
Расчет содержания жира по соотношению талии к росту
Отношение талии к росту вычисляется путем деления окружности талии или размера талии на рост. Данные используются для оценки содержания жира в организме человека. Этот онлай калькулятор поможет расчитать содержание жира в теле по отношению талии к росту
Калькулятор талии к росту
Онлайн калькулятор расчета содержания жира в теле по соотношению талии к росту для мужчин и женщин
Формула расчета соотношения талии к росту
Соотношение талии к росту = ( Окружность талии / Рост) x 100
Таблица соотношения талии к роста
| Для мужчин | |
|---|---|
| Соотношение меньше 35 | Аномально Худой |
| от 35 до 43 | Чрезмерно Худой |
| от 43 до 46 | Стройный и здоровый |
| от 46 до 53 | Здоровый, Нормальный, Идеальны Вес |
| от 53 до 58 | |
| от 58 до 63 | Чрезвычайно Избыточный вес / Тучный |
| больше 63 | Чрезмерно Тучный |
| For Women | |
|---|---|
| Соотношение меньше 35 | Аномально Худая |
| от 35 до 42 | Чрезмерно Худая |
| от 42 до 46 | Стройная и здоровая |
| от 46 до 49 | Здоровый, Нормальный, Идеальны Вес |
| от 49 до 54 | Избыточный вес |
| от 54 до 58 | Чрезвычайно Избыточный вес / Тучная |
| больше 58 | Чрезмерно Тучная |
Пример расчета талии к росты
Найти соотношение талии к росту у мужчины при его окружность талии 92 см и росте 182 см?Получаем,
- Окружность талии = 92 см
- Рост = 182 см
Решение:
Подставим данные значения в формулу
Соотношение талии к росту
= ( Окружность талии / Рост ) x 100
= 92 / 182 x 100
= 50.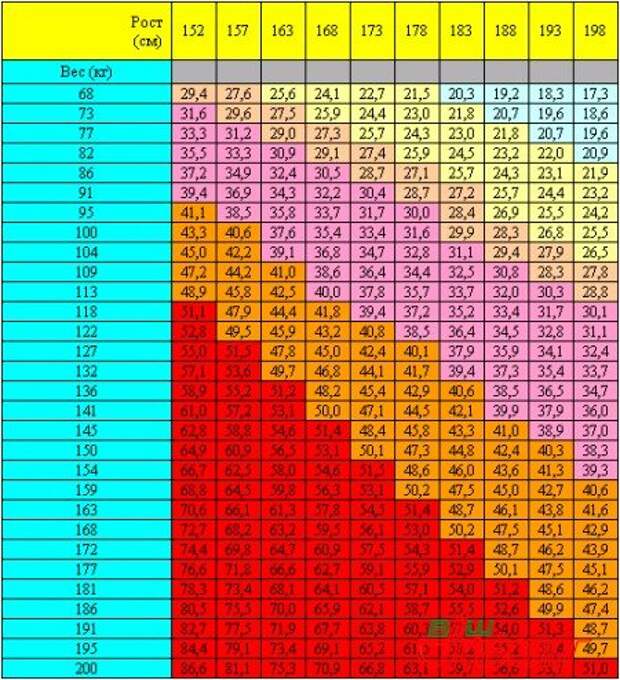 55
55
Как подобрать ростовку сноуборда
Перейти в каталог Выбора сноубордов
Первый вопрос, с которым сталкивается абсолютно каждый начинающий сноубордист, естественно заключается в верном подборе ростовки доски. Как правильно подобрать размеры сноуборда, чтобы на нем было удобно кататься и быстро прогрессировать?
Чтобы правильно выбрать ростовку, необходимо правильно понимать воздействие райдера на поведение сноуборда на спуске. Больше всего на поведение доски влияет вес райдера, именно он распределяется по длине канта. Рост райдера напрямую оказывает гораздо меньшее влияние, смещая центр тяжести чуть выше или ниже. У высокого человека центр тяжести повышается, становится проще «завалить» сноуборд на кант. Зато от роста зависит вес сноубордиста, поэтому, можно говорить, что рост оказывает косвенное влияние на выбор длины сноуборда.
Таким образом, правильнее выбирать сноуборд именно по весу, с небольшой поправкой на пол, рост и условия катания. Но, выбор по росту, также имеет право на существование, о нем поговорим чуть позже. А сейчас приведем методы выбора длины сноуборда от веса райдера.
Но, выбор по росту, также имеет право на существование, о нем поговорим чуть позже. А сейчас приведем методы выбора длины сноуборда от веса райдера.
1. Выбор ростовки сноуборда в зависимости от веса райдера.
Вообще, следует сказать, что производители сноубордов всегда на своей продукции размещают размерную сетку, в которой указано, какой длины сноуборд лучше выбрать в зависимости от вашего веса.
К этим советам стоит прислушиваться, но часто складывается ситуация, как на картинке ниже: имея вес 67-68 кг, райдер может выбрать абсолютно любую ростовку доски (кроме 160см) представленной ниже (BURTON Descedant). В данном случае, советуем абстрагироваться от весовых диапазонов и выбирать сноуборд по среднему значению веса спортсмена. Так, в нашем случае, получится ряд средних: 59, 66, 68, 70, 75 и 79 кг. Соответственно, к какому из значений ближе ваш вес – ту ростовку и следует выбирать.
Существуют формулы, по которым можно рассчитать зависимость длины сноуборда от веса райдера.
Для мужчин формула выглядит следующим образом: Ростовка = 136 + (Вес х 0.3).
Формула для женщин: Ростовка = 127 + (Вес х 0.4).
По данным формулам рассчитывается базовая ростовка сноуборда, которая подходит для катания на средних скоростях по небольшим ухоженным склонам.
Для не подготовленных небольших склонов нужно к базовой ростовке добавить 2-3 см. Для гор и глубокого снега (фрирайд, бэккантри) можно добавить 5-8 см.
Планируете кататься в Ленобласти по небольшим горкам — отнимайте 1-2 см. Для катания в парке и фристайла отнимайте 3-5 см.
Высоким людям можно добавить 2 см, низким – отнять 2 см.
Пример. Мужчина весом 75 кг, среднего роста, планирует кататься в Ленобласти на небольших горках. Базовая ростовка =136+(75х0.3)=158,5 см. Отнимаем 1-2 см (катание на небольших горках), получаем 156,5-157,5 см. Выбираем сноуборд с ростовкой, ближайшей к вычисленному значению.
2. Выбор ростовки сноуборда в зависимости от роста райдера.
Существует универсальный метод определения ростовки сноуборда по росту сноубордиста, с поправкой на телосложение и условия катания:
- Вычитаем от вашего роста 15 см.
- Если у вас плотное телосложение, прибавляем +5 см, если худое, то вычитаем -5 см.
- Если вы только начинающий райдер, то вычитаем -10 см, если у вас средний уровень, то вычитаем -5 см.
- Если вы планируете кататься в глубоком снегу (фрирайд, бэккантри), то следует прибавить +5 см, если вы планируете заниматься фристайлом, следует отнять -5 см.
Так, например, рассуждает райдер Иннокентий с ростом 182 см и весом 77 кг, выбирая себе доску: вычту из своего роста 15см = 167 см. Это моя первая доска (начинающий райдер: -10см), я обладаю нормальным телосложением (нет поправки, т.к. не плотное и не худое) и я планирую кататься на склонах Санкт-Петербурга (нет поправки, т.к. не фрирайд и не фристайл): 167-10=157 см.
В общих словах, следует сказать, что короткие доски считаются более маневренными – ими проще управлять, легче делать трюки, они прощают некоторые ошибки, допускаемые неопытными райдерами во время спусков, но при этом чуть хуже держат скорость.
Сноуборды с большими ростовками, напротив, лучше держат скорость и ведут себя стабильнее и увереннее на серьезных скоростях, но при этом они менее маневренные.
В любом случае, подбор ростовки – это индивидуальный процесс и зависит от специфических предпочтений спортсменов. Именно поэтому, если у вас возникли какие-либо вопросы относительно выбора подходящей ростовки, мы всегда и с радостью поможем сделать правильный выбор!
Перейти в каталог Выбора сноубордов
Таблица размеров (мужских, женских, детских) футболок
Главная Футболки для сублимации оптом, печать на сублимационных футболках Таблица размеров (мужских, женских, детских) футболок
|
Таблица размеров мужских футболок SPORTPOLYESTER, «Ложная сетка», футболок Поло
Определение вашего размера* |
Маркировка наших футболок |
Размеры наших футболок |
||||
Ваш обхват груди, |
Ваш рост, |
Ваш размер, |
Российский стандарт |
Международный стандарт |
Ширина футболки, |
Длина футболки, |
86-90 |
152-164 |
44 |
44 |
XS |
49 |
64 |
90-94 |
158-170 |
46 |
46 |
S |
51 |
66 |
94-98 |
164-176 |
48 |
48 |
M |
53 |
68 |
98-102 |
170-182 |
50 | 50 |
L |
55 |
70 |
102-106 |
176-188 |
52 |
52 |
XL |
57 |
72 |
106-110 |
182-194 |
54 |
54 |
2XL |
59 |
74 |
110-114 |
188-200 |
56 |
56 |
3XL |
61 |
77 |
114-118 |
200 и более |
58 |
58 |
4XL |
63 |
79 |
118-122 |
60 |
60 |
5XL |
65 |
81 |
|
Таблица размеров мужских двухслойных, хлопковых футболок
Определение вашего размера* |
Маркировка наших футболок |
Размеры наших футболок |
||||
Ваш обхват груди, |
Ваш рост, |
Ваш размер |
Российский стандарт |
Международный стандарт |
Ширина футболки, |
Длина футболки, |
86-90 |
152-164 |
44 |
44 |
XS |
48 |
64 |
90-94 |
158-170 |
46 |
46 |
S |
50 |
66 |
94-98 |
164-176 |
48 |
48 |
M |
52 |
68 |
98-102 |
170-182 |
50 |
50 |
L |
54 |
70 |
102-106 |
176-188 |
52 |
52 |
XL |
56 |
72 |
106-110 |
182-194 |
54 |
54 |
2XL |
58 |
74 |
110-114 |
188-200 |
56 |
56 |
3XL |
60 |
76 |
114-118 |
200 и более |
58 |
58 |
4XL |
62 |
78 |
118-122 |
60 |
60 |
5XL |
64 |
80 |
|
Таблица размеров мужских толстовок из ткани ФУТЕР
Определение вашего размера* |
Маркировка |
Размеры наших изделий |
|||||
Ваш обхват груди, |
Ваш рост, |
Ваш размер |
Российский стандарт |
Международный стандарт |
Ширина изделия, |
Длина изделия, |
Длина рукава, |
86-90 |
152-164 |
44 |
44 |
XS |
|||
90-94 |
158-170 |
46 |
46 |
S |
54 |
66 |
61 |
94-98 |
164-176 |
48 |
48 |
M |
56 |
68 |
63 |
98-102 |
170-182 |
50 |
50 |
L |
58 |
70 |
65 |
102-106 |
176-188 |
52 |
52 |
XL |
60 |
72 |
67 |
106-110 |
182-194 |
54 |
54 |
2XL |
62 |
74 |
69 |
110-114 |
188-200 |
56 |
56 |
3XL |
|||
114-118 |
200 и более |
58 |
58 |
4XL |
|||
118-122 |
60 |
60 |
5XL |
||||
*Как определить свой размер:
Чтобы определить свой размер, предлагаем вам взять сантиметровую ленту и измерить ей ваш обхват груди.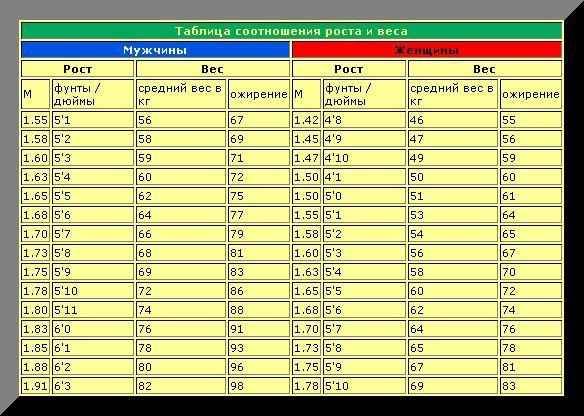 Спереди сантиметровая лента должна проходить по наиболее выступающим точкам груди, сбоку — под подмышечными впадинами. Измерение проводят в спокойном состоянии не на вдохе и выдохе.
Спереди сантиметровая лента должна проходить по наиболее выступающим точкам груди, сбоку — под подмышечными впадинами. Измерение проводят в спокойном состоянии не на вдохе и выдохе.
|
Таблица размеров женских футболок SPORTPOLYESTER
Определение вашего размера* |
Маркировка наших футболок |
Размеры наших футболок |
||||||
Ваш обхват груди, см |
Ваш рост, см |
Ваш размер |
Международный стандарт |
Российский стандарт |
Ширина футболки, см |
Длина футболки, см |
Ширина по талии, см |
Длина рукава, см |
82-85 |
146-158 |
42 |
XS |
42 |
40 |
56 |
35 |
13 |
86-89 |
152-164 |
44 |
S |
44 |
42 |
58 |
37 |
14 |
90-93 |
158-170 |
46 |
M |
46 |
44 |
60 |
39 |
15 |
94-97 |
164-176 |
48 |
L |
48 |
46 |
62 |
41 |
16 |
98-101 |
170-182 |
50 |
XL |
50 |
48 |
64 |
43 |
17 |
102-105 |
176-188 |
52 |
2XL |
52 |
50 |
66 |
45 |
18 |
106-109 |
176-188 |
54 |
3XL |
54 |
52 |
68 |
47 |
19 |
Таблица размеров женских двухслойных футболок
Определение вашего размера* |
Маркировка наших футболок |
Размеры наших футболок |
||||||
Ваш обхват груди, см |
Ваш рост, см |
Ваш размер |
Международный стандарт |
Российский стандарт |
Ширина футболки, см |
Длина футболки, см |
Ширина по талии, см |
Длина рукава, см |
82-85 |
146-158 |
42 |
XS |
42 |
40 |
58 |
35 |
13 |
86-89 |
152-164 |
44 |
S |
44 |
42 |
60 |
37 |
14 |
90-93 |
158-170 |
46 |
M |
46 |
44 |
62 |
39 |
15 |
94-97 |
164-176 |
48 |
L |
48 |
46 |
64 |
41 |
16 |
98-101 |
170-182 |
50 |
XL |
50 |
48 |
66 |
43 |
17 |
102-105 |
176-188 |
52 |
2XL |
52 |
50 |
68 |
45 |
18 |
106-109 |
176-188 |
54 |
3XL |
54 |
52 |
70 |
47 |
19 |
*Как определить свой размер:
Чтобы определить свой размер, вам нужно взять сантиметровую ленту и измерить ей ваш обхват груди.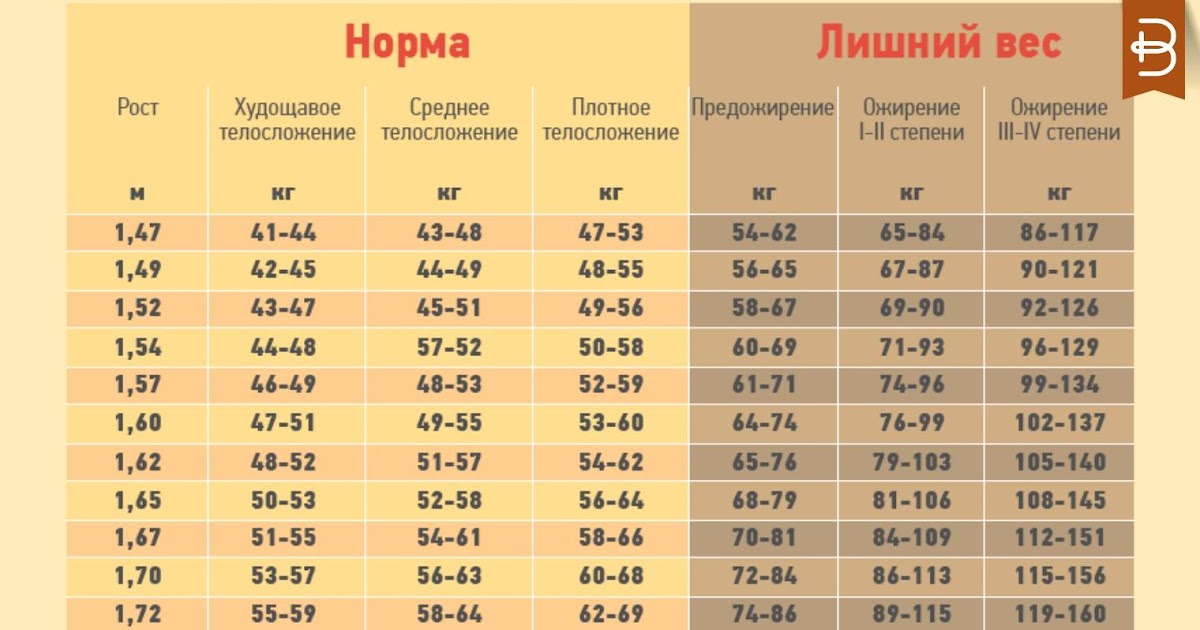 Спереди сантиметровая лента должна проходить по наиболее выступающим точкам груди, сбоку — под подмышечными впадинами. Измерение проводят в спокойном состоянии не на вдохе и выдохе.
Спереди сантиметровая лента должна проходить по наиболее выступающим точкам груди, сбоку — под подмышечными впадинами. Измерение проводят в спокойном состоянии не на вдохе и выдохе.
|
Таблица размеров детских футболок SPORTPOLYESTER, двухслойных, «Ложная сетка»
Определение размера вашего ребенка* |
Маркировка наших футболок |
Размеры наших футболок |
|||||
Возраст |
Обхват груди |
Рост ребенка, |
Размер одежды |
Российский стандарт |
Международный стандарт |
Ширина футболки, |
Длина футболки, |
2 года |
55-58 |
92-98 |
26 |
26 |
98 |
29 |
37 |
3-4 года |
57-62 |
98-110 |
28 |
28 |
104 |
31 |
40 |
4-5 лет |
61-66 |
110-116 |
30 |
30 |
116 |
34 |
44 |
5-6 лет |
65-70 |
116-128 |
34 |
34 |
128 |
37 |
49 |
7-8 лет |
69-74 |
128-134 |
36 |
36 |
134 |
40 |
51 |
9-10 лет |
73-78 |
134-146 |
38 |
38 |
140 |
43 |
55 |
11-12 лет |
77-82 |
146-152 |
40 |
40 |
152 |
46 |
59 |
*Как определить размер вашего ребенка:
Чтобы определить размер ребенка, вам нужно знать его возраст и рост. Еще, вам нужно взять сантиметровую ленту и измерить ей обхват груди ребенка. Спереди сантиметровая лента должна проходить по наиболее выступающим точкам груди, сбоку — под подмышечными впадинами. Измерение проводят в спокойном состоянии не на вдохе и выдохе.
Еще, вам нужно взять сантиметровую ленту и измерить ей обхват груди ребенка. Спереди сантиметровая лента должна проходить по наиболее выступающим точкам груди, сбоку — под подмышечными впадинами. Измерение проводят в спокойном состоянии не на вдохе и выдохе.
