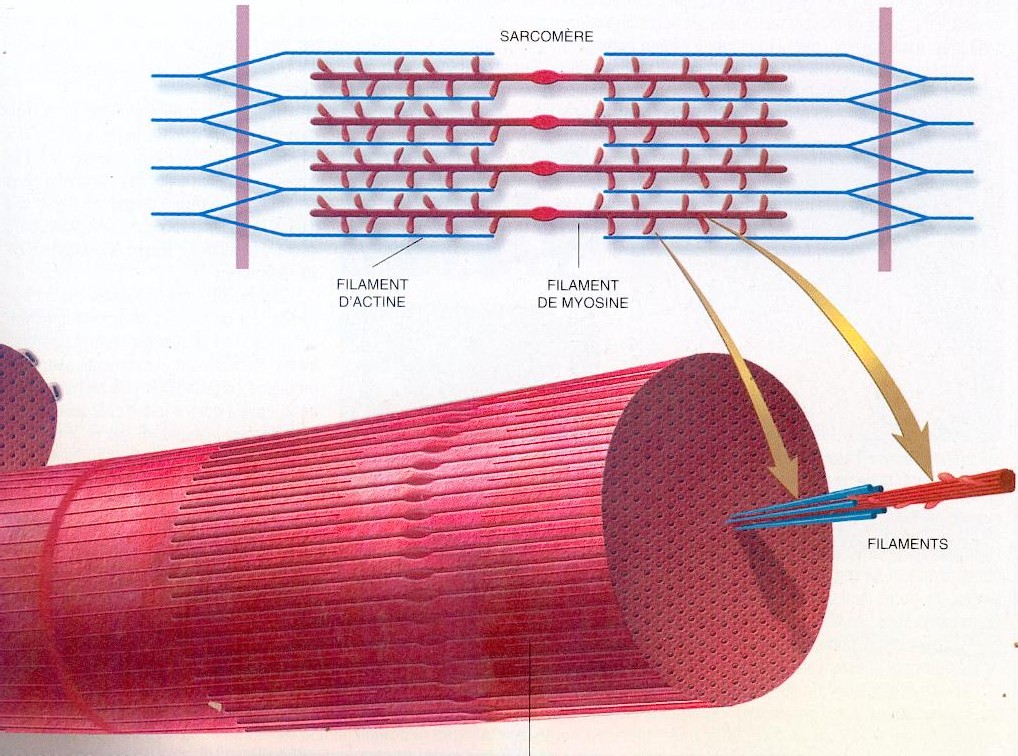 В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы.
В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова.
Гладкая мышечная ткань эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с железистыми секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках — сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках — сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Гладкая мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах.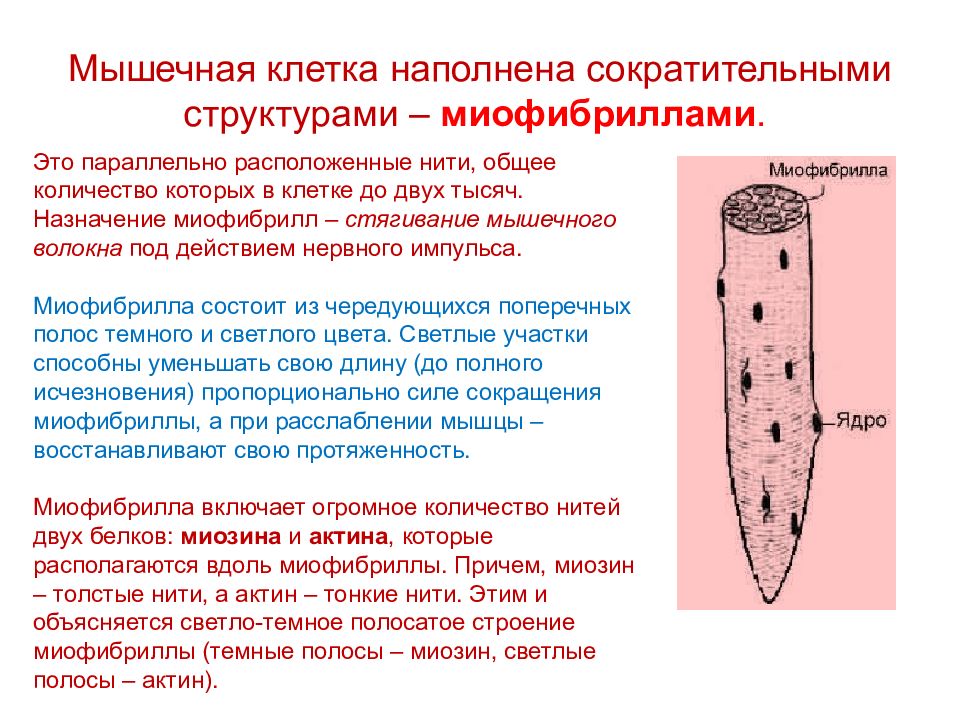 В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
Некоторые термины из практической медицины:
- лейомиома — доброкачественная опухоль, развивающаяся из гладкой мышечной ткани;
- миогелез — образование в мышцах болезненных очагов уплотнения, обусловленное переходом коллоидов миофибрилл в фазу геля, их гомогенизацией и восковидным некрозом; наблюдается, напр., при охлаждении тела, травмах;
- миоциты Аничкова — клетки с характерным расположением ядерного хроматина в виде зубчатой полоски, проявляющие фагоцитарную активность; встречаются в миокарде, напр. при миокардитах;
Скайраннинг — советы чайникам | АЛЬПИНДУСТРИЯ
17 ноября 2011 | Иван Григорьев
Советы и инструкции
Начало
Часть II
Продолжаю серию статей, посвящённых физиологии скайраннинга. Сегодня мы рассмотрим классификацию мышечных волокон и их изменение под воздействием различных видов нагрузки. К сожалению, многие люди не знакомы с этой информацией. Это очень грустно, потому что альпинисты должны быть сведущи в вопросах физиологии и подготовки.
Сегодня мы рассмотрим классификацию мышечных волокон и их изменение под воздействием различных видов нагрузки. К сожалению, многие люди не знакомы с этой информацией. Это очень грустно, потому что альпинисты должны быть сведущи в вопросах физиологии и подготовки.
Что такое альпинизм – это в первую очередь выносливость, терпилово, акклиматизация, технические прибамбасы. Всё это невозможно без грамотной подготовки — скалолазной, велосипедной, беговой, лыжной, кому как приятнее, но бег — это всё-таки основа. С помощью бега вы всегда сможете подготовить своё сердце и мышцы к любому восхождению. Вы можете лазать 8а, это даст вам преимущество на скальном маршруте, но в горы всё-таки ходят. Именно поэтому я так надеюсь, что скайраннинг станет локомотивом профессиональной горной подготовки, так же, как в своё время стали скалолазание и ледолазание в подготовке к прохождению более сложных участков маршрутов.
Информация, публикуемая ниже, написана замечательным физиологом, нашим соотечественником Виктором Николаевичем Селуяновым. Его статьи доступны в интернете, как и большинство других материалов на тему физиологии и тренировок, но к сожалению, человек так устроен, что часто не в состоянии найти нужную информацию: некоторые материалы кажутся слишком сложными, некоторые слишком объёмными или простыми. Так произошло с моим тренером, который 12 лет бегал марафоны, «убивал» себя и не имел ни малейшего представления о физиологии. Он знал что такое интервальная работа, пано, темповик, углеводный обмен, длительная, в каких объёмах и когда нужно выполнять, но всё это происходило на уровне подсознания. Несколько лет назад он нашёл Селуяновские статьи, точнее их порекомендовала Ирина Реутович, которая тренировалась – нет, не по ним, Ирина Владимировна тренировалась всегда по интуиции, у неё очень сильный организм. А статьи она дала для того, чтобы Михаил Викторович (мой тренер) «разобрался» в них. Вот так, всей компанией мы стали зубрить термины и купили пульсометры. Результат оказался потрясающим.
Вот так, всей компанией мы стали зубрить термины и купили пульсометры. Результат оказался потрясающим.
Если раньше наши тренировки носили угнетающий характер, мы молотили, что есть мочи, считали, что этого не достаточно, и молотили ещё. То теперь, помимо того, что мы не чувствовали такой усталости, за счёт того, что грамотно распределяли нагрузку, наши результаты полезли вверх. Через несколько месяцев после начала тренировок по Селуянову мы уже могли бежать по 4 минуты на км на пульсе всего-лишь 140 ударов. Правда, потом мы стали готовиться к суточному бегу и в результате измотали себя, потому что не следили за мышцами, но уже тогда мы поняли что к чему. Именно поэтому я публикую то, что проверено мной и моими друзьями, на себе, за годы тренировок.
Помимо Селуяновских статей я хотел порекомендовать ещё две книги: первая Питт Фитзингер и Скотт Дуглас «Бег по шоссе для серьёзных бегунов» и вторая Дэнни Дрэйер «Ци бег». Последнюю кстати я прочитал по рекомендации известного российского марафонца Леонида Швецова. В его словах звучали сожаления, что эта книга не попалась ему тогда, когда он только начинал свою беговую карьеру. Если у кого-то возникнут проблемы с поиском этих материалов, я могу выслать их по электронной почте, пишите.
В его словах звучали сожаления, что эта книга не попалась ему тогда, когда он только начинал свою беговую карьеру. Если у кого-то возникнут проблемы с поиском этих материалов, я могу выслать их по электронной почте, пишите.
А теперь, собственно, по делу.
Словарь чайника
МВ – мышечное волокно (волокна)
ММВ – медленные мышечные волокна
БМВ – быстрые мышечные волокна
ОМВ – окислительные мышечные волокна
ГМВ – гликолитические мышечные волокна
АэП – аэробный порог
АнП – анаэробный порог
МПК – максимальное потребление кислорода
КФ — креатинфосфат
АТФ – аденозинтрифосфорная кислота (основная «энергетическая валюта» клетки)
Миофибриллы — сократимые элементы мышечной клетки (цилиндрические нити толщиной 1 — 2 мкм, идущие вдоль от одного конца мышечного волокна до другого), сокращаются в присутствии АТФ.
Митохондрии – клеточные органеллы (элементы), в которых синтезируется АТФ за счет окислительного фосфорилирования.
Окислительное фосфорилирование – функция клеточного дыхания, при которой происходит синтез АТФ (идет в митохондриях).
Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) представляет собой серию химических реакций, протекающих в митохондриях, и является общим конечным путем окисления углеводов, липидов и белков.
Миокард – сердечная мышца
Миокардиоцит – клетка миокарда
Классификация мышечных волокон. Изменение мышечной композиции под действием тренировки
Остановимся подробнее на классификации мышечных волокон. Первый способ — на быстрые мышечные волокна (БМВ) и медленные мышечные волокна (ММВ), эта классификация идет по ферменту АТФаза миофибрилл (сократительных элементов), тип которого может быть быстрым или медленным. Отсюда быстро сокращающиеся и медленно сокращающиеся МВ.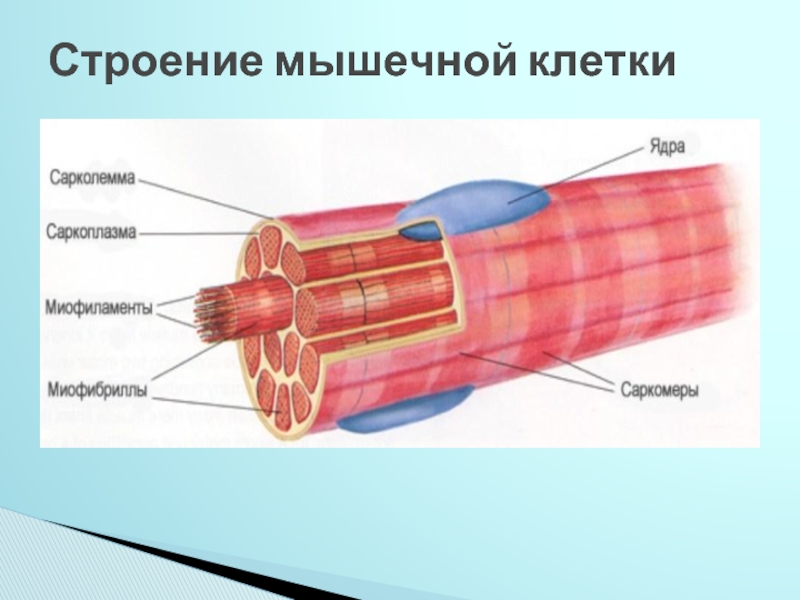 Соотношение быстрых и медленных волокон определяется наследственной информацией, и изменить его мы практически не можем.
Соотношение быстрых и медленных волокон определяется наследственной информацией, и изменить его мы практически не можем.
Второй способ – разделение МВ на окислительные и гликолитические, а они делятся уже не по миофибрилле, а по количеству митохондрий (структур клетки, где происходит потребление кислорода). Если есть митохондрии, то МВ окислительные, мало митохондрий или почти нет — гликолитические. Способность МВ к гликолизу также наследуется и определяется количеством ферментов гликолитического типа. Но вот количество митохондрий достаточно легко изменяется под воздействием тренировок. И с увеличением числа митохондрий МВ, бывшее гликолитическим, становится окислительным.
К сожалению, в этом вопросе существует путаница. Обычно смешивают обе классификации. Говорят о медленных, а подразумевают окислительные, смешивают гликолитические и быстрые. На самом деле медленные тоже могут быть гликолитическими, хотя этот вариант в литературе не описывается. Но мы знаем, что если человек лежит в больнице предоперационный период, а потом ещё и послеоперационный период, то потом уже и встать не может, ходить не может.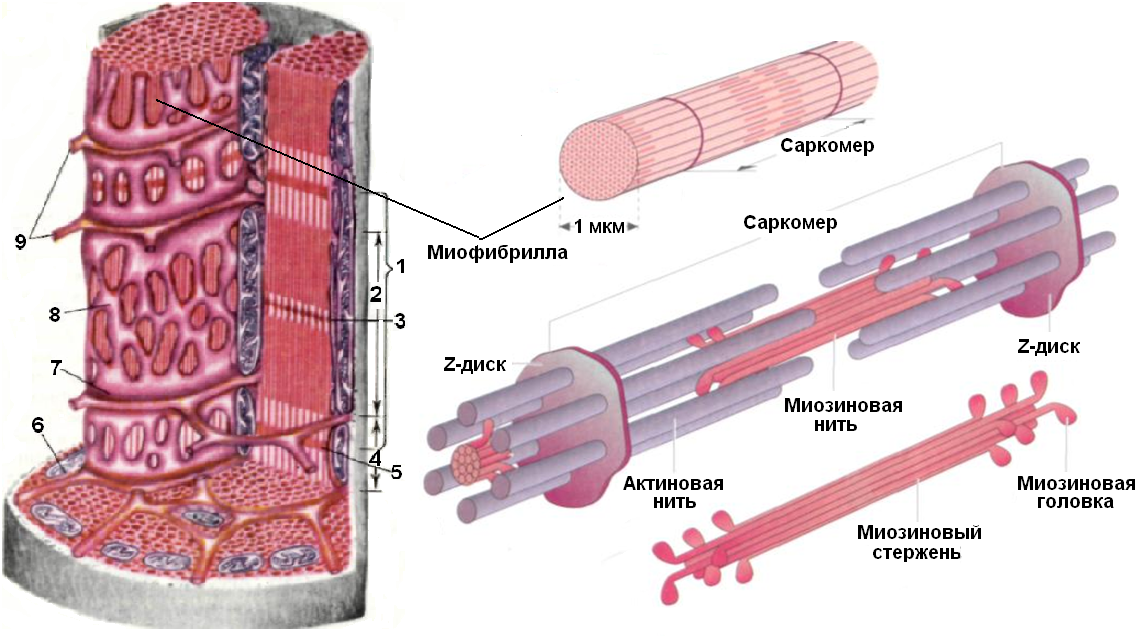 Первая причина — координация нарушается, а вторая причина — мышцы уходят. И самое главное, уходят, прежде всего, митохондрии из ММВ (период их «полураспада» всего 20 — 24 дня). Если человек пролежал 50 дней, то от митохондрий почти ничего не останется, МВ превратятся в медленные гликолитические, поскольку медленные или быстрые наследуется, а митохондрии создаются. (Быстрые МВ при правильных тренировках также могут стать окислительными).
Первая причина — координация нарушается, а вторая причина — мышцы уходят. И самое главное, уходят, прежде всего, митохондрии из ММВ (период их «полураспада» всего 20 — 24 дня). Если человек пролежал 50 дней, то от митохондрий почти ничего не останется, МВ превратятся в медленные гликолитические, поскольку медленные или быстрые наследуется, а митохондрии создаются. (Быстрые МВ при правильных тренировках также могут стать окислительными).
Поэтому с точки зрения тренировочного процесса для данного спортсмена не интересно деление МВ на медленные и быстрые – это имеет значение на этапе отбора. Вся логика построения тренировки идет не с точки зрения сокращения мышц по скорости, а направлена на превращение ГМВ в окислительные. Ибо в этом случае мы изменяем конкретного человека.
Цель тренировки в циклических видах спорта — создавать митохондрии. Только митохондрии потребляют кислород, значит, спортивная форма растет по мере накопления митохондрий. Возьмем мышечное волокно. У него есть миофибриллы, каждая миофибрилла оплетается митохондриями, и больше определенного предела они не могут образоваться, только в один слой, если условно так говорить. В конце концов, эти МВ накапливают столько митохондрий, что больше ничего прибавить не могут. ММВ быстро выходят на предел подготовленности, и дальше весь процесс роста спортивной формы идет через то, что мы гликолитические превращаем в окислительные. (Низкопороговые МВ потому и окислительные, что постоянно работают при любых интенсивностях с максимальной для них мощностью). Перестаём тренироваться или, например, начинаем низкопороговые тренировать, тогда высокопороговые митохондрии теряют. Весь смысл набора спортивной формы — набрать митохондрии в МВ высокопороговых двигательных единиц, другого пути нет. Все только этим и занимаются, а думают об интервальной тренировке и еще о чем-то, то есть о формальности. А суть тренировки — поменять содержание мышечных волокон, то есть добавить митохондрий.
В конце концов, эти МВ накапливают столько митохондрий, что больше ничего прибавить не могут. ММВ быстро выходят на предел подготовленности, и дальше весь процесс роста спортивной формы идет через то, что мы гликолитические превращаем в окислительные. (Низкопороговые МВ потому и окислительные, что постоянно работают при любых интенсивностях с максимальной для них мощностью). Перестаём тренироваться или, например, начинаем низкопороговые тренировать, тогда высокопороговые митохондрии теряют. Весь смысл набора спортивной формы — набрать митохондрии в МВ высокопороговых двигательных единиц, другого пути нет. Все только этим и занимаются, а думают об интервальной тренировке и еще о чем-то, то есть о формальности. А суть тренировки — поменять содержание мышечных волокон, то есть добавить митохондрий.
Вот вы начинаете правильно тренироваться и набираете митохондрий всё больше, больше и больше, мышцы переходят из формы гликолитической в окислительную, то есть с обилием митохондрий. И когда все мышечные волокна становятся окислительными — это предел спортивной формы, больше ничего не получится. Хотя тут есть одна хитрость. Дело в том, что окислительные волокна потребляют только жиры (пока есть запас жиров), а мощность при окислении жиров теряется. Отсюда получается некий парадокс — не надо делать так, чтобы мышцы были только окислительные, надо оставить немного гликолитических, иначе будете на жирах бежать, а мощность функционирования на жирах меньше примерно на 15%. Тогда те же самые мышцы будут более мощно работать. Понятно, что к лыжному спорту это тоже относится.
Хотя тут есть одна хитрость. Дело в том, что окислительные волокна потребляют только жиры (пока есть запас жиров), а мощность при окислении жиров теряется. Отсюда получается некий парадокс — не надо делать так, чтобы мышцы были только окислительные, надо оставить немного гликолитических, иначе будете на жирах бежать, а мощность функционирования на жирах меньше примерно на 15%. Тогда те же самые мышцы будут более мощно работать. Понятно, что к лыжному спорту это тоже относится.
Влияние гликолитических и окислительных мышечных волокон на результат
Так вот, вы начинаете бежать среднюю дистанцию, разбегаетесь, и выходите на порог анаэробного обмена, он как раз соответствует моменту, когда функционирует все ОМВ и даже часть гликолитических. При этом получается, что человек выходит на крейсерскую скорость. Если у него только ОМВ, то он так и будет стабильно молотить. Прибавить не может и убавить не может (убавить, конечно, может, но это ему не надо, а прибавить не может, потому что не чем добавить), он прибежит с той же самой скоростью на финиш.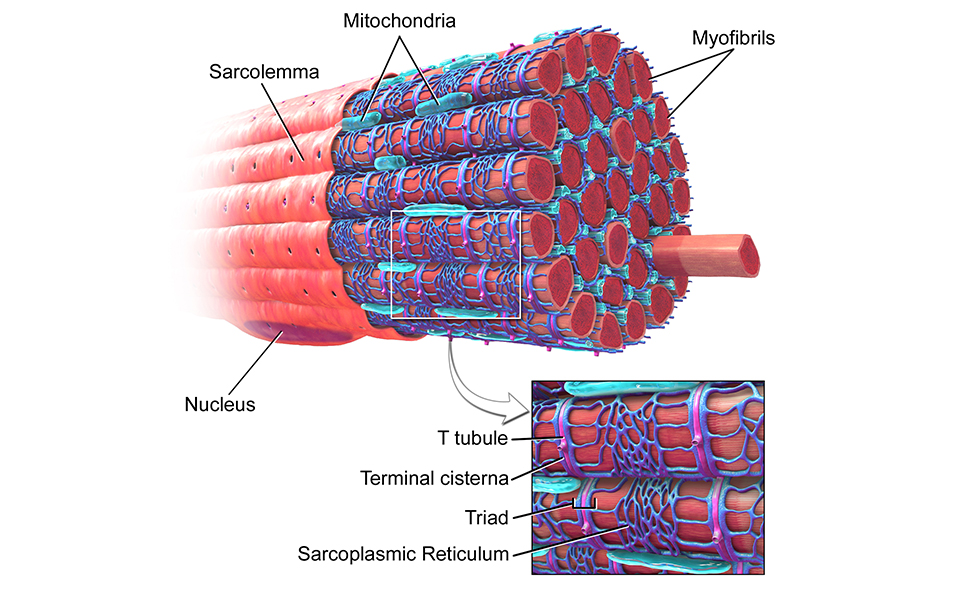 Если с ним будет бежать точно такой же человек, но у которого будет запас ГМВ, то он на финише всегда прибавит. Значит, получается, средневик — это человек, у которого есть запас мышечных волокон, которые он может включить в работу, и лучше быстрых гликолитических, тогда финиш будет еще быстрее. Так же и у лыжников: тот, у кого есть запас ГМВ, на финише выиграет, если дистанция будет ровная. Но, увы, так не бывает.
Если с ним будет бежать точно такой же человек, но у которого будет запас ГМВ, то он на финише всегда прибавит. Значит, получается, средневик — это человек, у которого есть запас мышечных волокон, которые он может включить в работу, и лучше быстрых гликолитических, тогда финиш будет еще быстрее. Так же и у лыжников: тот, у кого есть запас ГМВ, на финише выиграет, если дистанция будет ровная. Но, увы, так не бывает.
Снова перейдем на более простой вид спорта, велосипедный (мне ближе). Рассмотрим спортсмена, у которого ОМВ только 15-20%, остальные — гликолитические. На равнине он набирает критическую скорость, превышает её, и начинает постепенно закисляться. Проходит 5-6 минут, он попадает в мертвую точку, пульс запредельный, дышать невозможно. Спортсмен начинает мощность снижать, и через 2-3 км выходит, наконец, на ту самую скорость, которая нужна. Вот классический вариант развития физиологических процессов на равнине. А если это не равнина, а холмистая местность, и холмы короткие, по длине такие, что на подъем затрачивается не больше 30 секунд? Тогда в этот холм спортсмен включает свои ГМВ, их хватает ровно на 30 секунд.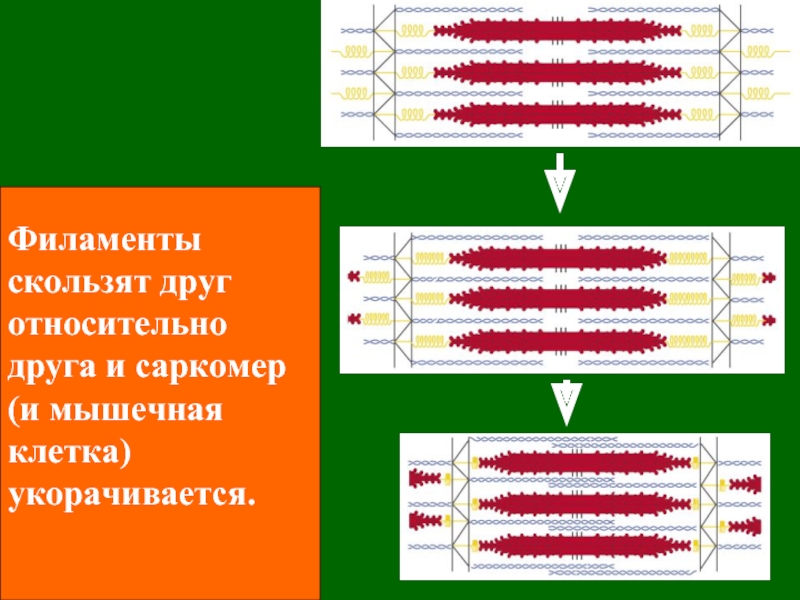 В холм влетает, скорость большая, а со спуска работать уже практически не надо, ГМВ восстанавливаются, потом опять подъём, спуск и т.д. При этом он может влететь в этот подъём быстро и мощно, а другой, у кого только одни окислительные, такой мощности не получит, попытается отыграть на спуске, но это очень трудно и особенно добавить не удастся. В этих условиях спортсмен, у которого много ГМВ, начинает выигрывать.
В холм влетает, скорость большая, а со спуска работать уже практически не надо, ГМВ восстанавливаются, потом опять подъём, спуск и т.д. При этом он может влететь в этот подъём быстро и мощно, а другой, у кого только одни окислительные, такой мощности не получит, попытается отыграть на спуске, но это очень трудно и особенно добавить не удастся. В этих условиях спортсмен, у которого много ГМВ, начинает выигрывать.
Рассмотрим двух спортсменов в равных условиях, но у первого мышцы покрупнее (больше ГМВ), а у второго поменьше. Если это равнина, первый, скорее всего, выиграет, потому что включит на финише гликолитические волокна. По дистанции они будут одинаково ехать, а на финише первый выиграет с разницей в 1-2 секунды. Если холмистая местность, но с короткими холмами, выиграет первый, у которого больше гликолитических МВ, может еще больше выиграет, потому что он в каждую горку 1-2 секунды отыграет, а со спуска еще быстрее уедет. Но как только горка превращается в минутную, то на первой он 2 секунды отыграет за 30 секунд, второй немножко отстал, а потом на следующей горке второй ему 10 секунд ввезет, потому что у первого ГМВ перестанут нормально работать, закислятся, а у второго ничего не закисляется, он со стабильной скоростью до верха и доедет.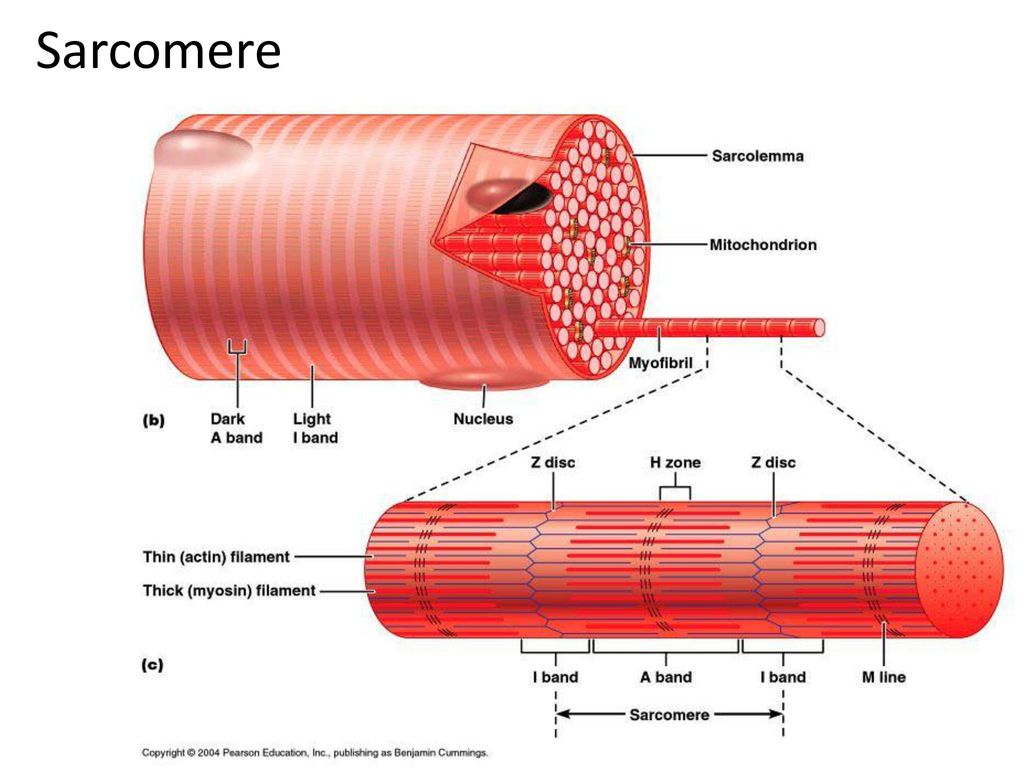
Вот тут эти нюансы и возникают.
