
Микрофибриллы — строение, вид, химический состав, свойства, функции, разновидности, вики — Wiki-Med
Основная статья: Субмембранный комплексСодержание (план)
Строение микрофибрилл
С помощью иммунохимических методов удалось выяснить, что в состав микрофибриллярных структур входят основные и вспомогательные сократимые белки, характерные для мышечных клеток: актин, миозин, α-актинин и тропомиозин. Эти белки в клетках образуют два типа структур: располагающуюся в основном под мембраной сложную сеть микрофиламентов и так называемые «нити натяжения», или stress-fibers — тесно связанные длинные пучки микрофиламентов. Названные структуры различаются между собой по чувствительности к цитохалазину — яду, специфически действующему на актиновые фибриллы. Под воздействием цитохалазина сеть микрофиламентов разрушается в течение нескольких десятков минут, для дезорганизации stress-fibers необходимо 1-2 суток. «Тянущая» роль stress-fibers отчетливо продемонстрирована в экспериментах по перерезке их лазерным лучом, при этом края клетки медленно растягиваются.
Белки, составляющие сеть микрофиламентов, располагаются строго определенным образом. Так, при обработке культивируемых in vitro клеток первичной культуры фибробластов флюоресцирующими антителами к актину на препаратах выявляется сложная сеть светящихся фибрилл, при этом свечение наблюдается и по всей длине фибрилл, и в местах их пересечения (рис. 5, А). При обработке клеток антителами к α-актинину свечение отмечается только в области пересечения актиновых микрофибрилл (рис. 5, Б). Наконец, обработка клеток антителами к тропомиозину позволяет получить следующую картину: как и в случае использования антител к актину, свечение наблюдается по всей длине микрофибрилл, но отсутствует в области их пересечения (рис. 5, В). Весьма вероятно, что эти данные отражают истинную локализацию изучаемых белков в сложной «сеточке» микрофибрилл, тем более что подобное взаимное расположение сократимых белков характерно и для мышечных клеток. Материал с сайта http://wiki-med.com
Функции микрофибрилл
Оказалось, что микрофибриллярная система отличается большой динамичностью и пластичностью. Она существенно изменяется при изменении состояния клеток в культуре: в процессе прикрепления и перемещения клеток по субстрату, а также при образовании временных и постоянных межклеточных контактов. В процессе прикрепления клеток к субстрату постепенно меняется их форма — от округлой (типичной для взвешенного состояния) до полного распластывания по субстрату. У прикрепленных клеток микрофибриллярная система становится значительно мощнее и переплетение микрофиламентов приобретает другой характер. Эти иммуноцитохимические данные были подтверждены многочисленными морфологическими исследованиями, выполненными с помощью обычной и высоковольтной электронной микроскопии. Обнаружилось, что в местах контакта клеток между собой и субстратом в субмембранной гиалоплазме образуется мощная сеть микрофибрилл, которая отсутствует в других участках периферической гиалоплазмы. Это становится возможным .за счет самосборки микрофибрилл, слагаемых фибриллярным актином (F-актином), из мономерного глобулярного актина (G-актина), сосредоточенного в периферической гиалоплазме.
Она существенно изменяется при изменении состояния клеток в культуре: в процессе прикрепления и перемещения клеток по субстрату, а также при образовании временных и постоянных межклеточных контактов. В процессе прикрепления клеток к субстрату постепенно меняется их форма — от округлой (типичной для взвешенного состояния) до полного распластывания по субстрату. У прикрепленных клеток микрофибриллярная система становится значительно мощнее и переплетение микрофиламентов приобретает другой характер. Эти иммуноцитохимические данные были подтверждены многочисленными морфологическими исследованиями, выполненными с помощью обычной и высоковольтной электронной микроскопии. Обнаружилось, что в местах контакта клеток между собой и субстратом в субмембранной гиалоплазме образуется мощная сеть микрофибрилл, которая отсутствует в других участках периферической гиалоплазмы. Это становится возможным .за счет самосборки микрофибрилл, слагаемых фибриллярным актином (F-актином), из мономерного глобулярного актина (G-актина), сосредоточенного в периферической гиалоплазме.
МЫШЕЧНАЯ ТКАНЬ — ГИСТОЛОГИЯ
МЫШЕЧНАЯ ТКАНЬ — ГИСТОЛОГИЯ ГИСТОЛОГИЯ В ТАБЛИЦАХ И СХЕМАХ МЫШЕЧНАЯ ТКАНЬ
СТРОЕНИЕ МИОФИБРИЛЛ
СТРОЕНИЕ МИОФИБРИЛЛ
- в цитоплазме имеется большое количество миофибрилл, обеспечивающих сокращение; миофибриллы состоят из актиновых (тонких) и миозиновых (толстых) микрофибрилл
- актиновая микрофибрилла (тонкая)
- представляет собой тонкую нить
- основу актиновой микрофибриллы составляет белок актин, который имеет фибриллярную структуру
- на актине есть места для связывания миозина
- в поперечнополосатой мышечной ткани к актину присоединены еще несколько белков,
образующих тропонин-тропомиозиновый комплекс:
- тропомиозин — закрывает на молекуле актина места для связывания с миозином
- тропонин С — присоединяет ионы кальция; после присоединения кальция сдвигает молекулу тропомиозина с ее первоначального расположения, что приводит к открытию на молекуле актина мест для связывания с миозином
- тропонин Т и тропонин I — выполняют структурную функцию
- в гладкой мышечной ткани тропонин-тропомиозинового комплекса нет
- актиновые микрофибриллы прикрепляются к цитоскелету клетки в области Z-линий с помощью специальных белков, таких как альфа-актинин, виментин, десмин
- миозиновая микрофибрилла (толстая)
- представляет собой толстую нить
- построена из молекул миозина, имеется множество типов миозина с разной скоростью расщепления АТФ, что обуславливает отличия в скорости сокращения разных мышечных волокон
- молекула миозина похожа на клюшку для игры в гольф (или уж на худой конец — в хоккей), в ней различают головку (это та часть клюшки, которая ударяет по мячу или шайбе) и (рукоятка клюшки)
- миозиновая микрофибрилла представляет собой пучек таких клюшек, связанных за рукоятки, причем часть головок смотрит в одну сторону, а часть — в другую (передне-заднее направление)
- участки миозиновых микрофибрилл, где находятся головки, вставлены между актиновыми микрофибриллами
- миозиновые микрофибриллы прикрепляются к цитоскелету клетки в области линии М (середина полоски Н)
- головка миозина может: 1)поворачиваться, 2)прикрепляться к актину, 3)расщеплять АТФ, то есть является АТФ-азой
- головка миозина может присоединяться к актину только тогда, когда она содержит АДФ и Фосфат (продукты распада АТФ)
- головка миозина, соединенная с актином, может совершать гребковое движение только в момент, когда от нее отсоединяются АДФ и Фосфат
- головка миозина может отсоединиться от актина только тогда, когда она присоединяет к себе молекулу АТФ
- в гладкой мышечной ткани головка миозина имеет легкие цепи, которые должны сначала фосфорилироваться, для того чтобы она смогла расщеплять и присоединять АТФ и взаимодействовать с актином
- миофибриллы строго ориентированы вдоль волокна
- актиновые и миозиновые микрофибриллы располагаются параллельно друг другу
- благодаря строгой ориентации миофибрилл мышечное волокно и кардиомиоциты имеют поперечную исчерченность
- поперечная исчерченность — это чередование светлых и темных полос или дисков на протяжении миофибрилл
- миофибрилла устроена так, что по ее длине имеются участки актиновых микрофибрилл, между которыми располагаются участки миозионвых микрофибрилл, и миозиновые микрофибриллы на небольшое расстояние проникают в пространства между актиновыми; так, что на концах актиновых и миозиновых участков имеются области, где есть и актиновые, и миозиновые микрофибриллы
- одним концом актиновые микрофибриллы прикрепляются к цитоскелету, это место называется Z-линией,
- в своей середине миозиновые микрофибриллы скрепляются с цитоскелетом, это место называется М-линией
- различают следующие виды дисков, полосок и линий на миофибриллах:
- I-диск (изотропный) — светлый диск, в пределах которого располагаются только актиновые микрофибриллы
- А-диск (анизотропный) — темный диск, в области которого располагаются актиновые и миозиновые микрофибриллы
- Н-нолоска — светлая полоса, располагающаяся в середине А-диска, здесь имеются только миозиновые микрофибриллы
- М-линия — находится в середине Н-полоски, здесь прикрепляются миозиновые микрофибриллы
- Z-линия — находится в середине I-диска, здесь прикрепляются актиновые микрофибриллы с помощью белков альфа-актинина, виментина и десмина
- саркомер — это участок мышечного волокна между двумя соседними Z-линиями, структурно-функциональная единица поперечнополосатой мышечной ткани
- в гладкомышечных клетках миофибриллы расположены беспорядочно, актиновые микрофибриллы одним своим концом прикрепляются к специальным областям внутренней поверхности цитомембраны, а другим — к миозину, миозиновые микрофибриллы прикрепляются к специальным местам в цитозоле клетки
© A Gunin; histol@mail.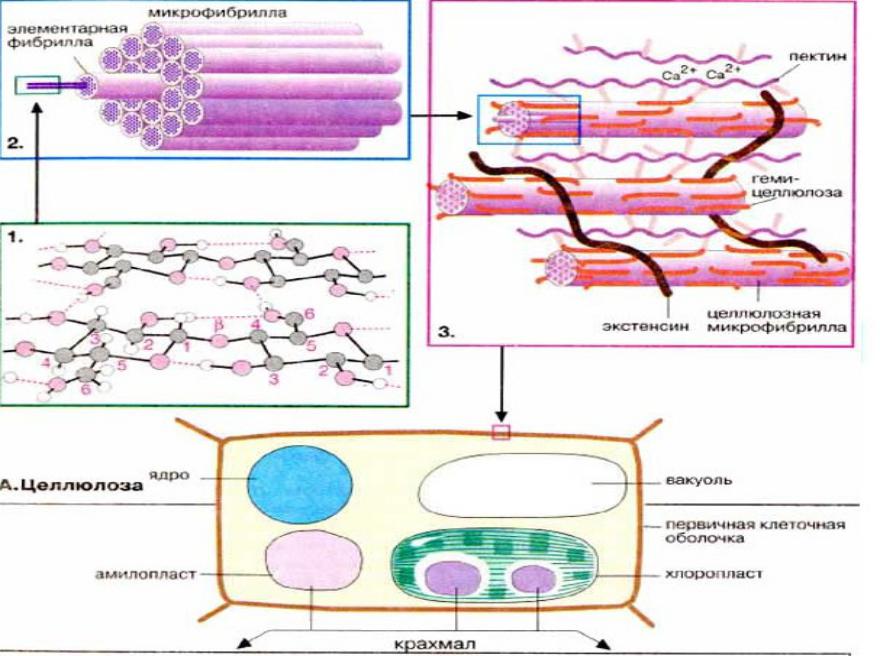 ru
ru
Как растянуть железобетон – Наука – Коммерсантъ
Система, с помощью которой растения увеличиваются в размерах, состоит из большого количества полимеров и ферментов. Пока воспроизвести эту систему искусственно не удается — более того, не все детали процесса понятны. Но когда это получится, строительная индустрия получит значительное технологическое ускорение.
Само слово «растение» подразумевает, что это нечто, способное расти. Рост — это необратимое увеличение объема или массы живого организма. Здесь мы ограничимся рассмотрением роста растений на клеточном уровне, имея в виду только увеличение объема клеток.
В отличие от клеток животных, которые отделены от внешней среды только тонкой жировой пленкой — мембраной, растительные клетки помимо нее имеют еще один внешний слой — клеточную стенку (рис. 1А). Компоненты растительных клеточных стенок известны большинству из нас как пищевые волокна, которые так приветствуются диетологами в рационе. Но кроме того, древесина, бумага, вата, льняные, хлопковые и бамбуковые ткани — все это растительные клеточные стенки и материалы, которым нет альтернативы.
Клеточная стенка в основном состоит из полимеров углеводной природы — полисахаридов. Неизменным компонентом всех растительных клеточных стенок является целлюлоза — наиболее распространенное органическое соединение на Земле. Целлюлоза состоит из глюкозы, молекулы которой соединены друг с другом последовательно в цепочку длиной до 10000 единиц. За счет периодичности своего строения цепочки целлюлозы могут взаимодействовать друг с другом продольно и частично кристаллизоваться. Несколько дюжин цепочек соединяются вместе, образуя стержневидные структуры, называемые микрофибриллами. Целлюлозные микрофибриллы — это «арматура» клеточной стенки (рис. 1Б). Сравнение тем более уместно, если учесть, что прочность целлюлозы сопоставима с прочностью стали.
Помимо целлюлозы в растительных клеточных стенках присутствуют также гемицеллюлозы и пектины. Гемицеллюлозы (греч. hemi- — полу-) были названы так из-за определенного сходства с целлюлозой. Основная цепь (остов) этих полисахаридов по своей форме похожа на целлюлозу.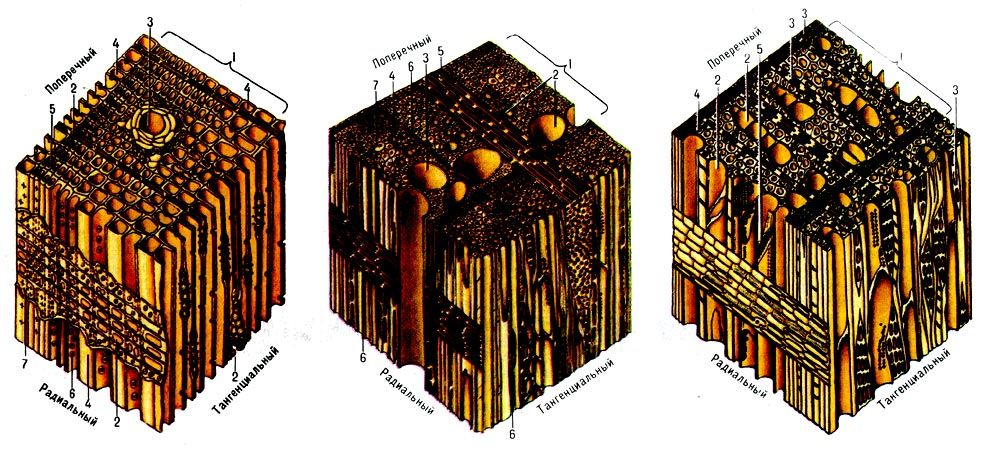
Пектины отличаются от остальных полисахаридов растительных клеточных стенок тем, что их остов отрицательно заряжен. В результате взаимодействия с положительно заряженными ионами кальция или бора пектиновые вещества образуют гели, что отражено в их названии (греч. пектос — свернувшийся, замерзший), а в повседневной жизни можно наблюдать при желировании джемов или мармелада из различных фруктов.
Итак, несущая конструкция растительных клеточных стенок образована целлюлозными микрофибриллами и соединяющими их гемицеллюлозами (рис. 1Б, В). Она уложена в гель, который формируют пектиновые вещества. В определенном смысле ее устройство можно сравнить с железобетоном, в котором элементы арматуры обеспечивают необходимую прочность, а бетон — совместную работу армирующих элементов и их защиту от повреждений.
Рост клеток происходит под действием их внутреннего гидростатического давления. У растений оно намного выше, чем у животных, и составляет в среднем 5–10 атмосфер. Клеточные стенки, «надутые» внутриклеточным давлением, функционируют как гидростатический скелет растения. Поэтому, когда растениям не хватает воды, они поникают: давление снижается, и скелет перестает работать.
Чтобы заставить клетки увеличить свой объем, можно было бы поднять в них давление, подобно тому, как мы заставляем «расти» воздушный шар, надувая его. Но растения поступают иначе. Во-первых, учитывая «железобетонную» прочность стенок, давление пришлось бы повысить еще на несколько атмосфер, чтобы достичь хоть какого-то роста, а это энергозатратно и чревато разрывами при контакте клеток с окружающими предметами.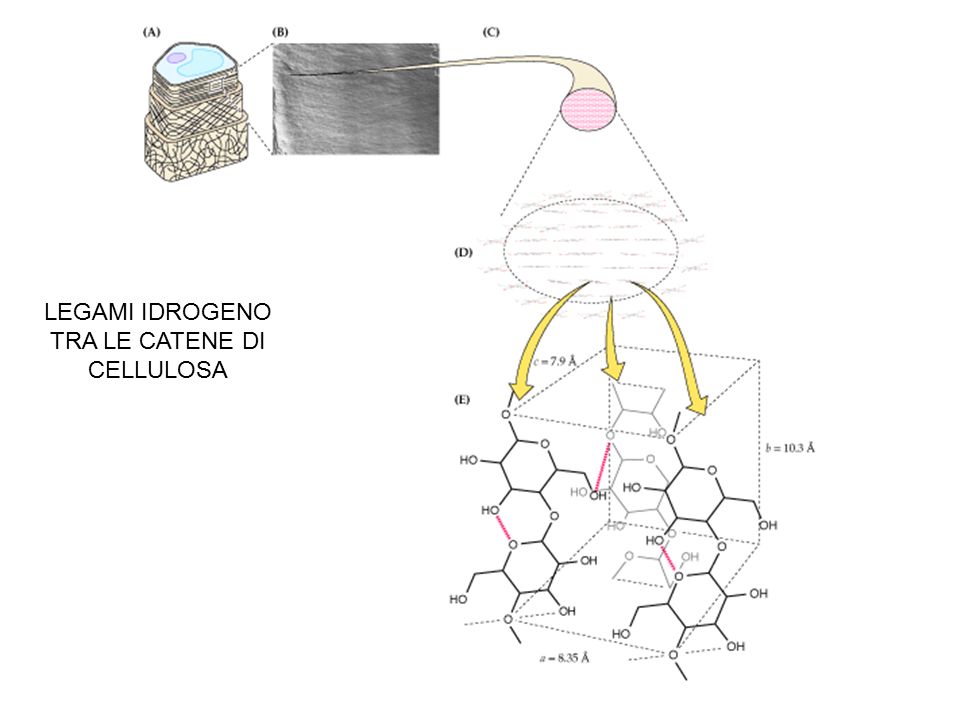 Во-вторых, давление действует одинаково во всех направлениях, а значит, все взрослые клетки были бы шарообразными, тогда как для выполнения своих функций им часто необходимо иметь сложную форму. Не имея возможности обеспечить необходимые скорость и направление роста клеток за счет изменения давления, растение управляет запуском, развитием и остановкой роста, регулируя механические свойства клеточных стенок.
Во-вторых, давление действует одинаково во всех направлениях, а значит, все взрослые клетки были бы шарообразными, тогда как для выполнения своих функций им часто необходимо иметь сложную форму. Не имея возможности обеспечить необходимые скорость и направление роста клеток за счет изменения давления, растение управляет запуском, развитием и остановкой роста, регулируя механические свойства клеточных стенок.
Использование нанодатчиков позволило установить, что растительные клетки, медленно увеличивающие свой объем или закончившие этот процесс, имеют более упругие клеточные стенки, чем те клетки, которые активно растут. Более того, разные участки клеточных стенок одной клетки могут иметь разную упругость, и, соответственно, именно те части, упругость которых ниже, будут преимущественно растягиваться. За счет этого растительные клетки могут приобретать сложные формы.
Одну из главных ролей в деле определения направления роста клеток и их конечной формы играет ориентация микрофибрилл целлюлозы (рис.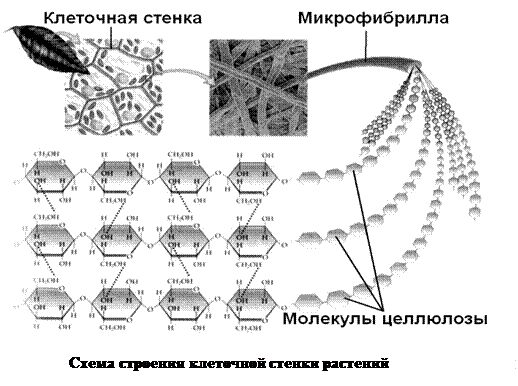 2). Рост в основном идет в направлении, перпендикулярном расположению микрофибрилл. По мере увеличения объема клетки расстояние между микрофибриллами растет, но одновременно изнутри прибывают новые порции компонентов стенки, что позволяет ей сохранять необходимую толщину и прочность.
2). Рост в основном идет в направлении, перпендикулярном расположению микрофибрилл. По мере увеличения объема клетки расстояние между микрофибриллами растет, но одновременно изнутри прибывают новые порции компонентов стенки, что позволяет ей сохранять необходимую толщину и прочность.
Можно предположить несколько вариантов того, как именно достигается увеличение расстояния между микрофибриллами и растяжение стенки (рис. 3). Частичное разрушение микрофибрилл (рис. 3А) может позволить это сделать, однако это ослабит стенку, и такой сценарий никогда не реализуется в растениях за неимением подходящих ферментов. Можно разрушить не целлюлозу, а часть гемицеллюлоз (рис. 3Б), чтобы микрофибриллы могли передвинуться друг относительно друга. Это также ослабит материал, хоть и в меньшей степени. Его прочность может быть восстановлена за счет прибытия новых порций полисахаридов, которые закрепят микрофибриллы на новом расстоянии друг от друга. Вариантом этого же сценария можно считать особый тип реакции, когда гемицеллюлозы после возникновения разрыва на остове связываются с доступной молекулой другого полисахарида (рис. 3В). Еще один возможный механизм — это изменение степени взаимодействия между микрофибриллами и гемицеллюлозами (рис. 3Г). По-видимому, именно так действуют на клеточную стенку белки экспансины. Экспансины появляются в тех органах и тканях, которые приступают к активному росту. Приложение «компресса» с экспансинами приводит к снижению упругости клеточных стенок в этом месте и вызывает вспучивание ткани, то есть ускоряет рост отдельных клеток.
3В). Еще один возможный механизм — это изменение степени взаимодействия между микрофибриллами и гемицеллюлозами (рис. 3Г). По-видимому, именно так действуют на клеточную стенку белки экспансины. Экспансины появляются в тех органах и тканях, которые приступают к активному росту. Приложение «компресса» с экспансинами приводит к снижению упругости клеточных стенок в этом месте и вызывает вспучивание ткани, то есть ускоряет рост отдельных клеток.
Экспансины не обладают ферментативной активностью, и механизм их действия неясен. Дополнительно облегчить реорганизацию целлюлозно-гемицеллюлозной сети можно, уменьшив прочность геля, в котором она располагается (рис. 3Д). Этот механизм реализуется за счет модификации пектинов, которая лишает их заряда и возможности к взаимодействию с ионами кальция.
Все эти варианты в той или иной мере участвуют в регуляции механических свойств клеточных стенок и роста клеток, однако до полного понимания этих процессов нам еще очень далеко. Сейчас мы не можем синтезировать в пробирке даже самый простой полисахарид — целлюлозу, не говоря о создании многокомпонентного и динамичного углеводного ансамбля, которым является клеточная стенка. Когда мы поймем, как все это устроено, возможно, мы научимся создавать материалы, сравнимые с теми, которые делают растения.
Когда мы поймем, как все это устроено, возможно, мы научимся создавать материалы, сравнимые с теми, которые делают растения.
Автор благодарит Российский научный фонд за финансовую поддержку тематики (номер проекта — 18-14-00168) и коллег за критику и советы.
Людмила Козлова, кандидат биологических наук, старший научный сотрудник Казанского института биохимии и биофизики Федерального исследовательского центра «Казанский научный центр Российской академии наук»
Строение волоса и подбор процедур. Статьи компании «KERATIN-PROF Беларусь»
Немного информации о строении волос, обязательна к изучению начинающим и будущим мастерам. Также в статье Вы узнаете, как научится подбирать процедуры по типам и состоянию волос
Чтобы иметь возможность правильно подобрать составы для процедур, нужно понимать, как устроены волосы, от чего и как они разрушаются и как их можно восстановить.
Начнем со строения.
Мы будем говорить о той части волоса, которая считается мертвой, то есть наружном стержне.
В процентном соотношении волос имеет следующий химический состав:
78%-кератин
6%-липиды
1%-пигмент
15%-вода
Часть волоса, расположенная над поверхностью эпидермиса, называется стержнем. Невидимая часть, расположенная в толще кожи, называется корнем (луковицей).
Современные средства по уходу за волосами бренда эффективно работают на стержне волоса. Волос состоит из следующих составляющих:
1. Кутикула представляет собой внешнюю оболочку из ороговевших чешуек. Она служит защитой для внутренних слоев от внешних воздействий. Именно кутикула обеспечивает блеск и шелковистость волос.
2. Кортекс (корковый слой волоса) находится непосредственно под кутикулой. Он состоит из множества ороговевших веретенообразных клеток, которые можно сравнить с нитью. Эти переплетенные «ниточки» образуют микрофибриллы волоса, которые составляют крупные волокна-макрофибриллы. Чтобы миллионы микрофибрилл не распадались между собой на молекулярном уровне, существуют поперечные связи: дисульфидные, водородные и солевые.
Из-за особенностей в строении кортекса волосы могут быть прямыми и кудрявыми, которые в свою очередь наследуются генетически. В клетках этого слоя находятся пигменты, придающие волосам цвет. Кортекс также отвечает за прочность и эластичность. В данном слое происходит химический процесс выпрямления, разглаживания или окрашивания волос.
3. Медула (сердцевина) — это стержень волоса, который состоит из 2-4 слоев, не ороговевших клеток различной структуры с кератиноподобным веществом. Медула отвечает за терморегуляцию, объем и придает силу волосам. В наше время сердцевина у многих прерывистая или полностью отсутствует
Чешуйчатый слой предохраняет нижележащие клетки кортекса от пагубного действия условий внешней среды. Кутикула волоса защищает кортекс и медуллу как панцирь. Она образована перекрывающимися слоями прозрачных пластинок кератина, напоминающих чешую. Кутикула препятствует механическим и химическим воздействиям на волос. Неповрежденная кутикула хорошо отражает свет: волосы блестят, не ломаются. Средства с высоким РН (щелочные — мыло, например), расслабляют чешуйки и заставляют их открываться и пропускать жидкость внутрь кортекса. Средства с низким уровнем РН (кислые) захлопывают чешуйки, делают их плотнее и прочнее.
Средства с высоким РН (щелочные — мыло, например), расслабляют чешуйки и заставляют их открываться и пропускать жидкость внутрь кортекса. Средства с низким уровнем РН (кислые) захлопывают чешуйки, делают их плотнее и прочнее.
Вот здесь кроется первый важный момент, который нужно понимать: чешуйки кутикулы раскрываются и закрываются от изменения уровня РН!!! Например, РН воды в море чаще имеет щелочную среду, РН воды в кране может иметь щелочную среду, и большинство шампуней (особенно масс-маркет) — это щелочи. Значит, если ваши волосы подверглись воздействию щелочи, то вам необходимо обработать их кислым средством, чтобы они закрылись (например, бальзам). Волосы с раскрытыми чешуйками кутикулы очень хрупкие, легко разрушаются и, конечно же, не блестят.
Второй важный момент: если вас интересует красота волос, их блеск, то вам нужно позаботиться о состоянии кутикулы!
Кутикулу тоже можно механически повредить, например, грубым расчесыванием, горячими щипцами. В этом случае даже при обработке специальными средствами она не сможет обеспечить полную защиту кортекса, и волос начнет разрушаться. Поэтому очень важно бережно обращаться с волосами, мягко расчесывать их без резких движений, а также как можно реже подвергать их нагреванию и сушке. В этом случае рекомендуется использовать термозащитные спреи и лосьоны.
В этом случае даже при обработке специальными средствами она не сможет обеспечить полную защиту кортекса, и волос начнет разрушаться. Поэтому очень важно бережно обращаться с волосами, мягко расчесывать их без резких движений, а также как можно реже подвергать их нагреванию и сушке. В этом случае рекомендуется использовать термозащитные спреи и лосьоны.
Теперь перейдем к кортексу, среднему слою, который составляет примерно 90% волоса человека. Именно в этом слое происходят изменения при окрашивании или химии. От состояния кортекса зависят сила, упругость и прочность волоса. Волос с хорошо развитым корковым слоем значительно прочнее волоса со слабо развитым слоем.
Корковый слой окружает сердцевину. Он представляет собой плотно прилегающие одна к другой клетки, продольно вытянутые и расположенные по длине волоса. При рассмотрении под электронным микроскопом видно, что эти клетки состоят из тончайших волокон, фибрилл, а те — из еще более тонких волокон, и так далее, пока не дойдем до молекулярного уровня. Здесь можно провести аналогию со строением каната, который сплетен из более тонких волокон. Волокна кортекса состоят в основном из белка — кератина. Белок состоит из длинных цепей аминокислот, которые в свою очередь состоят из химических элементов. Основными химическими элементами, из которых состоит человеческий волос являются: углерод, кислород, водород, азот и сера. Из этих элементов так же состоят кожа и ногти.
Здесь можно провести аналогию со строением каната, который сплетен из более тонких волокон. Волокна кортекса состоят в основном из белка — кератина. Белок состоит из длинных цепей аминокислот, которые в свою очередь состоят из химических элементов. Основными химическими элементами, из которых состоит человеческий волос являются: углерод, кислород, водород, азот и сера. Из этих элементов так же состоят кожа и ногти.
Белки — это длинные аминокислоты, соединенные между собой на концах прочными белковыми связями. Связываясь друг с другом, они образуют длинные, спиральные, сложные аминокислотные цепочки. Эти спирали образуют микрофибриллы, которые перекручиваясь, образуют волокна кортекса. Итак, мы разобрались, как волокна образуются в длину. Но что удерживает их вместе, как в тугом канате? Чтобы миллионы полипептидных цепочек не распадались, между ними также существуют поперечные связки или мостики. Связки эти удерживают волокна вместе на молекулярном уровне.
Эти связи имеют огромное значение для таких услуг, как термическая укладка, химическая завивка и химическое выпрямление волос. Основными поперечными связями являются: водородные, ионные или солевые и дисульфидные
Основными поперечными связями являются: водородные, ионные или солевые и дисульфидные
Водородные связи легко разрушаются водой и температурой. И хотя по отдельности водородные связи очень слабые, их очень много в волосах, поэтому они дают примерно одну треть силы волоса. Водородные связи легко разрушаются при намокании волос. Это позволяет волосам легко растягиваться. При высыхании и охлаждении водородные связи восстанавливаются и фиксируют форму волос в том положении в котором волосы оказались. Так, например, завивка волос на обычных бигуди и высушивание их естественным образом приводит к разрушению водородных связей, благодаря чему, волосы из прямых становятся закрученными, но через некоторое время или под воздействием влажной среды, разрушенные водородные связи восстанавливаются и волосы снова приобретают прежнюю форму.
Ионные или солевые связи так же являются слабыми физическими связями, образующимися поперек волоса между соседними белковыми цепочками. Ионные связи зависят от pH на волосах, поэтому они легко разрушаются сильными щелочными и кислыми растворами.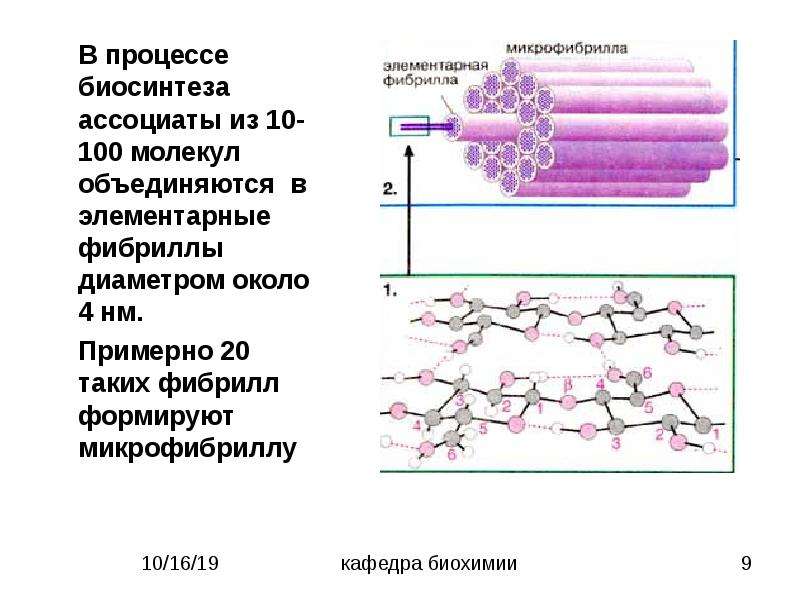 Но их в волосах так же много, поэтому оказывают значительное влияние на структуру и свойства волос. Солевые связи восстанавливаются при нормальном рН волос.
Но их в волосах так же много, поэтому оказывают значительное влияние на структуру и свойства волос. Солевые связи восстанавливаются при нормальном рН волос.
Дисульфидная связь — это сильная химическая связь, которая очень отличается от физических водородной и ионной. Она соединяет атомы серы двух соседних аминокислот, чтобы создать одну, объединяет две белковые цепочки в единое целое.
Хотя самих дисульфидных связей гораздо меньше, чем водородных и ионных, они очень сильные и не разрушаются водой. Разорвать дисульфидную связь можно с помощью препаратов для химической завивки и выпрямления. Нормальное температурное воздействие, как, например, при термоукладке, не разрушает дисульфидных связей. Однако очень сильные воздействия температуры, как при воздействии кипятка и некоторых высокотемпературных инструментов для укладки, нарушают эти связи. Яркий пример такого сильного воздействия — утюжки для выпрямления волос, температура которых достигает более 200 градусов.
Если дисульфидные связи были разорваны и не восстановлены химическим путем, то волос остается поврежденным. Со временем он становится хрупким, ломким, пористым, сильно обезвоженным и истонченным. Это приводит к посеченным концам, волосы тяжелее расчесываются и становятся на ощупь жесткими и сухими.
Со временем он становится хрупким, ломким, пористым, сильно обезвоженным и истонченным. Это приводит к посеченным концам, волосы тяжелее расчесываются и становятся на ощупь жесткими и сухими.
Итак, очередной вывод: если вас беспокоят сила и упругость ваших волос, то вам нужно задуматься о восстановлении кортекса волоса, то есть восстановлении дисульфидных связей (всегда используйте специальные средства при химическом воздействии или окрашивании! а также специальные шампуни и бальзамы, например, с гематином) и восстановлении волокон кортекса
Кстати, кератины в составе шампуней не способны проникнуть вглубь волоса, поэтому восстанавливают лишь частично, заполняя пустоты у внешней оболочки непосредственно под чешуйками кутикулы. При несильном повреждении волос, они могут дать восстанавливающий эффект, но при глубоких повреждения — не очень эффективны.
Кроме всего прочего в клетках коркового слоя обычно располагаются зерна красящего вещества — пигмента. При окрашивании этот пигмент разрушается и заменяется искусственным, но искусственный пигмент не является таким устойчивым, поэтому легко вымывается из кортекса волоса при мытье головы (после окрашивания постарайтесь не использовать сульфатные шампуни, это позволит дольше сохранить цвет), оставляя после себя пустоты с воздухом, что делает волос более хрупким.
Вообще, конечно, восстановление волос — это комплексная терапия, включающая в себя как лечение, восстановление, так и защиту волос от дальнейших повреждений. Она требует дополнительных мер, ухаживающих средств и большого желания и терпения.
С ЧЕГО НАЧАТЬ?
После детального изучения строения волос, нужно научиться понимать, как правильно подбирать процедуру по типу и состоянию волос клиента.
1. После знакомства с клиентом необходимо узнать у него «историю» его волос. Какие были окрашивания, какой оксид использовался. Окрашивали ли волосы хной? Были ли смывки/осветления. Делал ли клиент процедуры кератинового выпрямления/ботокса ранее? Как реагировали на процедуру волосы и сколько состав срок носки
2. Также уточняйте у клиента, не принимает ли он серьезные гормональные препараты, о беременности, о кормлении грудью
Далее проведите визуальный осмотр волос клиента, сделайте тест на степень повреждения волос (намочите волос и аккуратно растяните его).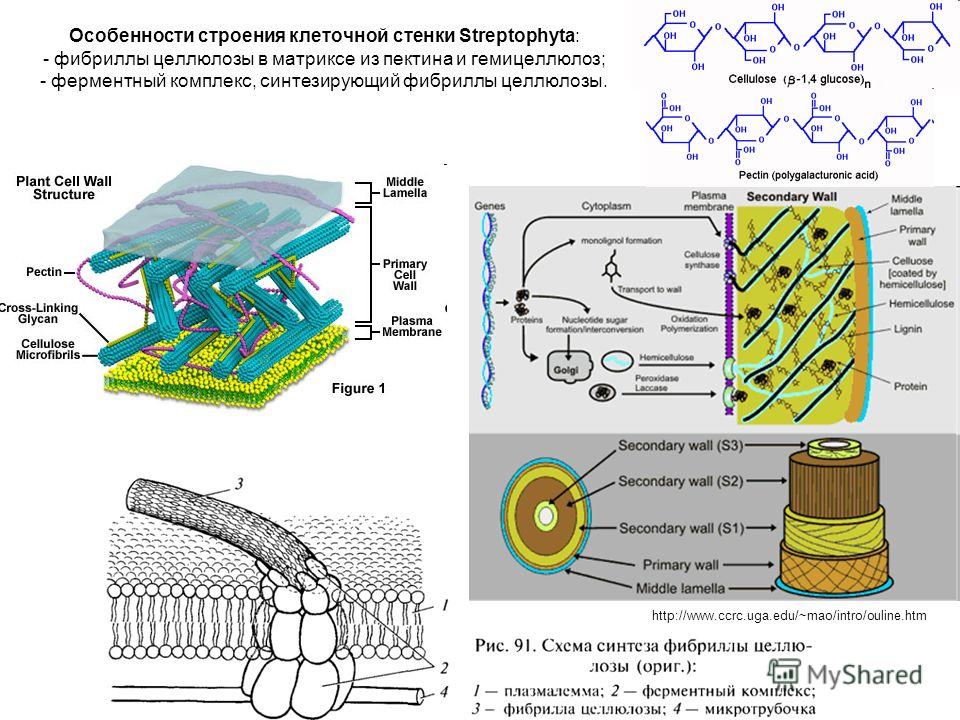 Если волосы растягиваются и не возвращаются в прежнее состояние-таким волосам не рекомендуется проводить «горячие» процедуры, т.е. процедуры с использованием утюжка, также можно проверить волосы на пористость.
Если волосы растягиваются и не возвращаются в прежнее состояние-таким волосам не рекомендуется проводить «горячие» процедуры, т.е. процедуры с использованием утюжка, также можно проверить волосы на пористость.
В случае если волосы сильно повреждены, а именно рвутся-рекомендуется провести процедуру сос-реконструкции холодным способом. Можно использоваться составы Felps SOS, ампулы от Dr.Therapy, каутеризацию от Gold Spell и другие составы для реконструкции.Такие составы направлены именно на восстановление дисульфидных связей волоса. Эти процедуры можно делать с помощью инфракрасного утюжка, иногда требуется курс восстановительных процедур (необходимо смотреть за состоянием волос). Курс может составлять 2-5 процедур.
После курса реконструкции, когда волосы перестали рваться, а при растягивании возвращаются в прежнюю форму-можно выполнять более серьезные восстановительные процедуры с использованием утюжка.
БОТОКС ИЛИ КЕРАТИН?
В целом это довольно схожие процедуры, большинство современных составов для ботокса по компонентам близки к составам для кератинового выпрямления волос.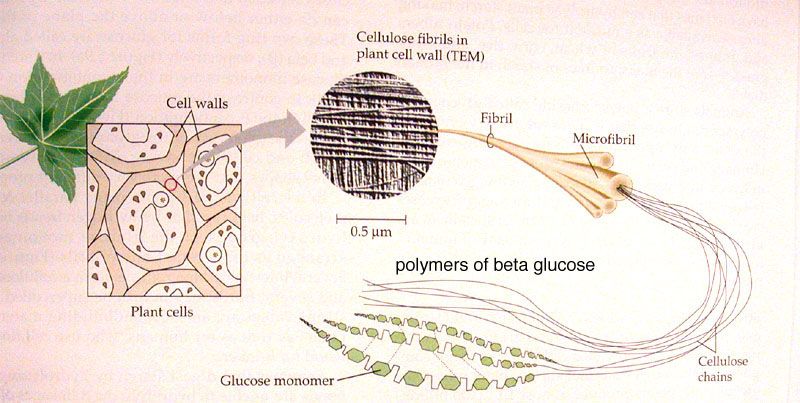 Как правило, в ботоксах содержится большее количество ухаживающих, увлажняющих компонентов (масла, кодиционеры), чем в кератине. За счет этого составы для ботокса дают больший эффект увлажнения и восстановления волос.
Как правило, в ботоксах содержится большее количество ухаживающих, увлажняющих компонентов (масла, кодиционеры), чем в кератине. За счет этого составы для ботокса дают больший эффект увлажнения и восстановления волос.
Кератин в первую очередь направлен на выпрямление волос, содержит большее количество выпрямляющих компонентов. Современные составы для выпрямления не сильно уступают ботоксам в содержании увлажняющих компонентов. Поэтому при правильно подобранном по типу волос составу можно получить отличный результат выпрямления и восстановления/увлажнения волос.
На какие волосы рекомендуется делать именно ботокс:
Окрашенные на высоких оксида (блонд), волосы после смывки, осветления, других сложных окрашиваний-после оценки состояния волос. На волосы после химической завивки (также в зависимости от состояния волос). Также ботокс рекомендован на ПРЯМЫЕ от природы волосы (натуральные, окрашенные), так как такие волосы нет смысла перегружать «тяжелыми» составами для выпрямления волос, а также не подвергать волосы серьезной термообработке, как при выпрямлении волос.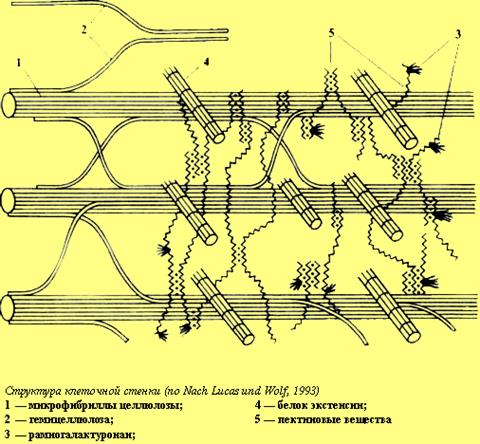 С ботоксом можно работать на более низких температурах.
С ботоксом можно работать на более низких температурах.
Также ботокс можно делать на вьющиеся волосы, для структуризации кудрей, устранения пушистости, увлажнения. Важно при этом подобрать ботокс без эффекта выпрямления. Есть довольно сильные составы для ботокса, которые смогут на некоторое время выпрямить кудри.
Основной показатель выполнения процедуры кератинового выпрямления-вьющиеся волосы, которые клиент хочет выпрямить, при этом часто пользуется утюжком и регулярно травмирует их. Далее стоит учитывать состояние волос, убедиться, что волосы смогут выдержать необходимую для термообработки состава температуру (оптимальная температура утюжка при работе с кератином 210-230 градусов). В зависимости от состояния волос температура может быть ниже, но это уже может сказаться на эффекте выпрямления и сроке носки состава. Поэтому, если Вы видите, что волосы не выдержат высокую температуру-предварительно подготовьте их, сделайте более щадящую процедуру-ботокс или реконструкцию. После этих процедур волосы смогут без труда выдержать серьезную термообработку и можно сделать выпрямление волос через некоторое время.
После этих процедур волосы смогут без труда выдержать серьезную термообработку и можно сделать выпрямление волос через некоторое время.
Строение целлюлозных микрофибрилл — Справочник химика 21
Строение целлюлозных микрофибрилл [c.237] Многие полисахариды, присутствующие в растениях, обнаружены в клеточных стенках. Стенки представляют собой сложную переплетенную решетку из целлюлозных микрофибрилл, окруженную аморфным межклеточным веществом. Основными компонентами межклеточного вещества являются пектиновые вещества, лигнин и гемицеллюлозы. Гемицеллюлозы — это те полисахариды клеточной стенки, которые не относятся ни к пектиновым веществам, ни к целлюлозе. Гемицеллюлозы представляют собой настолько разнородную и условно определяемую группу, что целесообразность такого термина вызывает большое сомнение. В препаратах гемицеллюлоз чаще всего присутствуют ксиланы, маннаны, глюкоманнаны, галактаны и арабаны. Часто в них находятся также Г-рамноза и В-галактуроновая кислота. В большинстве препаратов преобладают ксиланы, хотя гемицеллюлоза из некоторых видов древесины и семян особенно богата маннанами. Число и тип соединений, присутствующих в препаратах гемицеллюлозы из разных тканей, значительно различаются между собой кроме того, наблюдаются большие различия в относительном содержании компонентов и в деталях их строения. [c.175]
В большинстве препаратов преобладают ксиланы, хотя гемицеллюлоза из некоторых видов древесины и семян особенно богата маннанами. Число и тип соединений, присутствующих в препаратах гемицеллюлозы из разных тканей, значительно различаются между собой кроме того, наблюдаются большие различия в относительном содержании компонентов и в деталях их строения. [c.175]
Свойства целлюлозы определяются не только строением ее отдельных цепных молекул, но и взаимным их расположением, т. е. надмолекулярной и морфологической структурой волокна. Изучение строения целлюлозного волокна в основном при помощи физических методов исследования (метода двойного лучепреломления, микроскопии и электронной микроскопии, инфракрасной спектроскопии, рентгенографии и электронографии) привело к созданию теории ориентированного (аморфно-кри-сталлического) строения целлюлозы. В клеточных стенках древесины целлюлоза находится в виде тончайших волоконцев — целлюлозных микрофибрилл.
 Длинные цепные молекулы целлюлозы проходят вдоль микрофибрилл на ряде участков ориентированно (т. е. параллельно друг другу и на близких расстояниях), а на ряде других участков их ориентация менее совершенна. Участки целлюлозы, в которых существует совершенный порядок в трех пространственных направлениях (т. е. совершенная ориентация), называют ориентированными участками, кристаллитами, или мицеллами (в современном понимании). Длина этих участков около 500—600 А, ширина 50— 100 А. Участки, в которых совершенный порядок отсутствует и сохраняется лишь общая продольная направленность цепей, называются неориентированными, или аморфными (рис. 35). Ориентация цепей в кристаллитах поддерживается за счет сил межмолекулярного взаимодействия — сил Ван-дер-Ваальса и, [c.67]
Длинные цепные молекулы целлюлозы проходят вдоль микрофибрилл на ряде участков ориентированно (т. е. параллельно друг другу и на близких расстояниях), а на ряде других участков их ориентация менее совершенна. Участки целлюлозы, в которых существует совершенный порядок в трех пространственных направлениях (т. е. совершенная ориентация), называют ориентированными участками, кристаллитами, или мицеллами (в современном понимании). Длина этих участков около 500—600 А, ширина 50— 100 А. Участки, в которых совершенный порядок отсутствует и сохраняется лишь общая продольная направленность цепей, называются неориентированными, или аморфными (рис. 35). Ориентация цепей в кристаллитах поддерживается за счет сил межмолекулярного взаимодействия — сил Ван-дер-Ваальса и, [c.67]Клеточная стенка образована сетью целлюлозных микрофибрилл. Длина фибрилл и строение сети могут существенно различаться у разных видов и в разных тканях, но диаметр фибрилл обычно не [c.142]
Следующий за ним слой 82 образован параллельно расположенными микрофибриллами целлюлозы, идущими почти параллельно оси волокна в виде крупной спирали, имеющей угол наклона к оси волокна от 10 до 20 .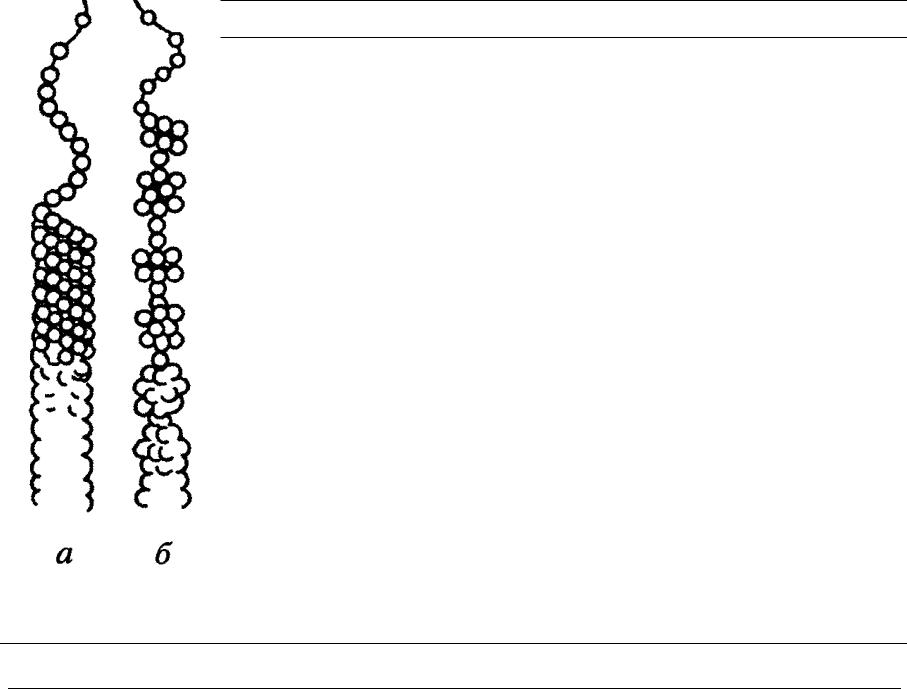 Между слоями 81 и 82 имеется резко выраженный переход. В этом месте целлюлозная матрица имеет рыхлое строение и при поперечных срезах тупым ножом часто [c.319]
Между слоями 81 и 82 имеется резко выраженный переход. В этом месте целлюлозная матрица имеет рыхлое строение и при поперечных срезах тупым ножом часто [c.319]
Согласно теории аморфно-кристаллического строения целлюлозы, ее цепные макромолекулы проходят в продольном направлении микрофибрилл, образуя чередующиеся участки, различающиеся степенью упорядоченности, — кристаллические и аморфные. Эти участки, если принять структурное понятие фазы, можно рассматривать как кристаллическую и аморфную фазы. Поверхность раздела между фазами отсутствует. Кристаллические участки постепенно переходят в аморфные, а последние снова в кристаллические. Поскольку длина макромолекул целлюлозы (2,5 мкм и более) намного превышает длину кристаллических участков, каждая целлюлозная цепь проходит последовательно ряд кристаллических и аморфных участков. Кристаллические участки называют также кристаллитами (см. также 5.3.3 и рис. 5.7). [c.237]
Общая схема структуры целлюлозы в настоящее время представляется следующей.
 Пучки цепных макромолекул соединяются, образуя микрофибриллы, которые, в свою очередь, соединяются в фибриллы. Последние расположены под углом друг к другу или в виде спиралей определенного строения, образуя слои в клеточной стенке древесины или в целлюлозных волокнах хлопка. Эти слои могут перемежаться со слоями нецеллюлозного вещества, такого как лигнин и пектин. [c.260]
Толстая стенка растительной клетки (рис. 1-3) устроена необычайно сложно [ИЗ—116]. Благодаря ее сложному строению растения обладают прочностью и жесткостью, а их клетки способны к быстрому удлинению в период роста. Норткот [ИЗ] сравнил строение стенки растений с фибраглассом — пластиком, армированным стекловолокном. Так, в стенке клетки находятся микрофибриллы, состоящие из целлюлозы и других полисахаридов, которые погружены в матрикс, также состоящий в основном из полисахаридов. На ранних стадиях роста зеленых растений закладывается первичная клеточная стенка, содержащая свободно переплетенные целлюлозные волокна диаметром приблизительно 10 нм, центральная часть которых (- 4 нм) имеет кристаллическую структуру.
Пучки цепных макромолекул соединяются, образуя микрофибриллы, которые, в свою очередь, соединяются в фибриллы. Последние расположены под углом друг к другу или в виде спиралей определенного строения, образуя слои в клеточной стенке древесины или в целлюлозных волокнах хлопка. Эти слои могут перемежаться со слоями нецеллюлозного вещества, такого как лигнин и пектин. [c.260]
Толстая стенка растительной клетки (рис. 1-3) устроена необычайно сложно [ИЗ—116]. Благодаря ее сложному строению растения обладают прочностью и жесткостью, а их клетки способны к быстрому удлинению в период роста. Норткот [ИЗ] сравнил строение стенки растений с фибраглассом — пластиком, армированным стекловолокном. Так, в стенке клетки находятся микрофибриллы, состоящие из целлюлозы и других полисахаридов, которые погружены в матрикс, также состоящий в основном из полисахаридов. На ранних стадиях роста зеленых растений закладывается первичная клеточная стенка, содержащая свободно переплетенные целлюлозные волокна диаметром приблизительно 10 нм, центральная часть которых (- 4 нм) имеет кристаллическую структуру. Такие целлюлозные волокна содержат 8000—12 000 остатков глюкозы. [c.395]
Такие целлюлозные волокна содержат 8000—12 000 остатков глюкозы. [c.395]Микрофибриллы целлюлозы — Справочник химика 21
Было показано, что микрофибриллы представляют собой каркас, скелет разросшейся первичной клеточной оболочки. Если из таких оболочек удалить все остальные компоненты, кроме целлюлозы, то это почти не повлияет на форму клетки и на механические свойства стенок [15, 23]. В настоящее время общепринято, что в оболочке меристематических клеток, у которых первичная клеточная оболочка не развита, микрофибриллы целлюлозы сами по себе не мешают удлинению клетки в направлении главной оси растения и в то же время препятствуют увеличению размеров клетки в направлении, перпендикулярном ее оси. Такое различие имеет огромное значение для растения, так как приводит к развитию удлиненных, вытянутых клеток, ориентированных в направлении роста. Таким образом, расположение микрофибрилл является основным фак- [c.