Отношение массы мозга плода к массе печени как мера задержки внутриутробного развития: анализ 182 вскрытий мертворожденных
Новорожденных в нижнем 10-м квантиле массы тела называют малыми для гестационного возраста (SGA; 1). Некоторые из них по своему телосложению малы, но другие не смогли реализовать свой потенциал роста из-за задержки внутриутробного развития (IUGR; 2). Младенцы с задержкой роста имеют повышенную перинатальную заболеваемость и смертность (1, 3). Выявление эффектов ЗВУР также важно для мертворожденного плода во время вскрытия, потому что оно может внести большой вклад в понимание этиологии их летального исхода, а его присутствие может быть полезным для руководства пренатальным уходом при будущих беременностях.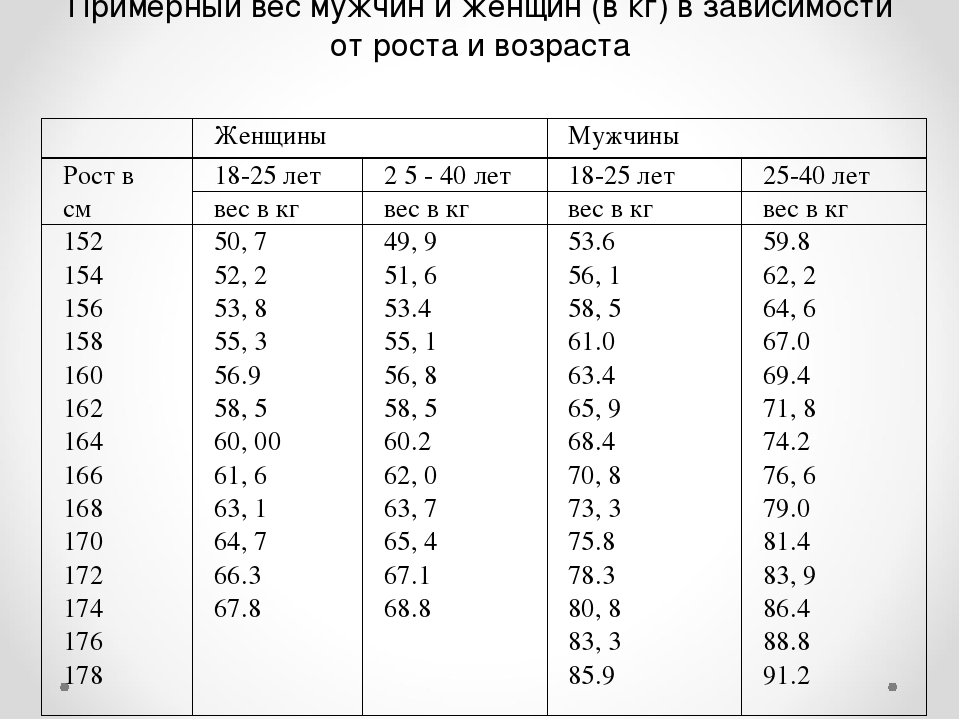
Распространенной схемой классификации плодов с задержкой роста является разделение на симметричные (тип I, пропорциональный, внутренний, гипопластический) и асимметричные (тип II, непропорциональный, внешний, гипотрофический) (1, 4, 5, 6 ). Симметричная задержка роста объясняется инфекцией до 14 недель беременности (, например, , ЦМВ, токсоплазмоз, краснуха) и различными синдромами пороков развития, особенно трисомиями 13, 18 и 21. Плоды с симметричной задержкой роста, согласно теории, кажутся маленькими, но иметь нормальные пропорции тела.Вероятность того, что у них может быть более поздний гестационный рост, чтобы компенсировать преждевременный гестационный отказ в росте, считается мала. Асимметричная задержка роста обычно возникает из-за плацентарной недостаточности материнской гипертонии, многоплодной беременности, материнского недоедания или материнского курения. Плоды с асимметричным ростом кажутся непропорциональными, с сохранением развития мозга за счет других соматических тканей (1, 3, 4, 5, 7, 8, 9).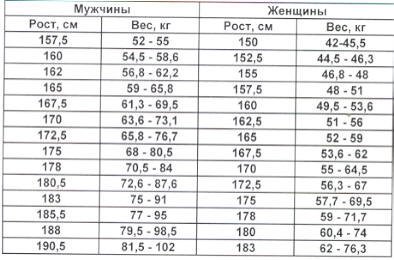
Ряд различных ультразвуковых индексов был использован для обнаружения плода SGA во время развития in utero .К ним относятся отношение головы к окружности живота, отношение средней мозговой артерии к кровотоку в пупочной артерии, а также показатели худобы плода, такие как весовой индекс (7, 10, 11, 12, 13, 14, 15, 16). ). Во время вскрытия прозектор может непосредственно взвесить плод и произвести соответствующие измерения, но как конституционально маленький плод отличается от плода с ЗВУР? Для этого использовали соотношение посмертного мозга плода к массе печени (5). Плоды с асимметричной задержкой роста, согласно теории, должны демонстрировать относительное сохранение массы мозга наряду с габитусом тонкого тела.Мозг плода обычно примерно в три раза превышает вес печени, но при ЗВУР асимметричного типа это значение может увеличиваться в два или три раза. Считается, что это соотношение относительно не зависит от срока гестации и теоретически обеспечивает измерение ЗВУР независимо от массы тела плода (5).
Данные, собранные у 231 мертворожденного плода при патологоанатомическом исследовании в больнице Кристиана, были проанализированы для анализа характеристик соотношения массы мозга / печени. Чтобы попытаться измерить значение отношения для обнаружения ЗВУР, была исследована его связь с низкой массой тела плода, с историей матери и с индексом веса.
Тенденции роста в Англии и Шотландии, 1972–1994 годы
Рост английских и шотландских детей в возрасте от 5 до 11 лет увеличился с 1972 по 1980 год, 1 но с 1979 по 1986 год мало изменился.2 Однако при сравнении наблюдалось более позднее увеличение. Данные за 1990 год с данными, собранными в 1982 году. 3 Увеличение веса и толщины кожной складки трицепса было обнаружено у белых английских детей с 1972 по 1980 год, а также увеличения веса, соотношения веса и роста и толщины кожной складки трицепса у шотландских детей за тот же период.4 Дальнейшее увеличение этих переменных было обнаружено для всех групп с 1982 по 1990 год.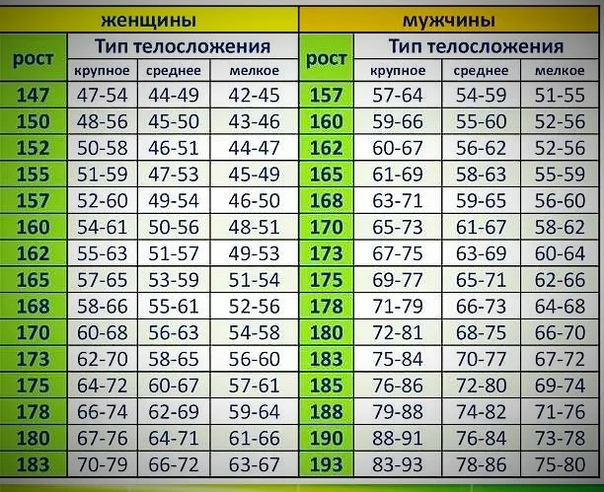
Все эти результаты были взяты из Национального исследования здоровья и роста (NSHG), мониторингового исследования здоровья и роста детей младшего школьного возраста, которое имело смешанный лонгитюдный дизайн.5 Ранние анализы основывались на данных за все годы с 1972 по 1980 г., но в более поздних анализах рассматривались данные по непересекающимся группам детей, измеренным за три разных года, чтобы можно было проанализировать объясняющие факторы.Однако случайно может быть выбран год ниже или выше среднего тренда. Сбор данных в NSHG прекратился в 1994 году. Тенденции роста, веса, веса для индекса роста и кожной складки трицепса представлены по годам, в основном в графической форме, за весь период исследования, с 1972 по 1994 год.
Объекты и методы
NSHG обследовала школьников государственных начальных школ в Англии и Шотландии в течение 23 лет, начиная с 1972 года и заканчивая 1994 годом. Районы были выбраны путем стратифицированной выборки, а затем выбранные школы в этих районах посещались ежегодно до 1982 года, а затем — раз в два года.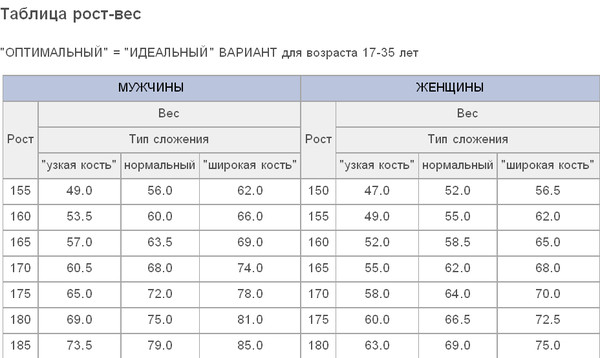 6 В начале исследования выборка из 22 районов в Англии была направлена на предпочтение более бедных районов, но Рона и Альтман обнаружили, что распределение социальных слоев в английских районах было близко к таковому в Англии и Уэльсе.7 Шесть районов в Шотландии. были отобраны отдельно в 1972 г .; поскольку позже было признано, что эта выборка была слишком маленькой, в 1983 г. были добавлены еще восемь областей. С 1972 г. школы выбирались из той же области или группы, чтобы заменить школы, выбывшие из исследования.
6 В начале исследования выборка из 22 районов в Англии была направлена на предпочтение более бедных районов, но Рона и Альтман обнаружили, что распределение социальных слоев в английских районах было близко к таковому в Англии и Уэльсе.7 Шесть районов в Шотландии. были отобраны отдельно в 1972 г .; поскольку позже было признано, что эта выборка была слишком маленькой, в 1983 г. были добавлены еще восемь областей. С 1972 г. школы выбирались из той же области или группы, чтобы заменить школы, выбывшие из исследования.
Все дети в выбранных школах были измерены и взвешены, если родители ребенка не отказались.В эти анализы были включены только дети в возрасте от 5 до 10 лет, которых полевые исследователи считали белыми. Результаты представлены по годам и возрастным группам. Дети младше 5 лет и старше 10,99 года были исключены, потому что мало детей были младше 4,5 лет или старше 11,5 лет.
Рост измерялся на специально разработанном ростометре Holtain с использованием метода, описанного Таннером и др. .8. Рост регистрировался с точностью до последних 0,5 см с 1972 по 1976 год и до последнего 0.1 см с 1977 г. 0,25 или 0,05 см, в зависимости от ситуации, были добавлены для исправления этих смещений. Детей измеряли один раз школьная медсестра, которая читала весы, и ассистент, который растягивал ребенка, под наблюдением обученного полевого работника. В первый год обучения детей растягивал либо полевой работник, либо ассистент. С 1973 по 1976 год детей растягивал ассистент, а с 1977 года — полевой работник. Медсестра также провела измерения веса и трицепса.Детей взвешивали только в трусах, вес фиксировали до последних полных 100 г. Коррекции веса не производилось, так как поправка на 50 г считалась незначительной. Механические весы использовались до 1984 года. Электронные цифровые весы были введены с 1985 года.
.8. Рост регистрировался с точностью до последних 0,5 см с 1972 по 1976 год и до последнего 0.1 см с 1977 г. 0,25 или 0,05 см, в зависимости от ситуации, были добавлены для исправления этих смещений. Детей измеряли один раз школьная медсестра, которая читала весы, и ассистент, который растягивал ребенка, под наблюдением обученного полевого работника. В первый год обучения детей растягивал либо полевой работник, либо ассистент. С 1973 по 1976 год детей растягивал ассистент, а с 1977 года — полевой работник. Медсестра также провела измерения веса и трицепса.Детей взвешивали только в трусах, вес фиксировали до последних полных 100 г. Коррекции веса не производилось, так как поправка на 50 г считалась незначительной. Механические весы использовались до 1984 года. Электронные цифровые весы были введены с 1985 года.
Измерение трицепса проводилось в соответствии с рекомендациями Таннера и Уайтхауса9, за исключением того, что средняя точка между кончиком акромиона и локтевым суставом была отмечена рукой, висящей прямо, а не согнутой. Это означало, что кожная складка трицепса измерялась немного выше того места, которое можно было бы измерить, если бы рука была согнута. С 1972 по 1977 год было проведено два измерения трицепса, каждое записано с точностью до целых 0,2 мм, и для анализа использовалось среднее из двух измерений. Начиная с 1978 г. было проведено только одно измерение, так как два измерения, выполненные одним и тем же измерителем, не привели к небольшому уменьшению вариации. Внутриклассовая корреляция по двум измерениям одним и тем же измерителем в 1972 г. составила 0,99; это тоже было 0.99 для двух измерений, выполненных разными замерщиками.3 Измерение для контроля качества было проведено полевым исследователем у каждого 10-го ребенка. Ежегодно проводилось как минимум одно испытание полевых работников, чтобы гарантировать единообразие методов измерения между полевыми работниками.
Это означало, что кожная складка трицепса измерялась немного выше того места, которое можно было бы измерить, если бы рука была согнута. С 1972 по 1977 год было проведено два измерения трицепса, каждое записано с точностью до целых 0,2 мм, и для анализа использовалось среднее из двух измерений. Начиная с 1978 г. было проведено только одно измерение, так как два измерения, выполненные одним и тем же измерителем, не привели к небольшому уменьшению вариации. Внутриклассовая корреляция по двум измерениям одним и тем же измерителем в 1972 г. составила 0,99; это тоже было 0.99 для двух измерений, выполненных разными замерщиками.3 Измерение для контроля качества было проведено полевым исследователем у каждого 10-го ребенка. Ежегодно проводилось как минимум одно испытание полевых работников, чтобы гарантировать единообразие методов измерения между полевыми работниками.
СТАТИСТИЧЕСКИЕ МЕТОДЫ
Подходящее преобразование было использовано для нормализации веса и измерений трицепса. Вес был преобразован с использованием log 10 (вес-9), это преобразование, как было показано ранее, нормализует вес и стабилизирует дисперсию.10 Толщина кожной складки трицепса была проанализирована по логарифмической шкале; средние были антилогарифмическими, чтобы получить среднее геометрическое, а средняя разница и доверительный интервал (ДИ) обратно преобразованы в процентное увеличение. Журнал индекса 10 (вес-9) / рост 3,7 использовался в качестве подходящей меры веса для индекса роста для этого возрастного диапазона10.
Вес был преобразован с использованием log 10 (вес-9), это преобразование, как было показано ранее, нормализует вес и стабилизирует дисперсию.10 Толщина кожной складки трицепса была проанализирована по логарифмической шкале; средние были антилогарифмическими, чтобы получить среднее геометрическое, а средняя разница и доверительный интервал (ДИ) обратно преобразованы в процентное увеличение. Журнал индекса 10 (вес-9) / рост 3,7 использовался в качестве подходящей меры веса для индекса роста для этого возрастного диапазона10.
95% доверительный интервал для разницы между средними значениями для 1972 и 1994 гг. Для роста, log 10 (вес — 9), log 10 (измерение кожной складки трицепса) и веса для индекса роста для каждой возрастной группы для английского и английского языков. Шотландские мальчики и девочки просчитывались.Изменение журнала 10 (измерение кожной складки трицепса) и его CI были обратно преобразованы, чтобы дать процентное изменение и 95% CI. Из-за смещенного преобразования журнала для веса только приблизительный CI может быть задан для изменения на шкале с обратным преобразованием. Это было рассчитано путем усреднения двух CI, один рассчитан путем добавления пределов CI на преобразованной шкале к среднему значению 1972 года, обратного преобразования и вычитания обратно преобразованного среднего значения 1972 года, второй — из эквивалентного расчета, начиная со среднего значения 1994 года.Средний рост, log 10 (вес -9), log 10 (измерение кожной складки трицепса) и вес для индекса роста были рассчитаны для всех лет с 1972 по 1982 год, а затем раз в два года до 1994 года. Были обследованы 14 районов Шотландии. за двухлетний период, с 1983 по 1984 год, и так далее до 1993-1994 годов. Эти данные использовались для построения графиков с линейной интерполяцией, соединяющих одну точку данных с другой.
Из-за смещенного преобразования журнала для веса только приблизительный CI может быть задан для изменения на шкале с обратным преобразованием. Это было рассчитано путем усреднения двух CI, один рассчитан путем добавления пределов CI на преобразованной шкале к среднему значению 1972 года, обратного преобразования и вычитания обратно преобразованного среднего значения 1972 года, второй — из эквивалентного расчета, начиная со среднего значения 1994 года.Средний рост, log 10 (вес -9), log 10 (измерение кожной складки трицепса) и вес для индекса роста были рассчитаны для всех лет с 1972 по 1982 год, а затем раз в два года до 1994 года. Были обследованы 14 районов Шотландии. за двухлетний период, с 1983 по 1984 год, и так далее до 1993-1994 годов. Эти данные использовались для построения графиков с линейной интерполяцией, соединяющих одну точку данных с другой.
Результаты
Ответ на измерения у белых детей был высоким на протяжении всего исследования, более 95% почти за все годы, как сообщается в ряде статей. 1
3 Каждый год участвовал полный набор территорий, за исключением 1977, 1980, 1981, 1990 и 1994 годов, когда участвовала только 21 английская территория, и в 1982 году, когда были включены только пять шотландских областей.
1
3 Каждый год участвовал полный набор территорий, за исключением 1977, 1980, 1981, 1990 и 1994 годов, когда участвовала только 21 английская территория, и в 1982 году, когда были включены только пять шотландских областей.
ВЫСОТА
Рост у английских и шотландских мальчиков и девочек умеренно линейно увеличивался в течение всего периода времени (рис. 1A и 1B, 2A и 2B). Меньший размер выборки с 1972 по 1982 год очевиден из большего разброса по годам на графиках для шотландских детей, которые показали больший общий прирост, чем английские дети (таблицы 1 и 2).Английские мальчики и девочки показали больший рост в более старшем, чем младшем возрасте, но для детей, живущих в Шотландии, этот рост был более значительным, чем для английских детей, которые существенно не различались (p> 0,05) между возрастными группами. Среднее увеличение составило 2,67 см (95% ДИ от 2,36 до 2,98 см) у шотландских детей по сравнению с 1,39 см (95% ДИ от 1,18 до 1,59 см) у английских детей.
Рисунок 1 Средний рост английских мальчиков (A) и девочек (B), 1972–1994 гг.
Средний рост шотландских мальчиков (A) и девочек (B), 1972–1994 гг.
Таблица 1Средний рост английских детей в 1972 и 1994 годах, по возрастным группам, разнице и 95% ДИ
Таблица 2Средний рост шотландских детей в 1972 и 1994 годах, по возрастным группам, разнице и 95% ДИ
ВЕС
Вес увеличился линейно по преобразованной шкале (рис. 3A и 3B, 4A и 4B). Шотландские дети прибавили в весе больше, чем английские. Как и в случае с ростом, наблюдалась тенденция к увеличению среди английских детей с возрастной группой, но не значительно среди шотландских детей.Обратное преобразование среднего логарифма 10 (вес-9) в таблицах 3 и 4 показывает, что увеличение было от 0,2 до 2,3 кг в зависимости от возрастной группы у английских детей и от 1,0 до 3,3 кг у шотландских детей.
Рисунок 3Среднее значение 10 (вес −9) английских мальчиков (A) и девочек (B), с 1972 по 1994 год.
Рисунок 4 Среднее значение 10 (вес −9) шотландских мальчиков (A) и девочек (B), 1972–1994 гг.