Теперь переключимся на лыжи. Если спринт будет с короткими подъёмами или же длинная дистанция с короткими подъёмами, выиграет тот, у кого есть запас ГМВ и очень большой. Но в лыжном спорте коротких подъемов почти не бывает. А как только подъёмы по длительности уходят за 30 секунд, всё меняется, к 40-й секунде ноги начинают здорово болеть, а к 1 минуте дыхание резко учащается, потому что ГМВ начинают накапливать ионы водорода, молочную кислоту, начинается значительное выделение углекислого газа, он заставляет интенсивно дышать, пульс за 200 и страшные мучения. Если всё время выходить на пульс 200-240, повторять его по ходу гонки 10 – 15 — 20 раз, то и соперника не увидишь… (состояние будет предельно тяжелым).
Физиология мышечного сокращения. Закон рекрутирования мышечных волокон
Напомним современные знания физиологии мышечного сокращения. Начнем с учебных знаний. В учебнике пишется, что существует процесс сокращения мышцы, и он обеспечивается некими механизмами энергообеспечения. Сам механизм сокращения связан с затратой молекул АТФ, молекулы АТФ должны быть внутри синтезированы с помощью молекулы КФ, а свободный креатин и свободный фосфат являются стимулом для разворачивания либо анаэробного гликолиза, либо аэробного гликолиза, либо окисления жиров. Вот классическая схема, современная, которая сейчас принята. Эта уточненная схема предложена Саксом, нашим ученым (у Чазова работает), для миокарда. В схеме существует КФ шунт, или, другими словами, все метаболические и энергетические пути, гликолиз и окисление жиров идут только через ресинтез КФ, а уже КФ идет непосредственно на ресинтез АТФ. Вот современные учебные знания.
Сам механизм сокращения связан с затратой молекул АТФ, молекулы АТФ должны быть внутри синтезированы с помощью молекулы КФ, а свободный креатин и свободный фосфат являются стимулом для разворачивания либо анаэробного гликолиза, либо аэробного гликолиза, либо окисления жиров. Вот классическая схема, современная, которая сейчас принята. Эта уточненная схема предложена Саксом, нашим ученым (у Чазова работает), для миокарда. В схеме существует КФ шунт, или, другими словами, все метаболические и энергетические пути, гликолиз и окисление жиров идут только через ресинтез КФ, а уже КФ идет непосредственно на ресинтез АТФ. Вот современные учебные знания.
В соответствии с ними, если спортсмен начинает двигаться в режиме «во всю», в течение примерно 15 секунд тратятся запасы АТФ и КФ (фосфагенов). Потом должен развернуться процесс, который стимулируется свободным креатином. Это, в первую очередь, процесс анаэробного гликолиза, который продолжается одну, может быть, полторы минуты, а вслед за этим должен развернуться процесс окислительного фосфорилирования, то есть начинается уже аэробный гликолиз.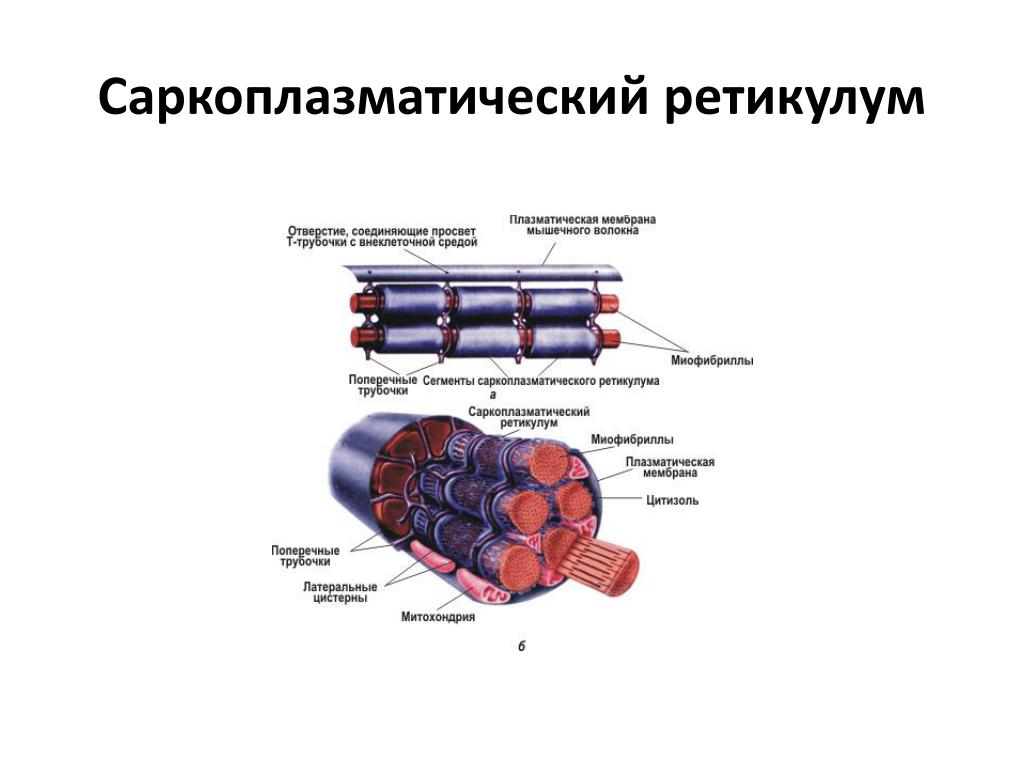 У нормального человека запасы углеводов снижаются где-то после 20-30 мин, а полностью заканчиваются через 45 мин. И только тогда, когда заканчиваются запасы углеводов в мышце и глюкоза в крови, начинает интенсивно развиваться процесс, связанный с окислением жиров. В случае передвижения со средней интенсивностью, при недостатке кислорода в крови, разворачивается анаэробный гликолиз. Это классическая схема.
У нормального человека запасы углеводов снижаются где-то после 20-30 мин, а полностью заканчиваются через 45 мин. И только тогда, когда заканчиваются запасы углеводов в мышце и глюкоза в крови, начинает интенсивно развиваться процесс, связанный с окислением жиров. В случае передвижения со средней интенсивностью, при недостатке кислорода в крови, разворачивается анаэробный гликолиз. Это классическая схема.
Но эта схема не выдерживает критики, когда мы переходим с представлений уровня пробирки или одного единственного мышечного волокна к мышце в целом. Для единственного изолированного МВ это более или менее правильное описание. Но у нас не одно МВ, а множество, еще есть множество мышц и, следовательно, в нашу модель мы должны включить и эти элементы. Кроме того, у нас есть ОМВ и ГМВ, у нас есть те МВ, которые раньше рекрутируются при определенной интенсивности: если интенсивность меняется, то дополнительные мышечные волокна включаются. Короче говоря, есть закон рекрутирования МВ. Если все эти компоненты учесть, то мы построим новую модель, которая состоит из центральной нервной системы, которая управляет мотонейронами в спинном мозге, а мотонейроны управляют мышцами. И вот в зависимости от импульсации, которая идет сверху, рекрутируются сначала низкопороговые двигательные единицы, а потом всё более высокопороговые, когда, допустим, увеличивается сила отталкивания. И в этом случае получается совсем другая картина.
Если все эти компоненты учесть, то мы построим новую модель, которая состоит из центральной нервной системы, которая управляет мотонейронами в спинном мозге, а мотонейроны управляют мышцами. И вот в зависимости от импульсации, которая идет сверху, рекрутируются сначала низкопороговые двигательные единицы, а потом всё более высокопороговые, когда, допустим, увеличивается сила отталкивания. И в этом случае получается совсем другая картина.
Например, вы начинаете двигаться с усилием 50% от максимума, максимум — это спринт (3-7 секунд), а 50% — это, условно говоря, бег на 1500 м или на 3000 м. Что будет происходить в организме? Вы рекрутируете столько мышечных волокон, сколько необходимо, чтобы держать скорость. Допустим, у вас 75% ОМВ. Допустим, вы рекрутировали половину всех мышечных волокон. Рекрутированные ОМВ отрабатывают 15 сек за счет АТФ и КФ, затем мощность их начинает падать где-то наполовину, и дальше эти ОМВ работают только в аэробном режиме, и пока используют только жиры.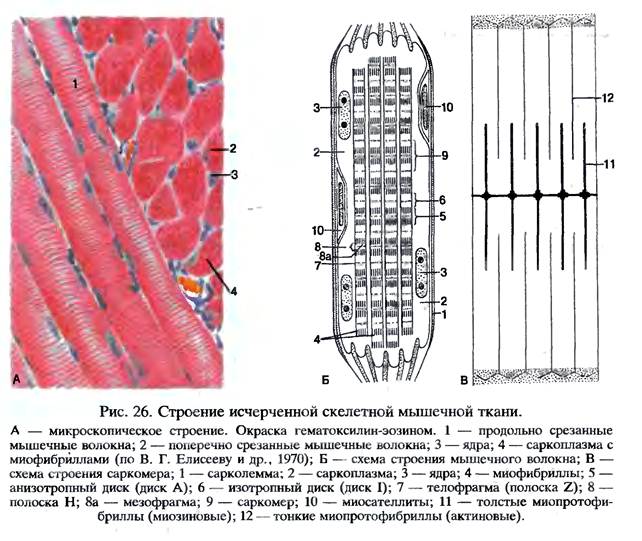 Не через 40 минут, а прямо сейчас, на 1-й минуте будут работать за счет окисления жиров! Потому что в ОМВ митохондрии, когда работают, выделяют наружу цитрат, который ингибирует (подавляет) гликолиз, поэтому могут окисляться только жиры (химию процесса окисления описывает цикл Кребса). Значит, не прошло и 15 секунд, как начали окисляться жиры. И вот мощность упала, а вам-то задание держать 50% от максимальной. Тогда вы обязаны рекрутировать еще порцию мышечных волокон. Допустим, дополнительные 25% вы рекрутируете, тоже окислительные, только они еще не работали, и они свои первые 15 секунд отрабатывают на АТФ и КФ. Получается, что на АТФ и КФ вы бежите уже не 15 секунд, а 30. То есть вы 15 секунд бежали на АТФ рекрутированных вначале МВ, и еще 15 секунд на следующих, но часть работы уже выполняется за счет аэробной продукции. Эти окислительные включились в работу, истратили свои запасы АТФ и КФ, не полностью, а наполовину, а вот эта половина поддерживается за счёт ресинтеза, то есть уже за счёт окислительных процессов, за счет жиров.
Не через 40 минут, а прямо сейчас, на 1-й минуте будут работать за счет окисления жиров! Потому что в ОМВ митохондрии, когда работают, выделяют наружу цитрат, который ингибирует (подавляет) гликолиз, поэтому могут окисляться только жиры (химию процесса окисления описывает цикл Кребса). Значит, не прошло и 15 секунд, как начали окисляться жиры. И вот мощность упала, а вам-то задание держать 50% от максимальной. Тогда вы обязаны рекрутировать еще порцию мышечных волокон. Допустим, дополнительные 25% вы рекрутируете, тоже окислительные, только они еще не работали, и они свои первые 15 секунд отрабатывают на АТФ и КФ. Получается, что на АТФ и КФ вы бежите уже не 15 секунд, а 30. То есть вы 15 секунд бежали на АТФ рекрутированных вначале МВ, и еще 15 секунд на следующих, но часть работы уже выполняется за счет аэробной продукции. Эти окислительные включились в работу, истратили свои запасы АТФ и КФ, не полностью, а наполовину, а вот эта половина поддерживается за счёт ресинтеза, то есть уже за счёт окислительных процессов, за счет жиров. И при заданной 50-процентной мощности вы обеспечиваете где-то 30-35% за счет окислительного фосфорилирования. При такой мощности где-то через 30-40 секунд вы выходите на предельные возможности этой мышцы в потреблении кислорода (она равна как раз 35% от максимальной мощности, которую эта мышца может развить). Это соответствует как раз АнП. Если нарисовать кривую потребления кислорода, то вы обнаружите плато, которое будет соответствовать АнП уже через 40 сек.
И при заданной 50-процентной мощности вы обеспечиваете где-то 30-35% за счет окислительного фосфорилирования. При такой мощности где-то через 30-40 секунд вы выходите на предельные возможности этой мышцы в потреблении кислорода (она равна как раз 35% от максимальной мощности, которую эта мышца может развить). Это соответствует как раз АнП. Если нарисовать кривую потребления кислорода, то вы обнаружите плато, которое будет соответствовать АнП уже через 40 сек.
Далее спортсмен будет рекрутировать ГМВ, но маленькими порциями, исходя из нормы мощности, которую вы задали. Вот он в течение минуты будет рекрутировать гликолитические. Они тоже сначала на АТФ и КФ работают, а потом за счет гликолиза. Включенные ГМВ минуту отработают, закислятся и снизят мощность почти до нуля. Поэтому придется включать новые ГМВ до тех пор, пока у вас есть их запас. Если он у вас большой, то можно минуты 3-4 так поработать. А тот, у кого запаса ГМВ нет, начнет снижать мощность, и откажется от выполнения задания.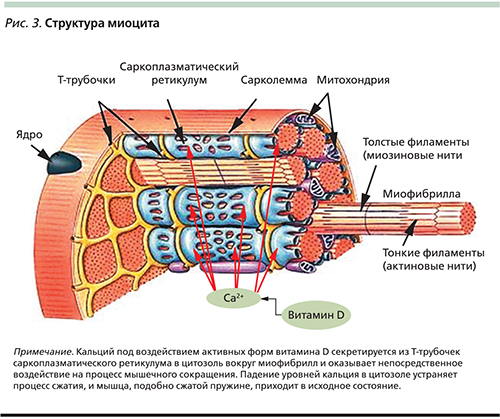
В итоге у тех, у кого ОМВ много, а гликолитических мало, кривая мощности поднимется, продержится где-то минуты полторы – две, и обязательно упадет на уровень АнП, и так будет держаться долго. Тот, кто имеет больший запас ГМВ, при прочих равных условиях сможет дольше проработать на высокой мощности, и на какой-то определенной дистанции выиграет. Получается, что человек, имеющий много ГМВ, но мало окислительных, на относительно коротких дистанциях, допустим, 1 — 1,5 минуты, ещё может выигрывать запасом гликолитических. Но чем длиннее дистанция, тем менее важна вот эта лишняя мышечная масса (ГМВ). И когда время на дистанции уходит, допустим, за 5 минут, то получается, что надо на себе везти лишнюю массу.
Из-за чего появляется специализация
В велосипедном спорте на равнине лишний вес не имеет принципиального значения. А если это гора, то даже в велосипедном спорте начинает играть роль собственный вес, спортсмен начинает тратить энергию на то, чтобы везти в подъем лишнюю мышечную массу.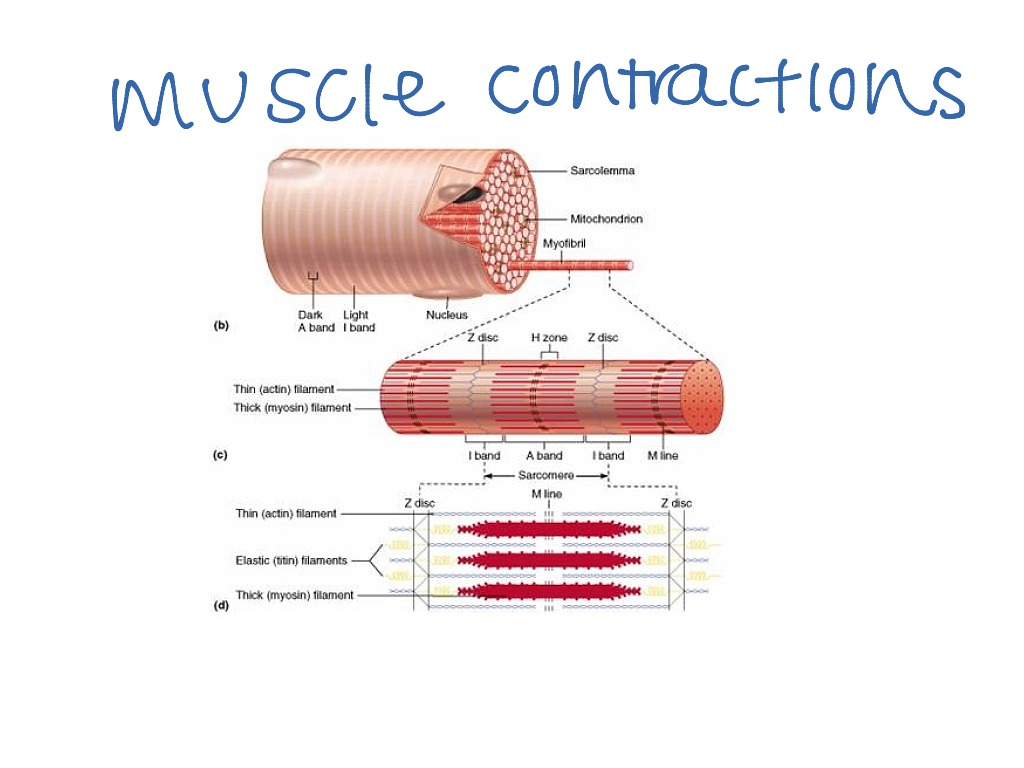 Поэтому чем длиннее дистанция, тем «вреднее» эта лишняя масса, и надо от нее всеми способами избавляться.
Поэтому чем длиннее дистанция, тем «вреднее» эта лишняя масса, и надо от нее всеми способами избавляться.
То же и в конькобежном спорте. Конечно, спортсмен в основном работает против ветра, но еще надо много энергии тратить, чтобы перемещать свое тело поперек дорожки, держать позу, а именно — везти свой собственный вес. Значит, и здесь вес начинает играть свою роль. Поэтому, если конькобежец везет на себе лишнюю мышечную массу, она мешает. На дистанциях в 500 и 1000 м некоторая «лишняя» масса помогает, потому что мощный толчок помогают сделать еще и мышцы рук и туловища. Но чем длиннее дистанция, тем больше «лишняя» масса мешает. Поэтому там, где возникают проблемы «лишней» массы, и появляется какая-то специализация (спринтер — стайер). Но иногда это не принципиально, если у спортсмена сильные мышцы ног с большой долей ОМВ (как у Хайдена).
Как и везде, существует простая модель и сложная. В сложной модели, вы видите, процессы по-другому разворачиваются, и даже можно объяснить, зачем нужны гликолитические волокна.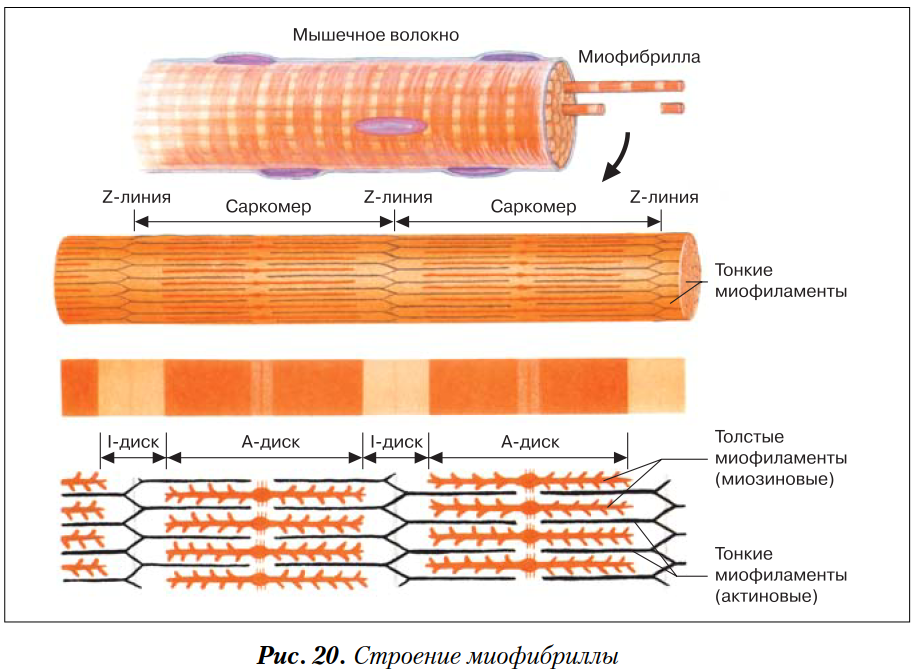 Пока дистанция относительно короткая, и если эта лишняя масса не очень мешает, то это очень выгодно. Чем длиннее дистанция и чем больше нагрузка, связанная с преодолением собственного веса, тем вреднее становится избыток ГМВ.
Пока дистанция относительно короткая, и если эта лишняя масса не очень мешает, то это очень выгодно. Чем длиннее дистанция и чем больше нагрузка, связанная с преодолением собственного веса, тем вреднее становится избыток ГМВ.
Центральные и периферические аэробные компоненты, их вклад в работоспособность
Теперь рассмотрим зависимость работоспособности от центрального и периферического факторов (сердечно-сосудистой системы и мышц). Если рассматривать какое-то конкретное двигательное действие — велосипед, коньки, легкую атлетику (бег) или лыжи, то мы увидим, что в каждом конкретном упражнении участвуют определенные мышечные группы. Если посчитать их массу, то окажется, что в велосипедном спорте одна мышечная масса, в легкой атлетике побольше, а в лыжном спорте еще больше. Возникает вопрос: сколько эти мышцы потребляют кислорода? Чисто теоретически это очень просто посчитать: 1 кг мышечной массы, если она находится на пределе подготовленности, потребляет кислорода где-то 0,2-0,3 л/мин, если в работе участвуют все ОМВ. Дальше надо просто умножить эту цифру на ту массу, которая есть, при условии, что она максимально подготовлена. Что значит максимально подготовлена?
Дальше надо просто умножить эту цифру на ту массу, которая есть, при условии, что она максимально подготовлена. Что значит максимально подготовлена?
Внутри этой мышечной массы одни ОМВ, миофибриллы и митохондрии находятся в таком соотношении, что больше уже ничего прибавить нельзя (миофибриллы все оплетены митохондриями, как в миокарде). И тогда получается, что для потребления 3 л кислорода нужно иметь 10 кг активной мышечной массы, а если нужно потреблять 6 л — достаточно иметь только 20 кг активной мышечной массы.
Теперь посчитаем, сколько кислорода может доставить сердце. Если принять, что 1 литр крови переносит 160 мл кислорода (при нормальном уровне гемоглобина), то, умножив это количество на минутный объём кровообращения, мы получим возможности сердца по доставке кислорода. У обычного человека, мужчины, ударный объём составляет порядка 120-130 мл за один выброс крови. При пульсе 190 ударов в минуту получим 190 уд/мин * 130 мл * 160 мл = около 4 л/мин. Всё так и считается, достаточно просто.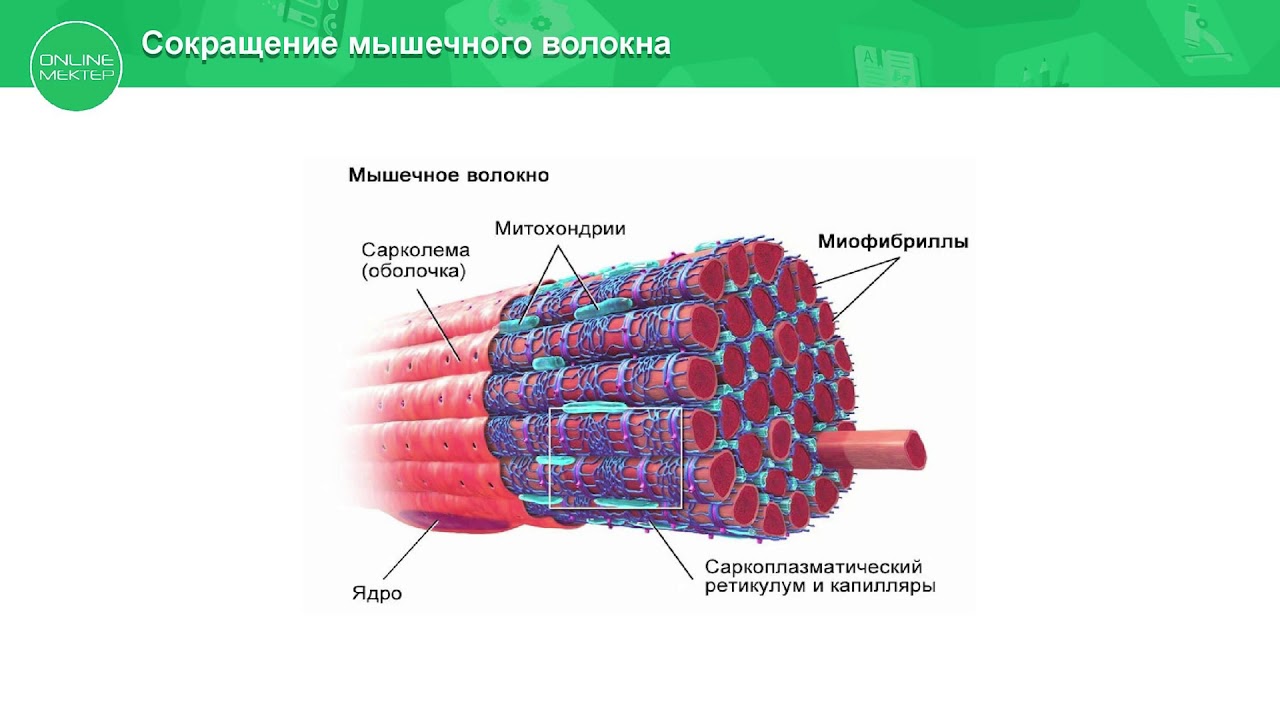 У супер-атлетов за один ударный цикл выбрасывается 240 мл, это соответствует 7-8 л/мин кислорода.
У супер-атлетов за один ударный цикл выбрасывается 240 мл, это соответствует 7-8 л/мин кислорода.
Мы определили, что 20 кг мышечной массы могут потребить около 6 литров кислорода в минуту. Если у лыжника на ногах мышечная масса 20 — 25 кг, и к этому добавить мышцы живота, спины, рук, то мы уйдем за цифру 30 с лишним килограммов. Сделаем поправку на то, что не вся эта мышечная масса будет потреблять кислород на пределе возможностей, и получим, что 40 кг активных мышц могут потребить кислорода около 8 л/мин. Вот столько должно перекачать сердце, чтобы полностью обеспечить мышцы кислородом, если эти мышцы максимально готовы.
Таким образом, мы получили два предела. Первый — из литературы известно, что перекачать 8 л/мин кислорода через организм с помощью сердца — это цифра предельная, этой цифры практически ни у кого нет. В то же время, 8 л/мин кислорода потребить мышцами — таких цифр тоже никто ещё не зафиксировал. Обычно потребляют где-то 6 л/мин, ну — 6,5 л/мин, цифры в 7 л/мин кислорода почти не появляется.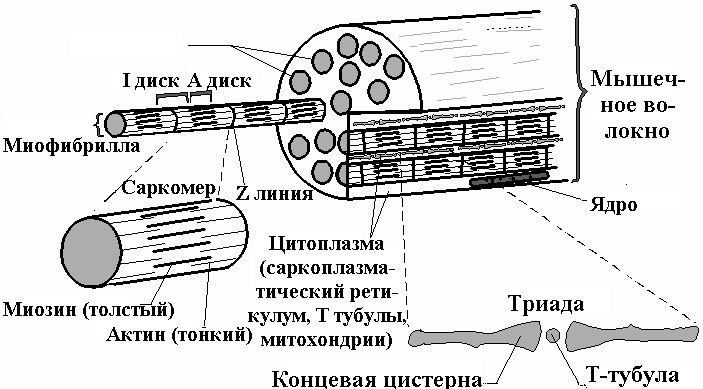
Тестирование уровней потребления кислорода поможет построить планы тренировок
Поскольку работоспособность может лимитировать либо одно, либо другое, то для того, чтобы разобраться с тем, чего не хватает конкретному спортсмену — его надо обязательное тестировать. Например, мы начинаем тестировать лыжников на уровне сборной команды, и получаем очень печальные цифры. Фиксируем показатели неоднократного победителя крупных российских марафонов (спортсмен каждый год в «десятке» на чемпионатах России), и видим: мышцы ног потребляют кислорода только 3,5 л/мин на уровне АнП — это результат порядка 1 разряда в велосипедном спорте. А лыжник должен потреблять ногами столько же, сколько велосипедист МСМК, причем это абсолютная цифра, не на килограмм веса. (В велосипедном спорте это не принципиально, там больше важно, что приходится на лобовую площадь.)