 88]
Первый наружный слой вторичной стенки 81 откладывается непосредственно на первичную стенку. Он иногда называется переходным, так как его структура является промежуточной между структурами первичной и вторичной стенки. Микрофибриллы целлюлозы в этом слое лежат параллельно друг другу под углом около 50° к оси волокна. Электронномикроскопические исследования показали, что слой 81 состоит из двух или более слоев с перекрестной фибриллярной структурой. [c.319]
88]
Первый наружный слой вторичной стенки 81 откладывается непосредственно на первичную стенку. Он иногда называется переходным, так как его структура является промежуточной между структурами первичной и вторичной стенки. Микрофибриллы целлюлозы в этом слое лежат параллельно друг другу под углом около 50° к оси волокна. Электронномикроскопические исследования показали, что слой 81 состоит из двух или более слоев с перекрестной фибриллярной структурой. [c.319] В бумажном производстве для обеспечения образования большого числа межволоконных связей широко применяется жирный помол массы, в основе которого лежит фибрилляция и набухание поверхностных слоев волокон. Этот процесс протекает наиболее быстро, если между микрофибриллами и отдельными пучками макромолекул целлюлозы располагаются гемицеллюлозы. Они, по-видимому, способствуют более быстрому отрыванию части микрофибрилл целлюлозы от поверхности целлюлозных волокон и вызывают их быструю гидратацию. В результате этого поверхность волокон покрывается слизью, способствующей склеиванию отдельных волокон и образованию тяжей между волокнами после их высыхания. [c.388]
В результате этого поверхность волокон покрывается слизью, способствующей склеиванию отдельных волокон и образованию тяжей между волокнами после их высыхания. [c.388]
Нативное состояние, в котором находятся ксиланы и маннаны в древесине, остается пока неизвестным. Однако тот факт, что кристаллизация их происходит лишь только после их химической модификации, а именно после удаления боковых цепей, позволяет предположить, что гемицеллюлозы выполняют роль аморфного наполнителя , занимая пространство между микрофибриллами целлюлозы. [c.19]
Тонкая первичная стенка обычно состоит из неупорядоченной решетки микрофибрилл целлюлозы, заполненной гемицеллюлозами, а позднее и лигнином. Первичная клеточная стенка в молодых растущих клетках расширяется до тех пор, пока не прекратится рост их и не начнет откладываться вторичная стенка. [c.319]
Следующий за ним слой 82 образован параллельно расположенными микрофибриллами целлюлозы, идущими почти параллельно оси волокна в виде крупной спирали, имеющей угол наклона к оси волокна от 10 до 20 . Между слоями 81 и 82 имеется резко выраженный переход. В этом месте целлюлозная матрица имеет рыхлое строение и при поперечных срезах тупым ножом часто [c.319]
Между слоями 81 и 82 имеется резко выраженный переход. В этом месте целлюлозная матрица имеет рыхлое строение и при поперечных срезах тупым ножом часто [c.319]
Слои вторичной стенки хорошо различимы на микрофотографиях, полученных в поляризованном свете, благодаря различной ориентации в разных слоях микрофибрилл целлюлозы (см. 8.6.2), обладающей вследствие кристаллической структуры двойным лучепреломлением. Слои 8 , 82 и 8з(Т) существенно различаются по толщине 8 и 8з(Т) тонкие, а 8а толстый и образует основную массу клеточной стенки. Во всех этих слоях уже преобладает целлюлоза. Слой 81 имеет толщину 0,1…0,3 мкм в зависимости от части годичного кольца (поздняя или ранняя) и древесной породы. Толщина слоя 82 составляет в среднем 2…6 мкм с колебаниями от 1 мкм (в ранней древесине) до 7…9 мкм (в поздней древесине). Слой 8з(Т) самый тонкий (0,1.. .0,2 мкм) строение его в значительной степени зависит от древесной породы. [c.216]
Еще до начала разделения материнской клетки на клеточную пластинку начинают откладываться микрофибриллы целлюлозы.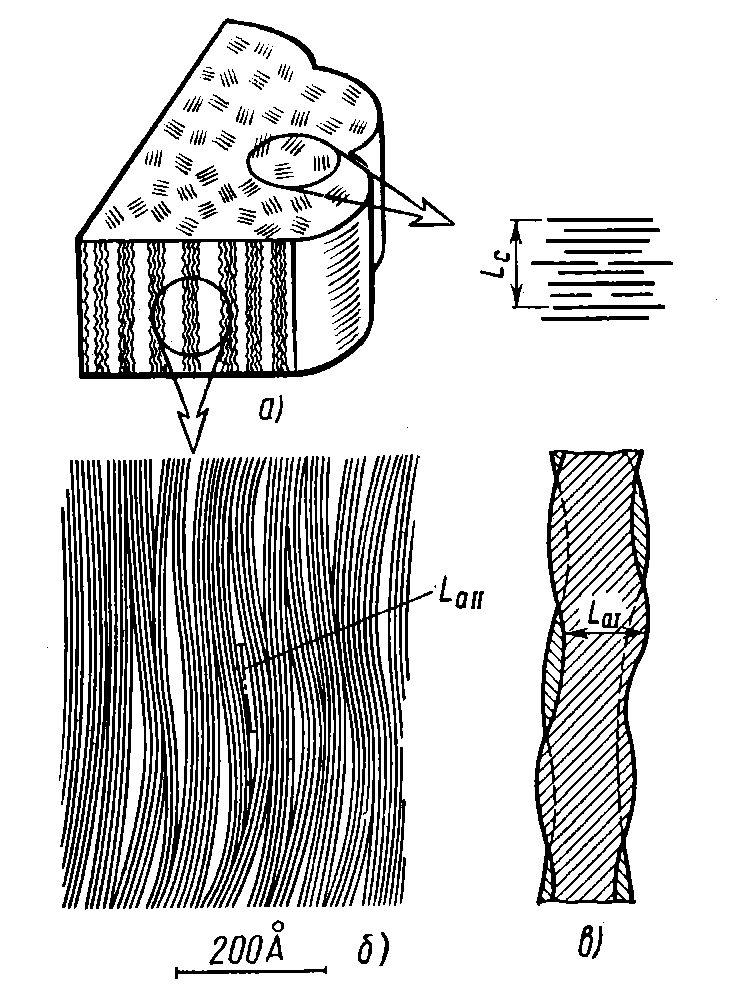 При цитокинезе каждый протопласт выделяет новую первичную оболочку, окружая себя ею. По окончании цикла деления клетка переходит в состояние, называемое рост растяжением . [c.28]
При цитокинезе каждый протопласт выделяет новую первичную оболочку, окружая себя ею. По окончании цикла деления клетка переходит в состояние, называемое рост растяжением . [c.28]
Аморфные области в микрофибриллах целлюлозы изучены недостаточно, и сведения об их надмолекулярной структуре ограничены. Уподоблять структуру аморфных областей структуре препаратов полностью аморфной (аморфизированной) целлюлозы неправомерно. В аморфных областях целлюлозы по сравнению с полностью разупорядоченным состоянием конформационные превращения макромолекул целлюлозы более ограничены. [c.245]
Вторичная стенка. В ней сосредоточена основная масса вещества стенки. Она состоит из каркаса, построенного из микрофибрилл целлюлозы, образующих под различными углами к оси клетки спиральные слои наружный 5ь средний 5 и внутренний 5з. [c.280]
Химический состав отдельных слоев клеточной стенки некоторых растительных материалов приводится далее (см.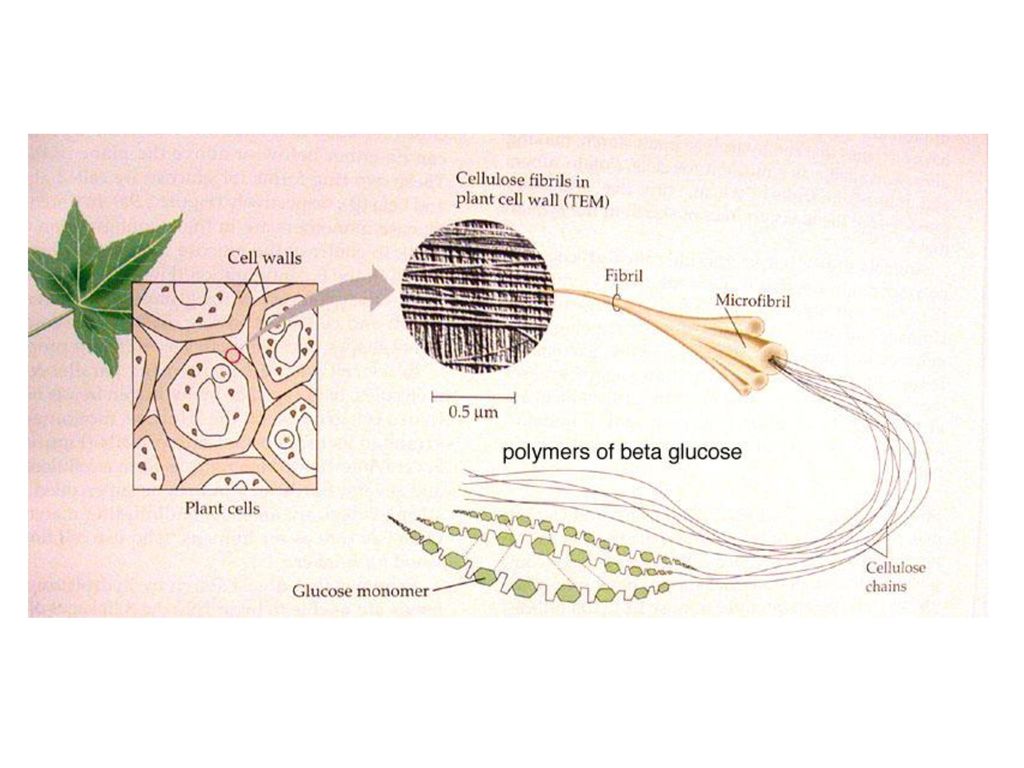 ниже, табл. 1.3, 1.6, 1.7), однако здесь мы рассмотрим расположение в этих слоях микрофибрилл целлюлозы. В первичной оболочке мнкро-фибриллы целлюлозы расположены беспорядочно и образуют характерную для первичной оболочки дисперсную текстуру. Они способны смещаться каждая в отдельности, не мешая друг другу и образуя многослойную сеть [8, с. 29]. Отмечается, что степень полимеризации и кристалличности целлюлозы в первичной оболочке гораздо меньше, чем во вторичной оболочке. Микрофибриллы во вторичной оболочке ориентированы в основном параллельно друг другу, что обусловливает наибольшее нх уплотнение и высокую механическую прочность растительного материала на разрыв. В слое 5[ направление фибрилл почти перпендикулярно оси клетки, в слое они образуют с осью клетки острый (5—30°) угол. [c.13]
ниже, табл. 1.3, 1.6, 1.7), однако здесь мы рассмотрим расположение в этих слоях микрофибрилл целлюлозы. В первичной оболочке мнкро-фибриллы целлюлозы расположены беспорядочно и образуют характерную для первичной оболочки дисперсную текстуру. Они способны смещаться каждая в отдельности, не мешая друг другу и образуя многослойную сеть [8, с. 29]. Отмечается, что степень полимеризации и кристалличности целлюлозы в первичной оболочке гораздо меньше, чем во вторичной оболочке. Микрофибриллы во вторичной оболочке ориентированы в основном параллельно друг другу, что обусловливает наибольшее нх уплотнение и высокую механическую прочность растительного материала на разрыв. В слое 5[ направление фибрилл почти перпендикулярно оси клетки, в слое они образуют с осью клетки острый (5—30°) угол. [c.13]
Образуются также ковалентные, нековалентные и водородные связи. ГМЦ вступают в связь с полисахаридами на поверхности микрофибрилл целлюлозы, с протеинами и предшественниками лигнина. Характер этих связей зависит от локализации ГЛ 1Ц в клеточной стенке, что более подробно рассмотрим далее. [c.27]
Характер этих связей зависит от локализации ГЛ 1Ц в клеточной стенке, что более подробно рассмотрим далее. [c.27]
С СП 130—180 осах[c.304]
ЛОЗЫ, и связь между волокнами становится прочнее. В этих процессах участвуют и микрофибриллы целлюлозы [315]. [c.398]
Таким образом, сам факт обнаружения положительного двойного лучепреломления в растущей клеточной пластинке является веским доказательством присутствия в ней микрофибрилл целлюлозы. Точный химический состав клеточных пластинок, однако, остается неизвестным, так как изолировать их пока не удалось. [c.88]
Ф и г. 41. Схематическое изображение различных типов укладки микрофибрилл целлюлозы. Максимальный показатель преломления (п ,) соответствует направлению преимущественной ориентации микрофибрилл, а меньшие показатели преломления (п ) — взаимноперпендикулярным направлениям в плоскости, перпендикулярной [c.89]
Микрофибриллы целлюлозы окружены чрезвычайно густым аморфным полисахаридным гелем. Простые расчеты показывают, что концентрация полисахаридов в стенке составляет как минимум 10—20% по весу. Так как 1%-ный агаровый гель уже представляет собой твердую массу, то 10—20%-ный гель должен, по-видимому, быть совершенно непроницаемым, однако мы знаем, что это не так. [c.91]
Простые расчеты показывают, что концентрация полисахаридов в стенке составляет как минимум 10—20% по весу. Так как 1%-ный агаровый гель уже представляет собой твердую массу, то 10—20%-ный гель должен, по-видимому, быть совершенно непроницаемым, однако мы знаем, что это не так. [c.91]
В последнем слое вторичной стенки 83 микрофибриллы целлюлозы обычно расположены в виде плоской спирали, как в слое 8]. Эта стенка называется иногда третичной, она имеет небольшую толщину и ее внутренняя поверхность, направленная в сторону клеточной полости, иногда покрыта наплывами. Микрофйбриллы в этом слое расположены параллельно и не так плотно, как в слое 82, и угол, который они образуют с осью волокна, изменяется в широких пределах 40].—————————————— [c.319]
Рис 1 3 Модель микрофибриллы целлюлозы (по Фрей ВяееЛМРу) а — общий вид, 6 — поперечный разрез [c.
 11]
11]Стенкн сосудов и паренхимных клеток по ориентации микрофибрилл несколько отличаются от стенок волокон. Определенные особенности в ориентации целлюлозных микрофибрилл характерны для клеточных стенок реактивной древесины. В стенках трахеид сжатой древесины в слое 82 угол ориентации близок к 45°, т.е. намного больше, чем у нормальной древесины. В стенках волокон тяговой древесины в О-слое микрофибриллы ориентированы почти параллельно оси волокна. Изучение окаймленных пор показало, что в торусах мембран наблюдается кольцевая ориентация микрофибрилл целлюлозы, а в окружающей торус маргинальной (краевой) зоне мембраны тяжи микрофибрилл ориентированы радиально и удерживают торус. В заболонной древесине торусы не лигнифицнрованы отложение в них лигнина происходит при образовании ядровой или спелой древесины. [c.222]
В холоцеллюлозе к трудногидролизуемым полисахаридам относится целлюлоза как кристаллический полимер, а легкогидролизуемыми полисахаридами являются гемицеллюлозы как аморфные полимеры. Разбавленные кислоты способны проникать только в аморфные полисахариды, входящие в состав лигноуглеводной матрицы и в аморфную часть целлюлозы, также легкогидролизующуюся. К трудногидролизуемым полисахаридам относятся и целлюлозаны — гемицеллюлозы, совместно закристаллизованные с целлюлозой в паракристаллической части ее микрофибрилл. Целлюлозаны — это те же самые гемицеллюлозы, т.е. ксиланы и маннаны, но гидролизующиеся только вместе с целлюлозой и труднее извлекаемые щелочами. Поэтому в технической целлюлозе, выделенной из древесины любым варочным процессом, а также в альфа-целлюлозе, выделенной из холоцеллюлозы, всегда остается примесь остаточных гемицеллюлоз. Состав холоцеллюлозы с учетом различной гидролизуемости полисахаридов показан в виде схемы на рис. 11.3. [c.285]
Разбавленные кислоты способны проникать только в аморфные полисахариды, входящие в состав лигноуглеводной матрицы и в аморфную часть целлюлозы, также легкогидролизующуюся. К трудногидролизуемым полисахаридам относятся и целлюлозаны — гемицеллюлозы, совместно закристаллизованные с целлюлозой в паракристаллической части ее микрофибрилл. Целлюлозаны — это те же самые гемицеллюлозы, т.е. ксиланы и маннаны, но гидролизующиеся только вместе с целлюлозой и труднее извлекаемые щелочами. Поэтому в технической целлюлозе, выделенной из древесины любым варочным процессом, а также в альфа-целлюлозе, выделенной из холоцеллюлозы, всегда остается примесь остаточных гемицеллюлоз. Состав холоцеллюлозы с учетом различной гидролизуемости полисахаридов показан в виде схемы на рис. 11.3. [c.285]
Первичная стенка. Это наружный слой, который образовался на стадии деления живых клеток. Он очень тонок, состоит из редкой сетки микрофибрилл целлюлозы, находящихся в аморфном веществе, состоящем, вероятно, из пептина и гемицеллюлоз.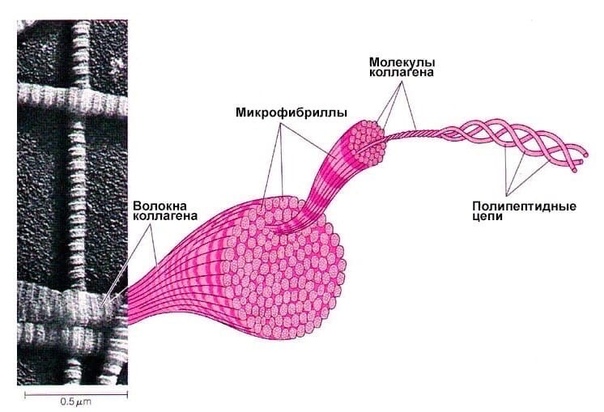 Целлюлоза имеет два уровня надмолекулярных образований элементарные фибриллы, поперечный размер которых 6,0-7,0 нм [57] и микрофибриллы, образованные несколькими элементарными фибриллами, диаметр которых колеблется от 10 до 40 нм [58]. Содержание целлюлозы в первичной стенке менее 12 %. [c.280]
Целлюлоза имеет два уровня надмолекулярных образований элементарные фибриллы, поперечный размер которых 6,0-7,0 нм [57] и микрофибриллы, образованные несколькими элементарными фибриллами, диаметр которых колеблется от 10 до 40 нм [58]. Содержание целлюлозы в первичной стенке менее 12 %. [c.280]
В некоторых случаях возможно и большее число стадий ферментативного-расщепления. Так, при гидролизе целлюлозы различают С -цел-люлазы, катализирующие расщепление микрофибрилл целлюлозы и переход ее в растворимое состояние, Сд.-целлюлазы, превращающие целлюлозу в смесь целлодекстринов, содержащих от двух до пяти моносахаридных остатков, и целлобиазу ф-глюкозидазу), катализирующую расщепление целлодекстринов до глюкозы. [c.619]
Колвин Дж Р Структура и образование целлюлозных микрофибрилл // Целлюлоза и ее производные / Ред Н Байклз, Л Сегал М, 1974 Т 2 С 20-46 [c.32]
Снижение среднего размера пор после ФГ целлюлозы осины до потери массы 0,6%, но мнению авторов [60], вызвано образованием микротрещин, которые открывают поры в клеточной оболочке, возникшие при химической обработке. Под сканирующим микроскопом наблюдается фибрилляция волокон, вызванная ФГ, Предполагается, что ксилан, покрывающий микрофибриллы целлюлозы, гидролизуется в большей степени, чем это можно обнаружить по количеству растворившихся при ФГ ксилозы и олигосахаридов кснланового ряда. Возможно, что ксилан гидролизуется частично и притом остается в клеточной стенке волокна. Так как ксилан, окружающий микрофибриллы целлюлозы, определяет силы когезии между волокнами, то частичный гидролиз его влияет на бумагообразующие свойства целлюлозной массы. [c.235]
Под сканирующим микроскопом наблюдается фибрилляция волокон, вызванная ФГ, Предполагается, что ксилан, покрывающий микрофибриллы целлюлозы, гидролизуется в большей степени, чем это можно обнаружить по количеству растворившихся при ФГ ксилозы и олигосахаридов кснланового ряда. Возможно, что ксилан гидролизуется частично и притом остается в клеточной стенке волокна. Так как ксилан, окружающий микрофибриллы целлюлозы, определяет силы когезии между волокнами, то частичный гидролиз его влияет на бумагообразующие свойства целлюлозной массы. [c.235]
Интересно, что механизм синтеза ГМЦ сугцественно отличается от синтеза целлюлозы. Последний протекает на плазмалемме [34] по механизму, обеспечивающему отложение целлюлозы в виде микрофибрилл определенного размера путем аппозиции. На поверхности илазмалеммы обнаружены глобулы, которые получили название терминальных комплексов синтеза микрофибрилл целлюлозы. Эти комплексы связаны с растущими концами микрофибрилл целлюлозы и дают начало одной микрофибрилле целлюлозы.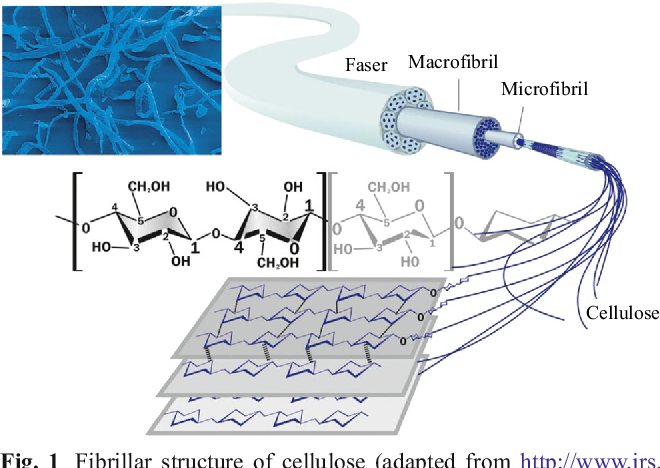 [c.26]
[c.26]
На рис. 1.2 показана схема расположения химических компонентов в первичной оболочке, предложенная Алберсхеймом [25]. Согласно эггой схеме, микрофибриллы целлюлозы покрыты моно-молекулярным слоем ксилоглюкана. Глюкозная часть молекулы ксилоглюкана расположена параллельно молекулам целлюлозы и образует с ней довольно стабильные водородные связи. [c.30]
На рис. 3.1 показано, что молекулы лигнина или части макромолекул этого полимера занимают пространство между микрофибриллами целлюлозы, но не касаются их поверхности, так как поверхность фибрилл покрыта СоЮем ГМЦ. [c.149]
Интересным свойством маннана можно считать образование кристаллитов на поверхности микрофибрилл целлюлозы [12]. При этом образуется так называемая структура шиш-кебаб (см. рис. 3.2), в которой микрофибрилла целлюлоз имеет вид нити, на которую нанизаны перпендикулярно ориентированные слоистые кристаллы маннана размером около 1000 А. Таким образом, ка поверхности микрофибрилл целлюлозы I находятся многочисленные центры кристаллизации, способствующие образованию кристаллов маннана.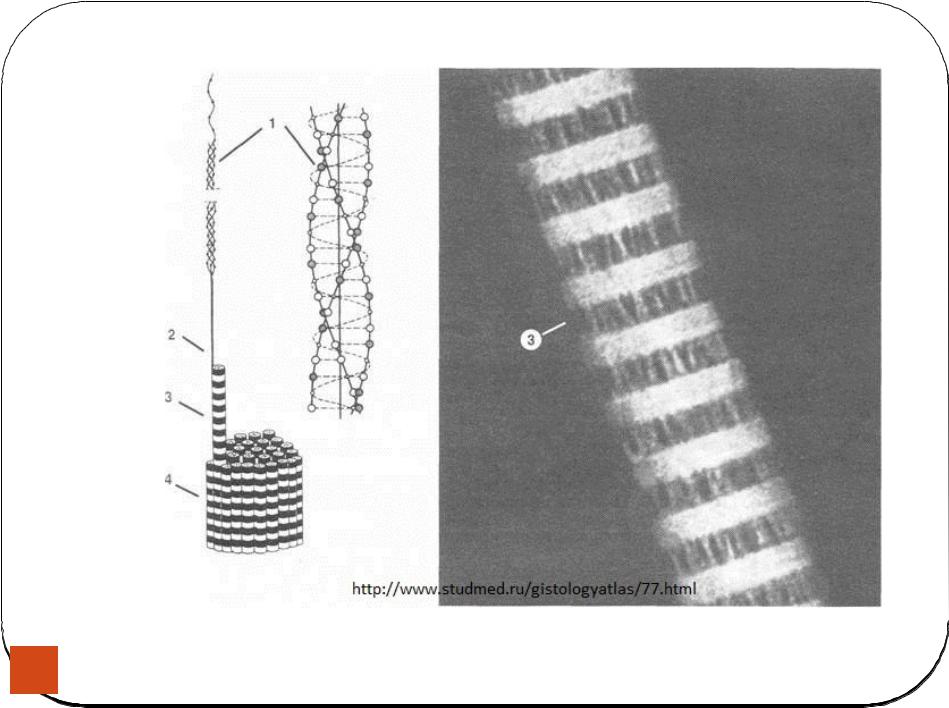 На этих активных центрах из-за сферических факторов кристаллы расположены параллельно друг к другу. Предполагается, что целлюлоза способна ориентировать маннан во время биосинтеза [13]. Небольшую примесь маннозы в гидролизатах -целлюлозы можно объяснить не включением молекул маннозы в молекулы полиглюкана, а образованием кристаллов маннана на поверхности фибрилл. [c.158]
На этих активных центрах из-за сферических факторов кристаллы расположены параллельно друг к другу. Предполагается, что целлюлоза способна ориентировать маннан во время биосинтеза [13]. Небольшую примесь маннозы в гидролизатах -целлюлозы можно объяснить не включением молекул маннозы в молекулы полиглюкана, а образованием кристаллов маннана на поверхности фибрилл. [c.158]
Высказывалось иредиоложение [366], что при кислой сульфитной варке деградация линейных фрагментов ГМЦ происходит с такой скоростью, что они успевают достичь микрофибрилл целлюлозы и сорбироваться на них до того, как их длина станет меньше необходимой для адсорбции. Прн двухступенчатых варках первая нейтральная или щелочная ступень, возможно, регулирует скорость последующего гидролиза. [c.307]
Мора и соавт. [678, 679] исходят из того, что ксилан, расположенный между микрофибриллами целлюлозы, играет вансную-роль в когезии клеточной стенки. Глюкоманнан более тесно ассоциирован с целлюлозой, чем ксилан.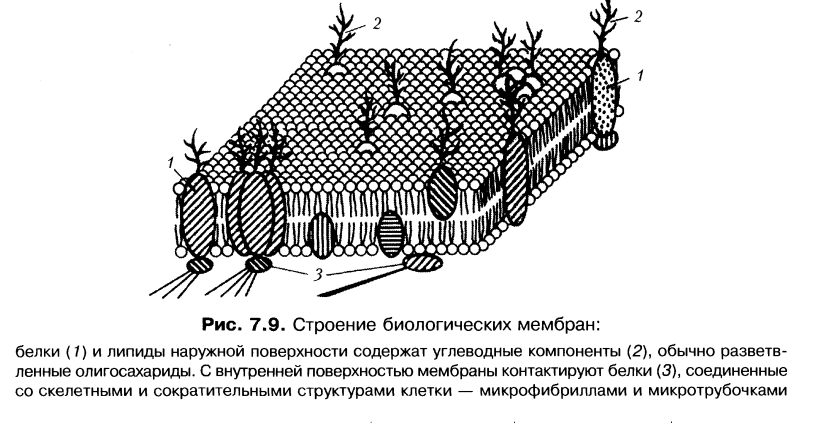 [c.381]
[c.381]
Располагаясь между микрофибриллами илп отдельными пучками макромолекул целлюлозы, ГМЦ, как предиолагается, способствуют отрыву части микрофибрилл целлюлозы от поверхности волокон в ироцессе размола и вызывают их быструю гидратацию. В результате иоверхность волокон покрывается слизью, сиособствующей склеиванию отдельных волокон и образованию тяжей между ними в ироцессе сушки бумаги [286, 315]. [c.385]
Тщательный анализ причин возникновения зернистости на снимках микрофибрилл целлюлозы проведен в работе [52]. В микроскопе с высоким разрешением исследовали ультратон-кие срезы, подвергнутые негативному контрастированию. Автор [c.104]
Действительно ли микротрубочки имеют прямое отношение к синтезу микрофибрилл целлюлозы, неизвестно, но сходство ориентации этих элементов заставляет призадуматься. Эльбейну и сотр. [6] удалось синтезировать целлюлозу с помощью фермента, локализованного в частицах, оседающих при центрифугировании с фракцией митохондрий. Это свидетельствует о том, что целлюлоза образуется в цитоплазме и притом, по-видимому, не в микротрубочках, хотя и эта возможность не может быть полностью отброшена. Аппарат Гольджи также следует принимать во внимание как возможное место синтеза полисахаридов [20]. [c.90]
Это свидетельствует о том, что целлюлоза образуется в цитоплазме и притом, по-видимому, не в микротрубочках, хотя и эта возможность не может быть полностью отброшена. Аппарат Гольджи также следует принимать во внимание как возможное место синтеза полисахаридов [20]. [c.90]
Имеются данные, говорящие о том, что микрофибриллы целлюлозы удлиняются путем нарастания с конца. Однако остается совершенно неясным, каким образом синтезирующие клетчатку цитоплазматические ферменты катализируют концевой рост микрофибрилл в клеточной оболочке. Возможно, частицы Эльбей-на локализованы в оболочке. Трудно также объяснить, как микротрубочки кортекса могут направлять ориентированный синтез микрофибрилл оболочки ведь эти системы, хотя они и расположены сходным образом, разделены плазматической мембраной. Еще труднее понять, как кортикальные микротрубочки могут [c.90]
Вторичные клеточные оболочки, как правило, значительно толще первичных. Они характеризуются отложением гораздо больших количеств целлюлозы и значительных количеств лигнина (о химии лигнина см. гл. 23). Полагают, что лигнин откладывается поверх микрофибрилл целлюлозы, причем этот процесс (одревеснение) начинается в области срединной пластинки и затем постененно распространяется в направлении клеточной мембраны. [c.92]
Лекция 3. Строение П.А.К. — BioBox All Secrets
ПАК – это субмембранная система клетки, в которой можно выделить: наружную мембрану или плазмалемму, внутреннею мембрану или гликокаликс, субмембранный опорно-сократительный аппарат.
ПЛАЗМАЛЕММА.
занимает центральную часть ПАКа, построена по типичной жидкостно мозаичной модели.
ГЛИКОКАЛИКС.
В основном включает в себя углеводный компонент: полисахариды, олигосахариды, гликопротеины, гликолипиды, а также наружные домены интегральных и полуинтегральных и периферических белков. Гликокаликс выполняет маркерную функцию, функцию индивидуализации, а также способен участвовать в образование клеточных контактов, кроме того, как производное гликокаликса формируется клеточная стенка у растений, а также белки соединительной ткани, например, коллаген и эластин. Гликокаликс может выполнять ферментативные функции, примером является фермент гидролаза, встроенная в гликокаликс, которая участвует в процессах пристеночного пищеварения.
СОСА.
СОСА включает в себя:
-
периферическую геалоплазму
-
белки опорно-сократительной системы
периферическая гиалоплазма отличается от основной определенной концентрацией ферментативных комплексов. Здесь располагаются белки и ферменты, которые способствуют транспорту через мембрану, здесь также локализована фермент аденилатциклаза, которая участвует в работе рецепторных систем вторичных посредников, здесь локализованы ферменты гликолиза. Кроме того, в периферической гиалоплазме находятся секреторные гранулы, которые способны выводится из клетки после поступления из клетки сигнала.
к белкам опорно-сократительной системы относятся:
-
тонкие фибриллы
-
микрофибриллы
-
скелетные фибриллы
-
микротрубочки
Тонкие фибриллы.
Первичная структура белков неизвестна, функции неизвестны. Однако выяснено, что тонкие филоменты способны делить клетку на функциональные отсеки или компартменты.
К тонким фибриллам могут прикрепляться ферментативные комплексы, различные органоиды (рибосомы, митохондрии)
Микрофибриллы.
Микрофибриллы состоят из белка актина и имеют диаметр примерно 5-7 нм. В клетке актин может существовать в двух вариантах: глобулярный актин или G-актин, фибриллярный или F-актин. При определенных условиях G-актин может полимеризоваться, и приобретать вид двойной спирали.
F-актин нестабилен и его структура стабилизируется другим белком тропомиоезином (это фибриллярный белок, который укладывается на структуру α-спирали и стабилизирует ее. Актиновые фибриллы способны не только к сборке, но и к разборке с разных концов молекул. Сборка и разборка контролируется определенными белками, которые называются кэп-белками. Актиновые фибриллы могут сшиваться и взаимодействовать между собой при помощи определенных белков. Сшивание молекул может происходит по середине или концами актиновых фибрилл, в результате образуется пучок микрофибрилл, который в зависимости от количества сшивающих белков может быть рыхлым или плотным. Кроме того к актиновым фибриллам могут якорные белки, при помощи которых фибриллы взаимодействуют с белками плазмолеммы, частично ограничивая их подвижность. в клетке актиновые фибриллы взаимодействуют с белком миозином. Причем выделяется одноголовочный и двуголовочный миозин. Одноголовочный встречается редко , в основном в кардиомиоцитах. Двуголовочный характерен для всех остальных клеток.
Головки обладают АТФ-азной активностью, это значит, что при расщеплении молекул АТФ головки могут менять свою конформацию. Расщепление молекул АТФ проходит в несколько этапов, причем конформационные изменения головки способны вызывать движения в шарнирной части, если молекула миозина прикреплена головкой к молекуле актина, то движение головки вызывает движение молекулы миозина по актину.
Головка миозина содержит в себе центр связывающий АТФ, а также несколько центров для взаимодействия с актиновыми фибриллами. Находясь в связанном с АТФ состоянии головка миозина не обладает сродством к F-актину. Гидролиз АТФ приводит к тому, что в АТФ-азном центре оказывается комплекс АДФ+Fн. Образование этого комплекса так изменяет конформацию головки, что она способна связаться с фибриллярным актином. Эта связь приводит к дальнейшему изменению конформации головки, в результате чего АДФ и неорганический фосфат уходят из АТФ-азного центра. В этот момент наблюдается смещение головки миозина относительно ее стержня, и головка продвигается по F-актину.
В норме в клетки в состав актомиозиновой системы входят не отдельные молекулы миозина, а их функциональные объединения, которые получили название биполярные миозиновые филаменты. В этом случае молекула миозина своими головками прикреплена к разным нитям фибриллярного актина, а хвосты миозина сцеплены.
Особенно сильно АМС развита в клетках поперечноисчерченой мускулатуры. Единицей строения клетки скелетной мускулатуры является сакромер, который ограничен белковыми полосками.
АМС участвует в образование на клетки временных и постоянных выростов. К временным выростам относятся псевдоподии или ложноножки, которые характерны для защитных клеток организма фагоцитов. К постоянным относятся микроворсинки, которые возникают в тонком кишечнике.
Актиновые фибриллы образуют стресс-фибриллы, которые способны удерживать форму клетки при меняющимся астматическом давлении. При делении клеток АМС участвует в формировании пояска деления. Поясок деления необходим для деления цитоплазмы.
Патологии.
Обнаружены химические вещества, которые способны влиять на полимеризацию микрофибрилл, например, метаболиты некоторых грибов – цитохалазины способны присоединяться к молекулам G-актина. Такой комплекс присоединяется к концу микрофибриллы и препятствует ее полимеризации, что приводит к разрушению микрофибриллы. Токсин бледной поганки фаллоидин взаимодействует сразу с F-актином вызывая его суперстабилизацию. В этом случае F-актин теряет способность к внутриклеточным перестройкам.
Встречаются наследственные патологии, которые обусловлены дефектами актин-связывающих белков, к такой патологии относятся один из видов миодистрофии – миодистрофия Дюшена. Причиной является патология гена локализованного в Х-хромосоме, поэтому данное заболевание проявляется в основном у мальчиков. Это мышечная слабость, причем к 8-13 годам они теряют способность ходить, погибают от остановки дыхания.
Скелетные фибриллы.
Является универсальным элементом СОСА и представляет собой белковые нити диаметром около 10нм. Характеризуются повышенной устойчивостью к действию химических и физических факторов, поэтому основной функцией является структурная и опорная. Участвуют в формировании цитоскелета клетки, способны поддерживать определенные части клетки, например, длинные отростки нейрона. В большом количестве встречаются в зоне контактов между клетками. Скелетные фибриллы представляют собой белки различной первичной структуры, но одинаковой третичной и четвертичной. На третичном уровне организации белок скелетные фибриллы представляет собой гомотетромер. Скелетные фибриллы образуются за счет взаимодействия нескольких гомотетромеров между собой по принципу кирпичной кладки.
Такая структура в длину может расти до бесконечности (в зависимости от размеров клетки), а в ширину не более чем до 8 протофибрилл. Примером белков скелетных фибрилл является белки кератины, которые особенно в большом количестве встречаются в наружных слоях эпидермиса и производных эпидермиса. Увеличение скелетных фибрилл в клетки приводит к нарушению клеточных функций и, следовательно, к различным патологиям, например, в сердечной мышцы к различным кардиомиодистрофиям, увеличение фибрилл в нейронах головного мозга приводит к различным формам старческого слабоумия. Увеличение скелетных фибрилл может происходить под действием различных химических факторов, например, алкоголя. Увеличение скелетных фибрилл в клетках печени приводит к циррозу печени у алкоголиков. Увеличение скелетных фибрилл в нейронах приводит к деградации личности. Первичная структура скелетных фибрилл в клетках различных тканей различна, поэтому скелетные фибриллы можно использовать для диагностики опухолей и на основании диагностики делать вывод является ли данная опухоль первичной, либо это метастаза.
Микротрубочки.
Это полые белковые структуры диаметром 22-25нм и шириной примерно 6нм.
Микротрубочки состоят из белков тубулинов. Как правило, с микротрубочками связаны так называемые ассоциативные белки или МАР белки.
В клетке встречаются 3 вида тубулинов α, β, γ, причем основная масса примерно 99% приходится на α и β тубулин. Γ-тубулин в составе микротрубочек не встречается, он располагается в клеточном центре и образует ЦОМТ. Считается, что γ-тубулин необходим для начала сборки микротрубочек. Тубулины являются ГТФ-связывающими белками, поэтому в присутствии ГТФ и ионов магния α и β тубулины активируются и способны объединятся в стабильные гетеродимеры. Затем в ЦОМТах при наличии ГТФ и магния начинается сборка гетеродимеров, которые реагируют между собой с образованием протофибрилл. Формируется так называемый тубулиновый коврик, который в длину может расти до бесконечности, а в ширину не более чем до 13 протофибрилл. После этого края коврика слипаются, и он превращается в полую трубочку. В микротрубочке выделяют + и — концы, на + конце в основном идет полимеризация микротрубочек, на –конце в основном деполимеризация. Этот процесс (сборки и разборки) регулируется специальными белками и в норме в клетки процесс сборки преобладает над процессом разборки. Сборка и разборка микротрубочек зависит от концентрации различных веществ, например, ионов кальция. При избыточной концентрации кальция в клетки процесс деполимеризации микротрубочек активируется. В настоящее время считают, что количество кальция в клетке является основным механизмом, определяющим процессы перестройки микротрубочковых систем. При действии алкоголя и растительного алкалоида колхицина микротрубочки разрушаются.
Основными функциями микротрубочек является структурная и опорная функции. Существуют структуры, в которых цитоскелет образуется за счет микротрубочек, например, тромбоциты. Кроме того микротрубочки объединяются с белками МАР и могут участвовать в выполнении многих важных функций в клетке. МАР-белки способны регулировать сборку и разборку микротрубочек, в частности в соединении с некоторыми МАР-белками микротрубочки приобретают устойчивость к действию ионов кальция и низких температур. Важнейшими среди МАР-белков являются белки транслокаторы, которые вместе с микротрубочками образуют тубулин транслокаторную систему клетки. Транслокаторы по строению сходны с миозином. В них выделяют стержень и головку, причем головка обладает АТФ-азной активностью, следовательно, при изменении конформации транслокатор способен перемещаться по микротрубочке.
Выделяют 3 группы транслокаторов:
-
кинезины
-
динеины
-
динамины
кинезины способны перемещаться по микротрубочке только от – к + концу, поэтому они обеспечивают транспорт веществ от центра клеток к ПАКу, это так называемый антреградный транспорт. В данном случае транслокатор движется по микротрубочке за счет конформационные изменений головки, а к стержню прикреплен немембранный пузырек с упакованным веществом.
С помощью данного вида транспорта транспортируются медиаторы в нейроны, а также пигменты в пигментных клетках
Динеины участвуют в формировании тубулин-динеиновой системы, которая осуществляет ретроградный транспорт. Помимо этого динеины участвую в образовании локомоторных структур клеток: ресничек и жгутиков.
Динамины обладают ГТФ-азной активностью, обеспечивают ретроградный транспорт. Функции динаминов изучены плохо.
Важнейшей функцией тубулин-транслокаторной системы является формирование двигательных систем в клетке, которые у всех эукориот построены по единой схеме.
Существуют наследственные патологии, связанные с нарушением структуры тубулина. Они практически не совместимы с жизнью, однако существует патологии тубулин-денеиновой системы, которые касаются непосредственно жгутиков и ресничек. При этом нарушается структура ресничного эпителия, выстилающего носоглотку, дыхательные пути и полость среднего уха, что приводит к хроническим бронхитам и отитам мужчины с синдромом неподвижных ресничек, как правило, стерильны.
Все компоненты СОСА характеризуются структурным единством, которое проявляется в том случае, что фибриллярные белки могут взаимодействовать друг с другом. Например, микрофибриллы могут образовывать пучки, которые могут взаимодействовать с микротрубочками и скелетными фибриллами. Все компоненты СОСА способны взаимодействовать с интегральными белками плазмолеммы, а плазмолемма структурно связана с гликокаликсом, поэтому для ПАКа характерно структурное единство.
Структура микрофибрилл целлюлозы в первичных клеточных стенках колленхимы | Физиология растений
Аннотация
В первичных стенках растущих клеток растений глюкозно-полимерная целлюлоза собрана в длинные микрофибриллы диаметром несколько нанометров. Жесткость и ориентация этих микрофибрилл контролируют рост клеток; Следовательно, синтез целлюлозы является ключевым фактором роста и морфогенеза растений. Колленхима сельдерея ( Apium graveolens ) является полезной модельной системой для изучения структуры микрофибрилл первичной стенки, поскольку ее микрофибриллы ориентированы с необычной однородностью, что облегчает проведение спектроскопических и дифракционных экспериментов.Используя комбинацию методов рассеяния рентгеновских лучей и нейтронов с колебательной спектроскопией и спектроскопией ядерного магнитного резонанса, мы показываем, что микрофибриллы колленхимы сельдерея имели средний диаметр от 2,9 до 3,0 нм, с наиболее вероятной структурой, содержащей 24 цепи в поперечном сечении, расположенные в восьми атомах водорода. -связанные листы из трех цепей с обширным беспорядком в боковой упаковке, конформации и водородной связи. Аналогичная структура из 18 цепей и структуры из 24 цепей разной формы хуже соответствовали данным.Конформационный беспорядок в основном ограничивался поверхностными цепями, а беспорядок в упаковке цепей — нет. То есть по положению и ориентации поверхностные цепи соответствовали неупорядоченной решетке, составляющей ядро каждой микрофибриллы. Были доказательства того, что соседние микрофибриллы нековалентно агрегированы вместе на части своей длины, предполагая, что необходимость разрушения этих агрегатов может быть сдерживающим фактором роста и гидролиза целлюлозы для производства биотоплива.
Рост и форма растений контролируются точно ориентированным расширением стенок отдельных клеток. Движущая сила для размножения клеток является осмотической, но скорость и направление экспансии контролируются механическими свойствами клеточной стенки (Szymanski and Cosgrove, 2009). Расширяющиеся первичные клеточные стенки представляют собой нанокомпозитные материалы, в которых длинные микрофибриллы целлюлозы диаметром несколько нанометров проходят через гидратированную матрицу ксилоглюканов, пектинов и других полимеров (Knox, 2008; Mohnen, 2008; Szymanski and Cosgrove, 2009; Scheller. и Ульвсков, 2010).Нативные микрофибриллы целлюлозы частично кристаллические (Nishiyama, 2009; Fernandes et al., 2011). Раньше считалось, что целлюлоза первичной стенки имеет уникальную кристаллическую структуру, называемую целлюлозой IV 1 (Dinand et al., 1996), но данные ЯМР предполагают наличие форм, подобных лучше охарактеризованным кристаллическим формам целлюлозы Iα и Iβ вместе с крупными кристаллическими формами целлюлозы. количества менее упорядоченной целлюлозы (Wickholm et al., 1998; Sturcová et al., 2004; Wada et al., 2004). Тем не менее, целлюлоза гораздо более упорядочена, чем любой другой компонент первичной клеточной стенки (Bootten et al., 2004), в соответствии с его ключевой ролью обеспечения силы и контроля роста.
Жесткость клеточной стенки наибольшая в направлении микрофибрилл целлюлозы, где рост является направленным, а преобладающая ориентация микрофибрилл обычно поперечна направлению роста (Green, 1999; MacKinnon et al., 2006; Szymanski and Cosgrove, 2009 г.). Расширение клеточной стенки требует либо увеличения расстояния между микрофибриллами (Marga et al., 2005), либо проскальзывания между ними (Cosgrove, 2005), либо того и другого, и микрофибриллы переориентируются в направлении роста (Anderson et al., 2010). Полимерные поперечные мостики между микрофибриллами (McCann et al., 1990), как полагают, противостоят этим деформациям наноструктуры клеточной стенки и, таким образом, контролируют скорость роста. До недавнего времени наибольшее внимание уделялось мостиковым соединениям ксилоглюканов, водородным связям с поверхностями микрофибрилл (Scheller, Ulvskov, 2010). Однако есть свидетельства того, что не все ксилоглюканы расположены надлежащим образом (Fujino et al., 2000; Park and Cosgrove, 2012a) и что могут быть задействованы другие мостиковые полимеры (Zykwinska et al., 2007). Также было высказано предположение, что пучки агрегированных микрофибрилл, а не отдельные микрофибриллы, могут быть ключевыми структурными единицами в первичных клеточных стенках (Anderson et al., 2010), как в древесине (Fahlén and Salmén, 2005; Fernandes et al., 2011). ). В таком случае отдельные микрофибриллы могут соединяться между пучками микрофибрилл. Таким образом, рост растительных клеток не совсем понятен, и нам нужна дополнительная информация о том, как контролируется ориентация целлюлозы, и о природе мостиковых полимеров, о поверхностях целлюлозы, с которыми эти полимеры связываются, и о сцеплении между поверхностями микрофибрилл, которые могут опосредовать агрегацию.
Микрофибриллы целлюлозы синтезируются на поверхности клетки большими ферментными комплексами, имеющими гексагональную симметрию, иногда называемыми «розетками» (Somerville, 2006). Каждый комплекс содержит несколько синтаз целлюлозы, которые различаются между первичными клеточными стенками и древесиной, хотя внешний вид комплексов похож (Somerville, 2006; Atanassov et al., 2009). Одновременный синтез с одного и того же конца всех цепей в микрофибрилле нативной целлюлозы является причиной их параллельности (Nishiyama et al., 2002, 2003), в отличие от энтропийно предпочтительной антипараллельной структуры, обнаруженной в искусственных целлюлозах, таких как вискоза (Langan et al., 2001). Очевидно, что количество цепей в микрофибрилле и количество целлюлозосинтаз в синтетическом комплексе взаимосвязаны. Принято считать, что количество цепочек делится на шесть, что соответствует симметрии гексагональной розетки и 36-цепочечным моделям (Himmel et al., 2007), ограниченным гидрофильными гранями кристаллов [110] и [1-10], поскольку в водорослевых целлюлозах (Bergenstråhle et al., 2008), получили широкое распространение. Сборка и ориентация целлюлозы взаимосвязаны, поскольку некоторые мутанты целлюлозосинтазы имеют фенотипы, дефектные по ориентации целлюлозы и растительной форме, а также обедненные содержанием целлюлозы (Paredez et al., 2008). У некоторых других мутантных линий, по-видимому, затрагивается кристалличность микрофибрилл (Fujita et al., 2011; Harris et al., 2012; Sánchez-Rodríguez et al., 2012).
Следовательно, детальное понимание структуры микрофибрилл целлюлозы первичной стенки поможет нам понять синтез целлюлозы, а также рост и структурную механику живых растений (Burgert and Fratzl, 2009).Первичные клеточные стенки и их целлюлозные скелеты также влияют на такие характеристики качества пищевых продуктов, как хрусткость салата из овощей и яблок ( Malus domestica ; Jarvis, 2011). Когда биотопливо производится из лигноцеллюлозной биомассы, лигнификация приводит к устойчивости (Himmel et al., 2007), но некоторые типы клеток у видов Miscanthus , просо ( Panicum virgatum ) и пожнивные остатки имеют только первичные стенки. без лигнина, и сопротивляемость этому зависит от природы микрофибрилл целлюлозы (Beckham et al., 2011).
Относительно подробная структура недавно была предложена для микрофибрилл древесины ели ( Picea spp.) (Fernandes et al., 2011), которые имеют диаметр 3,0 нм, что позволяет разместить только около 24 цепочек целлюлозы. Данные рентгеновской дифракции подтверждают «прямоугольную» форму (Matthews et al., 2006), ограниченную гранями [010] и [200]. Наблюдался значительный беспорядок, увеличивающийся к поверхности, и микрофибриллы были агрегированы в пучки размером примерно 15-20 нм в поперечнике, причем некоторые, но не все, боковые границы раздела были устойчивы к воде (Fernandes et al., 2011). Неупорядоченные домены являются особенностью других прочных биологических материалов, таких как шелк паука (van Beek et al., 2002).
Следовательно, представляет интерес, могут ли какие-либо из этих свойств древесной целлюлозы также быть обнаружены в микрофибриллах целлюлозы первичных (растущих) клеточных стенок. Было бы особенно полезно охарактеризовать нарушение, которое, как известно, присутствует в микрофибриллах первичной стенки, то есть определить, чем целлюлоза, которая не определяется как «кристаллическая», отличается от кристаллической целлюлозы.Многие эксперименты, ведущие к созданию структуры древесной целлюлозы, зависели от исключительно однородной ориентации микрофибрилл целлюлозы (Sturcová et al., 2004; Fernandes et al., 2011). Однако в растущих клеточных стенках микрофибриллы ориентированы неравномерно. Когда микрофибриллы сначала откладываются на внутренней поверхности первичной клеточной стенки, их ориентация обычно перпендикулярна направлению роста, но когда клеточная стенка расширяется, микрофибриллы переориентируются таким образом, что распределение ориентации интегрируется по толщине расширенной клеточной стенки. клеточная стенка становится все более близкой к случайной (Cosgrove, 2005; MacKinnon et al., 2006).
Эта техническая проблема не относится к клеточным стенкам колленхимы сельдерея ( Apium graveolens ), которые похожи по составу на другие первичные клеточные стенки, но имеют их микрофибриллы, ориентированные относительно равномерно вдоль оси клетки (Sturcová et al., 2004 ; Kennedy et al., 2007a, 2007b). Некоторая структурная информация о целлюлозе колленхимы сельдерея уже была получена в результате спектроскопических экспериментов и экспериментов по рассеянию (Sturcová et al., 2004; Kennedy et al., 2007a, 2007b), подтверждая беспорядок, ожидаемый в целлюлозе первичной стенки.Некоторые из этих экспериментов были аналогичны тому, что было сделано с еловой целлюлозой (Fernandes et al., 2011), но недостаточно данных, чтобы указать количество цепей в каждой микрофибрилле первичной стенки, природу и расположение нарушения, а также наличие или отсутствие прямого контакта микрофибрилл. Здесь мы сообщаем об экспериментах по рассеянию рентгеновских лучей и нейтронов, а также спектроскопических экспериментах, направленных на решение этих вопросов и приводящих к предложенной структуре микрофибрилл целлюлозы первичной стенки. Характеристика структуры, содержащей столько беспорядка, представляла необычные проблемы, но беспорядок, по-видимому, является центральным для загадочной способности целлюлозы первичной стенки обеспечивать высокую прочность, но при этом позволять и контролировать рост.