Обратно преобразованный средний преобразованный вес (BTM) и разница для английских детей в 1972 и 1994 годах по возрасту
Таблица 4Обратно преобразованный средний преобразованный вес (BTM) и разница для шотландских детей в 1972 и 1994 годах по возрасту
ВЕС ДЛЯ ИНДЕКСА ВЫСОТЫ
Индекс веса и роста был рассчитан таким образом, чтобы он не зависел от возраста 10, но это не исключает большего увеличения в более старшем возрасте.На рисунках 5A, B и 6A и B показан индекс массы тела по отношению к росту в трех возрастных группах: 5 и 6 лет вместе, 7 и 8 лет, 9 и 10 лет. У самых молодых английских мальчиков наблюдается небольшой спад, у 7-8-летних остается неизменным, а у самых старших — рост. У шотландских мальчиков 5 и 6 лет остались прежними, было небольшое увеличение средней полосы, в то время как 9 и 10 лет показали явный рост, особенно с 1982 года. У девочек как в Англии, так и в Шотландии с 1980 года сложилась четкая закономерность: самая младшая возрастная группа изменилась меньше всего, если вообще изменилась, средняя полоса показывает явный рост, а самые старшие девочки, 9 и 10-летние, имеют самый большой рост. В ходе исследования девочки показали больший рост, чем мальчики, а шотландские дети показали гораздо больший прирост веса по отношению к индексу роста, чем английские дети, начиная с более низкого уровня и заканчивая более тяжелым для своего роста, чем их английские сверстницы (таблицы 5 и 6).
В ходе исследования девочки показали больший рост, чем мальчики, а шотландские дети показали гораздо больший прирост веса по отношению к индексу роста, чем английские дети, начиная с более низкого уровня и заканчивая более тяжелым для своего роста, чем их английские сверстницы (таблицы 5 и 6).
Средний вес для роста английских мальчиков (A) и девочек (B), 1972–1994 гг.
Рисунок 6Средний вес для роста шотландских мальчиков (A) и девочек (B), 1972–1994 гг.
Таблица 5Средний вес для роста английских детей в 1972 и 1994 годах, по возрастным группам, разнице и 95% ДИ
Таблица 6Средний вес для роста шотландских детей в 1972 и 1994 годах, по возрастным группам, разнице и 95% доверительному интервалу
ТРИЦЕПС ТОЛЩИНА КОЖИ
Толщина кожной складки трицепса в целом показала четкую тенденцию к увеличению (рис. 7A и 7B, 8A и 8B), которая была наибольшей у детей старшего возраста, но также заметна у самых маленьких.В Шотландии эта тенденция больше, чем в Англии. Шотландские девушки показали наибольший рост (таблица 8). Число девочек младшего возраста увеличилось более чем на 10%, а девочек 9 и 10 лет — на 19% и 22% соответственно. Девятилетние шотландские мальчики показали прирост чуть менее 24% (таблица 8), что было большим приростом, чем в любой другой группе, но прирост для других шотландских мальчиков был меньше, чем для шотландских девочек. Среди английских мальчиков старшего возраста увеличилось почти на 17%, а в самых младших — примерно на 4% (таблица 7).У английских девочек рост был наименьшим (таблица 7), но тенденция к росту все еще очевидна (рис. 7B), даже у самых молодых англичанок. Однако теперь есть предположение, что с 1986 года, особенно у шотландских детей, рост, возможно, прекратился. Любопытный рост, наиболее заметный в Англии, произошел в 1977 и 1978 годах в младших возрастных группах. Это повторилось снова в 1986 году у детей всех возрастов, причем в Шотландии это было гораздо заметнее, чем в Англии. Ни то, ни другое нельзя объяснить изменениями в школах, полевых работниках или процедурах.
Шотландские девушки показали наибольший рост (таблица 8). Число девочек младшего возраста увеличилось более чем на 10%, а девочек 9 и 10 лет — на 19% и 22% соответственно. Девятилетние шотландские мальчики показали прирост чуть менее 24% (таблица 8), что было большим приростом, чем в любой другой группе, но прирост для других шотландских мальчиков был меньше, чем для шотландских девочек. Среди английских мальчиков старшего возраста увеличилось почти на 17%, а в самых младших — примерно на 4% (таблица 7).У английских девочек рост был наименьшим (таблица 7), но тенденция к росту все еще очевидна (рис. 7B), даже у самых молодых англичанок. Однако теперь есть предположение, что с 1986 года, особенно у шотландских детей, рост, возможно, прекратился. Любопытный рост, наиболее заметный в Англии, произошел в 1977 и 1978 годах в младших возрастных группах. Это повторилось снова в 1986 году у детей всех возрастов, причем в Шотландии это было гораздо заметнее, чем в Англии. Ни то, ни другое нельзя объяснить изменениями в школах, полевых работниках или процедурах.
Среднее значение 10 (кожная складка) английских мальчиков (A) и девочек (B), с 1972 по 1994 год.
Рисунок 8Среднее значение 10 (кожная складка) шотландских мальчиков (A) и девочек (B), с 1972 по 1944 год.
Таблица 8Средняя геометрическая толщина кожной складки (GM) трехглавой мышцы шотландских детей в 1972 и 1994 годах, по возрастным группам, процентному изменению и 95% доверительному интервалу
Таблица 7Средняя геометрическая толщина кожной складки трицепса (GM) для английских детей в 1972 и 1994 годах, по возрастным группам, процентному изменению и 95% ДИ
Обсуждение
Рисунки 1A и 1B, 2A и 2B показывают, что тенденция к увеличению высоты продолжается.Различные оценки тенденций роста за год или за когорту приведены в таблице 9. Прирост за год рассчитывался как разница между средним ростом всех детей в начале соответствующего периода и в конце, деленным на количество лет. Только оценки с 1972 по 1980 год 4 с трендом, измеренным в см на когорту, основывались на трендах за несколько последовательных лет. Теперь ясно, что вывод Чинна и др. о том, что тенденция роста прекратилась, было ошибочным 2, поскольку оценки 1979–1986 годов не соответствовали остальным.Эти оценки были основаны на школах всего в 13 английских областях и трех шотландских областях, которые участвовали в исследовании в период с 1972 по 1986 год, и, следовательно, были подвержены большему разбросу выборки. Это было предназначено для уменьшения возможной систематической ошибки путем исключения замененных школ и районов, но Чинн и Рона не обнаружили разницы в тенденциях между замененными и незамещенными территориями.3 Хотя оценки, основанные на данных за несколько последовательных лет, могут быть предпочтительнее различий между средними значениями для всего за два года проблема уменьшается по мере увеличения временного разрыва.Из-за смешанных продольных данных тенденции, основанные на последовательных годах, не могут быть легко объединены по возрастным группам. Средние оценки с 1972 по 1994 год близки к оценке с 1972 по 1980 год, за исключением шотландских девочек, число которых в старших возрастных группах за последние годы, похоже, увеличилось больше.
Теперь ясно, что вывод Чинна и др. о том, что тенденция роста прекратилась, было ошибочным 2, поскольку оценки 1979–1986 годов не соответствовали остальным.Эти оценки были основаны на школах всего в 13 английских областях и трех шотландских областях, которые участвовали в исследовании в период с 1972 по 1986 год, и, следовательно, были подвержены большему разбросу выборки. Это было предназначено для уменьшения возможной систематической ошибки путем исключения замененных школ и районов, но Чинн и Рона не обнаружили разницы в тенденциях между замененными и незамещенными территориями.3 Хотя оценки, основанные на данных за несколько последовательных лет, могут быть предпочтительнее различий между средними значениями для всего за два года проблема уменьшается по мере увеличения временного разрыва.Из-за смешанных продольных данных тенденции, основанные на последовательных годах, не могут быть легко объединены по возрастным группам. Средние оценки с 1972 по 1994 год близки к оценке с 1972 по 1980 год, за исключением шотландских девочек, число которых в старших возрастных группах за последние годы, похоже, увеличилось больше.
Оценки годовых линейных вековых тенденций роста (см) для английских и шотландских детей
Увеличение роста с возрастом также было обнаружено в других странах, хотя и в исследованиях, которые были завершены до или вскоре после начала нашего собственного.В Швеции Сернеруд и Линдгрен обнаружили, что, хотя 7-летние мальчики в Стокгольме, родившиеся в 1963 году, были почти того же роста, что и мальчики, родившиеся в 1953 году, рост 10-летних увеличивался на 0,07 см в год11. девочки увеличились на 0,04 см в год. Скорость роста у шведских мальчиков аналогична скорости роста английских мальчиков, но меньше, чем у английских девочек (таблица 9). Опрос школьников в возрасте от 8 до 14 лет в Осло, который начался в 1920 году и закончился в 1975 году, показал, что рост вырос на 0.На 055 см в год у мальчиков и на 0,06 см в год у девочек в период с 1955 по 1975 год.12
Предыдущие выводы NSHG в отношении веса и кожной складки трицепса были подтверждены и подвергнуты дальнейшей количественной оценке. Все группы показали увеличение веса, но у английских мальчиков это в основном соответствовало их росту. В других группах вес увеличился больше, чем ожидалось для их роста, о чем свидетельствует увеличение веса по индексу роста, которое было самым большим для шотландских девочек.Ранее сообщалось о более значительных тенденциях соотношения веса и роста у девочек и у шотландских детей 3, и был сделан вывод о том, что шотландские мальчики стали тяжелее и толще, чем их английские сверстники; теперь очевидно, что шотландские девушки также обогнали англичанок.
Все группы показали увеличение веса, но у английских мальчиков это в основном соответствовало их росту. В других группах вес увеличился больше, чем ожидалось для их роста, о чем свидетельствует увеличение веса по индексу роста, которое было самым большим для шотландских девочек.Ранее сообщалось о более значительных тенденциях соотношения веса и роста у девочек и у шотландских детей 3, и был сделан вывод о том, что шотландские мальчики стали тяжелее и толще, чем их английские сверстники; теперь очевидно, что шотландские девушки также обогнали англичанок.
В США не было обнаружено изменений в весе или измерении кожной складки трицепса у 8-летних детей между двумя национальными обследованиями здоровья и питания (NHANES I и II), которые проводились с 1971 по 1974 и с 1976 по 80 годы соответственно.13 Однако короткий промежуток времени между этими двумя исследованиями, возможно, не позволил развиться обнаруживаемым изменениям.
В исследовании, завершившемся через три года после начала нашего, у школьников Осло не было обнаружено увеличения веса. стать несколько стройнее. Вес также оставался неизменным у 7-10-летних детей в Стокгольме в период с 1953 по 1963 год.11Причины этого, если они могут быть установлены, могут быть полезны при планировании интервенций в Великобритании.
стать несколько стройнее. Вес также оставался неизменным у 7-10-летних детей в Стокгольме в период с 1953 по 1963 год.11Причины этого, если они могут быть установлены, могут быть полезны при планировании интервенций в Великобритании.
В предыдущих отчетах не освещались тенденции с указанными здесь возрастными группами, хотя Чинн и Рона сообщили о более значительных тенденциях роста в возрасте 8 лет, чем в возрасте 5 лет, и положительных тенденциях в темпах роста1. тенденция, приводящая к увеличению роста взрослого человека, и тенденция, обусловленная более ранним созреванием, хотя увеличение роста у детей в возрасте до 8 лет и увеличение роста взрослого человека, зарегистрированное в Великобритании в период с 1980 по 1991 год, благоприятствует первому.14 Невозможно различить эффекты времени и когорты, которые смешаны и не могут быть интерпретированы отдельно для непрерывного измерения.
Индекс массы тела к росту дает нечувствительную оценку ожирения, особенно у детей, поскольку этот комплексный индекс также включает вклад костей, мышц и воды. Несмотря на его нечувствительность, заметное увеличение индекса массы тела к росту было продемонстрировано у 9-10-летних и, в меньшей степени, у 7-8-летних.То, что тенденция соотношения веса и роста в основном связана с изменениями в полноте, было подтверждено постоянным увеличением толщины кожной складки трицепса у английских и шотландских мальчиков и девочек. Увеличение толщины кожной складки трицепса было более выражено у детей старшего возраста, но также очевидно в младшей возрастной группе (5 и 6 лет). Оба типа оценок четко продемонстрировали, что прибавка в полноте была больше у шотландских детей, чем у английских детей. Важным следствием наших результатов является то, что для целей мониторинга индекс массы тела к росту может дать ложную уверенность, поскольку для демонстрации тенденции требуются долгосрочные наблюдения и большие выборки.Поэтому мы рекомендуем использовать толщину кожной складки, предпочтительно трехглавой и подлопаточной, для наблюдения за ожирением у детей.
Несмотря на его нечувствительность, заметное увеличение индекса массы тела к росту было продемонстрировано у 9-10-летних и, в меньшей степени, у 7-8-летних.То, что тенденция соотношения веса и роста в основном связана с изменениями в полноте, было подтверждено постоянным увеличением толщины кожной складки трицепса у английских и шотландских мальчиков и девочек. Увеличение толщины кожной складки трицепса было более выражено у детей старшего возраста, но также очевидно в младшей возрастной группе (5 и 6 лет). Оба типа оценок четко продемонстрировали, что прибавка в полноте была больше у шотландских детей, чем у английских детей. Важным следствием наших результатов является то, что для целей мониторинга индекс массы тела к росту может дать ложную уверенность, поскольку для демонстрации тенденции требуются долгосрочные наблюдения и большие выборки.Поэтому мы рекомендуем использовать толщину кожной складки, предпочтительно трехглавой и подлопаточной, для наблюдения за ожирением у детей.
Увеличение веса в зависимости от индекса роста и толщины кожной складки трицепса не предвещает ничего хорошего для целей снижения ожирения 15, как ранее комментировали Чинн и Рона 3, которые не обнаружили устойчивых различий в тенденциях по социальным группам. Хотя есть некоторые предположения, что у шотландских детей толщина кожной складки трицепса могла достигнуть плато, нет никаких доказательств прекращения тенденции соотношения веса к индексу роста.Представление результатов за 23 года подтверждает суть предыдущих отчетов, и в свете исследования Министерства здравоохранения 16, которое показало, что доля мужчин и женщин с ожирением в Великобритании увеличилась вдвое в период с 1980 по 1993 год, до 13% и 16%, соответственно, могут стать дополнительным стимулом для разработки профилактических мероприятий против ожирения для всего населения.
Хотя есть некоторые предположения, что у шотландских детей толщина кожной складки трицепса могла достигнуть плато, нет никаких доказательств прекращения тенденции соотношения веса к индексу роста.Представление результатов за 23 года подтверждает суть предыдущих отчетов, и в свете исследования Министерства здравоохранения 16, которое показало, что доля мужчин и женщин с ожирением в Великобритании увеличилась вдвое в период с 1980 по 1993 год, до 13% и 16%, соответственно, могут стать дополнительным стимулом для разработки профилактических мероприятий против ожирения для всего населения.
Благодарности
Авторы благодарят коллег по команде, а также всех родителей, детей и помощников в изучаемой области.Исследование финансировалось Министерством здравоохранения и Департаментом дома и здравоохранения Шотландии.
Таблица средней длины и веса плода
С самого начала беременности дети растут с разной скоростью, поэтому эти цифры являются средними. Фактическая длина и вес вашего ребенка могут существенно отличаться. Размер родителей и даже расовая принадлежность могут сделать эти цифры несущественными для вашего ребенка.
Размер родителей и даже расовая принадлежность могут сделать эти цифры несущественными для вашего ребенка.Не беспокойтесь, если УЗИ покажет, что ваш ребенок намного меньше или больше. (Ваша акушерка или врач сообщат вам, стоит ли вам беспокоиться о размерах вашего ребенка.)
К доношенному сроку ваш ребенок может в конечном итоге весить менее 5 фунтов или более 9. Обратите внимание, что для целей этой таблицы младенцы измеряются от макушки (или вершины) головы до крупа (или внизу) примерно до 20 недель. После этого их измеряют с головы до пят. (Это связано с тем, что в первой половине беременности ножки ребенка прижаты к его туловищу, и их очень трудно измерить.)
Если у вас есть вопрос о свиданиях, прочитайте нашу статью о том, как BabyCenter Canada определяет, насколько вы беременны.
С восьми до примерно 20 недель вашего ребенка будут измерять от макушки до крупа, как показано ниже:
| Неделя беременности | Длина (дюймы) | Вес (унции) | Длина (см) | Масса (г) |
|---|---|---|---|---|
| 8 недель | 0,63 дюйма | 0,0417 унции | 1 грамм | |
| 9 недель | 0,90 дюйма | 0,07 унции | 2,3 см | 2 грамма |
| 10 недель | 1. 22 дюйма 22 дюйма | 0,14 унции | 3,1 см | 4 грамма |
| 11 недель | 1,61 дюйма | 0,25 унции | 4,1 см | 7 граммов |
| 12 недель | 5,4 см | 14 граммов | ||
| 13 недель | 2,91 дюйма | 0,81 унции | 7,4 см | 23 грамма |
| 14 недель | 3.42 дюйма | 1,52 унции | 8,7 см | 43 грамма |
| 15 недель | 3,98 дюйма | 2,47 унции | 10,1 см | 70 граммов |
| 16 недель | 11,6 см | 100 грамм | ||
| 17 недель | 5,12 дюйма | 4,94 унции | 13 см | 140 грамм |
| 18 недель | 5.59 дюймов | 6,70 унций | 14,2 см | 190 граммов |
| 19 недель | 6,02 дюйма | 8,47 унций | 15,3 см | 240 граммов |
| 208 недель | 16,4 см | 300 грамм |
Затем примерно с 20 недель вашего ребенка будут измерять от макушки до пятки:
| 20 недель | 10. 08 дюймов 08 дюймов | 10,58 унций | 25,6 см | 300 граммов | |||
| 21 неделя | 10,51 дюйма | 12,70 унций | 26,7 см | 360 граммов | недель | 27,8 см | 430 граммов |
| 23 недели | 11,38 дюйма | 1,10 фунта | 28,9 см | 501 грамм | |||
| 24 недели | 11.81 дюйм | 1,32 фунта | 30 см | 600 грамм | |||
| 25 недель | 13,62 дюйма | 1,46 фунта | 34,6 см | 660 граммов | |||
| 26 недель | 35,6 см | 760 граммов | |||||
| 27 недель | 14,41 дюйма | 1,93 фунта | 36,6 см | 875 граммов | |||
| 28 недель | 14.80 дюймов | 2,22 фунта | 37,6 см | 1005 граммов | |||
| 29 недель | 15,2 дюйма | 2,54 фунта | 38,6 см | 1153 грамма | |||
| фунта | 3017 недель | 39,9 см | 1319 граммов | ||||
| 31 неделя | 16,18 дюйма | 3,31 фунта | 41,1 см | 1502 грамма | |||
| 32 недели | 16. 69 дюймов 69 дюймов | 3,75 фунта | 42,4 см | 1702 грамма | |||
| 33 недели | 17,20 дюйма | 4,23 фунта | 43,7 см | фунтов 1918 граммов | |||
| 347 недель | 45 см | 2146 грамм | |||||
| 35 недель | 18,19 дюйма | 5,25 фунта | 46,2 см | 2383 грамма | |||
| 36 недель | 18.66 дюймов | 5,78 фунтов | 47,4 см | 2622 грамма | |||
| 37 недель | 19,13 дюйма | 6,30 фунтов | 48,6 см | 2859 граммов | 9017 9017 | 9017 9017 | 9017 | 49,8 см | 3083 грамма |
| 39 недель | 19,96 дюймов | 7,25 фунтов | 50,7 см | 3288 граммов | |||
| 40 недель | 20.17 дюймов | 51,5 см | 3685 грамм | ||||
| 43 недели | 20,20 дюйма | 8,19 фунта | 51,3 см | 3717 грамм |
Проверено для Канады, Lennox, Andrea
Ссылки
Источники: Doublet PM, Benson CB, Nadel AS, et al: «Улучшенная таблица массы тела при рождении для новорожденных, полученных в результате гестации, датированной ранним ультразвуковым исследованием. «Журнал ультразвуковой медицины. 16: 241, 1997.
«Журнал ультразвуковой медицины. 16: 241, 1997.
Hadlock FP, Shah YP, Kanon DJ и др. «Длина крупа коронки плода: переоценка связи с менструальным возрастом с помощью ультразвуковой радиологии с высоким разрешением в реальном времени». 182: 501, 1992.