Спрашивается, а какое у него сердце? Если взять график ступенчатого теста, то на начальном участке, когда рекрутируются только ОМВ, наблюдается некая прямая между пульсом и мощностью. Потом эта кривая зависимости (потом кривая получается) начинает как-то изменяться. И, как правило, происходит увеличение темпа прироста пульса. Если продолжить начальный отрезок линии дальше, и вывести на пульс 190, то можно предсказать, что бы было с этим человеком, если бы он вышел на пульс 190, и при этом у него были бы только ОМВ. И тогда мы определили бы потенциальные возможности сердца по доставке кислорода к мышцам. (Подробнее об этом можно прочитать в следующем номере в разделе, посвященном интерпретации данных ступенчатого теста).
Потом эта кривая зависимости (потом кривая получается) начинает как-то изменяться. И, как правило, происходит увеличение темпа прироста пульса. Если продолжить начальный отрезок линии дальше, и вывести на пульс 190, то можно предсказать, что бы было с этим человеком, если бы он вышел на пульс 190, и при этом у него были бы только ОМВ. И тогда мы определили бы потенциальные возможности сердца по доставке кислорода к мышцам. (Подробнее об этом можно прочитать в следующем номере в разделе, посвященном интерпретации данных ступенчатого теста).
Так вот, потенциальная производительность сердца оказывается у него 7 л/мин. Это означает, что наш спортсмен имеет прекрасное сердце, огромное сердце, его тренировать специально не надо, а мышцы, прежде всего ног, — очень слабые, они в очень плохом состоянии, их надо готовить, чтобы они соответствовали нормативам международного класса.
Чтобы этот лыжник показал хорошие результаты, ему надо где-то 4,5 л/мин потреблять ногами. С показателем 4,5 л/мин он бы в сборной уже устойчиво стоял. При этом пульс у него при потреблении кислорода 4,5 л/мин должен быть не 190 уд/мин, а 150, потому что должен быть запас, на котором руки будут работать. Хорошо, предположим, мы с ним в тесте получаем 4,5 л/мин на пульсе 150 уд/мин, и после этого начинается закисление, и он отказывается от работы. Тогда мы говорим, что ноги у него в хорошем состоянии (4,5 л/мин для лыжника вполне достаточно). Потом начинаем тестировать руки, и оказывается, что руки у него потребляют где-то 1,5 л/мин, больше не будут потреблять (это из нашего опыта известно). Он потребляет руками 1,5 л/мин, мы прибавляем их к 4,5 л/мин ног, и получаем потребление кислорода равное 6 л/мин. Затем делим на его вес 70 кг и получаем 85 мл/кг/мин — это уровень олимпийских достижений.
При этом пульс у него при потреблении кислорода 4,5 л/мин должен быть не 190 уд/мин, а 150, потому что должен быть запас, на котором руки будут работать. Хорошо, предположим, мы с ним в тесте получаем 4,5 л/мин на пульсе 150 уд/мин, и после этого начинается закисление, и он отказывается от работы. Тогда мы говорим, что ноги у него в хорошем состоянии (4,5 л/мин для лыжника вполне достаточно). Потом начинаем тестировать руки, и оказывается, что руки у него потребляют где-то 1,5 л/мин, больше не будут потреблять (это из нашего опыта известно). Он потребляет руками 1,5 л/мин, мы прибавляем их к 4,5 л/мин ног, и получаем потребление кислорода равное 6 л/мин. Затем делим на его вес 70 кг и получаем 85 мл/кг/мин — это уровень олимпийских достижений.
Дальше разбираемся, что с ним нужно делать, чтобы достичь таких показателей. Так вот, первый вывод: поскольку сердце у него большое, и может перекачать кислорода 7 л/мин, то этому человеку не надо делать вкатывание. Под вкатыванием понимаются объёмные длительные тренировки продолжительностью от 3 до 6-8 часов в день на относительно низком пульсе (100-150 уд/мин, близко к 120).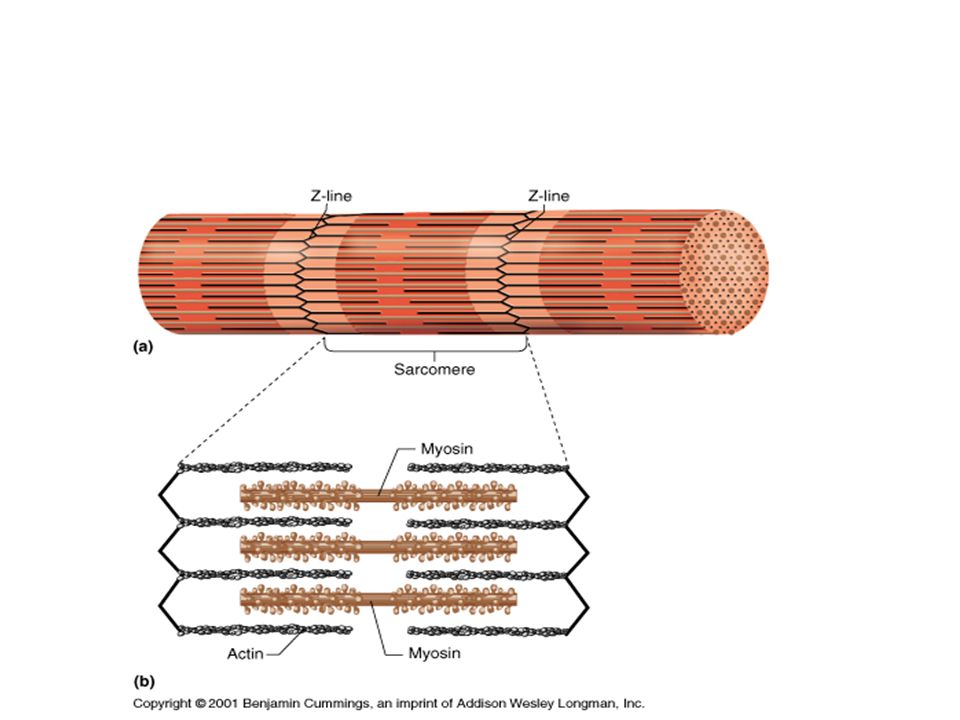 Если человек будет по 8 часов в день кататься при таком пульсе, то сердце начнет дилятировать (расширяться) и может значительно увеличиться в объеме. А этому человеку нужно заниматься в первую очередь мышцами ног — именно они ограничивают его возможности.
Если человек будет по 8 часов в день кататься при таком пульсе, то сердце начнет дилятировать (расширяться) и может значительно увеличиться в объеме. А этому человеку нужно заниматься в первую очередь мышцами ног — именно они ограничивают его возможности.
А у другого может оказаться наоборот. Вот вам следующий пример: еще один молодой перспективный лыжник, мы его тестируем, у него картина такая: пульс 190 уд/мин и 4,5 л/мин потребляет ногами, но пульс-то — 190. Всё, ему руки нельзя добавлять, он на пределе, сердце маленькое, слабое. Это было как раз в 2000 году, когда он ряд гонок выиграл и, как говорят, «капнул». Его больше в сборную брать не стали — сердце не держит. Никто же этого не знает, но чувствуют — спортсмен начинает проигрывать, не держит тренировочных нагрузок. Сердце маленькое. Наконец, дали ему отдохнуть, выбросили все объёмные нагрузки, оставили только интенсивные, спринтерского характера. Сердце постепенно вылечилось, за 4-5 месяцев стало нормальным, стало свои 8 л/мин качать, вместо 4,5 л/мин. Потребление кислорода в руках добавили, чуть ли не в 2 раза, а ноги у него и так хорошие. Он свои 4,5 л/мин ногами как потреблял, так и потребляет, но на пульсе не 190 уд/мин, а 160, потом руки еще добавляет, и он выходит на пульс 190, на этом пульсе можно бежать 10 км. У него был явный недостаток сердца, но причина не в том, что сердце плохое, ему надо было просто дать восстановиться, чтобы прекратились дистрофические явления, и он вернулся в нормальное состояние.
Потребление кислорода в руках добавили, чуть ли не в 2 раза, а ноги у него и так хорошие. Он свои 4,5 л/мин ногами как потреблял, так и потребляет, но на пульсе не 190 уд/мин, а 160, потом руки еще добавляет, и он выходит на пульс 190, на этом пульсе можно бежать 10 км. У него был явный недостаток сердца, но причина не в том, что сердце плохое, ему надо было просто дать восстановиться, чтобы прекратились дистрофические явления, и он вернулся в нормальное состояние.
Заключение
Хочется добавить о роли интуиции и знания. Про роль знания особо говорить не станем – этому посвящена вся статья. Что касается интуиции, приведем выдержку из книги Виктора Николаевича «Подготовка бегуна на средние дистанции»:
«Принцип интуиции. Каждый спортсмен должен опираться в тренировке не только на правила, но и на интуицию, поскольку имеются индивидуальные особенности адаптационных реакций».
Принцип интуиции можно переформулировать иначе – «Природа «умнее» любого ученого». Поэтому можно планировать тренировочный процесс, но при его реализации спортсмен должен «прислушиваться» к своим ощущениям, сопоставлять их с ожиданиями своими и тренера. Разумеется, необходимо как можно чаще контролировать состояние готовности различных систем и органов. Эта информация, вместе со знаниями, является основой продуктивной интуиции, творческих озарений в построении тренировочного процесса. В связи с этим действительно можно принять утверждение «Знание слепо без интуиции», когда идет работа с конкретным спортсменом, когда приходится вводить коррекцию в тренировочный процесс.
Поэтому можно планировать тренировочный процесс, но при его реализации спортсмен должен «прислушиваться» к своим ощущениям, сопоставлять их с ожиданиями своими и тренера. Разумеется, необходимо как можно чаще контролировать состояние готовности различных систем и органов. Эта информация, вместе со знаниями, является основой продуктивной интуиции, творческих озарений в построении тренировочного процесса. В связи с этим действительно можно принять утверждение «Знание слепо без интуиции», когда идет работа с конкретным спортсменом, когда приходится вводить коррекцию в тренировочный процесс.
Тяжёлый изнурительный бег вверх по горной тропе, использовавшийся Бьорном Дали и Вегардом Ульвангом, чем-то напоминает бег по холмам в Новозеландии учеников Лидьярда.
При подготовке статьи использованы разработки В. Н. Селуянова
Обсудить на форуме
комментарии к статье
Пока нет комментариев
РАЗНИЦА МЕЖДУ МИОФИБРИЛЛАМИ И МЫШЕЧНЫМ ВОЛОКНОМ | СРАВНИТЕ РАЗНИЦУ МЕЖДУ ПОХОЖИМИ ТЕРМИНАМИ — НАУКА
Есть три типа мышечных тканей; сердечные мышцы, скелетные мышцы и гладкие мышцы.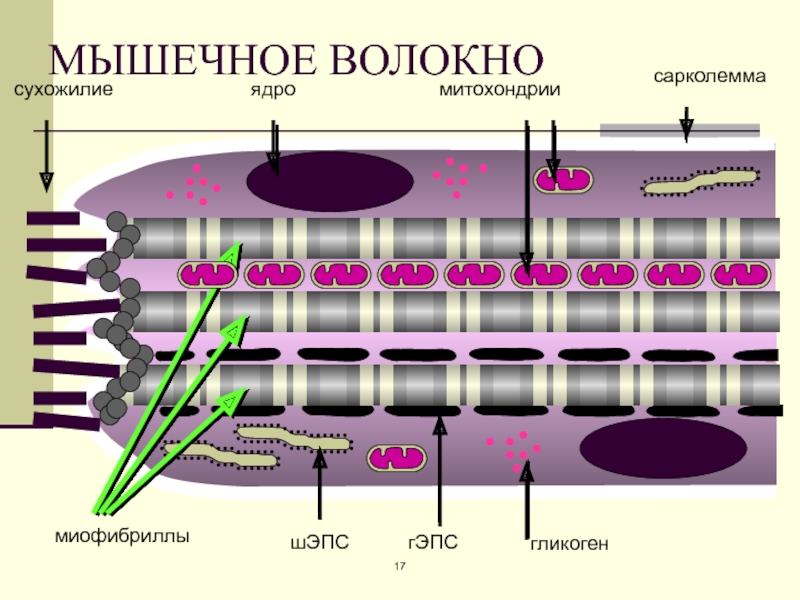 Каждый тип имеет определенную структуру и роль в мышечной системе. Гладкие мышцы образуют такие органы, как мочевой пуз
Каждый тип имеет определенную структуру и роль в мышечной системе. Гладкие мышцы образуют такие органы, как мочевой пуз
Есть три типа мышечных тканей; сердечные мышцы, скелетные мышцы и гладкие мышцы. Каждый тип имеет определенную структуру и роль в мышечной системе. Гладкие мышцы образуют такие органы, как мочевой пузырь и желудок. Сердечные мышцы сокращаются и перекачивают кровь по всему телу. Скелетные мышцы помогают костям и другим структурам двигаться. Эти мышцы состоят из длинных пучков клеток, называемых мышечными волокнами или миоцитами. Мышечные волокна состоят из тысяч миофибрилл. Ключевое различие между миофибриллами и мышечными волокнами заключается в том, что миофибрилла — это основная стержнеобразная единица мышечного волокна пока мышечное волокно — это трубчатые клетки мышцы.
1. Обзор и основные отличия
2. Что такое мышечное волокно
3.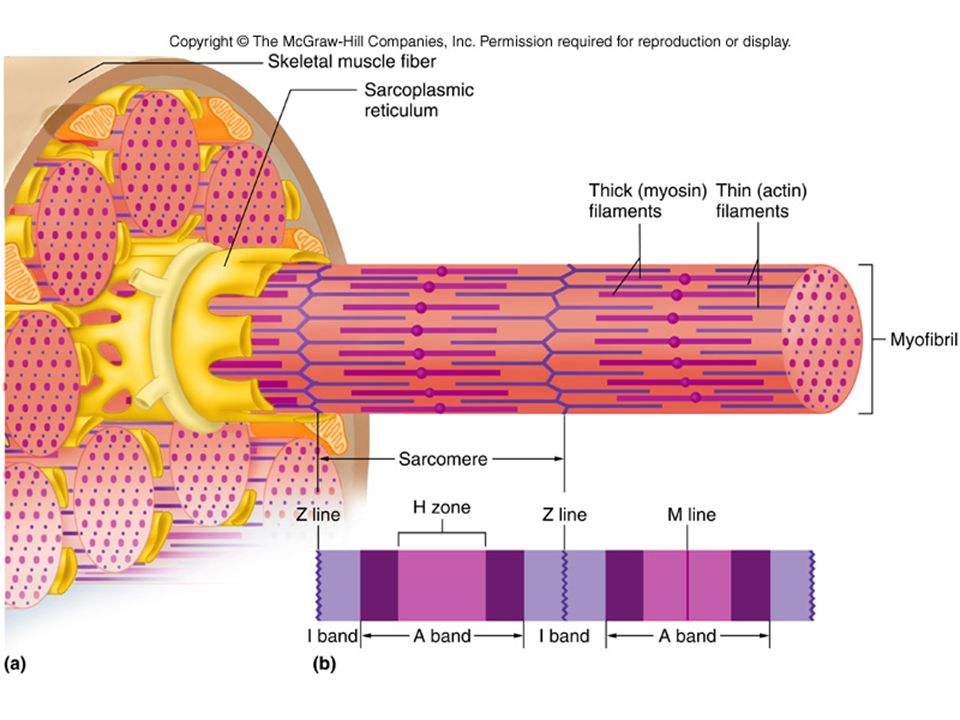 Что такое миофибрилла
Что такое миофибрилла
4. Сходства между миофибриллами и мышечными волокнами
5. Сравнение бок о бок — миофибриллы и мышечные волокна в табличной форме
6. Резюме
Что такое мышечное волокно?
Скелетные мышцы составляют большую часть мышечной системы. Скелетные мышцы защищены соединительной мышечной тканью, которая называется эпимизий. Мышца состоит из пучков трубчатых мышечных клеток. Эти трубчатые клетки известны как мышечные волокна или миоциты. Связки мышечных волокон известны как пучки. Один пучок мышечных волокон защищен соединительной тканью, известной как перимизий. Внутри перимизии находится множество мышечных волокон. Каждый пучок может содержать от 10 до 100 мышечных волокон. Крупные сильные мышцы имеют большое количество мышечных волокон внутри каждого пучка. Мышцы меньшего размера содержат меньшее количество мышечных волокон в пучке.
Мышечные ткани и мышечные волокна образуются из мезодермального слоя эмбриональных половых клеток в процессе, называемом миогенезом. Каждое мышечное волокно покрыто волокнистой соединительной тканью, называемой эндомизий. Диаметр мышечных волокон может составлять от 10 до 80 микрометров, и их можно увеличивать до 30 см в длину.
Каждое мышечное волокно покрыто волокнистой соединительной тканью, называемой эндомизий. Диаметр мышечных волокон может составлять от 10 до 80 микрометров, и их можно увеличивать до 30 см в длину.
Мышечные волокна состоят из многочисленных стержневидных единиц или цилиндрических органелл, называемых миофибриллами. Каждое мышечное волокно содержит от сотен до тысяч миофибрилл, которые представляют собой пучки белков миозина и актина, проходящие по всей длине мышечного волокна. Миофибриллы важны для сокращения мышц.
Типы мышечных волокон
Есть три основных типа клеток скелетных мышц или мышечных волокон. Это волокна типа I, волокна типа IIa и волокна типа IIb. Волокна типа I также известны как медленно сокращающиеся волокна или красные медленные волокна. Волокна типа IIa известны как быстрые окислительные волокна или красные быстрые волокна. Волокна типа IIb известны как быстрые гликолитические волокна или белые быстрые волокна.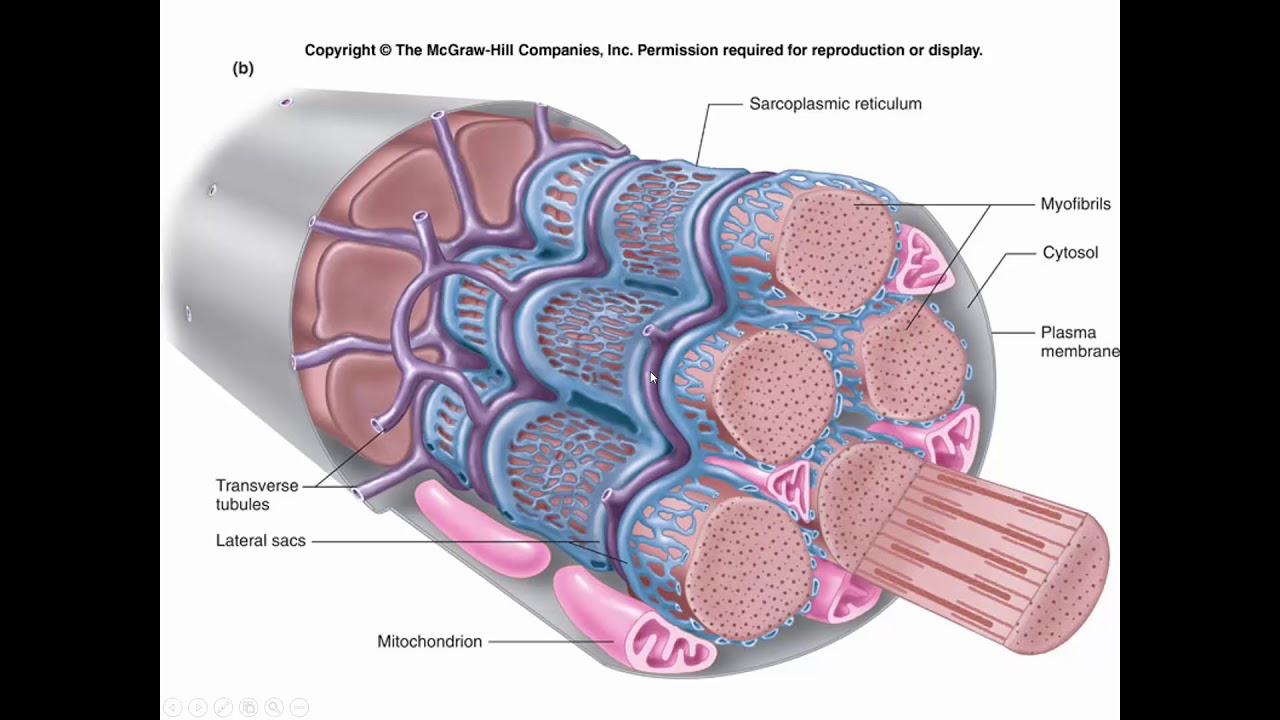 Каждый тип имеет разные функциональные характеристики.
Каждый тип имеет разные функциональные характеристики.
Что такое миофибрилла?
Миофибрилла или мышечное волокно — это основная палочковидная единица мышечной клетки. В мышечной клетке есть сотни миофибрилл, идущих параллельно друг другу. Миофибриллы состоят в основном из белков актина и миозина. Некоторые другие типы белков также присутствуют в миофибриллах. Эти белки организованы в толстые и тонкие длинные нити, называемые миофиламенты. Тонкие миофиламенты состоят в основном из белка актина, а толстые — из белка миозина. Эти два типа миофиламентов проходят по длине миофибриллы в участках, называемых саркомерами. Миофибриллы состоят из повторяющихся участков саркомеров. Эти саркомеры выглядят как чередующиеся темные и светлые полосы под микроскопом и отвечают за сокращение мышц.
Каковы сходства между миофибриллами и мышечными волокнами?
- Мышечные волокна и миофибриллы ответственны за сокращение мышц.
- Оба типа имеют трубчатую форму.

- Оба типа расположены параллельно внутри мышцы.

Какая связь между миофибриллами и мышечным волокном?
- Миофибриллы — это основные единицы мышечных волокон. Одно мышечное волокно содержит сотни миофибрилл.
В чем разница между миофибриллами и мышечным волокном?
Миофибриллы против мышечного волокна | |
| Миофибрилла — это основная стержнеобразная единица мышечного волокна. | Мышечное волокно — клетка мышцы трубчатой формы. |
| Сочинение | |
| Миофибриллы состоят из двух типов миофиламентов, называемых тонкими и толстыми нитями. | Мышечное волокно состоит из множества миофибрилл. |
| Природа | |
| Миофибрилла — это цилиндрическая органелла. | Мышечное волокно — это клетка с ядром и другими органеллами, включая митохондрии. |
Резюме — миофибриллы против мышечных волокон
Мышечное волокно — это основная единица мышцы.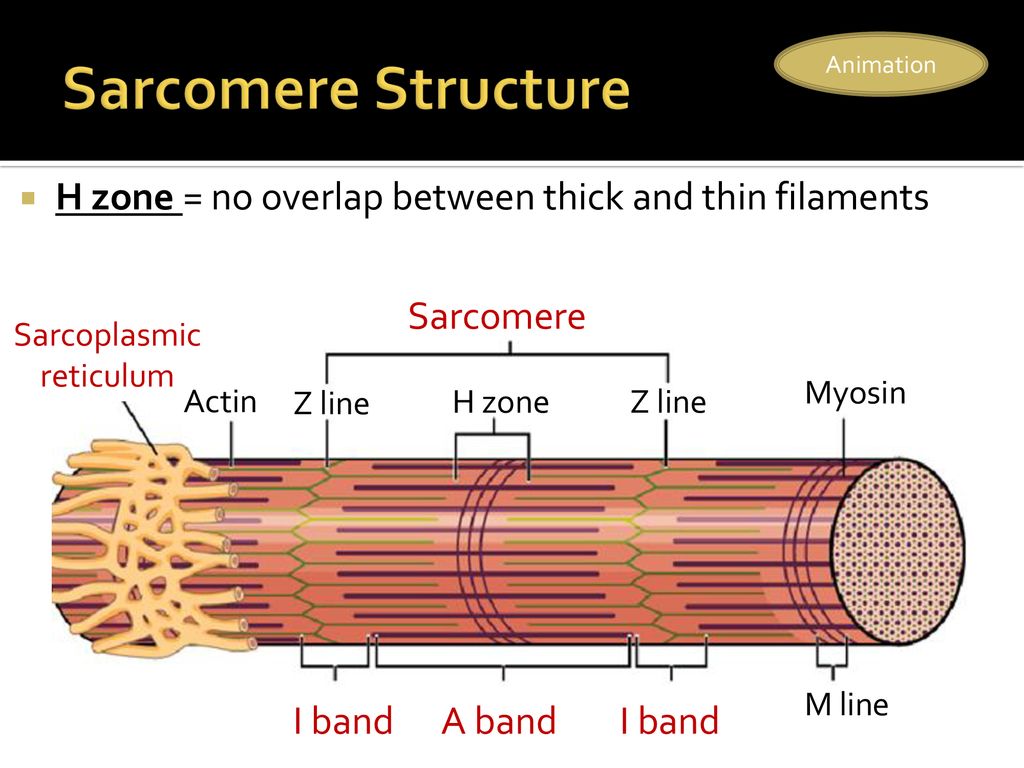 Они расположены пучками внутри мышцы. Мышечные волокна состоят из множества миофибрилл. Миофибриллы — это основные единицы мышечных волокон. Они состоят из тонких и толстых белковых нитей, называемых миофиламентами. Основными белками миофиламентов являются актин и миозин. Миофиламенты состоят из повторяющихся участков, называемых саркомерами. Их саркомеры отвечают за сокращение мышц. Мышечные волокна и миофибриллы вместе помогают сокращению мышц.
Они расположены пучками внутри мышцы. Мышечные волокна состоят из множества миофибрилл. Миофибриллы — это основные единицы мышечных волокон. Они состоят из тонких и толстых белковых нитей, называемых миофиламентами. Основными белками миофиламентов являются актин и миозин. Миофиламенты состоят из повторяющихся участков, называемых саркомерами. Их саркомеры отвечают за сокращение мышц. Мышечные волокна и миофибриллы вместе помогают сокращению мышц.
Скачать PDF-версию «Миофибриллы против мышечных волокон»
Вы можете скачать PDF-версию этой статьи и использовать ее в автономных целях в соответствии с примечаниями к цитированию. Пожалуйста, скачайте PDF-версию здесь. Разница между миофибриллами и мышечными волокнами.
Ссылки:
«Миофибриллы». Википедия. Фонд Викимедиа, 7 июля 2017 г. Web. Доступна здесь. 08 июля 2017.
«Структура мышц». Структура мышцы (и связанных соединительных тканей). N.p., n.d. Интернет. Доступна здесь. 08 июля 2017.
Изображение предоставлено:
1. «1022 мышечных волокон (малых)» от OpenStax — (CC BY 4.0) через Commons Wikimedia
«1022 мышечных волокон (малых)» от OpenStax — (CC BY 4.0) через Commons Wikimedia
2. «Миофибрилла» Слэшма из англоязычной Википедии (CC BY-SA 3.0) через Commons Wikimedia.
ПОЧЕМУ БОЛЯТ МЫШЦЫ ПОСЛЕ ТРЕНИРОВКИ?
Каждая скелетная мышца состоит из пучков мышечных волокон, которые содержат множество сократительных нитей- миофибрилл. Миофибриллы это постоянные специализированные структуры в клетках поперечнополосатых мышц, служащие для сокращения мышечных волокон.
Миофибриллы разделены перегородками (Z-пластинами) на саркомеры. Саркомеры это базовые сократительные единицы мышц. Мышцы сокращаются из-за укорочения множества саркомер, которые последовательно соединены в миофибриллах. Это описание очень упрощенно, но основной принцип передан верно.
Так вот, причиной мышечных болей считается разрыв миофибрилл в области Z-пластин. Мышечное перенапряжение в ходе выполнения упражнения вызывает перегрузку по Z-линии, вызывающую деформацию и разрыв. Организм отвечает на такую травму воспалением, как защитной реакцией. В процессе воспаления организм за счет действия макрофагов очищает место воспаления от разрушенных или негодных к дальнейшей эксплуатации клеток, таким образом подготавливая ткани к регенерации.
Мышечное перенапряжение в ходе выполнения упражнения вызывает перегрузку по Z-линии, вызывающую деформацию и разрыв. Организм отвечает на такую травму воспалением, как защитной реакцией. В процессе воспаления организм за счет действия макрофагов очищает место воспаления от разрушенных или негодных к дальнейшей эксплуатации клеток, таким образом подготавливая ткани к регенерации.
Боли в мышцах мы обязаны именно мышечным микротравмам и последующему воспалительному процессу в них. Боль в мышцах обычно запаздывает (на 8-48 часов) потому, что воспалительный процесс в поврежденной мышце развивается постепенно в зависимости от степени повреждения.
При этом мышечная боль возникает только в том случае если изменение в тренировочной нагрузке превышает 10% от привычной. Если тренироваться регулярно и тренировочный объем повышать постепенно, то мышечных болей можно будет избежать. Именно по этой причине рекомендуют повышать недельный объем не более чем на 10%.
Вы спросите, а как же распространенное мнение о том, что это молочная кислота провоцирует мышечную боль?
Так я вам отвечу, что это полная ерунда!