РЕЗУЛЬТАТЫ
Диаметр микрофибриллы и расстояние от малоуглового рассеяния нейтронов
Когда микрофибриллы коллагена или целлюлозы фиксированного диаметра упакованы в массивы с некоторой степенью регулярности, может наблюдаться малоугловая дифракция рентгеновских лучей или нейтронов (брэгговское рассеяние) от самих микрофибрилл (Hulmes et al., 1995; Fernandes et al. др., 2011). Это рассеяние происходит под меньшими углами, чем обычная дифракция на кристаллических плоскостях, и накладывается на малоугловое рассеяние, возникающее не из-за дифракции, а из-за текстуры и формы фибрилл и пустот между ними (форм-фактор).Если упаковка не является достаточно регулярной, чтобы образовать кристаллическую сверхрешетку, наблюдается только один пик Брэгга, соответствующий расстоянию между центрами фибрилл (Hulmes et al., 1995; Kennedy et al., 2007a).
Используя малоугловое рассеяние рентгеновских лучей (SAXS), наблюдали брэгговское отражение от микрофибрилл целлюлозы внутри интактных клеточных стенок колленхимы сельдерея. Расстояние между микрофибриллами увеличивается при гидратации (Kennedy et al., 2007b). Моделирование этих результатов показало, что наблюдаемый пик Брэгга может быть вызван рядом комбинаций расстояния между микрофибриллами и их диаметра.Следовательно, невозможно однозначно определить диаметр микрофибрилл (Kennedy et al., 2007a).
В древесине ели микрофибриллы слишком тесно агрегированы, чтобы обеспечить адекватный контраст для малоуглового брэгговского рассеяния. Эту проблему удалось решить путем дейтерирования поверхности и использования нейтронов вместо рентгеновских лучей (Fernandes et al., 2011). Природа контраста различается между источниками рентгеновского излучения и нейтронов. Рентгеновский контраст зависит в основном от локальной электронной плотности, тогда как контраст рассеяния нейтронов зависит от локального элементного состава и плотности.Одним из преимуществ этого метода является очень разные сечения рассеяния нейтронов водородом и дейтерием, что позволяет ввести контраст, например, за счет дейтериевого обмена в оксиде дейтерия (D 2 O; Horikawa et al., 2009; Matthews и др., 2011).
Экваториальный пик Брэгга был хорошо виден на изображениях малоуглового рассеяния нейтронов (МУРН) древесины ели, насыщенной D 2 O (Fernandes et al., 2011). Интенсивность пика Брэгга уменьшалась, а его расстояние ( q ) от центра дифракционной картины увеличивалось при сушке до тех пор, пока на картине SANS от полностью сухой дейтерированной древесины пик не был слабо виден при q = 2.1 нм −1 . Низкий контраст в этих условиях соответствовал контакту между соседними дейтерированными на поверхности микрофибриллами, так что расстояние между центрами 3,0 нм соответствовало по существу среднему диаметру микрофибрилл (Fernandes et al., 2011).
Подобные эксперименты SANS были проведены на стенках клеток из колленхимы сельдерея и на целлюлозе, выделенной из этих клеточных стенок (рис. 1). Пик брэгговского рассеяния в области q = 1 нм -1 , наложенный на экспоненциальный фон от некогерентного рассеяния, был очевиден как для стенок клеток, так и для изолированной целлюлозы, насыщенной чистым D 2 O, чистой водой или ряд смесей (рис.1; Дополнительный рис. S1). В соответствии с рассеянием от целлюлозы, а не от других полимеров, не наблюдалось пика Брэгга для смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Которая соответствует теоретической плотности длины рассеяния. целлюлозы (рис. 1).
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна.C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл. D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив.Вертикальный масштаб больше, чем в C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна. C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл.D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив. Вертикальная шкала больше, чем C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
В отличие от древесины ели, сухая целлюлоза сельдерея дает слабый пик Брэгга без дейтерирования поверхности микрофибрилл. В сухом состоянии пик Брэгга был при q = 2.11 нм -1 для целлюлозы в клеточных стенках или q = 2,14 нм -1 для изолированной целлюлозы (рис. 1), что соответствует межцентровым расстояниям в реальном пространстве 3,0 и 2,9 нм, соответственно. . Интенсивность Брэгга была менее 1% от наблюдаемой после регидратации. Этот очень низкий контраст соответствовал плотной упаковке микрофибрилл, вероятно, без каких-либо других полимеров между ними по крайней мере на некоторой части их длины. Расстояние от центра до центра должно приблизительно соответствовать среднему диаметру микрофибриллы.Несмотря на свою низкую интенсивность, пик Брэгга был легко различим на картинах МУРН, поскольку было очень мало помех от некогерентного рассеяния (рис. 1).
В гидратированном состоянии значение q пика Брэгга от микрофибрилл in situ в стенках клеток будет соответствовать реальному расстоянию ( d ) 7,1 ± 1,3 нм -1 , в то время как q Значение пика Брэгга для гидратированной целлюлозы соответствует реальному интервалу 6.0 ± 1,5 нм -1 . Эти номинальные расстояния d не означают, что все микрофибриллы разделены до такой степени при гидратации. Гидратация также значительно увеличивала контраст SANS за счет введения воды или D 2 O между микрофибриллами и, в случае D 2 O, за счет дейтерирования их поверхностей и нецеллюлозных полимеров. Отсюда следует, что при любом распределении расстояний между микрофибриллами более широкие промежутки будут значительно перепредставлены в шаблоне SANS. Номинальные интервалы, наблюдаемые после гидратации, немного превышали расчетные (Kennedy et al., 2007b) методом SAXS, что согласуется с разной основой рентгеновского контраста, но не было никакой разницы между расстояниями, наблюдаемыми в сухом состоянии с рентгеновскими лучами (Kennedy et al., 2007a) и с нейтронами (рис. 1). . Хотя на основе рентгеновских данных невозможно было сделать вывод о том, что расстояние, наблюдаемое в сухом состоянии, соответствует микрофибриллам, находящимся в прямом контакте друг с другом, этот вывод кажется вероятным на основе объединенных данных SAXS и SANS. . Гидратация четко разделяла микрофибриллы, но, вероятно, в разной степени по их длине (Kennedy et al., 2007b), возможно, в зависимости от сродства нецеллюлозных полисахаридов к воде (Jarvis, 1992).
Размеры микрофибрилл и нарушения в результате широкоугольного рассеяния рентгеновских лучей
Широкоугольные картины рассеяния рентгеновских лучей или нейтронов от природной целлюлозы достаточно хорошо разрешены для кристаллографического решения структуры, только если целлюлоза исключительно упорядочена и хорошо ориентирована и если микрофибриллы достаточно толстые, чтобы минимизировать уширение Шеррера, которое возникает в результате ограниченное количество плоскостей решетки.Большая часть целлюлозы колленхимы сельдерея была относительно хорошо ориентирована внутри клеточных стенок (рис. 2). Вклад широкоугольного рентгеновского рассеяния (WAXS) от незначительной изотропно ориентированной фракции целлюлозы был удален на стадии вычитания фона. Существенный вклад изотропного рассеяния на связанной воде также был удален при вычитании фона. Рассеяние от воды было идентифицировано экспериментом, в котором менялось содержание воды (дополнительный рис. S2).
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок WAXS из хорошо ориентированного целлюлозного компонента был размыт в радиальном направлении из-за уширения Шеррера, уширения, связанного с беспорядком, или обоих. Отражение 200 было при q = 15,5 нм -1 , что соответствует несколько большему расстоянию между слоями, чем в кристаллических целлюлозах, и отражения 1-10 и 110 были объединены. Эти особенности наблюдались на плохо ориентированных дифрактограммах от того, что было названо целлюлозой IV 1 (Dinand et al., 1996) или неупорядоченную целлюлозу Iβ (Wada et al., 2004) в первичных клеточных стенках. Для единообразия в этой статье используется обозначение решетки Iβ, но без всякого указания на то, что эта форма представляет собой наиболее близкую аналогию со средней структурой.
Количественная оценка беспорядка и вычисление размеров Шеррера по уширению этих отражений было затруднено из-за сильного перекрытия между отражениями 1-10 и 110 и чрезвычайно низкой интенсивности отражения 400, качественного индикатора большего бокового беспорядка, чем в хвойная древесина.При длительном времени сбора данных (16–24 ч) можно было получить достаточное отношение сигнал / шум для оценки уширения отражения 400. Гидратация за счет увеличения моноклинного угла и, таким образом, разделения отражений 1-10 и 110 упростила моделирование соответствующей ширины этих двух отражений в дифракционной картине от древесины ели (Fernandes et al., 2011). Однако в целлюлозе сельдерея гидратация немного увеличивала интервал [200] d , но не увеличивала значительно угол моноклинности.Процедура Fernandes et al. (2011) был использован для выделения эффекта одной формы бокового беспорядка, которая приводит к асимметрии радиальных пиков. Предполагая, что оставшиеся симметричные радиальные ширины отражений 1-10 и 110 были равны, наиболее подходящее значение было на 22% больше, чем радиальная ширина отражения 200 при моноклинном угле 95 °. Исходя из этого, средние размеры элементарной ячейки в сухом состоянии составляли a = 0,81 нм, b = 0,80 нм, c = 1,03 нм (с сохранением индекса целлюлозы Iβ для ясности, даже если а> b).Меньшие углы моноклинности и большая ширина, или наоборот, почти так же точно соответствовали наблюдаемым профилям интенсивности.
Еще одна форма беспорядка, которая, наряду с небольшими размерами решетки, способствует радиальному уширению, была смоделирована как «паракристаллическая» (Fernandes et al., 2011), что означает, что ширина отражений увеличивается с квадратом порядка отражения. или с q 2 d . Используя наиболее подходящие данные и построив график ширины δ q отражений против q 2 d , как описано (Fernandes et al., 2011), ширина симметричных компонент рефлексов 200 и 400 давала фактор остаточной разупорядоченности г = 0,03 и размер Шеррера 3,2 нм, перпендикулярный плоскостям колец в сухом состоянии. Средняя ширина отражений 1-10 и 110 лежит явно выше δ q по сравнению с графиком q 2 d для отражений 200 и 400. Подразумевается, что средний размер Шеррера в направлениях, нормальных к плоскостям решетки 1-10 и 110 был меньше, чем средний размер Шеррера в направлении, нормальном к плоскости решетки 200, как в еловой целлюлозе.Однако низкая интенсивность отражения 400 и, как следствие, сложность оценки вклада, связанного с беспорядком, в общее уширение сделали этот вывод менее надежным, чем для еловой целлюлозы.
Характеристика неупорядоченных компонентов с помощью дейтерирования и широкоугольного рассеяния нейтронов
В кристаллографическом исследовании целлюлозы Iα и Iβ (Nishiyama et al., 2002, 2003) разница в дифракции нейтронов, обусловленная полным высокотемпературным дейтерированием гидроксильных групп, использовалась для определения положений протонов и, следовательно, геометрии водородных связей. .Также должна быть возможность использовать сильное различие между рассеянием нейтронов дейтерием и протонами для определения вклада в рассеяние неупорядоченных областей, доступных для дейтерирования в более мягких условиях температуры окружающей среды, используемых в экспериментах с SANS и инфракрасным преобразованием Фурье (FTIR). Как может быть показано с помощью FTIR (см. Ниже), гидроксильные группы на нецеллюлозных полисахаридах доступны для дейтерирования, как и обращенные наружу гидроксилы на поверхности целлюлозы, если они не заблокированы агрегацией (Horikawa et al., 2009). Для неупорядоченных областей внутри микрофибрилл, чтобы допустить D 2 O, они должны иметь более широкие интервалы решетки, чем кристаллическая целлюлоза (Matthews et al., 2011).
На рис. 3А показано, что наблюдались видимые изменения в картине дифракции нейтронов при обратимом дейтерировании при температуре окружающей среды. Изменения включали увеличение или уменьшение интенсивности отдельных отражений, а также изменения диффузного рассеяния. В принципе, изменение интенсивности для любого отражения может быть положительным, отрицательным или нейтральным в зависимости от влияния дейтерирования на структурный фактор.В качестве руководства, интенсивности основных экваториальных и осевых отражений для целлюлозы Iβ в нативной (H) и пердейтерированной (D) формах были рассчитаны с использованием Shelxl-97 (Sheldrick, 2008) в пакете WinGX (Farrugia, 1999), из статическая модель, использующая опубликованные координаты атомов (Nishiyama et al., 2002). Из трех основных экваториальных отражений, определяющих геометрию боковой упаковки целлюлозных цепей, влияние дейтерирования на интенсивность было предсказано положительным для 200 и сильно отрицательным для 110 и 1-10.Было предсказано, что осевое отражение 004 будет увеличиваться по интенсивности при дейтерировании, в то время как отражение 001 отсутствовало как в D, так и в H-формах. Эти прогнозы согласуются с наблюдаемыми нейтронографическими изображениями для высококристаллической целлюлозы Iβ (Nishiyama et al., 2002).
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина). Ось волокна вертикальна.B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина).Ось волокна вертикальна. B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Наблюдаемый эффект поверхностного дейтерирования микрофибрилл был положительным для отражений 200 и 004, отрицательным для 110 и нейтральным для 1-10.Профиль вычитания (D — H) для экваториальных отражений (рис. 3B) показал, что их положение и ширина, соответствующие цепям, доступным для дейтерирования, не заметно отличались от фракции, которая не подвергалась дейтерированию. В азимутальном направлении экваториальная интенсивность показывала более широкий нижний хвост распределения ориентации в D, чем в H-дифракционной картине (рис. 3E), подразумевая, что некоторые из цепей, доступных для дейтерирования, были менее хорошо ориентированы, чем недоступные цепочки целлюлозы.
На оси волокна (рис. 3D) отражение 004 в D-форме было центрировано при немного меньшем q , чем в H-форме, что означает, что некоторые из цепочек, доступных для дейтерирования, были более протяженными, чем недоступные цепи. Разница в среднем расстоянии d составляла всего около 0,5%, а радиальная ширина этого отражения была одинаковой как для D, так и для H-формы. Отражение 002 было сильнее в D-форме, а отражение 001, отсутствующее в H-дифракционной картине, появилось в профиле разности осей.Наличие этого отражения 001 после дейтерирования означает, что некоторые из дейтерированных цепей не имели такого же продольного смещения, как чистая целлюлоза Iβ или целлюлоза Iα.
Почти изотропные изменения диффузного рассеяния при дейтерировании привели к появлению широкого кольца повышенной интенсивности в области 15 нм -1 < q <20 нм -1 , а ниже 15 нм -1 , была уменьшена диффузная интенсивность (рис. 3В). Изотропные компоненты, ответственные за эти особенности диффузного рассеяния, включают нецеллюлозные компоненты, которые обмениваются с D 2 O и небольшими количествами остаточной связанной воды.
Эксперименты по спиновой диффузии протонов и пространственные отношения между упорядоченными и неупорядоченными цепями
Скорость, с которой ядерный магнетизм уравновешивается за счет спиновой диффузии между наноразмерными доменами, может использоваться для оценки расстояния между этими доменами. Эксперименты по спиновой диффузии протонов, основанные на этом принципе, были применены к древесине (Newman, 1992; Altaner et al., 2006) и бактериальной целлюлозе (Masuda et al., 2003). Использовалось двумерное (2D) представление этого эксперимента (Fernandes et al., 2011), чтобы изучить расположение упорядоченных и неупорядоченных областей в еловой целлюлозе, используя различия в их пространственном отделении от лигнина. Совершенно другой эксперимент по спин-диффузии 13 C на соломе кукурузы ( Zea mays ) (Foston et al., 2012) привел к аналогичным выводам.
Целлюлоза колленхимы сельдерея не лигнифицирована, но содержит небольшие остаточные количества гемицеллюлоз глюкоманнана и ксилоглюкана, которые, обладая относительно высокой термической подвижностью, могут использоваться вместо лигнина в качестве резервуаров спина протонов.На рисунке 4 показан 2D-спектр этого эксперимента. Перекрестные пики указывают на домены, которые удовлетворяют двум условиям: (1) домены различаются по подвижности и, следовательно, по намагниченности в начале периода смешения спинов протонов в последовательности импульсов; и (2) домены достаточно разделены в пространстве, чтобы потребовалось измеримое время, несколько миллисекунд, для уравновешивания намагниченности протонов посредством спиновой диффузии. Приведенное ниже назначение сигналов основано на Sturcová et al. (2004).
Рисунок 4.
2D-представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, отслеживаемых по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены. [См. Онлайн-статью для цветной версии этого рисунка.]
Рис. 4.
Двумерное представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, мониторинг которого проводился по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены.[Цветную версию этого рисунка см. В онлайн-статье.]
Одномерные спектры C-ЯМР 13 были аналогичны тем, которые наблюдались ранее (Sturcová et al., 2004), с сигналами C-4 от кристаллической целлюлозы (89 –90 частей на миллион [частей на миллион]) и неупорядоченной целлюлозы (84–85 частей на миллион) в примерном соотношении 1: 2. В 2D-спектре присутствовали кросс-пики между C-4 кристаллической целлюлозы и C-4 неупорядоченной целлюлозы, но они были относительно слабыми. Также присутствовал кросс-пик между C-6 кристаллической целлюлозы (65 ppm) и C-4 неупорядоченной целлюлозы (84–85 ppm).Перекрестные пики от гемицеллюлозы C-1 (101 ppm) и C-4 (83 ppm) до кристаллической целлюлозы C-4 были сильнее, чем от неупорядоченной целлюлозы C-4, и перекрестные пики от гемицеллюлозы C-1 и C-4 к кристаллической целлюлозе С-6 были еще сильнее. Наиболее сильные кросс-пики связаны с сигналами C-2, C-5 и C-6 от неупорядоченной целлюлозы при 73, 75 и 62 м.д., но каждый из этих сигналов, очевидно, также содержит вклад гемицеллюлоз и, следовательно, не может быть определен. однозначно трактуется.
Эти наблюдения согласуются с моделями, в которых неупорядоченная и кристаллическая целлюлоза не смешиваются, а находятся в непосредственной пространственной близости внутри каждой микрофибриллы, а гемицеллюлозы расположены вне микрофибрилл, вероятно, частично за пределами каждого небольшого пучка микрофибрилл, но разница в кросс-пике интенсивность кристаллической и неупорядоченной целлюлозы предполагает, что одна группа гемицеллюлоз была в тесной связи с поверхностями микрофибрилл.
Поляризованный дейтерированный FTIR и модели водородной связи
Поляризованный FTIR предоставляет информацию о направлении водородных связей. Исследования FTIR на древесине ели показали, что гидроксильные группы на неупорядоченной целлюлозе на поверхности микрофибрилл были доступны для дейтериевого обмена, за исключением тех случаев, когда микрофибриллы были плотно агрегированы вместе. Также были доступны гидроксильные группы на гемицеллюлозах. Обмен дейтерия сдвигает полосу растяжения, связанную с каждой гидроксильной группой, на более низкую частоту в 1 раз.34, из-за большей массы атома дейтерия, но сохраняет порядок полос, потому что структура не изменилась (Sturcová et al., 2003).
На фиг. 5 показано, что дейтерирование либо клеточных стенок колленхимы сельдерея, либо изолированной целлюлозы было обширным, но меньше, чем количество, предсказанное из соотношения 2: 1 неупорядоченной целлюлозы к кристаллической, оцененного с использованием сигналов C-4 в спектрах ЯМР. Образец полос растяжения O-D легче всего сравнить с рисунком полос растяжения O-H, если шкала частот искусственно сдвинута в 1 раз.34 (рис. 5). Эти две модели явно отличались. Полосы растяжения O-D были менее ориентированы в продольном направлении, чем полосы растяжения O-H, показывая, что меньшее количество водородных связей в дейтерированной фракции было направлено вдоль оси микрофибрилл, а больше — наружу. Как и в еловой целлюлозе (Fernandes et al., 2011), дейтерирование снизило интенсивность выступающего плеча на 3400 см -1 в области растяжения O-H, что соответствует гидроксильным группам, более слабо связанным водородными связями и более направленным наружу, чем в среднем.После сдвига оси частот на на 1,34 раза максимум довольно широкой группы перекрывающихся полос валентных колебаний OD совпал с этим плечом, которое первоначально было отнесено к аномальной части Iα-подобного спектра целлюлозы (Sturcová et al., 2004), но позже было показано, что в еловой целлюлозе они соответствуют недоступным гидроксильным группам в неупорядоченных областях (Fernandes et al., 2011). Отнесение к неупорядоченным доменам, а не к целлюлозе Iα, подтверждается данными, представленными здесь.Только часть интенсивности на 3 400 см -1 была смещена дейтерированием, подразумевая, что некоторые из этих неупорядоченных цепей в целлюлозе колленхимы сельдерея доступны для дейтерирования, а некоторые — нет.
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой. Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию.Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В онлайн-статье.]
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой.Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию. Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В статье в Интернете.]
ОБСУЖДЕНИЕ
Структура из микрофибриллы
Целлюлоза, которую можно условно назвать кристаллической, и неупорядоченная целлюлоза, отличная от известных кристаллических форм целлюлозы Iα и Iβ, были идентифицированы всеми использованными методами.Из этого не следует, что одни и те же неупорядоченные фракции наблюдались всеми этими методами (т.е. что цепи, идентифицированные как неупорядоченные, в одних и тех же отношениях отличались от известных кристаллических форм). Наблюдались конформационный беспорядок, беспорядок упаковки и беспорядок водородных связей.
Спектры 13 C-ЯМР целлюлозы колленхимы сельдерея (рис. 4) идентифицировали цепи, которые отличались по конформации как от целлюлозы Iα, так и от целлюлозы Iβ, имеющие некоторую свободу вращения вокруг связи C5-C6 и имеющие C-6 В конформации преобладают гош-гош ( gg ) и гош-транс ( gt ) формы (Sturcová et al., 2004), как и в еловой целлюлозе (Fernandes et al., 2011), хотя соотношение gg — gt значительно отличалось от еловой (Sturcová et al., 2004). Спектры дейтерирования FTIR целлюлозы сельдерея показали, что некоторые домены имели паттерны водородных связей, отличные от наблюдаемых в целлюлозе Iα и целлюлозе Iβ, с пониженной средней прочностью водородных связей (более высокая средняя частота растяжения O-H) и меньшей общей тенденцией к осевой ориентации водорода. Изменение ориентации водородных связей было очевидным как в цепях, доступных для дейтериевого обмена, так и в недоступных цепях, и соответствовало переходу от транс-гош ( tg ) конформации C-6, обнаруженной в кристаллической целлюлозе, к gg . и конформеры GT .Подобные наблюдения на еловой целлюлозе были рационализированы с точки зрения модели, в которой ядро каждой микрофибриллы было занято в основном кристаллической целлюлозой, а поверхность микрофибрилл была занята цепями целлюлозы с несколько иной конформацией и водородными связями, некоторые из которых были доступны для обмена дейтерия, а другие были представлены. недоступны из-за боковой агрегации микрофибрилл. Подобная модель согласуется с FTIR-спектрами сельдерея (рис. 5), более ранними данными ЯМР (Sturcová et al., 2004), а также данные спин-диффузионного ЯМР, представленные здесь (рис. 4). По данным ЯМР спин-решеточной релаксации 13 C (Viëtor et al., 2002), целлюлоза колленхимы сельдерея имеет более высокую долю конформера C-6 gg, чем еловая целлюлоза. Размеры элементарной ячейки значительно различались между этими двумя видами целлюлозы, среднее расстояние между листами было больше в целлюлозе колленхимы сельдерея, а осевой размер был немного меньше. Молекулярное моделирование предложило связь между конформацией C-6 и шагом решетки (Matthews et al., 2012). Как и в случае с еловой целлюлозой, было невозможно и, возможно, бессмысленно различать формы целлюлозы Iα и Iβ с помощью ЯМР при наличии такого большого конформационного нарушения (Matthews et al., 2011).
Модели микрофибрилл с неупорядоченными поверхностями неоднократно предлагались на основе данных ЯМР (Newman, 1992; Viëtor et al., 2002; Bergenstråhle et al., 2008), но было неясно, как они должны согласовываться с полученными моделями. от дифракции рентгеновских лучей, которая более чувствительна к структуре упаковки цепей, чем к конформации цепи или водородным связям.Данные широкоугольного рассеяния нейтронов (WANS) (рис.3) показали, что в доменах, доступных для дейтерирования, цепи были упакованы латерально таким же образом и почти с такими же средними расстояниями, как и в кристаллических областях, несмотря на различия в конформации. и водородные связи, продемонстрированные спектроскопическими экспериментами, несмотря на значительные статистические различия в упаковке цепей повсюду в микрофибрилле. Повсюду в каждой микрофибрилле листы цепочек были уложены друг на друга с более широким средним интервалом, чем в более кристаллических целлюлозах.Дальнейшее следствие состоит в том, что большинство цепей, доступных для дейтерирования, были расположены на поверхностях микрофибрилл, а не в «аморфных» сегментах микрофибриллы, занимающих всю ширину микрофибриллы, потому что цепи внутри таких сегментов должны быть неплотно упакованы, чтобы вода могла стекать. входить.
Судя по осевому отражению 004, доступные для дейтерирования цепи оказались немного более вытянутыми в продольном направлении, чем недоступные цепи, возможно, из-за скручивания микрофибрилл.Продольное смещение было частично нарушено в цепочках, доступных для дейтерирования, что усложняло различение форм Iα и Iβ.
Широкий диапазон расстояний между боковыми цепями в нецеллюлозной фракции, доступный для дейтерирования, может быть выведен из диффузной, неориентированной части нейтронографических изображений. Вода, связанная с нецеллюлозной фракцией, рассеивала рентгеновские лучи аналогично жидкой воде, предполагая аналогичную функцию распределения пар, но из-за пониженной тепловой подвижности величина ее вклада в структуру WAXS была больше, чем ожидалось для объемной жидкой воды.
Диаметр и форма микрофибриллы
Данные SANS предполагают диаметр микрофибрилл, приблизительно равный межцентровому расстоянию от 2,9 до 3,0 нм в сухом состоянии. Предполагая круглое поперечное сечение и учитывая относительно рыхлую упаковку цепей, наблюдаемую при широкоугольном рассеянии, этот номинальный диаметр будет соответствовать 21–22 цепям. Микрофибриллы, конечно, не имеют круглой формы, и фактическое количество цепочек зависит от формы поперечного сечения.Наблюдения WANS подтвердили, что поверхностные цепочки располагались на той же неупорядоченной решетке, что и внутренние цепочки, и должны быть включены в расчеты размеров Шеррера из WAXS.
Межцентровое расстояние SANS и поперечные размеры по Шерреру были аналогичны тем, которые наблюдались для еловой целлюлозы (Fernandes et al., 2011). Размер [200] Шеррера был больше, чем среднее значение размеров [1-10] и [110], что согласуется с моделью прямоугольной микрофибриллы (Matthews et al., 2006; Fernandes et al., 2011), который имеет относительно гидрофобные [200] грани кристаллов, открытые сверху и снизу (рис. 6), вместо диагональных [1-10] и [110] граней кристаллов, открытых в целлюлозах водорослей (Baker et al. ., 1997). Однако опора для прямоугольной, а не ромбовидной модели менее надежна, чем для ели, из-за сложности коррекции беспорядка с использованием очень слабого отражения 400 и из-за проблем, присущих оценке отдельных ширины перекрывающихся 1-10 и 110 отражений.Учитывая наблюдаемое обширное нарушение, нельзя даже с уверенностью сказать, что микрофибриллы первичной стенки имеют постоянную форму поперечного сечения.
Рисунок 6.
Модели поперечного сечения микрофибрилл колленхимы сельдерея. A, 24-цепочечная ромбовидная модель, 2,9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться.Расчетные длины столбцов [200], [1-10] и [110] были следующими, соответственно: модель A, 2,5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 6.
Модели для поперечного сечения микрофибрилл колленхимы сельдерея. A, модель ромбовидной формы с 24 цепями, 2.9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться. Расчетные длины столбцов [200], [1-10] и [110] были соответственно следующими: модель A, 2.5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В статье в Интернете.]
Прямоугольное поперечное сечение с уложенными друг на друга листами из трех цепей будет соответствовать одной трети цепей, находящихся в центральное положение в каждом листе, конформационно сходное с кристаллическими целлюлозами, как наблюдалось с помощью ЯМР.Наблюдение единственного малоуглового пика Брэгга согласуется со слегка диспергированными расстояниями между микрофибриллами, которые можно было бы ожидать от примерно прямоугольных микрофибрилл, возможно, скрученных, находящихся в прерывистом контакте друг с другом. Могут быть приспособлены как постоянные, так и слегка изменяющиеся диаметры микрофибрилл, а также небольшое количество микрофибрилл, значительно отличающихся от доминирующих размеров.
Число цепей на микрофибриллу имеет отношение к структуре комплексов, которые синтезируют целлюлозу во время роста.Если количество цепей кратно шести, что соответствует геометрии синтетического комплекса, тогда стопка из восьми трехцепочечных листов (24 цепи) даст наиболее близкое совпадение с размером Шеррера 200 (рис. 6B), как и для еловая целлюлоза (Fernandes et al., 2011). Однако в нашем случае неопределенность довольно велика, и уровень беспорядка упаковки предполагает, что возможны нерегулярные и переменные сечения. Модель из 18 цепей (6 × 3) не исключена для еловой целлюлозы и согласуется с данными, представленными здесь (рис.6C), если размер 200 по Шерреру был увеличен дополнительными цепями гемицеллюлозы, как это было предложено для ели (Fernandes et al., 2011), или увеличен путем наложения одной микрофибриллы поверх другой на короткие расстояния.
Кристалличность
Что означает «кристалличность», зависит от методов, используемых для обнаружения компонентов, которые отличаются от кристаллической целлюлозы. Между спектроскопическими методами обнаружения цепей, отличающихся по конформации от общепринятых форм кристаллической целлюлозы и, следовательно, демонстрирующих различные модели водородных связей, наблюдалась некоторая степень согласованности.Это включало нецеллюлозные полисахариды, и, в частности, в спектрах FTIR было трудно провести четкое различие между вкладом нецеллюлозных полисахаридов и некоторой неупорядоченной целлюлозы.
Эксперименты с дейтерированием WANS показали, что эти конформационные различия мало влияют на упаковку целлюлозных цепей или дифракцию. Структура микрофибрилл явно содержала значительный беспорядок в боковой упаковке цепей, но этот беспорядок распространялся по всей структуре и не ограничивался доступными для дейтерия поверхностями.Кристалличность, оцениваемая дифракционными методами, — это не то же самое, что кристалличность, оцениваемая спектроскопически. В обоих случаях кажущаяся кристалличность будет увеличиваться с увеличением диаметра микрофибрилл, но как прямое следствие уменьшения уширения по Шерреру в дифракционных экспериментах и из-за уменьшения отношения поверхности к объему в спектроскопических экспериментах, которые позволяют различать поверхностные цепи.
Агрегация микрофибрилл
Агрегация микрофибрилл в более крупные пучки хорошо известна для древесины хвойных пород (Wickholm et al., 1998; Фахлен и Сальмен, 2005 г .; Fernandes et al., 2011). Фибриллярные единицы шириной от 5 до 10 нм, иногда до 40 нм, наблюдались с помощью электронной микроскопии (McCann et al., 1990; Sugimoto et al., 2000), но их интерпретация осложняется возможным включением нецеллюлозных полимеров. . Представленные здесь данные включают две прямые линии доказательств агрегации микрофибрилл первичной стенки в колленхиме. Доказательства малоуглового брэгговского рассеяния рентгеновских лучей (Kennedy et al., 2007a) и нейтронов ясно показали, что агрегация существует. Диапазон расстояний между микрофибриллами было трудно сделать вывод, потому что интенсивность рассеяния сильно увеличивалась с разделением микрофибрилл, так что хорошо разделенные микрофибриллы были чрезмерно представлены на диаграммах рассеяния. Низкий контраст МУРН в сухом состоянии и сходство межцентрового расстояния МУРН со средними размерами Шеррера WAXS предполагают, что некоторые сегменты микрофибрилл оставались в прямом контакте друг с другом, тогда как другие сегменты микрофибрилл становились хорошо разделенными при гидратации.
Спектры дейтерирования FTIR как от стенок клеток колленхимы, так и от изолированной целлюлозы включали полосы растяжения гидроксила с центром на 3 400 см -1 с нейтральной поляризацией. Эти гидроксильные полосы растяжения были отнесены к гидроксильным группам, отличным от всех групп Iα и Iβ кристаллических форм целлюлозы, более ориентированным наружу по ориентации и частично, но не полностью доступным для дейтерирования. Принимая во внимание также данные SANS и ЯМР, соответствующие гидроксильные группы могут быть отнесены к поверхностям микрофибрилл.Поскольку эти поверхности были только частично доступны для D 2 O, отсюда следует, что микрофибриллы были агрегированы вместе на части своей длины с границами раздела, непроницаемыми для воды. Эти особенности были обнаружены в древесине ели, но были также различия: древесина ели имеет разные нецеллюлозные полимеры, что, по-видимому, приводит к более тесному связыванию между поверхностями микрофибрилл и к напряжениям, которые искажают геометрию элементарной ячейки при гидратации (Fernandes et al., 2011).
Полоса валентных колебаний гидроксила 3,400 см -1 в FTIR-спектре, перемещенная до 2,540 см -1 за счет обмена D 2 O, ранее была неправильно отнесена к аберрантной форме целлюлозы Iα (Sturcová et al., 2004), но, по-видимому, является маркером поверхностных цепей и наличия агрегации микрофибрилл, если эти поверхностные цепи недоступны для дейтерирования. Интенсивность этой полосы сильно уменьшилась, когда целлюлоза из колленхимы сельдерея была подвергнута гидротермальному отжигу, что привело к слиянию микрофибрилл в более крупные кристаллические единицы с уменьшенным соотношением поверхности к объему (Sturcová et al., 2004). Интенсивность FTIR на уровне 3400 см или около него -1 была зарегистрирована в препаратах клеточных стенок, например, арабидопсиса дикого типа ( Arabidopsis thaliana ; MacKinnon et al., 2006), предполагая, что эта форма агрегации не является уникальной особенностью целлюлозы колленхимы сельдерея, но более широко распространена в первичных клеточных стенках.
Влияние на рост растений
Размер и структура микрофибрилл первичной стенки дают важные ключи к разгадке природы ферментных комплексов, ответственных за ориентированный синтез целлюлозы и, следовательно, за направленную экспансию клеток. Размножение растительных клеток требует разрушения сети нековалентно сшитых микрофибрилл в клеточной стенке.Понимание топологии этой сети, следовательно, необходимо, и недавно принятая идея о том, что ксилоглюканы просто покрывают и связывают поверхности микрофибрилл, была поставлена под сомнение (Bootten et al., 2004; Park and Cosgrove, 2012a). Данные спин-диффузии ЯМР (рис. 4) показали, что некоторые цепи гемицеллюлозы, вероятно, ксилоглюканы, располагались близко к поверхностям микрофибрилл и, таким образом, вероятно, лежали между микрофибриллами внутри агрегатов микрофибрилл. Однако другие сегменты микрофибрилл, по-видимому, находятся в прямом контакте без промежуточных полимеров, демонстрируя нерегулярный характер агрегации, которого можно было бы ожидать, если микрофибриллы были скрученными и нецилиндрическими, и предполагая, что взаимодействия целлюлоза-целлюлоза, а также ксилоглюкан-целлюлоза должны быть разбиты на отдельные агрегированные микрофибриллы.Др. Механизмы необходимы для отделения одного пучка микрофибрилл от следующего во время роста путем разрушения ксилоглюкана или др. Мостиков между пучками микрофибрилл (Anderson et al., 2010). Агрегация микрофибрилл вдоль части их длины, в то время как другие части их длины остаются свободными для разделения или в них вставлены другие полисахариды, поднимает интересные вопросы о действии экспансинов (Cosgrove, 2005) и о том, как действует действие определенных глюканов. гидролазы взаимодействуют с таковой ксилоглюкан-специфической эндоглюканазы (Park and Cosgrove, 2012b).
Интересен высокий уровень беспорядка на границах раздела, наиболее важных для роста, между микрофибриллами и нецеллюлозными полимерами, а также между одной микрофибриллой и другой. Примеры прочных биологических материалов из царства животных показывают, что неупорядоченные домены могут вносить вклад в ударную вязкость (высокую энергию разрушения) при внешнем стрессе (van Beek et al., 2002). В первичных клеточных стенках устойчивость к разрушению должна сочетаться с контролируемой ферментативно-опосредованной продуктивностью, по-видимому, на неупорядоченных границах раздела, что обеспечивает рост.
Последствия разложения целлюлозы
Доступность микрофибрилл целлюлозы для целлюлаз является важным фактором превращения лигноцеллюлозы в биотопливо и еще более важна в случае нелгнифицированной биомассы. Нестабильность целлюлозы часто связывают с ее кристалличностью (Arantes and Saddler, 2010), а «декристаллизацию» иногда называют целью процессов предварительной обработки (Beckham et al., 2011), без полного согласия относительно того, что означают эти термины.Степень, в которой микрофибриллы целлюлозы имеют относительно гидрофобную [200] открытую поверхность, будет положительно влиять на начальную стадию деградации целлюлазами, которые специфически связываются с этой поверхностью (Dagel et al., 2011; Liu et al., 2011). Было бы интересно узнать больше о специфичности связывания оксидаз целлюлозы GH61 / CBM33 (Forsberg et al., 2011; Quinlan et al., 2011), некоторые из которых также связываются с одной и той же поверхностью (Li et al., 2012). Агрегация микрофибрилл целлюлозы — это фактор сопротивляемости, который заслуживает большего внимания, и теперь к нему можно подойти с помощью экспериментов по дейтерированию FTIR.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Было выделенонитей колленхимы из зрелых черешков сельдерея ( Apium graveolens ), клеточные стенки и их целлюлозная фракция были приготовлены, как описано (Sturcová et al., 2004), и хранились в сухом виде. Подробнее см. Дополнительные материалы и методы S1.
SANS
ЭкспериментыSANS были выполнены на канале D11 в Институте Лауэ-Ланжевена на клеточных стенках колленхимы сельдерея и изолированной целлюлозе.Подробнее об эксперименте см. Дополнительные материалы и методы S1.
WAXS
дифрактограммы рентгеновских лучей получали при температуре окружающей среды с использованием дифрактометра Rigaku R-axis / RAPID для рентгеновских пластин с источником излучения с молибденовой рентгеновской эмиссией Kα (λ = 0,07071 нм). Пучок коллимировался до диаметра 0,5 мм. Образцы имели толщину 0,7 мм в направлении, параллельном пучку, а их другие размеры превышали диаметр пучка. Дифрактограммы были собраны в режиме перпендикулярного пропускания, за исключением экспериментов с наклоном для измерения осевых отражений.Расстояние от центра дифракционной картины было выражено как q = 4πsinθ / λ, где 2θ — угол рассеяния. Для получения дополнительных сведений об экспериментальных методах и методах обработки данных (Fernandes et al., 2011) см. Дополнительные материалы и методы S1.
WANS
Дифракция нейтронов проводилась на канале D19 в Институте Лауэ-Ланжевена при средней длине волны нейтронов 0,242 нм и расстоянии от образца до детектора 0,756 м. Образцы размером 25 × 15 × 1 мм собирали из нескольких выровненных нитей колленхимы и выдерживали при контролируемом давлении воды или паров D 2 O в проточном контейнере со стенками из алюминиевой фольги.Подробнее см. Дополнительные материалы и методы S1.
ЯМР
A 1 H Эксперимент по спиновой диффузии Гольдмана-Шена с детектированием по спектру 13 C после кросс-поляризации был проведен на клеточных стенках колленхимы сельдерея, гидратированных 0,2 см. 3 г −1 D 2 O. Экспериментальные процедуры и анализ данных, включая 2D-представление данных, были такими, как описано (Fernandes et al., 2011).
Дейтерирование FTIR
Стенки клеток колленхимы сельдерея и изолированную целлюлозу помещали в проточную ячейку с окнами из фторида бария и заменяли воздухом, насыщенным О 2 О, до тех пор, пока спектры не перестали изменяться. Затем газовую линию переключали на азот, тщательно осушая, пропуская через сухое молекулярное сито 4А, а затем пентоксид фосфора (Sicapent; Aldrich). Спектры FTIR собирали с помощью спектрометра Thermo Nicolet Nexus, оборудованного приставкой для микроскопа Nicolet Continuum, имеющей охлаждаемый жидким азотом детектор теллурида кадмия ртути (Fernandes et al., 2011).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Rigaku, Ltd. за предоставленный инструмент и Fujiang Zhang за получение некоторых спектров FTIR.
Глоссарий
SAXS
Малоугловое рассеяние рентгеновских лучей
D 2 O
SANS
Малоугловое рассеяние нейтронов
WAXS 9407
широкоугольное рассеивание
WANS
широкоугольное рассеяние нейтронов
FTIR
Инфракрасное преобразование Фурье
2D
ppm
LITERATURE CITED
(
2006
)Пространственные отношения между полимерами в ели ситкинской: исследования спиновой диффузии протонов
.Holzforschung
60
:665
—673
(
2010
)Визуализация в реальном времени переориентации целлюлозы во время расширения клеточной стенки в корнях Arabidopsis
.Plant Physiol
152
:787
—796
(
2009
)Выяснение механизмов сборки и субъединичного взаимодействия целлюлозосинтазного комплекса вторичных клеточных стенок Arabidopsis
.J Biol Chem
284
:3833
—3841
(
1997
)Атомно-силовая микроскопия высокого разрешения микрокристаллов нативной целлюлозы I Valonia
.J Struct Biol
119
:129
—138
(
2011
)Происхождение стойкости биомассы на молекулярном уровне: свободные энергии декристаллизации для четырех обычных полиморфов целлюлозы
.J Phys Chem B
115
:4118
—4127
(
2008
)Динамика границ раздела целлюлоза-вода: времена спин-решеточной релаксации ЯМР, рассчитанные на основе атомистического компьютерного моделирования
.J Phys Chem B
112
:2590
—2595
(
2004
)Твердотельный 13 C-ЯМР-спектроскопия показывает, что ксилоглюканы в первичных клеточных стенках маша (Vigna radiata L.) происходят в разных доменах: новая модель взаимодействий ксилоглюкан-целлюлоза в клеточной стенке
.J Exp Bot
55
:571
—583
(
2009
)Растения контролируют свойства и работу своих органов посредством ориентации фибрилл целлюлозы в их клеточных стенках
.Интегр Комп Биол
49
:69
—79
(
2005
)Рост клеточной стенки растений
.Nat Rev Mol Cell Biol
6
:850
—861
(
2011
)Визуализация на месте отдельных углеводсвязывающих модулей на микрофибриллах целлюлозы
.J Phys Chem B
115
:635
—641
(
1996
)Паренхиматозная клеточная целлюлоза из жома сахарной свеклы: получение и свойства
.Целлюлоза
3
:183
—188
(
2005
)Распределение пор и матрицы в стенке волокна, выявленное с помощью атомно-силовой микроскопии и анализа изображений
.Биомакромолекулы
6
:433
—438
(
1999
)Пакет WinGX для кристаллографии низкомолекулярных монокристаллов
.J Appl Cryst
32
:837
—838
(
2011
)Наноструктура микрофибрилл целлюлозы в древесине ели
.Proc Natl Acad Sci USA
108
:E1195
—E1203
(
2011
)Расщепление целлюлозы белком CBM33
.Protein Sci
20
:1479
—1483
(
2012
)Твердотельное селективное (13) C возбуждение и спин-диффузионный ЯМР для определения пространственных размеров в стенках растительных клеток
.J Agric Food Chem
60
:1419
—1427
(
2000
)Характеристика поперечных связей между микрофибриллами целлюлозы и их появление во время элонгационного роста в эпикотиле гороха
.Физиология растительных клеток
41
:486
—494
(
2011
)Кортикальные микротрубочки оптимизируют кристалличность клеточной стенки для обеспечения однонаправленного роста Arabidopsis
.Завод J
66
:915
—928
(
1999
)Выражение паттерна в растениях: сочетание молекулярных и основанных на исчислении биофизических парадигм
.Am J Bot
86
:1059
—1076
и другие. (
2012
)Кристалличность микрофибрилл целлюлозы снижается за счет мутации С-концевых остатков трансмембранной области CESA1A903V и CESA3T942I синтазы целлюлозы
.Proc Natl Acad Sci USA
109
:4098
—4103
(
2007
)Устойчивость биомассы: инженерные установки и ферменты для производства биотоплива
.Наука
315
:804
—807
(
2009
)Различия между сортами в размерах микрофибрилл целлюлозы, наблюдаемые с помощью инфракрасной спектроскопии
.Целлюлоза
16
:1
—8
(
1995
)Радиальная упаковка, порядок и беспорядок в фибриллах коллагена
.Biophys J
68
:1661
—1670
(
1992
)Контроль толщины клеточных стенок колленхимы пектинами
.Планта
187
:218
—220
(
2011
)Стенки растительных клеток: супрамолекулярные ансамбли
.Food Hydrocoll
25
:257
—262
(
2007a
)Диаметр микрофибрилл в целлюлозе колленхимы сельдерея: рассеяние рентгеновских лучей и данные ЯМР
.Целлюлоза
14
:235
—246
(
2007b
)Влияние гидратации на расстояние между микрофибриллами целлюлозы первичной стенки: исследование малоуглового рентгеновского рассеяния
.Целлюлоза
14
:401
—408
(
2008
)Выявление структурного и функционального разнообразия клеточных стенок растений
.Curr Opin Plant Biol
11
:308
—313
(
2001
)Рентгеновская структура мерсеризованной целлюлозы II при разрешении 1 Å
.Биомакромолекулы
2
:410
—416
(
2012
)Структурная основа нацеливания на субстрат и катализа грибковыми полисахаридмонооксигеназами
.Структура
20
:1051
—1061
(
2011
)Целлобиогидролаза гидролизует кристаллическую целлюлозу на гидрофобных поверхностях
.J Biol Chem
286
:11195
—11201
(
2006
)Структура и анизотропия клеточной стенки в procuste , мутанте целлюлозосинтазы Arabidopsis thaliana
.Планта
224
:438
—448
(
2005
)Расширение клеточной стенки приводит к координатному разделению параллельных микрофибрилл: данные сканирующей электронной микроскопии и атомно-силовой микроскопии
.Завод J
43
:181
—190
(
2003
)Твердотельный 13 C и 1 H спин-диффузионный ЯМР-анализ структуры микрофибрилл бактериальной целлюлозы
.Твердотельный ядерный магнитный резонон
23
:198
—212
(
2012
)Сравнение моделирования бета целлюлозы I с тремя силовыми полями углеводов
.J Chem Theory Comput
8
:735
—748
(
2011
)Высокотемпературные свойства целлюлозы I
.J Phys Chem B
115
:2155
—2166
(
2006
)Исследования микрокристаллической целлюлозы Ibeta
с помощью компьютерного моделирования.Углеводородный остаток
341
:138
—152
(
1990
)Прямая визуализация поперечных связей в первичной клеточной стенке растения
.J Cell Sci
96
:323
—334
(
2008
)Структура и биосинтез пектина
.Curr Opin Plant Biol
11
:266
—277
(
1992
)Ядерно-магнитно-резонансное исследование пространственных соотношений между химическими компонентами в стенках деревянных клеток
.Holzforschung
46
:205
—210
(
2009
)Структура и свойства микрофибриллы целлюлозы
.J Wood Sci
55
:241
—249
(
2002
)Кристаллическая структура и система водородных связей в целлюлозе Ibeta по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
124
:9074
—9082
(
2003
)Кристаллическая структура и система водородных связей в целлюлозе I (альфа) по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
125
:14300
—14306
(
2008
)Генетические доказательства того, что активность синтазы целлюлозы влияет на организацию кортикального массива микротрубочек
.Plant Physiol
147
:1723
—1734
(
2012a
)Изменения биомеханических свойств клеточной стенки у ксилоглюкан-дефицитного мутанта xxt1 / xxt2 Arabidopsis
.Plant Physiol
158
:465
—475
(
2012b
)Пересмотренная архитектура первичных клеточных стенок, основанная на биомеханических изменениях, индуцированных субстрат-специфическими эндоглюканазами
.Plant Physiol
158
:1933
—1943
и другие. (
2011
)Анализ окислительного разложения целлюлозы металлоферментом меди, который использует компоненты биомассы
.Proc Natl Acad Sci USA
108
:15079
—15084
и другие.(
2012
)CHITINASE-LIKE1 / POM-POM1 и его гомолог CTL2 представляют собой взаимодействующие с глюканом белки, важные для биосинтеза целлюлозы у Arabidopsis
.Plant Cell
24
:589
—607
(
2008
)Краткая история SHELX
.Acta Crystallogr A
64
:112
—122
(
2006
)Синтез целлюлозы у высших растений
.Annu Rev Cell Dev Biol
22
:53
—78
(
2004
)Структурные детали кристаллической целлюлозы из высших растений
.Биомакромолекулы
5
:1333
—1339
(
2003
)Поляризованная колебательная спектроскопия волоконных полимеров: водородные связи в целлюлозе II
.Биомакромолекулы
4
:1589
—1595
(
2000
)Новые методы позволяют проводить сравнительный анализ ориентации микротрубочек, текстуры стенок и скорости роста интактных корней Arabidopsis
.Plant Physiol
124
:1493
—1506
(
2009
)Динамическая координация систем цитоскелета и клеточной стенки во время морфогенеза растительных клеток
.Curr Biol
19
:R800
—R811
(
2002
)Молекулярная структура шелка драглайна пауков: складывание и ориентация белкового остова
.Proc Natl Acad Sci USA
99
:10266
—10271
(
2002
)Конформационные особенности кристаллической целлюлозы высших растений
.Завод J
30
:721
—731
(
2004
)Полиморфизм целлюлозы семейства I: повторное исследование целлюлозы IVI
.Биомакромолекулы
5
:1385
—1391
(
1998
)Определение некристаллических форм в целлюлозе I методом спектроскопии ЯМР CP / MAS C-13
.Углеводородный остаток
312
:123
—129
(
2007
)Организация боковых цепей пектинового арабинана и галактана в ассоциации с микрофибриллами целлюлозы в первичных клеточных стенках и связанных моделях предусматривается
.J Exp Bot
58
:1795
—1802
Заметки автора
© Американское общество биологов растений, 2013 г. Все права защищены.
© Автор (ы) 2013. Опубликовано Oxford University Press от имени Американского общества биологов растений. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0 /), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
Структура микрофибрилл целлюлозы в первичных клеточных стенках колленхимы | Физиология растений
Аннотация
В первичных стенках растущих клеток растений глюкозно-полимерная целлюлоза собрана в длинные микрофибриллы диаметром несколько нанометров. Жесткость и ориентация этих микрофибрилл контролируют рост клеток; Следовательно, синтез целлюлозы является ключевым фактором роста и морфогенеза растений.Колленхима сельдерея ( Apium graveolens ) является полезной модельной системой для изучения структуры микрофибрилл первичной стенки, поскольку ее микрофибриллы ориентированы с необычной однородностью, что облегчает проведение спектроскопических и дифракционных экспериментов. Используя комбинацию методов рассеяния рентгеновских лучей и нейтронов с колебательной спектроскопией и спектроскопией ядерного магнитного резонанса, мы показываем, что микрофибриллы колленхимы сельдерея имели средний диаметр от 2,9 до 3,0 нм, с наиболее вероятной структурой, содержащей 24 цепи в поперечном сечении, расположенные в восьми атомах водорода. -связанные листы из трех цепей с обширным беспорядком в боковой упаковке, конформации и водородной связи.Аналогичная структура из 18 цепей и структуры из 24 цепей разной формы хуже соответствовали данным. Конформационный беспорядок в основном ограничивался поверхностными цепями, а беспорядок в упаковке цепей — нет. То есть по положению и ориентации поверхностные цепи соответствовали неупорядоченной решетке, составляющей ядро каждой микрофибриллы. Были доказательства того, что соседние микрофибриллы нековалентно агрегированы вместе на части своей длины, предполагая, что необходимость разрушения этих агрегатов может быть сдерживающим фактором роста и гидролиза целлюлозы для производства биотоплива.