Usher R, McLean F. «Внутриутробный рост живорожденных кавказских младенцев на уровне моря: стандарты, полученные на основе измерений в 7 измерениях младенцев, рожденных между 25 и 44 неделями беременности». Педиатрия. т.74, 1969.
Wigglesworth JS. Перинатальная патология, второе издание.W.B. Компания Сондерс. 1996. стр. 24.
Классификация мертворождений в популяционных данных и роль ограничения роста плода: на примере RECODE | BMC по беременности и родам
Gordijn SJ, Korteweg FJ, Erwich JJ, Holm JP, van Diem MT, Bergman KA, Timmer A: Многослойный подход к анализу перинатальной смертности с использованием различных систем классификации. Eur J Obstet Gynecol Reprod Biol. 2009, 144 (2): 99-104. 10.1016 / j.ejogrb.2009.01.012.
10.1016 / j.ejogrb.2009.01.012.
Артикул PubMed Google ученый
Редди У. М., Гольденберг Р., Сильвер Р., Смит Г. К., Паули Р. М., Вапнер Р. Дж., Гардози Дж., Пинар Х., Графе М., Купферминк М. и др.: Классификация мертворождений — разработка международного консенсуса в отношении исследований: резюме национального института семинара по детскому здоровью и человеческому развитию. Obstet Gynecol. 2009, 114 (4): 901-914. 10.1097 / АОГ.0b013e3181b8f6e4.
Артикул PubMed PubMed Central Google ученый
Flenady V, Froen JF, Pinar H, Torabi R, Saastad E, Guyon G, Russell L, Charles A, Harrison C, Chauke L, et al: Оценка систем классификации мертворожденных.BMC Беременность и роды. 2009, 9: 24-10.1186 / 1471-2393-9-24.
Артикул PubMed PubMed Central Google ученый
Фленади В. , Миддлтон П., Смит Г.К., Дюк В., Эрвич Дж., Хонг Т. Ю., Нилсон Дж., Эззати М., Купманс Л., Элвуд Д. и др. Мертворождения: путь вперед в странах с высоким уровнем доходов. Ланцет. 2011, 377 (9778): 1703-1717. 10.1016 / S0140-6736 (11) 60064-0.
, Миддлтон П., Смит Г.К., Дюк В., Эрвич Дж., Хонг Т. Ю., Нилсон Дж., Эззати М., Купманс Л., Элвуд Д. и др. Мертворождения: путь вперед в странах с высоким уровнем доходов. Ланцет. 2011, 377 (9778): 1703-1717. 10.1016 / S0140-6736 (11) 60064-0.
Артикул PubMed Google ученый
Gardosi J, Kady SM, McGeown P, Francis A, Tonks A: Классификация мертворождений по соответствующему состоянию на момент смерти (ReCoDe): популяционное когортное исследование. BMJ. 2005, 331 (7525): 1113-1117. 10.1136 / bmj.38629.587639.7 С.
Артикул PubMed PubMed Central Google ученый
Чан А., Кинг Дж. Ф., Фленади В., Хаслам Р. Х., Тудехоп Д. И.: Классификация перинатальной смерти: разработка классификаций Австралии и Новой Зеландии.J Педиатр детского здоровья. 2004, 40 (7): 340-347. 10.1111 / j.1440-1754.2004.00398.x.
CAS Статья PubMed Google ученый
Froen JF, Pinar H, Flenady V, Bahrin S, Charles A, Chauke L, Day K, Duke CW, Facchinetti F, Fretts RC и др .: Причины смерти и связанные с ними состояния (Codac): a утилитарный подход к классификации перинатальной смерти. BMC Беременность и роды. 2009, 9: 22-10.1186 / 1471-2393-9-22.
Артикул PubMed PubMed Central Google ученый
Дадли Д.Д., Голденберг Р., Конвей Д., Сильвер Р.М., Сааде Г.Р., Варнер М.В., Пинар Х., Кустан Д., Буковски Р., Столл Б. и др.: Новая система для определения причин мертворождения. Obstet Gynecol. 2010, 116 (2): 254-260. 10.1097 / AOG.0b013e3181e7d975.
Артикул PubMed Google ученый
Gardosi J, Madurasinghe V, Williams M, Malik A, Francis A: Материнские и плодовые факторы риска мертворождения: популяционное исследование.BMJ. 2013, 346: f108-10.1136 / bmj.f108.
Артикул PubMed PubMed Central Google ученый

Froen JF, Gardosi JO, Thurmann A, Francis A, Stray-Pedersen B: Ограничение роста плода при внезапной внутриутробной необъяснимой смерти. Acta Obstet Gynecol Scand. 2004, 83 (9): 801-807.
Артикул PubMed Google ученый
Кидрон Д., Бернхейм Дж., Авирам Р.: Плацентарные данные, способствующие внутриутробной гибели плода, исследование 120 мертворожденных между 23 и 40 неделями беременности.Плацента. 2009, 30 (8): 700-704. 10.1016 / j.placenta.2009.05.009.
CAS Статья PubMed Google ученый
Korteweg FJ, Gordijn SJ, Timmer A, Holm JP, Ravise JM, Erwich JJ: Плацентарная причина внутриутробной гибели плода зависит от используемой системы классификации перинатальной смертности. Плацента. 2008, 29 (1): 71-80. 10.1016 / j.placenta.2007.07.003.
CAS Статья PubMed Google ученый

Rey S, Hoffmann P, Arnould P, Jouk PS, Tronc C, Cans C: Tendances et caractéristiques de la mortinatalité dans trois départements alpins (Исер, Савойя и Верхняя Савойя). Revue de médecine périnatale. 2009, 1 (4): 200-207.
Артикул Google ученый
ВОЗ: Международная статистическая классификация болезней и связанных с ними проблем со здоровьем. Десятая редакция. 1993, Женева, Швейцария: Всемирная организация здравоохранения, 129-2
Google ученый
Gardosi J, Mongelli M, Wilcox M, Chang A: регулируемый стандарт веса плода. Ультразвуковой акушерский гинекол. 1995, 6 (3): 168-174. 10.1046 / j.1469-0705.1995.06030168.x.
CAS Статья PubMed Google ученый
Ego A, Subtil D, Grange G, Thiebaugeorges O, Senat MV, Vayssiere C, Zeitlin J: Индивидуальные и популяционные стандарты веса при рождении для выявления младенцев с задержкой роста: французское многоцентровое исследование. Am J Obstet Gynecol. 2006, 194 (4): 1042-1049. 10.1016 / j.ajog.2005.10.816.
Am J Obstet Gynecol. 2006, 194 (4): 1042-1049. 10.1016 / j.ajog.2005.10.816.
Артикул PubMed Google ученый
Блондель Б., Нортон Дж., Дю Мазобрун С., Бреарт Г.: Разработка основных показателей перинатального здоровья в метрополии Франции в период с 1995 по 1998 гг. Результаты национального перинатального обследования. Журнал гинекологии, акушерства и биологии репродукции. 2001, 30 (6): 552-564.
CAS PubMed Google ученый
Hadlock FP, Harrist RB, Martinez-Poyer J: Внутриутробный анализ роста плода: ультразвуковой стандарт веса. Радиология. 1991, 181 (1): 129-133.
CAS Статья PubMed Google ученый
Vergani P, Cozzolino S, Pozzi E, Cuttin MS, Greco M, Ornaghi S, Lucchini V: Определение причин мертворождения: сравнение четырех систем классификации. Am J Obstet Gynecol. 2008, 199 (3): e311-314.
2008, 199 (3): e311-314.
Артикул Google ученый
Cousens S, Blencowe H, Stanton C, Chou D, Ahmed S, Steinhardt L, Creanga AA, Tuncalp O, Balsara ZP, Gupta S и др .: Национальные, региональные и мировые оценки показателей мертворождаемости в 2009 году с тенденциями с 1995 года. : систематический анализ. Ланцет. 2011, 377 (9774): 1319-1330. 10.1016 / S0140-6736 (10) 62310-0.
Артикул PubMed Google ученый
Винбо И.Г., Серениус Ф.Х., Далквист Г.Г., Каллен Б.А.: NICE, новая классификация причин смерти для мертворождений и неонатальных смертей.Классификация неонатальной и внутриутробной смерти по этиологии. Int J Epidemiol. 1998, 27 (3): 499-504. 10.1093 / ije / 27.3.499.
CAS Статья PubMed Google ученый
Froen JF, Gordijn SJ, Abdel-Aleem H, Bergsjo P, Betran A, Duke CW, Fauveau V, Flenady V, Hinderaker SG, Hofmeyr GJ и др. : Считать мертворожденные, заставлять цифры говорить — проблемы в сборе данных о мертворожденных. BMC Беременность и роды.2009, 9: 58-10.1186 / 1471-2393-9-58.
: Считать мертворожденные, заставлять цифры говорить — проблемы в сборе данных о мертворожденных. BMC Беременность и роды.2009, 9: 58-10.1186 / 1471-2393-9-58.
Артикул PubMed PubMed Central Google ученый
Lawn JE, Blencowe H, Pattinson R, Cousens S, Kumar R, Ibiebele I, Gardosi J, Day LT, Stanton C: S: Где? Когда? Почему? Как сделать подсчет данных ?. Ланцет. 2011, 377 (9775): 1448-1463. 10.1016 / S0140-6736 (10) 62187-3.
Артикул PubMed Google ученый
Gardosi J, Figueras F, Clausson B, Francis A: Индивидуальный потенциал роста: международный исследовательский инструмент для изучения эпидемиологии роста плода. Педиатр Перинат Эпидемиол. 2011, 25 (1): 2-10. 10.1111 / j.1365-3016.2010.01166.x.
Артикул PubMed Google ученый
Коул С.К., Эй Э. Н., Томсон А. М.: Классификация перинатальной смерти: акушерский подход. Br J Obstet Gynaecol. 1986, 93 (12): 1204-1212. 10.1111 / j.1471-0528.1986.tb07853.x.
Br J Obstet Gynaecol. 1986, 93 (12): 1204-1212. 10.1111 / j.1471-0528.1986.tb07853.x.
CAS Статья PubMed Google ученый
Korteweg FJ, Gordijn SJ, Timmer A, Erwich JJ, Bergman KA, Bouman K, Ravise JM, Heringa MP, Holm JP: Тюльпановая классификация перинатальной смертности: введение и междисциплинарное соглашение между экспертами. Бьог. 2006, 113 (4): 393-401. 10.1111 / j.1471-0528.2006.00881.x.
CAS Статья PubMed Google ученый
Wigglesworth JS: Мониторинг перинатальной смертности. Патофизиологический подход. Ланцет. 1980, 2 (8196): 684-686.
CAS Статья PubMed Google ученый
Гордон А., Джеффри Х.Э .: Классификация и описание мертворождений в Новом Южном Уэльсе, 2002–2004 гг. Med J Aust. 2008, 188 (11): 645-648.
PubMed Google ученый
Froen JF, Arnestad M, Frey K, Vege A, Saugstad OD, Stray-Pedersen B: Факторы риска внезапной внутриутробной необъяснимой смерти: эпидемиологические характеристики одиночных случаев в Осло, Норвегия, 1986–1995.Am J Obstet Gynecol. 2001, 184 (4): 694-702. 10.1067 / моб.2001.110697.
CAS Статья PubMed Google ученый
Хуанг Д.Й., Ашер Р.Х., Крамер М.С., Ян Х., Морин Л., Фреттс Р.С.: Детерминанты необъяснимой дородовой смерти плода. Obstet Gynecol. 2000, 95 (2): 215-221. 10.1016 / S0029-7844 (99) 00536-0.
CAS PubMed Google ученый
Gardosi J: Систематические обзоры: недостаточно данных для обоснования медицины.Br J Obstet Gynaecol. 1998, 105 (1): 1-5. 10.1111 / j.1471-0528.1998.tb09339.x.
CAS Статья PubMed Google ученый
Практические основы, важность дородовой оценки веса плода, стандартные кривые роста плода
 webmd.com»> Джолли М.К., Себире, штат Нью-Джерси, Харрис Дж. П., Реган Л., Робинсон С. Факторы риска макросомии и ее клинические последствия: исследование 350 311 беременностей. евро J Obstet Gynecol Reprod Biol .10 ноября 2003 г. 111 (1): 9-14. [Медлайн].
webmd.com»> Джолли М.К., Себире, штат Нью-Джерси, Харрис Дж. П., Реган Л., Робинсон С. Факторы риска макросомии и ее клинические последствия: исследование 350 311 беременностей. евро J Obstet Gynecol Reprod Biol .10 ноября 2003 г. 111 (1): 9-14. [Медлайн].Паттерсон Р.М., Прихода Т.Дж., Гиббс К.Э., Вуд Р. Анализ процентиля массы тела при рождении как предиктор перинатального исхода. Акушерский гинекол . 1986 Октябрь 68 (4): 459-63. [Медлайн].
Williams RL, Creasy RK, Cunningham GC, Hawes WE, Norris FD, Tashiro M. Рост плода и перинатальная жизнеспособность в Калифорнии. Акушерский гинекол . 1982 Май. 59 (5): 624-32. [Медлайн].
Wilcox M, Gardosi J, Mongelli M, Ray C., Johnson I.Вес при рождении от беременностей, датированных ультразвуковым исследованием в мультикультурном британском населении. BMJ . 1993, 4 сентября. 307 (6904): 588-91. [Медлайн].
BMJ . 1993, 4 сентября. 307 (6904): 588-91. [Медлайн].
Battaglia FC, Frazier TM, Hellegers AE. Масса тела при рождении, гестационный возраст и исход беременности, с особым вниманием к младенцам с высокой массой тела при рождении — низким гестационным возрастом. Педиатрия . 1966, 37 марта (3): 417-22. [Медлайн].
Нокон Дж. Дж., Маккензи Д. К., Томас Л. Дж., Ханселл Р. С..Дистоция плеча: анализ рисков и акушерские маневры. Am J Obstet Gynecol . 1993 июн. 168 (6 Pt 1): 1732-7; обсуждение 1737-9. [Медлайн].
Мешари А.А., Де Силва С., Рахман И. Макросомия плода — риски для матери и исход для плода. Int J Gynaecol Obstet . 1990 июл.32 (3): 215-22. [Медлайн].
Langer O, Berkus MD, Huff RW, Samueloff A. Дистоция плеча: следует ли родить плод с массой тела более 4000 граммов путем кесарева сечения ?. Am J Obstet Gynecol . 1991, октябрь 165 (4, часть 1): 831-7. [Медлайн].
Am J Obstet Gynecol . 1991, октябрь 165 (4, часть 1): 831-7. [Медлайн].
Бенедетти Т.Дж., Габбе С.Г. Дистоция плеча: осложнение макросомии плода и продолжительного второго периода родов с родоразрешением в среднем тазе. Акушерский гинекол . 1978 ноябрь 52 (5): 526-9. [Медлайн].
Baskett TF, Allen AC. Перинатальные последствия дистоции плеча. Акушерский гинекол . 1995 Июль 86 (1): 14-7. [Медлайн].
Horger EO III, Miller MC III, Conner ED.Связь большой массы тела при рождении с сахарным диабетом матери. Акушерский гинекол . 1975 Февраль 45 (2): 150-4. [Медлайн].
Бойд ME, Usher RH, McLean FH. Макросомия плода: прогноз, риски, предлагаемые методы лечения. Акушерский гинекол . 1983 июн. 61 (6): 715-22. [Медлайн].
[Медлайн].
Грегори К.Д., Генри О.А., Рамиконе Э., Чан Л.С., Платт Л.Д. Осложнения матери и ребенка у новорожденных с нормальным и высоким весом в зависимости от способа родоразрешения. Акушерский гинекол .1998, октябрь 92 (4, часть 1): 507-13. [Медлайн].
Гольдич И.М., Киркман К. Крупный плод. Управление и результат. Акушерский гинекол . 1978 июл.52 (1): 26-30. [Медлайн].
Lazer S, Biale Y, Mazor M, Lewenthal H, Insler V. Осложнения, связанные с макросомным плодом. Дж Репрод Мед . 1986 июн.31 (6): 501-5. [Медлайн].
Parks DG, Ziel HK. Макросомия. Предлагаемое показание к первичному кесареву сечению. Акушерский гинекол . 1978 Октябрь 52 (4): 407-9. [Медлайн].
 webmd.com»> Несбитт Т.С., Гилберт В.М., Херрхен Б. Дистоция плеча и связанные факторы риска у макросомных младенцев, рожденных в Калифорнии. Am J Obstet Gynecol . 1998, август 179 (2): 476-80. [Медлайн].
webmd.com»> Несбитт Т.С., Гилберт В.М., Херрхен Б. Дистоция плеча и связанные факторы риска у макросомных младенцев, рожденных в Калифорнии. Am J Obstet Gynecol . 1998, август 179 (2): 476-80. [Медлайн].Wilcox AJ, Skjaerven R. Вес при рождении и перинатальная смертность: влияние гестационного возраста. Am J Общественное здравоохранение . 1992 марта 82 (3): 378-82. [Медлайн].
Червенак Ю.Л., Дивон М.Ю., Хирш Дж., Гирц Б.А., Лангер О.Макросомия при беременности после родов: показан ли плановый ультразвуковой скрининг? Am J Obstet Gynecol . 1989 Сентябрь 161 (3): 753-6. [Медлайн].
Basso O, Wilcox AJ, Weinberg CR. Вес при рождении и смертность: причинно-следственная связь или противоречие ?. Am J Epidemiol . 2006 15 августа. 164 (4): 303-11. [Медлайн].
 webmd.com»> Lee AC, Kozuki N, Cousens S, et al. Оценки бремени и последствий для младенцев, рожденных малым для гестационного возраста, в странах с низким и средним уровнем дохода с использованием стандарта INTERGROWTH-21 st : анализ наборов данных CHERG. BMJ . 2017 г. 17 августа. 358: j3677. [Медлайн].
webmd.com»> Lee AC, Kozuki N, Cousens S, et al. Оценки бремени и последствий для младенцев, рожденных малым для гестационного возраста, в странах с низким и средним уровнем дохода с использованием стандарта INTERGROWTH-21 st : анализ наборов данных CHERG. BMJ . 2017 г. 17 августа. 358: j3677. [Медлайн].Хваджа СС, Аль-Сибай Х., Аль-Сулейман С.А. Макросомный младенчески-акушерский исход. Саудовская медицина J . 1986. 7: 74-9.
Cheung TH, Leung A, Chang A. Макросомные младенцы. Aust N Z J Obstet Gynaecol . 1990 ноября 30 (4): 319-22. [Медлайн].
Acker DB, Sachs BP, Friedman EA. Факторы риска дистоции плеча. Акушерский гинекол .1985 декабрь 66 (6): 762-8. [Медлайн].
Иффи Л., Бримакомб М., Апуццио Дж. Дж., Варади В., Портуондо Н., Надь Б. Риск необратимой травмы плода, связанной с дистоцией плеча, в зависимости от веса при рождении. евро J Obstet Gynecol Reprod Biol . 2008, январь, 136 (1): 53-60. [Медлайн].
Дж., Варади В., Портуондо Н., Надь Б. Риск необратимой травмы плода, связанной с дистоцией плеча, в зависимости от веса при рождении. евро J Obstet Gynecol Reprod Biol . 2008, январь, 136 (1): 53-60. [Медлайн].
Домбровски М.П., Вулф Х.М., Бранс Ю.В., Салех А.А., Сокол Р.Дж. Морфометрия новорожденных. Связь с акушерскими, педиатрическими и менструальными оценками гестационного возраста. Ам Дж. Дис Детский .1992 июл. 146 (7): 852-6. [Медлайн].
Zhang J, Bowes WA Jr. Распределение веса при рождении и гестационного возраста по расе, полу и паритету среди населения Соединенных Штатов. Акушерский гинекол . 1995 августа 86 (2): 200-8. [Медлайн].