90 % молочной кислоты утилизируется организмом еще во время тренировки, оставшиеся же 10 % полностью утилизируются организмом в течении ближайшего часа после окончания тренировки..jpg/1280px-1022_Muscle_Fibers_(small).jpg) Молочная кислота тут ни при чем.
Молочная кислота тут ни при чем.
На фото, сделанном с помощью электронного микроскопа (методом биопсии тканей), вы можете увидеть как выглядят мышцы до произведенного усилия (часть фото А) и после произведенного усилия (часть фото В). Наглядно видно какие структурные разрушения в мышечных клетках происходят. Именно эти разрушения и вызывают воспалительную реакцию организма выливающуюся в болезненные ощущения в мышцах.
Соответствующие исследования:
http://www.ncbi.nlm.nih.gov/pubmed/7250326
http://www.ncbi.nlm.nih.gov/pubmed/6698677
http://www.ncbi.nlm.nih.gov/pubmed/6629599
Мышечные диски — Справочник химика 21
Мышечные волокна построены из продольно расположенных фибрилл (миофибрилл) диаметром ок.
 1000 нм, в к-рых чередуются светлые и темные диски (соотв. I и А-диски рис. 2). В середине диска I расположена пластинка [c.93]
1000 нм, в к-рых чередуются светлые и темные диски (соотв. I и А-диски рис. 2). В середине диска I расположена пластинка [c.93]В световом микроскопе в миофибриллах видны поперечные линии, отстоящие друг от друга на расстоянии примерно 2,5 мкм (рис. 4-21 и 4-22). Область между двумя плотными Z-пластинками, называемая саркомером, является главным сократительным элементом мышечной клетки. В центре саркомера имеется плотная анизотропная полоса (обладающая сильным двойным лучепреломлением), получившая название А-диска. Продолжением Z-пластинок являются менее интенсивные [c.318]
Миофибриллы — это сократительные элементы мышечного волокна, количество которых может достигать нескольких тысяч. Под микроскопом миофибриллы имеют поперечную исчерченность в виде повторяющихся темных и светлых участков или дисков (рис. 115). Темные участки, или Л-диски в центре имеют более светлую /-/-зону, посредине которой проходит темная М-линия. Светлые участки, или /-диски в центре пересекаются узкой 7-линией (2-мембраной). [c.292]
[c.292]
Общий процесс сокращения, проявляющийся в укорочении мышечного волокна и развитии напряжения, является результатом суммирования одновременного образования большого числа спаек по всей длине миофибрилл, вовлеченных в процесс сокращения возбужденной мышцы. Величина напряжения в сокращающейся мышце будет пропорциональна количеству поперечных спаек или площади их наложения в пределах каждого саркомера, что диктуется нервной системой (рис. 120). При значительном растяжении мышцы (длине саркомера более 3,65 мкм) тонкие нити полностью выходят за пределы дисков А и напряжение в мышце отсутствует, По мере вхождения тонких нитей между толстыми и увеличения площади их наложения друг на друга напряжение в мышце постепенно увеличивается, достигая максимума при длине саркомера от 2,25 до 2,00 мм. При более значительном сокращении мышцы тонкие нити перекрываются в центре дисков А и сжимаются в зоне Н, образуя полосу сокращения. Напряжение мышцы в этой стадии сокращения быстро снижается. [c.302]
[c.302]
Светлые участки миофибрилл состоят из центральных частей тонких нитей. Они сравнительно легко пропускают лучи света или поток электронов, так как не обладают двойным лучепреломлением и называются изотропными, или светлыми, дисками (1-диски). В середине пучка тонких нитей поперечно располагается тонкая пластинка из белка, которая фиксирует положение мышечных нитей в пространстве. Эта пластинка хорошо видна под микроскопом в виде линии, идущей поперек 1-диска, и названа Z-пластинкой, или Z-линией (см. рис. 9 и 10). [c.128]
При уменьшении длины саркомера сжимается только 1-диск, тогда как плотный А-диск не изменяет своих размеров. Это легко объяснить, предположив, что сокращение миофибриллы происходит в результате скольжения толстых филаментов относительно тонких, причем длина тех и других остается неизменной (рис. 10-5). Эта простая модель скользящих нитей, впервые предложенная в 1954 г., имела основополагающее значение для понимания природы мышечного сокращения.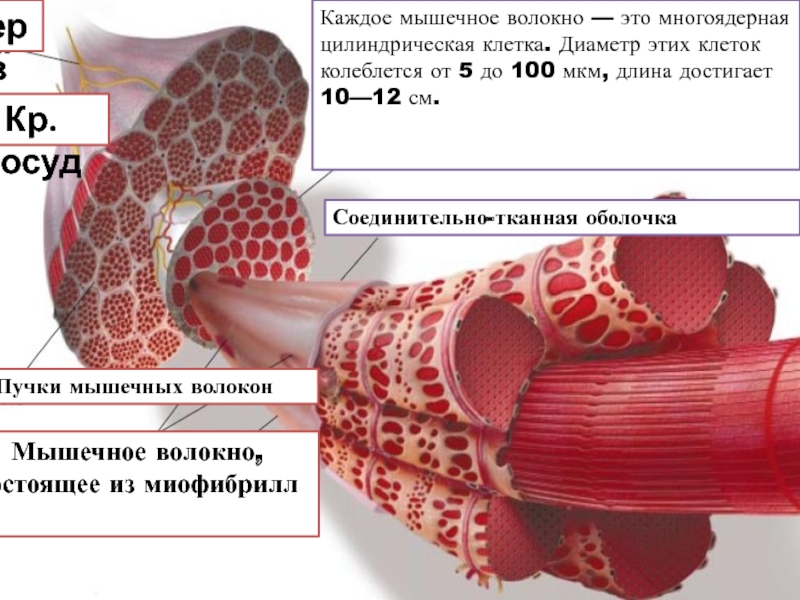 В частности, она привлекла внимание к молекулярным механизмам скольжения филаментов. [c.77]
В частности, она привлекла внимание к молекулярным механизмам скольжения филаментов. [c.77]
Саркомер—это функциональная единица мышцы. Саркомеры следуют друг за другом вдоль оси фибриллы, повторяясь через каждые 1500 — 2300 нм (рис. 56.1). При исследовании миофибриллы в электронном микроскопе выявляется чередование темных и светлых дисков (диски А и I). Центральная область диска А (зона Н) выглядит менее плотной, чем остальная его часть. Диск I делит пополам очень плотная и узкая линия Z. Эти детали мышечной структуры представлены на рис. 56.2. [c.333]
Саркоплазма скелетных мышц содержит большие запасы гликогена, локализованного в гранулах, примыкающих к 1-диску. Высвобождение глюкозы из гликогена зависит от специфического мышечного фермента гликогенфосфорилазы (см. гл. 19). Для того чтобы обеспечить образование глюкозо- [c.340]
Следует отметить, что не во всех мышечных клетках организма процесс сопряжения происходит, как в кардиомиоците.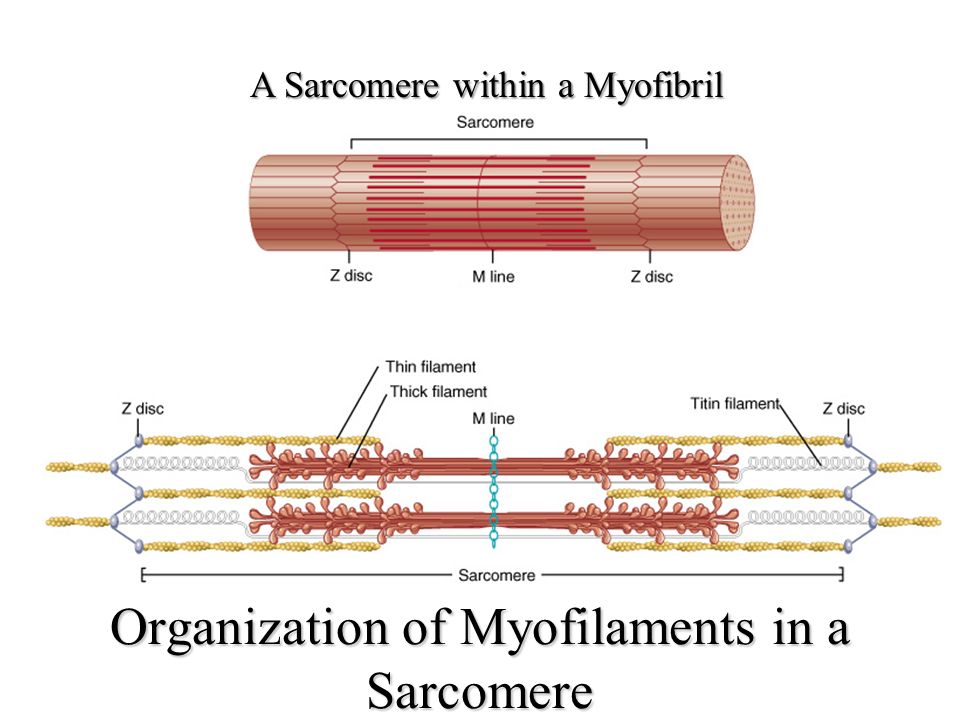 Так, в скелетных мышцах теплокровных потенциал действия короткий (2-3 мс) и медленный поток ионов кальция в них отсутствует. В этих клетках сильно развита Т-система поперечных трубочек, подходящих непосредственно к саркомерам близко к /-дискам (см. рис. 7.11). Изменения мембранного потенциала во время деполяризации через Т-систему передается в таких клетках непосредственно на мембрану СР, вызывая залповое высвобождение ионов Са + и дальнейшую активацию сокращения (3, 4, 5). [c.161]
Так, в скелетных мышцах теплокровных потенциал действия короткий (2-3 мс) и медленный поток ионов кальция в них отсутствует. В этих клетках сильно развита Т-система поперечных трубочек, подходящих непосредственно к саркомерам близко к /-дискам (см. рис. 7.11). Изменения мембранного потенциала во время деполяризации через Т-систему передается в таких клетках непосредственно на мембрану СР, вызывая залповое высвобождение ионов Са + и дальнейшую активацию сокращения (3, 4, 5). [c.161]
Она оказалась поистине удивительной. Кружки и эллипсы, видные на одиночных срезах, в действительности были сечением через различные элементы единой трехмерной конструкции, пронизывающей всю мышечную клетку. Конструкция состояла из сетей, расположенных вблизи Z-дисков, и колонн, перпендикулярных сетям и соединяющих их друг с другом. Ни одной шарообразной митохондрии, не связанной с основным каркасом, обнаружено не было. [c.176]
Митохондрии на уровне линии 2-диска представлены типичным митохондриальным ретикулумом — единой митохондриальной системой.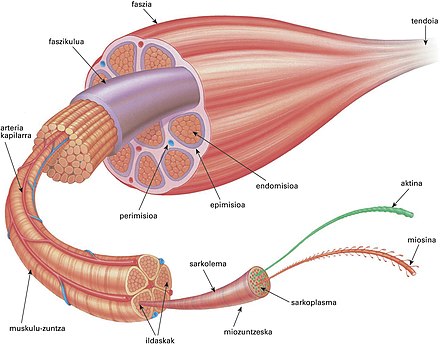 Такой этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных «поэтажных пластов митохондриального ретикулума. [c.23]
Такой этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных «поэтажных пластов митохондриального ретикулума. [c.23]
Что же за мотор приводит в движение бактериальные жгутики На представленной выще электронно-микроскопической фотографии виден крючок , прикрепленный к проходящей сквозь клеточную стенку палочке, которая в свою очередь прикреплена к тонкому диску (М-кольцу), погруженному в цитоплазматическую мембрану. Схематически это устройство у грамположительных бактерий показано выще. По-видимому, между М-кольцом и S-кольцом, укрепленным в клеточной стенке, возникает крутящий момент. Данные о наличии в моторе белков типа мышечных отсутствуют, и было высказано предположение о том, что из внутриклеточ- [c.283]
Согласно скользящей модели, напряжение, развиваемое мышцей, целиком определяется нитями актина и миозина и 7-дисками. Все эти элементы не вполне жестки, они обладают определенной податливостью.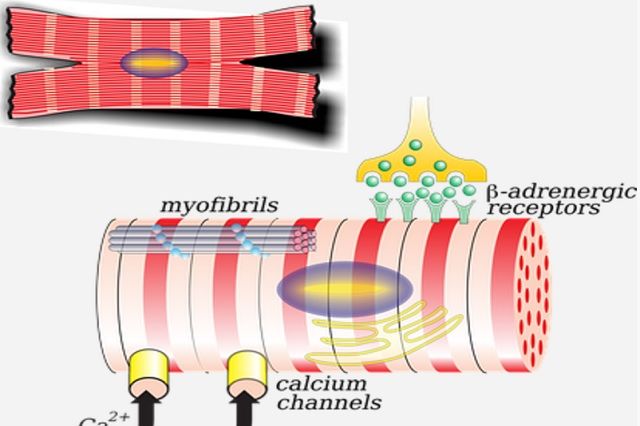 Конечные саркомеры мышечного волокна связаны с соединительной тканью сухожилий, и здесь также имеется податливость, пластичность. Одновременно эти элементы вносят некоторую упругость в движение мышцы. Однако общий вклад упругих и пластических деформаций не превышает 3% развиваемого мышцей напряжения. Все же следует рассматривать мышцу как вязкоупругое тело. Как мы увидим, уравнение Хилла списывает только вязкое течение в мышце. [c.401]
Конечные саркомеры мышечного волокна связаны с соединительной тканью сухожилий, и здесь также имеется податливость, пластичность. Одновременно эти элементы вносят некоторую упругость в движение мышцы. Однако общий вклад упругих и пластических деформаций не превышает 3% развиваемого мышцей напряжения. Все же следует рассматривать мышцу как вязкоупругое тело. Как мы увидим, уравнение Хилла списывает только вязкое течение в мышце. [c.401]
Хроническое отравление. Животные. У собак после 7-месячной имплантации дисков металлического Т. в мышечную ткань не выявлено раздражающего или кожнорезорбтивно-го действия, либо нарушения процесса заживления ран металл инкапсулировался фиброзной тканью. Отсутствие реакции костной ткани установлено через 120—180 дней после фиксации металлом переломов костей у собак. Трехнедельное интратрахеальное введение 75 мг метатитаната бария не выявило у морских свинок никаких фибротических реакций в легких в течение последующего 12-месячного наблюдения.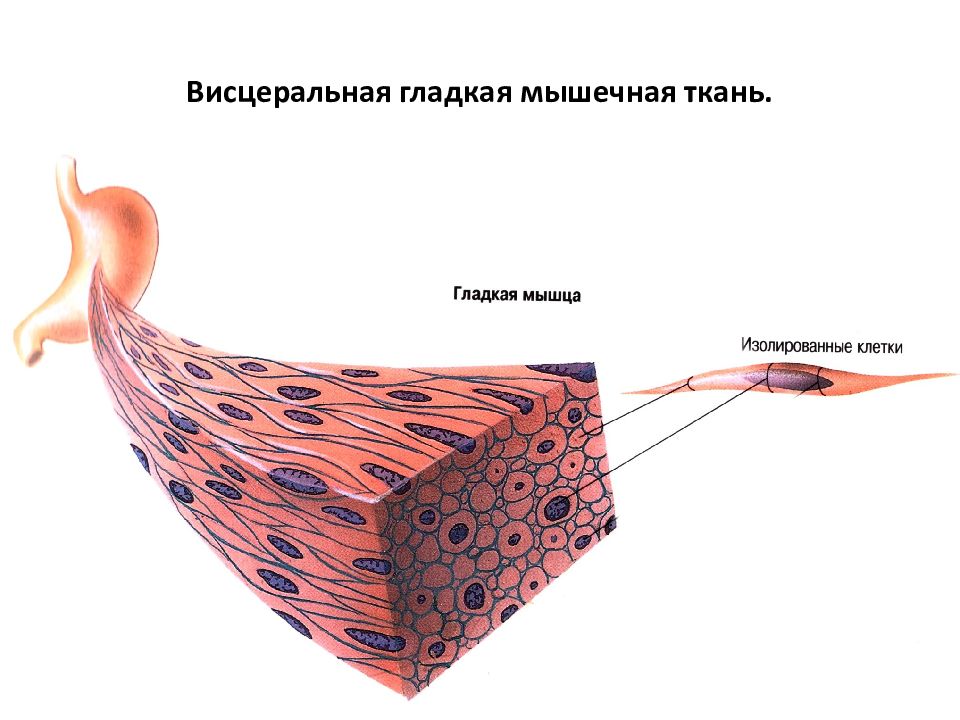 Трехмесячная ежедневная по 6 ч ингаляция волокон октатитаната калия в дозах (2,9-7-41,8) 10 волокон в 1 л вызывала дозозависимый фиброз легких у крыс, хомяков и морских свинок через 15—24 мес. Аналогичный фиброз вызывало у крыс ин- [c.440]
Трехмесячная ежедневная по 6 ч ингаляция волокон октатитаната калия в дозах (2,9-7-41,8) 10 волокон в 1 л вызывала дозозависимый фиброз легких у крыс, хомяков и морских свинок через 15—24 мес. Аналогичный фиброз вызывало у крыс ин- [c.440]
В заключение можно указать на морфологические изменения, происходящие в мышцах груди и конечностей мух, затравленных 5%-пым водным раствором дуолитовой эмульсии (препарат ДДТ ). Степень парализованности соответствовала изменениям, происходившим в структуре мышц. В ядрах мышечных волокон ДДТ вызывал образование скоплений хроматина, которые окрашиваются красителем в яркий цвет. В поперечно-полосатых волокнах отмечалось исчезновение отдельных темных дисков фибриллы на концах мышечных волокон слипались в широкие лепты. Некоторые изменения отмечались и в морфологии мышц, как, например, появление мышечных волокон с несколькими концентрическими слоями фибрилл. [c.118]
В этой связи весьма интересны исследования Ш т ю б е л я (11923 г.
 ), Муральта (1933 г.), Шмидта (1937 г.) и др. над мышечным волокном. Известно, что миофибриллы слагаются из правильно чередующихся участков (см. рис. 102), один из которых при обычных условиях микроскопического наблюдения кажутся более темными, другие более светлыми. Это чередование и обусловливает поперечную исчерченность мышечного волокна. Светлые и темные участки называются дисками и являются основными структурными элементами -миофибриллы. При исследовании в -поляризованном свете темные диски обнаруживают положительное двойное [c.328]
), Муральта (1933 г.), Шмидта (1937 г.) и др. над мышечным волокном. Известно, что миофибриллы слагаются из правильно чередующихся участков (см. рис. 102), один из которых при обычных условиях микроскопического наблюдения кажутся более темными, другие более светлыми. Это чередование и обусловливает поперечную исчерченность мышечного волокна. Светлые и темные участки называются дисками и являются основными структурными элементами -миофибриллы. При исследовании в -поляризованном свете темные диски обнаруживают положительное двойное [c.328] Поляризационные исследования Штюбеля показали, что диски Ц состояли из палочкообразных упорядоченных мицелл белка миозина, расположенных длинником по оси мышечного волокна. Эти мицеллы обладают, кроме того, положительным собственным преломлением. Следует сказать, что подобную картину дает и электронный микроскоп. На основании всестороннего г изучения строения миофибрилл различными методами установлено, что каждая мышечная фибрилла строится из вытянутых по ее длиннику молекул белка миозина, обладающих упорядоченной складчатостью. [c.383]
[c.383]
В жировом теле, трахейных матрицах и в гиподерме пораженных насекомых появлялись пятигранные (в проекции) полиэдры. С развитием болезни вирусом поражаются и другие органы. Смит и Ксерос [232] обнаружили полиэдры также в шелкоотделительных железах, в мышечных тканях, в нервных ганглиях и в имагиналь-ных дисках. В материалах, который исследовала Лотмар, имелась двойная инфекция — цитоплазменного и ядерного вирусов, поэтому она обнаружила полиэдры также в кишечнике и в мальпигиевых сосудах гусениц. [c.108]
Стенка сердца образована сердечными мышечными волокнами, соединительной тканью и мелкими кровеносными сосудами. Каждое мышечное волокно (кардиомиоцит) содержит одно или два ядра, множество крупных митохондрий и множество параллельных друг другу миофибрилл. Миофибриллы образованы актиновыми и миозиновыми нитями (миофиламентами), которые обеспечивают сокращение кардиомиоци-та подобно тому, как это происходит в скелетной мышце (разд. 18.4). В принципе внутреннее строение кардиомиоцитов такое же, как у волокон скелетных мышц, поэтому под микроскопом они также выглядят поперечно-полосатыми (рис. 14.15 и 14.16). Темные полосы, называемые интеркалярными или вставочными дисками, представляют собой поверхностные клеточные мембраны, отделяющие одну мышечную клетку от другой. Мембраны модифицированы, что по- [c.155]
18.4). В принципе внутреннее строение кардиомиоцитов такое же, как у волокон скелетных мышц, поэтому под микроскопом они также выглядят поперечно-полосатыми (рис. 14.15 и 14.16). Темные полосы, называемые интеркалярными или вставочными дисками, представляют собой поверхностные клеточные мембраны, отделяющие одну мышечную клетку от другой. Мембраны модифицированы, что по- [c.155]
Микроскопическое изучение строения миофибрилл показало, что они состоят из чередующихся светлых и темных участков, или дисков. В мышечных клетках миофибриллы располагаются таким образом, что светлые и темные участки рядом расположе1шых миофибрилл совпадают, что создает видимую под микроскопом поперечную исчертан-ность всего мышечного волокна (рис. 9). [c.127]
Актиновые филаменты заякорены своими плюс-концами в Z-диске, где их удерживают в правильно организованной решетке другие белки. Из них лучше всего охарактеризован а-актинин — актин-связывающий белок, имеющийся в большинстве животных клеток.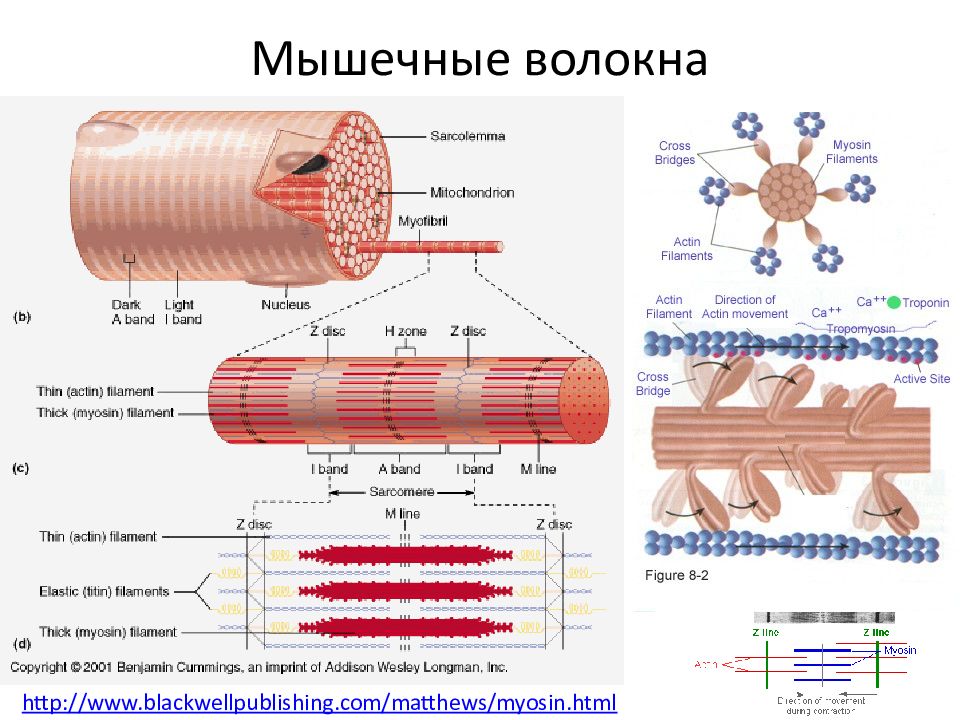 В мышечных клетках он находится в области Z-диска. Очищенный а-актинин — биполярная палочковидная молекула (рис. 11-20), которая может связывать актиновые филаменты в параллельные пучки. Аналогичную функцию в случае миозина может выполнять белок миомезин, который сшивает соседние миозиновые филаменты в области М-линии (посередине биполярного толстого филамента собирая их в гексагональную упаковку. Стабилизирует упаковку миозиновых филаментов еще одна группа миозин-связывающих белков, выявляемых при окраске антителами как серия из 11 регулярно расположенных бледных полосок по обе стороны от М-линии. [c.267]
В мышечных клетках он находится в области Z-диска. Очищенный а-актинин — биполярная палочковидная молекула (рис. 11-20), которая может связывать актиновые филаменты в параллельные пучки. Аналогичную функцию в случае миозина может выполнять белок миомезин, который сшивает соседние миозиновые филаменты в области М-линии (посередине биполярного толстого филамента собирая их в гексагональную упаковку. Стабилизирует упаковку миозиновых филаментов еще одна группа миозин-связывающих белков, выявляемых при окраске антителами как серия из 11 регулярно расположенных бледных полосок по обе стороны от М-линии. [c.267]
В мышечных клетках есть также целая система очень плохо растворимых белковых филаментов, которые можно выделить лишь после полной экстракции миозина и актина из саркомера концентрированным раствором йодистого калия. Одна группа таких филаментов, построенных из очень крупного белка, названного титином. тянется параллельно толстым и тонким филаментам и соединяет толстые филаменты с Z-диском.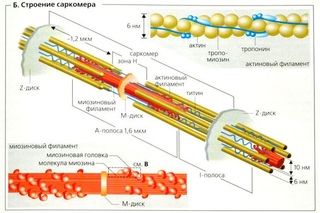 Хитиновые филаменты очень эластичны и, по-видимому, действуют как пружины, центрируя толстые филаменты между Z-дисками (рис. 11-21). Еще одна группа нерастворимых нитей — это промежуточные филаменты (разд. 11.5), которые расположены между Z-дисками соседних миофибрилл. Предполагается, что они удерживают саркомеры в определенных пространственных отношениях между собой и соединяют миофибриллы с плазматической мембраной мышечной клетки. [c.267]
Хитиновые филаменты очень эластичны и, по-видимому, действуют как пружины, центрируя толстые филаменты между Z-дисками (рис. 11-21). Еще одна группа нерастворимых нитей — это промежуточные филаменты (разд. 11.5), которые расположены между Z-дисками соседних миофибрилл. Предполагается, что они удерживают саркомеры в определенных пространственных отношениях между собой и соединяют миофибриллы с плазматической мембраной мышечной клетки. [c.267]
Миозин является белком многих качеств. В сокращении скелетных, сердечных и гладких мышц и во внутриклеточных движениях он одновременно выполняет, по крайней мере, три ключевых функции — структурную, аллостерическую и ферментативную. Наиболее полезная информация о функциях миозина была получена при исследовании поперечнополосатых скелетных мышц, сокращающихся произвольно, а также аналогичных тканей беспозвоночных, прежде всего летательных мышц насекомых. Электронно-микроскопическое изучение продольных и поперечных тонких срезов скелетных мышц, впервые проведенное в 1953 г. X. Хаксли, выявило высокий уровень их структурной организации [439]. Уже в следующем году X. Хаксли вместе с Дж. Хенсоном предложили так называемую модель скользящих нитей, которая имела основополагающее значение для понимания природы и молекулярного механизма мышечных сокращений [440]. Скелетные мышцы — это пучки мышечных волокон, наиболее крупным повторяющимся структурным элементом которых является миофибрилла — цилиндрическая нить диаметра 1-2 мкм (1000-2000 А), идущая от одного конца клетки до другого. Миофибрилла, в свою очередь, содержит белковые филамен-ты двух типов толстые и тонкие. Основной белок толстых нитей — миозин, тонких — актин. Миозиновые и актиновые филаменты в миофиб-рилле строго упорядочены. Функциональной сократительной единицей миофибриллы является саркомера, имеющая длину около 2,5 мкм и разделяющаяся на I- и А-диски (рис. 1.31). Толстые филаменты (длина 1,6 мкм и толщина 0,015 мкм) тянутся от одного края А-диска до другого, а тонкие (длина 1,0 мкм и толщина 0,008 мкм) идут от [c.