Рост и форма растений контролируются точно ориентированным расширением стенок отдельных клеток. Движущая сила для размножения клеток является осмотической, но скорость и направление экспансии контролируются механическими свойствами клеточной стенки (Szymanski and Cosgrove, 2009). Расширяющиеся первичные клеточные стенки представляют собой нанокомпозитные материалы, в которых длинные микрофибриллы целлюлозы диаметром несколько нанометров проходят через гидратированную матрицу ксилоглюканов, пектинов и других полимеров (Knox, 2008; Mohnen, 2008; Szymanski and Cosgrove, 2009; Scheller. и Ульвсков, 2010).Нативные микрофибриллы целлюлозы частично кристаллические (Nishiyama, 2009; Fernandes et al., 2011). Раньше считалось, что целлюлоза первичной стенки имеет уникальную кристаллическую структуру, называемую целлюлозой IV 1 (Dinand et al., 1996), но данные ЯМР предполагают наличие форм, подобных лучше охарактеризованным кристаллическим формам целлюлозы Iα и Iβ вместе с крупными кристаллическими формами целлюлозы. количества менее упорядоченной целлюлозы (Wickholm et al., 1998; Sturcová et al., 2004; Wada et al., 2004). Тем не менее, целлюлоза гораздо более упорядочена, чем любой другой компонент первичной клеточной стенки (Bootten et al., 2004), в соответствии с его ключевой ролью обеспечения силы и контроля роста.
Жесткость клеточной стенки наибольшая в направлении микрофибрилл целлюлозы, где рост является направленным, а преобладающая ориентация микрофибрилл обычно поперечна направлению роста (Green, 1999; MacKinnon et al., 2006; Szymanski and Cosgrove, 2009 г.). Расширение клеточной стенки требует либо увеличения расстояния между микрофибриллами (Marga et al., 2005), либо проскальзывания между ними (Cosgrove, 2005), либо того и другого, и микрофибриллы переориентируются в направлении роста (Anderson et al., 2010). Полимерные поперечные мостики между микрофибриллами (McCann et al., 1990), как полагают, противостоят этим деформациям наноструктуры клеточной стенки и, таким образом, контролируют скорость роста. До недавнего времени наибольшее внимание уделялось мостиковым соединениям ксилоглюканов, водородным связям с поверхностями микрофибрилл (Scheller, Ulvskov, 2010). Однако есть свидетельства того, что не все ксилоглюканы расположены надлежащим образом (Fujino et al., 2000; Park and Cosgrove, 2012a) и что могут быть задействованы другие мостиковые полимеры (Zykwinska et al., 2007). Также было высказано предположение, что пучки агрегированных микрофибрилл, а не отдельные микрофибриллы, могут быть ключевыми структурными единицами в первичных клеточных стенках (Anderson et al., 2010), как в древесине (Fahlén and Salmén, 2005; Fernandes et al., 2011). ). В таком случае отдельные микрофибриллы могут соединяться между пучками микрофибрилл. Таким образом, рост растительных клеток не совсем понятен, и нам нужна дополнительная информация о том, как контролируется ориентация целлюлозы, и о природе мостиковых полимеров, о поверхностях целлюлозы, с которыми эти полимеры связываются, и о сцеплении между поверхностями микрофибрилл, которые могут опосредовать агрегацию.
Микрофибриллы целлюлозы синтезируются на поверхности клетки большими ферментными комплексами, имеющими гексагональную симметрию, иногда называемыми «розетками» (Somerville, 2006). Каждый комплекс содержит несколько синтаз целлюлозы, которые различаются между первичными клеточными стенками и древесиной, хотя внешний вид комплексов похож (Somerville, 2006; Atanassov et al., 2009). Одновременный синтез с одного и того же конца всех цепей в микрофибрилле нативной целлюлозы является причиной их параллельности (Nishiyama et al., 2002, 2003), в отличие от энтропийно предпочтительной антипараллельной структуры, обнаруженной в искусственных целлюлозах, таких как вискоза (Langan et al., 2001). Очевидно, что количество цепей в микрофибрилле и количество целлюлозосинтаз в синтетическом комплексе взаимосвязаны. Принято считать, что количество цепочек делится на шесть, что соответствует симметрии гексагональной розетки и 36-цепочечным моделям (Himmel et al., 2007), ограниченным гидрофильными гранями кристаллов [110] и [1-10], поскольку в водорослевых целлюлозах (Bergenstråhle et al., 2008), получили широкое распространение. Сборка и ориентация целлюлозы взаимосвязаны, поскольку некоторые мутанты целлюлозосинтазы имеют фенотипы, дефектные по ориентации целлюлозы и растительной форме, а также обедненные содержанием целлюлозы (Paredez et al., 2008). У некоторых других мутантных линий, по-видимому, затрагивается кристалличность микрофибрилл (Fujita et al., 2011; Harris et al., 2012; Sánchez-Rodríguez et al., 2012).
Следовательно, детальное понимание структуры микрофибрилл целлюлозы первичной стенки поможет нам понять синтез целлюлозы, а также рост и структурную механику живых растений (Burgert and Fratzl, 2009).Первичные клеточные стенки и их целлюлозные скелеты также влияют на такие характеристики качества пищевых продуктов, как хрусткость салата из овощей и яблок ( Malus domestica ; Jarvis, 2011). Когда биотопливо производится из лигноцеллюлозной биомассы, лигнификация приводит к устойчивости (Himmel et al., 2007), но некоторые типы клеток у видов Miscanthus , просо ( Panicum virgatum ) и пожнивные остатки имеют только первичные стенки. без лигнина, и сопротивляемость этому зависит от природы микрофибрилл целлюлозы (Beckham et al., 2011).
Относительно подробная структура недавно была предложена для микрофибрилл древесины ели ( Picea spp.) (Fernandes et al., 2011), которые имеют диаметр 3,0 нм, что позволяет разместить только около 24 цепочек целлюлозы. Данные рентгеновской дифракции подтверждают «прямоугольную» форму (Matthews et al., 2006), ограниченную гранями [010] и [200]. Наблюдался значительный беспорядок, увеличивающийся к поверхности, и микрофибриллы были агрегированы в пучки размером примерно 15-20 нм в поперечнике, причем некоторые, но не все, боковые границы раздела были устойчивы к воде (Fernandes et al., 2011). Неупорядоченные домены являются особенностью других прочных биологических материалов, таких как шелк паука (van Beek et al., 2002).
Следовательно, представляет интерес, могут ли какие-либо из этих свойств древесной целлюлозы также быть обнаружены в микрофибриллах целлюлозы первичных (растущих) клеточных стенок. Было бы особенно полезно охарактеризовать нарушение, которое, как известно, присутствует в микрофибриллах первичной стенки, то есть определить, чем целлюлоза, которая не определяется как «кристаллическая», отличается от кристаллической целлюлозы.Многие эксперименты, ведущие к созданию структуры древесной целлюлозы, зависели от исключительно однородной ориентации микрофибрилл целлюлозы (Sturcová et al., 2004; Fernandes et al., 2011). Однако в растущих клеточных стенках микрофибриллы ориентированы неравномерно. Когда микрофибриллы сначала откладываются на внутренней поверхности первичной клеточной стенки, их ориентация обычно перпендикулярна направлению роста, но когда клеточная стенка расширяется, микрофибриллы переориентируются таким образом, что распределение ориентации интегрируется по толщине расширенной клеточной стенки. клеточная стенка становится все более близкой к случайной (Cosgrove, 2005; MacKinnon et al., 2006).
Эта техническая проблема не относится к клеточным стенкам колленхимы сельдерея ( Apium graveolens ), которые похожи по составу на другие первичные клеточные стенки, но имеют их микрофибриллы, ориентированные относительно равномерно вдоль оси клетки (Sturcová et al., 2004 ; Kennedy et al., 2007a, 2007b). Некоторая структурная информация о целлюлозе колленхимы сельдерея уже была получена в результате спектроскопических экспериментов и экспериментов по рассеянию (Sturcová et al., 2004; Kennedy et al., 2007a, 2007b), подтверждая беспорядок, ожидаемый в целлюлозе первичной стенки.Некоторые из этих экспериментов были аналогичны тому, что было сделано с еловой целлюлозой (Fernandes et al., 2011), но недостаточно данных, чтобы указать количество цепей в каждой микрофибрилле первичной стенки, природу и расположение нарушения, а также наличие или отсутствие прямого контакта микрофибрилл. Здесь мы сообщаем об экспериментах по рассеянию рентгеновских лучей и нейтронов, а также спектроскопических экспериментах, направленных на решение этих вопросов и приводящих к предложенной структуре микрофибрилл целлюлозы первичной стенки. Характеристика структуры, содержащей столько беспорядка, представляла необычные проблемы, но беспорядок, по-видимому, является центральным для загадочной способности целлюлозы первичной стенки обеспечивать высокую прочность, но при этом позволять и контролировать рост.
РЕЗУЛЬТАТЫ
Диаметр микрофибриллы и расстояние от малоуглового рассеяния нейтронов
Когда микрофибриллы коллагена или целлюлозы фиксированного диаметра упакованы в массивы с некоторой степенью регулярности, может наблюдаться малоугловая дифракция рентгеновских лучей или нейтронов (брэгговское рассеяние) от самих микрофибрилл (Hulmes et al., 1995; Fernandes et al. др., 2011). Это рассеяние происходит под меньшими углами, чем обычная дифракция на кристаллических плоскостях, и накладывается на малоугловое рассеяние, возникающее не из-за дифракции, а из-за текстуры и формы фибрилл и пустот между ними (форм-фактор).Если упаковка не является достаточно регулярной, чтобы образовать кристаллическую сверхрешетку, наблюдается только один пик Брэгга, соответствующий расстоянию между центрами фибрилл (Hulmes et al., 1995; Kennedy et al., 2007a).
Используя малоугловое рассеяние рентгеновских лучей (SAXS), наблюдали брэгговское отражение от микрофибрилл целлюлозы внутри интактных клеточных стенок колленхимы сельдерея. Расстояние между микрофибриллами увеличивается при гидратации (Kennedy et al., 2007b). Моделирование этих результатов показало, что наблюдаемый пик Брэгга может быть вызван рядом комбинаций расстояния между микрофибриллами и их диаметра.Следовательно, невозможно однозначно определить диаметр микрофибрилл (Kennedy et al., 2007a).
В древесине ели микрофибриллы слишком тесно агрегированы, чтобы обеспечить адекватный контраст для малоуглового брэгговского рассеяния. Эту проблему удалось решить путем дейтерирования поверхности и использования нейтронов вместо рентгеновских лучей (Fernandes et al., 2011). Природа контраста различается между источниками рентгеновского излучения и нейтронов. Рентгеновский контраст зависит в основном от локальной электронной плотности, тогда как контраст рассеяния нейтронов зависит от локального элементного состава и плотности.Одним из преимуществ этого метода является очень разные сечения рассеяния нейтронов водородом и дейтерием, что позволяет ввести контраст, например, за счет дейтериевого обмена в оксиде дейтерия (D 2 O; Horikawa et al., 2009; Matthews и др., 2011).
Экваториальный пик Брэгга был хорошо виден на изображениях малоуглового рассеяния нейтронов (МУРН) древесины ели, насыщенной D 2 O (Fernandes et al., 2011). Интенсивность пика Брэгга уменьшалась, а его расстояние ( q ) от центра дифракционной картины увеличивалось при сушке до тех пор, пока на картине SANS от полностью сухой дейтерированной древесины пик не был слабо виден при q = 2.1 нм −1 . Низкий контраст в этих условиях соответствовал контакту между соседними дейтерированными на поверхности микрофибриллами, так что расстояние между центрами 3,0 нм соответствовало по существу среднему диаметру микрофибрилл (Fernandes et al., 2011).
Подобные эксперименты SANS были проведены на стенках клеток из колленхимы сельдерея и на целлюлозе, выделенной из этих клеточных стенок (рис. 1). Пик брэгговского рассеяния в области q = 1 нм -1 , наложенный на экспоненциальный фон от некогерентного рассеяния, был очевиден как для стенок клеток, так и для изолированной целлюлозы, насыщенной чистым D 2 O, чистой водой или ряд смесей (рис.1; Дополнительный рис. S1). В соответствии с рассеянием от целлюлозы, а не от других полимеров, не наблюдалось пика Брэгга для смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Которая соответствует теоретической плотности длины рассеяния. целлюлозы (рис. 1).
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна.C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл. D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив.Вертикальный масштаб больше, чем в C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна. C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл.D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив. Вертикальная шкала больше, чем C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
В отличие от древесины ели, сухая целлюлоза сельдерея дает слабый пик Брэгга без дейтерирования поверхности микрофибрилл. В сухом состоянии пик Брэгга был при q = 2.11 нм -1 для целлюлозы в клеточных стенках или q = 2,14 нм -1 для изолированной целлюлозы (рис. 1), что соответствует межцентровым расстояниям в реальном пространстве 3,0 и 2,9 нм, соответственно. . Интенсивность Брэгга была менее 1% от наблюдаемой после регидратации. Этот очень низкий контраст соответствовал плотной упаковке микрофибрилл, вероятно, без каких-либо других полимеров между ними по крайней мере на некоторой части их длины. Расстояние от центра до центра должно приблизительно соответствовать среднему диаметру микрофибриллы.Несмотря на свою низкую интенсивность, пик Брэгга был легко различим на картинах МУРН, поскольку было очень мало помех от некогерентного рассеяния (рис. 1).
В гидратированном состоянии значение q пика Брэгга от микрофибрилл in situ в стенках клеток будет соответствовать реальному расстоянию ( d ) 7,1 ± 1,3 нм -1 , в то время как q Значение пика Брэгга для гидратированной целлюлозы соответствует реальному интервалу 6.0 ± 1,5 нм -1 . Эти номинальные расстояния d не означают, что все микрофибриллы разделены до такой степени при гидратации. Гидратация также значительно увеличивала контраст SANS за счет введения воды или D 2 O между микрофибриллами и, в случае D 2 O, за счет дейтерирования их поверхностей и нецеллюлозных полимеров. Отсюда следует, что при любом распределении расстояний между микрофибриллами более широкие промежутки будут значительно перепредставлены в шаблоне SANS. Номинальные интервалы, наблюдаемые после гидратации, немного превышали расчетные (Kennedy et al., 2007b) методом SAXS, что согласуется с разной основой рентгеновского контраста, но не было никакой разницы между расстояниями, наблюдаемыми в сухом состоянии с рентгеновскими лучами (Kennedy et al., 2007a) и с нейтронами (рис. 1). . Хотя на основе рентгеновских данных невозможно было сделать вывод о том, что расстояние, наблюдаемое в сухом состоянии, соответствует микрофибриллам, находящимся в прямом контакте друг с другом, этот вывод кажется вероятным на основе объединенных данных SAXS и SANS. . Гидратация четко разделяла микрофибриллы, но, вероятно, в разной степени по их длине (Kennedy et al., 2007b), возможно, в зависимости от сродства нецеллюлозных полисахаридов к воде (Jarvis, 1992).
Размеры микрофибрилл и нарушения в результате широкоугольного рассеяния рентгеновских лучей
Широкоугольные картины рассеяния рентгеновских лучей или нейтронов от природной целлюлозы достаточно хорошо разрешены для кристаллографического решения структуры, только если целлюлоза исключительно упорядочена и хорошо ориентирована и если микрофибриллы достаточно толстые, чтобы минимизировать уширение Шеррера, которое возникает в результате ограниченное количество плоскостей решетки.Большая часть целлюлозы колленхимы сельдерея была относительно хорошо ориентирована внутри клеточных стенок (рис. 2). Вклад широкоугольного рентгеновского рассеяния (WAXS) от незначительной изотропно ориентированной фракции целлюлозы был удален на стадии вычитания фона. Существенный вклад изотропного рассеяния на связанной воде также был удален при вычитании фона. Рассеяние от воды было идентифицировано экспериментом, в котором менялось содержание воды (дополнительный рис. S2).
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок WAXS из хорошо ориентированного целлюлозного компонента был размыт в радиальном направлении из-за уширения Шеррера, уширения, связанного с беспорядком, или обоих. Отражение 200 было при q = 15,5 нм -1 , что соответствует несколько большему расстоянию между слоями, чем в кристаллических целлюлозах, и отражения 1-10 и 110 были объединены. Эти особенности наблюдались на плохо ориентированных дифрактограммах от того, что было названо целлюлозой IV 1 (Dinand et al., 1996) или неупорядоченную целлюлозу Iβ (Wada et al., 2004) в первичных клеточных стенках. Для единообразия в этой статье используется обозначение решетки Iβ, но без всякого указания на то, что эта форма представляет собой наиболее близкую аналогию со средней структурой.
Количественная оценка беспорядка и вычисление размеров Шеррера по уширению этих отражений было затруднено из-за сильного перекрытия между отражениями 1-10 и 110 и чрезвычайно низкой интенсивности отражения 400, качественного индикатора большего бокового беспорядка, чем в хвойная древесина.При длительном времени сбора данных (16–24 ч) можно было получить достаточное отношение сигнал / шум для оценки уширения отражения 400. Гидратация за счет увеличения моноклинного угла и, таким образом, разделения отражений 1-10 и 110 упростила моделирование соответствующей ширины этих двух отражений в дифракционной картине от древесины ели (Fernandes et al., 2011). Однако в целлюлозе сельдерея гидратация немного увеличивала интервал [200] d , но не увеличивала значительно угол моноклинности.Процедура Fernandes et al. (2011) был использован для выделения эффекта одной формы бокового беспорядка, которая приводит к асимметрии радиальных пиков. Предполагая, что оставшиеся симметричные радиальные ширины отражений 1-10 и 110 были равны, наиболее подходящее значение было на 22% больше, чем радиальная ширина отражения 200 при моноклинном угле 95 °. Исходя из этого, средние размеры элементарной ячейки в сухом состоянии составляли a = 0,81 нм, b = 0,80 нм, c = 1,03 нм (с сохранением индекса целлюлозы Iβ для ясности, даже если а> b).Меньшие углы моноклинности и большая ширина, или наоборот, почти так же точно соответствовали наблюдаемым профилям интенсивности.
Еще одна форма беспорядка, которая, наряду с небольшими размерами решетки, способствует радиальному уширению, была смоделирована как «паракристаллическая» (Fernandes et al., 2011), что означает, что ширина отражений увеличивается с квадратом порядка отражения. или с q 2 d . Используя наиболее подходящие данные и построив график ширины δ q отражений против q 2 d , как описано (Fernandes et al., 2011), ширина симметричных компонент рефлексов 200 и 400 давала фактор остаточной разупорядоченности г = 0,03 и размер Шеррера 3,2 нм, перпендикулярный плоскостям колец в сухом состоянии. Средняя ширина отражений 1-10 и 110 лежит явно выше δ q по сравнению с графиком q 2 d для отражений 200 и 400. Подразумевается, что средний размер Шеррера в направлениях, нормальных к плоскостям решетки 1-10 и 110 был меньше, чем средний размер Шеррера в направлении, нормальном к плоскости решетки 200, как в еловой целлюлозе.Однако низкая интенсивность отражения 400 и, как следствие, сложность оценки вклада, связанного с беспорядком, в общее уширение сделали этот вывод менее надежным, чем для еловой целлюлозы.
Характеристика неупорядоченных компонентов с помощью дейтерирования и широкоугольного рассеяния нейтронов
В кристаллографическом исследовании целлюлозы Iα и Iβ (Nishiyama et al., 2002, 2003) разница в дифракции нейтронов, обусловленная полным высокотемпературным дейтерированием гидроксильных групп, использовалась для определения положений протонов и, следовательно, геометрии водородных связей. .Также должна быть возможность использовать сильное различие между рассеянием нейтронов дейтерием и протонами для определения вклада в рассеяние неупорядоченных областей, доступных для дейтерирования в более мягких условиях температуры окружающей среды, используемых в экспериментах с SANS и инфракрасным преобразованием Фурье (FTIR). Как может быть показано с помощью FTIR (см. Ниже), гидроксильные группы на нецеллюлозных полисахаридах доступны для дейтерирования, как и обращенные наружу гидроксилы на поверхности целлюлозы, если они не заблокированы агрегацией (Horikawa et al., 2009). Для неупорядоченных областей внутри микрофибрилл, чтобы допустить D 2 O, они должны иметь более широкие интервалы решетки, чем кристаллическая целлюлоза (Matthews et al., 2011).
На рис. 3А показано, что наблюдались видимые изменения в картине дифракции нейтронов при обратимом дейтерировании при температуре окружающей среды. Изменения включали увеличение или уменьшение интенсивности отдельных отражений, а также изменения диффузного рассеяния. В принципе, изменение интенсивности для любого отражения может быть положительным, отрицательным или нейтральным в зависимости от влияния дейтерирования на структурный фактор.В качестве руководства, интенсивности основных экваториальных и осевых отражений для целлюлозы Iβ в нативной (H) и пердейтерированной (D) формах были рассчитаны с использованием Shelxl-97 (Sheldrick, 2008) в пакете WinGX (Farrugia, 1999), из статическая модель, использующая опубликованные координаты атомов (Nishiyama et al., 2002). Из трех основных экваториальных отражений, определяющих геометрию боковой упаковки целлюлозных цепей, влияние дейтерирования на интенсивность было предсказано положительным для 200 и сильно отрицательным для 110 и 1-10.Было предсказано, что осевое отражение 004 будет увеличиваться по интенсивности при дейтерировании, в то время как отражение 001 отсутствовало как в D, так и в H-формах. Эти прогнозы согласуются с наблюдаемыми нейтронографическими изображениями для высококристаллической целлюлозы Iβ (Nishiyama et al., 2002).
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина). Ось волокна вертикальна.B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина).Ось волокна вертикальна. B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Наблюдаемый эффект поверхностного дейтерирования микрофибрилл был положительным для отражений 200 и 004, отрицательным для 110 и нейтральным для 1-10.Профиль вычитания (D — H) для экваториальных отражений (рис. 3B) показал, что их положение и ширина, соответствующие цепям, доступным для дейтерирования, не заметно отличались от фракции, которая не подвергалась дейтерированию. В азимутальном направлении экваториальная интенсивность показывала более широкий нижний хвост распределения ориентации в D, чем в H-дифракционной картине (рис. 3E), подразумевая, что некоторые из цепей, доступных для дейтерирования, были менее хорошо ориентированы, чем недоступные цепочки целлюлозы.
На оси волокна (рис. 3D) отражение 004 в D-форме было центрировано при немного меньшем q , чем в H-форме, что означает, что некоторые из цепочек, доступных для дейтерирования, были более протяженными, чем недоступные цепи. Разница в среднем расстоянии d составляла всего около 0,5%, а радиальная ширина этого отражения была одинаковой как для D, так и для H-формы. Отражение 002 было сильнее в D-форме, а отражение 001, отсутствующее в H-дифракционной картине, появилось в профиле разности осей.Наличие этого отражения 001 после дейтерирования означает, что некоторые из дейтерированных цепей не имели такого же продольного смещения, как чистая целлюлоза Iβ или целлюлоза Iα.
Почти изотропные изменения диффузного рассеяния при дейтерировании привели к появлению широкого кольца повышенной интенсивности в области 15 нм -1 < q <20 нм -1 , а ниже 15 нм -1 , была уменьшена диффузная интенсивность (рис. 3В). Изотропные компоненты, ответственные за эти особенности диффузного рассеяния, включают нецеллюлозные компоненты, которые обмениваются с D 2 O и небольшими количествами остаточной связанной воды.
Эксперименты по спиновой диффузии протонов и пространственные отношения между упорядоченными и неупорядоченными цепями
Скорость, с которой ядерный магнетизм уравновешивается за счет спиновой диффузии между наноразмерными доменами, может использоваться для оценки расстояния между этими доменами. Эксперименты по спиновой диффузии протонов, основанные на этом принципе, были применены к древесине (Newman, 1992; Altaner et al., 2006) и бактериальной целлюлозе (Masuda et al., 2003). Использовалось двумерное (2D) представление этого эксперимента (Fernandes et al., 2011), чтобы изучить расположение упорядоченных и неупорядоченных областей в еловой целлюлозе, используя различия в их пространственном отделении от лигнина. Совершенно другой эксперимент по спин-диффузии 13 C на соломе кукурузы ( Zea mays ) (Foston et al., 2012) привел к аналогичным выводам.
Целлюлоза колленхимы сельдерея не лигнифицирована, но содержит небольшие остаточные количества гемицеллюлоз глюкоманнана и ксилоглюкана, которые, обладая относительно высокой термической подвижностью, могут использоваться вместо лигнина в качестве резервуаров спина протонов.На рисунке 4 показан 2D-спектр этого эксперимента. Перекрестные пики указывают на домены, которые удовлетворяют двум условиям: (1) домены различаются по подвижности и, следовательно, по намагниченности в начале периода смешения спинов протонов в последовательности импульсов; и (2) домены достаточно разделены в пространстве, чтобы потребовалось измеримое время, несколько миллисекунд, для уравновешивания намагниченности протонов посредством спиновой диффузии. Приведенное ниже назначение сигналов основано на Sturcová et al. (2004).
Рисунок 4.
2D-представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, отслеживаемых по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены. [См. Онлайн-статью для цветной версии этого рисунка.]
Рис. 4.
Двумерное представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, мониторинг которого проводился по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены.[Цветную версию этого рисунка см. В онлайн-статье.]
Одномерные спектры C-ЯМР 13 были аналогичны тем, которые наблюдались ранее (Sturcová et al., 2004), с сигналами C-4 от кристаллической целлюлозы (89 –90 частей на миллион [частей на миллион]) и неупорядоченной целлюлозы (84–85 частей на миллион) в примерном соотношении 1: 2. В 2D-спектре присутствовали кросс-пики между C-4 кристаллической целлюлозы и C-4 неупорядоченной целлюлозы, но они были относительно слабыми. Также присутствовал кросс-пик между C-6 кристаллической целлюлозы (65 ppm) и C-4 неупорядоченной целлюлозы (84–85 ppm).Перекрестные пики от гемицеллюлозы C-1 (101 ppm) и C-4 (83 ppm) до кристаллической целлюлозы C-4 были сильнее, чем от неупорядоченной целлюлозы C-4, и перекрестные пики от гемицеллюлозы C-1 и C-4 к кристаллической целлюлозе С-6 были еще сильнее. Наиболее сильные кросс-пики связаны с сигналами C-2, C-5 и C-6 от неупорядоченной целлюлозы при 73, 75 и 62 м.д., но каждый из этих сигналов, очевидно, также содержит вклад гемицеллюлоз и, следовательно, не может быть определен. однозначно трактуется.
Эти наблюдения согласуются с моделями, в которых неупорядоченная и кристаллическая целлюлоза не смешиваются, а находятся в непосредственной пространственной близости внутри каждой микрофибриллы, а гемицеллюлозы расположены вне микрофибрилл, вероятно, частично за пределами каждого небольшого пучка микрофибрилл, но разница в кросс-пике интенсивность кристаллической и неупорядоченной целлюлозы предполагает, что одна группа гемицеллюлоз была в тесной связи с поверхностями микрофибрилл.
Поляризованный дейтерированный FTIR и модели водородной связи
Поляризованный FTIR предоставляет информацию о направлении водородных связей. Исследования FTIR на древесине ели показали, что гидроксильные группы на неупорядоченной целлюлозе на поверхности микрофибрилл были доступны для дейтериевого обмена, за исключением тех случаев, когда микрофибриллы были плотно агрегированы вместе. Также были доступны гидроксильные группы на гемицеллюлозах. Обмен дейтерия сдвигает полосу растяжения, связанную с каждой гидроксильной группой, на более низкую частоту в 1 раз.34, из-за большей массы атома дейтерия, но сохраняет порядок полос, потому что структура не изменилась (Sturcová et al., 2003).
На фиг. 5 показано, что дейтерирование либо клеточных стенок колленхимы сельдерея, либо изолированной целлюлозы было обширным, но меньше, чем количество, предсказанное из соотношения 2: 1 неупорядоченной целлюлозы к кристаллической, оцененного с использованием сигналов C-4 в спектрах ЯМР. Образец полос растяжения O-D легче всего сравнить с рисунком полос растяжения O-H, если шкала частот искусственно сдвинута в 1 раз.34 (рис. 5). Эти две модели явно отличались. Полосы растяжения O-D были менее ориентированы в продольном направлении, чем полосы растяжения O-H, показывая, что меньшее количество водородных связей в дейтерированной фракции было направлено вдоль оси микрофибрилл, а больше — наружу. Как и в еловой целлюлозе (Fernandes et al., 2011), дейтерирование снизило интенсивность выступающего плеча на 3400 см -1 в области растяжения O-H, что соответствует гидроксильным группам, более слабо связанным водородными связями и более направленным наружу, чем в среднем.После сдвига оси частот на на 1,34 раза максимум довольно широкой группы перекрывающихся полос валентных колебаний OD совпал с этим плечом, которое первоначально было отнесено к аномальной части Iα-подобного спектра целлюлозы (Sturcová et al., 2004), но позже было показано, что в еловой целлюлозе они соответствуют недоступным гидроксильным группам в неупорядоченных областях (Fernandes et al., 2011). Отнесение к неупорядоченным доменам, а не к целлюлозе Iα, подтверждается данными, представленными здесь.Только часть интенсивности на 3 400 см -1 была смещена дейтерированием, подразумевая, что некоторые из этих неупорядоченных цепей в целлюлозе колленхимы сельдерея доступны для дейтерирования, а некоторые — нет.
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой. Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию.Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В онлайн-статье.]
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой.Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию. Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В статье в Интернете.]
ОБСУЖДЕНИЕ
Структура из микрофибриллы
Целлюлоза, которую можно условно назвать кристаллической, и неупорядоченная целлюлоза, отличная от известных кристаллических форм целлюлозы Iα и Iβ, были идентифицированы всеми использованными методами.Из этого не следует, что одни и те же неупорядоченные фракции наблюдались всеми этими методами (т.е. что цепи, идентифицированные как неупорядоченные, в одних и тех же отношениях отличались от известных кристаллических форм). Наблюдались конформационный беспорядок, беспорядок упаковки и беспорядок водородных связей.
Спектры 13 C-ЯМР целлюлозы колленхимы сельдерея (рис. 4) идентифицировали цепи, которые отличались по конформации как от целлюлозы Iα, так и от целлюлозы Iβ, имеющие некоторую свободу вращения вокруг связи C5-C6 и имеющие C-6 В конформации преобладают гош-гош ( gg ) и гош-транс ( gt ) формы (Sturcová et al., 2004), как и в еловой целлюлозе (Fernandes et al., 2011), хотя соотношение gg — gt значительно отличалось от еловой (Sturcová et al., 2004). Спектры дейтерирования FTIR целлюлозы сельдерея показали, что некоторые домены имели паттерны водородных связей, отличные от наблюдаемых в целлюлозе Iα и целлюлозе Iβ, с пониженной средней прочностью водородных связей (более высокая средняя частота растяжения O-H) и меньшей общей тенденцией к осевой ориентации водорода. Изменение ориентации водородных связей было очевидным как в цепях, доступных для дейтериевого обмена, так и в недоступных цепях, и соответствовало переходу от транс-гош ( tg ) конформации C-6, обнаруженной в кристаллической целлюлозе, к gg . и конформеры GT .Подобные наблюдения на еловой целлюлозе были рационализированы с точки зрения модели, в которой ядро каждой микрофибриллы было занято в основном кристаллической целлюлозой, а поверхность микрофибрилл была занята цепями целлюлозы с несколько иной конформацией и водородными связями, некоторые из которых были доступны для обмена дейтерия, а другие были представлены. недоступны из-за боковой агрегации микрофибрилл. Подобная модель согласуется с FTIR-спектрами сельдерея (рис. 5), более ранними данными ЯМР (Sturcová et al., 2004), а также данные спин-диффузионного ЯМР, представленные здесь (рис. 4). По данным ЯМР спин-решеточной релаксации 13 C (Viëtor et al., 2002), целлюлоза колленхимы сельдерея имеет более высокую долю конформера C-6 gg, чем еловая целлюлоза. Размеры элементарной ячейки значительно различались между этими двумя видами целлюлозы, среднее расстояние между листами было больше в целлюлозе колленхимы сельдерея, а осевой размер был немного меньше. Молекулярное моделирование предложило связь между конформацией C-6 и шагом решетки (Matthews et al., 2012). Как и в случае с еловой целлюлозой, было невозможно и, возможно, бессмысленно различать формы целлюлозы Iα и Iβ с помощью ЯМР при наличии такого большого конформационного нарушения (Matthews et al., 2011).
Модели микрофибрилл с неупорядоченными поверхностями неоднократно предлагались на основе данных ЯМР (Newman, 1992; Viëtor et al., 2002; Bergenstråhle et al., 2008), но было неясно, как они должны согласовываться с полученными моделями. от дифракции рентгеновских лучей, которая более чувствительна к структуре упаковки цепей, чем к конформации цепи или водородным связям.Данные широкоугольного рассеяния нейтронов (WANS) (рис.3) показали, что в доменах, доступных для дейтерирования, цепи были упакованы латерально таким же образом и почти с такими же средними расстояниями, как и в кристаллических областях, несмотря на различия в конформации. и водородные связи, продемонстрированные спектроскопическими экспериментами, несмотря на значительные статистические различия в упаковке цепей повсюду в микрофибрилле. Повсюду в каждой микрофибрилле листы цепочек были уложены друг на друга с более широким средним интервалом, чем в более кристаллических целлюлозах.Дальнейшее следствие состоит в том, что большинство цепей, доступных для дейтерирования, были расположены на поверхностях микрофибрилл, а не в «аморфных» сегментах микрофибриллы, занимающих всю ширину микрофибриллы, потому что цепи внутри таких сегментов должны быть неплотно упакованы, чтобы вода могла стекать. входить.
Судя по осевому отражению 004, доступные для дейтерирования цепи оказались немного более вытянутыми в продольном направлении, чем недоступные цепи, возможно, из-за скручивания микрофибрилл.Продольное смещение было частично нарушено в цепочках, доступных для дейтерирования, что усложняло различение форм Iα и Iβ.
Широкий диапазон расстояний между боковыми цепями в нецеллюлозной фракции, доступный для дейтерирования, может быть выведен из диффузной, неориентированной части нейтронографических изображений. Вода, связанная с нецеллюлозной фракцией, рассеивала рентгеновские лучи аналогично жидкой воде, предполагая аналогичную функцию распределения пар, но из-за пониженной тепловой подвижности величина ее вклада в структуру WAXS была больше, чем ожидалось для объемной жидкой воды.
Диаметр и форма микрофибриллы
Данные SANS предполагают диаметр микрофибрилл, приблизительно равный межцентровому расстоянию от 2,9 до 3,0 нм в сухом состоянии. Предполагая круглое поперечное сечение и учитывая относительно рыхлую упаковку цепей, наблюдаемую при широкоугольном рассеянии, этот номинальный диаметр будет соответствовать 21–22 цепям. Микрофибриллы, конечно, не имеют круглой формы, и фактическое количество цепочек зависит от формы поперечного сечения.Наблюдения WANS подтвердили, что поверхностные цепочки располагались на той же неупорядоченной решетке, что и внутренние цепочки, и должны быть включены в расчеты размеров Шеррера из WAXS.
Межцентровое расстояние SANS и поперечные размеры по Шерреру были аналогичны тем, которые наблюдались для еловой целлюлозы (Fernandes et al., 2011). Размер [200] Шеррера был больше, чем среднее значение размеров [1-10] и [110], что согласуется с моделью прямоугольной микрофибриллы (Matthews et al., 2006; Fernandes et al., 2011), который имеет относительно гидрофобные [200] грани кристаллов, открытые сверху и снизу (рис. 6), вместо диагональных [1-10] и [110] граней кристаллов, открытых в целлюлозах водорослей (Baker et al. ., 1997). Однако опора для прямоугольной, а не ромбовидной модели менее надежна, чем для ели, из-за сложности коррекции беспорядка с использованием очень слабого отражения 400 и из-за проблем, присущих оценке отдельных ширины перекрывающихся 1-10 и 110 отражений.Учитывая наблюдаемое обширное нарушение, нельзя даже с уверенностью сказать, что микрофибриллы первичной стенки имеют постоянную форму поперечного сечения.
Рисунок 6.
Модели поперечного сечения микрофибрилл колленхимы сельдерея. A, 24-цепочечная ромбовидная модель, 2,9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться.Расчетные длины столбцов [200], [1-10] и [110] были следующими, соответственно: модель A, 2,5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 6.
Модели для поперечного сечения микрофибрилл колленхимы сельдерея. A, модель ромбовидной формы с 24 цепями, 2.9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться. Расчетные длины столбцов [200], [1-10] и [110] были соответственно следующими: модель A, 2.5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В статье в Интернете.]
Прямоугольное поперечное сечение с уложенными друг на друга листами из трех цепей будет соответствовать одной трети цепей, находящихся в центральное положение в каждом листе, конформационно сходное с кристаллическими целлюлозами, как наблюдалось с помощью ЯМР.Наблюдение единственного малоуглового пика Брэгга согласуется со слегка диспергированными расстояниями между микрофибриллами, которые можно было бы ожидать от примерно прямоугольных микрофибрилл, возможно, скрученных, находящихся в прерывистом контакте друг с другом. Могут быть приспособлены как постоянные, так и слегка изменяющиеся диаметры микрофибрилл, а также небольшое количество микрофибрилл, значительно отличающихся от доминирующих размеров.
Число цепей на микрофибриллу имеет отношение к структуре комплексов, которые синтезируют целлюлозу во время роста.Если количество цепей кратно шести, что соответствует геометрии синтетического комплекса, тогда стопка из восьми трехцепочечных листов (24 цепи) даст наиболее близкое совпадение с размером Шеррера 200 (рис. 6B), как и для еловая целлюлоза (Fernandes et al., 2011). Однако в нашем случае неопределенность довольно велика, и уровень беспорядка упаковки предполагает, что возможны нерегулярные и переменные сечения. Модель из 18 цепей (6 × 3) не исключена для еловой целлюлозы и согласуется с данными, представленными здесь (рис.6C), если размер 200 по Шерреру был увеличен дополнительными цепями гемицеллюлозы, как это было предложено для ели (Fernandes et al., 2011), или увеличен путем наложения одной микрофибриллы поверх другой на короткие расстояния.
Кристалличность
Что означает «кристалличность», зависит от методов, используемых для обнаружения компонентов, которые отличаются от кристаллической целлюлозы. Между спектроскопическими методами обнаружения цепей, отличающихся по конформации от общепринятых форм кристаллической целлюлозы и, следовательно, демонстрирующих различные модели водородных связей, наблюдалась некоторая степень согласованности.Это включало нецеллюлозные полисахариды, и, в частности, в спектрах FTIR было трудно провести четкое различие между вкладом нецеллюлозных полисахаридов и некоторой неупорядоченной целлюлозы.
Эксперименты с дейтерированием WANS показали, что эти конформационные различия мало влияют на упаковку целлюлозных цепей или дифракцию. Структура микрофибрилл явно содержала значительный беспорядок в боковой упаковке цепей, но этот беспорядок распространялся по всей структуре и не ограничивался доступными для дейтерия поверхностями.Кристалличность, оцениваемая дифракционными методами, — это не то же самое, что кристалличность, оцениваемая спектроскопически. В обоих случаях кажущаяся кристалличность будет увеличиваться с увеличением диаметра микрофибрилл, но как прямое следствие уменьшения уширения по Шерреру в дифракционных экспериментах и из-за уменьшения отношения поверхности к объему в спектроскопических экспериментах, которые позволяют различать поверхностные цепи.
Агрегация микрофибрилл
Агрегация микрофибрилл в более крупные пучки хорошо известна для древесины хвойных пород (Wickholm et al., 1998; Фахлен и Сальмен, 2005 г .; Fernandes et al., 2011). Фибриллярные единицы шириной от 5 до 10 нм, иногда до 40 нм, наблюдались с помощью электронной микроскопии (McCann et al., 1990; Sugimoto et al., 2000), но их интерпретация осложняется возможным включением нецеллюлозных полимеров. . Представленные здесь данные включают две прямые линии доказательств агрегации микрофибрилл первичной стенки в колленхиме. Доказательства малоуглового брэгговского рассеяния рентгеновских лучей (Kennedy et al., 2007a) и нейтронов ясно показали, что агрегация существует. Диапазон расстояний между микрофибриллами было трудно сделать вывод, потому что интенсивность рассеяния сильно увеличивалась с разделением микрофибрилл, так что хорошо разделенные микрофибриллы были чрезмерно представлены на диаграммах рассеяния. Низкий контраст МУРН в сухом состоянии и сходство межцентрового расстояния МУРН со средними размерами Шеррера WAXS предполагают, что некоторые сегменты микрофибрилл оставались в прямом контакте друг с другом, тогда как другие сегменты микрофибрилл становились хорошо разделенными при гидратации.
Спектры дейтерирования FTIR как от стенок клеток колленхимы, так и от изолированной целлюлозы включали полосы растяжения гидроксила с центром на 3 400 см -1 с нейтральной поляризацией. Эти гидроксильные полосы растяжения были отнесены к гидроксильным группам, отличным от всех групп Iα и Iβ кристаллических форм целлюлозы, более ориентированным наружу по ориентации и частично, но не полностью доступным для дейтерирования. Принимая во внимание также данные SANS и ЯМР, соответствующие гидроксильные группы могут быть отнесены к поверхностям микрофибрилл.Поскольку эти поверхности были только частично доступны для D 2 O, отсюда следует, что микрофибриллы были агрегированы вместе на части своей длины с границами раздела, непроницаемыми для воды. Эти особенности были обнаружены в древесине ели, но были также различия: древесина ели имеет разные нецеллюлозные полимеры, что, по-видимому, приводит к более тесному связыванию между поверхностями микрофибрилл и к напряжениям, которые искажают геометрию элементарной ячейки при гидратации (Fernandes et al., 2011).
Полоса валентных колебаний гидроксила 3,400 см -1 в FTIR-спектре, перемещенная до 2,540 см -1 за счет обмена D 2 O, ранее была неправильно отнесена к аберрантной форме целлюлозы Iα (Sturcová et al., 2004), но, по-видимому, является маркером поверхностных цепей и наличия агрегации микрофибрилл, если эти поверхностные цепи недоступны для дейтерирования. Интенсивность этой полосы сильно уменьшилась, когда целлюлоза из колленхимы сельдерея была подвергнута гидротермальному отжигу, что привело к слиянию микрофибрилл в более крупные кристаллические единицы с уменьшенным соотношением поверхности к объему (Sturcová et al., 2004). Интенсивность FTIR на уровне 3400 см или около него -1 была зарегистрирована в препаратах клеточных стенок, например, арабидопсиса дикого типа ( Arabidopsis thaliana ; MacKinnon et al., 2006), предполагая, что эта форма агрегации не является уникальной особенностью целлюлозы колленхимы сельдерея, но более широко распространена в первичных клеточных стенках.
Влияние на рост растений
Размер и структура микрофибрилл первичной стенки дают важные ключи к разгадке природы ферментных комплексов, ответственных за ориентированный синтез целлюлозы и, следовательно, за направленную экспансию клеток. Размножение растительных клеток требует разрушения сети нековалентно сшитых микрофибрилл в клеточной стенке.Понимание топологии этой сети, следовательно, необходимо, и недавно принятая идея о том, что ксилоглюканы просто покрывают и связывают поверхности микрофибрилл, была поставлена под сомнение (Bootten et al., 2004; Park and Cosgrove, 2012a). Данные спин-диффузии ЯМР (рис. 4) показали, что некоторые цепи гемицеллюлозы, вероятно, ксилоглюканы, располагались близко к поверхностям микрофибрилл и, таким образом, вероятно, лежали между микрофибриллами внутри агрегатов микрофибрилл. Однако другие сегменты микрофибрилл, по-видимому, находятся в прямом контакте без промежуточных полимеров, демонстрируя нерегулярный характер агрегации, которого можно было бы ожидать, если микрофибриллы были скрученными и нецилиндрическими, и предполагая, что взаимодействия целлюлоза-целлюлоза, а также ксилоглюкан-целлюлоза должны быть разбиты на отдельные агрегированные микрофибриллы.Др. Механизмы необходимы для отделения одного пучка микрофибрилл от следующего во время роста путем разрушения ксилоглюкана или др. Мостиков между пучками микрофибрилл (Anderson et al., 2010). Агрегация микрофибрилл вдоль части их длины, в то время как другие части их длины остаются свободными для разделения или в них вставлены другие полисахариды, поднимает интересные вопросы о действии экспансинов (Cosgrove, 2005) и о том, как действует действие определенных глюканов. гидролазы взаимодействуют с таковой ксилоглюкан-специфической эндоглюканазы (Park and Cosgrove, 2012b).
Интересен высокий уровень беспорядка на границах раздела, наиболее важных для роста, между микрофибриллами и нецеллюлозными полимерами, а также между одной микрофибриллой и другой. Примеры прочных биологических материалов из царства животных показывают, что неупорядоченные домены могут вносить вклад в ударную вязкость (высокую энергию разрушения) при внешнем стрессе (van Beek et al., 2002). В первичных клеточных стенках устойчивость к разрушению должна сочетаться с контролируемой ферментативно-опосредованной продуктивностью, по-видимому, на неупорядоченных границах раздела, что обеспечивает рост.
Последствия разложения целлюлозы
Доступность микрофибрилл целлюлозы для целлюлаз является важным фактором превращения лигноцеллюлозы в биотопливо и еще более важна в случае нелгнифицированной биомассы. Нестабильность целлюлозы часто связывают с ее кристалличностью (Arantes and Saddler, 2010), а «декристаллизацию» иногда называют целью процессов предварительной обработки (Beckham et al., 2011), без полного согласия относительно того, что означают эти термины.Степень, в которой микрофибриллы целлюлозы имеют относительно гидрофобную [200] открытую поверхность, будет положительно влиять на начальную стадию деградации целлюлазами, которые специфически связываются с этой поверхностью (Dagel et al., 2011; Liu et al., 2011). Было бы интересно узнать больше о специфичности связывания оксидаз целлюлозы GH61 / CBM33 (Forsberg et al., 2011; Quinlan et al., 2011), некоторые из которых также связываются с одной и той же поверхностью (Li et al., 2012). Агрегация микрофибрилл целлюлозы — это фактор сопротивляемости, который заслуживает большего внимания, и теперь к нему можно подойти с помощью экспериментов по дейтерированию FTIR.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Было выделенонитей колленхимы из зрелых черешков сельдерея ( Apium graveolens ), клеточные стенки и их целлюлозная фракция были приготовлены, как описано (Sturcová et al., 2004), и хранились в сухом виде. Подробнее см. Дополнительные материалы и методы S1.
SANS
ЭкспериментыSANS были выполнены на канале D11 в Институте Лауэ-Ланжевена на клеточных стенках колленхимы сельдерея и изолированной целлюлозе.Подробнее об эксперименте см. Дополнительные материалы и методы S1.
WAXS
дифрактограммы рентгеновских лучей получали при температуре окружающей среды с использованием дифрактометра Rigaku R-axis / RAPID для рентгеновских пластин с источником излучения с молибденовой рентгеновской эмиссией Kα (λ = 0,07071 нм). Пучок коллимировался до диаметра 0,5 мм. Образцы имели толщину 0,7 мм в направлении, параллельном пучку, а их другие размеры превышали диаметр пучка. Дифрактограммы были собраны в режиме перпендикулярного пропускания, за исключением экспериментов с наклоном для измерения осевых отражений.Расстояние от центра дифракционной картины было выражено как q = 4πsinθ / λ, где 2θ — угол рассеяния. Для получения дополнительных сведений об экспериментальных методах и методах обработки данных (Fernandes et al., 2011) см. Дополнительные материалы и методы S1.
WANS
Дифракция нейтронов проводилась на канале D19 в Институте Лауэ-Ланжевена при средней длине волны нейтронов 0,242 нм и расстоянии от образца до детектора 0,756 м. Образцы размером 25 × 15 × 1 мм собирали из нескольких выровненных нитей колленхимы и выдерживали при контролируемом давлении воды или паров D 2 O в проточном контейнере со стенками из алюминиевой фольги.Подробнее см. Дополнительные материалы и методы S1.
ЯМР
A 1 H Эксперимент по спиновой диффузии Гольдмана-Шена с детектированием по спектру 13 C после кросс-поляризации был проведен на клеточных стенках колленхимы сельдерея, гидратированных 0,2 см. 3 г −1 D 2 O. Экспериментальные процедуры и анализ данных, включая 2D-представление данных, были такими, как описано (Fernandes et al., 2011).
Дейтерирование FTIR
Стенки клеток колленхимы сельдерея и изолированную целлюлозу помещали в проточную ячейку с окнами из фторида бария и заменяли воздухом, насыщенным О 2 О, до тех пор, пока спектры не перестали изменяться. Затем газовую линию переключали на азот, тщательно осушая, пропуская через сухое молекулярное сито 4А, а затем пентоксид фосфора (Sicapent; Aldrich). Спектры FTIR собирали с помощью спектрометра Thermo Nicolet Nexus, оборудованного приставкой для микроскопа Nicolet Continuum, имеющей охлаждаемый жидким азотом детектор теллурида кадмия ртути (Fernandes et al., 2011).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Rigaku, Ltd. за предоставленный инструмент и Fujiang Zhang за получение некоторых спектров FTIR.