Бреннер В.Е., Эдельман Д.А., Хендрикс СН. Стандарт роста плода для Соединенных Штатов Америки. Am J Obstet Gynecol . 1976 г. , 1 ноября. 126 (5): 555-64. [Медлайн].
, 1 ноября. 126 (5): 555-64. [Медлайн].
Фреман М.Г., Грейвс В.Л., Томпсон Р.Л.Таблицы массы тела при рождении и гестационного возраста для неимущих негров и европеоидов. Педиатрия . 1970 июл.46 (1): 9-15. [Медлайн].
Amini SB, Catalano PM, Hirsch V, Mann LI. Анализ массы тела при рождении по гестационному возрасту с использованием компьютеризированной базы перинатальных данных, 1975–1992 годы. Акушерский гинекол . 1994, март 83 (3): 342-52. [Медлайн].
Arbuckle TE, Wilkins R, Sherman GJ. Процентили веса при рождении по гестационному возрасту в Канаде. Акушерский гинекол .1993, январь, 81 (1): 39-48. [Медлайн].
Грюнвальд П. Рост человеческого плода. I. Нормальный рост и его вариации. Am J Obstet Gynecol . 1966 15 апреля. 94 (8): 1112-9. [Медлайн].
1966 15 апреля. 94 (8): 1112-9. [Медлайн].
Бабсон С.Г., Берман Р.Э., Лессель Р. Рост плода. Вес при рождении живорожденных для гестационного возраста белых младенцев из среднего класса. Педиатрия . 1970 июн. 45 (6): 937-44. [Медлайн].
Александр Г.Р., Химес Дж. Х., Кауфман РБ, Мор Дж., Коган М.Национальный эталон США по развитию плода. Акушерский гинекол . 1996 Февраль 87 (2): 163-8. [Медлайн].
Lubchenco LO, Hansman C, Dressler M, Boyd E. Внутриутробный рост, рассчитанный на основе данных о массе живорожденных при рождении на сроках от 24 до 42 недель беременности. Педиатрия . 1963, ноябрь 32: 793-800. [Медлайн].
Cheng MC, Chew PC, Ratnam SS. Распределение массы тела при рождении у сингапурских китайских, малайских и индийских младенцев от 34 до 42 недель беременности. Дж. Обстет Гинеколь Бр. Коммонв . 1972 Февраль 79 (2): 149-53. [Медлайн].
Дж. Обстет Гинеколь Бр. Коммонв . 1972 Февраль 79 (2): 149-53. [Медлайн].
Ван Х, Гайер Б., Пейдж ДМ. Различия в гестационном весе при рождении среди китайцев, японцев и белых американцев. Int J Epidemiol . 1994 23 февраля (1): 119-28. [Медлайн].
Sterky G. Шведские стандартные кривые для внутриматочного роста. Педиатрия . 1970 Июль 46 (1): 7-8. [Медлайн].
Дэвид Р.Дж.Кривые внутриутробного роста населения на основе компьютеризированных свидетельств о рождении. Саут Мед Дж. . 1983 ноябрь 76 (11): 1401-6. [Медлайн].
Ott WJ. Задержка внутриутробного развития и преждевременные роды. Am J Obstet Gynecol . 1993 июнь 168 (6 Pt 1): 1710-5; обсуждение 1715-7. [Медлайн].
 webmd.com»> Томсон AM, Биллевич WZ, Hytten FE. Оценка роста плода. Дж. Обстет Гинеколь Бр. Коммонв . 1968 Сентябрь 75 (9): 903-16.[Медлайн].
webmd.com»> Томсон AM, Биллевич WZ, Hytten FE. Оценка роста плода. Дж. Обстет Гинеколь Бр. Коммонв . 1968 Сентябрь 75 (9): 903-16.[Медлайн].Вильяр Дж., Шейх Исмаил Л., Виктора С. и др. От Международного консорциума по развитию плода и новорожденного в 21 веке (INTERGROWTH-21st). Международные стандарты веса, длины и окружности головы новорожденных в разбивке по гестационному возрасту и полу: перекрестное исследование новорожденных в рамках проекта INTERGROWTH-21st. Ланцет . Сентябрь 2014 г. 384 (9946): 857-68.
Mikolajczyk RT, Zhang J, Betran AP, et al. Глобальный справочник по процентилям веса плода и веса при рождении. Ланцет . 2011 28 мая. 377 (9780): 1855-61. [Медлайн].
Кэмпбелл С., Warsof SL, Литтл Д., Купер DJ. Регулярное ультразвуковое обследование для прогнозирования гестационного возраста. Акушерский гинекол . 1985 Май. 65 (5): 613-20. [Медлайн].
Акушерский гинекол . 1985 Май. 65 (5): 613-20. [Медлайн].
Наум Г.Г., Станислав Х., Хаффакер Б.Дж. Прибавка веса плода в срок: линейная с минимальной зависимостью от материнского ожирения. Am J Obstet Gynecol . 1995 Май. 172 (5): 1387-94. [Медлайн].
Ebomoyi E, Adetoro OO, Wickremasinghe AR. Вес при рождении и социобиологические факторы в Илорине, Нигерия. J Biosoc Sci . 1991, 23 октября (4): 417-23. [Медлайн].
Унгер С., Вайзер Дж. К., Маккалоу РЭ, Кифер С., Мур LG. Высота, низкий вес при рождении и младенческая смертность в Колорадо. ЯМА . 17 июня 1988 г. 259 (23): 3427-32. [Медлайн].
Shiono PH, Klebanoff MA, Graubard BI, et al. Вес при рождении у женщин разных национальностей. ЯМА . 1986, 3 января. 255 (1): 48-52. [Медлайн].
255 (1): 48-52. [Медлайн].
Дженсен GM, Мур LG. Влияние высоты и других факторов риска на массу тела при рождении: независимые или интерактивные эффекты ?. Am J Общественное здравоохранение . 1997 июн. 87 (6): 1003-7. [Медлайн].
Халид МЭ, Али МЭ, Али KZ. Масса доношенного при рождении и морфология плаценты на большой и малой высоте. Int J Gynaecol Obstet . 1997 июн. 57 (3): 259-65. [Медлайн].
Янси М.К., Мур Дж., Брэди К., Миллиган Д., Стрампель В.Влияние высоты на газы пуповинной крови. Акушерский гинекол . 1992 апр. 79 (4): 571-4. [Медлайн].
Маккалоу РЭ, Ривз Дж. Т.. Задержка роста плода и повышенная младенческая смертность при высоком альтитиде. Арк Энвирон Здоровье . 1977 январь-февраль. 32 (1): 36-9. [Медлайн].
[Медлайн].
Ип Р. Высота и масса тела при рождении. Педиатр Дж. . 1987 декабрь 111 (6, часть 1): 869-76. [Медлайн].
Наум Г.Г., Станислав Х.Гемоглобин, высота над уровнем моря и масса тела при рождении: влияет ли анемия матери во время беременности на рост плода ?. Дж Репрод Мед . 2004 апр. 49 (4): 297-305. [Медлайн].
Наум Г.Г., Фам К.К., Станислав Х. Прогнозирование веса при рождении у испаноязычных женщин с использованием уравнения, основанного на характеристиках матери. евро J Obstet Gynecol Reprod Biol . 2004, 10 февраля. 112 (2): 145-50. [Медлайн].
Наум Г.Г., Станислав Х. Валидация уравнения прогнозирования массы тела при рождении на основе характеристик матери. Дж Репрод Мед . 2002 Сентябрь 47 (9): 752-60. [Медлайн].
[Медлайн].
Наум Г.Г. Выявление и лечение макросомии плода. Contemp Ob / Gyn . 2000 июнь. 89-119. [Полный текст].
Наум Г.Г. Макросомия плода: выявление, риски и лечение. Последипломный акушерский гинекол . 2000. 20 (10): 1-8.
Наум Г.Г., Станислав Х., Хаффакер Б.Дж. Точный прогноз массы тела при рождении на основе измеряемых в перспективе характеристик матери. Дж Репрод Мед . 1999 августа 44 (8): 705-12. [Медлайн].
Ричардс М., Харди Р., Кух Д., Уодсворт, штат Мэн. Вес при рождении и когнитивные функции в когорте родившихся в Британии 1946 года: продольное популяционное исследование. BMJ . 2001, 27 января. 322 (7280): 199-203. [Медлайн].
Gardosi J, Mongelli M, Wilcox M, Chang A. Регулируемый эталон веса плода. Ультразвуковой акушерский гинеколь . 1995 Сентябрь 6 (3): 168-74. [Медлайн].
Регулируемый эталон веса плода. Ультразвуковой акушерский гинеколь . 1995 Сентябрь 6 (3): 168-74. [Медлайн].
Викстром I, Бергстрем Р., Баккетейг Л., Якобсен Г., Линдмарк Г.Прогнозирование высокой массы тела при рождении на основе характеристик матери, высоты дна симфиза и ультразвуковой биометрии. Гинекол Обстет Инвест . 1993. 35 (1): 27-33. [Медлайн].
Моррисон Дж., Уильямс ГМ, Наджман Дж. М., Андерсон МД. Влияние роста и веса отца на вес при рождении. Aust N Z J Obstet Gynaecol . 1991 Май. 31 (2): 114-6. [Медлайн].
Wilcox MA, Newton CS, Johnson IR. Влияние отца на массу тела при рождении. Acta Obstet Gynecol Scand . 1995, январь, 74 (1): 15-8. [Медлайн].
Причард CW, Сазерленд HW, Карр-Хилл, РА. Вес при рождении и рост по отцовской линии. Br J Obstet Gynaecol . 1983 Февраль 90 (2): 156-61. [Медлайн].
Br J Obstet Gynaecol . 1983 Февраль 90 (2): 156-61. [Медлайн].
Кому WW, Cheung W, Kwok JS. Рост и вес отца как детерминанты веса при рождении в китайском населении. Ам Дж. Перинатол . 1998. 15 (9): 545-8. [Медлайн].
McKeown T, Record RG.Влияние внутриутробной среды на корреляцию между массой тела при рождении и ростом родителей. Ам Дж Хам Генет . 1954 Декабрь 6 (4): 457-63. [Медлайн].
Morton NE. Наследование массы тела человека при рождении. Энн Хам Генет . 1955 20 октября (2): 125-34. [Медлайн].
Рейтер RC, Джонс GR, Мур TR. Раса по отцовской принадлежности, масса тела при рождении и риск дистоции среди малазийских гравида. Am J Gynecol Health . 1991. 5: 104-8.
Klebanoff MA, Mednick BR, Schulsinger C, Secher NJ, Shiono PH. Влияние отца на массу тела ребенка при рождении. Am J Obstet Gynecol . 1998 Май. 178 (5): 1022-6. [Медлайн].
Наум Г.Г., Станислав Х. Связь отцовских факторов с массой тела при рождении. Дж Репрод Мед . 2003 декабрь 48 (12): 963-8. [Медлайн].
Lam ETC, Black JM, Little KD, Ausherman J, Rafiroiu C. Влияние физических упражнений на массу тела при рождении: метаанализ. Ам Дж. Исследования в области здравоохранения . 2002. 18 (1): 38-45.
Perkins CCD, Пиварник Дж. М., Найджел Панет, Штейн А. Д.. Физическая активность и рост плода во время беременности. Акушерский гинекол . 2007. 109: 81-7.
Линдси М.К., Грейвс В., Кляйн Л. Взаимосвязь одного значения теста на ненормальную толерантность к глюкозе и осложнений беременности. Акушерский гинекол . 1989, январь 73 (1): 103-6. [Медлайн].
Sermer M, Naylor CD, Gare DJ и др.Влияние увеличения непереносимости углеводов на исходы беременности и родов у 3637 женщин без гестационного диабета. Проект гестационного диабета трех больниц Торонто. Am J Obstet Gynecol . 1995 Июль 173 (1): 146-56. [Медлайн].
Leikin EL, Jenkins JH, Pomerantz GA, Klein L. Аномальные скрининговые тесты на глюкозу во время беременности: фактор риска макросомии плода. Акушерский гинекол . 1987 апр. 69 (4): 570-3. [Медлайн].
Mello G, Parretti E, Mecacci F, eet al.Факторы риска макросомии плода: важность положительного орального теста на глюкозу. евро J Эндокринол . 1997 Июль 137 (1): 27-33. [Медлайн].
Рис Э.А., Сиван Э., Фрэнсис Г., Хомко С.Дж. Исходы беременности среди женщин с диабетическим микрососудистым заболеванием и без него (классы Уайта от B до FR) по сравнению с контрольной группой без диабета. Ам Дж. Перинатол . 1998. 15 (9): 549-55. [Медлайн].
Andrews MC, Jones HW Jr. Нарушение репродуктивной функции матки единорога: задержка внутриутробного развития, бесплодие и повторный аборт в пяти случаях. Am J Obstet Gynecol . 1982 15 сентября. 144 (2): 173-6. [Медлайн].
Haelterman E, Breart G, Paris-Llado J, Dramaix M, Tchobroutsky C. Влияние неосложненной хронической гипертензии на риск родов с малым для гестационного возраста рождением. Am J Epidemiol . 1997 15 апреля. 145 (8): 689-95. [Медлайн].
Velentgas P, Benga-De E, Williams MA. Хроническая гипертензия, гипертензия, вызванная беременностью, и низкий вес при рождении. Эпидемиология .1994 Май. 5 (3): 345-8. [Медлайн].
Xiong X, Mayes D, Demianczuk N, et al. Влияние гипертонии, вызванной беременностью, на рост плода. Am J Obstet Gynecol . 1999, январь 180 (1, часть 1): 207-13. [Медлайн].
Эскенази Б., Фенстер Л., Сидней С., Элкин Е.П. Задержка развития плода у повторнородящих и нерожавших женщин с преэклампсией. Am J Obstet Gynecol . 1993 ноябрь 169 (5): 1112-8. [Медлайн].
Педерсен Дж.Вес и длина тела при рождении новорожденных от матерей с диабетом. Acta Endocrinol (Копен) . 1954 16 августа (4): 330-42. [Медлайн].
Берк М.А., Мимуни Ф., Миодовник М., Герцберг В., Валак Дж. Макросомия у младенцев инсулинозависимых матерей с диабетом. Педиатрия . 1989 июн.83 (6): 1029-34. [Медлайн].
Шах ПС. Отцовские факторы и низкий вес при рождении, преждевременные роды и роды с малым весом для гестационного возраста: систематический обзор. Am J Obstet Gynecol .2010 Февраль 202 (2): 103-23. [Медлайн].
Keirse MJ. Эпидемиология и этиология задержки роста ребенка. Clin Obstet Gynaecol . 1984 августа, 11 (2): 415-36. [Медлайн].
Наум Г.Г., Хаффакер Б.Дж. Расовые различия в результатах перорального скринингового теста на глюкозу: установление расовых критериев отклонений во время беременности. Акушерский гинекол . 1993 Апрель, 81 (4): 517-22. [Медлайн].
Голденберг Р.Л., Кливер С.П., Каттер Г.Р. и др.Черно-белые различия в антропометрических измерениях новорожденных. Акушерский гинекол . 1991, ноябрь 78 (5, часть 1): 782-8. [Медлайн].
Hulsey TC, Levkoff AH, Александр ГР. Вес при рождении новорожденных от черных и белых матерей без осложнений беременности. Am J Obstet Gynecol . 1991 Май. 164 (5 Пет 1): 1299-302. [Медлайн].
Breschi MC, Seghieri G, Bartolomei G, Gironi A, Baldi S, Ferrannini E. Отношение веса при рождении к концентрации глюкозы в плазме крови и концентрации инсулина матери во время нормальной беременности. Диабетология . 1993 Декабрь 36 (12): 1315-21. [Медлайн].
Милетик Т., Стойни Э. Влияние веса матери до беременности, роста и индекса массы тела на массу тела при рождении новорожденных мужского и женского пола. Coll Antropol . 29 июня 2005 г. (1): 263-6. [Медлайн].
Johnsen S, Rasmussen S, Wilsgaard T, Sollien R, Kiserud T. Продольные контрольные диапазоны для расчетной массы плода. Acta Obstet Gynecol . 2006. 85: 286-297.
Кливер С.П., Голденберг Р.Л., Каттер Г.Р., Хоффман Х.Дж., Дэвис Р.О., Нельсон К.Г. Влияние курения сигарет на неонатальные антропометрические измерения. Акушерский гинекол . 1995 апр. 85 (4): 625-30. [Медлайн].
Horta BL, Victora CG, Menezes AM, Halpern R, Barros FC. Низкая масса тела при рождении, преждевременные роды и задержка внутриутробного развития в связи с курением матери. Педиатр перинат эпидемиологии . 1997 г., 11 (2): 140-51.[Медлайн].
Матай М., Скиннер А., Лоутон К., Вайндлинг А.М. Курение матери, уровни котинина в моче и масса тела при рождении. Aust N Z J Obstet Gynaecol . 1990 Февраль 30 (1): 33-6. [Медлайн].
Канди Т., Гэмбл Г., Мануэль А., Тауненд К., Робертс А. Детерминанты веса при рождении у женщин с установленным и гестационным диабетом. Aust N Z J Obstet Gynaecol . 1993 августа 33 (3): 249-54. [Медлайн].
Loeffler FE.Прогнозирование клинической массы плода. Дж. Обстет Гинеколь Бр. Коммонв . 1967 Октябрь 74 (5): 675-7. [Медлайн].
Ong HC, Sen DK. Клиническая оценка массы плода. Am J Obstet Gynecol . 1972 г., 1. 112 (7): 877-80. [Медлайн].
Ханретти К.П., Нилсон Дж. П., Флеминг Э. Повторная оценка клинической оценки веса плода: сравнение с ультразвуком. Дж. Обстет Гинеколь . 1990. 10: 199-201.
Bossak WS, Spellacy WN.Точность определения веса плода при пальпации живота. Дж Репрод Мед . 1972, 9 августа (2): 58-60. [Медлайн].
Кауфманн Р.С., Макбрайд П., Аманква К.С., Хаффман Д.Г. Влияние малых степеней непереносимости глюкозы на частоту макросомии новорожденных. Акушерский гинекол . 1992 июл. 80 (1): 97-101. [Медлайн].
Berkus MD, Langer O. Тест толерантности к глюкозе: степень нарушения глюкозы коррелирует с неонатальным исходом. Акушерский гинекол . 1993 г., 81 (3): 344-8. [Медлайн].
Чаухан С.П., Латтон П.М., Бейли К.Дж., Герьери Дж. П., Моррисон Дж. Клинические, сонографические и родовые оценки веса новорожденного у новорожденных во время родов. Акушерский гинекол . 1992 июн. 79 (6): 956-8. [Медлайн].
Herrero RL, Fitzsimmons J. Расчетный вес плода: оценка матери и врача. Дж Репрод Мед . 1999 Август 44 (8): 674-8. [Медлайн].
Чаухан С.П., Салливан, Калифорния, Луттон ТС, Маганн Е.Ф., Моррисон Дж.Оценка веса при рождении роженицами при переношенной беременности. Дж Перинатол . 1995 май-июнь. 15 (3): 192-4. [Медлайн].
Наум Г.Г. Прогнозирование веса плода. Стоит ли обучать маневрам Леопольда студентам-медикам и медицинскому персоналу? Дж Репрод Мед . 2002 Апрель, 47 (4): 271-8. [Медлайн].
Шепард MJ, Ричардс VA, Берковиц RL, Warsof SL, Hobbins JC. Оценка двух уравнений для прогнозирования веса плода с помощью ультразвука. Am J Obstet Gynecol . 1982 г., 1. 142 (1): 47-54. [Медлайн].
Hadlock FP, Harrist RB, Carpenter RJ, Deter RL, Park SK. Сонографическая оценка веса плода. Значение длины бедра в дополнение к измерениям головы и живота. Радиология . 1984 Февраль 150 (2): 535-40. [Медлайн].