X. Хаксли, выявило высокий уровень их структурной организации [439]. Уже в следующем году X. Хаксли вместе с Дж. Хенсоном предложили так называемую модель скользящих нитей, которая имела основополагающее значение для понимания природы и молекулярного механизма мышечных сокращений [440]. Скелетные мышцы — это пучки мышечных волокон, наиболее крупным повторяющимся структурным элементом которых является миофибрилла — цилиндрическая нить диаметра 1-2 мкм (1000-2000 А), идущая от одного конца клетки до другого. Миофибрилла, в свою очередь, содержит белковые филамен-ты двух типов толстые и тонкие. Основной белок толстых нитей — миозин, тонких — актин. Миозиновые и актиновые филаменты в миофиб-рилле строго упорядочены. Функциональной сократительной единицей миофибриллы является саркомера, имеющая длину около 2,5 мкм и разделяющаяся на I- и А-диски (рис. 1.31). Толстые филаменты (длина 1,6 мкм и толщина 0,015 мкм) тянутся от одного края А-диска до другого, а тонкие (длина 1,0 мкм и толщина 0,008 мкм) идут от [c. 120]
120]
Обратный пьезоэффект — изменение линейных размеров кристаллов под действием электрического поля 26, 76]. Пьезоэлектрнками являются ориентированные пленкн ДНК (80, 81], древесина [2], поли- у-бензил-/.-глутамат [81], холестерин [26], мочевина [26], лактоза, нуклеопротеиды [57, 72], сухие и невысушенные кости человека [69, 70] (рнс. 9.4), сухожилия [98], диски поперечнополосатых мышечных волокон [33], коллагеновые структуры [71]. [c.158]
Миофибриллы, образующие сердечную мышцу, ветвятся каждая мышечная клетка контактирует через вставочные диски с несколькими клетками. Миофибрил-ла представляет собой ряд последовательно расположенных саркомеров. В 2-линиях центральные области содержат а-актинин и актин, эти области окружены десминовыми филаментами. Отходящие от 2-дисков тонкие нити состоят из сердечномышечного а-актина и покрыты сер- [c.49]
Высокая степень регулярности строения мышц привлекла внимание к процессу их развития как к модели для изучения путей возникновения пространственной организации. Перераспределение некоторых мышечных белков во время развития мышц прослежено в деталях. Так, а-актинин в развивающихся скелетномышечных клетках расположен сначала так же, как в фибробластах и гладкомышечных клетках, т. е. в виде точечных -скоплений вдоль волокон натяжения, а примерно на четвертый день, когда начинается формирование саркомеров, он обнаруживается в центральных областях г-дисков. На ранних стадиях развития культивируемых мышечных клеток филамин тоже распределен, как в фибробластах, т.е. вдоль волокон натяжения затем он на несколько дней исчезает, а потом появляется на периферии 2-дисков. В период исчезновения филамина происходит переключение его экспрессии на ранних стадиях синтезируется белок, сходный с филамином фибробластов и гладких мышц или даже идентичный ему, а тот белок, который появляется позднее и локализуется на периферии 2-дисков, является уже другой, отличающейся по своим биохимическим свойствам формой филамина [81]. Десмин не обнаруживается в мышечных клетках до тех пор, пока они не перестают делиться и не начинают экспрессировать специфические мышечные белки.
Перераспределение некоторых мышечных белков во время развития мышц прослежено в деталях. Так, а-актинин в развивающихся скелетномышечных клетках расположен сначала так же, как в фибробластах и гладкомышечных клетках, т. е. в виде точечных -скоплений вдоль волокон натяжения, а примерно на четвертый день, когда начинается формирование саркомеров, он обнаруживается в центральных областях г-дисков. На ранних стадиях развития культивируемых мышечных клеток филамин тоже распределен, как в фибробластах, т.е. вдоль волокон натяжения затем он на несколько дней исчезает, а потом появляется на периферии 2-дисков. В период исчезновения филамина происходит переключение его экспрессии на ранних стадиях синтезируется белок, сходный с филамином фибробластов и гладких мышц или даже идентичный ему, а тот белок, который появляется позднее и локализуется на периферии 2-дисков, является уже другой, отличающейся по своим биохимическим свойствам формой филамина [81]. Десмин не обнаруживается в мышечных клетках до тех пор, пока они не перестают делиться и не начинают экспрессировать специфические мышечные белки.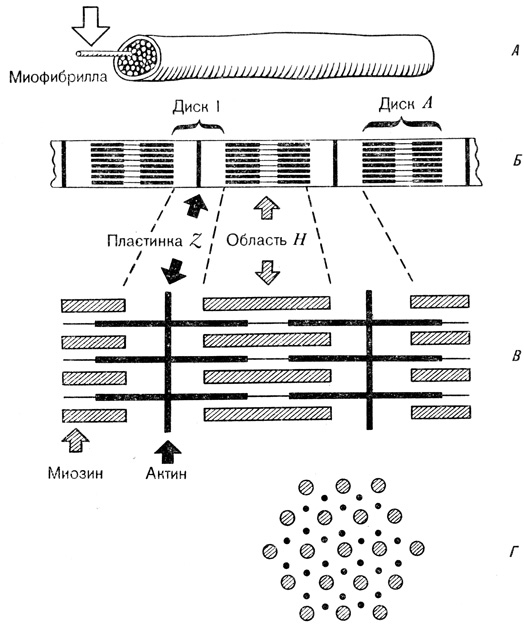 В одноядерных миобластах десмин образуется лишь в небольшом количестве, синтез его происходит главным образом в многоядерных мышечных трубках. В течение первых нескольких дней после начала его экспрессии десмин выявляется в мышечных клетках в виде системы филаментов, сходной, например, с сетью [c.52]
В одноядерных миобластах десмин образуется лишь в небольшом количестве, синтез его происходит главным образом в многоядерных мышечных трубках. В течение первых нескольких дней после начала его экспрессии десмин выявляется в мышечных клетках в виде системы филаментов, сходной, например, с сетью [c.52]
Для правильного развития мышцы важное значение имеют два несократительных белка. На протяжении всего развития в ней присутствуют тубулин и микротрубочки. Микротрубочки ориентированы обычно параллельно длинной оси развивающейся мышечной клетки, нарушение их функционирования отрицательно сказывается на мышечном развитии (к этому вопросу мы еще вернемся позднее). Другой несократительный белок развивающейся мышечной клетки — это виментин. В диффузно распределенном виде он сохраняется в клетке даже после начала формирования саркомеров и появления в них -актинина. Характер распределения виментина в мышце остается, впрочем, предметом споров. Одна группа исследователей находит виментин в -дисках, причем примерно тогда же, когда в них есть и десмин [84].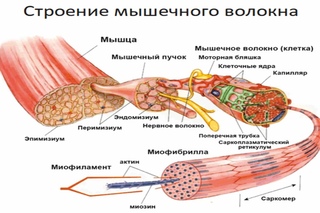 Однако двум другим лабораториям обнаружить виментин в сар-комерах не удается [85, 86]. [c.53]
Однако двум другим лабораториям обнаружить виментин в сар-комерах не удается [85, 86]. [c.53]
На стадии 4—12 сомитов в развивающемся сердце человека в кардиомиоцитах появляются миофибриллы. Сначала специфические соединения между миоцитами отсутствуют и клетки ведут себя как когезивный пласт. Вскоре образуются апикальные комплексы, которые развиваются во вставочные диски. Примерно в возрасте 22 дней начинаются синхронизированные сокращения мышечных клеток, причем электрическая связь осуществляется через нексусы.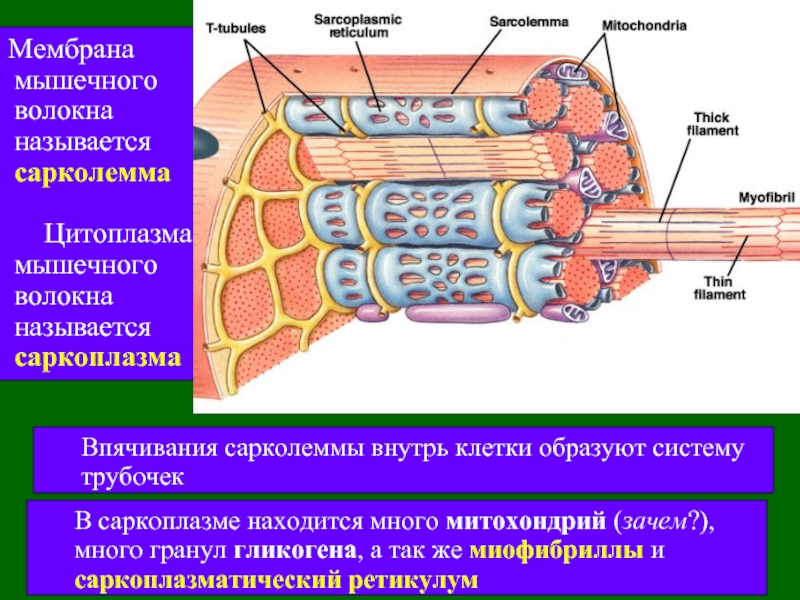 [c.90]
[c.90]
Разница между миофибриллой и мышечным волокном (Наука и природа)
Ключевая разница — миофибриллы против мышечного волокнаЕсть три типа мышечных тканей; сердечные мышцы, скелетные мышцы и гладкие мышцы. Каждый тип имеет определенную структуру и роль в мышечной системе. Гладкие мышцы создают такие органы, как мочевой пузырь и желудок. Сердечные мышцы сокращаются и перекачивают кровь по всему телу. Скелетные мышцы помогают костям и другим структурам в их движениях. Эти мышцы состоят из длинных пучков клеток, называемых мышечными волокнами или миоцитами. Мышечные волокна состоят из тысяч миофибрилл. Основное различие между миофибриллой и мышечным волокном состоит в том, что миофибрилла является основным стержнеобразным звеном мышечного волокна пока мышечное волокно представляет собой трубчатые клетки мышц.
СОДЕРЖАНИЕ
1. Обзор и основные отличия
2. Что такое мышечное волокно
3.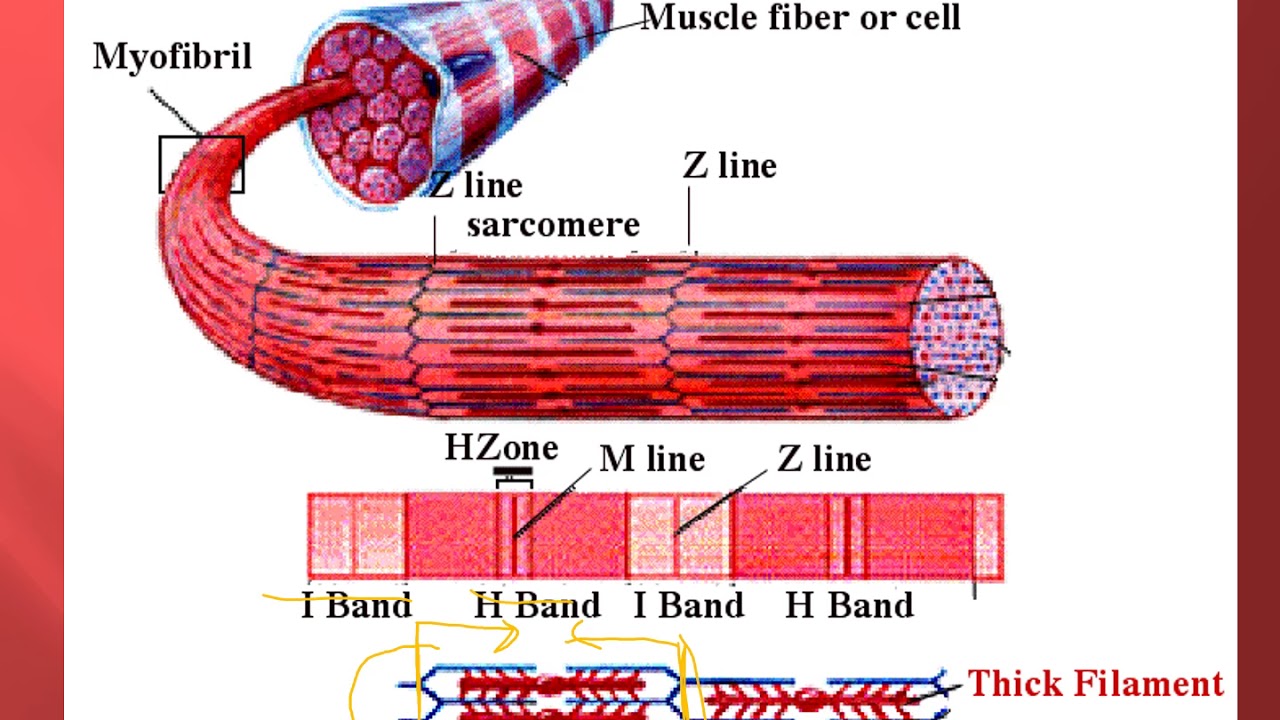 Что такое миофибрилла
Что такое миофибрилла
4. Сходство между миофибриллой и мышечным волокном
5. Сравнение бок о бок — миофибрилла и мышечное волокно в форме таблицы
6. Резюме
Что такое мышечное волокно?
Скелетные мышцы представляют собой большую часть мышечной системы. Скелетные мышцы защищены соединительной мышечной тканью epimysium. Мышца состоит из пучков трубчатых мышечных клеток. Эти трубчатые клетки известны как мышечные волокна или миоциты. Связки мышечных волокон известны как fasciculi. Один пучок мышечных волокон защищен соединительной тканью, известной как perimysium. В перимисии много мышечных волокон. Каждый пучок может содержать от 10 до 100 мышечных волокон. Большие, сильные мышцы имеют большое количество мышечных волокон внутри каждого пучка. Меньшие мышцы содержат меньшее количество мышечных волокон в пучке.
Мышечные ткани и мышечные волокна формируются из мезодермального слоя зародышевых клеток за счет процесса, называемого миогенезом.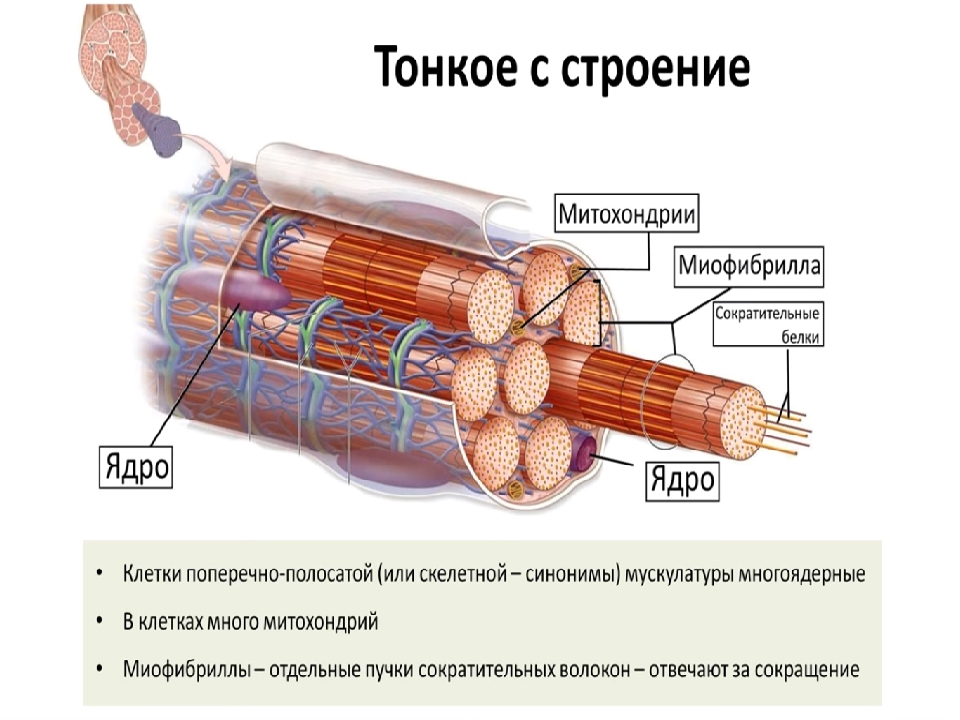 Каждое мышечное волокно покрыто волокнистой соединительной тканью эндомизий. Диаметр мышечных волокон может варьироваться от 10 до 80 микрометров, а их длина может быть увеличена до 30 см..
Каждое мышечное волокно покрыто волокнистой соединительной тканью эндомизий. Диаметр мышечных волокон может варьироваться от 10 до 80 микрометров, а их длина может быть увеличена до 30 см..
Мышечное волокно состоит из многочисленных стержнеобразных единиц или цилиндрических органелл, называемых миофибриллами. Каждое мышечное волокно содержит от сотен до тысяч миофибрилл, которые представляют собой пучки белков миозина и актина, проходящих по длине мышечного волокна. Миофибриллы важны для сокращения мышц.
Рисунок 01: Структура мышечных волокон
Типы мышечных волокон
Существует три основных типа скелетных мышечных клеток или мышечных волокон. Это волокна типа I, волокна типа IIa и волокна типа IIb. Волокна типа I также известны как медленные волокна или красные медленные волокна. Волокна типа IIa известны как быстрые окислительные волокна или красные быстрые волокна. Волокна типа IIb известны как быстрые гликолитические волокна или белые быстрые волокна.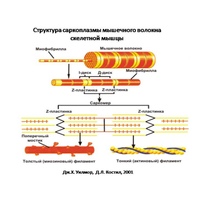 Каждый тип имеет разные характеристики в функции.
Каждый тип имеет разные характеристики в функции.
Что такое миофибрилла?
Миофибрилла или мышечная фибрилла является основным стержнеобразным элементом мышечной клетки. В мышечной клетке сотни миофибрилл проходят параллельно друг другу. Миофибриллы состоят в основном из актиновых и миозиновых белков. Некоторые другие типы белков также присутствуют в миофибриллах. Эти белки организованы в толстые и тонкие длинные нити, называемые миофиламентов. Тонкие миофиламенты состоят в основном из актинового белка, в то время как толстые филаменты состоят из миозинового белка. Эти два типа миофиламентов проходят по длине миофибрилл в срезах, называемых саркомерами. Миофибриллы состоят из повторяющихся участков саркомеров. Эти саркомеры появляются под микроскопом в виде чередующихся темных и светлых полос и отвечают за сокращение мышц.
Рисунок 02: Структура миофибрилл
Каковы сходства между миофибриллой и мышечным волокном?
- Мышечные волокна и миофибриллы ответственны за мышечные сокращения.
- Оба типа имеют трубчатую форму.
- Оба типа располагаются параллельно внутри мышцы.
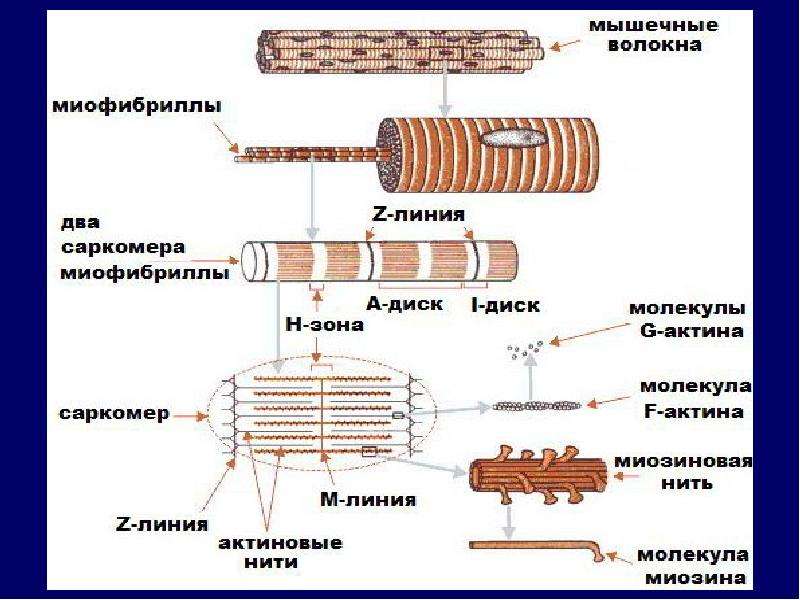
Какова связь между миофибриллой и мышечным волокном??
- Миофибриллы являются основными единицами мышечных волокон. Одно мышечное волокно содержит сотни миофибрилл.
В чем разница между миофибриллой и мышечным волокном?
Миофибрил против мышечного волокна | |
| Миофибрилла — базовая стержнеобразная единица мышечного волокна. | Мышечное волокно представляет собой трубчатую клетку мышц. |
| Сочинение | |
| Миофибрилла состоит из двух типов миофиламентов, называемых тонкими и толстыми нитями. | Мышечное волокно состоит из множества миофибрилл. |
| Природа | |
| Миофибрилла — это цилиндрическая органелла. | Мышечное волокно — это клетка с ядром и другими органеллами, включая митохондрии.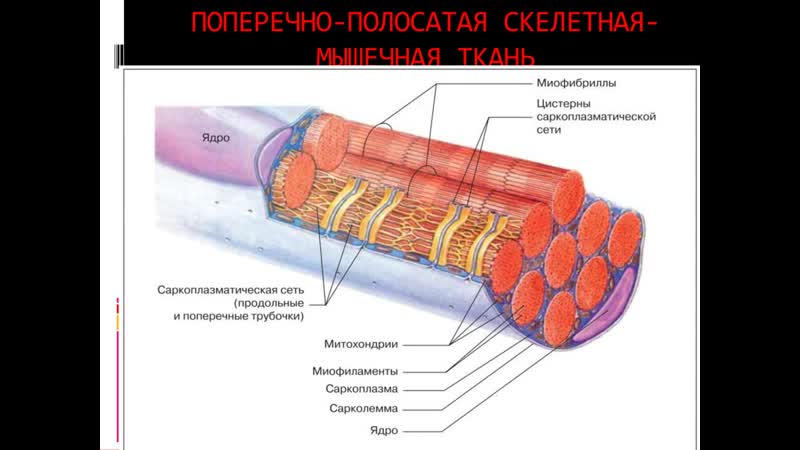 |
Резюме — Миофибрилла против мышечного волокна
Мышечное волокно является основной единицей мышц. Они расположены в пучках внутри мышц. Мышечное волокно состоит из множества миофибрилл. Миофибриллы являются основными единицами мышечных волокон. Они состоят из тонких и толстых белковых нитей, называемых миофиламентами. Основными белками в миофиламентах являются актин и миозин. Миофиламенты состоят из повторяющихся секций, называемых саркомерами. Их саркомеры отвечают за сокращение мышц. Мышечные волокна и миофибриллы в совокупности помогают сокращению мышц.
Скачать PDF версию Миофибрилла против мышечного волокна
Вы можете скачать PDF версию этой статьи и использовать ее в автономном режиме, как указано в примечаниях. Пожалуйста, загрузите PDF версию здесь. Разница между миофибриллой и мышечным волокном.
Ссылки:
«Миофибриллы.» Wikipedia. Фонд Викимедиа, 07 июля 2017 года. Интернет. Доступна здесь. 08 июля 2017.
«Структура мышц». Структура мышц (и связанных с ними соединительных тканей). Н.п., н.д. Web. Доступна здесь. 08 июля 2017.
Структура мышц (и связанных с ними соединительных тканей). Н.п., н.д. Web. Доступна здесь. 08 июля 2017.
Изображение предоставлено:
1. «1022 мышечных волокон (маленькие)» по OpenStax — (CC BY 4.0) через Commons Wikimedia
2. «Миофибрилла» от Slashme из Википедии на английском языке (CC BY-SA 3.0) через Викисклад Commons
Миофибриллы — обзор | ScienceDirect Topics
Миофибриллярная и саркомерная структура
В миоцитах миофибриллы представляют собой наиболее распространенную органеллу, занимающую примерно 50–60% цитоплазмы. Миофибриллы состоят из перекрывающихся толстых и тонких миофиламентов, организованных в отдельные повторяющиеся единицы, называемые саркомерами. Z-диски (также называемые Z-линиями; Z на рис. 5.1, 5.2), состоящие в основном из α-актинина, образуют границы саркомеров и служат местом прикрепления тонких филаментов.Высокомощные ЭМ поперечного сечения Z-дисков выявляют решетчатые узоры, описанные как маленькие квадраты и плетение корзины (4).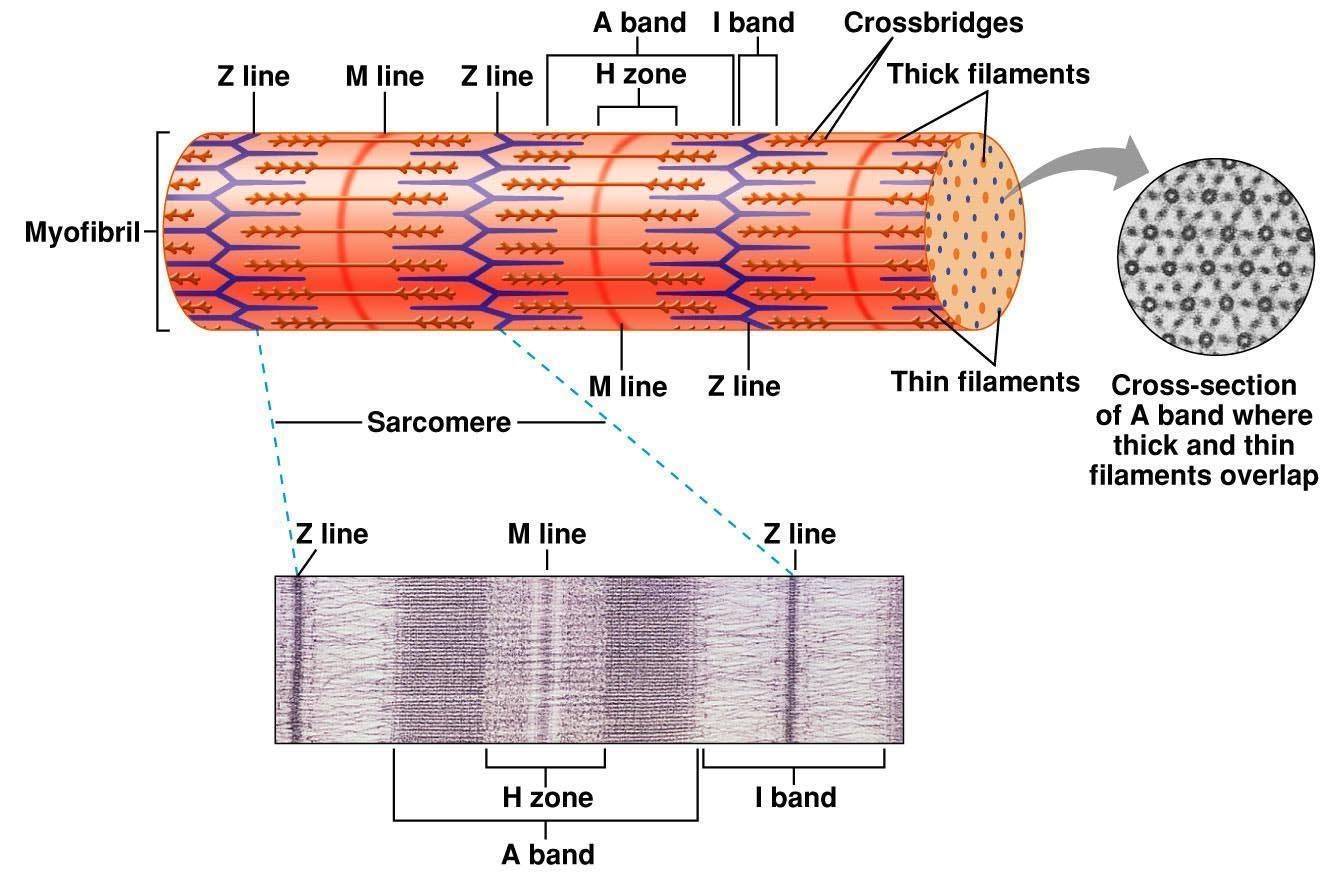 Хотя эти структуры выглядят как линии на двухмерных микрофотографиях, термин «диск» лучше описывает их трехмерную архитектуру. Во время сердечного цикла длина саркомера изменяется от ~ 2,2 мкм в конце диастолы до ~ 1,5 мкм в конце систолы в результате скольжения толстых и тонких филаментов относительно друг друга. Сокращение миоцитов приводит к изменению рисунка исчерченности в зависимости от степени перекрытия между толстыми и тонкими филаментами.
Хотя эти структуры выглядят как линии на двухмерных микрофотографиях, термин «диск» лучше описывает их трехмерную архитектуру. Во время сердечного цикла длина саркомера изменяется от ~ 2,2 мкм в конце диастолы до ~ 1,5 мкм в конце систолы в результате скольжения толстых и тонких филаментов относительно друг друга. Сокращение миоцитов приводит к изменению рисунка исчерченности в зависимости от степени перекрытия между толстыми и тонкими филаментами.
Тонкие филаменты (видимые с помощью ПЭМ, * на рис. 5.4c), ~приблизительно 1 мкм в длину и 8 нм в ширину, вставляются в Z-диски на концах каждого саркомера и поляризованы в противоположных направлениях на каждом стороне Z-диска. Они содержат в основном актин, который взаимодействует с миозином толстого филамента во время сокращения. Тонкие филаменты также содержат регуляторные белки тропонин и тропомиозин. Толстые филаменты, расположенные в середине саркомера, имеют длину примерно 1.5 мкм в длину и 15 нм в ширину. Толстые филаменты прикреплены к Z-дискам титиновыми филаментами. М-линии (одиночная стрелка на рис. 5.2; М на рис. 5.4в), расположенные в середине толстых филаментов (также в середине саркомера), содержат креатинкиназу, миомезин и М-белок. Поперечные мостики в области М-линии соединяют соседние толстые филаменты. Титин, крупнейший известный белок, проходит от М-линии вдоль толстого филамента, затем к Z-диску, но с трудом визуализируется с помощью ЭМ. Титин имеет область с двунаправленным пружинным действием, ответственным за пассивное натяжение выше длины провисания (т.г. в покое, длина ненагруженного саркомера ~ 1,90 мкм у большинства млекопитающих) и восстановление длины ниже провисшей длины (5). Здесь стоит отметить, что мы постоянно наблюдали длину ненагруженного саркомера в покое ~ 2,0 мкм в изолированных миоцитах человека (6).