Глоссарий
SAXS
Малоугловое рассеяние рентгеновских лучей
D 2 O
SANS
Малоугловое рассеяние нейтронов
WAXS 9407
широкоугольное рассеивание
WANS
широкоугольное рассеяние нейтронов
FTIR
Инфракрасное преобразование Фурье
2D
ppm
LITERATURE CITED
(
2006
)Пространственные отношения между полимерами в ели ситкинской: исследования спиновой диффузии протонов
.Holzforschung
60
:665
—673
(
2010
)Визуализация в реальном времени переориентации целлюлозы во время расширения клеточной стенки в корнях Arabidopsis
.Plant Physiol
152
:787
—796
(
2009
)Выяснение механизмов сборки и субъединичного взаимодействия целлюлозосинтазного комплекса вторичных клеточных стенок Arabidopsis
.J Biol Chem
284
:3833
—3841
(
1997
)Атомно-силовая микроскопия высокого разрешения микрокристаллов нативной целлюлозы I Valonia
.J Struct Biol
119
:129
—138
(
2011
)Происхождение стойкости биомассы на молекулярном уровне: свободные энергии декристаллизации для четырех обычных полиморфов целлюлозы
.J Phys Chem B
115
:4118
—4127
(
2008
)Динамика границ раздела целлюлоза-вода: времена спин-решеточной релаксации ЯМР, рассчитанные на основе атомистического компьютерного моделирования
.J Phys Chem B
112
:2590
—2595
(
2004
)Твердотельный 13 C-ЯМР-спектроскопия показывает, что ксилоглюканы в первичных клеточных стенках маша (Vigna radiata L.) происходят в разных доменах: новая модель взаимодействий ксилоглюкан-целлюлоза в клеточной стенке
.J Exp Bot
55
:571
—583
(
2009
)Растения контролируют свойства и работу своих органов посредством ориентации фибрилл целлюлозы в их клеточных стенках
.Интегр Комп Биол
49
:69
—79
(
2005
)Рост клеточной стенки растений
.Nat Rev Mol Cell Biol
6
:850
—861
(
2011
)Визуализация на месте отдельных углеводсвязывающих модулей на микрофибриллах целлюлозы
.J Phys Chem B
115
:635
—641
(
1996
)Паренхиматозная клеточная целлюлоза из жома сахарной свеклы: получение и свойства
.Целлюлоза
3
:183
—188
(
2005
)Распределение пор и матрицы в стенке волокна, выявленное с помощью атомно-силовой микроскопии и анализа изображений
.Биомакромолекулы
6
:433
—438
(
1999
)Пакет WinGX для кристаллографии низкомолекулярных монокристаллов
.J Appl Cryst
32
:837
—838
(
2011
)Наноструктура микрофибрилл целлюлозы в древесине ели
.Proc Natl Acad Sci USA
108
:E1195
—E1203
(
2011
)Расщепление целлюлозы белком CBM33
.Protein Sci
20
:1479
—1483
(
2012
)Твердотельное селективное (13) C возбуждение и спин-диффузионный ЯМР для определения пространственных размеров в стенках растительных клеток
.J Agric Food Chem
60
:1419
—1427
(
2000
)Характеристика поперечных связей между микрофибриллами целлюлозы и их появление во время элонгационного роста в эпикотиле гороха
.Физиология растительных клеток
41
:486
—494
(
2011
)Кортикальные микротрубочки оптимизируют кристалличность клеточной стенки для обеспечения однонаправленного роста Arabidopsis
.Завод J
66
:915
—928
(
1999
)Выражение паттерна в растениях: сочетание молекулярных и основанных на исчислении биофизических парадигм
.Am J Bot
86
:1059
—1076
и другие. (
2012
)Кристалличность микрофибрилл целлюлозы снижается за счет мутации С-концевых остатков трансмембранной области CESA1A903V и CESA3T942I синтазы целлюлозы
.Proc Natl Acad Sci USA
109
:4098
—4103
(
2007
)Устойчивость биомассы: инженерные установки и ферменты для производства биотоплива
.Наука
315
:804
—807
(
2009
)Различия между сортами в размерах микрофибрилл целлюлозы, наблюдаемые с помощью инфракрасной спектроскопии
.Целлюлоза
16
:1
—8
(
1995
)Радиальная упаковка, порядок и беспорядок в фибриллах коллагена
.Biophys J
68
:1661
—1670
(
1992
)Контроль толщины клеточных стенок колленхимы пектинами
.Планта
187
:218
—220
(
2011
)Стенки растительных клеток: супрамолекулярные ансамбли
.Food Hydrocoll
25
:257
—262
(
2007a
)Диаметр микрофибрилл в целлюлозе колленхимы сельдерея: рассеяние рентгеновских лучей и данные ЯМР
.Целлюлоза
14
:235
—246
(
2007b
)Влияние гидратации на расстояние между микрофибриллами целлюлозы первичной стенки: исследование малоуглового рентгеновского рассеяния
.Целлюлоза
14
:401
—408
(
2008
)Выявление структурного и функционального разнообразия клеточных стенок растений
.Curr Opin Plant Biol
11
:308
—313
(
2001
)Рентгеновская структура мерсеризованной целлюлозы II при разрешении 1 Å
.Биомакромолекулы
2
:410
—416
(
2012
)Структурная основа нацеливания на субстрат и катализа грибковыми полисахаридмонооксигеназами
.Структура
20
:1051
—1061
(
2011
)Целлобиогидролаза гидролизует кристаллическую целлюлозу на гидрофобных поверхностях
.J Biol Chem
286
:11195
—11201
(
2006
)Структура и анизотропия клеточной стенки в procuste , мутанте целлюлозосинтазы Arabidopsis thaliana
.Планта
224
:438
—448
(
2005
)Расширение клеточной стенки приводит к координатному разделению параллельных микрофибрилл: данные сканирующей электронной микроскопии и атомно-силовой микроскопии
.Завод J
43
:181
—190
(
2003
)Твердотельный 13 C и 1 H спин-диффузионный ЯМР-анализ структуры микрофибрилл бактериальной целлюлозы
.Твердотельный ядерный магнитный резонон
23
:198
—212
(
2012
)Сравнение моделирования бета целлюлозы I с тремя силовыми полями углеводов
.J Chem Theory Comput
8
:735
—748
(
2011
)Высокотемпературные свойства целлюлозы I
.J Phys Chem B
115
:2155
—2166
(
2006
)Исследования микрокристаллической целлюлозы Ibeta
с помощью компьютерного моделирования.Углеводородный остаток
341
:138
—152
(
1990
)Прямая визуализация поперечных связей в первичной клеточной стенке растения
.J Cell Sci
96
:323
—334
(
2008
)Структура и биосинтез пектина
.Curr Opin Plant Biol
11
:266
—277
(
1992
)Ядерно-магнитно-резонансное исследование пространственных соотношений между химическими компонентами в стенках деревянных клеток
.Holzforschung
46
:205
—210
(
2009
)Структура и свойства микрофибриллы целлюлозы
.J Wood Sci
55
:241
—249
(
2002
)Кристаллическая структура и система водородных связей в целлюлозе Ibeta по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
124
:9074
—9082
(
2003
)Кристаллическая структура и система водородных связей в целлюлозе I (альфа) по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
125
:14300
—14306
(
2008
)Генетические доказательства того, что активность синтазы целлюлозы влияет на организацию кортикального массива микротрубочек
.Plant Physiol
147
:1723
—1734
(
2012a
)Изменения биомеханических свойств клеточной стенки у ксилоглюкан-дефицитного мутанта xxt1 / xxt2 Arabidopsis
.Plant Physiol
158
:465
—475
(
2012b
)Пересмотренная архитектура первичных клеточных стенок, основанная на биомеханических изменениях, индуцированных субстрат-специфическими эндоглюканазами
.Plant Physiol
158
:1933
—1943
и другие. (
2011
)Анализ окислительного разложения целлюлозы металлоферментом меди, который использует компоненты биомассы
.Proc Natl Acad Sci USA
108
:15079
—15084
и другие.(
2012
)CHITINASE-LIKE1 / POM-POM1 и его гомолог CTL2 представляют собой взаимодействующие с глюканом белки, важные для биосинтеза целлюлозы у Arabidopsis
.Plant Cell
24
:589
—607
(
2008
)Краткая история SHELX
.Acta Crystallogr A
64
:112
—122
(
2006
)Синтез целлюлозы у высших растений
.Annu Rev Cell Dev Biol
22
:53
—78
(
2004
)Структурные детали кристаллической целлюлозы из высших растений
.Биомакромолекулы
5
:1333
—1339
(
2003
)Поляризованная колебательная спектроскопия волоконных полимеров: водородные связи в целлюлозе II
.Биомакромолекулы
4
:1589
—1595
(
2000
)Новые методы позволяют проводить сравнительный анализ ориентации микротрубочек, текстуры стенок и скорости роста интактных корней Arabidopsis
.Plant Physiol
124
:1493
—1506
(
2009
)Динамическая координация систем цитоскелета и клеточной стенки во время морфогенеза растительных клеток
.Curr Biol
19
:R800
—R811
(
2002
)Молекулярная структура шелка драглайна пауков: складывание и ориентация белкового остова
.Proc Natl Acad Sci USA
99
:10266
—10271
(
2002
)Конформационные особенности кристаллической целлюлозы высших растений
.Завод J
30
:721
—731
(
2004
)Полиморфизм целлюлозы семейства I: повторное исследование целлюлозы IVI
.Биомакромолекулы
5
:1385
—1391
(
1998
)Определение некристаллических форм в целлюлозе I методом спектроскопии ЯМР CP / MAS C-13
.Углеводородный остаток
312
:123
—129
(
2007
)Организация боковых цепей пектинового арабинана и галактана в ассоциации с микрофибриллами целлюлозы в первичных клеточных стенках и связанных моделях предусматривается
.J Exp Bot
58
:1795
—1802
Заметки автора
© Американское общество биологов растений, 2013 г. Все права защищены.
© Автор (ы) 2013. Опубликовано Oxford University Press от имени Американского общества биологов растений. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0 /), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
Структура микрофибрилл целлюлозы в первичных клеточных стенках колленхимы | Физиология растений
Аннотация
В первичных стенках растущих клеток растений глюкозно-полимерная целлюлоза собрана в длинные микрофибриллы диаметром несколько нанометров. Жесткость и ориентация этих микрофибрилл контролируют рост клеток; Следовательно, синтез целлюлозы является ключевым фактором роста и морфогенеза растений.Колленхима сельдерея ( Apium graveolens ) является полезной модельной системой для изучения структуры микрофибрилл первичной стенки, поскольку ее микрофибриллы ориентированы с необычной однородностью, что облегчает проведение спектроскопических и дифракционных экспериментов. Используя комбинацию методов рассеяния рентгеновских лучей и нейтронов с колебательной спектроскопией и спектроскопией ядерного магнитного резонанса, мы показываем, что микрофибриллы колленхимы сельдерея имели средний диаметр от 2,9 до 3,0 нм, с наиболее вероятной структурой, содержащей 24 цепи в поперечном сечении, расположенные в восьми атомах водорода. -связанные листы из трех цепей с обширным беспорядком в боковой упаковке, конформации и водородной связи.Аналогичная структура из 18 цепей и структуры из 24 цепей разной формы хуже соответствовали данным. Конформационный беспорядок в основном ограничивался поверхностными цепями, а беспорядок в упаковке цепей — нет. То есть по положению и ориентации поверхностные цепи соответствовали неупорядоченной решетке, составляющей ядро каждой микрофибриллы. Были доказательства того, что соседние микрофибриллы нековалентно агрегированы вместе на части своей длины, предполагая, что необходимость разрушения этих агрегатов может быть сдерживающим фактором роста и гидролиза целлюлозы для производства биотоплива.
Рост и форма растений контролируются точно ориентированным расширением стенок отдельных клеток. Движущая сила для размножения клеток является осмотической, но скорость и направление экспансии контролируются механическими свойствами клеточной стенки (Szymanski and Cosgrove, 2009). Расширяющиеся первичные клеточные стенки представляют собой нанокомпозитные материалы, в которых длинные микрофибриллы целлюлозы диаметром несколько нанометров проходят через гидратированную матрицу ксилоглюканов, пектинов и других полимеров (Knox, 2008; Mohnen, 2008; Szymanski and Cosgrove, 2009; Scheller. и Ульвсков, 2010).Нативные микрофибриллы целлюлозы частично кристаллические (Nishiyama, 2009; Fernandes et al., 2011). Раньше считалось, что целлюлоза первичной стенки имеет уникальную кристаллическую структуру, называемую целлюлозой IV 1 (Dinand et al., 1996), но данные ЯМР предполагают наличие форм, подобных лучше охарактеризованным кристаллическим формам целлюлозы Iα и Iβ вместе с крупными кристаллическими формами целлюлозы. количества менее упорядоченной целлюлозы (Wickholm et al., 1998; Sturcová et al., 2004; Wada et al., 2004). Тем не менее, целлюлоза гораздо более упорядочена, чем любой другой компонент первичной клеточной стенки (Bootten et al., 2004), в соответствии с его ключевой ролью обеспечения силы и контроля роста.
Жесткость клеточной стенки наибольшая в направлении микрофибрилл целлюлозы, где рост является направленным, а преобладающая ориентация микрофибрилл обычно поперечна направлению роста (Green, 1999; MacKinnon et al., 2006; Szymanski and Cosgrove, 2009 г.). Расширение клеточной стенки требует либо увеличения расстояния между микрофибриллами (Marga et al., 2005), либо проскальзывания между ними (Cosgrove, 2005), либо того и другого, и микрофибриллы переориентируются в направлении роста (Anderson et al., 2010). Полимерные поперечные мостики между микрофибриллами (McCann et al., 1990), как полагают, противостоят этим деформациям наноструктуры клеточной стенки и, таким образом, контролируют скорость роста. До недавнего времени наибольшее внимание уделялось мостиковым соединениям ксилоглюканов, водородным связям с поверхностями микрофибрилл (Scheller, Ulvskov, 2010). Однако есть свидетельства того, что не все ксилоглюканы расположены надлежащим образом (Fujino et al., 2000; Park and Cosgrove, 2012a) и что могут быть задействованы другие мостиковые полимеры (Zykwinska et al., 2007). Также было высказано предположение, что пучки агрегированных микрофибрилл, а не отдельные микрофибриллы, могут быть ключевыми структурными единицами в первичных клеточных стенках (Anderson et al., 2010), как в древесине (Fahlén and Salmén, 2005; Fernandes et al., 2011). ). В таком случае отдельные микрофибриллы могут соединяться между пучками микрофибрилл. Таким образом, рост растительных клеток не совсем понятен, и нам нужна дополнительная информация о том, как контролируется ориентация целлюлозы, и о природе мостиковых полимеров, о поверхностях целлюлозы, с которыми эти полимеры связываются, и о сцеплении между поверхностями микрофибрилл, которые могут опосредовать агрегацию.
Микрофибриллы целлюлозы синтезируются на поверхности клетки большими ферментными комплексами, имеющими гексагональную симметрию, иногда называемыми «розетками» (Somerville, 2006). Каждый комплекс содержит несколько синтаз целлюлозы, которые различаются между первичными клеточными стенками и древесиной, хотя внешний вид комплексов похож (Somerville, 2006; Atanassov et al., 2009). Одновременный синтез с одного и того же конца всех цепей в микрофибрилле нативной целлюлозы является причиной их параллельности (Nishiyama et al., 2002, 2003), в отличие от энтропийно предпочтительной антипараллельной структуры, обнаруженной в искусственных целлюлозах, таких как вискоза (Langan et al., 2001). Очевидно, что количество цепей в микрофибрилле и количество целлюлозосинтаз в синтетическом комплексе взаимосвязаны. Принято считать, что количество цепочек делится на шесть, что соответствует симметрии гексагональной розетки и 36-цепочечным моделям (Himmel et al., 2007), ограниченным гидрофильными гранями кристаллов [110] и [1-10], поскольку в водорослевых целлюлозах (Bergenstråhle et al., 2008), получили широкое распространение. Сборка и ориентация целлюлозы взаимосвязаны, поскольку некоторые мутанты целлюлозосинтазы имеют фенотипы, дефектные по ориентации целлюлозы и растительной форме, а также обедненные содержанием целлюлозы (Paredez et al., 2008). У некоторых других мутантных линий, по-видимому, затрагивается кристалличность микрофибрилл (Fujita et al., 2011; Harris et al., 2012; Sánchez-Rodríguez et al., 2012).
Следовательно, детальное понимание структуры микрофибрилл целлюлозы первичной стенки поможет нам понять синтез целлюлозы, а также рост и структурную механику живых растений (Burgert and Fratzl, 2009).Первичные клеточные стенки и их целлюлозные скелеты также влияют на такие характеристики качества пищевых продуктов, как хрусткость салата из овощей и яблок ( Malus domestica ; Jarvis, 2011). Когда биотопливо производится из лигноцеллюлозной биомассы, лигнификация приводит к устойчивости (Himmel et al., 2007), но некоторые типы клеток у видов Miscanthus , просо ( Panicum virgatum ) и пожнивные остатки имеют только первичные стенки. без лигнина, и сопротивляемость этому зависит от природы микрофибрилл целлюлозы (Beckham et al., 2011).
Относительно подробная структура недавно была предложена для микрофибрилл древесины ели ( Picea spp.) (Fernandes et al., 2011), которые имеют диаметр 3,0 нм, что позволяет разместить только около 24 цепочек целлюлозы. Данные рентгеновской дифракции подтверждают «прямоугольную» форму (Matthews et al., 2006), ограниченную гранями [010] и [200]. Наблюдался значительный беспорядок, увеличивающийся к поверхности, и микрофибриллы были агрегированы в пучки размером примерно 15-20 нм в поперечнике, причем некоторые, но не все, боковые границы раздела были устойчивы к воде (Fernandes et al., 2011). Неупорядоченные домены являются особенностью других прочных биологических материалов, таких как шелк паука (van Beek et al., 2002).
Следовательно, представляет интерес, могут ли какие-либо из этих свойств древесной целлюлозы также быть обнаружены в микрофибриллах целлюлозы первичных (растущих) клеточных стенок. Было бы особенно полезно охарактеризовать нарушение, которое, как известно, присутствует в микрофибриллах первичной стенки, то есть определить, чем целлюлоза, которая не определяется как «кристаллическая», отличается от кристаллической целлюлозы.Многие эксперименты, ведущие к созданию структуры древесной целлюлозы, зависели от исключительно однородной ориентации микрофибрилл целлюлозы (Sturcová et al., 2004; Fernandes et al., 2011). Однако в растущих клеточных стенках микрофибриллы ориентированы неравномерно. Когда микрофибриллы сначала откладываются на внутренней поверхности первичной клеточной стенки, их ориентация обычно перпендикулярна направлению роста, но когда клеточная стенка расширяется, микрофибриллы переориентируются таким образом, что распределение ориентации интегрируется по толщине расширенной клеточной стенки. клеточная стенка становится все более близкой к случайной (Cosgrove, 2005; MacKinnon et al., 2006).
Эта техническая проблема не относится к клеточным стенкам колленхимы сельдерея ( Apium graveolens ), которые похожи по составу на другие первичные клеточные стенки, но имеют их микрофибриллы, ориентированные относительно равномерно вдоль оси клетки (Sturcová et al., 2004 ; Kennedy et al., 2007a, 2007b). Некоторая структурная информация о целлюлозе колленхимы сельдерея уже была получена в результате спектроскопических экспериментов и экспериментов по рассеянию (Sturcová et al., 2004; Kennedy et al., 2007a, 2007b), подтверждая беспорядок, ожидаемый в целлюлозе первичной стенки.Некоторые из этих экспериментов были аналогичны тому, что было сделано с еловой целлюлозой (Fernandes et al., 2011), но недостаточно данных, чтобы указать количество цепей в каждой микрофибрилле первичной стенки, природу и расположение нарушения, а также наличие или отсутствие прямого контакта микрофибрилл. Здесь мы сообщаем об экспериментах по рассеянию рентгеновских лучей и нейтронов, а также спектроскопических экспериментах, направленных на решение этих вопросов и приводящих к предложенной структуре микрофибрилл целлюлозы первичной стенки. Характеристика структуры, содержащей столько беспорядка, представляла необычные проблемы, но беспорядок, по-видимому, является центральным для загадочной способности целлюлозы первичной стенки обеспечивать высокую прочность, но при этом позволять и контролировать рост.
РЕЗУЛЬТАТЫ
Диаметр микрофибриллы и расстояние от малоуглового рассеяния нейтронов
Когда микрофибриллы коллагена или целлюлозы фиксированного диаметра упакованы в массивы с некоторой степенью регулярности, может наблюдаться малоугловая дифракция рентгеновских лучей или нейтронов (брэгговское рассеяние) от самих микрофибрилл (Hulmes et al., 1995; Fernandes et al. др., 2011). Это рассеяние происходит под меньшими углами, чем обычная дифракция на кристаллических плоскостях, и накладывается на малоугловое рассеяние, возникающее не из-за дифракции, а из-за текстуры и формы фибрилл и пустот между ними (форм-фактор).Если упаковка не является достаточно регулярной, чтобы образовать кристаллическую сверхрешетку, наблюдается только один пик Брэгга, соответствующий расстоянию между центрами фибрилл (Hulmes et al., 1995; Kennedy et al., 2007a).
Используя малоугловое рассеяние рентгеновских лучей (SAXS), наблюдали брэгговское отражение от микрофибрилл целлюлозы внутри интактных клеточных стенок колленхимы сельдерея. Расстояние между микрофибриллами увеличивается при гидратации (Kennedy et al., 2007b). Моделирование этих результатов показало, что наблюдаемый пик Брэгга может быть вызван рядом комбинаций расстояния между микрофибриллами и их диаметра.Следовательно, невозможно однозначно определить диаметр микрофибрилл (Kennedy et al., 2007a).
В древесине ели микрофибриллы слишком тесно агрегированы, чтобы обеспечить адекватный контраст для малоуглового брэгговского рассеяния. Эту проблему удалось решить путем дейтерирования поверхности и использования нейтронов вместо рентгеновских лучей (Fernandes et al., 2011). Природа контраста различается между источниками рентгеновского излучения и нейтронов. Рентгеновский контраст зависит в основном от локальной электронной плотности, тогда как контраст рассеяния нейтронов зависит от локального элементного состава и плотности.Одним из преимуществ этого метода является очень разные сечения рассеяния нейтронов водородом и дейтерием, что позволяет ввести контраст, например, за счет дейтериевого обмена в оксиде дейтерия (D 2 O; Horikawa et al., 2009; Matthews и др., 2011).
Экваториальный пик Брэгга был хорошо виден на изображениях малоуглового рассеяния нейтронов (МУРН) древесины ели, насыщенной D 2 O (Fernandes et al., 2011). Интенсивность пика Брэгга уменьшалась, а его расстояние ( q ) от центра дифракционной картины увеличивалось при сушке до тех пор, пока на картине SANS от полностью сухой дейтерированной древесины пик не был слабо виден при q = 2.1 нм −1 . Низкий контраст в этих условиях соответствовал контакту между соседними дейтерированными на поверхности микрофибриллами, так что расстояние между центрами 3,0 нм соответствовало по существу среднему диаметру микрофибрилл (Fernandes et al., 2011).
Подобные эксперименты SANS были проведены на стенках клеток из колленхимы сельдерея и на целлюлозе, выделенной из этих клеточных стенок (рис. 1). Пик брэгговского рассеяния в области q = 1 нм -1 , наложенный на экспоненциальный фон от некогерентного рассеяния, был очевиден как для стенок клеток, так и для изолированной целлюлозы, насыщенной чистым D 2 O, чистой водой или ряд смесей (рис.1; Дополнительный рис. S1). В соответствии с рассеянием от целлюлозы, а не от других полимеров, не наблюдалось пика Брэгга для смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Которая соответствует теоретической плотности длины рассеяния. целлюлозы (рис. 1).
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна.C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл. D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив.Вертикальный масштаб больше, чем в C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна. C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл.D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив. Вертикальная шкала больше, чем C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
В отличие от древесины ели, сухая целлюлоза сельдерея дает слабый пик Брэгга без дейтерирования поверхности микрофибрилл. В сухом состоянии пик Брэгга был при q = 2.11 нм -1 для целлюлозы в клеточных стенках или q = 2,14 нм -1 для изолированной целлюлозы (рис. 1), что соответствует межцентровым расстояниям в реальном пространстве 3,0 и 2,9 нм, соответственно. . Интенсивность Брэгга была менее 1% от наблюдаемой после регидратации. Этот очень низкий контраст соответствовал плотной упаковке микрофибрилл, вероятно, без каких-либо других полимеров между ними по крайней мере на некоторой части их длины. Расстояние от центра до центра должно приблизительно соответствовать среднему диаметру микрофибриллы.Несмотря на свою низкую интенсивность, пик Брэгга был легко различим на картинах МУРН, поскольку было очень мало помех от некогерентного рассеяния (рис. 1).
В гидратированном состоянии значение q пика Брэгга от микрофибрилл in situ в стенках клеток будет соответствовать реальному расстоянию ( d ) 7,1 ± 1,3 нм -1 , в то время как q Значение пика Брэгга для гидратированной целлюлозы соответствует реальному интервалу 6.0 ± 1,5 нм -1 . Эти номинальные расстояния d не означают, что все микрофибриллы разделены до такой степени при гидратации. Гидратация также значительно увеличивала контраст SANS за счет введения воды или D 2 O между микрофибриллами и, в случае D 2 O, за счет дейтерирования их поверхностей и нецеллюлозных полимеров. Отсюда следует, что при любом распределении расстояний между микрофибриллами более широкие промежутки будут значительно перепредставлены в шаблоне SANS. Номинальные интервалы, наблюдаемые после гидратации, немного превышали расчетные (Kennedy et al., 2007b) методом SAXS, что согласуется с разной основой рентгеновского контраста, но не было никакой разницы между расстояниями, наблюдаемыми в сухом состоянии с рентгеновскими лучами (Kennedy et al., 2007a) и с нейтронами (рис. 1). . Хотя на основе рентгеновских данных невозможно было сделать вывод о том, что расстояние, наблюдаемое в сухом состоянии, соответствует микрофибриллам, находящимся в прямом контакте друг с другом, этот вывод кажется вероятным на основе объединенных данных SAXS и SANS. . Гидратация четко разделяла микрофибриллы, но, вероятно, в разной степени по их длине (Kennedy et al., 2007b), возможно, в зависимости от сродства нецеллюлозных полисахаридов к воде (Jarvis, 1992).
Размеры микрофибрилл и нарушения в результате широкоугольного рассеяния рентгеновских лучей
Широкоугольные картины рассеяния рентгеновских лучей или нейтронов от природной целлюлозы достаточно хорошо разрешены для кристаллографического решения структуры, только если целлюлоза исключительно упорядочена и хорошо ориентирована и если микрофибриллы достаточно толстые, чтобы минимизировать уширение Шеррера, которое возникает в результате ограниченное количество плоскостей решетки.Большая часть целлюлозы колленхимы сельдерея была относительно хорошо ориентирована внутри клеточных стенок (рис. 2). Вклад широкоугольного рентгеновского рассеяния (WAXS) от незначительной изотропно ориентированной фракции целлюлозы был удален на стадии вычитания фона. Существенный вклад изотропного рассеяния на связанной воде также был удален при вычитании фона. Рассеяние от воды было идентифицировано экспериментом, в котором менялось содержание воды (дополнительный рис. S2).
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок WAXS из хорошо ориентированного целлюлозного компонента был размыт в радиальном направлении из-за уширения Шеррера, уширения, связанного с беспорядком, или обоих. Отражение 200 было при q = 15,5 нм -1 , что соответствует несколько большему расстоянию между слоями, чем в кристаллических целлюлозах, и отражения 1-10 и 110 были объединены. Эти особенности наблюдались на плохо ориентированных дифрактограммах от того, что было названо целлюлозой IV 1 (Dinand et al., 1996) или неупорядоченную целлюлозу Iβ (Wada et al., 2004) в первичных клеточных стенках. Для единообразия в этой статье используется обозначение решетки Iβ, но без всякого указания на то, что эта форма представляет собой наиболее близкую аналогию со средней структурой.
Количественная оценка беспорядка и вычисление размеров Шеррера по уширению этих отражений было затруднено из-за сильного перекрытия между отражениями 1-10 и 110 и чрезвычайно низкой интенсивности отражения 400, качественного индикатора большего бокового беспорядка, чем в хвойная древесина.При длительном времени сбора данных (16–24 ч) можно было получить достаточное отношение сигнал / шум для оценки уширения отражения 400. Гидратация за счет увеличения моноклинного угла и, таким образом, разделения отражений 1-10 и 110 упростила моделирование соответствующей ширины этих двух отражений в дифракционной картине от древесины ели (Fernandes et al., 2011). Однако в целлюлозе сельдерея гидратация немного увеличивала интервал [200] d , но не увеличивала значительно угол моноклинности.Процедура Fernandes et al. (2011) был использован для выделения эффекта одной формы бокового беспорядка, которая приводит к асимметрии радиальных пиков. Предполагая, что оставшиеся симметричные радиальные ширины отражений 1-10 и 110 были равны, наиболее подходящее значение было на 22% больше, чем радиальная ширина отражения 200 при моноклинном угле 95 °. Исходя из этого, средние размеры элементарной ячейки в сухом состоянии составляли a = 0,81 нм, b = 0,80 нм, c = 1,03 нм (с сохранением индекса целлюлозы Iβ для ясности, даже если а> b).Меньшие углы моноклинности и большая ширина, или наоборот, почти так же точно соответствовали наблюдаемым профилям интенсивности.
Еще одна форма беспорядка, которая, наряду с небольшими размерами решетки, способствует радиальному уширению, была смоделирована как «паракристаллическая» (Fernandes et al., 2011), что означает, что ширина отражений увеличивается с квадратом порядка отражения. или с q 2 d . Используя наиболее подходящие данные и построив график ширины δ q отражений против q 2 d , как описано (Fernandes et al., 2011), ширина симметричных компонент рефлексов 200 и 400 давала фактор остаточной разупорядоченности г = 0,03 и размер Шеррера 3,2 нм, перпендикулярный плоскостям колец в сухом состоянии. Средняя ширина отражений 1-10 и 110 лежит явно выше δ q по сравнению с графиком q 2 d для отражений 200 и 400. Подразумевается, что средний размер Шеррера в направлениях, нормальных к плоскостям решетки 1-10 и 110 был меньше, чем средний размер Шеррера в направлении, нормальном к плоскости решетки 200, как в еловой целлюлозе.Однако низкая интенсивность отражения 400 и, как следствие, сложность оценки вклада, связанного с беспорядком, в общее уширение сделали этот вывод менее надежным, чем для еловой целлюлозы.
Характеристика неупорядоченных компонентов с помощью дейтерирования и широкоугольного рассеяния нейтронов
В кристаллографическом исследовании целлюлозы Iα и Iβ (Nishiyama et al., 2002, 2003) разница в дифракции нейтронов, обусловленная полным высокотемпературным дейтерированием гидроксильных групп, использовалась для определения положений протонов и, следовательно, геометрии водородных связей. .Также должна быть возможность использовать сильное различие между рассеянием нейтронов дейтерием и протонами для определения вклада в рассеяние неупорядоченных областей, доступных для дейтерирования в более мягких условиях температуры окружающей среды, используемых в экспериментах с SANS и инфракрасным преобразованием Фурье (FTIR). Как может быть показано с помощью FTIR (см. Ниже), гидроксильные группы на нецеллюлозных полисахаридах доступны для дейтерирования, как и обращенные наружу гидроксилы на поверхности целлюлозы, если они не заблокированы агрегацией (Horikawa et al., 2009). Для неупорядоченных областей внутри микрофибрилл, чтобы допустить D 2 O, они должны иметь более широкие интервалы решетки, чем кристаллическая целлюлоза (Matthews et al., 2011).
На рис. 3А показано, что наблюдались видимые изменения в картине дифракции нейтронов при обратимом дейтерировании при температуре окружающей среды. Изменения включали увеличение или уменьшение интенсивности отдельных отражений, а также изменения диффузного рассеяния. В принципе, изменение интенсивности для любого отражения может быть положительным, отрицательным или нейтральным в зависимости от влияния дейтерирования на структурный фактор.В качестве руководства, интенсивности основных экваториальных и осевых отражений для целлюлозы Iβ в нативной (H) и пердейтерированной (D) формах были рассчитаны с использованием Shelxl-97 (Sheldrick, 2008) в пакете WinGX (Farrugia, 1999), из статическая модель, использующая опубликованные координаты атомов (Nishiyama et al., 2002). Из трех основных экваториальных отражений, определяющих геометрию боковой упаковки целлюлозных цепей, влияние дейтерирования на интенсивность было предсказано положительным для 200 и сильно отрицательным для 110 и 1-10.Было предсказано, что осевое отражение 004 будет увеличиваться по интенсивности при дейтерировании, в то время как отражение 001 отсутствовало как в D, так и в H-формах. Эти прогнозы согласуются с наблюдаемыми нейтронографическими изображениями для высококристаллической целлюлозы Iβ (Nishiyama et al., 2002).
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина). Ось волокна вертикальна.B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина).Ось волокна вертикальна. B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Наблюдаемый эффект поверхностного дейтерирования микрофибрилл был положительным для отражений 200 и 004, отрицательным для 110 и нейтральным для 1-10.Профиль вычитания (D — H) для экваториальных отражений (рис. 3B) показал, что их положение и ширина, соответствующие цепям, доступным для дейтерирования, не заметно отличались от фракции, которая не подвергалась дейтерированию. В азимутальном направлении экваториальная интенсивность показывала более широкий нижний хвост распределения ориентации в D, чем в H-дифракционной картине (рис. 3E), подразумевая, что некоторые из цепей, доступных для дейтерирования, были менее хорошо ориентированы, чем недоступные цепочки целлюлозы.
На оси волокна (рис. 3D) отражение 004 в D-форме было центрировано при немного меньшем q , чем в H-форме, что означает, что некоторые из цепочек, доступных для дейтерирования, были более протяженными, чем недоступные цепи. Разница в среднем расстоянии d составляла всего около 0,5%, а радиальная ширина этого отражения была одинаковой как для D, так и для H-формы. Отражение 002 было сильнее в D-форме, а отражение 001, отсутствующее в H-дифракционной картине, появилось в профиле разности осей.Наличие этого отражения 001 после дейтерирования означает, что некоторые из дейтерированных цепей не имели такого же продольного смещения, как чистая целлюлоза Iβ или целлюлоза Iα.
Почти изотропные изменения диффузного рассеяния при дейтерировании привели к появлению широкого кольца повышенной интенсивности в области 15 нм -1 < q <20 нм -1 , а ниже 15 нм -1 , была уменьшена диффузная интенсивность (рис. 3В). Изотропные компоненты, ответственные за эти особенности диффузного рассеяния, включают нецеллюлозные компоненты, которые обмениваются с D 2 O и небольшими количествами остаточной связанной воды.
Эксперименты по спиновой диффузии протонов и пространственные отношения между упорядоченными и неупорядоченными цепями
Скорость, с которой ядерный магнетизм уравновешивается за счет спиновой диффузии между наноразмерными доменами, может использоваться для оценки расстояния между этими доменами. Эксперименты по спиновой диффузии протонов, основанные на этом принципе, были применены к древесине (Newman, 1992; Altaner et al., 2006) и бактериальной целлюлозе (Masuda et al., 2003). Использовалось двумерное (2D) представление этого эксперимента (Fernandes et al., 2011), чтобы изучить расположение упорядоченных и неупорядоченных областей в еловой целлюлозе, используя различия в их пространственном отделении от лигнина. Совершенно другой эксперимент по спин-диффузии 13 C на соломе кукурузы ( Zea mays ) (Foston et al., 2012) привел к аналогичным выводам.
Целлюлоза колленхимы сельдерея не лигнифицирована, но содержит небольшие остаточные количества гемицеллюлоз глюкоманнана и ксилоглюкана, которые, обладая относительно высокой термической подвижностью, могут использоваться вместо лигнина в качестве резервуаров спина протонов.На рисунке 4 показан 2D-спектр этого эксперимента. Перекрестные пики указывают на домены, которые удовлетворяют двум условиям: (1) домены различаются по подвижности и, следовательно, по намагниченности в начале периода смешения спинов протонов в последовательности импульсов; и (2) домены достаточно разделены в пространстве, чтобы потребовалось измеримое время, несколько миллисекунд, для уравновешивания намагниченности протонов посредством спиновой диффузии. Приведенное ниже назначение сигналов основано на Sturcová et al. (2004).
Рисунок 4.
2D-представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, отслеживаемых по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены. [См. Онлайн-статью для цветной версии этого рисунка.]
Рис. 4.
Двумерное представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, мониторинг которого проводился по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены.[Цветную версию этого рисунка см. В онлайн-статье.]
Одномерные спектры C-ЯМР 13 были аналогичны тем, которые наблюдались ранее (Sturcová et al., 2004), с сигналами C-4 от кристаллической целлюлозы (89 –90 частей на миллион [частей на миллион]) и неупорядоченной целлюлозы (84–85 частей на миллион) в примерном соотношении 1: 2. В 2D-спектре присутствовали кросс-пики между C-4 кристаллической целлюлозы и C-4 неупорядоченной целлюлозы, но они были относительно слабыми. Также присутствовал кросс-пик между C-6 кристаллической целлюлозы (65 ppm) и C-4 неупорядоченной целлюлозы (84–85 ppm).Перекрестные пики от гемицеллюлозы C-1 (101 ppm) и C-4 (83 ppm) до кристаллической целлюлозы C-4 были сильнее, чем от неупорядоченной целлюлозы C-4, и перекрестные пики от гемицеллюлозы C-1 и C-4 к кристаллической целлюлозе С-6 были еще сильнее. Наиболее сильные кросс-пики связаны с сигналами C-2, C-5 и C-6 от неупорядоченной целлюлозы при 73, 75 и 62 м.д., но каждый из этих сигналов, очевидно, также содержит вклад гемицеллюлоз и, следовательно, не может быть определен. однозначно трактуется.
Эти наблюдения согласуются с моделями, в которых неупорядоченная и кристаллическая целлюлоза не смешиваются, а находятся в непосредственной пространственной близости внутри каждой микрофибриллы, а гемицеллюлозы расположены вне микрофибрилл, вероятно, частично за пределами каждого небольшого пучка микрофибрилл, но разница в кросс-пике интенсивность кристаллической и неупорядоченной целлюлозы предполагает, что одна группа гемицеллюлоз была в тесной связи с поверхностями микрофибрилл.
Поляризованный дейтерированный FTIR и модели водородной связи
Поляризованный FTIR предоставляет информацию о направлении водородных связей. Исследования FTIR на древесине ели показали, что гидроксильные группы на неупорядоченной целлюлозе на поверхности микрофибрилл были доступны для дейтериевого обмена, за исключением тех случаев, когда микрофибриллы были плотно агрегированы вместе. Также были доступны гидроксильные группы на гемицеллюлозах. Обмен дейтерия сдвигает полосу растяжения, связанную с каждой гидроксильной группой, на более низкую частоту в 1 раз.34, из-за большей массы атома дейтерия, но сохраняет порядок полос, потому что структура не изменилась (Sturcová et al., 2003).
На фиг. 5 показано, что дейтерирование либо клеточных стенок колленхимы сельдерея, либо изолированной целлюлозы было обширным, но меньше, чем количество, предсказанное из соотношения 2: 1 неупорядоченной целлюлозы к кристаллической, оцененного с использованием сигналов C-4 в спектрах ЯМР. Образец полос растяжения O-D легче всего сравнить с рисунком полос растяжения O-H, если шкала частот искусственно сдвинута в 1 раз.34 (рис. 5). Эти две модели явно отличались. Полосы растяжения O-D были менее ориентированы в продольном направлении, чем полосы растяжения O-H, показывая, что меньшее количество водородных связей в дейтерированной фракции было направлено вдоль оси микрофибрилл, а больше — наружу. Как и в еловой целлюлозе (Fernandes et al., 2011), дейтерирование снизило интенсивность выступающего плеча на 3400 см -1 в области растяжения O-H, что соответствует гидроксильным группам, более слабо связанным водородными связями и более направленным наружу, чем в среднем.После сдвига оси частот на на 1,34 раза максимум довольно широкой группы перекрывающихся полос валентных колебаний OD совпал с этим плечом, которое первоначально было отнесено к аномальной части Iα-подобного спектра целлюлозы (Sturcová et al., 2004), но позже было показано, что в еловой целлюлозе они соответствуют недоступным гидроксильным группам в неупорядоченных областях (Fernandes et al., 2011). Отнесение к неупорядоченным доменам, а не к целлюлозе Iα, подтверждается данными, представленными здесь.Только часть интенсивности на 3 400 см -1 была смещена дейтерированием, подразумевая, что некоторые из этих неупорядоченных цепей в целлюлозе колленхимы сельдерея доступны для дейтерирования, а некоторые — нет.
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой. Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию.Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В онлайн-статье.]
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой.Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию. Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В статье в Интернете.]
ОБСУЖДЕНИЕ
Структура из микрофибриллы
Целлюлоза, которую можно условно назвать кристаллической, и неупорядоченная целлюлоза, отличная от известных кристаллических форм целлюлозы Iα и Iβ, были идентифицированы всеми использованными методами.Из этого не следует, что одни и те же неупорядоченные фракции наблюдались всеми этими методами (т.е. что цепи, идентифицированные как неупорядоченные, в одних и тех же отношениях отличались от известных кристаллических форм). Наблюдались конформационный беспорядок, беспорядок упаковки и беспорядок водородных связей.
Спектры 13 C-ЯМР целлюлозы колленхимы сельдерея (рис. 4) идентифицировали цепи, которые отличались по конформации как от целлюлозы Iα, так и от целлюлозы Iβ, имеющие некоторую свободу вращения вокруг связи C5-C6 и имеющие C-6 В конформации преобладают гош-гош ( gg ) и гош-транс ( gt ) формы (Sturcová et al., 2004), как и в еловой целлюлозе (Fernandes et al., 2011), хотя соотношение gg — gt значительно отличалось от еловой (Sturcová et al., 2004). Спектры дейтерирования FTIR целлюлозы сельдерея показали, что некоторые домены имели паттерны водородных связей, отличные от наблюдаемых в целлюлозе Iα и целлюлозе Iβ, с пониженной средней прочностью водородных связей (более высокая средняя частота растяжения O-H) и меньшей общей тенденцией к осевой ориентации водорода. Изменение ориентации водородных связей было очевидным как в цепях, доступных для дейтериевого обмена, так и в недоступных цепях, и соответствовало переходу от транс-гош ( tg ) конформации C-6, обнаруженной в кристаллической целлюлозе, к gg . и конформеры GT .Подобные наблюдения на еловой целлюлозе были рационализированы с точки зрения модели, в которой ядро каждой микрофибриллы было занято в основном кристаллической целлюлозой, а поверхность микрофибрилл была занята цепями целлюлозы с несколько иной конформацией и водородными связями, некоторые из которых были доступны для обмена дейтерия, а другие были представлены. недоступны из-за боковой агрегации микрофибрилл. Подобная модель согласуется с FTIR-спектрами сельдерея (рис. 5), более ранними данными ЯМР (Sturcová et al., 2004), а также данные спин-диффузионного ЯМР, представленные здесь (рис. 4). По данным ЯМР спин-решеточной релаксации 13 C (Viëtor et al., 2002), целлюлоза колленхимы сельдерея имеет более высокую долю конформера C-6 gg, чем еловая целлюлоза. Размеры элементарной ячейки значительно различались между этими двумя видами целлюлозы, среднее расстояние между листами было больше в целлюлозе колленхимы сельдерея, а осевой размер был немного меньше. Молекулярное моделирование предложило связь между конформацией C-6 и шагом решетки (Matthews et al., 2012). Как и в случае с еловой целлюлозой, было невозможно и, возможно, бессмысленно различать формы целлюлозы Iα и Iβ с помощью ЯМР при наличии такого большого конформационного нарушения (Matthews et al., 2011).
Модели микрофибрилл с неупорядоченными поверхностями неоднократно предлагались на основе данных ЯМР (Newman, 1992; Viëtor et al., 2002; Bergenstråhle et al., 2008), но было неясно, как они должны согласовываться с полученными моделями. от дифракции рентгеновских лучей, которая более чувствительна к структуре упаковки цепей, чем к конформации цепи или водородным связям.Данные широкоугольного рассеяния нейтронов (WANS) (рис.3) показали, что в доменах, доступных для дейтерирования, цепи были упакованы латерально таким же образом и почти с такими же средними расстояниями, как и в кристаллических областях, несмотря на различия в конформации. и водородные связи, продемонстрированные спектроскопическими экспериментами, несмотря на значительные статистические различия в упаковке цепей повсюду в микрофибрилле. Повсюду в каждой микрофибрилле листы цепочек были уложены друг на друга с более широким средним интервалом, чем в более кристаллических целлюлозах.Дальнейшее следствие состоит в том, что большинство цепей, доступных для дейтерирования, были расположены на поверхностях микрофибрилл, а не в «аморфных» сегментах микрофибриллы, занимающих всю ширину микрофибриллы, потому что цепи внутри таких сегментов должны быть неплотно упакованы, чтобы вода могла стекать. входить.
Судя по осевому отражению 004, доступные для дейтерирования цепи оказались немного более вытянутыми в продольном направлении, чем недоступные цепи, возможно, из-за скручивания микрофибрилл.Продольное смещение было частично нарушено в цепочках, доступных для дейтерирования, что усложняло различение форм Iα и Iβ.
Широкий диапазон расстояний между боковыми цепями в нецеллюлозной фракции, доступный для дейтерирования, может быть выведен из диффузной, неориентированной части нейтронографических изображений. Вода, связанная с нецеллюлозной фракцией, рассеивала рентгеновские лучи аналогично жидкой воде, предполагая аналогичную функцию распределения пар, но из-за пониженной тепловой подвижности величина ее вклада в структуру WAXS была больше, чем ожидалось для объемной жидкой воды.
Диаметр и форма микрофибриллы
Данные SANS предполагают диаметр микрофибрилл, приблизительно равный межцентровому расстоянию от 2,9 до 3,0 нм в сухом состоянии. Предполагая круглое поперечное сечение и учитывая относительно рыхлую упаковку цепей, наблюдаемую при широкоугольном рассеянии, этот номинальный диаметр будет соответствовать 21–22 цепям. Микрофибриллы, конечно, не имеют круглой формы, и фактическое количество цепочек зависит от формы поперечного сечения.Наблюдения WANS подтвердили, что поверхностные цепочки располагались на той же неупорядоченной решетке, что и внутренние цепочки, и должны быть включены в расчеты размеров Шеррера из WAXS.
Межцентровое расстояние SANS и поперечные размеры по Шерреру были аналогичны тем, которые наблюдались для еловой целлюлозы (Fernandes et al., 2011). Размер [200] Шеррера был больше, чем среднее значение размеров [1-10] и [110], что согласуется с моделью прямоугольной микрофибриллы (Matthews et al., 2006; Fernandes et al., 2011), который имеет относительно гидрофобные [200] грани кристаллов, открытые сверху и снизу (рис. 6), вместо диагональных [1-10] и [110] граней кристаллов, открытых в целлюлозах водорослей (Baker et al. ., 1997). Однако опора для прямоугольной, а не ромбовидной модели менее надежна, чем для ели, из-за сложности коррекции беспорядка с использованием очень слабого отражения 400 и из-за проблем, присущих оценке отдельных ширины перекрывающихся 1-10 и 110 отражений.Учитывая наблюдаемое обширное нарушение, нельзя даже с уверенностью сказать, что микрофибриллы первичной стенки имеют постоянную форму поперечного сечения.
Рисунок 6.
Модели поперечного сечения микрофибрилл колленхимы сельдерея. A, 24-цепочечная ромбовидная модель, 2,9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться.Расчетные длины столбцов [200], [1-10] и [110] были следующими, соответственно: модель A, 2,5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 6.
Модели для поперечного сечения микрофибрилл колленхимы сельдерея. A, модель ромбовидной формы с 24 цепями, 2.9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться. Расчетные длины столбцов [200], [1-10] и [110] были соответственно следующими: модель A, 2.5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В статье в Интернете.]
Прямоугольное поперечное сечение с уложенными друг на друга листами из трех цепей будет соответствовать одной трети цепей, находящихся в центральное положение в каждом листе, конформационно сходное с кристаллическими целлюлозами, как наблюдалось с помощью ЯМР.Наблюдение единственного малоуглового пика Брэгга согласуется со слегка диспергированными расстояниями между микрофибриллами, которые можно было бы ожидать от примерно прямоугольных микрофибрилл, возможно, скрученных, находящихся в прерывистом контакте друг с другом. Могут быть приспособлены как постоянные, так и слегка изменяющиеся диаметры микрофибрилл, а также небольшое количество микрофибрилл, значительно отличающихся от доминирующих размеров.
Число цепей на микрофибриллу имеет отношение к структуре комплексов, которые синтезируют целлюлозу во время роста.Если количество цепей кратно шести, что соответствует геометрии синтетического комплекса, тогда стопка из восьми трехцепочечных листов (24 цепи) даст наиболее близкое совпадение с размером Шеррера 200 (рис. 6B), как и для еловая целлюлоза (Fernandes et al., 2011). Однако в нашем случае неопределенность довольно велика, и уровень беспорядка упаковки предполагает, что возможны нерегулярные и переменные сечения. Модель из 18 цепей (6 × 3) не исключена для еловой целлюлозы и согласуется с данными, представленными здесь (рис.6C), если размер 200 по Шерреру был увеличен дополнительными цепями гемицеллюлозы, как это было предложено для ели (Fernandes et al., 2011), или увеличен путем наложения одной микрофибриллы поверх другой на короткие расстояния.
Кристалличность
Что означает «кристалличность», зависит от методов, используемых для обнаружения компонентов, которые отличаются от кристаллической целлюлозы. Между спектроскопическими методами обнаружения цепей, отличающихся по конформации от общепринятых форм кристаллической целлюлозы и, следовательно, демонстрирующих различные модели водородных связей, наблюдалась некоторая степень согласованности.Это включало нецеллюлозные полисахариды, и, в частности, в спектрах FTIR было трудно провести четкое различие между вкладом нецеллюлозных полисахаридов и некоторой неупорядоченной целлюлозы.
Эксперименты с дейтерированием WANS показали, что эти конформационные различия мало влияют на упаковку целлюлозных цепей или дифракцию. Структура микрофибрилл явно содержала значительный беспорядок в боковой упаковке цепей, но этот беспорядок распространялся по всей структуре и не ограничивался доступными для дейтерия поверхностями.Кристалличность, оцениваемая дифракционными методами, — это не то же самое, что кристалличность, оцениваемая спектроскопически. В обоих случаях кажущаяся кристалличность будет увеличиваться с увеличением диаметра микрофибрилл, но как прямое следствие уменьшения уширения по Шерреру в дифракционных экспериментах и из-за уменьшения отношения поверхности к объему в спектроскопических экспериментах, которые позволяют различать поверхностные цепи.
Агрегация микрофибрилл
Агрегация микрофибрилл в более крупные пучки хорошо известна для древесины хвойных пород (Wickholm et al., 1998; Фахлен и Сальмен, 2005 г .; Fernandes et al., 2011). Фибриллярные единицы шириной от 5 до 10 нм, иногда до 40 нм, наблюдались с помощью электронной микроскопии (McCann et al., 1990; Sugimoto et al., 2000), но их интерпретация осложняется возможным включением нецеллюлозных полимеров. . Представленные здесь данные включают две прямые линии доказательств агрегации микрофибрилл первичной стенки в колленхиме. Доказательства малоуглового брэгговского рассеяния рентгеновских лучей (Kennedy et al., 2007a) и нейтронов ясно показали, что агрегация существует. Диапазон расстояний между микрофибриллами было трудно сделать вывод, потому что интенсивность рассеяния сильно увеличивалась с разделением микрофибрилл, так что хорошо разделенные микрофибриллы были чрезмерно представлены на диаграммах рассеяния. Низкий контраст МУРН в сухом состоянии и сходство межцентрового расстояния МУРН со средними размерами Шеррера WAXS предполагают, что некоторые сегменты микрофибрилл оставались в прямом контакте друг с другом, тогда как другие сегменты микрофибрилл становились хорошо разделенными при гидратации.
Спектры дейтерирования FTIR как от стенок клеток колленхимы, так и от изолированной целлюлозы включали полосы растяжения гидроксила с центром на 3 400 см -1 с нейтральной поляризацией. Эти гидроксильные полосы растяжения были отнесены к гидроксильным группам, отличным от всех групп Iα и Iβ кристаллических форм целлюлозы, более ориентированным наружу по ориентации и частично, но не полностью доступным для дейтерирования. Принимая во внимание также данные SANS и ЯМР, соответствующие гидроксильные группы могут быть отнесены к поверхностям микрофибрилл.Поскольку эти поверхности были только частично доступны для D 2 O, отсюда следует, что микрофибриллы были агрегированы вместе на части своей длины с границами раздела, непроницаемыми для воды. Эти особенности были обнаружены в древесине ели, но были также различия: древесина ели имеет разные нецеллюлозные полимеры, что, по-видимому, приводит к более тесному связыванию между поверхностями микрофибрилл и к напряжениям, которые искажают геометрию элементарной ячейки при гидратации (Fernandes et al., 2011).
Полоса валентных колебаний гидроксила 3,400 см -1 в FTIR-спектре, перемещенная до 2,540 см -1 за счет обмена D 2 O, ранее была неправильно отнесена к аберрантной форме целлюлозы Iα (Sturcová et al., 2004), но, по-видимому, является маркером поверхностных цепей и наличия агрегации микрофибрилл, если эти поверхностные цепи недоступны для дейтерирования. Интенсивность этой полосы сильно уменьшилась, когда целлюлоза из колленхимы сельдерея была подвергнута гидротермальному отжигу, что привело к слиянию микрофибрилл в более крупные кристаллические единицы с уменьшенным соотношением поверхности к объему (Sturcová et al., 2004). Интенсивность FTIR на уровне 3400 см или около него -1 была зарегистрирована в препаратах клеточных стенок, например, арабидопсиса дикого типа ( Arabidopsis thaliana ; MacKinnon et al., 2006), предполагая, что эта форма агрегации не является уникальной особенностью целлюлозы колленхимы сельдерея, но более широко распространена в первичных клеточных стенках.
Влияние на рост растений
Размер и структура микрофибрилл первичной стенки дают важные ключи к разгадке природы ферментных комплексов, ответственных за ориентированный синтез целлюлозы и, следовательно, за направленную экспансию клеток. Размножение растительных клеток требует разрушения сети нековалентно сшитых микрофибрилл в клеточной стенке.Понимание топологии этой сети, следовательно, необходимо, и недавно принятая идея о том, что ксилоглюканы просто покрывают и связывают поверхности микрофибрилл, была поставлена под сомнение (Bootten et al., 2004; Park and Cosgrove, 2012a). Данные спин-диффузии ЯМР (рис. 4) показали, что некоторые цепи гемицеллюлозы, вероятно, ксилоглюканы, располагались близко к поверхностям микрофибрилл и, таким образом, вероятно, лежали между микрофибриллами внутри агрегатов микрофибрилл. Однако другие сегменты микрофибрилл, по-видимому, находятся в прямом контакте без промежуточных полимеров, демонстрируя нерегулярный характер агрегации, которого можно было бы ожидать, если микрофибриллы были скрученными и нецилиндрическими, и предполагая, что взаимодействия целлюлоза-целлюлоза, а также ксилоглюкан-целлюлоза должны быть разбиты на отдельные агрегированные микрофибриллы.Др. Механизмы необходимы для отделения одного пучка микрофибрилл от следующего во время роста путем разрушения ксилоглюкана или др. Мостиков между пучками микрофибрилл (Anderson et al., 2010). Агрегация микрофибрилл вдоль части их длины, в то время как другие части их длины остаются свободными для разделения или в них вставлены другие полисахариды, поднимает интересные вопросы о действии экспансинов (Cosgrove, 2005) и о том, как действует действие определенных глюканов. гидролазы взаимодействуют с таковой ксилоглюкан-специфической эндоглюканазы (Park and Cosgrove, 2012b).
Интересен высокий уровень беспорядка на границах раздела, наиболее важных для роста, между микрофибриллами и нецеллюлозными полимерами, а также между одной микрофибриллой и другой. Примеры прочных биологических материалов из царства животных показывают, что неупорядоченные домены могут вносить вклад в ударную вязкость (высокую энергию разрушения) при внешнем стрессе (van Beek et al., 2002). В первичных клеточных стенках устойчивость к разрушению должна сочетаться с контролируемой ферментативно-опосредованной продуктивностью, по-видимому, на неупорядоченных границах раздела, что обеспечивает рост.