Hadlock FP, Harrist RB, Sharman RS, Deter RL, Park SK. Оценка веса плода по измерениям головы, тела и бедра — перспективное исследование. Am J Obstet Gynecol . 1985, 1 февраля. 151 (3): 333-7. [Медлайн].
Sabbagha RE, Minogue J, Tamura RK, Hungerford SA. Оценка массы тела при рождении с использованием формул для ультразвукового исследования, предназначенная для крупных, подходящих и малых для гестационного возраста плодов. Am J Obstet Gynecol . 1989 Apr.160 (4): 854-60; обсуждение 860-2. [Медлайн].
Warsof SL, Gohari P, Berkowitz RL, Hobbins JC. Оценка веса плода с помощью компьютерного анализа. Am J Obstet Gynecol . 1977 15 августа 128 (8): 881-92. [Медлайн].
Warsof SL, Wolf P, Coulehan J, Queenan JT. Сравнение формул оценки веса плода с измерением головы и без него. Акушерский гинекол . 1986 апр. 67 (4): 569-73. [Медлайн].
Noumi G, Collado-Khoury F, Bombard A, Julliard K, Weiner Z. Клиническая и сонографическая оценка веса плода, проводимая резидентами во время родов. Am J Obstet Gynecol .2005 Май. 192 (5): 1407-9. [Медлайн].
Watson WJ, Soisson AP, Harlass FE. Предполагаемый вес доношенного плода. Точность УЗИ по сравнению с клиническим обследованием. Дж Репрод Мед . 1988 Апрель, 33 (4): 369-71. [Медлайн].
Чаухан С.П., Латтон ТС, Бейли К.Дж., Моррисон Дж. Прогноз массы тела при рождении во время родов: клиническая и сонографическая оценка, основанная только на длине бедренной кости. Акушерский гинекол . 1993 Май. 81 (5 (Pt 1)): 695-7.[Медлайн].
Чаухан С.П., Перри К.Г., Маганн Э.Ф. Оценка массы тела при рождении во время родов: клинические и сонографические модели с использованием от одного до четырех параметров плода. Дж Матерн Фетал Инвест . 1994. 4: 263-7.
Chauhan SP, Cowan BD, Magann EF, Bradford TH, Roberts WE, Morrison JC. Обнаружение макросомного плода во время родов: клиническая картина по сравнению с 8 сонографическими моделями. Aust N Z J Obstet Gynaecol . 1995 августа 35 (3): 266-70.[Медлайн].
Сармандал П., Бейли С.М., Грант Дж. Сравнение трех методов оценки вариаций между наблюдателями, примененных к ультразвуковому измерению плода в третьем триместре. Br J Obstet Gynaecol . 1989 ноябрь 96 (11): 1261-5. [Медлайн].
Чаухан С.П., Хендрикс Н.В., Маганн Е.Ф., Моррисон Дж. К., Кенни С.П., Дево Л.Д. Ограничения клинических и сонографических оценок массы тела при рождении: опыт с 1034 роженицами. Акушерский гинекол .1998, январь, 91 (1): 72-7. [Медлайн].
Hendrix NW, Grady CS, Chauhan SP. Клиническая и сонографическая оценка веса при рождении у доношенных родов. Рандомизированное клиническое испытание. Дж Репрод Мед . 2000 апр. 45 (4): 317-22. [Медлайн].
Дар П., Вайнер И., Софрин О., Сакс Г. С., Буковский И., Ариэли С. Клинические и сонографические оценки веса плода при активных родах с разрывом плодных оболочек. Дж Репрод Мед . 2000 Май. 45 (5): 390-4.[Медлайн].
Johnstone FD, Prescott RJ, Steel JM, Mao JH, Chambers S, Muir N. Клиническое и ультразвуковое прогнозирование макросомии при диабетической беременности. Br J Obstet Gynaecol . 1996 августа 103 (8): 747-54. [Медлайн].
Этридж Дж. К. Младший, Луис Дж. М., Мерсер Б.М. Точность определения веса плода с помощью УЗИ при ближайших родах. J Matern Fetal Neonatal Med . 2013 19 сентября [Medline].
Барел О, Маймон Р., Вакнин З., Товбин Дж., Сморгик Н.Сонографическая оценка веса плода — это что-то большее, чем просто измерения плода ?. Пренат Диагностика . 2013 г. 4 октября [Medline].
Наум Г.Г., Фам К.К., МакХью Дж.П. Ультразвуковое предсказание веса при рождении у латиноамериканских женщин. Точность в поликлинике. Дж Репрод Мед . 2003 января. 48 (1): 13-22. [Медлайн].
Platek DN, Divon MY, Anyaegbunam A, Merkatz IR. Ультрасонографическая оценка веса плода персоналом дома во время родов. Am J Obstet Gynecol . 1991, октябрь 165 (4, часть 1): 842-5. [Медлайн].
Кэмпбелл С., Уилкин Д. Ультразвуковое измерение окружности живота плода при оценке веса плода. Br J Obstet Gynaecol . 1975 сентябрь 82 (9): 689-97. [Медлайн].
Хиггинботтом Дж., Слейтер Дж., Портер Дж., Уитфилд С.Р. Оценка веса плода по ультразвуковому измерению окружности туловища. Br J Obstet Gynaecol . 1975 сен.82 (9): 698-701. [Медлайн].
Jordaan HV. Оценка веса плода с помощью УЗИ. J Clin Ультразвук . 1983 фев-март. 11 (2): 59-66. [Медлайн].
Ву Дж.С., Ван CW, Чо КМ. Компьютерная оценка ультразвукового прогноза веса плода с использованием уравнений множественной регрессии с учетом и без длины бедренной кости плода. J Ультразвук Med . 1985, 4 февраля (2): 65-7. [Медлайн].
Винцилеос AM, Кэмпбелл WA, Родис JF, Bors-Koefoed R, Nochimson DJ.Формулы оценки веса плода с измерением окружности головы, живота, бедра и бедра. Am J Obstet Gynecol . 1987 Август 157 (2): 410-4. [Медлайн].
Hsieh FJ, Chang FM, Huang HC, Lu CC, Ko TM, Chen HY. Компьютерный анализ для прогнозирования веса плода путем ультразвукового сравнения бипариетального диаметра (BPD), окружности живота (AC) и длины бедра (FL). Тайвань Йи Сюэ Хуэй За Чжи . 1987 сентябрь 86 (9): 957-64. [Медлайн].
Шинозука Н., Окай Т., Кохзума С. и др.Формулы для оценки веса плода с помощью ультразвуковых измерений на основе удельного веса и объемов новорожденных. Am J Obstet Gynecol . 1987 ноябрь 157 (5): 1140-5. [Медлайн].
Ott WJ, Doyle S, Flamm S, Wittman J. Точная ультразвуковая оценка веса плода. Перспективный анализ новых ультразвуковых формул. Ам Дж. Перинатол . 1986, 3 (4) октября: 307-10. [Медлайн].
Combs CA, Jaekle RK, Rosenn B, Pope M, Miodovnik M, Siddiqi TA.Сонографическая оценка веса плода на основе модели объема плода. Акушерский гинекол . 1993 Сентябрь 82 (3): 365-70. [Медлайн].
Наум Г.Г., Станислав Х. Ультрасонографическое предсказание массы тела при рождении: насколько это точно ?. Am J Obstet Gynecol . 2003 Февраль 188 (2): 566-74. [Медлайн].
Смит Г.К., Смит М.Ф., Макней МБ, Флеминг Дж.Э. Связь между окружностью живота плода и массой тела при рождении: данные по 3512 беременностям. Br J Obstet Gynaecol . 1997 Февраль 104 (2): 186-90. [Медлайн].
Abramowicz JS, Sherer DM, Bar-Tov E, Woods JR. Отщечный диаметр при ультразвуковой оценке роста плода. Am J Obstet Gynecol . 1991, октябрь 165 (4, часть 1): 846-52. [Медлайн].
Santolaya-Forgas J, Meyer WJ, Gauthier DW, Kahn D. Отношение подкожной клетчатки плода к длине бедра во время родов: ключ ультразвукового исследования к макросомии плода. Am J Obstet Gynecol . 1994 Октябрь 171 (4): 1072-5. [Медлайн].
Суд А.К., Янси М., Ричардс Д. Прогнозирование макросомии плода с использованием толщины мягких тканей плечевой кости. Акушерский гинекол . 1995 июн. 85 (6): 937-40. [Медлайн].
Андерсон Н.Г., Джолли И.Дж., Уэллс Дж. Сонографическая оценка веса плода: сравнение систематической ошибки, точности и постоянства с использованием 12 различных формул. Ультразвуковой акушерский гинеколь . 2007 Август.30 (2): 173-9. [Медлайн].
Наум Г.Г., Станислав Х. Одно только УЗИ уступает комбинированным методам прогнозирования веса плода. Ультразвуковой акушерский гинеколь . 2007 30 ноября (6): 913-4. [Медлайн].
Jaeschke R, Guyatt GH, Sackett DL. Справочники по медицинской литературе. III. Как пользоваться статьей о диагностическом тесте. Б. Каковы результаты и помогут ли они мне в уходе за пациентами? Рабочая группа по доказательной медицине. ЯМА . 1994, 2 марта. 271 (9): 703-7. [Медлайн].
Scioscia M, Vimercati A, Ceci O, Vicino M, Selvaggi LE. Оценка веса при рождении с помощью двумерного ультразвукового исследования: критическая оценка его точности. Акушерский гинекол . 2008, январь 111 (1): 57-65. [Медлайн].
Колман А., Махарадж Д., Хаттон Дж., Туохи Дж. Надежность ультразвуковой оценки веса плода при доношенных одноплодных беременностях. N Z Med J . 2006 г.119 (1241): U2146. [Медлайн].
Чаухан С.П., Хендрикс Н.В., Маганн Э.Ф., Моррисон Дж. К., Скардо Дж. А., Бергелла В. Обзор сонографической оценки веса плода: капризы точности. J Matern Fetal Neonatal Med . 2005 октября 18 (4): 211-20. [Медлайн].
Benacerraf BR, Gelman R, Frigoletto FD Jr. Сонографически оцененный вес плода: точность и ограничение. Am J Obstet Gynecol . 1988 ноябрь 159 (5): 1118-21. [Медлайн].
Halaska MG, Vlk R, Feldmar P, Hrehorcak M, Krcmar M, Mlcochova H. Предсказание веса при рождении при рождении с использованием УЗИ и характеристик матери. евро J Obstet Gynecol Reprod Biol . 2006 сентябрь-октябрь. 128 (1-2): 231-5. [Медлайн].
Sherman DJ, Arieli S, Tovbin J, Siegel G, Caspi E, Bukovsky I. Сравнение клинической и ультразвуковой оценки веса плода. Акушерский гинекол . 1998 Февраль 91 (2): 212-7. [Медлайн].
Fox NS, Bhavsar V, Saltzman DH, Rebarber A, Chasen ST.Влияние индекса массы тела матери на клиническую оценку массы плода при доношенной беременности. Акушерский гинекол . 2009 Март 113 (3): 641-5. [Медлайн].
Наум Г.Г., Станислав Х. Компьютеризированный метод точного прогнозирования макросомии плода за 11 недель до родов. евро J Obstet Gynecol Reprod Biol . 2007 августа 133 (2): 148-56. [Медлайн].
Чаухан С.П., Гробман В.А., Герман Р.А., Чаухан В.Б., Чанг Дж., Маганн Э.Ф.Подозрение и лечение макросомного плода: обзор. Am J Obstet Gynecol . 2005 Август 193 (2): 332-46. [Медлайн].
Сокол Р.Ж., Чик Л., Домбровский М.П., Задор И.Е. Правильная идентификация макросомного плода: улучшение прогнозов на основе ультразвукового исследования. Am J Obstet Gynecol . 2000 июн.182 (6): 1489-95. [Медлайн].
Coomarasamy A, Connock M, Thornton J, Khan KS. Точность ультразвуковой биометрии в прогнозировании макросомии: систематический количественный обзор. БЖОГ . 2005 ноябрь 112 (11): 1461-6. [Медлайн].
Combs CA, Singh NB, Khoury JC. Выборная индукция в сравнении со спонтанными родами после сонографической диагностики макросомии плода. Акушерский гинекол . 1993 апр. 81 (4): 492-6. [Медлайн].
Гонен О., Розен Д. Д., Долфин З., Теппер Р., Марков С., Фейгин М. Д. Индукция родов по сравнению с выжидательной тактикой при макросомии: рандомизированное исследование. Акушерский гинекол . 1997 июн. 89 (6): 913-7.[Медлайн].
Friesen CD, Miller AM, Rayburn WF. Влияние спонтанных или искусственных родов на рождение макросомного плода. Ам Дж. Перинатол . 1995 12 января (1): 63-6. [Медлайн].
Weeks JW, Pitman T, Spinnato JA 2 место. Макросомия плода: влияет ли дородовое прогнозирование на путь родов и исход родов ?. Am J Obstet Gynecol . 1995 Октябрь 173 (4): 1215-9. [Медлайн].
Фавр Р., Бадер А.М., Нисанд Г.Проспективное исследование по оценке веса плода по окружности конечностей, полученной с помощью трехмерного ультразвукового исследования. Ультразвуковой акушерский гинеколь . 1995 6 августа (2): 140-4. [Медлайн].
Сонг ТБ, Мур Т.Р., Ли Джи, Ким Й.Х., Ким Е.К. Прогнозирование веса плода путем измерения объема бедра с помощью трехмерного ультразвукового исследования. Акушерский гинекол . 2000 августа 96 (2): 157-61. [Медлайн].
Baker PN, Johnson IR, Gowland PA, et al. Оценка веса плода с помощью эхопланарной магнитно-резонансной томографии. Ланцет . 1994 12 марта. 343 (8898): 644-5. [Медлайн].
Kolderup LB, Ларос РК, Musci TJ. Частота стойких родовых травм у макросомных младенцев: связь с способом родов. Am J Obstet Gynecol . 1997 Июль 177 (1): 37-41. [Медлайн].
липопротеинов в Staphylococcus aureus опосредуют воспаление с помощью TLR2 и железозависимый рост in vivo
Abstract
Липопротеины (Lpp) являются лигандами TLR2 и передают сигнал через адаптер MyD88.Как часть оболочки бактериальных клеток, Lpp в основном участвуют в получении питательных веществ для Staphylococcus aureus . Влияние Lpp на активацию TLR2-MyD88 для S. aureus при системной инфекции неизвестно. S. aureus штамм SA113, дефицитный по ферменту, кодируемому геном пролипопротеиндиацилглицерилтрансферазы (Δ lgt ), который присоединяет липидный якорь к про-Lpp, был использован для изучения преимуществ и затрат созревания Lpp. Lpp в S. aureus индуцировал ранние и сильные цитокины с помощью передачи сигналов TLR2-MyD88 в перитонеальных макрофагах мыши.Lpp через TLR2 вносит вклад в патогенез сепсиса у мышей C57BL / 6 с IL-1β, хемокин-опосредованным воспалением и высоким числом бактерий. В отсутствие MyD88-опосредованного воспаления Lpp позволял бактериальному очищению печени, лишенной инфильтрирующих клеток, но все же давал сильное преимущество в росте у мышей, которое, как было показано, зависело от поглощения и хранения железа in vitro и in vivo. Для бактерий, ограниченных железом, преимущество роста, связанного с Lpp, было очевидным при инфицировании мышей MyD88 — / — , но не C57BL / 6.С другой стороны, перегрузка железом хозяина восстанавливала дефицит роста Δ lgt у MyD88 — / — , но не у иммунокомпетентных мышей C57BL / 6. Эти результаты показывают, что усвоение железа улучшается за счет Lpp S. aureus , но этому противодействует воспаление. Таким образом, липидное заякоривание является эволюционным преимуществом для S. aureus , так как оно сохраняет важные белки для лучшего выживания при инфекции.
S Taphylococcus aureus — частая причина опасного для жизни сепсиса.Он выражает несколько факторов вирулентности, которые способствуют его выживанию в организме хозяина и помогают уклоняться от иммунных ответов. Липопротеины стафилококка (Lpp) 3 составляют большое семейство заякоренных в мембране белков. В геноме S. aureus > 50 генов несут сигнальную последовательность типа II, типичную для Lpp, 35 из которых могут быть связаны с известной или предсказанной функцией; многие из них являются частью переносчиков ABC и участвуют в усвоении питательных веществ (1). Преобладающими Lpp являются SitC (2), который является связывающим белком стафилококкового транспортера железа SitABC; PrsA, пептидил-пролил цис — / транс -изомераза, участвующая в укладке белка; и OppA, олигопептидная пермеаза (1).Помимо SitC, несколько охарактеризованных стафилококковых Lpp участвуют в транспорте железа в качестве сидерофорсвязывающих частей переносчиков ABC (FhuD1 / FhuD2, SirA, SstD) (2, 3, 4, 5) и в качестве связывающих белков гемового железа и трансферрина железа (IsdE и HtsA) (6, 7).
Lpp синтезируются как пре-Lpp с N-концевой сигнальной последовательностью, содержащей консервативный C-концевой липобокс. Фрагмент диацилглицерила переносится на инвариантный С-концевой цистеин липопротеиндиацилглицерилтрансферазой (Lgt).Сигнальный пептид про-Lpp отщепляется липопротеиновой сигнальной пептидазой (LspA) (8). В то время как у грамотрицательных бактерий Lpp дополнительно модифицируется в диацилглицерил-цистеине с помощью N -ацилтрансферазы (Lnt), гомолог Lnt не был идентифицирован в S. aureus , что позволяет предположить диацилированный Lpp. N-концевая часть изолированного стафилококкового Lpp была охарактеризована как диацилированный белок (9), тогда как другое исследование нашло доказательства триацилирования Lpp SitC еще неизвестным ферментом (10).
Lpp очищен от нескольких патогенов, включая S. aureus , а также синтетические липопептиды Pam 3 CysSerLys 4 (Pam 3 Cys) (11, 12) и Pam 2 CysSerLys 4 (Pam 2 Cys), отражая три- и диацилированный Lpp бактерий, как известно, активирует TLR2 (10, 13, 14, 15, 16, 17, 18). Три липидные цепи Pam 3 Cys опосредуют гетеродимеризацию рецепторов TLR1 и TLR2 (12, 19). Сигнал диацилированного Lpp с помощью димеров TLR2 / 6 и моделирование кристаллизации предсказали связывание липидных цепей с TLR2 и стабилизацию с помощью TLR6 (19).Активация TLR2 синтетическими липопептидами приводит к индукции цитокинов, хемокинов, адгезинов и нитритов в макрофагах и эпителиальных клетках (11). Сложность распознавания TLR2 становится очевидной, поскольку было показано, что стафилококковые липотейхоевые кислоты (LTA) запускают иммунный ответ (20, 21, 22). Однако Lpp, а не LTA, по-видимому, являются доминирующими стимулами TLR2 у штаммов S. aureus (1, 10, 13, 18, 23).
Инактивация созревания Lpp у S. aureus , Mycobacterium tuberculosis , Listeria monocytogenes , Streptococcus pneumoniae и Bacillus subtilis приводила к снижению роста в условиях стресса или в присутствии фагоцитов in vitro (1 , 23, 24, 25, 26, 27).Более того, было обнаружено, что мутанты с делецией гена S. aureus и стрептококков группы B lgt (Δ lgt ) высвобождают большое количество пре-Lpp, возможно, за счет выделения предшественника Lpp (1, 28). Созревание Lpp необходимо для вирулентности M. tuberculosis и S. pneumoniae in vivo (25, 26, 29). Напротив, мутанты Δ lgt у стрептококков S. aureus и группы B были гипервирулентными in vivo и индуцировали меньше цитокинов in vitro (1, 23, 28).