М-линии (одиночная стрелка на рис. 5.2; М на рис. 5.4в), расположенные в середине толстых филаментов (также в середине саркомера), содержат креатинкиназу, миомезин и М-белок. Поперечные мостики в области М-линии соединяют соседние толстые филаменты. Титин, крупнейший известный белок, проходит от М-линии вдоль толстого филамента, затем к Z-диску, но с трудом визуализируется с помощью ЭМ. Титин имеет область с двунаправленным пружинным действием, ответственным за пассивное натяжение выше длины провисания (т.г. в покое, длина ненагруженного саркомера ~ 1,90 мкм у большинства млекопитающих) и восстановление длины ниже провисшей длины (5). Здесь стоит отметить, что мы постоянно наблюдали длину ненагруженного саркомера в покое ~ 2,0 мкм в изолированных миоцитах человека (6).
Саркомер разделен на A (длинная полоса на рис. 5.1), I (короткая полоса на рис. 5.1), H- и Z-диски (Z на рис. 5.1, 5.2). Более темная А-полоса сохраняет постоянную длину во время сокращения в первую очередь из-за латеральной регистрации толстых филаментов, но также содержит различное количество перекрывающихся тонких филаментов в зависимости от фазы сокращения. Как указано выше, Z-диски отмечают границу саркомера и сохраняют постоянную ширину во время сокращения. Более легкая I-полоса состоит только из тонких филаментов и уменьшается в ширину при сокращении. H-полоса занимает центральную часть A-полосы и содержит только толстые филаменты без перекрытия тонких филаментов. В скелетных мышцах хорошо видны изменения ширины Н-диапазона во время разных фаз сокращения саркомера. Ширина Н-диапазона увеличивается во время релаксации по мере того, как тонкие филаменты соскальзывают от центра саркомера.Однако изменения ширины Н-диапазона плохо видны на электронных микрофотографиях сердечной мышцы. Причина не ясна, но считается, что она связана с изменением длины тонкой нити. Бледные линии по обе стороны от М-линии в сердечной мышце известны как псевдо-H-полосы (двойная стрелка на рис. 5.2) и не меняют свою ширину во время расслабления. Псевдо-H-полосы представляют собой область толстого филамента, которая содержит только миозиновые хвосты и лишена миозиновых головок, которые взаимодействуют с тонкими филаментами.
Как указано выше, Z-диски отмечают границу саркомера и сохраняют постоянную ширину во время сокращения. Более легкая I-полоса состоит только из тонких филаментов и уменьшается в ширину при сокращении. H-полоса занимает центральную часть A-полосы и содержит только толстые филаменты без перекрытия тонких филаментов. В скелетных мышцах хорошо видны изменения ширины Н-диапазона во время разных фаз сокращения саркомера. Ширина Н-диапазона увеличивается во время релаксации по мере того, как тонкие филаменты соскальзывают от центра саркомера.Однако изменения ширины Н-диапазона плохо видны на электронных микрофотографиях сердечной мышцы. Причина не ясна, но считается, что она связана с изменением длины тонкой нити. Бледные линии по обе стороны от М-линии в сердечной мышце известны как псевдо-H-полосы (двойная стрелка на рис. 5.2) и не меняют свою ширину во время расслабления. Псевдо-H-полосы представляют собой область толстого филамента, которая содержит только миозиновые хвосты и лишена миозиновых головок, которые взаимодействуют с тонкими филаментами. В поперечных срезах ПЭМ (рис. 5.4а) в различных областях саркомера наблюдаются толстые и/или тонкие филаменты. Поперечные мостики М-линии соединяют толстые филаменты в центре саркомера (не показаны).
В поперечных срезах ПЭМ (рис. 5.4а) в различных областях саркомера наблюдаются толстые и/или тонкие филаменты. Поперечные мостики М-линии соединяют толстые филаменты в центре саркомера (не показаны).
Миофибриллы — обзор | ScienceDirect Topics
6.3.2 Структура мышц и водоудерживающая способность
Миофибриллы составляют большую часть мышечных клеток. Эти органеллы составляют более 80% объема мышечной клетки. До 85% воды в мышечной клетке содержится в миофибриллах.Большая часть этой воды удерживается капиллярными силами, возникающими из-за расположения толстых и тонких нитей внутри миофибриллы. Было показано, что в живых мышцах саркомеры остаются изоволюметрическими во время сокращения и расслабления (Millman et al., 1981, 1983). Это указывает на то, что в физиологических условиях количество воды, удерживаемой в волокнистой структуре мышечной клетки, не обязательно должно изменяться. Однако на расположение этой воды могут влиять изменения объема по мере окоченения мышц.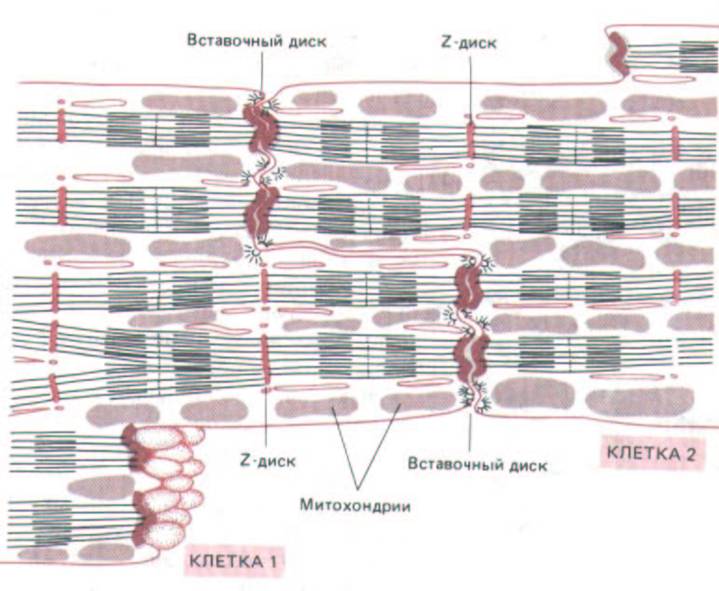 По мере ригидности мышц между толстыми и тонкими филаментами образуются поперечные мостики, тем самым уменьшая доступное пространство для пребывания воды (Offer and Trinick, 1983). Показано, что при снижении рН мышц свиньи от физиологических значений до 5,2–5,5 (вблизи изоэлектрической точки миозина) расстояние между толстыми филаментами уменьшается в среднем на 2,5 нм (Diesbourg et al., 1988). ). Это может вытеснить саркоплазматическую жидкость между миофиламентами в экстрамиофибриллярное пространство.Фактически, из интрамиофибриллярного пространства может быть потеряно достаточно жидкости, чтобы увеличить экстрамиофибриллярный объем в 1,6 раза больше, чем его объем до ригидности (Bendall and Swatland, 1988).
По мере ригидности мышц между толстыми и тонкими филаментами образуются поперечные мостики, тем самым уменьшая доступное пространство для пребывания воды (Offer and Trinick, 1983). Показано, что при снижении рН мышц свиньи от физиологических значений до 5,2–5,5 (вблизи изоэлектрической точки миозина) расстояние между толстыми филаментами уменьшается в среднем на 2,5 нм (Diesbourg et al., 1988). ). Это может вытеснить саркоплазматическую жидкость между миофиламентами в экстрамиофибриллярное пространство.Фактически, из интрамиофибриллярного пространства может быть потеряно достаточно жидкости, чтобы увеличить экстрамиофибриллярный объем в 1,6 раза больше, чем его объем до ригидности (Bendall and Swatland, 1988).
Во время развития окоченения общий диаметр мышечных клеток уменьшается (Hegarty, 1970; Swatland and Belfry, 1985) и, вероятно, является результатом передачи латерального сокращения миофибрилл на всю клетку (Diesbourg et al. , 1988). Кроме того, во время развития окоченения саркомеры могут укорачиваться; это также еще больше уменьшает пространство, доступное для воды внутри миофибриллы. Фактически было показано, что потери сока увеличиваются линейно с уменьшением длины саркомеров в мышечных клетках (Honikel et al., 1986). Совсем недавно высокочувствительные низкопольные исследования ядерного магнитного резонанса (ЯМР) использовались для более полного понимания взаимосвязи между структурой мышечных клеток и распределением воды (Straadt et al., 2007; Bertram et al., ). 2002; Бертрам и Андерсен, 2007). Эти исследования показывают, что внутри миофибриллы пропорционально меньше воды удерживается в А-полосе с высокой плотностью белка, чем в менее плотной I-полосе.Это наблюдение может помочь объяснить, почему более короткие саркомеры (особенно в мышцах с холодовым укорочением) часто связаны с повышенной потерей жидкости. По мере того как миофибрилла укорачивается и наступает окоченение, укорочение саркомера приводит к укорочению и последующему снижению объема области I-диапазона в миофибрилле. Потеря объема в этой миофибриллярной области (где может находиться много воды) в сочетании с рН-индуцированным латеральным сокращением миофибрилл приводит к вытеснению воды из миофибриллярной структуры в экстрамиофибриллярные пространства внутри мышечной клетки (Bendall and Swatland, 1988).
Фактически было показано, что потери сока увеличиваются линейно с уменьшением длины саркомеров в мышечных клетках (Honikel et al., 1986). Совсем недавно высокочувствительные низкопольные исследования ядерного магнитного резонанса (ЯМР) использовались для более полного понимания взаимосвязи между структурой мышечных клеток и распределением воды (Straadt et al., 2007; Bertram et al., ). 2002; Бертрам и Андерсен, 2007). Эти исследования показывают, что внутри миофибриллы пропорционально меньше воды удерживается в А-полосе с высокой плотностью белка, чем в менее плотной I-полосе.Это наблюдение может помочь объяснить, почему более короткие саркомеры (особенно в мышцах с холодовым укорочением) часто связаны с повышенной потерей жидкости. По мере того как миофибрилла укорачивается и наступает окоченение, укорочение саркомера приводит к укорочению и последующему снижению объема области I-диапазона в миофибрилле. Потеря объема в этой миофибриллярной области (где может находиться много воды) в сочетании с рН-индуцированным латеральным сокращением миофибрилл приводит к вытеснению воды из миофибриллярной структуры в экстрамиофибриллярные пространства внутри мышечной клетки (Bendall and Swatland, 1988). ).Таким образом, вполне вероятно, что постепенная мобилизация воды изнутри миофибриллы в другие пространства внутри клетки может быть ключевой в обеспечении значительного источника капель.
).Таким образом, вполне вероятно, что постепенная мобилизация воды изнутри миофибриллы в другие пространства внутри клетки может быть ключевой в обеспечении значительного источника капель.
Важно отметить, что само по себе сморщивание миофибрилл не может быть причиной перемещения жидкости во внеклеточное пространство и, в конечном счете, из мышцы. Миофибриллы связаны друг с другом и с клеточной мембраной белковыми связями (Wang and Ramirez-Mitchell, 1983). Эти связи, если они сохраняются интактными в посмертной мышце, передают уменьшение диаметра миофибрилл на мышечную клетку (Kristensen and Purslow, 2001; Melody et al., 2004; Morrison и др., , 1998; Diesbourg и др., , 1988). Таким образом, усадка миофибрилл может привести к уменьшению общего диаметра мышечной клетки, создавая каналы между клетками и между пучками клеток, которые могут вытекать из продукта воронкой (Offer and Knight, 1988). Внеклеточное пространство вокруг мышечных волокон постоянно увеличивается вплоть до 24 часов после смерти, но промежутки между пучками мышечных волокон немного уменьшаются между 9 и 24 часами после смерти, возможно, из-за оттока жидкости из этих основных каналов (Schafer et al. , 2002). Эти связи между соседними миофибриллами и между миофибриллами и клеточной мембраной состоят из нескольких белков, которые связаны со структурами промежуточных филаментов и структурами, известными как костамеры. Костамеры обеспечивают структурный каркас, отвечающий за прикрепление миофибрилл к сарколемме. Белки, которые составляют или связаны с промежуточными филаментами и костамерами, включают (среди др.) десмин, филамин и синемин, дистрофин, талин и винкулин (Greaser, 1991).Если костамеры и костамерные связи остаются неповрежденными во время превращения мышц в мясо, сокращение миофибрилл по мере того, как мышцы становятся ригорными, будет передаваться на всю клетку через эти белковые связи и в конечном итоге приведет к уменьшению объема самой мышечной клетки (Melody ). et al., 2004; Offer and Knight, 1988b; Kristensen and Purslow, 2001). Таким образом, процесс ригора может приводить к мобилизации воды не только из миофибрилл, но и из экстрамиофибриллярных пространств по мере сужения общего объема клетки.
, 2002). Эти связи между соседними миофибриллами и между миофибриллами и клеточной мембраной состоят из нескольких белков, которые связаны со структурами промежуточных филаментов и структурами, известными как костамеры. Костамеры обеспечивают структурный каркас, отвечающий за прикрепление миофибрилл к сарколемме. Белки, которые составляют или связаны с промежуточными филаментами и костамерами, включают (среди др.) десмин, филамин и синемин, дистрофин, талин и винкулин (Greaser, 1991).Если костамеры и костамерные связи остаются неповрежденными во время превращения мышц в мясо, сокращение миофибрилл по мере того, как мышцы становятся ригорными, будет передаваться на всю клетку через эти белковые связи и в конечном итоге приведет к уменьшению объема самой мышечной клетки (Melody ). et al., 2004; Offer and Knight, 1988b; Kristensen and Purslow, 2001). Таким образом, процесс ригора может приводить к мобилизации воды не только из миофибрилл, но и из экстрамиофибриллярных пространств по мере сужения общего объема клетки. Фактически уменьшение диаметра мышечных клеток наблюдалось в посмертных мышцах (Offer and Cousins, 1992). Эта вода, которая удаляется из миофибрилл и, в конечном счете, из мышечной клетки, в конечном итоге собирается во внеклеточном пространстве. Несколько исследований показали, что между мышечными клетками и между мышечными пучками в период после окоченения образуются промежутки (Offer and Cousins, 1992; Offer et al., 1989). Эти промежутки между мышечными пучками являются основными каналами, по которым из мяса может вытекать очищение; некоторые исследователи фактически назвали их «капельными каналами».
Фактически уменьшение диаметра мышечных клеток наблюдалось в посмертных мышцах (Offer and Cousins, 1992). Эта вода, которая удаляется из миофибрилл и, в конечном счете, из мышечной клетки, в конечном итоге собирается во внеклеточном пространстве. Несколько исследований показали, что между мышечными клетками и между мышечными пучками в период после окоченения образуются промежутки (Offer and Cousins, 1992; Offer et al., 1989). Эти промежутки между мышечными пучками являются основными каналами, по которым из мяса может вытекать очищение; некоторые исследователи фактически назвали их «капельными каналами».
Недавние исследования показали, что снижает деградацию белков, которые связывают миофибриллы с сарколеммой (таких как десмин), что приводит к усиленному сокращению мышечной клетки, что в конечном итоге приводит к потере влаги (Morrison et al ., 1998; Huff-Lonergan and Lonergan, 2005; Bee et al., 2007, 2004). Если эти миофибриллярные связи не разрушаются относительно рано в посмертном периоде, остается меньше места для воды, вытесненной из миофибрилл во время развития окоченения, внутри клеток.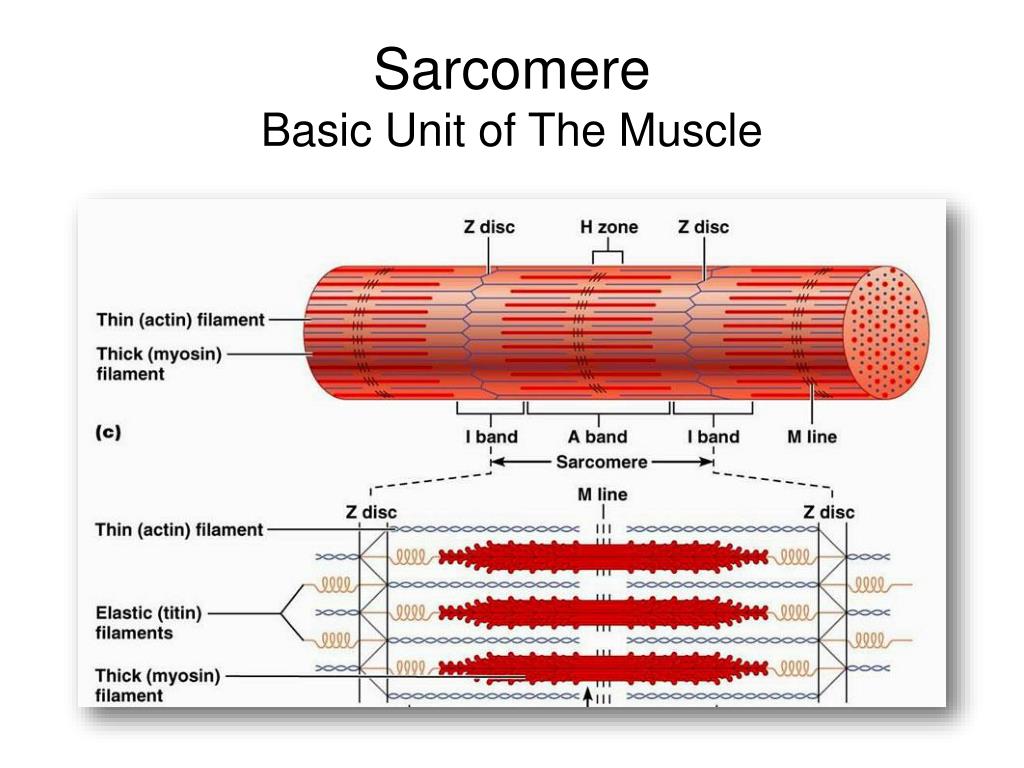 Деградация белка промежуточных филаментов, десмина, может происходить уже через 45 минут после обескровливания в некоторых мышцах с более высокой водоудерживающей способностью (Melody et al., 2004). Этот временной интервал идеально подходит для удержания жидкости, вытесненной из миофибриллы. Несколько исследований показали, что существует значительная корреляция между разложением десмина и водоудерживающей способностью свинины (Bee et al., 2007, 2004; Gardner et al., 2005, 2006; Melody et al., 2004; Чжан и др., 2006). Деградация некоторых костамерных белков также может играть роль в поддержании жидкости внутри клетки (Kristensen and Purslow, 2001; Zhang et al., 2006). Костамерные белки помогают связывать белки промежуточных филаментов с саркомером; таким образом, их деградация также может позволить устранить некоторые структурные ограничения. Было показано, что костамерные белки винкулин и талин в большей степени разлагаются в свинине, которая обладает более высокой водоудерживающей способностью.
Деградация белка промежуточных филаментов, десмина, может происходить уже через 45 минут после обескровливания в некоторых мышцах с более высокой водоудерживающей способностью (Melody et al., 2004). Этот временной интервал идеально подходит для удержания жидкости, вытесненной из миофибриллы. Несколько исследований показали, что существует значительная корреляция между разложением десмина и водоудерживающей способностью свинины (Bee et al., 2007, 2004; Gardner et al., 2005, 2006; Melody et al., 2004; Чжан и др., 2006). Деградация некоторых костамерных белков также может играть роль в поддержании жидкости внутри клетки (Kristensen and Purslow, 2001; Zhang et al., 2006). Костамерные белки помогают связывать белки промежуточных филаментов с саркомером; таким образом, их деградация также может позволить устранить некоторые структурные ограничения. Было показано, что костамерные белки винкулин и талин в большей степени разлагаются в свинине, которая обладает более высокой водоудерживающей способностью.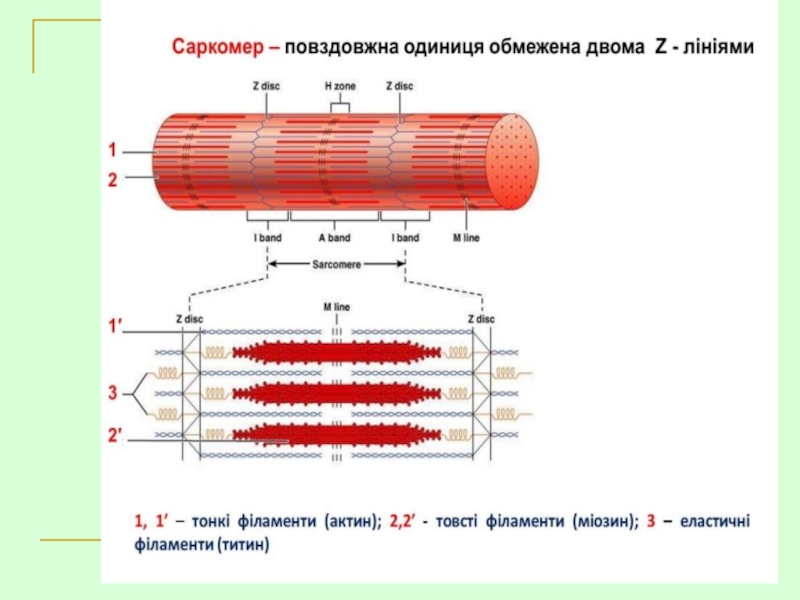 Таким образом, представляется, что деградация межмиофибриллярных и костамерных связей относительно скоро после обескровливания действительно может улучшить водоудерживающую способность (Bee et al., 2007, 2004; Gardner et al., 2005; Melody). и др., 2004; Хафф-Лонерган и Лонерган, 2005). Это может происходить за счет ограничения степени, в которой латеральное сокращение миофибрилл передается на общую мышечную клетку, что может позволить, по крайней мере, в некоторой степени, внутриклеточное пространство быть в некоторой степени независимым от объема или сокращения миофибрилл (Хафф-Лонерган и Лонерган, 2005).
Таким образом, представляется, что деградация межмиофибриллярных и костамерных связей относительно скоро после обескровливания действительно может улучшить водоудерживающую способность (Bee et al., 2007, 2004; Gardner et al., 2005; Melody). и др., 2004; Хафф-Лонерган и Лонерган, 2005). Это может происходить за счет ограничения степени, в которой латеральное сокращение миофибрилл передается на общую мышечную клетку, что может позволить, по крайней мере, в некоторой степени, внутриклеточное пространство быть в некоторой степени независимым от объема или сокращения миофибрилл (Хафф-Лонерган и Лонерган, 2005).
Важно отметить, что общее расщепление мышечных белков не обязательно положительно влияет на влагоудерживающую способность свежего мяса. Например, расщепление белка интегрина может не способствовать водоудерживающей способности (Lawson, 2004; Zhang et al., 2006). Интегрины представляют собой семейство рецепторов адгезии, которые опосредуют взаимодействия между внеклеточным матриксом и клеточным цитоскелетом.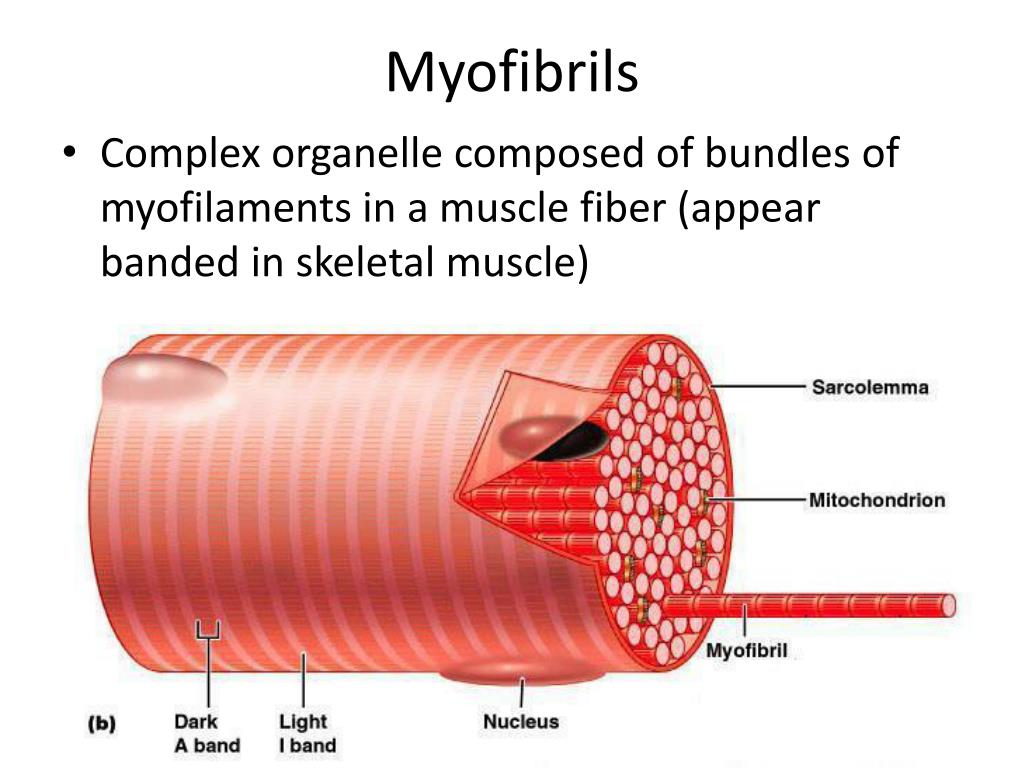 Было показано, что деградация интегрина в свинине связана с повышенной потерей воды в свежей свинине (Zhang et al., 2006). Также было показано, что деградация интегрина связана с образованием капельных каналов в свинине (Lawson, 2004). Таким образом, образование капель, вероятно, является результатом специфических структурных изменений в мышечной клетке. Таким образом, возможно, что деградация белков промежуточных филаментов, таких как десмин, и ассоциированных с мембраной белков, таких как интегрин, может играть очень разные роли в опосредовании водоудерживающей способности свинины. Например, деградация десмина на ранней стадии после смерти может снять ограничения внутри клетки, которые ограничивают пространство, доступное для воды/жидкости, которая была вытеснена из миофибриллы во время окоченения.И наоборот, деградация белка, такого как интегрин, может фактически способствовать образованию капельных каналов и, таким образом, может фактически улучшить способность влаги «уходить» из мышечной клетки (Lawson, 2004; Zhang et al.
Было показано, что деградация интегрина в свинине связана с повышенной потерей воды в свежей свинине (Zhang et al., 2006). Также было показано, что деградация интегрина связана с образованием капельных каналов в свинине (Lawson, 2004). Таким образом, образование капель, вероятно, является результатом специфических структурных изменений в мышечной клетке. Таким образом, возможно, что деградация белков промежуточных филаментов, таких как десмин, и ассоциированных с мембраной белков, таких как интегрин, может играть очень разные роли в опосредовании водоудерживающей способности свинины. Например, деградация десмина на ранней стадии после смерти может снять ограничения внутри клетки, которые ограничивают пространство, доступное для воды/жидкости, которая была вытеснена из миофибриллы во время окоченения.И наоборот, деградация белка, такого как интегрин, может фактически способствовать образованию капельных каналов и, таким образом, может фактически улучшить способность влаги «уходить» из мышечной клетки (Lawson, 2004; Zhang et al.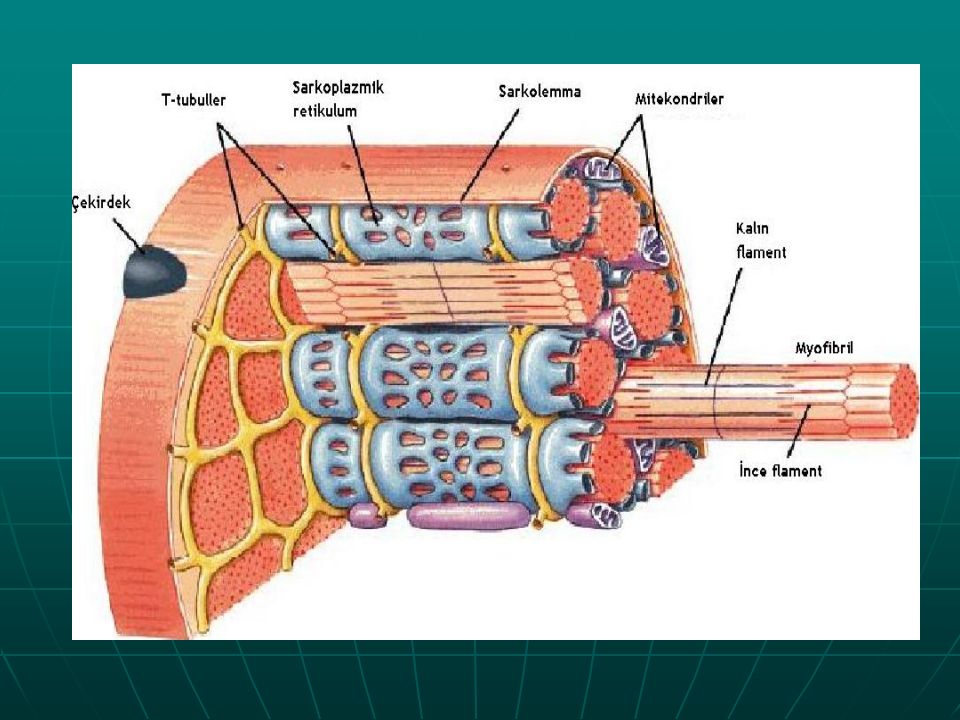 , 2006).