Последствия разложения целлюлозы
Доступность микрофибрилл целлюлозы для целлюлаз является важным фактором превращения лигноцеллюлозы в биотопливо и еще более важна в случае нелгнифицированной биомассы. Нестабильность целлюлозы часто связывают с ее кристалличностью (Arantes and Saddler, 2010), а «декристаллизацию» иногда называют целью процессов предварительной обработки (Beckham et al., 2011), без полного согласия относительно того, что означают эти термины.Степень, в которой микрофибриллы целлюлозы имеют относительно гидрофобную [200] открытую поверхность, будет положительно влиять на начальную стадию деградации целлюлазами, которые специфически связываются с этой поверхностью (Dagel et al., 2011; Liu et al., 2011). Было бы интересно узнать больше о специфичности связывания оксидаз целлюлозы GH61 / CBM33 (Forsberg et al., 2011; Quinlan et al., 2011), некоторые из которых также связываются с одной и той же поверхностью (Li et al., 2012). Агрегация микрофибрилл целлюлозы — это фактор сопротивляемости, который заслуживает большего внимания, и теперь к нему можно подойти с помощью экспериментов по дейтерированию FTIR.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Было выделенонитей колленхимы из зрелых черешков сельдерея ( Apium graveolens ), клеточные стенки и их целлюлозная фракция были приготовлены, как описано (Sturcová et al., 2004), и хранились в сухом виде. Подробнее см. Дополнительные материалы и методы S1.
SANS
ЭкспериментыSANS были выполнены на канале D11 в Институте Лауэ-Ланжевена на клеточных стенках колленхимы сельдерея и изолированной целлюлозе.Подробнее об эксперименте см. Дополнительные материалы и методы S1.
WAXS
дифрактограммы рентгеновских лучей получали при температуре окружающей среды с использованием дифрактометра Rigaku R-axis / RAPID для рентгеновских пластин с источником излучения с молибденовой рентгеновской эмиссией Kα (λ = 0,07071 нм). Пучок коллимировался до диаметра 0,5 мм. Образцы имели толщину 0,7 мм в направлении, параллельном пучку, а их другие размеры превышали диаметр пучка. Дифрактограммы были собраны в режиме перпендикулярного пропускания, за исключением экспериментов с наклоном для измерения осевых отражений.Расстояние от центра дифракционной картины было выражено как q = 4πsinθ / λ, где 2θ — угол рассеяния. Для получения дополнительных сведений об экспериментальных методах и методах обработки данных (Fernandes et al., 2011) см. Дополнительные материалы и методы S1.
WANS
Дифракция нейтронов проводилась на канале D19 в Институте Лауэ-Ланжевена при средней длине волны нейтронов 0,242 нм и расстоянии от образца до детектора 0,756 м. Образцы размером 25 × 15 × 1 мм собирали из нескольких выровненных нитей колленхимы и выдерживали при контролируемом давлении воды или паров D 2 O в проточном контейнере со стенками из алюминиевой фольги.Подробнее см. Дополнительные материалы и методы S1.
ЯМР
A 1 H Эксперимент по спиновой диффузии Гольдмана-Шена с детектированием по спектру 13 C после кросс-поляризации был проведен на клеточных стенках колленхимы сельдерея, гидратированных 0,2 см. 3 г −1 D 2 O. Экспериментальные процедуры и анализ данных, включая 2D-представление данных, были такими, как описано (Fernandes et al., 2011).
Дейтерирование FTIR
Стенки клеток колленхимы сельдерея и изолированную целлюлозу помещали в проточную ячейку с окнами из фторида бария и заменяли воздухом, насыщенным О 2 О, до тех пор, пока спектры не перестали изменяться. Затем газовую линию переключали на азот, тщательно осушая, пропуская через сухое молекулярное сито 4А, а затем пентоксид фосфора (Sicapent; Aldrich). Спектры FTIR собирали с помощью спектрометра Thermo Nicolet Nexus, оборудованного приставкой для микроскопа Nicolet Continuum, имеющей охлаждаемый жидким азотом детектор теллурида кадмия ртути (Fernandes et al., 2011).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Rigaku, Ltd. за предоставленный инструмент и Fujiang Zhang за получение некоторых спектров FTIR.
Глоссарий
SAXS
Малоугловое рассеяние рентгеновских лучей
D 2 O
SANS
Малоугловое рассеяние нейтронов
WAXS 9407
широкоугольное рассеивание
WANS
широкоугольное рассеяние нейтронов
FTIR
Инфракрасное преобразование Фурье
2D
ppm
LITERATURE CITED
(
2006
)Пространственные отношения между полимерами в ели ситкинской: исследования спиновой диффузии протонов
.Holzforschung
60
:665
—673
(
2010
)Визуализация в реальном времени переориентации целлюлозы во время расширения клеточной стенки в корнях Arabidopsis
.Plant Physiol
152
:787
—796
(
2009
)Выяснение механизмов сборки и субъединичного взаимодействия целлюлозосинтазного комплекса вторичных клеточных стенок Arabidopsis
.J Biol Chem
284
:3833
—3841
(
1997
)Атомно-силовая микроскопия высокого разрешения микрокристаллов нативной целлюлозы I Valonia
.J Struct Biol
119
:129
—138
(
2011
)Происхождение стойкости биомассы на молекулярном уровне: свободные энергии декристаллизации для четырех обычных полиморфов целлюлозы
.J Phys Chem B
115
:4118
—4127
(
2008
)Динамика границ раздела целлюлоза-вода: времена спин-решеточной релаксации ЯМР, рассчитанные на основе атомистического компьютерного моделирования
.J Phys Chem B
112
:2590
—2595
(
2004
)Твердотельный 13 C-ЯМР-спектроскопия показывает, что ксилоглюканы в первичных клеточных стенках маша (Vigna radiata L.) происходят в разных доменах: новая модель взаимодействий ксилоглюкан-целлюлоза в клеточной стенке
.J Exp Bot
55
:571
—583
(
2009
)Растения контролируют свойства и работу своих органов посредством ориентации фибрилл целлюлозы в их клеточных стенках
.Интегр Комп Биол
49
:69
—79
(
2005
)Рост клеточной стенки растений
.Nat Rev Mol Cell Biol
6
:850
—861
(
2011
)Визуализация на месте отдельных углеводсвязывающих модулей на микрофибриллах целлюлозы
.J Phys Chem B
115
:635
—641
(
1996
)Паренхиматозная клеточная целлюлоза из жома сахарной свеклы: получение и свойства
.Целлюлоза
3
:183
—188
(
2005
)Распределение пор и матрицы в стенке волокна, выявленное с помощью атомно-силовой микроскопии и анализа изображений
.Биомакромолекулы
6
:433
—438
(
1999
)Пакет WinGX для кристаллографии низкомолекулярных монокристаллов
.J Appl Cryst
32
:837
—838
(
2011
)Наноструктура микрофибрилл целлюлозы в древесине ели
.Proc Natl Acad Sci USA
108
:E1195
—E1203
(
2011
)Расщепление целлюлозы белком CBM33
.Protein Sci
20
:1479
—1483
(
2012
)Твердотельное селективное (13) C возбуждение и спин-диффузионный ЯМР для определения пространственных размеров в стенках растительных клеток
.J Agric Food Chem
60
:1419
—1427
(
2000
)Характеристика поперечных связей между микрофибриллами целлюлозы и их появление во время элонгационного роста в эпикотиле гороха
.Физиология растительных клеток
41
:486
—494
(
2011
)Кортикальные микротрубочки оптимизируют кристалличность клеточной стенки для обеспечения однонаправленного роста Arabidopsis
.Завод J
66
:915
—928
(
1999
)Выражение паттерна в растениях: сочетание молекулярных и основанных на исчислении биофизических парадигм
.Am J Bot
86
:1059
—1076
и другие. (
2012
)Кристалличность микрофибрилл целлюлозы снижается за счет мутации С-концевых остатков трансмембранной области CESA1A903V и CESA3T942I синтазы целлюлозы
.Proc Natl Acad Sci USA
109
:4098
—4103
(
2007
)Устойчивость биомассы: инженерные установки и ферменты для производства биотоплива
.Наука
315
:804
—807
(
2009
)Различия между сортами в размерах микрофибрилл целлюлозы, наблюдаемые с помощью инфракрасной спектроскопии
.Целлюлоза
16
:1
—8
(
1995
)Радиальная упаковка, порядок и беспорядок в фибриллах коллагена
.Biophys J
68
:1661
—1670
(
1992
)Контроль толщины клеточных стенок колленхимы пектинами
.Планта
187
:218
—220
(
2011
)Стенки растительных клеток: супрамолекулярные ансамбли
.Food Hydrocoll
25
:257
—262
(
2007a
)Диаметр микрофибрилл в целлюлозе колленхимы сельдерея: рассеяние рентгеновских лучей и данные ЯМР
.Целлюлоза
14
:235
—246
(
2007b
)Влияние гидратации на расстояние между микрофибриллами целлюлозы первичной стенки: исследование малоуглового рентгеновского рассеяния
.Целлюлоза
14
:401
—408
(
2008
)Выявление структурного и функционального разнообразия клеточных стенок растений
.Curr Opin Plant Biol
11
:308
—313
(
2001
)Рентгеновская структура мерсеризованной целлюлозы II при разрешении 1 Å
.Биомакромолекулы
2
:410
—416
(
2012
)Структурная основа нацеливания на субстрат и катализа грибковыми полисахаридмонооксигеназами
.Структура
20
:1051
—1061
(
2011
)Целлобиогидролаза гидролизует кристаллическую целлюлозу на гидрофобных поверхностях
.J Biol Chem
286
:11195
—11201
(
2006
)Структура и анизотропия клеточной стенки в procuste , мутанте целлюлозосинтазы Arabidopsis thaliana
.Планта
224
:438
—448
(
2005
)Расширение клеточной стенки приводит к координатному разделению параллельных микрофибрилл: данные сканирующей электронной микроскопии и атомно-силовой микроскопии
.Завод J
43
:181
—190
(
2003
)Твердотельный 13 C и 1 H спин-диффузионный ЯМР-анализ структуры микрофибрилл бактериальной целлюлозы
.Твердотельный ядерный магнитный резонон
23
:198
—212
(
2012
)Сравнение моделирования бета целлюлозы I с тремя силовыми полями углеводов
.J Chem Theory Comput
8
:735
—748
(
2011
)Высокотемпературные свойства целлюлозы I
.J Phys Chem B
115
:2155
—2166
(
2006
)Исследования микрокристаллической целлюлозы Ibeta
с помощью компьютерного моделирования.Углеводородный остаток
341
:138
—152
(
1990
)Прямая визуализация поперечных связей в первичной клеточной стенке растения
.J Cell Sci
96
:323
—334
(
2008
)Структура и биосинтез пектина
.Curr Opin Plant Biol
11
:266
—277
(
1992
)Ядерно-магнитно-резонансное исследование пространственных соотношений между химическими компонентами в стенках деревянных клеток
.Holzforschung
46
:205
—210
(
2009
)Структура и свойства микрофибриллы целлюлозы
.J Wood Sci
55
:241
—249
(
2002
)Кристаллическая структура и система водородных связей в целлюлозе Ibeta по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
124
:9074
—9082
(
2003
)Кристаллическая структура и система водородных связей в целлюлозе I (альфа) по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
125
:14300
—14306
(
2008
)Генетические доказательства того, что активность синтазы целлюлозы влияет на организацию кортикального массива микротрубочек
.Plant Physiol
147
:1723
—1734
(
2012a
)Изменения биомеханических свойств клеточной стенки у ксилоглюкан-дефицитного мутанта xxt1 / xxt2 Arabidopsis
.Plant Physiol
158
:465
—475
(
2012b
)Пересмотренная архитектура первичных клеточных стенок, основанная на биомеханических изменениях, индуцированных субстрат-специфическими эндоглюканазами
.Plant Physiol
158
:1933
—1943
и другие. (
2011
)Анализ окислительного разложения целлюлозы металлоферментом меди, который использует компоненты биомассы
.Proc Natl Acad Sci USA
108
:15079
—15084
и другие.(
2012
)CHITINASE-LIKE1 / POM-POM1 и его гомолог CTL2 представляют собой взаимодействующие с глюканом белки, важные для биосинтеза целлюлозы у Arabidopsis
.Plant Cell
24
:589
—607
(
2008
)Краткая история SHELX
.Acta Crystallogr A
64
:112
—122
(
2006
)Синтез целлюлозы у высших растений
.Annu Rev Cell Dev Biol
22
:53
—78
(
2004
)Структурные детали кристаллической целлюлозы из высших растений
.Биомакромолекулы
5
:1333
—1339
(
2003
)Поляризованная колебательная спектроскопия волоконных полимеров: водородные связи в целлюлозе II
.Биомакромолекулы
4
:1589
—1595
(
2000
)Новые методы позволяют проводить сравнительный анализ ориентации микротрубочек, текстуры стенок и скорости роста интактных корней Arabidopsis
.Plant Physiol
124
:1493
—1506
(
2009
)Динамическая координация систем цитоскелета и клеточной стенки во время морфогенеза растительных клеток
.Curr Biol
19
:R800
—R811
(
2002
)Молекулярная структура шелка драглайна пауков: складывание и ориентация белкового остова
.Proc Natl Acad Sci USA
99
:10266
—10271
(
2002
)Конформационные особенности кристаллической целлюлозы высших растений
.Завод J
30
:721
—731
(
2004
)Полиморфизм целлюлозы семейства I: повторное исследование целлюлозы IVI
.Биомакромолекулы
5
:1385
—1391
(
1998
)Определение некристаллических форм в целлюлозе I методом спектроскопии ЯМР CP / MAS C-13
.Углеводородный остаток
312
:123
—129
(
2007
)Организация боковых цепей пектинового арабинана и галактана в ассоциации с микрофибриллами целлюлозы в первичных клеточных стенках и связанных моделях предусматривается
.J Exp Bot
58
:1795
—1802
Заметки автора
© Американское общество биологов растений, 2013 г. Все права защищены.
© Автор (ы) 2013. Опубликовано Oxford University Press от имени Американского общества биологов растений. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0 /), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
Структура микрофибрилл целлюлозы в первичных клеточных стенках колленхимы | Физиология растений
Аннотация
В первичных стенках растущих клеток растений глюкозно-полимерная целлюлоза собрана в длинные микрофибриллы диаметром несколько нанометров. Жесткость и ориентация этих микрофибрилл контролируют рост клеток; Следовательно, синтез целлюлозы является ключевым фактором роста и морфогенеза растений.Колленхима сельдерея ( Apium graveolens ) является полезной модельной системой для изучения структуры микрофибрилл первичной стенки, поскольку ее микрофибриллы ориентированы с необычной однородностью, что облегчает проведение спектроскопических и дифракционных экспериментов. Используя комбинацию методов рассеяния рентгеновских лучей и нейтронов с колебательной спектроскопией и спектроскопией ядерного магнитного резонанса, мы показываем, что микрофибриллы колленхимы сельдерея имели средний диаметр от 2,9 до 3,0 нм, с наиболее вероятной структурой, содержащей 24 цепи в поперечном сечении, расположенные в восьми атомах водорода. -связанные листы из трех цепей с обширным беспорядком в боковой упаковке, конформации и водородной связи.Аналогичная структура из 18 цепей и структуры из 24 цепей разной формы хуже соответствовали данным. Конформационный беспорядок в основном ограничивался поверхностными цепями, а беспорядок в упаковке цепей — нет. То есть по положению и ориентации поверхностные цепи соответствовали неупорядоченной решетке, составляющей ядро каждой микрофибриллы. Были доказательства того, что соседние микрофибриллы нековалентно агрегированы вместе на части своей длины, предполагая, что необходимость разрушения этих агрегатов может быть сдерживающим фактором роста и гидролиза целлюлозы для производства биотоплива.
Рост и форма растений контролируются точно ориентированным расширением стенок отдельных клеток. Движущая сила для размножения клеток является осмотической, но скорость и направление экспансии контролируются механическими свойствами клеточной стенки (Szymanski and Cosgrove, 2009). Расширяющиеся первичные клеточные стенки представляют собой нанокомпозитные материалы, в которых длинные микрофибриллы целлюлозы диаметром несколько нанометров проходят через гидратированную матрицу ксилоглюканов, пектинов и других полимеров (Knox, 2008; Mohnen, 2008; Szymanski and Cosgrove, 2009; Scheller. и Ульвсков, 2010).Нативные микрофибриллы целлюлозы частично кристаллические (Nishiyama, 2009; Fernandes et al., 2011). Раньше считалось, что целлюлоза первичной стенки имеет уникальную кристаллическую структуру, называемую целлюлозой IV 1 (Dinand et al., 1996), но данные ЯМР предполагают наличие форм, подобных лучше охарактеризованным кристаллическим формам целлюлозы Iα и Iβ вместе с крупными кристаллическими формами целлюлозы. количества менее упорядоченной целлюлозы (Wickholm et al., 1998; Sturcová et al., 2004; Wada et al., 2004). Тем не менее, целлюлоза гораздо более упорядочена, чем любой другой компонент первичной клеточной стенки (Bootten et al., 2004), в соответствии с его ключевой ролью обеспечения силы и контроля роста.
Жесткость клеточной стенки наибольшая в направлении микрофибрилл целлюлозы, где рост является направленным, а преобладающая ориентация микрофибрилл обычно поперечна направлению роста (Green, 1999; MacKinnon et al., 2006; Szymanski and Cosgrove, 2009 г.). Расширение клеточной стенки требует либо увеличения расстояния между микрофибриллами (Marga et al., 2005), либо проскальзывания между ними (Cosgrove, 2005), либо того и другого, и микрофибриллы переориентируются в направлении роста (Anderson et al., 2010). Полимерные поперечные мостики между микрофибриллами (McCann et al., 1990), как полагают, противостоят этим деформациям наноструктуры клеточной стенки и, таким образом, контролируют скорость роста. До недавнего времени наибольшее внимание уделялось мостиковым соединениям ксилоглюканов, водородным связям с поверхностями микрофибрилл (Scheller, Ulvskov, 2010). Однако есть свидетельства того, что не все ксилоглюканы расположены надлежащим образом (Fujino et al., 2000; Park and Cosgrove, 2012a) и что могут быть задействованы другие мостиковые полимеры (Zykwinska et al., 2007). Также было высказано предположение, что пучки агрегированных микрофибрилл, а не отдельные микрофибриллы, могут быть ключевыми структурными единицами в первичных клеточных стенках (Anderson et al., 2010), как в древесине (Fahlén and Salmén, 2005; Fernandes et al., 2011). ). В таком случае отдельные микрофибриллы могут соединяться между пучками микрофибрилл. Таким образом, рост растительных клеток не совсем понятен, и нам нужна дополнительная информация о том, как контролируется ориентация целлюлозы, и о природе мостиковых полимеров, о поверхностях целлюлозы, с которыми эти полимеры связываются, и о сцеплении между поверхностями микрофибрилл, которые могут опосредовать агрегацию.
Микрофибриллы целлюлозы синтезируются на поверхности клетки большими ферментными комплексами, имеющими гексагональную симметрию, иногда называемыми «розетками» (Somerville, 2006). Каждый комплекс содержит несколько синтаз целлюлозы, которые различаются между первичными клеточными стенками и древесиной, хотя внешний вид комплексов похож (Somerville, 2006; Atanassov et al., 2009). Одновременный синтез с одного и того же конца всех цепей в микрофибрилле нативной целлюлозы является причиной их параллельности (Nishiyama et al., 2002, 2003), в отличие от энтропийно предпочтительной антипараллельной структуры, обнаруженной в искусственных целлюлозах, таких как вискоза (Langan et al., 2001). Очевидно, что количество цепей в микрофибрилле и количество целлюлозосинтаз в синтетическом комплексе взаимосвязаны. Принято считать, что количество цепочек делится на шесть, что соответствует симметрии гексагональной розетки и 36-цепочечным моделям (Himmel et al., 2007), ограниченным гидрофильными гранями кристаллов [110] и [1-10], поскольку в водорослевых целлюлозах (Bergenstråhle et al., 2008), получили широкое распространение. Сборка и ориентация целлюлозы взаимосвязаны, поскольку некоторые мутанты целлюлозосинтазы имеют фенотипы, дефектные по ориентации целлюлозы и растительной форме, а также обедненные содержанием целлюлозы (Paredez et al., 2008). У некоторых других мутантных линий, по-видимому, затрагивается кристалличность микрофибрилл (Fujita et al., 2011; Harris et al., 2012; Sánchez-Rodríguez et al., 2012).
Следовательно, детальное понимание структуры микрофибрилл целлюлозы первичной стенки поможет нам понять синтез целлюлозы, а также рост и структурную механику живых растений (Burgert and Fratzl, 2009).Первичные клеточные стенки и их целлюлозные скелеты также влияют на такие характеристики качества пищевых продуктов, как хрусткость салата из овощей и яблок ( Malus domestica ; Jarvis, 2011). Когда биотопливо производится из лигноцеллюлозной биомассы, лигнификация приводит к устойчивости (Himmel et al., 2007), но некоторые типы клеток у видов Miscanthus , просо ( Panicum virgatum ) и пожнивные остатки имеют только первичные стенки. без лигнина, и сопротивляемость этому зависит от природы микрофибрилл целлюлозы (Beckham et al., 2011).
Относительно подробная структура недавно была предложена для микрофибрилл древесины ели ( Picea spp.) (Fernandes et al., 2011), которые имеют диаметр 3,0 нм, что позволяет разместить только около 24 цепочек целлюлозы. Данные рентгеновской дифракции подтверждают «прямоугольную» форму (Matthews et al., 2006), ограниченную гранями [010] и [200]. Наблюдался значительный беспорядок, увеличивающийся к поверхности, и микрофибриллы были агрегированы в пучки размером примерно 15-20 нм в поперечнике, причем некоторые, но не все, боковые границы раздела были устойчивы к воде (Fernandes et al., 2011). Неупорядоченные домены являются особенностью других прочных биологических материалов, таких как шелк паука (van Beek et al., 2002).
Следовательно, представляет интерес, могут ли какие-либо из этих свойств древесной целлюлозы также быть обнаружены в микрофибриллах целлюлозы первичных (растущих) клеточных стенок. Было бы особенно полезно охарактеризовать нарушение, которое, как известно, присутствует в микрофибриллах первичной стенки, то есть определить, чем целлюлоза, которая не определяется как «кристаллическая», отличается от кристаллической целлюлозы.Многие эксперименты, ведущие к созданию структуры древесной целлюлозы, зависели от исключительно однородной ориентации микрофибрилл целлюлозы (Sturcová et al., 2004; Fernandes et al., 2011). Однако в растущих клеточных стенках микрофибриллы ориентированы неравномерно. Когда микрофибриллы сначала откладываются на внутренней поверхности первичной клеточной стенки, их ориентация обычно перпендикулярна направлению роста, но когда клеточная стенка расширяется, микрофибриллы переориентируются таким образом, что распределение ориентации интегрируется по толщине расширенной клеточной стенки. клеточная стенка становится все более близкой к случайной (Cosgrove, 2005; MacKinnon et al., 2006).
Эта техническая проблема не относится к клеточным стенкам колленхимы сельдерея ( Apium graveolens ), которые похожи по составу на другие первичные клеточные стенки, но имеют их микрофибриллы, ориентированные относительно равномерно вдоль оси клетки (Sturcová et al., 2004 ; Kennedy et al., 2007a, 2007b). Некоторая структурная информация о целлюлозе колленхимы сельдерея уже была получена в результате спектроскопических экспериментов и экспериментов по рассеянию (Sturcová et al., 2004; Kennedy et al., 2007a, 2007b), подтверждая беспорядок, ожидаемый в целлюлозе первичной стенки.Некоторые из этих экспериментов были аналогичны тому, что было сделано с еловой целлюлозой (Fernandes et al., 2011), но недостаточно данных, чтобы указать количество цепей в каждой микрофибрилле первичной стенки, природу и расположение нарушения, а также наличие или отсутствие прямого контакта микрофибрилл. Здесь мы сообщаем об экспериментах по рассеянию рентгеновских лучей и нейтронов, а также спектроскопических экспериментах, направленных на решение этих вопросов и приводящих к предложенной структуре микрофибрилл целлюлозы первичной стенки. Характеристика структуры, содержащей столько беспорядка, представляла необычные проблемы, но беспорядок, по-видимому, является центральным для загадочной способности целлюлозы первичной стенки обеспечивать высокую прочность, но при этом позволять и контролировать рост.
РЕЗУЛЬТАТЫ
Диаметр микрофибриллы и расстояние от малоуглового рассеяния нейтронов
Когда микрофибриллы коллагена или целлюлозы фиксированного диаметра упакованы в массивы с некоторой степенью регулярности, может наблюдаться малоугловая дифракция рентгеновских лучей или нейтронов (брэгговское рассеяние) от самих микрофибрилл (Hulmes et al., 1995; Fernandes et al. др., 2011). Это рассеяние происходит под меньшими углами, чем обычная дифракция на кристаллических плоскостях, и накладывается на малоугловое рассеяние, возникающее не из-за дифракции, а из-за текстуры и формы фибрилл и пустот между ними (форм-фактор).Если упаковка не является достаточно регулярной, чтобы образовать кристаллическую сверхрешетку, наблюдается только один пик Брэгга, соответствующий расстоянию между центрами фибрилл (Hulmes et al., 1995; Kennedy et al., 2007a).
Используя малоугловое рассеяние рентгеновских лучей (SAXS), наблюдали брэгговское отражение от микрофибрилл целлюлозы внутри интактных клеточных стенок колленхимы сельдерея. Расстояние между микрофибриллами увеличивается при гидратации (Kennedy et al., 2007b). Моделирование этих результатов показало, что наблюдаемый пик Брэгга может быть вызван рядом комбинаций расстояния между микрофибриллами и их диаметра.Следовательно, невозможно однозначно определить диаметр микрофибрилл (Kennedy et al., 2007a).
В древесине ели микрофибриллы слишком тесно агрегированы, чтобы обеспечить адекватный контраст для малоуглового брэгговского рассеяния. Эту проблему удалось решить путем дейтерирования поверхности и использования нейтронов вместо рентгеновских лучей (Fernandes et al., 2011). Природа контраста различается между источниками рентгеновского излучения и нейтронов. Рентгеновский контраст зависит в основном от локальной электронной плотности, тогда как контраст рассеяния нейтронов зависит от локального элементного состава и плотности.Одним из преимуществ этого метода является очень разные сечения рассеяния нейтронов водородом и дейтерием, что позволяет ввести контраст, например, за счет дейтериевого обмена в оксиде дейтерия (D 2 O; Horikawa et al., 2009; Matthews и др., 2011).
Экваториальный пик Брэгга был хорошо виден на изображениях малоуглового рассеяния нейтронов (МУРН) древесины ели, насыщенной D 2 O (Fernandes et al., 2011). Интенсивность пика Брэгга уменьшалась, а его расстояние ( q ) от центра дифракционной картины увеличивалось при сушке до тех пор, пока на картине SANS от полностью сухой дейтерированной древесины пик не был слабо виден при q = 2.1 нм −1 . Низкий контраст в этих условиях соответствовал контакту между соседними дейтерированными на поверхности микрофибриллами, так что расстояние между центрами 3,0 нм соответствовало по существу среднему диаметру микрофибрилл (Fernandes et al., 2011).
Подобные эксперименты SANS были проведены на стенках клеток из колленхимы сельдерея и на целлюлозе, выделенной из этих клеточных стенок (рис. 1). Пик брэгговского рассеяния в области q = 1 нм -1 , наложенный на экспоненциальный фон от некогерентного рассеяния, был очевиден как для стенок клеток, так и для изолированной целлюлозы, насыщенной чистым D 2 O, чистой водой или ряд смесей (рис.1; Дополнительный рис. S1). В соответствии с рассеянием от целлюлозы, а не от других полимеров, не наблюдалось пика Брэгга для смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Которая соответствует теоретической плотности длины рассеяния. целлюлозы (рис. 1).
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна.C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл. D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив.Вертикальный масштаб больше, чем в C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна. C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл.D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив. Вертикальная шкала больше, чем C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
В отличие от древесины ели, сухая целлюлоза сельдерея дает слабый пик Брэгга без дейтерирования поверхности микрофибрилл. В сухом состоянии пик Брэгга был при q = 2.11 нм -1 для целлюлозы в клеточных стенках или q = 2,14 нм -1 для изолированной целлюлозы (рис. 1), что соответствует межцентровым расстояниям в реальном пространстве 3,0 и 2,9 нм, соответственно. . Интенсивность Брэгга была менее 1% от наблюдаемой после регидратации. Этот очень низкий контраст соответствовал плотной упаковке микрофибрилл, вероятно, без каких-либо других полимеров между ними по крайней мере на некоторой части их длины. Расстояние от центра до центра должно приблизительно соответствовать среднему диаметру микрофибриллы.Несмотря на свою низкую интенсивность, пик Брэгга был легко различим на картинах МУРН, поскольку было очень мало помех от некогерентного рассеяния (рис. 1).
В гидратированном состоянии значение q пика Брэгга от микрофибрилл in situ в стенках клеток будет соответствовать реальному расстоянию ( d ) 7,1 ± 1,3 нм -1 , в то время как q Значение пика Брэгга для гидратированной целлюлозы соответствует реальному интервалу 6.0 ± 1,5 нм -1 . Эти номинальные расстояния d не означают, что все микрофибриллы разделены до такой степени при гидратации. Гидратация также значительно увеличивала контраст SANS за счет введения воды или D 2 O между микрофибриллами и, в случае D 2 O, за счет дейтерирования их поверхностей и нецеллюлозных полимеров. Отсюда следует, что при любом распределении расстояний между микрофибриллами более широкие промежутки будут значительно перепредставлены в шаблоне SANS. Номинальные интервалы, наблюдаемые после гидратации, немного превышали расчетные (Kennedy et al., 2007b) методом SAXS, что согласуется с разной основой рентгеновского контраста, но не было никакой разницы между расстояниями, наблюдаемыми в сухом состоянии с рентгеновскими лучами (Kennedy et al., 2007a) и с нейтронами (рис. 1). . Хотя на основе рентгеновских данных невозможно было сделать вывод о том, что расстояние, наблюдаемое в сухом состоянии, соответствует микрофибриллам, находящимся в прямом контакте друг с другом, этот вывод кажется вероятным на основе объединенных данных SAXS и SANS. . Гидратация четко разделяла микрофибриллы, но, вероятно, в разной степени по их длине (Kennedy et al., 2007b), возможно, в зависимости от сродства нецеллюлозных полисахаридов к воде (Jarvis, 1992).
Размеры микрофибрилл и нарушения в результате широкоугольного рассеяния рентгеновских лучей
Широкоугольные картины рассеяния рентгеновских лучей или нейтронов от природной целлюлозы достаточно хорошо разрешены для кристаллографического решения структуры, только если целлюлоза исключительно упорядочена и хорошо ориентирована и если микрофибриллы достаточно толстые, чтобы минимизировать уширение Шеррера, которое возникает в результате ограниченное количество плоскостей решетки.Большая часть целлюлозы колленхимы сельдерея была относительно хорошо ориентирована внутри клеточных стенок (рис. 2). Вклад широкоугольного рентгеновского рассеяния (WAXS) от незначительной изотропно ориентированной фракции целлюлозы был удален на стадии вычитания фона. Существенный вклад изотропного рассеяния на связанной воде также был удален при вычитании фона. Рассеяние от воды было идентифицировано экспериментом, в котором менялось содержание воды (дополнительный рис. S2).
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок WAXS из хорошо ориентированного целлюлозного компонента был размыт в радиальном направлении из-за уширения Шеррера, уширения, связанного с беспорядком, или обоих. Отражение 200 было при q = 15,5 нм -1 , что соответствует несколько большему расстоянию между слоями, чем в кристаллических целлюлозах, и отражения 1-10 и 110 были объединены. Эти особенности наблюдались на плохо ориентированных дифрактограммах от того, что было названо целлюлозой IV 1 (Dinand et al., 1996) или неупорядоченную целлюлозу Iβ (Wada et al., 2004) в первичных клеточных стенках. Для единообразия в этой статье используется обозначение решетки Iβ, но без всякого указания на то, что эта форма представляет собой наиболее близкую аналогию со средней структурой.
Количественная оценка беспорядка и вычисление размеров Шеррера по уширению этих отражений было затруднено из-за сильного перекрытия между отражениями 1-10 и 110 и чрезвычайно низкой интенсивности отражения 400, качественного индикатора большего бокового беспорядка, чем в хвойная древесина.При длительном времени сбора данных (16–24 ч) можно было получить достаточное отношение сигнал / шум для оценки уширения отражения 400. Гидратация за счет увеличения моноклинного угла и, таким образом, разделения отражений 1-10 и 110 упростила моделирование соответствующей ширины этих двух отражений в дифракционной картине от древесины ели (Fernandes et al., 2011). Однако в целлюлозе сельдерея гидратация немного увеличивала интервал [200] d , но не увеличивала значительно угол моноклинности.Процедура Fernandes et al. (2011) был использован для выделения эффекта одной формы бокового беспорядка, которая приводит к асимметрии радиальных пиков. Предполагая, что оставшиеся симметричные радиальные ширины отражений 1-10 и 110 были равны, наиболее подходящее значение было на 22% больше, чем радиальная ширина отражения 200 при моноклинном угле 95 °. Исходя из этого, средние размеры элементарной ячейки в сухом состоянии составляли a = 0,81 нм, b = 0,80 нм, c = 1,03 нм (с сохранением индекса целлюлозы Iβ для ясности, даже если а> b).Меньшие углы моноклинности и большая ширина, или наоборот, почти так же точно соответствовали наблюдаемым профилям интенсивности.
Еще одна форма беспорядка, которая, наряду с небольшими размерами решетки, способствует радиальному уширению, была смоделирована как «паракристаллическая» (Fernandes et al., 2011), что означает, что ширина отражений увеличивается с квадратом порядка отражения. или с q 2 d . Используя наиболее подходящие данные и построив график ширины δ q отражений против q 2 d , как описано (Fernandes et al., 2011), ширина симметричных компонент рефлексов 200 и 400 давала фактор остаточной разупорядоченности г = 0,03 и размер Шеррера 3,2 нм, перпендикулярный плоскостям колец в сухом состоянии. Средняя ширина отражений 1-10 и 110 лежит явно выше δ q по сравнению с графиком q 2 d для отражений 200 и 400. Подразумевается, что средний размер Шеррера в направлениях, нормальных к плоскостям решетки 1-10 и 110 был меньше, чем средний размер Шеррера в направлении, нормальном к плоскости решетки 200, как в еловой целлюлозе.Однако низкая интенсивность отражения 400 и, как следствие, сложность оценки вклада, связанного с беспорядком, в общее уширение сделали этот вывод менее надежным, чем для еловой целлюлозы.
Характеристика неупорядоченных компонентов с помощью дейтерирования и широкоугольного рассеяния нейтронов
В кристаллографическом исследовании целлюлозы Iα и Iβ (Nishiyama et al., 2002, 2003) разница в дифракции нейтронов, обусловленная полным высокотемпературным дейтерированием гидроксильных групп, использовалась для определения положений протонов и, следовательно, геометрии водородных связей. .Также должна быть возможность использовать сильное различие между рассеянием нейтронов дейтерием и протонами для определения вклада в рассеяние неупорядоченных областей, доступных для дейтерирования в более мягких условиях температуры окружающей среды, используемых в экспериментах с SANS и инфракрасным преобразованием Фурье (FTIR). Как может быть показано с помощью FTIR (см. Ниже), гидроксильные группы на нецеллюлозных полисахаридах доступны для дейтерирования, как и обращенные наружу гидроксилы на поверхности целлюлозы, если они не заблокированы агрегацией (Horikawa et al., 2009). Для неупорядоченных областей внутри микрофибрилл, чтобы допустить D 2 O, они должны иметь более широкие интервалы решетки, чем кристаллическая целлюлоза (Matthews et al., 2011).
На рис. 3А показано, что наблюдались видимые изменения в картине дифракции нейтронов при обратимом дейтерировании при температуре окружающей среды. Изменения включали увеличение или уменьшение интенсивности отдельных отражений, а также изменения диффузного рассеяния. В принципе, изменение интенсивности для любого отражения может быть положительным, отрицательным или нейтральным в зависимости от влияния дейтерирования на структурный фактор.В качестве руководства, интенсивности основных экваториальных и осевых отражений для целлюлозы Iβ в нативной (H) и пердейтерированной (D) формах были рассчитаны с использованием Shelxl-97 (Sheldrick, 2008) в пакете WinGX (Farrugia, 1999), из статическая модель, использующая опубликованные координаты атомов (Nishiyama et al., 2002). Из трех основных экваториальных отражений, определяющих геометрию боковой упаковки целлюлозных цепей, влияние дейтерирования на интенсивность было предсказано положительным для 200 и сильно отрицательным для 110 и 1-10.Было предсказано, что осевое отражение 004 будет увеличиваться по интенсивности при дейтерировании, в то время как отражение 001 отсутствовало как в D, так и в H-формах. Эти прогнозы согласуются с наблюдаемыми нейтронографическими изображениями для высококристаллической целлюлозы Iβ (Nishiyama et al., 2002).
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина). Ось волокна вертикальна.B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина).Ось волокна вертикальна. B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Наблюдаемый эффект поверхностного дейтерирования микрофибрилл был положительным для отражений 200 и 004, отрицательным для 110 и нейтральным для 1-10.Профиль вычитания (D — H) для экваториальных отражений (рис. 3B) показал, что их положение и ширина, соответствующие цепям, доступным для дейтерирования, не заметно отличались от фракции, которая не подвергалась дейтерированию. В азимутальном направлении экваториальная интенсивность показывала более широкий нижний хвост распределения ориентации в D, чем в H-дифракционной картине (рис. 3E), подразумевая, что некоторые из цепей, доступных для дейтерирования, были менее хорошо ориентированы, чем недоступные цепочки целлюлозы.
На оси волокна (рис. 3D) отражение 004 в D-форме было центрировано при немного меньшем q , чем в H-форме, что означает, что некоторые из цепочек, доступных для дейтерирования, были более протяженными, чем недоступные цепи. Разница в среднем расстоянии d составляла всего около 0,5%, а радиальная ширина этого отражения была одинаковой как для D, так и для H-формы. Отражение 002 было сильнее в D-форме, а отражение 001, отсутствующее в H-дифракционной картине, появилось в профиле разности осей.Наличие этого отражения 001 после дейтерирования означает, что некоторые из дейтерированных цепей не имели такого же продольного смещения, как чистая целлюлоза Iβ или целлюлоза Iα.
Почти изотропные изменения диффузного рассеяния при дейтерировании привели к появлению широкого кольца повышенной интенсивности в области 15 нм -1 < q <20 нм -1 , а ниже 15 нм -1 , была уменьшена диффузная интенсивность (рис. 3В). Изотропные компоненты, ответственные за эти особенности диффузного рассеяния, включают нецеллюлозные компоненты, которые обмениваются с D 2 O и небольшими количествами остаточной связанной воды.
Эксперименты по спиновой диффузии протонов и пространственные отношения между упорядоченными и неупорядоченными цепями
Скорость, с которой ядерный магнетизм уравновешивается за счет спиновой диффузии между наноразмерными доменами, может использоваться для оценки расстояния между этими доменами. Эксперименты по спиновой диффузии протонов, основанные на этом принципе, были применены к древесине (Newman, 1992; Altaner et al., 2006) и бактериальной целлюлозе (Masuda et al., 2003). Использовалось двумерное (2D) представление этого эксперимента (Fernandes et al., 2011), чтобы изучить расположение упорядоченных и неупорядоченных областей в еловой целлюлозе, используя различия в их пространственном отделении от лигнина. Совершенно другой эксперимент по спин-диффузии 13 C на соломе кукурузы ( Zea mays ) (Foston et al., 2012) привел к аналогичным выводам.
Целлюлоза колленхимы сельдерея не лигнифицирована, но содержит небольшие остаточные количества гемицеллюлоз глюкоманнана и ксилоглюкана, которые, обладая относительно высокой термической подвижностью, могут использоваться вместо лигнина в качестве резервуаров спина протонов.На рисунке 4 показан 2D-спектр этого эксперимента. Перекрестные пики указывают на домены, которые удовлетворяют двум условиям: (1) домены различаются по подвижности и, следовательно, по намагниченности в начале периода смешения спинов протонов в последовательности импульсов; и (2) домены достаточно разделены в пространстве, чтобы потребовалось измеримое время, несколько миллисекунд, для уравновешивания намагниченности протонов посредством спиновой диффузии. Приведенное ниже назначение сигналов основано на Sturcová et al. (2004).
Рисунок 4.
2D-представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, отслеживаемых по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены. [См. Онлайн-статью для цветной версии этого рисунка.]
Рис. 4.
Двумерное представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, мониторинг которого проводился по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены.[Цветную версию этого рисунка см. В онлайн-статье.]
Одномерные спектры C-ЯМР 13 были аналогичны тем, которые наблюдались ранее (Sturcová et al., 2004), с сигналами C-4 от кристаллической целлюлозы (89 –90 частей на миллион [частей на миллион]) и неупорядоченной целлюлозы (84–85 частей на миллион) в примерном соотношении 1: 2. В 2D-спектре присутствовали кросс-пики между C-4 кристаллической целлюлозы и C-4 неупорядоченной целлюлозы, но они были относительно слабыми. Также присутствовал кросс-пик между C-6 кристаллической целлюлозы (65 ppm) и C-4 неупорядоченной целлюлозы (84–85 ppm).Перекрестные пики от гемицеллюлозы C-1 (101 ppm) и C-4 (83 ppm) до кристаллической целлюлозы C-4 были сильнее, чем от неупорядоченной целлюлозы C-4, и перекрестные пики от гемицеллюлозы C-1 и C-4 к кристаллической целлюлозе С-6 были еще сильнее. Наиболее сильные кросс-пики связаны с сигналами C-2, C-5 и C-6 от неупорядоченной целлюлозы при 73, 75 и 62 м.д., но каждый из этих сигналов, очевидно, также содержит вклад гемицеллюлоз и, следовательно, не может быть определен. однозначно трактуется.
Эти наблюдения согласуются с моделями, в которых неупорядоченная и кристаллическая целлюлоза не смешиваются, а находятся в непосредственной пространственной близости внутри каждой микрофибриллы, а гемицеллюлозы расположены вне микрофибрилл, вероятно, частично за пределами каждого небольшого пучка микрофибрилл, но разница в кросс-пике интенсивность кристаллической и неупорядоченной целлюлозы предполагает, что одна группа гемицеллюлоз была в тесной связи с поверхностями микрофибрилл.
Поляризованный дейтерированный FTIR и модели водородной связи
Поляризованный FTIR предоставляет информацию о направлении водородных связей. Исследования FTIR на древесине ели показали, что гидроксильные группы на неупорядоченной целлюлозе на поверхности микрофибрилл были доступны для дейтериевого обмена, за исключением тех случаев, когда микрофибриллы были плотно агрегированы вместе. Также были доступны гидроксильные группы на гемицеллюлозах. Обмен дейтерия сдвигает полосу растяжения, связанную с каждой гидроксильной группой, на более низкую частоту в 1 раз.34, из-за большей массы атома дейтерия, но сохраняет порядок полос, потому что структура не изменилась (Sturcová et al., 2003).
На фиг. 5 показано, что дейтерирование либо клеточных стенок колленхимы сельдерея, либо изолированной целлюлозы было обширным, но меньше, чем количество, предсказанное из соотношения 2: 1 неупорядоченной целлюлозы к кристаллической, оцененного с использованием сигналов C-4 в спектрах ЯМР. Образец полос растяжения O-D легче всего сравнить с рисунком полос растяжения O-H, если шкала частот искусственно сдвинута в 1 раз.34 (рис. 5). Эти две модели явно отличались. Полосы растяжения O-D были менее ориентированы в продольном направлении, чем полосы растяжения O-H, показывая, что меньшее количество водородных связей в дейтерированной фракции было направлено вдоль оси микрофибрилл, а больше — наружу. Как и в еловой целлюлозе (Fernandes et al., 2011), дейтерирование снизило интенсивность выступающего плеча на 3400 см -1 в области растяжения O-H, что соответствует гидроксильным группам, более слабо связанным водородными связями и более направленным наружу, чем в среднем.После сдвига оси частот на на 1,34 раза максимум довольно широкой группы перекрывающихся полос валентных колебаний OD совпал с этим плечом, которое первоначально было отнесено к аномальной части Iα-подобного спектра целлюлозы (Sturcová et al., 2004), но позже было показано, что в еловой целлюлозе они соответствуют недоступным гидроксильным группам в неупорядоченных областях (Fernandes et al., 2011). Отнесение к неупорядоченным доменам, а не к целлюлозе Iα, подтверждается данными, представленными здесь.Только часть интенсивности на 3 400 см -1 была смещена дейтерированием, подразумевая, что некоторые из этих неупорядоченных цепей в целлюлозе колленхимы сельдерея доступны для дейтерирования, а некоторые — нет.
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой. Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию.Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В онлайн-статье.]
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой.Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию. Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В статье в Интернете.]
ОБСУЖДЕНИЕ
Структура из микрофибриллы
Целлюлоза, которую можно условно назвать кристаллической, и неупорядоченная целлюлоза, отличная от известных кристаллических форм целлюлозы Iα и Iβ, были идентифицированы всеми использованными методами.Из этого не следует, что одни и те же неупорядоченные фракции наблюдались всеми этими методами (т.е. что цепи, идентифицированные как неупорядоченные, в одних и тех же отношениях отличались от известных кристаллических форм). Наблюдались конформационный беспорядок, беспорядок упаковки и беспорядок водородных связей.
Спектры 13 C-ЯМР целлюлозы колленхимы сельдерея (рис. 4) идентифицировали цепи, которые отличались по конформации как от целлюлозы Iα, так и от целлюлозы Iβ, имеющие некоторую свободу вращения вокруг связи C5-C6 и имеющие C-6 В конформации преобладают гош-гош ( gg ) и гош-транс ( gt ) формы (Sturcová et al., 2004), как и в еловой целлюлозе (Fernandes et al., 2011), хотя соотношение gg — gt значительно отличалось от еловой (Sturcová et al., 2004). Спектры дейтерирования FTIR целлюлозы сельдерея показали, что некоторые домены имели паттерны водородных связей, отличные от наблюдаемых в целлюлозе Iα и целлюлозе Iβ, с пониженной средней прочностью водородных связей (более высокая средняя частота растяжения O-H) и меньшей общей тенденцией к осевой ориентации водорода. Изменение ориентации водородных связей было очевидным как в цепях, доступных для дейтериевого обмена, так и в недоступных цепях, и соответствовало переходу от транс-гош ( tg ) конформации C-6, обнаруженной в кристаллической целлюлозе, к gg . и конформеры GT .Подобные наблюдения на еловой целлюлозе были рационализированы с точки зрения модели, в которой ядро каждой микрофибриллы было занято в основном кристаллической целлюлозой, а поверхность микрофибрилл была занята цепями целлюлозы с несколько иной конформацией и водородными связями, некоторые из которых были доступны для обмена дейтерия, а другие были представлены. недоступны из-за боковой агрегации микрофибрилл. Подобная модель согласуется с FTIR-спектрами сельдерея (рис. 5), более ранними данными ЯМР (Sturcová et al., 2004), а также данные спин-диффузионного ЯМР, представленные здесь (рис. 4). По данным ЯМР спин-решеточной релаксации 13 C (Viëtor et al., 2002), целлюлоза колленхимы сельдерея имеет более высокую долю конформера C-6 gg, чем еловая целлюлоза. Размеры элементарной ячейки значительно различались между этими двумя видами целлюлозы, среднее расстояние между листами было больше в целлюлозе колленхимы сельдерея, а осевой размер был немного меньше. Молекулярное моделирование предложило связь между конформацией C-6 и шагом решетки (Matthews et al., 2012). Как и в случае с еловой целлюлозой, было невозможно и, возможно, бессмысленно различать формы целлюлозы Iα и Iβ с помощью ЯМР при наличии такого большого конформационного нарушения (Matthews et al., 2011).
Модели микрофибрилл с неупорядоченными поверхностями неоднократно предлагались на основе данных ЯМР (Newman, 1992; Viëtor et al., 2002; Bergenstråhle et al., 2008), но было неясно, как они должны согласовываться с полученными моделями. от дифракции рентгеновских лучей, которая более чувствительна к структуре упаковки цепей, чем к конформации цепи или водородным связям.Данные широкоугольного рассеяния нейтронов (WANS) (рис.3) показали, что в доменах, доступных для дейтерирования, цепи были упакованы латерально таким же образом и почти с такими же средними расстояниями, как и в кристаллических областях, несмотря на различия в конформации. и водородные связи, продемонстрированные спектроскопическими экспериментами, несмотря на значительные статистические различия в упаковке цепей повсюду в микрофибрилле. Повсюду в каждой микрофибрилле листы цепочек были уложены друг на друга с более широким средним интервалом, чем в более кристаллических целлюлозах.Дальнейшее следствие состоит в том, что большинство цепей, доступных для дейтерирования, были расположены на поверхностях микрофибрилл, а не в «аморфных» сегментах микрофибриллы, занимающих всю ширину микрофибриллы, потому что цепи внутри таких сегментов должны быть неплотно упакованы, чтобы вода могла стекать. входить.
Судя по осевому отражению 004, доступные для дейтерирования цепи оказались немного более вытянутыми в продольном направлении, чем недоступные цепи, возможно, из-за скручивания микрофибрилл.Продольное смещение было частично нарушено в цепочках, доступных для дейтерирования, что усложняло различение форм Iα и Iβ.
Широкий диапазон расстояний между боковыми цепями в нецеллюлозной фракции, доступный для дейтерирования, может быть выведен из диффузной, неориентированной части нейтронографических изображений. Вода, связанная с нецеллюлозной фракцией, рассеивала рентгеновские лучи аналогично жидкой воде, предполагая аналогичную функцию распределения пар, но из-за пониженной тепловой подвижности величина ее вклада в структуру WAXS была больше, чем ожидалось для объемной жидкой воды.
Диаметр и форма микрофибриллы
Данные SANS предполагают диаметр микрофибрилл, приблизительно равный межцентровому расстоянию от 2,9 до 3,0 нм в сухом состоянии. Предполагая круглое поперечное сечение и учитывая относительно рыхлую упаковку цепей, наблюдаемую при широкоугольном рассеянии, этот номинальный диаметр будет соответствовать 21–22 цепям. Микрофибриллы, конечно, не имеют круглой формы, и фактическое количество цепочек зависит от формы поперечного сечения.Наблюдения WANS подтвердили, что поверхностные цепочки располагались на той же неупорядоченной решетке, что и внутренние цепочки, и должны быть включены в расчеты размеров Шеррера из WAXS.
Межцентровое расстояние SANS и поперечные размеры по Шерреру были аналогичны тем, которые наблюдались для еловой целлюлозы (Fernandes et al., 2011). Размер [200] Шеррера был больше, чем среднее значение размеров [1-10] и [110], что согласуется с моделью прямоугольной микрофибриллы (Matthews et al., 2006; Fernandes et al., 2011), который имеет относительно гидрофобные [200] грани кристаллов, открытые сверху и снизу (рис. 6), вместо диагональных [1-10] и [110] граней кристаллов, открытых в целлюлозах водорослей (Baker et al. ., 1997). Однако опора для прямоугольной, а не ромбовидной модели менее надежна, чем для ели, из-за сложности коррекции беспорядка с использованием очень слабого отражения 400 и из-за проблем, присущих оценке отдельных ширины перекрывающихся 1-10 и 110 отражений.Учитывая наблюдаемое обширное нарушение, нельзя даже с уверенностью сказать, что микрофибриллы первичной стенки имеют постоянную форму поперечного сечения.
Рисунок 6.
Модели поперечного сечения микрофибрилл колленхимы сельдерея. A, 24-цепочечная ромбовидная модель, 2,9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться.Расчетные длины столбцов [200], [1-10] и [110] были следующими, соответственно: модель A, 2,5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 6.
Модели для поперечного сечения микрофибрилл колленхимы сельдерея. A, модель ромбовидной формы с 24 цепями, 2.9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться. Расчетные длины столбцов [200], [1-10] и [110] были соответственно следующими: модель A, 2.5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В статье в Интернете.]
Прямоугольное поперечное сечение с уложенными друг на друга листами из трех цепей будет соответствовать одной трети цепей, находящихся в центральное положение в каждом листе, конформационно сходное с кристаллическими целлюлозами, как наблюдалось с помощью ЯМР.Наблюдение единственного малоуглового пика Брэгга согласуется со слегка диспергированными расстояниями между микрофибриллами, которые можно было бы ожидать от примерно прямоугольных микрофибрилл, возможно, скрученных, находящихся в прерывистом контакте друг с другом. Могут быть приспособлены как постоянные, так и слегка изменяющиеся диаметры микрофибрилл, а также небольшое количество микрофибрилл, значительно отличающихся от доминирующих размеров.