Передача сигналов TLR2-MyD88, по-видимому, играет важную роль в системной инфекции S.aureus , поскольку мыши TLR2 — / — и MyD88 — / — погибают от инфекции (30). В исследовании описан гипервирулентный фенотип S. aureus Δ lgt при сепсисе (23). Однако еще предстоит выяснить, связан ли этот фенотип с нарушением передачи сигналов TLR2 у хозяина.
Таким образом, мы исследовали, почему созревание Lpp является преимуществом для S. aureus , хотя это созревание приводит к распознаванию TLR2, что приводит к сильному иммунному ответу и контролю инфекции.
Здесь мы показываем, что, с одной стороны, созревание Lpp важно для индукции TLR2-MyD88-опосредованного воспаления in vitro и при системной инфекции. Однако усиленное воспаление, вызванное SA113 дикого типа (wt), связано с более высокой бактериальной нагрузкой в органах инфицированных мышей C57BL / 6, TLR2 — / — и MyD88 — / — . Затем мы обнаруживаем, что, с другой стороны, созревание Lpp является фундаментальным для поглощения железа и тем самым способствует росту S. aureus . Таким образом, Lpp-опосредованный рост и выживаемость S.aureus противодействует MyD88-опосредованному воспалению in vivo.
Материалы и методы
Мыши
Мышей разводили в специальных условиях, свободных от патогенов, в Доме для животных Департамента биомедицины Университетской больницы Базеля в соответствии с положениями швейцарского ветеринарного законодательства. Мыши MyD88 — / — были любезным подарком от W. D. Hardt (ETH Zürich) и TLR2 — / — от W. J. Rieflin (Tularik), все они скрещивались на фоне C57BL / 6 в течение 10 поколений.Мышей умерщвляли CO 2 или i.p. для инъекций 500 мг / кг тиопентала (Abbott Laboratories).
Бактериальные штаммы и условия роста
SA113 wt (Американская коллекция типовых культур 35556), его изогенные мутанты lgt :: ermB (Δ lgt ) и SA113 Δ lgt , дополненный pRB lgt , описанные в подробно ранее (1), были использованы в этом исследовании. Newman wt (31) (собственная коллекция штаммов) и мутант Newman Δ lgt , полученные точно так же, как описано для SA113 (1), использовали для дополнительных экспериментов.
Бактерии выращивали в триптиказо-соевом бульоне в течение 7 часов и пересеивали в течение ночи в тех же условиях. Ночные культуры промывали 0,9% NaCl (Bichsel, Швейцария) и использовали при сепсисе. Каждый посевной материал оценивали по количеству КОЕ / мл на чашках с агаром Мюллера-Хинтона (MHA). Для активации NF-κB и индукции цитокинов бактерии дополнительно субкультивировали в триптиказо-соевом бульоне до конечной логарифмической фазы. При необходимости в соевый бульон с триптиказой добавляли 10 мкг / мл эритромицина.
Рост в условиях обеднения железа проводился, как ранее опубликовано (32), с изменениями.Вкратце, бактерии культивировали в RPMI 1640 с 250 мкМ дипиридила (DIP; Sigma-Aldrich) в течение ночи при 37 ° C без встряхивания, затем субкультивировали при 37 ° C со скоростью 120 об / мин в RPMI, обработанном в течение 1 часа 6% Chelex (Sigma-Aldrich). ) с добавлением двухвалентных катионов. Доступность железа восстанавливали с помощью 20 мкМ FeSO 4 , 20 мкМ FeCl 3 или 10% декстрана железа (Serumwerk Bernburg). КОЕ / мл оценивали путем посева серийных разведений на чашки с MHA.
Для выживания S. aureus в плазме мышиных мышей кровь собирали пункцией сердца в гирудине.Плазма была отделена от заражающих клеток центрифугированием и инфицирована штаммами S. aureus . Рост оценивали путем посева серийных разведений на чашки с MHA.
Получение перитонеальных макрофагов
Мышей вводили i.p. вводят 2 мл 3% тиогликолата (BD Biosciences). Через 3-4 дня клетки перитонеального экссудата собирали из брюшной полости путем промывания 6 мл холодной смеси RPMI 1640 Complete (5% FBS, 2 мМ глутамата, 1 мМ пирувата натрия, 1.5 мМ HEPES, заменимые аминокислоты). Эритроциты лизировали, и 1 × 10 6 клеток / мл высевали либо в 24-луночные планшеты для продукции цитокинов, либо на предметные стекла с 16-луночными камерами для анализов активации NF-κB. Прилипшие макрофаги инфицировали 1 × 10 7 КОЕ / мл жизнеспособных штаммов S. aureus в полной RPMI 1640.
Анализ цитокинов макрофагов
Супернатанты заменяли через 1 час после фагоцитоза на RPMI 1640 complete, содержащий 100 мкг / мл гентамицина.Супернатанты собирали через 2 и 18 ч после заражения, и TNF, IL-1β, IL-6, IL-10 и MIP-2 определяли с помощью ELISA в соответствии с инструкциями от BD Biosciences и для MIP-2 от R&D Systems.
Анализ транслокации NF-κB
Макрофаги стимулировали бактериями. Через 20 и 120 минут внеклеточные бактерии разлагались 20 ед. Лизостафина (Genmedics). Клетки промывали PBS и фиксировали в растворе полилизин-параформальдегида (2,14 г / л периодата, 13,7 г / л лизина, 2% параформальдегида).Клетки инкубировали с поликлональными кроличьими антителами против NF-κB p65 мыши (eBioscience) в PBS, содержащем 2% нормальной козьей сыворотки, и затем инкубировали с биотинилированными козьими антителами против кроличьих антител (Vector Laboratories). Добавляли раствор набора ABC (Vector Laboratories) и визуализировали с помощью раствора AEC (Dako). Ядра контрастировали гематоксилином, клетки помещали в глицериновый желатин Кайзера (Merck), и ядерную локализацию NF-κB анализировали с помощью световой микроскопии.
Модель сепсиса
Сепсис был индуцирован у самок мышей C57BL / 6, TLR2 — / — и MyD88 — / — в возрасте 6–8 недель. S. aureus (от 1 × 10 6 до 1 × 10 8 КОЕ) в 200 мкл 0,9% NaCl вводили в боковую хвостовую вену. Для перегрузки железом мышам вводили i.p. вводят 2 мг декстрана железа (Serumwerk Bernburg) за 3 ч до заражения. После заражения измеряли увеличение веса. Кровь собирали сердечной пункцией в EDTA. Животным вводили перфузию в левый желудочек раствором Рингера; селезенки, печень, обе почки и оба колена собирали и гомогенизировали в объеме 1 мл 0.9% NaCl. Цитокины и хемокины определяли в гомогенатах плазмы и селезенки с помощью ELISA (см. «Анализ цитокинов макрофагов»). Бактерии оценивали в гомогенатах крови и тканей путем серийных разведений в 0,9% NaCl, высеянных на чашки с MHA. Органы фиксировали параформальдегидом, заливали парафином и окрашивали H&E или берлинским синим для окрашивания железа. Инфильтраты, определяемые как скопление> 20 лейкоцитов без центральной некротической области, подсчитывали с помощью световой микроскопии при увеличении × 10 в трех случайно выбранных областях срезов органов, окрашенных H и E.Для криосрезов ткани замораживали и окрашивали иммуногистохимическим методом, как описано ранее (33), анти-CD45, анти-CD11b, анти-BM8 или анти-Gr-1 Abs. Микроскопический анализ выполняли с помощью программного обеспечения для анализа CellP в микроскопе Olympus BX61.
Стимуляция спленоцитов in vitro
Спленоциты инфицированных мышей выделяли через 1 день после инъекции, механически разрушали (размер ячеек 70 мкм) с последующим лизисом эритроцитов и завершением ресуспендирования в RPMI 1640. В 24-луночных планшетах 1 × 10 6 клеток инфицировали в течение 1 ч теми же жизнеспособными бактериями, которые использовались для инфекции, при соотношении бактерий / спленоцитов 1: 1.В течение следующих 18 часов инкубации к клеткам добавляли гентамицин для предотвращения роста бактерий. Супернатанты собирали и цитокины анализировали с помощью ELISA, как это делается для макрофагов (см. «Анализ цитокинов макрофагов»).
Анализ стрептонигрина и
55 Поглощение FeБактерии из ночных культур или ex vivo после гомогенизации почек инфицированных мышей C57BL / 6 подвергали воздействию 2 мкг / мл стрептонигрина (34) при 37 ° C в течение 30 мин. КОЕ / мл оценивали до и после воздействия путем посева серийных разведений на чашки с MHA.
Для экспериментов по поглощению Fe 55 бактерии выращивали в обработанной Chelex минимальной среде M9 (1% казаминокислоты и 5% раствор витамина MEM). Среду обрабатывали 200 мкМ DIP. Бактерии собирали при OD 578 0,8–1,0, дважды промывали транспортной средой M9 (обработанные Chelex соли 1 × M9, 1 мл 1 мМ MgSO 4 , 0,1% глюкозы) при 4 ° C и ресуспендировали. в транспортной среде М9 до OD 578 1.0. После инкубации при 37 ° C в течение 5 мин при встряхивании 55 Fe, восстановленный аскорбатом (100 мкМ (5.5 кБк / мл) / 100 мМ аскорбата) разбавляли в бактериальной суспензии в 100 раз. Образцы отбирали в указанное время, фильтровали на смешанных целлюлозных мембранных фильтрах GN-6 Metricel (Pall), дважды промывали 0,1 М раствором LiCl, сушили и считали с помощью жидкостного сцинтилляционного счетчика.
Статистический анализ
На графиках Каплана-Мейера для сравнения выживаемости при сепсисе среди инфицированных мышей использовали лог-ранговый тест. Двусторонний тест ANOVA использовали для статистического анализа цитокинов и NF-κB в макрофагах in vitro и для потери веса мышей после заражения.КОЕ в органах, цитокины в плазме или анализы рестимуляции спленоцитов анализировали с помощью Манна-Уитни. Статистический анализ был выполнен с помощью Prism 5.0a (программное обеспечение GraphPad). Значение p <0,05 считалось статистически значимым.
Результаты
Lpp вызывают воспаление через TLR2 и MyD88 in vitro
Мы сначала исследовали влияние Lpp на индукцию выбранных продуктов макрофагов, которые участвуют в патогенезе сепсиса (35). Поэтому мы инфицировали перитонеальные макрофаги мышей C57BL / 6 и TLR2 — / — жизнеспособными S.aureus SA113 wt и Δ lgt для оценки активации NF-κB, цитокинов и уровней MIP-2. SA113 wt, но не Δ lgt индуцировал NF-κB в макрофагах C57BL / 6 через 20 минут, тогда как инфицированные клетки TLR2 — / — не проявляли активации до 2 часов (рис. 1⇓ A ). TNF, IL-10, IL-6 и MIP-2 индуцировались SA113 wt через передачу сигналов TLR2 сразу после заражения (2 ч). Эти цитокины отсутствовали или были в меньшей степени индуцированы SA113 Δ lgt и / или в отсутствие TLR2 (рис.1⇓, B – D и F ). Через 2 часа IL-1β не обнаруживался (рис. 1⇓ E ). Поздно после инфицирования (18 ч) уровни TNF оставались высокими в присутствии Lpp; однако в их отсутствие TNF превышал уровень (рис. 1⇓ B ). IL-10, IL-6 и IL-1β оставались частично Lpp-TLR2-зависимыми, тогда как MIP-2 оставался независимым (рис. 1– C – F ). Результаты, полученные с SA113 Δ lgt -pRB lgt в клетках C57BL / 6, были сопоставимы с результатами для SA113 wt (рис.1⇓).
РИСУНОК 1.Lpp на жизнеспособном S. aureus индуцируют цитокины в макрофагах через TLR2. Перитонеальные макрофаги от мышей C57BL / 6 и TLR2 — / — были инфицированы SA113 wt (заполненные и полосатые столбцы соответственно), Δ lgt (пустые и пунктирные столбцы соответственно) и дополненным Δ lgt -pRB lgt (серые столбцы, только в C57BL / 6) при множественности инфицирования 10. A , Процент NF-κB-положительных клеток после 20 и 120 минут заражения. B – F , супернатанты анализировали с помощью ELISA на ( B ) TNF, ( C ) IL-10, ( D ) IL-6, ( E ) IL-1β и ( F). ) MIP-2 через 2 и 18 ч заражения. †, не обнаруживается. Данные представляют собой средние значения ± стандартное отклонение трех или более экспериментов. Существенные различия инфицированных wt макрофагов C57BL / 6 по сравнению со всеми другими группами указаны следующим образом: *, p <0,05; ∗∗, p <0,01; ∗∗∗, p <0,001.
Высвобождение цитокинов требовало MyD88-опосредованных сигналов в макрофагах, поскольку ни один из штаммов не индуцировал цитокины в макрофагах MyD88 — / — (данные не показаны).Взятые вместе, эти данные показывают, что Lpp SA113 индуцирует воспалительные медиаторы, которые имеют отношение к сепсису, посредством передачи сигналов TLR2-MyD88.
Стафилококковые Lpp увеличивают рост бактерий и воспалительную реакцию in vivo
Недавно Bubeck Wardenburg et al. показали лучшую выживаемость мышей C57BL / 6, инфицированных 5 × 10 6 КОЕ S. aureus Newman wt, чем мутантом Δ lgt . Авторы объяснили этот ответ более сильным иммунным ответом у S.aureus wt по сравнению с мышами, инфицированными Δ lgt (23). В наших исследованиях выживаемости с S. aureus SA113, 10 8 КОЕ приводили к аналогичной смерти у 20% мышей, инфицированных wt ( n = 9) и 25% у мышей, инфицированных Δ lgt ( n = 9) через 9 дней.
Затем мы исследовали роль Lpp в S. aureus на росте in vivo. Мыши C57BL / 6 были инфицированы 1 × 10 7 КОЕ SA113 wt и Δ lgt и показали аналогичную потерю веса в течение 9 дней заражения (рис.2⇓ А ). Через 1 день бактериальная нагрузка SA113 wt и Δ lgt была одинаковой во всех исследованных органах мышей C57BL / 6 (данные не показаны). Однако на 9 день после инфицирования бактериальная нагрузка SA113 wt в селезенке, почках и коленях мышей C57BL / 6 была значительно выше по сравнению с Δ lgt (рис. 2 B ), тогда как бактериальная нагрузка в печени была немного, хотя и незначительно, выше после заражения SA113 wt (рис. 2⇓ B ).
РИСУНОК 2.Lpp усиливают вирулентность S. aureus при сепсисе. Мыши A – F , C57BL / 6, инфицированные 1 × 10 7 КОЕ SA113 wt (заполнены) и SA113 Δ lgt (открытые). A , потеря веса мышей во время инфекции и ( B ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях на 9 день. C , плазменный уровень IL-6 1 и 9 дней после заражения. D . Через день после заражения изолированные спленоциты рестимулировали соответствующим штаммом в течение 24 часов in vitro.TNF, IL-6, IL-10, IL-1β и MIP-2 в супернатантах анализировали с помощью ELISA. E , Иммуногистохимия против CD45 и против BM8 замороженных срезов печени мышей, инфицированных SA113 wt. F и G , Гистологический анализ срезов печени, окрашенных H и E, после инфицирования SA113 wt и Δ lgt на 9 день и количественная оценка абсцессов и инфильтратов с помощью микроскопии. Данные представлены в виде средних значений ± стандартное отклонение для потери веса и цитокинов и медианы для КОЕ / мг и количества в срезах печени по крайней мере шести мышей на группу.Значимые различия между wt и Δ lgt указаны следующим образом: *, p <0,05; ∗∗, p <0,01.
Эти данные предполагают, что S. aureus Lpp полезны для роста бактерий в органах во время хронической инфекции.
Мы дополнительно изучили, связан ли рост бактерий с индукцией медиаторов воспаления Lpp in vivo. Поэтому мы определили цитокины в плазме мышей C57BL / 6 после инфицирования SA113 wt и Δ lgt .Через 1 час заражения уровни IL-6 были выше у SA113 wt по сравнению с Δ lgt (среднее ± стандартное отклонение, 671 ± 251 и 95 ± 164 пг / мл; p <0,05), но разница исчезла 4 ч после заражения (SA113 wt 450 ± 431 пг / мл против Δ lgt 234 ± 405 пг / мл). Через день после заражения TNF, IL-10, IL-1β и MIP-2 не обнаруживались в плазме (данные не показаны). Уровни IL-6 в плазме были сопоставимы после заражения SA113 wt и Δ lgt и значительно повысились до аналогичных уровней через 9 дней (рис.2⇑ C ), что указывает на отсутствие системной регуляции цитокинов стафилококковым Lpp в SA113. Для изучения органоспецифической индукции цитокинов, опосредованных Lpp, спленоциты выделяли через 1 день после инфицирования и рестимулировали в течение 24 часов in vitro тем же штаммом, который использовался in vivo. Оба штамма индуцировали все медиаторы в одинаковой степени, за исключением IL-1β, который продуцировался исключительно в ответ на SA113 wt (рис. 2⇑ D ), что позволяет предположить, что Lpp опосредует локальную регуляцию цитокинов в инфицированных органах.
Чтобы оценить важность стафилококкового Lpp в персистирующей тканевой инфекции, мы подсчитали абсцессы и инфильтраты в срезах печени мышей C57BL / 6, инфицированных wt и Δ lgt , на 9 день. Инфильтраты после инфицирования SA113 wt или Δ . lgt состояли из лейкоцитов CD45 + , макрофагов BM8 + (рис. 2⇑ E ) и моноцитов CD11b + , тогда как гранулоциты Gr-1 + рекрутировались редко (данные не показаны).Однако после инфицирования SA113 wt было обнаружено больше инфильтратов и абсцессов, чем Δ lgt (рис. 2, F и G ). Увеличение инфильтратов моноцитов в печени, инфицированной SA113 wt, коррелировало с повышенными уровнями MIP-2 (81 ± 32 против 41 ± 19 пг / мл, p <0,001) и KC (968 ± 454 против 617 ± 153 пг / мл, p <0,05) в селезенке после инфицирования SA113 wt по сравнению с Δ lgt на 9 день. Таким образом, зрелый Lpp в S. aureus обеспечивал лучший рост органов и вызывал раннее более сильное высвобождение IL-1β и индуцированные поздние хемокины, сопровождающиеся усилением моноцитарной инфильтрации.Результаты показывают, что Lpp поддерживает рост и выживаемость S. aureus , несмотря на то, что вызывает иммунный ответ.
Lpp обладают TLR2- и MyD88-независимым эффектом на рост бактерий in vivo.
Наши данные по цитокинам показали роль TLR2 в обнаружении стафилококкового Lpp после стимуляции макрофагов in vitro. Чтобы изучить вклад TLR2 в врожденный ответ in vivo, мы инфицировали мышей TLR2 — / — SA113 wt и Δ lgt . SA113 wt вызывал более высокую потерю веса (28%), чем Δ lgt (16%) у инфицированных мышей (рис.3⇓ А ). Бактериальная нагрузка в органах TLR2 — / — была выше после инфицирования SA113 wt, чем с Δ lgt (рис. 3⇓ B ). SA113 wt и Δ lgt не индуцировали TNF, IL-10, IL-1β и MIP-2 в плазме мышей, а IL-6 не зависел от Lpp (данные не показаны). В рестимулированных спленоцитах инфицированных мышей TLR2 — / — TNF, IL-6, IL-10, IL-1β и MIP-2 не зависели от стафилококкового Lpp и были ниже, чем в рестимулированных спленоцитах мышей C57BL / 6 (рис.3⇓ C ). По сравнению с мышами C57BL / 6 (рис. 2⇑, A, и B ), мыши TLR2 — / — показали значительно большую потерю веса (рис. 3⇓ A ) и значительно более высокую бактериальную нагрузку в почках и почках. колени (рис.3 B ) после заражения SA113 wt ( p <0,001 и p <0,01 соответственно), но не после заражения Δ lgt .