, 2006).
Введение
Мышцы представляют собой многоклеточные сократительные единицы. Они делятся на три типа:
- скелетные мышцы
- гладкая мышца
- сердечная мышца
Читая о каждом типе мышц, подумайте о сходствах и различиях между ними с точки зрения структуры и функций.
Скелетная мышца
Скелетная мышца в основном отвечает за движение скелета, но также находится в таких органах, как глазное яблоко и язык. Это произвольная мышца, поэтому она находится под сознательным контролем. Скелетные мышцы специализируются на быстром и сильном кратковременном сокращении.
Каждая мышечная клетка определяется клеточной мембраной (сарколеммой) и содержит множество ядер по всей своей длине. Ядра смещены периферически в пределах поперечного сечения цитоплазмы (саркоплазмы), в то время как большое количество продольных миофибрилл, групп расположенных сократительных белков, занимает большую часть центрального пространства.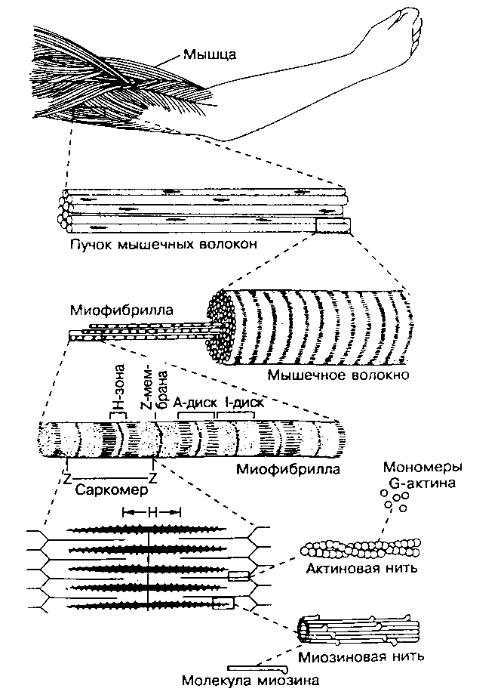 Миофибрилла содержит несколько важных гистологических ориентиров:
Миофибрилла содержит несколько важных гистологических ориентиров:
- Миофибрилла состоит из чередующихся тяжей. I-полосы (изотропные в поляризованном свете) кажутся светлыми, а А-полосы (анизотропные в поляризованном свете) кажутся темными. Чередование этих полос приводит к поперечно-полосатому виду скелетных мышц.
- Z-линии (Zwischenschieben) делят I-диапазоны пополам.
- Светлая полоса, называемая Н-диапазоном (Хеллер), находится внутри каждого А-диапазона.
- M-линия (Mittelschiebe) делит пополам каждый A-диапазон (и при этом делит пополам каждый H-диапазон).
Каждую миофибриллу можно рассматривать как серию сократительных единиц, называемых саркомерами, которые содержат два типа филаментов: толстые филаменты, состоящие из миозина, и тонкие филаменты, состоящие из актина. Отдельные филаменты не изменяют своей длины при мышечном сокращении; скорее тонкие нити скользят по толстым нитям, укорачивая саркомер. Природа этих нитей может быть понята в контексте гистологических ориентиров миофибриллы.
- Толстые филаменты представляют собой биполярный массив полимеризованных миозиновых моторов. Двигатели на одной стороне нити ориентированы в одном направлении, тогда как двигатели на другой стороне нити ориентированы в противоположном направлении. В центре нити отсутствуют двигатели; он содержит только спиральную спиральную область миозинов. Набор белков сшивает каждую миозиновую нить с соседними в центре нити. Эти белки составляют М-линию.
- Тонкие нити прикреплены к дискообразной зоне, которая гистологически выглядит как Z-линия.Z-линии содержат белки, которые связывают и стабилизируют плюс-концы актиновых филаментов. Z-линии также определяют границы каждого саркомера.
- I- и H-полосы представляют собой области, в которых толстые и тонкие филаменты не перекрываются (поэтому эти полосы кажутся более бледными под микроскопом). I-полоса содержит исключительно тонкие нити, тогда как H-полоса содержит исключительно толстые нити.
Скелетные мышцы делятся на два типа мышечных волокон:
- Медленно сокращающиеся (тип I) мышечные волокна сокращаются медленнее и зависят от аэробного метаболизма. Они содержат большое количество митохондрий и миоглобина, молекулы для хранения кислорода. Красноватый цвет миоглобина является причиной того, что эти волокна можно назвать красными волокнами. Эти мышцы могут поддерживать непрерывное сокращение и полезны в таких видах деятельности, как поддержание осанки.
- Быстросокращающиеся мышечные волокна (тип II) сокращаются быстрее из-за присутствия более быстрого миозина. Волокна типа II можно разделить на те, которые имеют большое количество митохондрий и миоглобина, и те, которые имеют мало митохондрий и мало миоглобина.Первые в основном используют аэробное дыхание для выработки энергии, тогда как вторые полагаются на гликолиз. Отсутствие миоглобина приводит к более бледному цвету, чем у медленно сокращающихся мышц, поэтому быстросокращающиеся волокна можно назвать белыми волокнами. Эти мышцы важны для интенсивных, но спорадических сокращений; например, те, которые имеют место в бицепсе.
 Они содержат большое количество митохондрий и миоглобина, молекулы для хранения кислорода. Красноватый цвет миоглобина является причиной того, что эти волокна можно назвать красными волокнами. Эти мышцы могут поддерживать непрерывное сокращение и полезны в таких видах деятельности, как поддержание осанки.
Они содержат большое количество митохондрий и миоглобина, молекулы для хранения кислорода. Красноватый цвет миоглобина является причиной того, что эти волокна можно назвать красными волокнами. Эти мышцы могут поддерживать непрерывное сокращение и полезны в таких видах деятельности, как поддержание осанки. Большинство мышц содержат смесь этих экстремальных типов волокон. У людей типы волокон невозможно различить на основе общего осмотра, но для различения волокон требуются специальные пятна или обработка.
У людей типы волокон невозможно различить на основе общего осмотра, но для различения волокон требуются специальные пятна или обработка.
Нервно-мышечное соединение и активация клеток скелетных мышц
Клетки скелетных мышц иннервируются двигательными нейронами. Двигательная единица определяется как нейрон и волокна, которые он иннервирует. Некоторые мотонейроны иннервируют одну или несколько мышечных клеток, тогда как другие мотонейроны могут иннервировать сотни мышечных клеток. Мышцы, требующие точного контроля, имеют моторные нейроны, иннервирующие меньше мышечных клеток; мышцы, участвующие в менее контролируемых движениях, могут иметь множество волокон, иннервируемых каждым нейроном.Моторные аксоны заканчиваются нервно-мышечным соединением на поверхности волокон скелетных мышц. Нервно-мышечное соединение состоит из пресинаптического нервного окончания и постсинаптического мышечного волокна. При деполяризации пресинаптические пузырьки, содержащие нейротрансмиттер ацетилхолин, сливаются с мембраной, высвобождая ацетилхолин в щель. Ацетилхолин связывается с рецепторами на постсинаптической мембране и вызывает деполяризацию мышечного волокна, что приводит к его сокращению. Как правило, один потенциал действия в нейроне высвобождает достаточно нейротрансмиттера, чтобы вызвать одно сокращение мышечного волокна.
Ацетилхолин связывается с рецепторами на постсинаптической мембране и вызывает деполяризацию мышечного волокна, что приводит к его сокращению. Как правило, один потенциал действия в нейроне высвобождает достаточно нейротрансмиттера, чтобы вызвать одно сокращение мышечного волокна.
В мышечных клетках сарколемма или плазматическая мембрана простирается поперечно в саркоплазму, чтобы окружить каждую миофибриллу, образуя систему Т-трубочек. Эти Т-трубочки обеспечивают синхронное сокращение всех саркомеров миофибриллы. Т-трубочки находятся на стыке А- и I-полос, и их просветы переходят во внеклеточное пространство. В таких соединениях Т-трубочки находятся в тесном контакте с саркоплазматическим ретикулумом, который образует сеть, окружающую каждую миофибриллу.Часть саркоплазматического ретикулума, связанная с Т-трубочками, называется терминальной цистерной из-за ее уплощенного цистернального расположения. Когда сигнал возбуждения поступает в нервно-мышечное соединение, деполяризация сарколеммы быстро проходит через систему Т-трубочек и вступает в контакт с саркоплазматическим ретикулумом, вызывая высвобождение кальция и приводя к мышечному сокращению.
Гладкая мускулатура
Гладкая мускулатура образует сократительную часть стенки пищеварительного тракта от средней части пищевода до внутреннего сфинктера заднего прохода.Он обнаруживается в стенках протоков желез, связанных с пищеварительным трактом, в стенках дыхательных путей от трахеи до альвеолярных ходов, в мочевыводящих и половых путях. Стенки артерий, вен и крупных лимфатических сосудов также содержат гладкие мышцы.
Гладкая мускулатура предназначена для медленных и продолжительных сокращений с небольшой силой. Вместо того, чтобы иметь двигательные единицы, все клетки в целой массе гладкой мускулатуры сокращаются вместе. Гладкой мышце присуща способность к сокращению, и вегетативная нервная система, гормоны и местные метаболиты могут влиять на ее сокращение.Поскольку гладкие мышцы не находятся под сознательным контролем, они являются непроизвольными мышцами.
Гладкомышечные волокна представляют собой удлиненные веретенообразные клетки с одним ядром.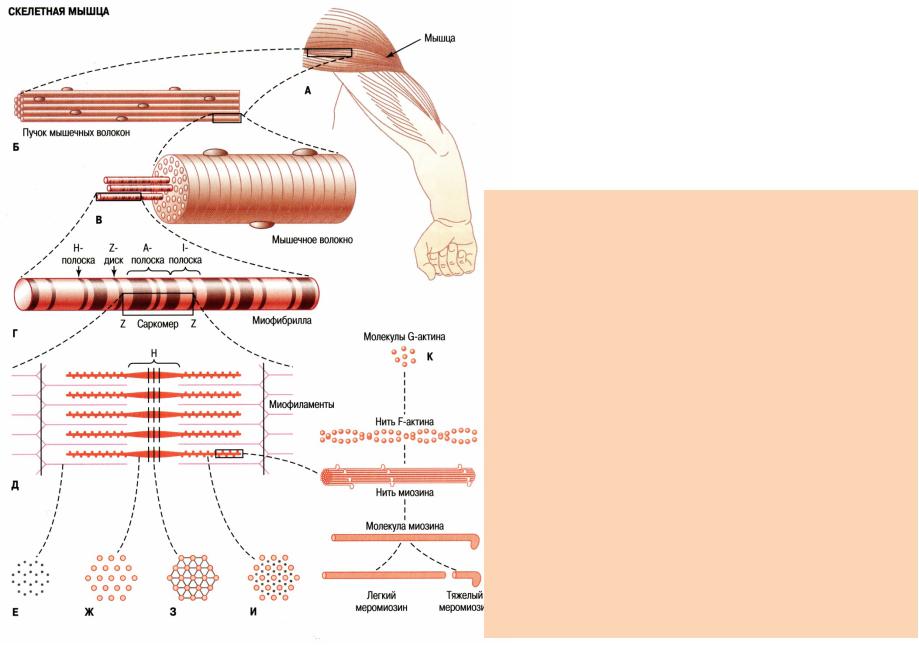 В целом они намного короче клеток скелетных мышц. Ядро расположено центрально, а саркоплазма заполнена фибриллами. Толстые (миозиновые) и тонкие (актиновые) филаменты разбросаны по всей саркоплазме и прикрепляются к адгезионным плотностям на клеточной мембране и к очаговым плотностям внутри цитоплазмы. Поскольку сократительные белки этих клеток не организованы в миофибриллы, как в скелетных и сердечных мышцах, они кажутся гладкими, а не исчерченными.
В целом они намного короче клеток скелетных мышц. Ядро расположено центрально, а саркоплазма заполнена фибриллами. Толстые (миозиновые) и тонкие (актиновые) филаменты разбросаны по всей саркоплазме и прикрепляются к адгезионным плотностям на клеточной мембране и к очаговым плотностям внутри цитоплазмы. Поскольку сократительные белки этих клеток не организованы в миофибриллы, как в скелетных и сердечных мышцах, они кажутся гладкими, а не исчерченными.
Гладкие мышечные волокна связаны друг с другом в нерегулярно разветвленные пучки, которые различаются по расположению от органа к органу. Эти пучки являются функциональными сократительными единицами. Между волокнами и пучками имеется также сеть поддерживающих коллагеновых тканей.
Сердечная мышца
Сердечная мышца имеет общие важные характеристики как со скелетными, так и с гладкими мышцами. Функционально сердечная мышца производит сильные сокращения, как и скелетная мышца. Тем не менее, он имеет встроенные механизмы, чтобы инициировать непрерывное сокращение, как гладкие мышцы. Скорость и сила сокращения не подлежат произвольному контролю, а зависят от вегетативной нервной системы и гормонов.
Скорость и сила сокращения не подлежат произвольному контролю, а зависят от вегетативной нервной системы и гормонов.
Гистологически сердечная мышца кажется поперечно-полосатой, как и скелетная мышца, из-за расположения сократительных белков. Он также имеет несколько уникальных структурных характеристик:
- Волокна сердечной мышцы расположены не просто параллельно. Вместо этого они разветвляются на концах, образуя соединения с несколькими соседними ячейками, что приводит к сложной трехмерной сети.
- Волокна сердечной мышцы представляют собой длинные цилиндрические клетки с одним или двумя ядрами. Ядра расположены центрально, как и у гладких мышц.
- Саркоплазма сердечной мышцы имеет большое количество митохондрий для удовлетворения потребности в энергии.
Коллагеновые ткани окружают отдельные волокна сердечной мышцы. В этой поддерживающей ткани имеется обильная васкуляризация, которая необходима для удовлетворения высоких метаболических потребностей сердечной мышцы.
Волокна сердечной мышцы соединены конец к концу с помощью специализированных областей соединения, называемых вставочными дисками.Вставочные диски обеспечивают закрепление миофибрилл и позволяют быстро распространять сократительные стимулы между клетками. Такое быстрое распространение сокращения позволяет сердечным мышцам действовать как функциональный синцитий. Вставочные диски содержат три типа межмембранного контакта:
- сращенные фасции, соединенные с актиновыми нитями для передачи сокращения
- десмосомы, соединяющиеся с промежуточными филаментами цитоскелета
- щелевые контакты, представляющие собой участки с низким электрическим сопротивлением, обеспечивающие распространение возбуждения
В дополнение к сократительным клеткам существует специализированная система, состоящая из модифицированных мышечных клеток, функция которых заключается в создании стимула для сердцебиения и проведении импульса к различным частям миокарда.Эта система состоит из синоатриального узла, атриовентрикулярного узла, пучка Гиса и волокон Пуркинье.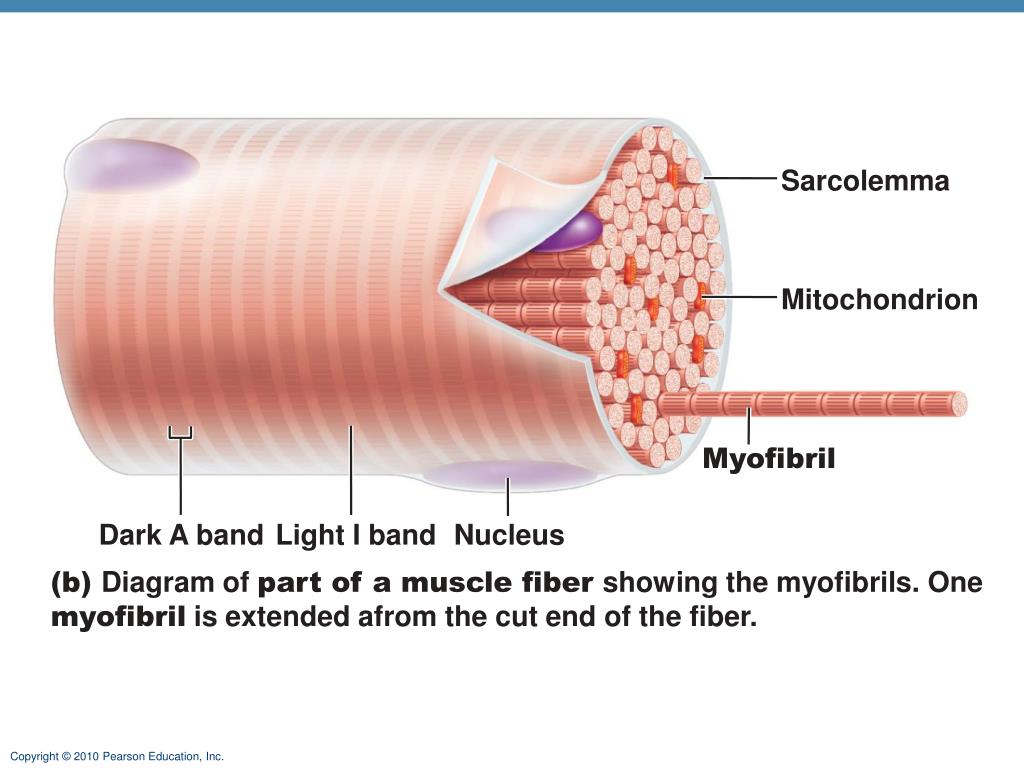
Увеличение мышечных клеток — ScienceDaily
Саркомеры представляют собой небольшие повторяющиеся субъединицы миофибрилл, длинные цилиндры, которые соединяются вместе, образуя мышечные волокна. Внутри саркомеров нити белков миозина и актина взаимодействуют, вызывая сокращение и расслабление мышц. До сих пор традиционные экспериментальные подходы к исследованию структуры и функции мышечной ткани проводились на реконструированных белковых комплексах или имели низкое разрешение.«Вместо этого электронная криотомография позволяет нам получать подробные трехмерные изображения замороженных мышц без артефактов», — говорит Раунсер.
Крио-ЭТ долгое время была устоявшейся, но нишевой методологией. Но недавние технические достижения в области электронной криомикроскопии (крио-ЭМ), а также новая разработка измельчения криосфокусированного ионного пучка (ФИП) подталкивают разрешение крио-ЭТ. Подобно крио-ЭМ, исследователи мгновенно замораживают биологический образец при очень низкой температуре (-175 °C). Благодаря этому процессу образец сохраняет свою гидратацию и тонкую структуру и остается близким к своему естественному состоянию.Затем применяется фрезерование FIB, чтобы сбрить лишний материал и получить идеальную толщину около 100 нанометров для просвечивающего электронного микроскопа, который получает несколько изображений, когда образец наклоняется вдоль оси. Наконец, вычислительные методы реконструируют трехмерное изображение с высоким разрешением.
Благодаря этому процессу образец сохраняет свою гидратацию и тонкую структуру и остается близким к своему естественному состоянию.Затем применяется фрезерование FIB, чтобы сбрить лишний материал и получить идеальную толщину около 100 нанометров для просвечивающего электронного микроскопа, который получает несколько изображений, когда образец наклоняется вдоль оси. Наконец, вычислительные методы реконструируют трехмерное изображение с высоким разрешением.
Команда Раунсера провела крио-ЭТ на миофибриллах мышей, выделенных в Королевском колледже, и получила разрешение в один нанометр (миллионную долю миллиметра, достаточно, чтобы увидеть тонкие структуры внутри белка): «Теперь мы можем рассмотреть миофибриллу в деталях. казалось невообразимым всего четыре года назад.Это захватывающе!», — говорит Раунсер.
Волокна в их естественном окружении
Расчетная реконструкция миофибрилл выявила трехмерную организацию саркомера, включая подобласти М-, А- и I-дисков, а также Z-диск, который неожиданно образует более неправильную сетку и принимает различные конформации.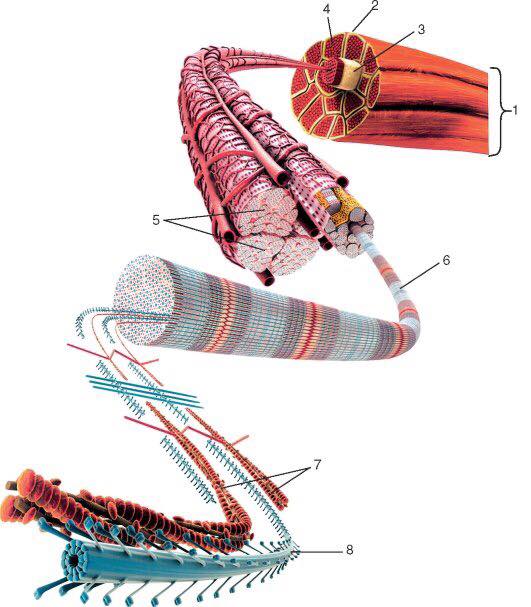 Ученые использовали образец с миозином, прочно связанным с актином, что представляет собой стадию сокращения мышц, называемую состоянием окоченения.И действительно, они впервые смогли визуализировать в нативной клетке, как две головки одного и того же миозина связываются с актиновым филаментом. Они также обнаружили, что двойная головка не только взаимодействует с одним и тем же актиновым филаментом, но также обнаруживается расщепленным между двумя актиновыми филаментами. Это никогда раньше не наблюдалось и показывает, что близость к следующей актиновой нити сильнее, чем кооперативный эффект между соседними головками.
Ученые использовали образец с миозином, прочно связанным с актином, что представляет собой стадию сокращения мышц, называемую состоянием окоченения.И действительно, они впервые смогли визуализировать в нативной клетке, как две головки одного и того же миозина связываются с актиновым филаментом. Они также обнаружили, что двойная головка не только взаимодействует с одним и тем же актиновым филаментом, но также обнаруживается расщепленным между двумя актиновыми филаментами. Это никогда раньше не наблюдалось и показывает, что близость к следующей актиновой нити сильнее, чем кооперативный эффект между соседними головками.
«Это только начало. Cryo-ET движется от нишевой к широко распространенной технологии в структурной биологии», — говорит Раунсер.«Скоро мы сможем исследовать мышечные заболевания на молекулярном и даже атомном уровне». Мышцы мыши очень похожи на человеческие, но ученые планируют исследовать мышечную ткань из биопсии или полученную из плюрипотентных стволовых клеток.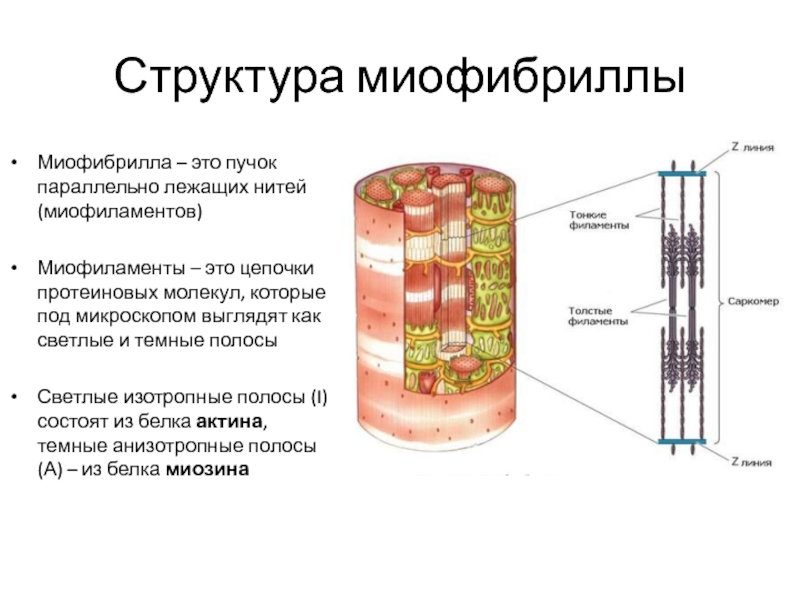
Источник истории:
Материалы предоставлены Институтом молекулярной физиологии Макса Планка . Примечание. Содержимое можно редактировать по стилю и длине.
38.4B: Волокна скелетных мышц — Биология LibreTexts
Скелетные мышцы состоят из поперечнополосатых субъединиц, называемых саркомерами, которые состоят из миофиламентов актина и миозина.
Цели обучения
- Описать структуру волокна скелетной мышцы
Ключевые моменты
- Мышцы состоят из длинных пучков миоцитов или мышечных волокон.
- Миоциты содержат тысячи миофибрилл.
- Каждая миофибрилла состоит из многочисленных саркомеров, функциональной сократительной области поперечно-полосатой мышцы. Саркомеры состоят из миофиламентов миозина и актина, которые взаимодействуют, используя модель скользящих филаментов и цикл поперечного мостика, чтобы сокращаться.
Основные термины
- саркоплазма : Цитоплазма миоцита.
- саркоплазматический ретикулум : Эквивалент гладкого эндоплазматического ретикулума в миоците.
- сарколемма : Клеточная мембрана миоцита.
- саркомер : функциональная сократительная единица миофибриллы поперечно-полосатой мышцы.

Структура волокон скелетных мышц
Миоциты, иногда называемые мышечными волокнами, составляют основную часть мышечной ткани.Они связаны вместе перимизием, оболочкой из соединительной ткани, в пучки, называемые пучками, которые, в свою очередь, связаны друг с другом, образуя мышечную ткань. Миоциты содержат многочисленные специализированные клеточные структуры, которые облегчают их сокращение и, следовательно, сокращение мышцы в целом.
Высокоспециализированная структура миоцитов привела к созданию терминологии, отличающей их от обычных клеток животных.
Родовая клетка > Миоцит
Цитоплазма > Саркоплазма
Клеточная мембрана > Сарколемма
Гладкий эндоплазматический ретикулум > Саркоплазматический ретикулум
Структура миоцитов
Миоциты могут быть невероятно большими, диаметром до 100 микрометров и длиной до 30 сантиметров.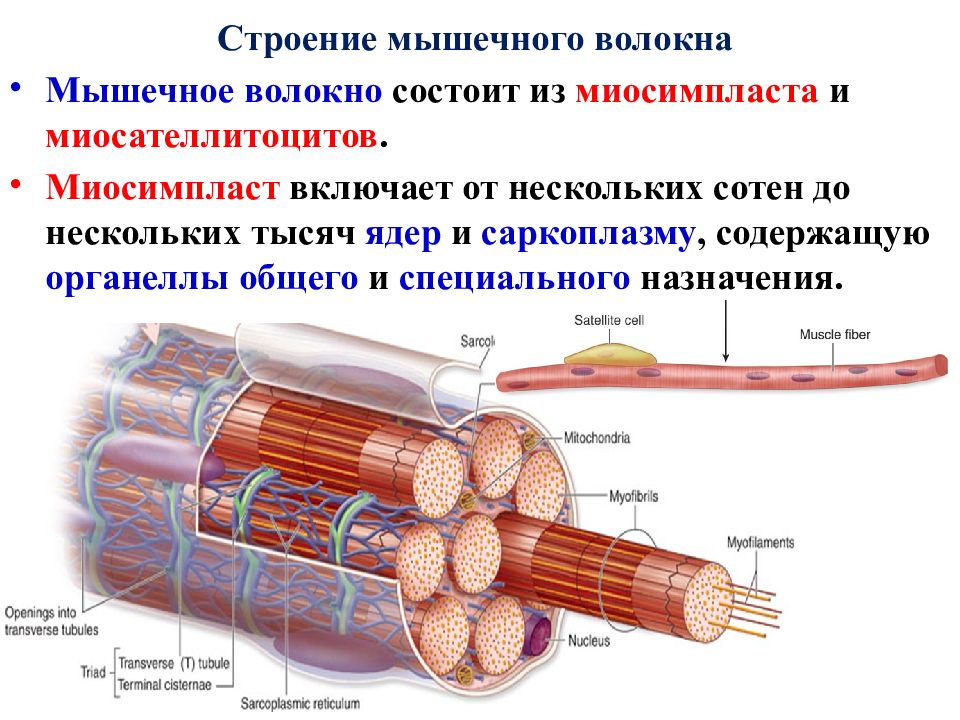 Саркоплазма богата гликогеном и миоглобином, которые хранят глюкозу и кислород, необходимые для выработки энергии, и почти полностью заполнена миофибриллами, длинными волокнами, состоящими из 90 263 миофиламентов, которые способствуют сокращению мышц.
Саркоплазма богата гликогеном и миоглобином, которые хранят глюкозу и кислород, необходимые для выработки энергии, и почти полностью заполнена миофибриллами, длинными волокнами, состоящими из 90 263 миофиламентов, которые способствуют сокращению мышц.
Сарколемма миоцитов содержит многочисленные инвагинации (ямки), называемые поперечными канальцами, которые обычно перпендикулярны длине миоцита. Поперечные канальцы играют важную роль в снабжении миоцита ионами Ca + , которые являются ключевыми для мышечного сокращения.