Число цепей на микрофибриллу имеет отношение к структуре комплексов, которые синтезируют целлюлозу во время роста.Если количество цепей кратно шести, что соответствует геометрии синтетического комплекса, тогда стопка из восьми трехцепочечных листов (24 цепи) даст наиболее близкое совпадение с размером Шеррера 200 (рис. 6B), как и для еловая целлюлоза (Fernandes et al., 2011). Однако в нашем случае неопределенность довольно велика, и уровень беспорядка упаковки предполагает, что возможны нерегулярные и переменные сечения. Модель из 18 цепей (6 × 3) не исключена для еловой целлюлозы и согласуется с данными, представленными здесь (рис.6C), если размер 200 по Шерреру был увеличен дополнительными цепями гемицеллюлозы, как это было предложено для ели (Fernandes et al., 2011), или увеличен путем наложения одной микрофибриллы поверх другой на короткие расстояния.
Кристалличность
Что означает «кристалличность», зависит от методов, используемых для обнаружения компонентов, которые отличаются от кристаллической целлюлозы. Между спектроскопическими методами обнаружения цепей, отличающихся по конформации от общепринятых форм кристаллической целлюлозы и, следовательно, демонстрирующих различные модели водородных связей, наблюдалась некоторая степень согласованности.Это включало нецеллюлозные полисахариды, и, в частности, в спектрах FTIR было трудно провести четкое различие между вкладом нецеллюлозных полисахаридов и некоторой неупорядоченной целлюлозы.
Эксперименты с дейтерированием WANS показали, что эти конформационные различия мало влияют на упаковку целлюлозных цепей или дифракцию. Структура микрофибрилл явно содержала значительный беспорядок в боковой упаковке цепей, но этот беспорядок распространялся по всей структуре и не ограничивался доступными для дейтерия поверхностями.Кристалличность, оцениваемая дифракционными методами, — это не то же самое, что кристалличность, оцениваемая спектроскопически. В обоих случаях кажущаяся кристалличность будет увеличиваться с увеличением диаметра микрофибрилл, но как прямое следствие уменьшения уширения по Шерреру в дифракционных экспериментах и из-за уменьшения отношения поверхности к объему в спектроскопических экспериментах, которые позволяют различать поверхностные цепи.
Агрегация микрофибрилл
Агрегация микрофибрилл в более крупные пучки хорошо известна для древесины хвойных пород (Wickholm et al., 1998; Фахлен и Сальмен, 2005 г .; Fernandes et al., 2011). Фибриллярные единицы шириной от 5 до 10 нм, иногда до 40 нм, наблюдались с помощью электронной микроскопии (McCann et al., 1990; Sugimoto et al., 2000), но их интерпретация осложняется возможным включением нецеллюлозных полимеров. . Представленные здесь данные включают две прямые линии доказательств агрегации микрофибрилл первичной стенки в колленхиме. Доказательства малоуглового брэгговского рассеяния рентгеновских лучей (Kennedy et al., 2007a) и нейтронов ясно показали, что агрегация существует. Диапазон расстояний между микрофибриллами было трудно сделать вывод, потому что интенсивность рассеяния сильно увеличивалась с разделением микрофибрилл, так что хорошо разделенные микрофибриллы были чрезмерно представлены на диаграммах рассеяния. Низкий контраст МУРН в сухом состоянии и сходство межцентрового расстояния МУРН со средними размерами Шеррера WAXS предполагают, что некоторые сегменты микрофибрилл оставались в прямом контакте друг с другом, тогда как другие сегменты микрофибрилл становились хорошо разделенными при гидратации.
Спектры дейтерирования FTIR как от стенок клеток колленхимы, так и от изолированной целлюлозы включали полосы растяжения гидроксила с центром на 3 400 см -1 с нейтральной поляризацией. Эти гидроксильные полосы растяжения были отнесены к гидроксильным группам, отличным от всех групп Iα и Iβ кристаллических форм целлюлозы, более ориентированным наружу по ориентации и частично, но не полностью доступным для дейтерирования. Принимая во внимание также данные SANS и ЯМР, соответствующие гидроксильные группы могут быть отнесены к поверхностям микрофибрилл.Поскольку эти поверхности были только частично доступны для D 2 O, отсюда следует, что микрофибриллы были агрегированы вместе на части своей длины с границами раздела, непроницаемыми для воды. Эти особенности были обнаружены в древесине ели, но были также различия: древесина ели имеет разные нецеллюлозные полимеры, что, по-видимому, приводит к более тесному связыванию между поверхностями микрофибрилл и к напряжениям, которые искажают геометрию элементарной ячейки при гидратации (Fernandes et al., 2011).
Полоса валентных колебаний гидроксила 3,400 см -1 в FTIR-спектре, перемещенная до 2,540 см -1 за счет обмена D 2 O, ранее была неправильно отнесена к аберрантной форме целлюлозы Iα (Sturcová et al., 2004), но, по-видимому, является маркером поверхностных цепей и наличия агрегации микрофибрилл, если эти поверхностные цепи недоступны для дейтерирования. Интенсивность этой полосы сильно уменьшилась, когда целлюлоза из колленхимы сельдерея была подвергнута гидротермальному отжигу, что привело к слиянию микрофибрилл в более крупные кристаллические единицы с уменьшенным соотношением поверхности к объему (Sturcová et al., 2004). Интенсивность FTIR на уровне 3400 см или около него -1 была зарегистрирована в препаратах клеточных стенок, например, арабидопсиса дикого типа ( Arabidopsis thaliana ; MacKinnon et al., 2006), предполагая, что эта форма агрегации не является уникальной особенностью целлюлозы колленхимы сельдерея, но более широко распространена в первичных клеточных стенках.
Влияние на рост растений
Размер и структура микрофибрилл первичной стенки дают важные ключи к разгадке природы ферментных комплексов, ответственных за ориентированный синтез целлюлозы и, следовательно, за направленную экспансию клеток. Размножение растительных клеток требует разрушения сети нековалентно сшитых микрофибрилл в клеточной стенке.Понимание топологии этой сети, следовательно, необходимо, и недавно принятая идея о том, что ксилоглюканы просто покрывают и связывают поверхности микрофибрилл, была поставлена под сомнение (Bootten et al., 2004; Park and Cosgrove, 2012a). Данные спин-диффузии ЯМР (рис. 4) показали, что некоторые цепи гемицеллюлозы, вероятно, ксилоглюканы, располагались близко к поверхностям микрофибрилл и, таким образом, вероятно, лежали между микрофибриллами внутри агрегатов микрофибрилл. Однако другие сегменты микрофибрилл, по-видимому, находятся в прямом контакте без промежуточных полимеров, демонстрируя нерегулярный характер агрегации, которого можно было бы ожидать, если микрофибриллы были скрученными и нецилиндрическими, и предполагая, что взаимодействия целлюлоза-целлюлоза, а также ксилоглюкан-целлюлоза должны быть разбиты на отдельные агрегированные микрофибриллы.Др. Механизмы необходимы для отделения одного пучка микрофибрилл от следующего во время роста путем разрушения ксилоглюкана или др. Мостиков между пучками микрофибрилл (Anderson et al., 2010). Агрегация микрофибрилл вдоль части их длины, в то время как другие части их длины остаются свободными для разделения или в них вставлены другие полисахариды, поднимает интересные вопросы о действии экспансинов (Cosgrove, 2005) и о том, как действует действие определенных глюканов. гидролазы взаимодействуют с таковой ксилоглюкан-специфической эндоглюканазы (Park and Cosgrove, 2012b).
Интересен высокий уровень беспорядка на границах раздела, наиболее важных для роста, между микрофибриллами и нецеллюлозными полимерами, а также между одной микрофибриллой и другой. Примеры прочных биологических материалов из царства животных показывают, что неупорядоченные домены могут вносить вклад в ударную вязкость (высокую энергию разрушения) при внешнем стрессе (van Beek et al., 2002). В первичных клеточных стенках устойчивость к разрушению должна сочетаться с контролируемой ферментативно-опосредованной продуктивностью, по-видимому, на неупорядоченных границах раздела, что обеспечивает рост.
Последствия разложения целлюлозы
Доступность микрофибрилл целлюлозы для целлюлаз является важным фактором превращения лигноцеллюлозы в биотопливо и еще более важна в случае нелгнифицированной биомассы. Нестабильность целлюлозы часто связывают с ее кристалличностью (Arantes and Saddler, 2010), а «декристаллизацию» иногда называют целью процессов предварительной обработки (Beckham et al., 2011), без полного согласия относительно того, что означают эти термины.Степень, в которой микрофибриллы целлюлозы имеют относительно гидрофобную [200] открытую поверхность, будет положительно влиять на начальную стадию деградации целлюлазами, которые специфически связываются с этой поверхностью (Dagel et al., 2011; Liu et al., 2011). Было бы интересно узнать больше о специфичности связывания оксидаз целлюлозы GH61 / CBM33 (Forsberg et al., 2011; Quinlan et al., 2011), некоторые из которых также связываются с одной и той же поверхностью (Li et al., 2012). Агрегация микрофибрилл целлюлозы — это фактор сопротивляемости, который заслуживает большего внимания, и теперь к нему можно подойти с помощью экспериментов по дейтерированию FTIR.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Было выделенонитей колленхимы из зрелых черешков сельдерея ( Apium graveolens ), клеточные стенки и их целлюлозная фракция были приготовлены, как описано (Sturcová et al., 2004), и хранились в сухом виде. Подробнее см. Дополнительные материалы и методы S1.
SANS
ЭкспериментыSANS были выполнены на канале D11 в Институте Лауэ-Ланжевена на клеточных стенках колленхимы сельдерея и изолированной целлюлозе.Подробнее об эксперименте см. Дополнительные материалы и методы S1.
WAXS
дифрактограммы рентгеновских лучей получали при температуре окружающей среды с использованием дифрактометра Rigaku R-axis / RAPID для рентгеновских пластин с источником излучения с молибденовой рентгеновской эмиссией Kα (λ = 0,07071 нм). Пучок коллимировался до диаметра 0,5 мм. Образцы имели толщину 0,7 мм в направлении, параллельном пучку, а их другие размеры превышали диаметр пучка. Дифрактограммы были собраны в режиме перпендикулярного пропускания, за исключением экспериментов с наклоном для измерения осевых отражений.Расстояние от центра дифракционной картины было выражено как q = 4πsinθ / λ, где 2θ — угол рассеяния. Для получения дополнительных сведений об экспериментальных методах и методах обработки данных (Fernandes et al., 2011) см. Дополнительные материалы и методы S1.
WANS
Дифракция нейтронов проводилась на канале D19 в Институте Лауэ-Ланжевена при средней длине волны нейтронов 0,242 нм и расстоянии от образца до детектора 0,756 м. Образцы размером 25 × 15 × 1 мм собирали из нескольких выровненных нитей колленхимы и выдерживали при контролируемом давлении воды или паров D 2 O в проточном контейнере со стенками из алюминиевой фольги.Подробнее см. Дополнительные материалы и методы S1.
ЯМР
A 1 H Эксперимент по спиновой диффузии Гольдмана-Шена с детектированием по спектру 13 C после кросс-поляризации был проведен на клеточных стенках колленхимы сельдерея, гидратированных 0,2 см. 3 г −1 D 2 O. Экспериментальные процедуры и анализ данных, включая 2D-представление данных, были такими, как описано (Fernandes et al., 2011).
Дейтерирование FTIR
Стенки клеток колленхимы сельдерея и изолированную целлюлозу помещали в проточную ячейку с окнами из фторида бария и заменяли воздухом, насыщенным О 2 О, до тех пор, пока спектры не перестали изменяться. Затем газовую линию переключали на азот, тщательно осушая, пропуская через сухое молекулярное сито 4А, а затем пентоксид фосфора (Sicapent; Aldrich). Спектры FTIR собирали с помощью спектрометра Thermo Nicolet Nexus, оборудованного приставкой для микроскопа Nicolet Continuum, имеющей охлаждаемый жидким азотом детектор теллурида кадмия ртути (Fernandes et al., 2011).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Rigaku, Ltd. за предоставленный инструмент и Fujiang Zhang за получение некоторых спектров FTIR.
Глоссарий
SAXS
Малоугловое рассеяние рентгеновских лучей
D 2 O
SANS
Малоугловое рассеяние нейтронов
WAXS 9407
широкоугольное рассеивание
WANS
широкоугольное рассеяние нейтронов
FTIR
Инфракрасное преобразование Фурье
2D
ppm
LITERATURE CITED
(
2006
)Пространственные отношения между полимерами в ели ситкинской: исследования спиновой диффузии протонов
.Holzforschung
60
:665
—673
(
2010
)Визуализация в реальном времени переориентации целлюлозы во время расширения клеточной стенки в корнях Arabidopsis
.Plant Physiol
152
:787
—796
(
2009
)Выяснение механизмов сборки и субъединичного взаимодействия целлюлозосинтазного комплекса вторичных клеточных стенок Arabidopsis
.J Biol Chem
284
:3833
—3841
(
1997
)Атомно-силовая микроскопия высокого разрешения микрокристаллов нативной целлюлозы I Valonia
.J Struct Biol
119
:129
—138
(
2011
)Происхождение стойкости биомассы на молекулярном уровне: свободные энергии декристаллизации для четырех обычных полиморфов целлюлозы
.J Phys Chem B
115
:4118
—4127
(
2008
)Динамика границ раздела целлюлоза-вода: времена спин-решеточной релаксации ЯМР, рассчитанные на основе атомистического компьютерного моделирования
.J Phys Chem B
112
:2590
—2595
(
2004
)Твердотельный 13 C-ЯМР-спектроскопия показывает, что ксилоглюканы в первичных клеточных стенках маша (Vigna radiata L.) происходят в разных доменах: новая модель взаимодействий ксилоглюкан-целлюлоза в клеточной стенке
.J Exp Bot
55
:571
—583
(
2009
)Растения контролируют свойства и работу своих органов посредством ориентации фибрилл целлюлозы в их клеточных стенках
.Интегр Комп Биол
49
:69
—79
(
2005
)Рост клеточной стенки растений
.Nat Rev Mol Cell Biol
6
:850
—861
(
2011
)Визуализация на месте отдельных углеводсвязывающих модулей на микрофибриллах целлюлозы
.J Phys Chem B
115
:635
—641
(
1996
)Паренхиматозная клеточная целлюлоза из жома сахарной свеклы: получение и свойства
.Целлюлоза
3
:183
—188
(
2005
)Распределение пор и матрицы в стенке волокна, выявленное с помощью атомно-силовой микроскопии и анализа изображений
.Биомакромолекулы
6
:433
—438
(
1999
)Пакет WinGX для кристаллографии низкомолекулярных монокристаллов
.J Appl Cryst
32
:837
—838
(
2011
)Наноструктура микрофибрилл целлюлозы в древесине ели
.Proc Natl Acad Sci USA
108
:E1195
—E1203
(
2011
)Расщепление целлюлозы белком CBM33
.Protein Sci
20
:1479
—1483
(
2012
)Твердотельное селективное (13) C возбуждение и спин-диффузионный ЯМР для определения пространственных размеров в стенках растительных клеток
.J Agric Food Chem
60
:1419
—1427
(
2000
)Характеристика поперечных связей между микрофибриллами целлюлозы и их появление во время элонгационного роста в эпикотиле гороха
.Физиология растительных клеток
41
:486
—494
(
2011
)Кортикальные микротрубочки оптимизируют кристалличность клеточной стенки для обеспечения однонаправленного роста Arabidopsis
.Завод J
66
:915
—928
(
1999
)Выражение паттерна в растениях: сочетание молекулярных и основанных на исчислении биофизических парадигм
.Am J Bot
86
:1059
—1076
и другие. (
2012
)Кристалличность микрофибрилл целлюлозы снижается за счет мутации С-концевых остатков трансмембранной области CESA1A903V и CESA3T942I синтазы целлюлозы
.Proc Natl Acad Sci USA
109
:4098
—4103
(
2007
)Устойчивость биомассы: инженерные установки и ферменты для производства биотоплива
.Наука
315
:804
—807
(
2009
)Различия между сортами в размерах микрофибрилл целлюлозы, наблюдаемые с помощью инфракрасной спектроскопии
.Целлюлоза
16
:1
—8
(
1995
)Радиальная упаковка, порядок и беспорядок в фибриллах коллагена
.Biophys J
68
:1661
—1670
(
1992
)Контроль толщины клеточных стенок колленхимы пектинами
.Планта
187
:218
—220
(
2011
)Стенки растительных клеток: супрамолекулярные ансамбли
.Food Hydrocoll
25
:257
—262
(
2007a
)Диаметр микрофибрилл в целлюлозе колленхимы сельдерея: рассеяние рентгеновских лучей и данные ЯМР
.Целлюлоза
14
:235
—246
(
2007b
)Влияние гидратации на расстояние между микрофибриллами целлюлозы первичной стенки: исследование малоуглового рентгеновского рассеяния
.Целлюлоза
14
:401
—408
(
2008
)Выявление структурного и функционального разнообразия клеточных стенок растений
.Curr Opin Plant Biol
11
:308
—313
(
2001
)Рентгеновская структура мерсеризованной целлюлозы II при разрешении 1 Å
.Биомакромолекулы
2
:410
—416
(
2012
)Структурная основа нацеливания на субстрат и катализа грибковыми полисахаридмонооксигеназами
.Структура
20
:1051
—1061
(
2011
)Целлобиогидролаза гидролизует кристаллическую целлюлозу на гидрофобных поверхностях
.J Biol Chem
286
:11195
—11201
(
2006
)Структура и анизотропия клеточной стенки в procuste , мутанте целлюлозосинтазы Arabidopsis thaliana
.Планта
224
:438
—448
(
2005
)Расширение клеточной стенки приводит к координатному разделению параллельных микрофибрилл: данные сканирующей электронной микроскопии и атомно-силовой микроскопии
.Завод J
43
:181
—190
(
2003
)Твердотельный 13 C и 1 H спин-диффузионный ЯМР-анализ структуры микрофибрилл бактериальной целлюлозы
.Твердотельный ядерный магнитный резонон
23
:198
—212
(
2012
)Сравнение моделирования бета целлюлозы I с тремя силовыми полями углеводов
.J Chem Theory Comput
8
:735
—748
(
2011
)Высокотемпературные свойства целлюлозы I
.J Phys Chem B
115
:2155
—2166
(
2006
)Исследования микрокристаллической целлюлозы Ibeta
с помощью компьютерного моделирования.Углеводородный остаток
341
:138
—152
(
1990
)Прямая визуализация поперечных связей в первичной клеточной стенке растения
.J Cell Sci
96
:323
—334
(
2008
)Структура и биосинтез пектина
.Curr Opin Plant Biol
11
:266
—277
(
1992
)Ядерно-магнитно-резонансное исследование пространственных соотношений между химическими компонентами в стенках деревянных клеток
.Holzforschung
46
:205
—210
(
2009
)Структура и свойства микрофибриллы целлюлозы
.J Wood Sci
55
:241
—249
(
2002
)Кристаллическая структура и система водородных связей в целлюлозе Ibeta по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
124
:9074
—9082
(
2003
)Кристаллическая структура и система водородных связей в целлюлозе I (альфа) по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
125
:14300
—14306
(
2008
)Генетические доказательства того, что активность синтазы целлюлозы влияет на организацию кортикального массива микротрубочек
.Plant Physiol
147
:1723
—1734
(
2012a
)Изменения биомеханических свойств клеточной стенки у ксилоглюкан-дефицитного мутанта xxt1 / xxt2 Arabidopsis
.Plant Physiol
158
:465
—475
(
2012b
)Пересмотренная архитектура первичных клеточных стенок, основанная на биомеханических изменениях, индуцированных субстрат-специфическими эндоглюканазами
.Plant Physiol
158
:1933
—1943
и другие. (
2011
)Анализ окислительного разложения целлюлозы металлоферментом меди, который использует компоненты биомассы
.Proc Natl Acad Sci USA
108
:15079
—15084
и другие.(
2012
)CHITINASE-LIKE1 / POM-POM1 и его гомолог CTL2 представляют собой взаимодействующие с глюканом белки, важные для биосинтеза целлюлозы у Arabidopsis
.Plant Cell
24
:589
—607
(
2008
)Краткая история SHELX
.Acta Crystallogr A
64
:112
—122
(
2006
)Синтез целлюлозы у высших растений
.Annu Rev Cell Dev Biol
22
:53
—78
(
2004
)Структурные детали кристаллической целлюлозы из высших растений
.Биомакромолекулы
5
:1333
—1339
(
2003
)Поляризованная колебательная спектроскопия волоконных полимеров: водородные связи в целлюлозе II
.Биомакромолекулы
4
:1589
—1595
(
2000
)Новые методы позволяют проводить сравнительный анализ ориентации микротрубочек, текстуры стенок и скорости роста интактных корней Arabidopsis
.Plant Physiol
124
:1493
—1506
(
2009
)Динамическая координация систем цитоскелета и клеточной стенки во время морфогенеза растительных клеток
.Curr Biol
19
:R800
—R811
(
2002
)Молекулярная структура шелка драглайна пауков: складывание и ориентация белкового остова
.Proc Natl Acad Sci USA
99
:10266
—10271
(
2002
)Конформационные особенности кристаллической целлюлозы высших растений
.Завод J
30
:721
—731
(
2004
)Полиморфизм целлюлозы семейства I: повторное исследование целлюлозы IVI
.Биомакромолекулы
5
:1385
—1391
(
1998
)Определение некристаллических форм в целлюлозе I методом спектроскопии ЯМР CP / MAS C-13
.Углеводородный остаток
312
:123
—129
(
2007
)Организация боковых цепей пектинового арабинана и галактана в ассоциации с микрофибриллами целлюлозы в первичных клеточных стенках и связанных моделях предусматривается
.J Exp Bot
58
:1795
—1802
Заметки автора
© Американское общество биологов растений, 2013 г. Все права защищены.
© Автор (ы) 2013. Опубликовано Oxford University Press от имени Американского общества биологов растений. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0 /), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
Структура микрофибрилл целлюлозы в первичных клеточных стенках колленхимы | Физиология растений
Аннотация
В первичных стенках растущих клеток растений глюкозно-полимерная целлюлоза собрана в длинные микрофибриллы диаметром несколько нанометров. Жесткость и ориентация этих микрофибрилл контролируют рост клеток; Следовательно, синтез целлюлозы является ключевым фактором роста и морфогенеза растений.Колленхима сельдерея ( Apium graveolens ) является полезной модельной системой для изучения структуры микрофибрилл первичной стенки, поскольку ее микрофибриллы ориентированы с необычной однородностью, что облегчает проведение спектроскопических и дифракционных экспериментов. Используя комбинацию методов рассеяния рентгеновских лучей и нейтронов с колебательной спектроскопией и спектроскопией ядерного магнитного резонанса, мы показываем, что микрофибриллы колленхимы сельдерея имели средний диаметр от 2,9 до 3,0 нм, с наиболее вероятной структурой, содержащей 24 цепи в поперечном сечении, расположенные в восьми атомах водорода. -связанные листы из трех цепей с обширным беспорядком в боковой упаковке, конформации и водородной связи.Аналогичная структура из 18 цепей и структуры из 24 цепей разной формы хуже соответствовали данным. Конформационный беспорядок в основном ограничивался поверхностными цепями, а беспорядок в упаковке цепей — нет. То есть по положению и ориентации поверхностные цепи соответствовали неупорядоченной решетке, составляющей ядро каждой микрофибриллы. Были доказательства того, что соседние микрофибриллы нековалентно агрегированы вместе на части своей длины, предполагая, что необходимость разрушения этих агрегатов может быть сдерживающим фактором роста и гидролиза целлюлозы для производства биотоплива.
Рост и форма растений контролируются точно ориентированным расширением стенок отдельных клеток. Движущая сила для размножения клеток является осмотической, но скорость и направление экспансии контролируются механическими свойствами клеточной стенки (Szymanski and Cosgrove, 2009). Расширяющиеся первичные клеточные стенки представляют собой нанокомпозитные материалы, в которых длинные микрофибриллы целлюлозы диаметром несколько нанометров проходят через гидратированную матрицу ксилоглюканов, пектинов и других полимеров (Knox, 2008; Mohnen, 2008; Szymanski and Cosgrove, 2009; Scheller. и Ульвсков, 2010).Нативные микрофибриллы целлюлозы частично кристаллические (Nishiyama, 2009; Fernandes et al., 2011). Раньше считалось, что целлюлоза первичной стенки имеет уникальную кристаллическую структуру, называемую целлюлозой IV 1 (Dinand et al., 1996), но данные ЯМР предполагают наличие форм, подобных лучше охарактеризованным кристаллическим формам целлюлозы Iα и Iβ вместе с крупными кристаллическими формами целлюлозы. количества менее упорядоченной целлюлозы (Wickholm et al., 1998; Sturcová et al., 2004; Wada et al., 2004). Тем не менее, целлюлоза гораздо более упорядочена, чем любой другой компонент первичной клеточной стенки (Bootten et al., 2004), в соответствии с его ключевой ролью обеспечения силы и контроля роста.
Жесткость клеточной стенки наибольшая в направлении микрофибрилл целлюлозы, где рост является направленным, а преобладающая ориентация микрофибрилл обычно поперечна направлению роста (Green, 1999; MacKinnon et al., 2006; Szymanski and Cosgrove, 2009 г.). Расширение клеточной стенки требует либо увеличения расстояния между микрофибриллами (Marga et al., 2005), либо проскальзывания между ними (Cosgrove, 2005), либо того и другого, и микрофибриллы переориентируются в направлении роста (Anderson et al., 2010). Полимерные поперечные мостики между микрофибриллами (McCann et al., 1990), как полагают, противостоят этим деформациям наноструктуры клеточной стенки и, таким образом, контролируют скорость роста. До недавнего времени наибольшее внимание уделялось мостиковым соединениям ксилоглюканов, водородным связям с поверхностями микрофибрилл (Scheller, Ulvskov, 2010). Однако есть свидетельства того, что не все ксилоглюканы расположены надлежащим образом (Fujino et al., 2000; Park and Cosgrove, 2012a) и что могут быть задействованы другие мостиковые полимеры (Zykwinska et al., 2007). Также было высказано предположение, что пучки агрегированных микрофибрилл, а не отдельные микрофибриллы, могут быть ключевыми структурными единицами в первичных клеточных стенках (Anderson et al., 2010), как в древесине (Fahlén and Salmén, 2005; Fernandes et al., 2011). ). В таком случае отдельные микрофибриллы могут соединяться между пучками микрофибрилл. Таким образом, рост растительных клеток не совсем понятен, и нам нужна дополнительная информация о том, как контролируется ориентация целлюлозы, и о природе мостиковых полимеров, о поверхностях целлюлозы, с которыми эти полимеры связываются, и о сцеплении между поверхностями микрофибрилл, которые могут опосредовать агрегацию.
Микрофибриллы целлюлозы синтезируются на поверхности клетки большими ферментными комплексами, имеющими гексагональную симметрию, иногда называемыми «розетками» (Somerville, 2006). Каждый комплекс содержит несколько синтаз целлюлозы, которые различаются между первичными клеточными стенками и древесиной, хотя внешний вид комплексов похож (Somerville, 2006; Atanassov et al., 2009). Одновременный синтез с одного и того же конца всех цепей в микрофибрилле нативной целлюлозы является причиной их параллельности (Nishiyama et al., 2002, 2003), в отличие от энтропийно предпочтительной антипараллельной структуры, обнаруженной в искусственных целлюлозах, таких как вискоза (Langan et al., 2001). Очевидно, что количество цепей в микрофибрилле и количество целлюлозосинтаз в синтетическом комплексе взаимосвязаны. Принято считать, что количество цепочек делится на шесть, что соответствует симметрии гексагональной розетки и 36-цепочечным моделям (Himmel et al., 2007), ограниченным гидрофильными гранями кристаллов [110] и [1-10], поскольку в водорослевых целлюлозах (Bergenstråhle et al., 2008), получили широкое распространение. Сборка и ориентация целлюлозы взаимосвязаны, поскольку некоторые мутанты целлюлозосинтазы имеют фенотипы, дефектные по ориентации целлюлозы и растительной форме, а также обедненные содержанием целлюлозы (Paredez et al., 2008). У некоторых других мутантных линий, по-видимому, затрагивается кристалличность микрофибрилл (Fujita et al., 2011; Harris et al., 2012; Sánchez-Rodríguez et al., 2012).
Следовательно, детальное понимание структуры микрофибрилл целлюлозы первичной стенки поможет нам понять синтез целлюлозы, а также рост и структурную механику живых растений (Burgert and Fratzl, 2009).Первичные клеточные стенки и их целлюлозные скелеты также влияют на такие характеристики качества пищевых продуктов, как хрусткость салата из овощей и яблок ( Malus domestica ; Jarvis, 2011). Когда биотопливо производится из лигноцеллюлозной биомассы, лигнификация приводит к устойчивости (Himmel et al., 2007), но некоторые типы клеток у видов Miscanthus , просо ( Panicum virgatum ) и пожнивные остатки имеют только первичные стенки. без лигнина, и сопротивляемость этому зависит от природы микрофибрилл целлюлозы (Beckham et al., 2011).
Относительно подробная структура недавно была предложена для микрофибрилл древесины ели ( Picea spp.) (Fernandes et al., 2011), которые имеют диаметр 3,0 нм, что позволяет разместить только около 24 цепочек целлюлозы. Данные рентгеновской дифракции подтверждают «прямоугольную» форму (Matthews et al., 2006), ограниченную гранями [010] и [200]. Наблюдался значительный беспорядок, увеличивающийся к поверхности, и микрофибриллы были агрегированы в пучки размером примерно 15-20 нм в поперечнике, причем некоторые, но не все, боковые границы раздела были устойчивы к воде (Fernandes et al., 2011). Неупорядоченные домены являются особенностью других прочных биологических материалов, таких как шелк паука (van Beek et al., 2002).
Следовательно, представляет интерес, могут ли какие-либо из этих свойств древесной целлюлозы также быть обнаружены в микрофибриллах целлюлозы первичных (растущих) клеточных стенок. Было бы особенно полезно охарактеризовать нарушение, которое, как известно, присутствует в микрофибриллах первичной стенки, то есть определить, чем целлюлоза, которая не определяется как «кристаллическая», отличается от кристаллической целлюлозы.Многие эксперименты, ведущие к созданию структуры древесной целлюлозы, зависели от исключительно однородной ориентации микрофибрилл целлюлозы (Sturcová et al., 2004; Fernandes et al., 2011). Однако в растущих клеточных стенках микрофибриллы ориентированы неравномерно. Когда микрофибриллы сначала откладываются на внутренней поверхности первичной клеточной стенки, их ориентация обычно перпендикулярна направлению роста, но когда клеточная стенка расширяется, микрофибриллы переориентируются таким образом, что распределение ориентации интегрируется по толщине расширенной клеточной стенки. клеточная стенка становится все более близкой к случайной (Cosgrove, 2005; MacKinnon et al., 2006).
Эта техническая проблема не относится к клеточным стенкам колленхимы сельдерея ( Apium graveolens ), которые похожи по составу на другие первичные клеточные стенки, но имеют их микрофибриллы, ориентированные относительно равномерно вдоль оси клетки (Sturcová et al., 2004 ; Kennedy et al., 2007a, 2007b). Некоторая структурная информация о целлюлозе колленхимы сельдерея уже была получена в результате спектроскопических экспериментов и экспериментов по рассеянию (Sturcová et al., 2004; Kennedy et al., 2007a, 2007b), подтверждая беспорядок, ожидаемый в целлюлозе первичной стенки.Некоторые из этих экспериментов были аналогичны тому, что было сделано с еловой целлюлозой (Fernandes et al., 2011), но недостаточно данных, чтобы указать количество цепей в каждой микрофибрилле первичной стенки, природу и расположение нарушения, а также наличие или отсутствие прямого контакта микрофибрилл. Здесь мы сообщаем об экспериментах по рассеянию рентгеновских лучей и нейтронов, а также спектроскопических экспериментах, направленных на решение этих вопросов и приводящих к предложенной структуре микрофибрилл целлюлозы первичной стенки. Характеристика структуры, содержащей столько беспорядка, представляла необычные проблемы, но беспорядок, по-видимому, является центральным для загадочной способности целлюлозы первичной стенки обеспечивать высокую прочность, но при этом позволять и контролировать рост.
РЕЗУЛЬТАТЫ
Диаметр микрофибриллы и расстояние от малоуглового рассеяния нейтронов
Когда микрофибриллы коллагена или целлюлозы фиксированного диаметра упакованы в массивы с некоторой степенью регулярности, может наблюдаться малоугловая дифракция рентгеновских лучей или нейтронов (брэгговское рассеяние) от самих микрофибрилл (Hulmes et al., 1995; Fernandes et al. др., 2011). Это рассеяние происходит под меньшими углами, чем обычная дифракция на кристаллических плоскостях, и накладывается на малоугловое рассеяние, возникающее не из-за дифракции, а из-за текстуры и формы фибрилл и пустот между ними (форм-фактор).Если упаковка не является достаточно регулярной, чтобы образовать кристаллическую сверхрешетку, наблюдается только один пик Брэгга, соответствующий расстоянию между центрами фибрилл (Hulmes et al., 1995; Kennedy et al., 2007a).
Используя малоугловое рассеяние рентгеновских лучей (SAXS), наблюдали брэгговское отражение от микрофибрилл целлюлозы внутри интактных клеточных стенок колленхимы сельдерея. Расстояние между микрофибриллами увеличивается при гидратации (Kennedy et al., 2007b). Моделирование этих результатов показало, что наблюдаемый пик Брэгга может быть вызван рядом комбинаций расстояния между микрофибриллами и их диаметра.Следовательно, невозможно однозначно определить диаметр микрофибрилл (Kennedy et al., 2007a).
В древесине ели микрофибриллы слишком тесно агрегированы, чтобы обеспечить адекватный контраст для малоуглового брэгговского рассеяния. Эту проблему удалось решить путем дейтерирования поверхности и использования нейтронов вместо рентгеновских лучей (Fernandes et al., 2011). Природа контраста различается между источниками рентгеновского излучения и нейтронов. Рентгеновский контраст зависит в основном от локальной электронной плотности, тогда как контраст рассеяния нейтронов зависит от локального элементного состава и плотности.Одним из преимуществ этого метода является очень разные сечения рассеяния нейтронов водородом и дейтерием, что позволяет ввести контраст, например, за счет дейтериевого обмена в оксиде дейтерия (D 2 O; Horikawa et al., 2009; Matthews и др., 2011).
Экваториальный пик Брэгга был хорошо виден на изображениях малоуглового рассеяния нейтронов (МУРН) древесины ели, насыщенной D 2 O (Fernandes et al., 2011). Интенсивность пика Брэгга уменьшалась, а его расстояние ( q ) от центра дифракционной картины увеличивалось при сушке до тех пор, пока на картине SANS от полностью сухой дейтерированной древесины пик не был слабо виден при q = 2.1 нм −1 . Низкий контраст в этих условиях соответствовал контакту между соседними дейтерированными на поверхности микрофибриллами, так что расстояние между центрами 3,0 нм соответствовало по существу среднему диаметру микрофибрилл (Fernandes et al., 2011).
Подобные эксперименты SANS были проведены на стенках клеток из колленхимы сельдерея и на целлюлозе, выделенной из этих клеточных стенок (рис. 1). Пик брэгговского рассеяния в области q = 1 нм -1 , наложенный на экспоненциальный фон от некогерентного рассеяния, был очевиден как для стенок клеток, так и для изолированной целлюлозы, насыщенной чистым D 2 O, чистой водой или ряд смесей (рис.1; Дополнительный рис. S1). В соответствии с рассеянием от целлюлозы, а не от других полимеров, не наблюдалось пика Брэгга для смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Которая соответствует теоретической плотности длины рассеяния. целлюлозы (рис. 1).
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна.C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл. D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив.Вертикальный масштаб больше, чем в C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 1.
SANS из клеточных стенок колленхимы сельдерея и целлюлозы. A, шаблон SANS для сухих стенок ячеек. B, шаблон SANS для сухой изолированной целлюлозы. В A и B ось волокна вертикальна. C, экваториальные профили рассеяния от клеточных стенок и изолированной целлюлозы. Подгоняемая линия представляет собой сумму экспоненциальной функции, соответствующей некогерентному рассеянию, и гауссовой функции, соответствующей экваториальному пику Брэгга, возникающему в результате дифракции от упаковки микрофибрилл.D, профили экваториального рассеяния от целлюлозы, насыщенной водой, D 2 O или смеси 35% [D 2 O] / [D 2 O + вода] (об. / Об.), Соответствующей целлюлозе, напротив. Вертикальная шкала больше, чем C в 150 раз. [Цветную версию этого рисунка см. В онлайн-статье]
В отличие от древесины ели, сухая целлюлоза сельдерея дает слабый пик Брэгга без дейтерирования поверхности микрофибрилл. В сухом состоянии пик Брэгга был при q = 2.11 нм -1 для целлюлозы в клеточных стенках или q = 2,14 нм -1 для изолированной целлюлозы (рис. 1), что соответствует межцентровым расстояниям в реальном пространстве 3,0 и 2,9 нм, соответственно. . Интенсивность Брэгга была менее 1% от наблюдаемой после регидратации. Этот очень низкий контраст соответствовал плотной упаковке микрофибрилл, вероятно, без каких-либо других полимеров между ними по крайней мере на некоторой части их длины. Расстояние от центра до центра должно приблизительно соответствовать среднему диаметру микрофибриллы.Несмотря на свою низкую интенсивность, пик Брэгга был легко различим на картинах МУРН, поскольку было очень мало помех от некогерентного рассеяния (рис. 1).
В гидратированном состоянии значение q пика Брэгга от микрофибрилл in situ в стенках клеток будет соответствовать реальному расстоянию ( d ) 7,1 ± 1,3 нм -1 , в то время как q Значение пика Брэгга для гидратированной целлюлозы соответствует реальному интервалу 6.0 ± 1,5 нм -1 . Эти номинальные расстояния d не означают, что все микрофибриллы разделены до такой степени при гидратации. Гидратация также значительно увеличивала контраст SANS за счет введения воды или D 2 O между микрофибриллами и, в случае D 2 O, за счет дейтерирования их поверхностей и нецеллюлозных полимеров. Отсюда следует, что при любом распределении расстояний между микрофибриллами более широкие промежутки будут значительно перепредставлены в шаблоне SANS. Номинальные интервалы, наблюдаемые после гидратации, немного превышали расчетные (Kennedy et al., 2007b) методом SAXS, что согласуется с разной основой рентгеновского контраста, но не было никакой разницы между расстояниями, наблюдаемыми в сухом состоянии с рентгеновскими лучами (Kennedy et al., 2007a) и с нейтронами (рис. 1). . Хотя на основе рентгеновских данных невозможно было сделать вывод о том, что расстояние, наблюдаемое в сухом состоянии, соответствует микрофибриллам, находящимся в прямом контакте друг с другом, этот вывод кажется вероятным на основе объединенных данных SAXS и SANS. . Гидратация четко разделяла микрофибриллы, но, вероятно, в разной степени по их длине (Kennedy et al., 2007b), возможно, в зависимости от сродства нецеллюлозных полисахаридов к воде (Jarvis, 1992).
Размеры микрофибрилл и нарушения в результате широкоугольного рассеяния рентгеновских лучей
Широкоугольные картины рассеяния рентгеновских лучей или нейтронов от природной целлюлозы достаточно хорошо разрешены для кристаллографического решения структуры, только если целлюлоза исключительно упорядочена и хорошо ориентирована и если микрофибриллы достаточно толстые, чтобы минимизировать уширение Шеррера, которое возникает в результате ограниченное количество плоскостей решетки.Большая часть целлюлозы колленхимы сельдерея была относительно хорошо ориентирована внутри клеточных стенок (рис. 2). Вклад широкоугольного рентгеновского рассеяния (WAXS) от незначительной изотропно ориентированной фракции целлюлозы был удален на стадии вычитания фона. Существенный вклад изотропного рассеяния на связанной воде также был удален при вычитании фона. Рассеяние от воды было идентифицировано экспериментом, в котором менялось содержание воды (дополнительный рис. S2).
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 2.
A, WAXS-изображение клеточных стенок колленхимы сельдерея. Ось волокна вертикальна. B — Экваториальный профиль WAXS с интенсивностью рассеяния, с симметричным подобранным компонентом и компонентом, обеспечивающим асимметрию. C, отражение 400, масштаб по вертикали увеличен в 50 раз. D, График δ q против q 2 d для основных экваториальных отражений. Пересечение линии, соединяющей отражения 200 и 400, использовалось для расчета размера Шеррера, перпендикулярного плоскости кристалла [200], а равный наклон (пунктирная линия) использовался для получения приблизительного среднего отрезка для 1-10 и 110 размышления.[См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок WAXS из хорошо ориентированного целлюлозного компонента был размыт в радиальном направлении из-за уширения Шеррера, уширения, связанного с беспорядком, или обоих. Отражение 200 было при q = 15,5 нм -1 , что соответствует несколько большему расстоянию между слоями, чем в кристаллических целлюлозах, и отражения 1-10 и 110 были объединены. Эти особенности наблюдались на плохо ориентированных дифрактограммах от того, что было названо целлюлозой IV 1 (Dinand et al., 1996) или неупорядоченную целлюлозу Iβ (Wada et al., 2004) в первичных клеточных стенках. Для единообразия в этой статье используется обозначение решетки Iβ, но без всякого указания на то, что эта форма представляет собой наиболее близкую аналогию со средней структурой.
Количественная оценка беспорядка и вычисление размеров Шеррера по уширению этих отражений было затруднено из-за сильного перекрытия между отражениями 1-10 и 110 и чрезвычайно низкой интенсивности отражения 400, качественного индикатора большего бокового беспорядка, чем в хвойная древесина.При длительном времени сбора данных (16–24 ч) можно было получить достаточное отношение сигнал / шум для оценки уширения отражения 400. Гидратация за счет увеличения моноклинного угла и, таким образом, разделения отражений 1-10 и 110 упростила моделирование соответствующей ширины этих двух отражений в дифракционной картине от древесины ели (Fernandes et al., 2011). Однако в целлюлозе сельдерея гидратация немного увеличивала интервал [200] d , но не увеличивала значительно угол моноклинности.Процедура Fernandes et al. (2011) был использован для выделения эффекта одной формы бокового беспорядка, которая приводит к асимметрии радиальных пиков. Предполагая, что оставшиеся симметричные радиальные ширины отражений 1-10 и 110 были равны, наиболее подходящее значение было на 22% больше, чем радиальная ширина отражения 200 при моноклинном угле 95 °. Исходя из этого, средние размеры элементарной ячейки в сухом состоянии составляли a = 0,81 нм, b = 0,80 нм, c = 1,03 нм (с сохранением индекса целлюлозы Iβ для ясности, даже если а> b).Меньшие углы моноклинности и большая ширина, или наоборот, почти так же точно соответствовали наблюдаемым профилям интенсивности.
Еще одна форма беспорядка, которая, наряду с небольшими размерами решетки, способствует радиальному уширению, была смоделирована как «паракристаллическая» (Fernandes et al., 2011), что означает, что ширина отражений увеличивается с квадратом порядка отражения. или с q 2 d . Используя наиболее подходящие данные и построив график ширины δ q отражений против q 2 d , как описано (Fernandes et al., 2011), ширина симметричных компонент рефлексов 200 и 400 давала фактор остаточной разупорядоченности г = 0,03 и размер Шеррера 3,2 нм, перпендикулярный плоскостям колец в сухом состоянии. Средняя ширина отражений 1-10 и 110 лежит явно выше δ q по сравнению с графиком q 2 d для отражений 200 и 400. Подразумевается, что средний размер Шеррера в направлениях, нормальных к плоскостям решетки 1-10 и 110 был меньше, чем средний размер Шеррера в направлении, нормальном к плоскости решетки 200, как в еловой целлюлозе.Однако низкая интенсивность отражения 400 и, как следствие, сложность оценки вклада, связанного с беспорядком, в общее уширение сделали этот вывод менее надежным, чем для еловой целлюлозы.
Характеристика неупорядоченных компонентов с помощью дейтерирования и широкоугольного рассеяния нейтронов
В кристаллографическом исследовании целлюлозы Iα и Iβ (Nishiyama et al., 2002, 2003) разница в дифракции нейтронов, обусловленная полным высокотемпературным дейтерированием гидроксильных групп, использовалась для определения положений протонов и, следовательно, геометрии водородных связей. .Также должна быть возможность использовать сильное различие между рассеянием нейтронов дейтерием и протонами для определения вклада в рассеяние неупорядоченных областей, доступных для дейтерирования в более мягких условиях температуры окружающей среды, используемых в экспериментах с SANS и инфракрасным преобразованием Фурье (FTIR). Как может быть показано с помощью FTIR (см. Ниже), гидроксильные группы на нецеллюлозных полисахаридах доступны для дейтерирования, как и обращенные наружу гидроксилы на поверхности целлюлозы, если они не заблокированы агрегацией (Horikawa et al., 2009). Для неупорядоченных областей внутри микрофибрилл, чтобы допустить D 2 O, они должны иметь более широкие интервалы решетки, чем кристаллическая целлюлоза (Matthews et al., 2011).
На рис. 3А показано, что наблюдались видимые изменения в картине дифракции нейтронов при обратимом дейтерировании при температуре окружающей среды. Изменения включали увеличение или уменьшение интенсивности отдельных отражений, а также изменения диффузного рассеяния. В принципе, изменение интенсивности для любого отражения может быть положительным, отрицательным или нейтральным в зависимости от влияния дейтерирования на структурный фактор.В качестве руководства, интенсивности основных экваториальных и осевых отражений для целлюлозы Iβ в нативной (H) и пердейтерированной (D) формах были рассчитаны с использованием Shelxl-97 (Sheldrick, 2008) в пакете WinGX (Farrugia, 1999), из статическая модель, использующая опубликованные координаты атомов (Nishiyama et al., 2002). Из трех основных экваториальных отражений, определяющих геометрию боковой упаковки целлюлозных цепей, влияние дейтерирования на интенсивность было предсказано положительным для 200 и сильно отрицательным для 110 и 1-10.Было предсказано, что осевое отражение 004 будет увеличиваться по интенсивности при дейтерировании, в то время как отражение 001 отсутствовало как в D, так и в H-формах. Эти прогнозы согласуются с наблюдаемыми нейтронографическими изображениями для высококристаллической целлюлозы Iβ (Nishiyama et al., 2002).
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина). Ось волокна вертикальна.B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Рисунок 3.
WANS из клеточных стенок колленхимы сельдерея. A, диаграммы рассеяния в H-форме (правая половина) и после дейтериевого обмена гидроксильных групп, доступных для воды (левая половина).Ось волокна вертикальна. B, разностная диаграмма рассеяния (D — H). C, экваториальные профили рассеяния от стенок ячеек в форме H, форме D и разности (D — H). D: профили рассеяния на оси волокна от стенок ячеек в форме H, форме D и разности (D — H). E: азимутальные профили рассеяния на отражении 200. [См. Онлайн-статью для цветной версии этого рисунка.]
Наблюдаемый эффект поверхностного дейтерирования микрофибрилл был положительным для отражений 200 и 004, отрицательным для 110 и нейтральным для 1-10.Профиль вычитания (D — H) для экваториальных отражений (рис. 3B) показал, что их положение и ширина, соответствующие цепям, доступным для дейтерирования, не заметно отличались от фракции, которая не подвергалась дейтерированию. В азимутальном направлении экваториальная интенсивность показывала более широкий нижний хвост распределения ориентации в D, чем в H-дифракционной картине (рис. 3E), подразумевая, что некоторые из цепей, доступных для дейтерирования, были менее хорошо ориентированы, чем недоступные цепочки целлюлозы.
На оси волокна (рис. 3D) отражение 004 в D-форме было центрировано при немного меньшем q , чем в H-форме, что означает, что некоторые из цепочек, доступных для дейтерирования, были более протяженными, чем недоступные цепи. Разница в среднем расстоянии d составляла всего около 0,5%, а радиальная ширина этого отражения была одинаковой как для D, так и для H-формы. Отражение 002 было сильнее в D-форме, а отражение 001, отсутствующее в H-дифракционной картине, появилось в профиле разности осей.Наличие этого отражения 001 после дейтерирования означает, что некоторые из дейтерированных цепей не имели такого же продольного смещения, как чистая целлюлоза Iβ или целлюлоза Iα.
Почти изотропные изменения диффузного рассеяния при дейтерировании привели к появлению широкого кольца повышенной интенсивности в области 15 нм -1 < q <20 нм -1 , а ниже 15 нм -1 , была уменьшена диффузная интенсивность (рис. 3В). Изотропные компоненты, ответственные за эти особенности диффузного рассеяния, включают нецеллюлозные компоненты, которые обмениваются с D 2 O и небольшими количествами остаточной связанной воды.
Эксперименты по спиновой диффузии протонов и пространственные отношения между упорядоченными и неупорядоченными цепями
Скорость, с которой ядерный магнетизм уравновешивается за счет спиновой диффузии между наноразмерными доменами, может использоваться для оценки расстояния между этими доменами. Эксперименты по спиновой диффузии протонов, основанные на этом принципе, были применены к древесине (Newman, 1992; Altaner et al., 2006) и бактериальной целлюлозе (Masuda et al., 2003). Использовалось двумерное (2D) представление этого эксперимента (Fernandes et al., 2011), чтобы изучить расположение упорядоченных и неупорядоченных областей в еловой целлюлозе, используя различия в их пространственном отделении от лигнина. Совершенно другой эксперимент по спин-диффузии 13 C на соломе кукурузы ( Zea mays ) (Foston et al., 2012) привел к аналогичным выводам.
Целлюлоза колленхимы сельдерея не лигнифицирована, но содержит небольшие остаточные количества гемицеллюлоз глюкоманнана и ксилоглюкана, которые, обладая относительно высокой термической подвижностью, могут использоваться вместо лигнина в качестве резервуаров спина протонов.На рисунке 4 показан 2D-спектр этого эксперимента. Перекрестные пики указывают на домены, которые удовлетворяют двум условиям: (1) домены различаются по подвижности и, следовательно, по намагниченности в начале периода смешения спинов протонов в последовательности импульсов; и (2) домены достаточно разделены в пространстве, чтобы потребовалось измеримое время, несколько миллисекунд, для уравновешивания намагниченности протонов посредством спиновой диффузии. Приведенное ниже назначение сигналов основано на Sturcová et al. (2004).
Рисунок 4.
2D-представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, отслеживаемых по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены. [См. Онлайн-статью для цветной версии этого рисунка.]
Рис. 4.
Двумерное представление твердотельного эксперимента по спин-диффузии 1 H на гидратированных клеточных стенках колленхимы сельдерея, мониторинг которого проводился по спектру 13 C. Спектр 13 C, измеренный с кросс-поляризацией и вращением под магическим углом, показан сверху и сбоку. Перекрестные пики в 2D-спектре представляют собой сигналы от ядер 13 C, которые расположены в доменах, которые различаются скоростью спин-спиновой релаксации 1 H и пространственно разделены.[Цветную версию этого рисунка см. В онлайн-статье.]
Одномерные спектры C-ЯМР 13 были аналогичны тем, которые наблюдались ранее (Sturcová et al., 2004), с сигналами C-4 от кристаллической целлюлозы (89 –90 частей на миллион [частей на миллион]) и неупорядоченной целлюлозы (84–85 частей на миллион) в примерном соотношении 1: 2. В 2D-спектре присутствовали кросс-пики между C-4 кристаллической целлюлозы и C-4 неупорядоченной целлюлозы, но они были относительно слабыми. Также присутствовал кросс-пик между C-6 кристаллической целлюлозы (65 ppm) и C-4 неупорядоченной целлюлозы (84–85 ppm).Перекрестные пики от гемицеллюлозы C-1 (101 ppm) и C-4 (83 ppm) до кристаллической целлюлозы C-4 были сильнее, чем от неупорядоченной целлюлозы C-4, и перекрестные пики от гемицеллюлозы C-1 и C-4 к кристаллической целлюлозе С-6 были еще сильнее. Наиболее сильные кросс-пики связаны с сигналами C-2, C-5 и C-6 от неупорядоченной целлюлозы при 73, 75 и 62 м.д., но каждый из этих сигналов, очевидно, также содержит вклад гемицеллюлоз и, следовательно, не может быть определен. однозначно трактуется.
Эти наблюдения согласуются с моделями, в которых неупорядоченная и кристаллическая целлюлоза не смешиваются, а находятся в непосредственной пространственной близости внутри каждой микрофибриллы, а гемицеллюлозы расположены вне микрофибрилл, вероятно, частично за пределами каждого небольшого пучка микрофибрилл, но разница в кросс-пике интенсивность кристаллической и неупорядоченной целлюлозы предполагает, что одна группа гемицеллюлоз была в тесной связи с поверхностями микрофибрилл.
Поляризованный дейтерированный FTIR и модели водородной связи
Поляризованный FTIR предоставляет информацию о направлении водородных связей. Исследования FTIR на древесине ели показали, что гидроксильные группы на неупорядоченной целлюлозе на поверхности микрофибрилл были доступны для дейтериевого обмена, за исключением тех случаев, когда микрофибриллы были плотно агрегированы вместе. Также были доступны гидроксильные группы на гемицеллюлозах. Обмен дейтерия сдвигает полосу растяжения, связанную с каждой гидроксильной группой, на более низкую частоту в 1 раз.34, из-за большей массы атома дейтерия, но сохраняет порядок полос, потому что структура не изменилась (Sturcová et al., 2003).
На фиг. 5 показано, что дейтерирование либо клеточных стенок колленхимы сельдерея, либо изолированной целлюлозы было обширным, но меньше, чем количество, предсказанное из соотношения 2: 1 неупорядоченной целлюлозы к кристаллической, оцененного с использованием сигналов C-4 в спектрах ЯМР. Образец полос растяжения O-D легче всего сравнить с рисунком полос растяжения O-H, если шкала частот искусственно сдвинута в 1 раз.34 (рис. 5). Эти две модели явно отличались. Полосы растяжения O-D были менее ориентированы в продольном направлении, чем полосы растяжения O-H, показывая, что меньшее количество водородных связей в дейтерированной фракции было направлено вдоль оси микрофибрилл, а больше — наружу. Как и в еловой целлюлозе (Fernandes et al., 2011), дейтерирование снизило интенсивность выступающего плеча на 3400 см -1 в области растяжения O-H, что соответствует гидроксильным группам, более слабо связанным водородными связями и более направленным наружу, чем в среднем.После сдвига оси частот на на 1,34 раза максимум довольно широкой группы перекрывающихся полос валентных колебаний OD совпал с этим плечом, которое первоначально было отнесено к аномальной части Iα-подобного спектра целлюлозы (Sturcová et al., 2004), но позже было показано, что в еловой целлюлозе они соответствуют недоступным гидроксильным группам в неупорядоченных областях (Fernandes et al., 2011). Отнесение к неупорядоченным доменам, а не к целлюлозе Iα, подтверждается данными, представленными здесь.Только часть интенсивности на 3 400 см -1 была смещена дейтерированием, подразумевая, что некоторые из этих неупорядоченных цепей в целлюлозе колленхимы сельдерея доступны для дейтерирования, а некоторые — нет.
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой. Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию.Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В онлайн-статье.]
Рисунок 5.
Поляризованные FTIR-спектры стенок клеток колленхимы сельдерея и изолированной целлюлозы, подвергнутых парофазному дейтериевому обмену и высушенных без контакта с водой.Сплошные линии показывают продольную поляризацию, а пунктирные линии показывают поперечную поляризацию. Для сравнения с областью растяжения O-H (3,000–3,500 см, –1 ), область растяжения O-D (2,200–2,700 см, –1 ) также показана со шкалой длин волн, увеличенной в 1,34 раза. Спектры разности (H — D) показаны ниже. [Цветную версию этого рисунка см. В статье в Интернете.]
ОБСУЖДЕНИЕ
Структура из микрофибриллы
Целлюлоза, которую можно условно назвать кристаллической, и неупорядоченная целлюлоза, отличная от известных кристаллических форм целлюлозы Iα и Iβ, были идентифицированы всеми использованными методами.Из этого не следует, что одни и те же неупорядоченные фракции наблюдались всеми этими методами (т.е. что цепи, идентифицированные как неупорядоченные, в одних и тех же отношениях отличались от известных кристаллических форм). Наблюдались конформационный беспорядок, беспорядок упаковки и беспорядок водородных связей.
Спектры 13 C-ЯМР целлюлозы колленхимы сельдерея (рис. 4) идентифицировали цепи, которые отличались по конформации как от целлюлозы Iα, так и от целлюлозы Iβ, имеющие некоторую свободу вращения вокруг связи C5-C6 и имеющие C-6 В конформации преобладают гош-гош ( gg ) и гош-транс ( gt ) формы (Sturcová et al., 2004), как и в еловой целлюлозе (Fernandes et al., 2011), хотя соотношение gg — gt значительно отличалось от еловой (Sturcová et al., 2004). Спектры дейтерирования FTIR целлюлозы сельдерея показали, что некоторые домены имели паттерны водородных связей, отличные от наблюдаемых в целлюлозе Iα и целлюлозе Iβ, с пониженной средней прочностью водородных связей (более высокая средняя частота растяжения O-H) и меньшей общей тенденцией к осевой ориентации водорода. Изменение ориентации водородных связей было очевидным как в цепях, доступных для дейтериевого обмена, так и в недоступных цепях, и соответствовало переходу от транс-гош ( tg ) конформации C-6, обнаруженной в кристаллической целлюлозе, к gg . и конформеры GT .Подобные наблюдения на еловой целлюлозе были рационализированы с точки зрения модели, в которой ядро каждой микрофибриллы было занято в основном кристаллической целлюлозой, а поверхность микрофибрилл была занята цепями целлюлозы с несколько иной конформацией и водородными связями, некоторые из которых были доступны для обмена дейтерия, а другие были представлены. недоступны из-за боковой агрегации микрофибрилл. Подобная модель согласуется с FTIR-спектрами сельдерея (рис. 5), более ранними данными ЯМР (Sturcová et al., 2004), а также данные спин-диффузионного ЯМР, представленные здесь (рис. 4). По данным ЯМР спин-решеточной релаксации 13 C (Viëtor et al., 2002), целлюлоза колленхимы сельдерея имеет более высокую долю конформера C-6 gg, чем еловая целлюлоза. Размеры элементарной ячейки значительно различались между этими двумя видами целлюлозы, среднее расстояние между листами было больше в целлюлозе колленхимы сельдерея, а осевой размер был немного меньше. Молекулярное моделирование предложило связь между конформацией C-6 и шагом решетки (Matthews et al., 2012). Как и в случае с еловой целлюлозой, было невозможно и, возможно, бессмысленно различать формы целлюлозы Iα и Iβ с помощью ЯМР при наличии такого большого конформационного нарушения (Matthews et al., 2011).
Модели микрофибрилл с неупорядоченными поверхностями неоднократно предлагались на основе данных ЯМР (Newman, 1992; Viëtor et al., 2002; Bergenstråhle et al., 2008), но было неясно, как они должны согласовываться с полученными моделями. от дифракции рентгеновских лучей, которая более чувствительна к структуре упаковки цепей, чем к конформации цепи или водородным связям.Данные широкоугольного рассеяния нейтронов (WANS) (рис.3) показали, что в доменах, доступных для дейтерирования, цепи были упакованы латерально таким же образом и почти с такими же средними расстояниями, как и в кристаллических областях, несмотря на различия в конформации. и водородные связи, продемонстрированные спектроскопическими экспериментами, несмотря на значительные статистические различия в упаковке цепей повсюду в микрофибрилле. Повсюду в каждой микрофибрилле листы цепочек были уложены друг на друга с более широким средним интервалом, чем в более кристаллических целлюлозах.Дальнейшее следствие состоит в том, что большинство цепей, доступных для дейтерирования, были расположены на поверхностях микрофибрилл, а не в «аморфных» сегментах микрофибриллы, занимающих всю ширину микрофибриллы, потому что цепи внутри таких сегментов должны быть неплотно упакованы, чтобы вода могла стекать. входить.
Судя по осевому отражению 004, доступные для дейтерирования цепи оказались немного более вытянутыми в продольном направлении, чем недоступные цепи, возможно, из-за скручивания микрофибрилл.Продольное смещение было частично нарушено в цепочках, доступных для дейтерирования, что усложняло различение форм Iα и Iβ.
Широкий диапазон расстояний между боковыми цепями в нецеллюлозной фракции, доступный для дейтерирования, может быть выведен из диффузной, неориентированной части нейтронографических изображений. Вода, связанная с нецеллюлозной фракцией, рассеивала рентгеновские лучи аналогично жидкой воде, предполагая аналогичную функцию распределения пар, но из-за пониженной тепловой подвижности величина ее вклада в структуру WAXS была больше, чем ожидалось для объемной жидкой воды.
Диаметр и форма микрофибриллы
Данные SANS предполагают диаметр микрофибрилл, приблизительно равный межцентровому расстоянию от 2,9 до 3,0 нм в сухом состоянии. Предполагая круглое поперечное сечение и учитывая относительно рыхлую упаковку цепей, наблюдаемую при широкоугольном рассеянии, этот номинальный диаметр будет соответствовать 21–22 цепям. Микрофибриллы, конечно, не имеют круглой формы, и фактическое количество цепочек зависит от формы поперечного сечения.Наблюдения WANS подтвердили, что поверхностные цепочки располагались на той же неупорядоченной решетке, что и внутренние цепочки, и должны быть включены в расчеты размеров Шеррера из WAXS.
Межцентровое расстояние SANS и поперечные размеры по Шерреру были аналогичны тем, которые наблюдались для еловой целлюлозы (Fernandes et al., 2011). Размер [200] Шеррера был больше, чем среднее значение размеров [1-10] и [110], что согласуется с моделью прямоугольной микрофибриллы (Matthews et al., 2006; Fernandes et al., 2011), который имеет относительно гидрофобные [200] грани кристаллов, открытые сверху и снизу (рис. 6), вместо диагональных [1-10] и [110] граней кристаллов, открытых в целлюлозах водорослей (Baker et al. ., 1997). Однако опора для прямоугольной, а не ромбовидной модели менее надежна, чем для ели, из-за сложности коррекции беспорядка с использованием очень слабого отражения 400 и из-за проблем, присущих оценке отдельных ширины перекрывающихся 1-10 и 110 отражений.Учитывая наблюдаемое обширное нарушение, нельзя даже с уверенностью сказать, что микрофибриллы первичной стенки имеют постоянную форму поперечного сечения.
Рисунок 6.
Модели поперечного сечения микрофибрилл колленхимы сельдерея. A, 24-цепочечная ромбовидная модель, 2,9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться.Расчетные длины столбцов [200], [1-10] и [110] были следующими, соответственно: модель A, 2,5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В онлайн-статье]
Рисунок 6.
Модели для поперечного сечения микрофибрилл колленхимы сельдерея. A, модель ромбовидной формы с 24 цепями, 2.9 × 2,7 нм. B, 24-цепочечная прямоугольная модель, 3,3 × 2,7 нм. C, прямоугольная модель из 18 цепочек, 2,5 × 2,7 нм. Длины столбцов WAXS (размеры по Шерреру) после коррекции беспорядка позволяют различать эти модели с осторожностью из-за сложности оценки ширины перекрывающихся отражений 1-10 и 110, аппроксимаций, участвующих в коррекции беспорядка, и возможность того, что поперечные сечения микрофибрилл могут различаться. Расчетные длины столбцов [200], [1-10] и [110] были соответственно следующими: модель A, 2.5, 2,8 и 2,5 нм; модель B, 3,2, 2,6 и 2,5 нм; модель C, 2,4, 2,5 и 2,3 нм. Наблюдаемые длины колонок были следующими: [200], 3,2 нм; [1-10] и [110], среднее значение 2,5 нм. Исходя из этого, наиболее подходящей моделью была модель B. [Цветную версию этого рисунка см. В статье в Интернете.]
Прямоугольное поперечное сечение с уложенными друг на друга листами из трех цепей будет соответствовать одной трети цепей, находящихся в центральное положение в каждом листе, конформационно сходное с кристаллическими целлюлозами, как наблюдалось с помощью ЯМР.Наблюдение единственного малоуглового пика Брэгга согласуется со слегка диспергированными расстояниями между микрофибриллами, которые можно было бы ожидать от примерно прямоугольных микрофибрилл, возможно, скрученных, находящихся в прерывистом контакте друг с другом. Могут быть приспособлены как постоянные, так и слегка изменяющиеся диаметры микрофибрилл, а также небольшое количество микрофибрилл, значительно отличающихся от доминирующих размеров.
Число цепей на микрофибриллу имеет отношение к структуре комплексов, которые синтезируют целлюлозу во время роста.Если количество цепей кратно шести, что соответствует геометрии синтетического комплекса, тогда стопка из восьми трехцепочечных листов (24 цепи) даст наиболее близкое совпадение с размером Шеррера 200 (рис. 6B), как и для еловая целлюлоза (Fernandes et al., 2011). Однако в нашем случае неопределенность довольно велика, и уровень беспорядка упаковки предполагает, что возможны нерегулярные и переменные сечения. Модель из 18 цепей (6 × 3) не исключена для еловой целлюлозы и согласуется с данными, представленными здесь (рис.6C), если размер 200 по Шерреру был увеличен дополнительными цепями гемицеллюлозы, как это было предложено для ели (Fernandes et al., 2011), или увеличен путем наложения одной микрофибриллы поверх другой на короткие расстояния.
Кристалличность
Что означает «кристалличность», зависит от методов, используемых для обнаружения компонентов, которые отличаются от кристаллической целлюлозы. Между спектроскопическими методами обнаружения цепей, отличающихся по конформации от общепринятых форм кристаллической целлюлозы и, следовательно, демонстрирующих различные модели водородных связей, наблюдалась некоторая степень согласованности.Это включало нецеллюлозные полисахариды, и, в частности, в спектрах FTIR было трудно провести четкое различие между вкладом нецеллюлозных полисахаридов и некоторой неупорядоченной целлюлозы.
Эксперименты с дейтерированием WANS показали, что эти конформационные различия мало влияют на упаковку целлюлозных цепей или дифракцию. Структура микрофибрилл явно содержала значительный беспорядок в боковой упаковке цепей, но этот беспорядок распространялся по всей структуре и не ограничивался доступными для дейтерия поверхностями.Кристалличность, оцениваемая дифракционными методами, — это не то же самое, что кристалличность, оцениваемая спектроскопически. В обоих случаях кажущаяся кристалличность будет увеличиваться с увеличением диаметра микрофибрилл, но как прямое следствие уменьшения уширения по Шерреру в дифракционных экспериментах и из-за уменьшения отношения поверхности к объему в спектроскопических экспериментах, которые позволяют различать поверхностные цепи.
Агрегация микрофибрилл
Агрегация микрофибрилл в более крупные пучки хорошо известна для древесины хвойных пород (Wickholm et al., 1998; Фахлен и Сальмен, 2005 г .; Fernandes et al., 2011). Фибриллярные единицы шириной от 5 до 10 нм, иногда до 40 нм, наблюдались с помощью электронной микроскопии (McCann et al., 1990; Sugimoto et al., 2000), но их интерпретация осложняется возможным включением нецеллюлозных полимеров. . Представленные здесь данные включают две прямые линии доказательств агрегации микрофибрилл первичной стенки в колленхиме. Доказательства малоуглового брэгговского рассеяния рентгеновских лучей (Kennedy et al., 2007a) и нейтронов ясно показали, что агрегация существует. Диапазон расстояний между микрофибриллами было трудно сделать вывод, потому что интенсивность рассеяния сильно увеличивалась с разделением микрофибрилл, так что хорошо разделенные микрофибриллы были чрезмерно представлены на диаграммах рассеяния. Низкий контраст МУРН в сухом состоянии и сходство межцентрового расстояния МУРН со средними размерами Шеррера WAXS предполагают, что некоторые сегменты микрофибрилл оставались в прямом контакте друг с другом, тогда как другие сегменты микрофибрилл становились хорошо разделенными при гидратации.
Спектры дейтерирования FTIR как от стенок клеток колленхимы, так и от изолированной целлюлозы включали полосы растяжения гидроксила с центром на 3 400 см -1 с нейтральной поляризацией. Эти гидроксильные полосы растяжения были отнесены к гидроксильным группам, отличным от всех групп Iα и Iβ кристаллических форм целлюлозы, более ориентированным наружу по ориентации и частично, но не полностью доступным для дейтерирования. Принимая во внимание также данные SANS и ЯМР, соответствующие гидроксильные группы могут быть отнесены к поверхностям микрофибрилл.Поскольку эти поверхности были только частично доступны для D 2 O, отсюда следует, что микрофибриллы были агрегированы вместе на части своей длины с границами раздела, непроницаемыми для воды. Эти особенности были обнаружены в древесине ели, но были также различия: древесина ели имеет разные нецеллюлозные полимеры, что, по-видимому, приводит к более тесному связыванию между поверхностями микрофибрилл и к напряжениям, которые искажают геометрию элементарной ячейки при гидратации (Fernandes et al., 2011).
Полоса валентных колебаний гидроксила 3,400 см -1 в FTIR-спектре, перемещенная до 2,540 см -1 за счет обмена D 2 O, ранее была неправильно отнесена к аберрантной форме целлюлозы Iα (Sturcová et al., 2004), но, по-видимому, является маркером поверхностных цепей и наличия агрегации микрофибрилл, если эти поверхностные цепи недоступны для дейтерирования. Интенсивность этой полосы сильно уменьшилась, когда целлюлоза из колленхимы сельдерея была подвергнута гидротермальному отжигу, что привело к слиянию микрофибрилл в более крупные кристаллические единицы с уменьшенным соотношением поверхности к объему (Sturcová et al., 2004). Интенсивность FTIR на уровне 3400 см или около него -1 была зарегистрирована в препаратах клеточных стенок, например, арабидопсиса дикого типа ( Arabidopsis thaliana ; MacKinnon et al., 2006), предполагая, что эта форма агрегации не является уникальной особенностью целлюлозы колленхимы сельдерея, но более широко распространена в первичных клеточных стенках.
Влияние на рост растений
Размер и структура микрофибрилл первичной стенки дают важные ключи к разгадке природы ферментных комплексов, ответственных за ориентированный синтез целлюлозы и, следовательно, за направленную экспансию клеток. Размножение растительных клеток требует разрушения сети нековалентно сшитых микрофибрилл в клеточной стенке.Понимание топологии этой сети, следовательно, необходимо, и недавно принятая идея о том, что ксилоглюканы просто покрывают и связывают поверхности микрофибрилл, была поставлена под сомнение (Bootten et al., 2004; Park and Cosgrove, 2012a). Данные спин-диффузии ЯМР (рис. 4) показали, что некоторые цепи гемицеллюлозы, вероятно, ксилоглюканы, располагались близко к поверхностям микрофибрилл и, таким образом, вероятно, лежали между микрофибриллами внутри агрегатов микрофибрилл. Однако другие сегменты микрофибрилл, по-видимому, находятся в прямом контакте без промежуточных полимеров, демонстрируя нерегулярный характер агрегации, которого можно было бы ожидать, если микрофибриллы были скрученными и нецилиндрическими, и предполагая, что взаимодействия целлюлоза-целлюлоза, а также ксилоглюкан-целлюлоза должны быть разбиты на отдельные агрегированные микрофибриллы.Др. Механизмы необходимы для отделения одного пучка микрофибрилл от следующего во время роста путем разрушения ксилоглюкана или др. Мостиков между пучками микрофибрилл (Anderson et al., 2010). Агрегация микрофибрилл вдоль части их длины, в то время как другие части их длины остаются свободными для разделения или в них вставлены другие полисахариды, поднимает интересные вопросы о действии экспансинов (Cosgrove, 2005) и о том, как действует действие определенных глюканов. гидролазы взаимодействуют с таковой ксилоглюкан-специфической эндоглюканазы (Park and Cosgrove, 2012b).
Интересен высокий уровень беспорядка на границах раздела, наиболее важных для роста, между микрофибриллами и нецеллюлозными полимерами, а также между одной микрофибриллой и другой. Примеры прочных биологических материалов из царства животных показывают, что неупорядоченные домены могут вносить вклад в ударную вязкость (высокую энергию разрушения) при внешнем стрессе (van Beek et al., 2002). В первичных клеточных стенках устойчивость к разрушению должна сочетаться с контролируемой ферментативно-опосредованной продуктивностью, по-видимому, на неупорядоченных границах раздела, что обеспечивает рост.
Последствия разложения целлюлозы
Доступность микрофибрилл целлюлозы для целлюлаз является важным фактором превращения лигноцеллюлозы в биотопливо и еще более важна в случае нелгнифицированной биомассы. Нестабильность целлюлозы часто связывают с ее кристалличностью (Arantes and Saddler, 2010), а «декристаллизацию» иногда называют целью процессов предварительной обработки (Beckham et al., 2011), без полного согласия относительно того, что означают эти термины.Степень, в которой микрофибриллы целлюлозы имеют относительно гидрофобную [200] открытую поверхность, будет положительно влиять на начальную стадию деградации целлюлазами, которые специфически связываются с этой поверхностью (Dagel et al., 2011; Liu et al., 2011). Было бы интересно узнать больше о специфичности связывания оксидаз целлюлозы GH61 / CBM33 (Forsberg et al., 2011; Quinlan et al., 2011), некоторые из которых также связываются с одной и той же поверхностью (Li et al., 2012). Агрегация микрофибрилл целлюлозы — это фактор сопротивляемости, который заслуживает большего внимания, и теперь к нему можно подойти с помощью экспериментов по дейтерированию FTIR.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Было выделенонитей колленхимы из зрелых черешков сельдерея ( Apium graveolens ), клеточные стенки и их целлюлозная фракция были приготовлены, как описано (Sturcová et al., 2004), и хранились в сухом виде. Подробнее см. Дополнительные материалы и методы S1.
SANS
ЭкспериментыSANS были выполнены на канале D11 в Институте Лауэ-Ланжевена на клеточных стенках колленхимы сельдерея и изолированной целлюлозе.Подробнее об эксперименте см. Дополнительные материалы и методы S1.
WAXS
дифрактограммы рентгеновских лучей получали при температуре окружающей среды с использованием дифрактометра Rigaku R-axis / RAPID для рентгеновских пластин с источником излучения с молибденовой рентгеновской эмиссией Kα (λ = 0,07071 нм). Пучок коллимировался до диаметра 0,5 мм. Образцы имели толщину 0,7 мм в направлении, параллельном пучку, а их другие размеры превышали диаметр пучка. Дифрактограммы были собраны в режиме перпендикулярного пропускания, за исключением экспериментов с наклоном для измерения осевых отражений.Расстояние от центра дифракционной картины было выражено как q = 4πsinθ / λ, где 2θ — угол рассеяния. Для получения дополнительных сведений об экспериментальных методах и методах обработки данных (Fernandes et al., 2011) см. Дополнительные материалы и методы S1.
WANS
Дифракция нейтронов проводилась на канале D19 в Институте Лауэ-Ланжевена при средней длине волны нейтронов 0,242 нм и расстоянии от образца до детектора 0,756 м. Образцы размером 25 × 15 × 1 мм собирали из нескольких выровненных нитей колленхимы и выдерживали при контролируемом давлении воды или паров D 2 O в проточном контейнере со стенками из алюминиевой фольги.Подробнее см. Дополнительные материалы и методы S1.
ЯМР
A 1 H Эксперимент по спиновой диффузии Гольдмана-Шена с детектированием по спектру 13 C после кросс-поляризации был проведен на клеточных стенках колленхимы сельдерея, гидратированных 0,2 см. 3 г −1 D 2 O. Экспериментальные процедуры и анализ данных, включая 2D-представление данных, были такими, как описано (Fernandes et al., 2011).
Дейтерирование FTIR
Стенки клеток колленхимы сельдерея и изолированную целлюлозу помещали в проточную ячейку с окнами из фторида бария и заменяли воздухом, насыщенным О 2 О, до тех пор, пока спектры не перестали изменяться. Затем газовую линию переключали на азот, тщательно осушая, пропуская через сухое молекулярное сито 4А, а затем пентоксид фосфора (Sicapent; Aldrich). Спектры FTIR собирали с помощью спектрометра Thermo Nicolet Nexus, оборудованного приставкой для микроскопа Nicolet Continuum, имеющей охлаждаемый жидким азотом детектор теллурида кадмия ртути (Fernandes et al., 2011).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Rigaku, Ltd. за предоставленный инструмент и Fujiang Zhang за получение некоторых спектров FTIR.
Глоссарий
SAXS
Малоугловое рассеяние рентгеновских лучей
D 2 O
SANS
Малоугловое рассеяние нейтронов
WAXS 9407
широкоугольное рассеивание
WANS
широкоугольное рассеяние нейтронов
FTIR
Инфракрасное преобразование Фурье
2D
ppm
LITERATURE CITED
(
2006
)Пространственные отношения между полимерами в ели ситкинской: исследования спиновой диффузии протонов
.Holzforschung
60
:665
—673
(
2010
)Визуализация в реальном времени переориентации целлюлозы во время расширения клеточной стенки в корнях Arabidopsis
.Plant Physiol
152
:787
—796
(
2009
)Выяснение механизмов сборки и субъединичного взаимодействия целлюлозосинтазного комплекса вторичных клеточных стенок Arabidopsis
.J Biol Chem
284
:3833
—3841
(
1997
)Атомно-силовая микроскопия высокого разрешения микрокристаллов нативной целлюлозы I Valonia
.J Struct Biol
119
:129
—138
(
2011
)Происхождение стойкости биомассы на молекулярном уровне: свободные энергии декристаллизации для четырех обычных полиморфов целлюлозы
.J Phys Chem B
115
:4118
—4127
(
2008
)Динамика границ раздела целлюлоза-вода: времена спин-решеточной релаксации ЯМР, рассчитанные на основе атомистического компьютерного моделирования
.J Phys Chem B
112
:2590
—2595
(
2004
)Твердотельный 13 C-ЯМР-спектроскопия показывает, что ксилоглюканы в первичных клеточных стенках маша (Vigna radiata L.) происходят в разных доменах: новая модель взаимодействий ксилоглюкан-целлюлоза в клеточной стенке
.J Exp Bot
55
:571
—583
(
2009
)Растения контролируют свойства и работу своих органов посредством ориентации фибрилл целлюлозы в их клеточных стенках
.Интегр Комп Биол
49
:69
—79
(
2005
)Рост клеточной стенки растений
.Nat Rev Mol Cell Biol
6
:850
—861
(
2011
)Визуализация на месте отдельных углеводсвязывающих модулей на микрофибриллах целлюлозы
.J Phys Chem B
115
:635
—641
(
1996
)Паренхиматозная клеточная целлюлоза из жома сахарной свеклы: получение и свойства
.Целлюлоза
3
:183
—188
(
2005
)Распределение пор и матрицы в стенке волокна, выявленное с помощью атомно-силовой микроскопии и анализа изображений
.Биомакромолекулы
6
:433
—438
(
1999
)Пакет WinGX для кристаллографии низкомолекулярных монокристаллов
.J Appl Cryst
32
:837
—838
(
2011
)Наноструктура микрофибрилл целлюлозы в древесине ели
.Proc Natl Acad Sci USA
108
:E1195
—E1203
(
2011
)Расщепление целлюлозы белком CBM33
.Protein Sci
20
:1479
—1483
(
2012
)Твердотельное селективное (13) C возбуждение и спин-диффузионный ЯМР для определения пространственных размеров в стенках растительных клеток
.J Agric Food Chem
60
:1419
—1427
(
2000
)Характеристика поперечных связей между микрофибриллами целлюлозы и их появление во время элонгационного роста в эпикотиле гороха
.Физиология растительных клеток
41
:486
—494
(
2011
)Кортикальные микротрубочки оптимизируют кристалличность клеточной стенки для обеспечения однонаправленного роста Arabidopsis
.Завод J
66
:915
—928
(
1999
)Выражение паттерна в растениях: сочетание молекулярных и основанных на исчислении биофизических парадигм
.Am J Bot
86
:1059
—1076
и другие. (
2012
)Кристалличность микрофибрилл целлюлозы снижается за счет мутации С-концевых остатков трансмембранной области CESA1A903V и CESA3T942I синтазы целлюлозы
.Proc Natl Acad Sci USA
109
:4098
—4103
(
2007
)Устойчивость биомассы: инженерные установки и ферменты для производства биотоплива
.Наука
315
:804
—807
(
2009
)Различия между сортами в размерах микрофибрилл целлюлозы, наблюдаемые с помощью инфракрасной спектроскопии
.Целлюлоза
16
:1
—8
(
1995
)Радиальная упаковка, порядок и беспорядок в фибриллах коллагена
.Biophys J
68
:1661
—1670
(
1992
)Контроль толщины клеточных стенок колленхимы пектинами
.Планта
187
:218
—220
(
2011
)Стенки растительных клеток: супрамолекулярные ансамбли
.Food Hydrocoll
25
:257
—262
(
2007a
)Диаметр микрофибрилл в целлюлозе колленхимы сельдерея: рассеяние рентгеновских лучей и данные ЯМР
.Целлюлоза
14
:235
—246
(
2007b
)Влияние гидратации на расстояние между микрофибриллами целлюлозы первичной стенки: исследование малоуглового рентгеновского рассеяния
.Целлюлоза
14
:401
—408
(
2008
)Выявление структурного и функционального разнообразия клеточных стенок растений
.Curr Opin Plant Biol
11
:308
—313
(
2001
)Рентгеновская структура мерсеризованной целлюлозы II при разрешении 1 Å
.Биомакромолекулы
2
:410
—416
(
2012
)Структурная основа нацеливания на субстрат и катализа грибковыми полисахаридмонооксигеназами
.Структура
20
:1051
—1061
(
2011
)Целлобиогидролаза гидролизует кристаллическую целлюлозу на гидрофобных поверхностях
.J Biol Chem
286
:11195
—11201
(
2006
)Структура и анизотропия клеточной стенки в procuste , мутанте целлюлозосинтазы Arabidopsis thaliana
.Планта
224
:438
—448
(
2005
)Расширение клеточной стенки приводит к координатному разделению параллельных микрофибрилл: данные сканирующей электронной микроскопии и атомно-силовой микроскопии
.Завод J
43
:181
—190
(
2003
)Твердотельный 13 C и 1 H спин-диффузионный ЯМР-анализ структуры микрофибрилл бактериальной целлюлозы
.Твердотельный ядерный магнитный резонон
23
:198
—212
(
2012
)Сравнение моделирования бета целлюлозы I с тремя силовыми полями углеводов
.J Chem Theory Comput
8
:735
—748
(
2011
)Высокотемпературные свойства целлюлозы I
.J Phys Chem B
115
:2155
—2166
(
2006
)Исследования микрокристаллической целлюлозы Ibeta
с помощью компьютерного моделирования.Углеводородный остаток
341
:138
—152
(
1990
)Прямая визуализация поперечных связей в первичной клеточной стенке растения
.J Cell Sci
96
:323
—334
(
2008
)Структура и биосинтез пектина
.Curr Opin Plant Biol
11
:266
—277
(
1992
)Ядерно-магнитно-резонансное исследование пространственных соотношений между химическими компонентами в стенках деревянных клеток
.Holzforschung
46
:205
—210
(
2009
)Структура и свойства микрофибриллы целлюлозы
.J Wood Sci
55
:241
—249
(
2002
)Кристаллическая структура и система водородных связей в целлюлозе Ibeta по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
124
:9074
—9082
(
2003
)Кристаллическая структура и система водородных связей в целлюлозе I (альфа) по данным синхротронной рентгеновской дифракции и дифракции нейтронов на волокне
.J Am Chem Soc
125
:14300
—14306
(
2008
)Генетические доказательства того, что активность синтазы целлюлозы влияет на организацию кортикального массива микротрубочек
.Plant Physiol
147
:1723
—1734
(
2012a
)Изменения биомеханических свойств клеточной стенки у ксилоглюкан-дефицитного мутанта xxt1 / xxt2 Arabidopsis
.Plant Physiol
158
:465
—475
(
2012b
)Пересмотренная архитектура первичных клеточных стенок, основанная на биомеханических изменениях, индуцированных субстрат-специфическими эндоглюканазами
.Plant Physiol
158
:1933
—1943
и другие. (
2011
)Анализ окислительного разложения целлюлозы металлоферментом меди, который использует компоненты биомассы
.Proc Natl Acad Sci USA
108
:15079
—15084
и другие.(
2012
)CHITINASE-LIKE1 / POM-POM1 и его гомолог CTL2 представляют собой взаимодействующие с глюканом белки, важные для биосинтеза целлюлозы у Arabidopsis
.Plant Cell
24
:589
—607
(
2008
)Краткая история SHELX
.Acta Crystallogr A
64
:112
—122
(
2006
)Синтез целлюлозы у высших растений
.Annu Rev Cell Dev Biol
22
:53
—78
(
2004
)Структурные детали кристаллической целлюлозы из высших растений
.Биомакромолекулы
5
:1333
—1339
(
2003
)Поляризованная колебательная спектроскопия волоконных полимеров: водородные связи в целлюлозе II
.Биомакромолекулы
4
:1589
—1595
(
2000
)Новые методы позволяют проводить сравнительный анализ ориентации микротрубочек, текстуры стенок и скорости роста интактных корней Arabidopsis
.Plant Physiol
124
:1493
—1506
(
2009
)Динамическая координация систем цитоскелета и клеточной стенки во время морфогенеза растительных клеток
.Curr Biol
19
:R800
—R811
(
2002
)Молекулярная структура шелка драглайна пауков: складывание и ориентация белкового остова
.Proc Natl Acad Sci USA
99
:10266
—10271
(
2002
)Конформационные особенности кристаллической целлюлозы высших растений
.Завод J
30
:721
—731
(
2004
)Полиморфизм целлюлозы семейства I: повторное исследование целлюлозы IVI
.Биомакромолекулы
5
:1385
—1391
(
1998
)Определение некристаллических форм в целлюлозе I методом спектроскопии ЯМР CP / MAS C-13
.Углеводородный остаток
312
:123
—129
(
2007
)Организация боковых цепей пектинового арабинана и галактана в ассоциации с микрофибриллами целлюлозы в первичных клеточных стенках и связанных моделях предусматривается
.J Exp Bot
58
:1795
—1802
Заметки автора
© Американское общество биологов растений, 2013 г. Все права защищены.
© Автор (ы) 2013. Опубликовано Oxford University Press от имени Американского общества биологов растений. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0 /), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
Микрофибрилла — обзор | Темы ScienceDirect
1 Введение и обоснование
Внеклеточные микрофибриллы присутствуют в большинстве систем органов и широко распространены в сердечно-сосудистой, глазной и скелетной тканях (Hubmacher & Reinhardt, 2011). Микрофибриллы всегда составляют внешнюю оболочку эластических волокон в кровеносных сосудах, легких, коже и других эластичных тканях, где они играют решающую роль в биогенезе и гомеостазе эластичных волокон (Wagenseil & Mecham, 2007).Микрофибриллы также обнаруживаются в отсутствие эластина во многих тканях либо в виде отдельных единиц, либо пересекаются с базальными мембранами. Микрофибриллы — это супрамолекулярные белковые комплексы, связанные с более чем 20 различными компонентами (Baldwin, Simpson, Steer, Cain, & Kielty, 2013). Строительными блоками скелета являются изоформы фибриллина. Семейство фибриллинов у человека состоит из трех высокогомологичных крупных белков (~ 350 кДа), фибриллина-1, -2 и -3, все они кодируются разными генами (Nagase, Nakayama, Nakajima, Kikuno, & Ohara, 2001; Pereira et al. al., 1993; Zhang et al., 1994). Как и многие другие внеклеточные гликопротеины, фибриллины состоят из отдельных доменов. Множественные тандемные массивы доменов, подобных эпидермальному фактору роста, составляют большую часть молекулы фибриллина. Большинство этих доменов EGF связывают кальций, который важен для защиты от протеолиза (Reinhardt, Ono, & Sakai, 1997), контроля взаимодействия лигандов (Lin et al., 2002; Marson et al., 2005; Rock et al., 2004). ; Tiedemann, Bätge, Müller, & Reinhardt, 2001) и структурная стабилизация (Downing et al., 1996; Рейнхардт, Мехлинг и др., 1997; Вернер, Нотт, Хэндфорд, Кэмпбелл и Даунинг, 2000). Другие домены в фибриллинах включают восемь цистеин-содержащих доменов, называемых доменами «ТВ» и «гибридными» доменами. Оба типа доменов обнаруживаются исключительно в фибриллинах и связанных с ними латентных трансформирующих факторах роста-β (TGF-β) -связывающих белках (LTBP) (Hyytiäinen, Penttinen, & Keski-Oja, 2004).
Сборка мономерного фибриллина в зрелые тканевые микрофибриллы — это иерархический процесс, требующий различных белков и множества молекулярных стадий (Sabatier et al., 2014). Критические молекулярные механизмы включают процессинг профибриллина во время секреции или сразу после нее (Ritty, Broekelmann, Tisdale, Milewicz, & Mecham, 1999; Wallis et al., 2003), латеральные самовзаимодействия, производящие единицу, которая напоминает бусину в шарике на поверхности. структура микрофибриллы а-струны (Ashworth, Kelly, Wilson, Shuttleworth, & Kielty, 1999; Hubmacher et al., 2008; Marson et al., 2005; Trask, Ritty, Broekelmann, Tisdale, & Mecham, 1999), взаимодействие с клеткой поверхностный гепарансульфат для концентрирования или выравнивания возникающих микрофибрилл (Cain et al., 2008; Sabatier et al., 2014; Tiedemann et al., 2001), самовзаимодействия N-to-C с удлинением микрофибрилл (Lin et al., 2002; Reinhardt et al., 1996; Sakai, Keene, Glanville, & Bächinger, 1991) и перенос на фибронектин. волокна для стабилизации, удлинения и взаимодействия с другими белками микрофибрилл (Kinsey et al., 2008; Sabatier et al., 2009, 2013). Функциональные взаимодействия с интегринами клеточной поверхности могут играть дополнительную, но менее определенную роль в сборке фибриллина (Bax et al., 2003; Jovanovic et al., 2007; Пфафф, Рейнхардт, Сакаи и Тимпл, 1996; Сакамото и др., 1996). Зрелые микрофибриллы сильно сшиты, что служит для стабилизации трехмерной структуры и обеспечения дополнительной прочности, позволяющей противостоять механическим силам в тканях. В микрофибриллах встречаются два основных типа поперечных связей: восстанавливаемые дисульфидные связи и невосстанавливаемые ɛ- (γ-глутамил) -лизиновые поперечные связи (Gibson, Kumaratilake, & Cleary, 1989; Qian & Glanville, 1997).
Мутации в генах фибриллина вызывают ряд заболеваний соединительной ткани, называемых фибриллинопатиями, которые характеризуются клиническими симптомами в сердечно-сосудистых, кожных, скелетных и глазных тканях (Robinson et al., 2006). Наиболее заметно, что мутации в гене фибриллина-1 приводят к синдрому Марфана, синдрому жесткой кожи, аутосомно-доминантному синдрому Вейля-Марчесани, гелеофизической и акромикрической дисплазии и родственным расстройствам (Collod-Beroud et al., 2003; Frederic et al., 2009) ; Hubmacher & Reinhardt, 2011). Синдром Марфана является наиболее распространенным (1 из 5000) и наиболее изученным из этих расстройств, при этом расслоение и разрыв стенки аорты представляет собой серьезное клиническое осложнение и главную причину смерти (Pyeritz & Dietz, 2002).На людях и мышах было показано, что нарушение регуляции активности TGF-β играет важную роль в патогенезе синдрома Марфана и связанных с ним расстройств (Franken et al., 2013; Matt et al., 2009; Neptune et al., 2003).
В связи с явной значимостью заболевания фибриллины и микрофибриллы часто подвергаются экспериментальному анализу in vivo, в культуре клеток и биохимических анализах in vitro. Здесь мы предлагаем обзор некоторых из наиболее часто используемых в этой области методов анализа фибриллинов и микрофибрилл.
Форма микрофибрилл целлюлозы природных растений
Нишияма, Й., Ланган, П. и Шанзи, Х. Кристаллическая структура и система водородных связей в целлюлозе Iβ по данным синхротронной дифракции рентгеновских лучей и нейтронного волокна. Журнал Американского химического общества 124 , 9074–9082 (2002).
Артикул PubMed CAS Google ученый
Нишияма Ю., Джонсон, Г. П., Френч, А.Д., Форсайт, В. Т. и Ланган, П. Нейтронная кристаллография, молекулярная динамика и квантово-механические исследования природы водородных связей в целлюлозе Iβ. Биомакромолекул 9 , 3133–3140 (2008).
Артикул PubMed CAS Google ученый
Нишияма, Й., Сугияма, Дж., Чанзи, Х. и Ланган, П. Кристаллическая структура и система водородных связей в целлюлозе Iα по данным синхротронной рентгеновской дифракции и дифракции нейтронных волокон. Журнал Американского химического общества 125 , 14300–14306 (2003).
Артикул PubMed CAS Google ученый
Кубицки Дж. Д., Мохамед М. Н.-А. И Уоттс, Х. Д. Квантово-механическое моделирование структур, энергетики и спектральных свойств целлюлозы Iα и Iβ. Целлюлоза 20 , 9–23 (2013).
Артикул CAS Google ученый
Ли, К. М. и др. . Сетка водородных связей и колебания натяжения ОН целлюлозы: сравнение компьютерного моделирования с поляризованными ИК- и SFG-спектрами. Журнал физической химии B 119 , 15138–15149 (2015).
Артикул PubMed CAS Google ученый
Косгроув, Д. Дж. Реконструкция наших моделей целлюлозы и сборки первичной клеточной стенки. Текущее мнение в биологии растений 22 , 122–131 (2014).
Артикул PubMed PubMed Central CAS Google ученый
Джарвис, М.С. Структура микрофибрилл нативной целлюлозы, отправная точка для производства наноцеллюлозы. Phil. Пер. R. Soc. А 376 , 20170045 (2018).
ADS Статья PubMed Google ученый
Ван Т., Янг Х., Кубицки Дж. Д. и Хонг М.Структурный полиморфизм целлюлозы в первичных клеточных стенках растений, исследованный с помощью высокопольной 2D твердотельной ЯМР-спектроскопии и расчетов функциональной теории плотности. Биомакромолекул (2016).
Харрис Д. М. и др. . Кристалличность микрофибрилл целлюлозы снижается путем мутации остатков C-концевых трансмембранных областей CESA1A903V и CESA3T942I целлюлозосинтазы. Труды Национальной академии наук 109 , 4098–4103 (2012).
ADS Статья Google ученый
Ли, К. М., Кафле, К., Хуанг, С. и Ким, С. Х. Мультимодальный широкополосный спектрометр генерации суммарной частоты колебаний (MM-BB-V-SFG) и микроскоп. Журнал физической химии B 120 , 102–116 (2015).
ADS Статья PubMed CAS Google ученый
Herth, W.И Вебер, Г. Возникновение предполагаемых синтезирующих целлюлозу «розеток» в плазматической мембране клеток суспензионной культуры Glycine max. Naturwissenschaften 71 , 153–154 (1984).
ADS Статья CAS Google ученый
Ньюман, Р. Х., Хилл, С. Дж. И Харрис, П. Дж. Данные широкоугольного рентгеновского рассеяния и твердотельного ядерного магнитного резонанса в сочетании с тестовыми моделями микрофибрилл целлюлозы в стенках клеток маша. Физиология растений 163 , 1558–1567 (2013).
Артикул PubMed PubMed Central CAS Google ученый
Фернандес, А. Н. и др. . Наноструктура микрофибрилл целлюлозы в древесине ели. Труды Национальной академии наук 108 , E1195 – E1203 (2011).
ADS Статья Google ученый
Никсон, Б. Т. и др. . Сравнительный структурный и вычислительный анализ поддерживает восемнадцать синтаз целлюлозы в комплексе синтеза целлюлозы растений. Научные отчеты 6 , 28696 (2016).
ADS Статья PubMed PubMed Central CAS Google ученый
Hill, J. L., Hammudi, M. B. & Tien, M. Комплекс целлюлозосинтазы Arabidopsis: предлагаемый гексамер тримеров CESA в эквимолярной стехиометрии. Растительная клетка 26 , 4834–4842 (2014).
Артикул PubMed PubMed Central CAS Google ученый
Вандаваси, В. Г. и др. . Структурное исследование каталитического домена CESA1 комплекса синтеза целлюлозы Arabidopsis: доказательства наличия тримеров CESA. Физиология растений 170 , 123–135 (2016).
Артикул PubMed CAS Google ученый
Янг, Х., Циммер, Дж., Инлинг, Ю. Г. и Кубики, Дж. Д. Как удлиняется целлюлоза · Исследование QM / MM молекулярного механизма полимеризации целлюлозы в бактериальной CESA. Журнал физической химии B 119 , 6525–6535 (2015).
Артикул PubMed CAS Google ученый
Sethaphong, L. et al. . Третичная модель растительной целлюлозосинтазы. Труды Национальной академии наук 110 , 7512–7517 (2013).
ADS Статья Google ученый
Морган, Дж. Л., Струмилло, Дж. И Циммер, Дж. Кристаллографический снимок синтеза целлюлозы и мембранной транслокации. Природа 493 , 181–186 (2013).
ADS Статья PubMed CAS Google ученый
Ли Ю., Лин М. и Дэвенпорт Дж. В. Исследования целлюлозы I из первых принципов: кристаллическая структура, межмолекулярные силы и взаимодействия с водой. Журнал физической химии C 115 , 11533–11539 (2011).
Артикул CAS Google ученый
Bučko, T., Tunega, D., Ángyán, J. G. & Hafner, J. Ab initio исследование структуры и взаимного превращения фаз природной целлюлозы. Журнал физической химии A 115 , 10097–10105 (2011).
ADS Статья PubMed CAS Google ученый
Голдберг, Р. Н. и др. . Термодинамическое исследование алломорфов целлюлозы: целлюлозы (am), целлюлозы Iβ (cr), целлюлозы II (cr) и целлюлозы III (cr). Журнал химической термодинамики 81 , 184–226 (2015).
Артикул CAS Google ученый
Knight, C. J. & Hub, J. S. WAXSiS: веб-сервер для расчета кривых SAXS / WAXS на основе молекулярной динамики явного растворителя. Исследование нуклеиновых кислот 43 , W225 – W230 (2015).
Артикул PubMed PubMed Central CAS Google ученый
Chen, P.-c. И Хаб, Дж. С. Проверка ансамблей решений на основе моделирования молекулярной динамики с помощью данных широкоугольного рентгеновского рассеяния. Биофизический журнал 107 , 435–447 (2014).
ADS Статья PubMed PubMed Central CAS Google ученый
Frisch, M. и др. . Gaussian 09, редакция B. 01. Gaussian, Inc., Уоллингфорд, Коннектикут (2010).
Watts, H. D., Mohamed, M. N. A. & Kubicki, J. D. A. DFT-исследование частот колебаний и химических сдвигов 13C ЯМР модельных фрагментов целлюлозы в зависимости от размера. Целлюлоза 21 , 53–70 (2014).
Артикул CAS Google ученый
Гримме, С., Хансен, А., Бранденбург, Дж. Г. и Баннварт, К. Методы электронной структуры среднего поля с поправкой на дисперсию. Химические обзоры 116 , 5105–5154 (2016).
Артикул PubMed CAS Google ученый
Гримме С., Эрлих С. и Геригк Л. Эффект функции затухания в теории функционала плотности с поправкой на дисперсию. Журнал вычислительной химии 32 , 1456–1465 (2011).
Артикул PubMed CAS Google ученый
Perdew, J. P. & Wang, Y. Точное и простое аналитическое представление энергии корреляции электронного газа. Physical Review B 45 , 13244 (1992).
ADS Статья CAS Google ученый
Чжао, Ю., Шульц, Н. Э. и Трухлар, Д. Г. Дизайн функционалов плотности путем объединения метода удовлетворения ограничений с параметризацией для термохимии, термохимической кинетики и нековалентных взаимодействий. Журнал химической теории и вычислений 2 , 364–382 (2006).
Артикул PubMed CAS Google ученый
Бек, А. Д. Новое смешение теорий Хартри – Фока и теории функционала локальной плотности. Журнал химической физики 98 , 1372–1377 (1993).
ADS Статья CAS Google ученый
Ли, К., Янг, В. и Парр, Р. Г. Развитие формулы корреляционной энергии Колле-Сальветти в функционал электронной плотности. Физический обзор B 37 , 785 (1988).
ADS Статья CAS Google ученый
Cances, E., Mennucci, B. & Tomasi, J. Новый формализм интегрального уравнения для модели поляризуемого континуума: теоретические основы и приложения к изотропным и анизотропным диэлектрикам. Журнал химической физики 107 , 3032–3041 (1997).
ADS Статья CAS Google ученый
Кубики, Дж. Д., Уоттс, Х. Д., Чжао, З. и Чжун, Л. Квантово-механические расчеты взаимодействий целлюлоза-вода: структуры, энергия, частоты колебаний и химические сдвиги ЯМР для поверхностей целлюлозы Iα и Iβ. Целлюлоза 21 , 909–926 (2014).
Артикул CAS Google ученый
Янг, Х. и др. . Структурные факторы, влияющие на химические сдвиги ЯМР 13С целлюлозы: компьютерное исследование. Целлюлоза , 1–14 (2017).
Ван Т. и Хонг М. Исследования структуры целлюлозы и взаимодействия с полисахаридами матрикса в первичных клеточных стенках растений методом ЯМР в твердом состоянии. Журнал экспериментальной ботаники 67 , 503–514 (2015).
Артикул PubMed CAS Google ученый
Kresse, G. & Furthmüller, J. Эффективные итерационные схемы для ab initio расчетов полной энергии с использованием базисного набора плоских волн. Физический обзор B 54 , 11169 (1996).
ADS Статья CAS Google ученый
Кресс, Г., Фуртмюллер, Дж. И Хафнер, Дж. Теория кристаллических структур селена и теллура: эффект поправок обобщенного градиента к приближению локальной плотности. Physical Review B 50 , 13181 (1994).
ADS Статья CAS Google ученый
Кресс, Г. и Хафнер, Дж. Ab initio молекулярная динамика для переходных металлов с открытой оболочкой. Physical Review B 48 , 13115 (1993).
ADS Статья CAS Google ученый
Kresse, G. & Hafner, J.Неэмпирическое молекулярно-динамическое моделирование перехода жидкий металл – аморфный полупроводник в германии. Physical Review B 49 , 14251 (1994).
ADS Статья CAS Google ученый
Саротти, А. М. и Пеллегринет, С. С. Мультистандартный подход к расчетам GIAO 13C ЯМР. Журнал органической химии 74 , 7254–7260 (2009).
Артикул PubMed CAS Google ученый
Watts, H. D., Mohamed, M. N. A. & Kubicki, J. D. Сравнение сдвигов ЯМР, рассчитанных по многостандартному и TMS-стандарту, для кониферилового спирта и применение многостандартного метода к димерам лигнина. Журнал физической химии B 115 , 1958–1970 (2011).
Артикул PubMed CAS Google ученый
Готлиб, Х. Э., Котляр, В. и Нудельман, А. Химические сдвиги ЯМР обычных лабораторных растворителей в виде следов примесей. Журнал органической химии 62 , 7512–7515 (1997).
Артикул PubMed CAS Google ученый
Adamo, C. & Barone, V. Функционалы обмена с улучшенным поведением на больших расстояниях и методы адиабатического соединения без регулируемых параметров: модели mPW и mPW1PW. Журнал химической физики 108 , 664–675 (1998).
ADS Статья CAS Google ученый
Рассолов В. А., Ратнер М. А., Попл Дж. А., Редферн П. К. и Кертисс Л. А. Базис 6-31G * для атомов третьей строки. Журнал вычислительной химии 22 , 976–984 (2001).
Артикул CAS Google ученый
Йоргенсен, В. Л., Чандрасекхар, Дж., Мадура, Дж. Д., Импи, Р. В. и Кляйн, М. Л. Сравнение простых потенциальных функций для моделирования жидкой воды. Журнал химической физики 79 , 926–935 (1983).
ADS Статья CAS Google ученый
Schrödinger Release 2014-1: Maestro, версия 9.7 (2014).
Бэнкс, Дж. Л. и др. . Комплексная программа моделирования, прикладная химическая теория (IMPACT). Журнал вычислительной химии 26 , 1752–1780 (2005).
Артикул PubMed PubMed Central CAS Google ученый
Кейс, Д. А. и др. . Янтарь 14 (2014).
Киршнер, К. Н. и др. . GLYCAM06: обобщаемое биомолекулярное силовое поле. Углеводы. Журнал вычислительной химии 29 , 622–655 (2008).
Артикул PubMed CAS Google ученый
Thomas, L.H. et al. . Дифракционное доказательство структуры микрофибрилл целлюлозы в бамбуке, модель целлюлозы травы и злаков. Биология растений BMC 15 , 153 (2015).
Артикул PubMed PubMed Central CAS Google ученый
Crawshaw, J., Bras, W., Mant, G. & Cameron, R. Одновременные исследования SAXS и WAXS изменений микроструктуры нативных целлюлозных волокон при набухании в водном гидроксиде натрия. Журнал прикладной науки о полимерах 83 , 1209–1218 (2002).
Артикул CAS Google ученый