Каждый миоцит содержит несколько ядер из-за их происхождения из нескольких миобластов, клеток-предшественников, дающих начало миоцитам. Эти миобласты расположены на периферии миоцита и уплощены таким образом, чтобы не влиять на сокращение миоцита.
Рисунок \(\PageIndex{1}\): Миоцит: клетка скелетных мышц : клетка скелетных мышц окружена плазматической мембраной, называемой сарколеммой, с цитоплазмой, называемой саркоплазмой. Мышечное волокно состоит из множества миофибрилл, упакованных в упорядоченные единицы.
Мышечное волокно состоит из множества миофибрилл, упакованных в упорядоченные единицы.Структура миофибрилл
Каждый миоцит может содержать многие тысячи миофибрилл. Миофибриллы идут параллельно миоциту и обычно проходят по всей его длине, прикрепляясь к сарколемме на обоих концах. Каждая миофибрилла окружена саркоплазматическим ретикулумом, который тесно связан с поперечными канальцами. Саркоплазматический ретикулум действует как приемник ионов Ca + , которые высвобождаются при передаче сигналов из поперечных канальцев.
Саркомеры
Миофибриллы состоят из длинных миофиламентов актина, миозина и других ассоциированных белков.Эти белки организованы в области, называемые саркомерами, функциональной сократительной областью миоцита. Внутри саркомера актина и миозина миофиламенты переплетаются друг с другом и скользят друг по другу посредством модели сокращения скользящих филаментов. Регулярная организация этих саркомеров придает скелетным и сердечным мышцам характерный вид исчерченности.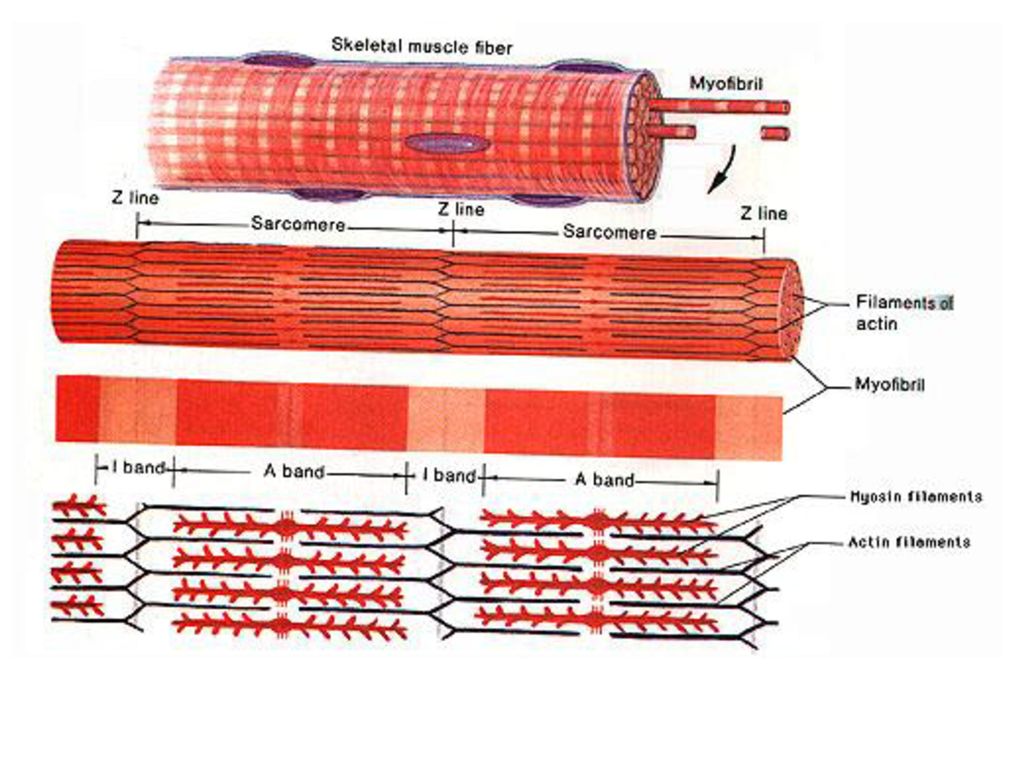
Саркомер : Саркомер представляет собой функциональную сократительную область миоцита и определяет область взаимодействия между набором толстых и тонких филаментов.
Миофиламенты (толстые и тонкие филаменты)
Миофибриллы состоят из более мелких структур, называемых миофиламентами. Различают два основных типа миофиламентов: толстые филаменты и тонкие филаменты. Толстые филаменты состоят в основном из миозиновых белков, хвосты которых связываются вместе, оставляя головки открытыми для переплетения тонких филаментов. Тонкие филаменты состоят из актина, тропомиозина и тропонина. Молекулярная модель сокращения, описывающая взаимодействие между актиновыми и миозиновыми миофиламентами, называется циклом перекрестного мостика.
Разница между миофибриллой и мышечным волокном
Ключевая разница — миофибрилла и мышечное волокно Существует три типа мышечных тканей; сердечные мышцы, скелетные мышцы и гладкие мышцы.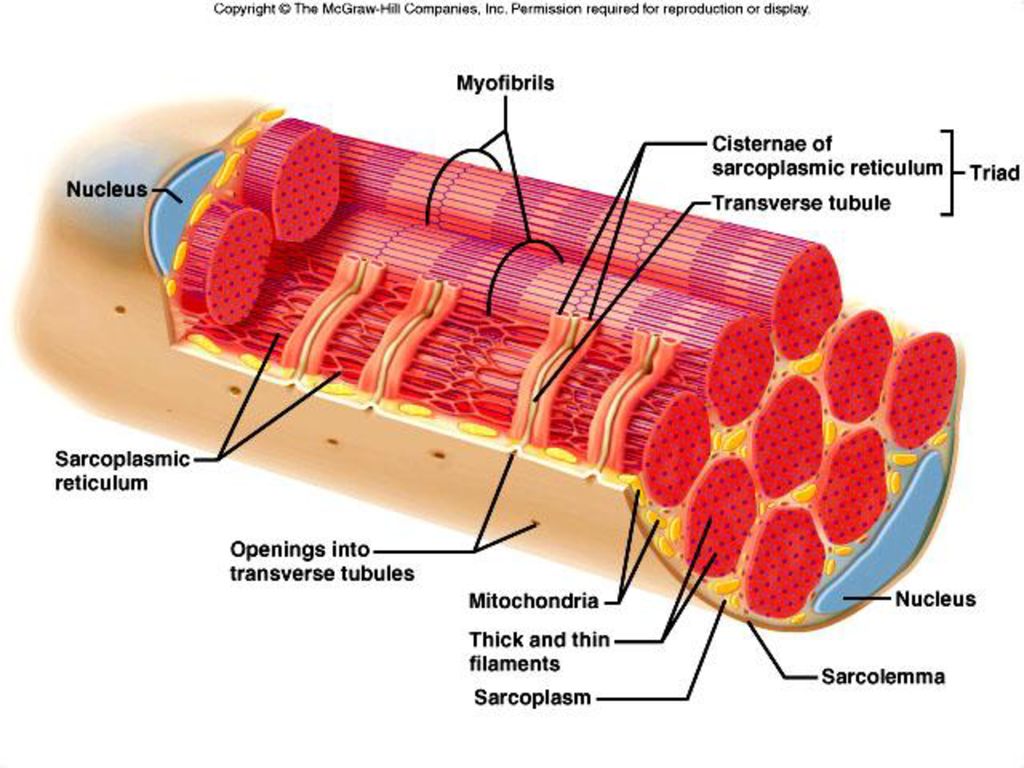 Каждый тип имеет определенную структуру и роль в мышечной системе. Гладкие мышцы образуют такие органы, как мочевой пузырь и желудок. Сердечные мышцы сокращаются и перекачивают кровь по всему телу. Скелетные мышцы помогают костям и другим структурам двигаться.Эти мышцы состоят из длинных пучков клеток, называемых мышечными волокнами или миоцитами. Мышечные волокна состоят из тысяч миофибрилл. Ключевое различие между миофибриллой и мышечным волокном заключается в том, что миофибрилла является основной стержнеобразной единицей мышечного волокна , тогда как мышечное волокно представляет собой трубчатые клетки мышцы.
Каждый тип имеет определенную структуру и роль в мышечной системе. Гладкие мышцы образуют такие органы, как мочевой пузырь и желудок. Сердечные мышцы сокращаются и перекачивают кровь по всему телу. Скелетные мышцы помогают костям и другим структурам двигаться.Эти мышцы состоят из длинных пучков клеток, называемых мышечными волокнами или миоцитами. Мышечные волокна состоят из тысяч миофибрилл. Ключевое различие между миофибриллой и мышечным волокном заключается в том, что миофибрилла является основной стержнеобразной единицей мышечного волокна , тогда как мышечное волокно представляет собой трубчатые клетки мышцы.
СОДЕРЖАНИЕ
1. Обзор и ключевые отличия
2. Что такое мышечное волокно
3. Что такое миофибрилла
4. Сходство между миофибриллой и мышечным волокном
5.Сравнение бок о бок — миофибриллы и мышечные волокна в табличной форме
6. Резюме
Что такое мышечное волокно?
Скелетные мышцы составляют большую часть мышечной системы.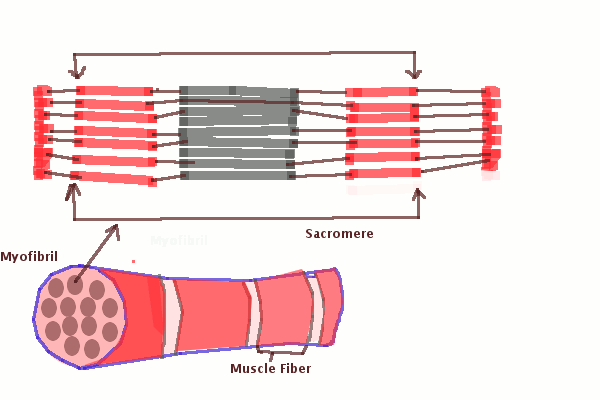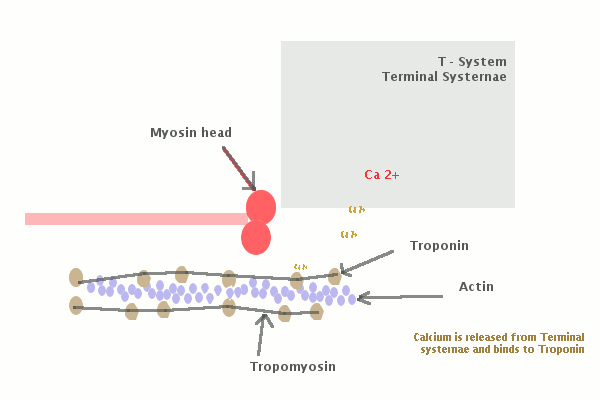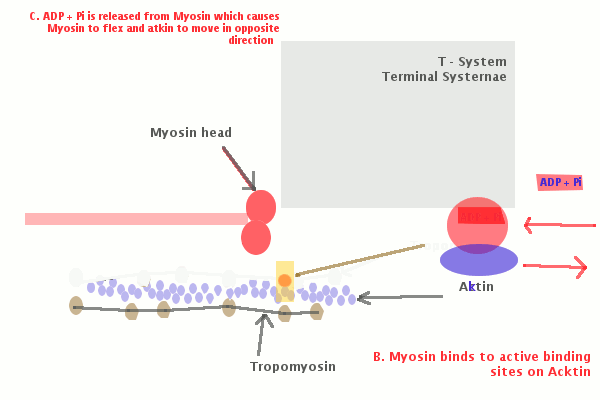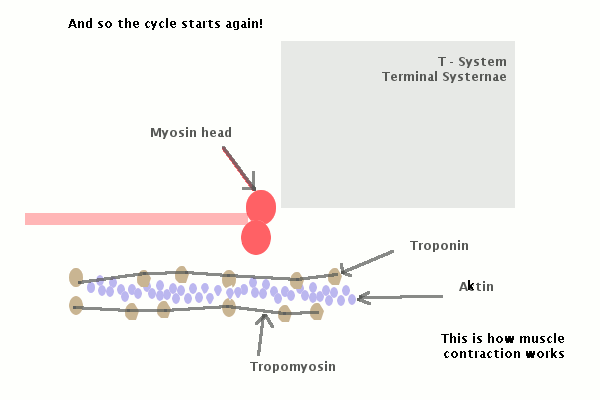 Скелетные мышцы защищены соединительной мышечной тканью, называемой epimysium . Мышца состоит из пучков трубчатых мышечных клеток. Эти трубчатые клетки известны как мышечные волокна или миоцитов . Пучки мышечных волокон известны как пучков . Один пучок мышечных волокон защищен соединительной тканью, известной как перимизий .Внутри перимизия много мышечных волокон. Каждый пучок может содержать от 10 до 100 мышечных волокон. Крупные, сильные мышцы имеют большое количество мышечных волокон внутри каждого пучка. Меньшие мышцы содержат меньшее количество мышечных волокон в пучке.
Скелетные мышцы защищены соединительной мышечной тканью, называемой epimysium . Мышца состоит из пучков трубчатых мышечных клеток. Эти трубчатые клетки известны как мышечные волокна или миоцитов . Пучки мышечных волокон известны как пучков . Один пучок мышечных волокон защищен соединительной тканью, известной как перимизий .Внутри перимизия много мышечных волокон. Каждый пучок может содержать от 10 до 100 мышечных волокон. Крупные, сильные мышцы имеют большое количество мышечных волокон внутри каждого пучка. Меньшие мышцы содержат меньшее количество мышечных волокон в пучке.
Мышечные ткани и мышечные волокна образуются из мезодермального слоя эмбриональных зародышевых клеток в процессе, называемом миогенезом. Каждое мышечное волокно покрыто волокнистой соединительной тканью, называемой эндомизием . Диаметр мышечных волокон может составлять от 10 до 80 микрометров, а их длина может достигать 30 см.
Мышечное волокно состоит из многочисленных палочковидных единиц или цилиндрических органелл, называемых миофибриллами. Каждое мышечное волокно содержит от сотен до тысяч миофибрилл, которые представляют собой пучки миозиновых и актиновых белков, проходящих по всей длине мышечного волокна. Миофибриллы играют важную роль в сокращении мышц.
Рисунок 01: Структура мышечных волокон
Типы мышечных волокон
Существует три основных типа клеток скелетных мышц или мышечных волокон. Это волокна типа I, волокна типа IIa и волокна типа IIb.Волокна типа I также известны как медленные волокна или красные медленные волокна. Волокна типа IIa известны как быстрые окислительные волокна или красные быстрые волокна. Волокна типа IIb известны как быстрые гликолитические волокна или белые быстрые волокна. Каждый тип имеет различные характеристики в функции.
Что такое миофибриллы?
Миофибрилла или мышечная фибрилла представляет собой основную палочковидную единицу мышечной клетки. В мышечной клетке сотни миофибрилл идут параллельно друг другу.Миофибриллы состоят в основном из белков актина и миозина. Некоторые другие типы белков также присутствуют в миофибриллах. Эти белки организованы в толстые и тонкие длинные нити, называемые миофиламентами . Тонкие миофиламенты состоят в основном из белка актина, тогда как толстые филаменты состоят из белка миозина. Эти два типа миофиламентов проходят по всей длине миофибриллы в секциях, называемых саркомерами. Миофибриллы состоят из повторяющихся участков саркомеров. Эти саркомеры выглядят под микроскопом как чередующиеся темные и светлые полосы и ответственны за мышечные сокращения.
В мышечной клетке сотни миофибрилл идут параллельно друг другу.Миофибриллы состоят в основном из белков актина и миозина. Некоторые другие типы белков также присутствуют в миофибриллах. Эти белки организованы в толстые и тонкие длинные нити, называемые миофиламентами . Тонкие миофиламенты состоят в основном из белка актина, тогда как толстые филаменты состоят из белка миозина. Эти два типа миофиламентов проходят по всей длине миофибриллы в секциях, называемых саркомерами. Миофибриллы состоят из повторяющихся участков саркомеров. Эти саркомеры выглядят под микроскопом как чередующиеся темные и светлые полосы и ответственны за мышечные сокращения.
Рисунок 02: Структура миофибриллы
Каковы сходства между миофибриллами и мышечными волокнами?
- Мышечные волокна и миофибриллы отвечают за сокращение мышц.
- Оба типа имеют трубчатую форму.
- Оба типа расположены внутри мышцы параллельно.
Какая связь между миофибриллами и мышечными волокнами?
- Миофибриллы являются основными единицами мышечных волокон. Одно мышечное волокно содержит сотни миофибрилл.
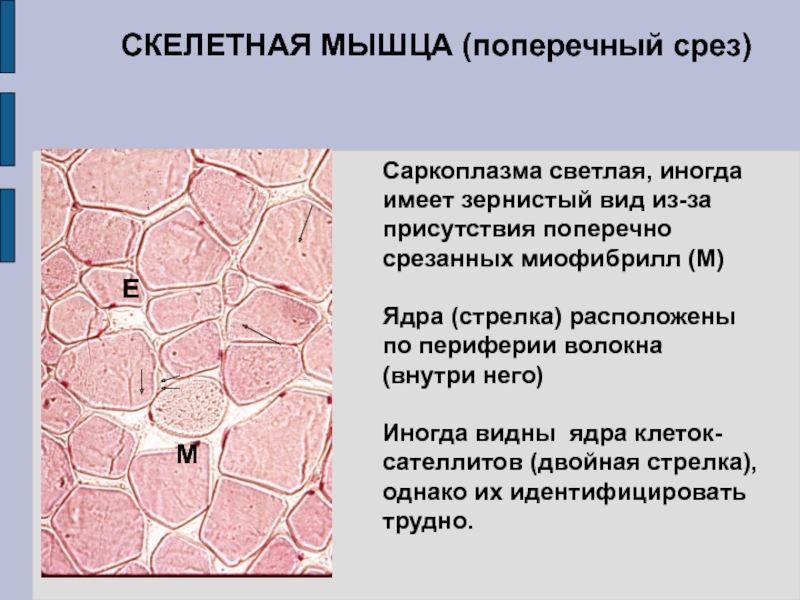 Одно мышечное волокно содержит сотни миофибрилл.
Одно мышечное волокно содержит сотни миофибрилл.В чем разница между миофибриллами и мышечными волокнами?
Миофибриллы против мышечных волокон | |
| Миофибрилла – основная палочковидная единица мышечного волокна. | Мышечное волокно представляет собой мышечную клетку трубчатой формы. |
| Состав | |
| Миофибриллы состоят из двух типов миофиламентов, называемых тонкими и толстыми филаментами. | Мышечное волокно состоит из многочисленных миофибрилл. |
| Природа | |
| Миофибрилла представляет собой цилиндрическую органеллу. | Мышечное волокно представляет собой клетку с ядром и другими органеллами, включая митохондрии. |
Резюме – Миофибриллы против мышечных волокон
Мышечное волокно является основной единицей мышцы. Они располагаются пучками внутри мышцы. Мышечные волокна состоят из многочисленных миофибрилл. Миофибриллы являются основными единицами мышечных волокон. Они состоят из тонких и толстых белковых нитей, называемых миофиламентами.Основными белками миофиламентов являются актин и миозин. Миофиламенты состоят из повторяющихся участков, называемых саркомерами. Их саркомеры отвечают за сокращение мышц. Мышечные волокна и миофибриллы в совокупности способствуют сокращению мышц.
Они располагаются пучками внутри мышцы. Мышечные волокна состоят из многочисленных миофибрилл. Миофибриллы являются основными единицами мышечных волокон. Они состоят из тонких и толстых белковых нитей, называемых миофиламентами.Основными белками миофиламентов являются актин и миозин. Миофиламенты состоят из повторяющихся участков, называемых саркомерами. Их саркомеры отвечают за сокращение мышц. Мышечные волокна и миофибриллы в совокупности способствуют сокращению мышц.
Загрузить PDF-версию исследования «Миофибриллы и мышечные волокна»
Вы можете загрузить PDF-версию этой статьи и использовать ее в автономном режиме в соответствии с примечаниями к цитированию. Загрузите PDF-версию здесь. Разница между миофибриллами и мышечными волокнами.
Каталожные номера:
«Миофибриллы». Википедия. Фонд Викимедиа, 7 июля 2017 г. Интернет. Доступно здесь. 8 июля 2017 г.
«Структура мышц». Структура мышц (и связанных с ними соединительных тканей). н.п., н.д. Веб. Доступно здесь. 08 июля 2017.
Веб. Доступно здесь. 08 июля 2017.
Изображение предоставлено:
1. «1022 мышечных волокна (маленькие)» Автор OpenStax — (CC BY 4.0) через Commons Wikimedia
2. «Myofibril» Автор Slashme из англоязычной Википедии (CC BY-SA 3.0) через Commons Wikimedia
лекция17
лекция1718 февраля 2005
Лекция 17
Чтение, глава 24 & 22
В.Энергия
E. Поток энергии в весы для организма
Все ваши клетки нуждаются в глюкозе и кислороде для аэробного дыхания
4. Правила глюкоза крови
г. Сахарный диабет сахарный диабет
Многие случаи
диабет не диагностируется. Чем раньше начато лечение, тем
лучше. Начальные симптомы диабета включают частое мочеиспускание,
повышенная жажда, сильное чувство голода, похудание, утомляемость,
раздражительность и нечеткость зрения.Если вы или кто-то из ваших знакомых демонстрирует
эти симптомы, врач может проверить способность вашего тела контролировать
уровень глюкозы. В долгосрочной перспективе дайбет может способствовать высокому
кровяное давление, болезни сердца, болезни почек и глаукома. Отложить
или избегать их, люди с диабетом контролируют уровень глюкозы в крови с помощью
специальная диета и/или с инъекциями инсулина.
В долгосрочной перспективе дайбет может способствовать высокому
кровяное давление, болезни сердца, болезни почек и глаукома. Отложить
или избегать их, люди с диабетом контролируют уровень глюкозы в крови с помощью
специальная диета и/или с инъекциями инсулина.
5. Использование АТФ мышечные клетки.
Мышцы и нервы клетки являются большими потребителями АТФ.Нервные клетки потребляют большое количество глюкоза, которую они используют для производства АТФ при аэробном дыхании. Мышечные клетки могут использовать как жирные кислоты, так и глюкозу для своих аэробных функций. дыхание.
АТФ используется для двоих вещи в мышечных клетках: активный транспорт кальция (Ca ++) и движение моторных белков. В нервах АТФ используется главным образом для активный транспорт натрия (Na + ) и калия (K + ), но также стимулирует секрецию нейротрансмиттера. химических веществ эндомембранной системой.
а.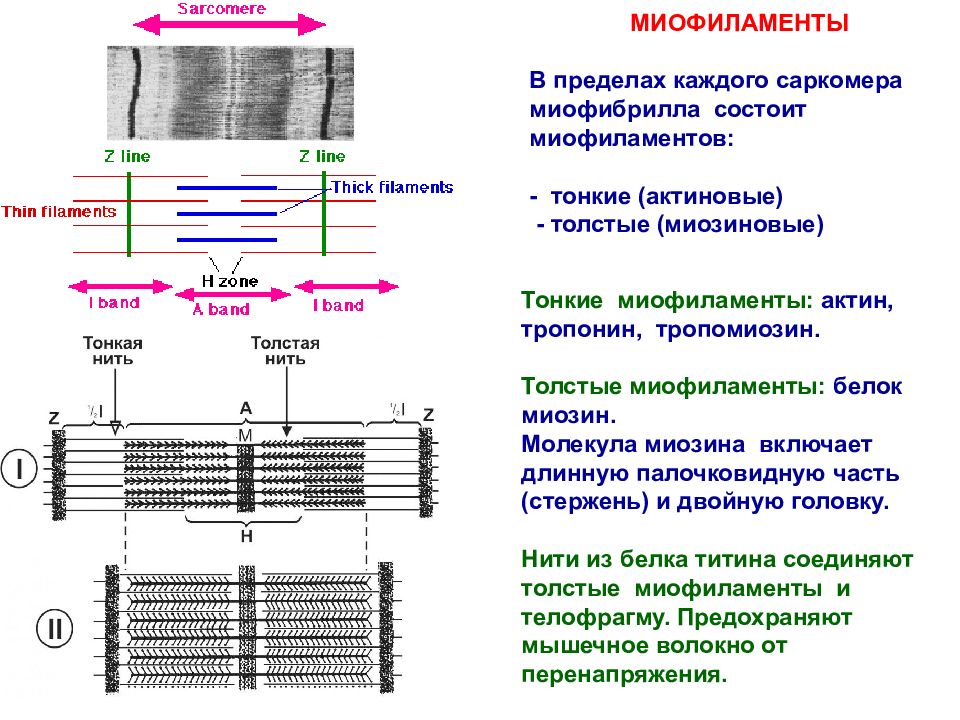 скелет
мышцы
скелет
мышцы
Скелетные мышцы те, над которыми у вас есть произвольный контроль с помощью вашего мозга и нервы. Они двигают кости вашего скелета, что заставляет вас переехать.
Клетки скелета мышцы необычны. Они являются результатом того, что несколько клеток имеют слились в длинное «мышечное волокно» с несколькими ядрами, которые происходит в начале развития.
Внутренние мышечные волокна представляет собой множество актиновых микрофиламентов, которые проходят вдоль мышечное волокно. Между этими актиновыми микрофиламентами расположены длинные тяжи миозин. Миозин является моторным белком, т.е. белком, который захватывает микротрубочки или микрофиламенты и перемещается по ним. актин-миозин структуры в мышечных волокнах называются «миофибриллами» (см. стр. 517–520). в вашем тексте изображения мышечных волокон и миофибриллы).
Когда мышца
контракты, это происходит потому, что миозиновые нити миофибрилл тянут
две актиновые микрофиламенты вместе. Сокращение происходит в ответ на
ионы кальция (Ca ++ ), которые связываются с актином
микрофиламенты и создают «зацепки» для миозина.
незащищенный.
Сокращение происходит в ответ на
ионы кальция (Ca ++ ), которые связываются с актином
микрофиламенты и создают «зацепки» для миозина.
незащищенный.
Кальций в мышцах волокна хранятся в сложном эндоплазматическом ретикулуме (ЭР), который называется «саркоплазматический ретикулум» (SR), чтобы отличить его от ER в другие типы клеток. Когда волокно скелетной мышцы получает нерв сигнал, SR высвобождает накопленный кальций, захватывает миозин пряди обнажаются, и мышечное волокно сокращается.В течение расслабление мышечного волокна, кальций закачивается обратно в SR при подготовке к следующему нервному сигналу.
АТФ используется мышцами
волокна двумя способами. Во-первых, он используется транспортными белками для
«активный транспорт» кальция в СР между сокращениями. В
покоя СР мышечного волокна имеет значительно более высокую концентрацию
кальций внутри, чем снаружи. При получении нервного сигнала кальций
каналы в СР ненадолго открываются и кальций устремляется в цитозоль
за счет «селективной диффузии», которая представляет собой диффузию одного типа растворенного вещества
через транспортный белок, не использующий АТФ. Мышца начинается
заключать соглашение.
Мышца начинается
заключать соглашение.
Второе использование для АТФ в мышечных волокнах позволяет миозиновым «рукам» отпустить актин после того, как они потянули один раз и схватили другую рукоять выше чтобы они могли снова потянуть и продолжить сокращение. Когда актин поручни подвергаются воздействию кальция, связывающегося с актиновыми микрофиламентами, миозин спонтанно захватывает актиновую опору и тянет один раз. Чтобы для того, чтобы отпустить эту рукоятку и снова потянуть, ATP должна обеспечить энергия для высвобождающего движения.
Таким образом, расходуется АТФ
с высокой скоростью за счет сокращения мышц. Потребность в АТФ в мышцах
клеток иллюстрируется явлением «трупного окоченения», которое
мышечная ригидность, возникающая в мертвых телах в течение короткого времени после
смерть. В этих мышцах АТФ полностью преобразуется в АДФ.
митохондрии не могут преобразовать АДФ обратно в АТФ, потому что нет
кислород доступен (дыхательные движения остановлены). В отсутствие
АТФ, транспортные белки Ca ++ перестают качать
Ca ++ в SR и Ca ++ в SR
постепенно просачивается, обнажая сайты связывания миозина.
актиновые филаменты миофибрилл.Белки миозина захватывают актин
и потяните один раз, но не можете отпустить и снова потянуть. Мышцы
поэтому оставайтесь неподвижными в позе смерти до связывания
миозин в актин начинает распадаться и они становятся рыхлыми
снова.
В отсутствие
АТФ, транспортные белки Ca ++ перестают качать
Ca ++ в SR и Ca ++ в SR
постепенно просачивается, обнажая сайты связывания миозина.
актиновые филаменты миофибрилл.Белки миозина захватывают актин
и потяните один раз, но не можете отпустить и снова потянуть. Мышцы
поэтому оставайтесь неподвижными в позе смерти до связывания
миозин в актин начинает распадаться и они становятся рыхлыми
снова.
.