РИСУНОК 3.TLR2 обнаружение Lpp оказывает незначительное влияние на S.aureus in vivo. A , Потеря веса и ( B ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях у мышей TLR2 — / — после инфицирования 1 × 10 7 SA113 wt (заполнено) и SA113 Δ lgt (открыто) на 9 день. C . Через день после инфицирования изолированные спленоциты рестимулировали соответствующим штаммом в течение 24 часов in vitro. TNF, IL-6, IL-10, IL-1β и MIP-2 в супернатантах анализировали с помощью ELISA. D , Потеря веса и ( E ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях у мышей MyD88 — / — с 1 × 10 6 SA113 wt (заполнено) и SA113 Δ lgt (открыто) в 9 день. F . Через день после заражения изолированные спленоциты рестимулировали соответствующим штаммом в течение 24 часов in vitro. TNF, IL-6, IL-10, IL-1β и MIP-2 в супернатантах анализировали с помощью ELISA. G , Гистологический анализ срезов печени, окрашенных H и E, через 9 дней после заражения. Данные для мышей TLR2 — / — и MyD88 — / — представлены как средние значения ± стандартное отклонение для потери веса и цитокинов и медианы для КОЕ / мг от пяти до шести мышей на группу. Значимые различия между wt и Δ lgt обозначены следующим образом: *, p <0.05; ∗∗, p <0,01; ∗∗∗, p <0,001.
MyD88 — / — Мыши не имеют воспалительной реакции макрофагов и погибают от инфекций, вызванных стафилококками (30). Поскольку Lpp S. aureus усиливал рост и воспаление у системно инфицированных мышей C57BL / 6, мы использовали MyD88 — / — мышей, чтобы избежать сильного воспаления, и проанализировали Lpp-опосредованный рост S. aureus . Зараженные мыши MyD88 — / — медленно теряли в весе, но потеря веса была более серьезной с SA113 wt, чем с Δ lgt (рис.3⇑ D ). Бактериальная нагрузка мышей, инфицированных SA113 wt, была выше во всех органах, чем Δ lgt (рис. 3⇑ E ). Интересно, что бактериальная нагрузка в печени была резко снижена по сравнению с мышами C57BL / 6 и TLR2 — / — , в то время как во всех других органах было увеличено количество бактерий. Рестимулированные спленоциты мышей MyD88 — / — не индуцировали или вызывали очень низкие уровни TNF, IL-6, IL-10, IL-1β и MIP-2 независимо от стафилококкового Lpp (рис. 3– F ).В соответствии с отсутствием цитокинов и хемокинов (данные не показаны), в печени не было гранулем (рис. 3⇑ G ). Таким образом, отсутствие воспаления сопровождалось не только увеличением бактериальной нагрузки, но и атипичным распределением S. aureus по органам. Однако, как суммировано в Таблице I⇓, Lpp обеспечивает лучший рост бактерий независимо от передачи сигналов TLR2 и MyD88 хозяина.
Таблица I.Рейтинг заболеваний для иллюстрации влияния стафилококкового Lpp и железа на потерю веса и бактериальную нагрузку у мышей C57BL / 6, TLR2 — / — и MyD88 — / — a
Созревание Lpp требуется для
S.aureus рост и поглощение железа in vitro и ex vivoМы показали, что различия в росте SA113 wt и Δ lgt in vivo не зависели от иммунных ответов, опосредованных Lpp-TLR2 —. Поскольку многие S. aureus Lpp участвуют в транспорте железа (1, 36), мы исследовали роль созревания Lpp в приобретении и росте железа in vitro. Как и ожидалось, ни SA113 wt, ни Δ lgt не росли в течение 48 часов в обедненном железом RPMI 1640 (не показано).Добавление в обедненный железом RPMI 1640 с FeSO 4 , FeCl 3 и декстраном железа обеспечило лучший рост SA113 wt, чем Δ lgt (рис. 4 A ). SA113 Δ lgt погиб в среде RPMI 1640 с добавкой FeSO 4 и рос хуже, чем вес в присутствии FeCl 3 или декстрана железа (рис. 4 A ). Это говорит о пользе экспрессии Lgt для роста и максимального усвоения железа.
РИСУНОК 4.Lpp улучшает получение железа в S.aureus . SA113 wt (заполненный) и Δ lgt (открытый) выращивали в обедненном железом RPMI 1640 с добавлением ( A ) 20 мкМ FeSO 4 (кружки), 20 мкМ FeCl 3 (квадраты) или 10 % декстрана железа (алмазы). B , Рост SA113 wt (заполненный) и Δ lgt (открытый) в плазме мышей C57BL / 6. C , чувствительность к стрептонигрину в течение 30 минут для SA113 wt и Δ lgt , выращенных в течение ночи в RPMI 1640 с 250 мкМ DIP. D , 55 Поглощение Fe SA113 wt (заполненный) и Δ lgt (открытый) в среде M9 с (черный) или без (серый) 200 мкМ DIP. E , Чувствительность к стрептонигрину в течение 30 минут объединенного SA113 wt и Δ lgt , выделенного из почек инфицированных мышей C57BL / 6. Данные показывают одного представителя из трех экспериментов.
Поскольку трансферрин насыщен на 60% у мышей (37) и помимо гема служит источником железа для S. aureus (7), мы исследовали, улучшает ли созревание Lpp использование железа, связанного с трансферрином, с использованием плазмы в качестве среды для роста. . Мы не обнаружили различий в росте SA113 wt и Δ lgt в плазме (рис.4⇑ B ).
Чтобы подтвердить, что улучшенный рост SA113 wt является следствием лучшего поглощения железа, мы использовали железозависимый антибиотик стрептонигрин. SA113 wt, выращенный в условиях ограниченного количества железа в RPMI-DIP, был более чувствителен к стрептонигрину и показал в 2,5 раза более высокое поглощение Fe 55 , чем Δ lgt или неограниченные штаммы SA113, что указывает на то, что Lpp улучшает поглощение железа (рис. 4⇑). , C и D ). Lpp также влиял на поглощение железа во время инфекции, поскольку ex vivo SA113 wt из почек C57BL / 6 был более чувствителен к стрептонигрину, чем Δ lgt (рис.4⇑ E ). Взятые вместе, наши результаты показывают, что созревание Lpp является предпосылкой для максимального усвоения железа S. aureus in vitro и in vivo.
Lpp обеспечивают рост бактерий, ограниченных железом.
S. aureus у мышей MyD88 — / —Усиление Lpp 55 Поглощение Fe S. aureus после ограничения железа in vitro, что свидетельствует об усилении регуляции фермента. Lpp в этих условиях. Таким образом, мы ожидали Lpp-опосредованного роста SA113 wt in vivo.Перед инфицированием мы культивировали SA113 wt и Δ lgt в условиях ограничения железа. Ограничение железа SA113 wt и Δ lgt вызывало временную потерю веса 10% и полное восстановление веса мышей C57BL / 6 на 9 день (фиг. 5⇓ A ). Снижение внутрибактериального железа устраняет различия между SA113 wt и Δ lgt в бактериальной нагрузке (рис. 5⇓ B ), а также в инфильтратах и абсцессах печени (данные не показаны). Это указывает на то, что Lpp-зависимое преимущество SA113 wt у мышей C57BL / 6 (рис.2⇑) не было связано с приобретением железа в иммунокомпетентном хозяине.
РИСУНОК 5.Истощение запасов железа увеличивает Lpp-опосредованную вирулентность S . aureus у мышей MyD88 — / — . A , Потеря веса и ( B ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях после инфицирования мышей C57BL / 6 4 × 10 7 КОЕ лишенного железа SA113 wt (заполнено) и Δ lgt (открыто) на 9 день. C , Потеря веса и ( D ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях после инфицирования мышей MyD88 — / — 1 × 10 6 КОЕ лишенного железа SA113 wt (заполненный) и Δ lgt (открытый) на 9 день.Данные представлены в виде среднего значения ± стандартное отклонение для потери веса и медианы для КОЕ / мг не менее четырех мышей на группу. Значимые различия между wt и Δ lgt указаны следующим образом: *, p <0,05; ∗∗, p <0,01; ∗∗∗, p <0,001.
Мы предположили, что ограниченный железом SA113 wt не может расти выше Δ lgt у мышей C57BL / 6 из-за ограничения железа, вызванного сильным MyD88-зависимым воспалением. Действительно, бактериальная нагрузка ограниченного железом SA113 wt была выше, чем Δ lgt после инфицирования мышей MyD88 — / — (рис.5⇑ D ). Мыши MyD88 — / — , инфицированные SA113 wt, показали потерю веса 11%, а мыши, инфицированные Δ lgt , сохранили вес (фиг. 5: C ). Инфильтраты и абсцессы в печени у обоих штаммов были редкими (не показаны). Эти результаты сведены в Таблицу I⇑. Они предполагают, что для стимулирующего рост эффекта Lpp S. aureus необходимо железо до инфицирования, чтобы обеспечить немедленную доступность железа в организме хозяина, который эффективно секвестрирует железо с помощью MyD88-зависимого механизма.
Lpp незаменимы для роста бактерий у перегруженных железом мышей MyD88
— / —Далее мы исследовали, устраняет ли перегрузка железом дефект роста SA113 Δ lgt в присутствии или в отсутствие передачи сигналов MyD88. Поэтому мы вводили декстран железа мышам C57BL / 6 и MyD88 — / — перед заражением штаммами SA113, выращенными в нормальных условиях. Во время инфицирования SA113 wt вызывал 27% -ную потерю веса у перегруженных железом мышей C57BL / 6, тогда как Δ lgt вызывал временную потерю веса на 20% и восстановление веса после 5-го дня (рис.6⇓ А ). Перегрузка железом не увеличивала вирулентность Δ lgt по сравнению с wt, о чем свидетельствует более низкая бактериальная нагрузка во всех органах и меньшее количество абсцессов в печени после инфицирования мышей C57BL / 6 (фиг. 6⇓, B – D ). Хотя SA113 Δ lgt вызывал такое же количество инфильтратов, как и wt (рис. 6 C ), преимущество Lpp наблюдалось по накоплению железа в инфильтратах после заражения SA113 wt (рис. 6 D ).
РИСУНОК 6.Избыток железа увеличивает вирулентность на S.aureus Δ lgt у мышей MyD88 — / — . A – D , Заражение перегруженных железом мышей C57BL / 6 4 × 10 7 КОЕ SA113 wt (заполнено) и Δ lgt (открыто). A , Потеря веса и ( B ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях на 9-й день. C , Количество абсцессов и инфильтратов срезов печени, окрашенных H и E, после заражения SA113 wt и Δ lgt на 9 день оценивали с помощью микроскопии. D , Окрашивание железом срезов печени после заражения SA113 wt и Δ lgt на день 9. E и F , Заражение перегруженных железом мышей MyD88 — / — 1 × 10 6 КОЕ SA113 wt (заполнено) и Δ lgt (открыто). A , Потеря веса и ( B ) бактериальная нагрузка в селезенке, печени, обеих почках и обоих коленях на 9-й день после инфицирования. Данные представлены в виде среднего значения ± стандартное отклонение для потери веса и цитокинов и медианы для КОЕ / мг и числа в срезах печени по крайней мере четырех мышей на группу.Значимые различия между wt и Δ lgt указаны следующим образом: *, p <0,05; ∗∗, p <0,01; ∗∗∗, p <0,001.
Напротив, у перегруженных железом мышей MyD88 — / — , SA113 wt и Δ lgt вызывали медленное прогрессирование потери веса до ~ 17% на 9 день (рис. 6⇑ E ). Количество бактерий SA113 Δ lgt достигло таких же уровней, как и у wt, во всех органах инфицированных мышей (рис. 6⇑ E ).Эти результаты суммированы в таблице I⇑ и показывают, что при высоких уровнях железа созревание Lpp необходимо для роста хозяина с дефицитом MyD88, но необходимо для роста в органах иммунокомпетентного хозяина.
Lpp в штамме Newman действуют провоспалительным образом и усиливают вирулентность у мышей MyD88
— / —Bubeck Wardenburg et al. наблюдали снижение вирулентности S. aureus Newman wt по сравнению с Δ lgt , что они объяснили уходом от защиты хозяина (23).Мы хотели оценить, был ли эффект Lpp, способствующий росту и усилению вирулентности, наблюдаемый с SA113, также наблюдался у штамма Newman, продуцирующего стафилококковый энтеротоксин B (38). С Newman wt и его изогенным мутантом Δ lgt мы обнаружили макрофаги, активированные Lpp- и TLR2-зависимым образом (данные не показаны). Результаты были аналогичны результатам, полученным с SA113 и опубликованным ранее, с убитыми нагреванием Newman wt и Δ lgt (23). Системное инфицирование 1 × 10 8 КОЕ Newman wt и Δ lgt привело к 100% и 20% гибели мышей C57BL / 6 в течение 24 часов, соответственно; однако все мыши, инфицированные Δ lgt , погибли в течение 32 часов (дополнительный рис.1 А ). 4 При несмертельном инокуляте 1 × 10 6 КОЕ на мышь Newman wt вызывал более серьезную потерю веса, чем Δ lgt (дополнительный рис. 1 B ). Newman wt вызывал более высокую бактериальную нагрузку в печени и почках и индуцировал больше IL-6 в плазме мышей, чем Δ lgt (дополнительная фигура 1, C и D ). В рестимулированных спленоцитах, выделенных через 1 день после инфицирования мышей C57BL / 6, TNF, IL-6, IL-10, IL-1β и MIP-2 частично или полностью зависели от стафилококкового Lpp (дополнительный рис.1 E ).
Чтобы проверить, может ли Lpp усиливать рост в отсутствие воспаления, мы заразили мышей MyD88 — / — с Newman wt и Δ lgt . Мыши MyD88 — / — , инфицированные Newman wt, умерли значительно раньше, а выжившие мыши потеряли больше веса, чем животные, инфицированные Δ lgt (дополнительная фигура 2, F и G ). Кроме того, после выращивания на обедненной железом среде мы наблюдали более высокую чувствительность к стрептонигрину Newman wt, чем Δ lgt in vitro (дополнительный рис.1 H ), что свидетельствует об улучшении усвоения железа Lpp в переносчиках ABC, как это наблюдается для штаммов SA113. Наши данные показывают, что Lpp обоих штаммов S. aureus , Newman и SA113, вызывают не только TLR2-опосредованный иммунный ответ, но также оказывают стимулирующее действие на рост.
Обсуждение
Мы оценили функцию зрелого Lpp в стафилококковом иммунопатогенезе. Используя низковирулентный штамм SA113 S. aureus , мы показали, что воспаление in vitro и у иммунокомпетентного хозяина in vivo зависит от Lpp.С одной стороны, мы показываем, что зрелые Lpp вызывают сильный цитокиновый и хемокиновый ответ за счет ранней активации TLR2. Lpp поздно активирует другие сигнальные рецепторы MyD88 (например, через IL-1β и IL-1R), что необходимо для мощной противомикробной защиты хозяина. С другой стороны, наши данные показывают, что зрелый Lpp обеспечивает лучший рост за счет приобретения железа. Доступность железа до инфицирования обеспечивала Lpp-опосредованное преимущество роста S. aureus в отсутствие передачи сигналов MyD88, которому противодействовало воспаление у иммунокомпетентных мышей C57BL / 6.Однако это преимущество было аннулировано, когда во время инфекции было доступно дополнительное железо, что позволяет предположить, что хозяин с дефицитом MyD88 не мог эффективно ограничивать железо.
Наше исследование показывает, что Lpp ответственны за раннюю индукцию цитокинов жизнеспособным S. aureus и усиливают позднее опосредованное IL-1β воспаление в макрофагах за счет передачи сигналов TLR2 in vitro. Наши данные согласуются с предыдущими сообщениями, показывающими, что Lpp жизнеспособного SA113 wt и убитого нагреванием Newman wt индуцируют цитокины в клеточной линии моноцитов и макрофагах, соответственно (1, 23).Самая сильная зависимость Lpp-TLR2 была обнаружена для IL-1β, который участвует в рекрутировании нейтрофилов и, следовательно, в уничтожении S. aureus при кожной инфекции (39). Поскольку мы обнаружили, что активация макрофагов полностью зависит от MyD88, S. aureus использует TLR2 в основном как ранний рецептор, а другие рецепторы, такие как IL-1R (40) и, возможно, TLR9, участвуют поздно (30, 41). Кроме того, мы подтвердили исключительную активацию TLR2 Lpp, а не LTA в жизнеспособном SA113, что уже наблюдалось после очистки стафилококкового LTA и Lpp (10, 13, 18).
TNF, IL-6, IL-10, IL-1β и MIP-2 являются важными медиаторами сепсиса (35) и были обнаружены в нашей модели системной инфекции. Используя SA113 wt, мы обнаружили, что Lpp индуцирует продукцию IL-1β в селезенке, последовательно задерживает хемокины в селезенке и сильное привлечение моноцитов в печень с образованием гранулем. IL-6 и другие цитокины в плазме не зависели от Lpp, за исключением очень ранней временной точки, равной 1 часу после инфицирования, что указывает на местную регуляцию воспаления с помощью Lpp.Локальная регуляция с помощью Lpp также может объяснять сравнимое течение болезни после инфицирования либо SA113 wt, либо Δ lgt . Интересно, что воспаление, связанное с Lpp, было связано с более высокой бактериальной нагрузкой в органах, демонстрируя, что в SA113 созревание Lpp участвует в сохранении инфекции S. aureus . Lpp у штамма Newman аналогичным образом способствовал вирулентности, которая проявлялась в более сильной потере веса, выраженном воспалении в селезенке, острой фазе реакции в плазме и преимуществе роста в печени и почках.Наши результаты не могут подтвердить ранее наблюдаемую повышенную вирулентность Δ lgt (23). Несоответствие может происходить из-за способа нанесения или из-за другого поколения мутантов Newman Δ lgt .
Насколько нам известно, исследование, посвященное вопросу о том, опосредуется ли ответ хозяина при системной инфекции стафилококковым Lpp или TLR2, отсутствует. На мышах TLR2 — / — мы показали, что распознавание стафилококкового Lpp с помощью TLR2 вызывает воспаление, ограничивает рост бактерий и улучшает восстановление веса у мышей, инфицированных SA113 wt.Заражение SA113 Δ lgt полностью не зависело от TLR2. В согласии с Takeuchi et al. (30) мы обнаружили, что передача сигналов Lpp-TLR2 ограничивает бактериальную нагрузку только в почках, но не в селезенке и печени. Мы предлагаем органоспецифический TLR2-опосредованный ответ, но в целом умеренную роль передачи сигналов TLR2 в сепсисе S. aureus , как ранее было опубликовано для стафилококковых кожных инфекций (40).
В соответствии с предыдущими исследованиями, S. aureus wt вызывал отчетливые клинические результаты у мышей MyD88 — / — с отсроченным появлением тяжелых симптомов и более высокой бактериальной нагрузкой по сравнению с мышами C57BL / 6 (30, 40).Неожиданно оказалось, что у мышей MyD88 — / — SA113 и Newman Δ lgt все еще вызывали более легкое заболевание, чем wt, за исключением вклада других Lpp-TLR2-независимых цитокинов в более низкую бактериальную нагрузку Δ lgt . В то время как в большинстве органов отсутствие MyD88 способствовало росту бактерий, количество бактерий SA113 было низким, а гранулемы отсутствовали в печени мышей MyD88 — / — . Нарушение индукции хемокинов и инфильтрация лейкоцитов, возможно, предотвратили скрытие бактерий у мышей MyD88 — / — и способствовали уничтожению бактерий в печени.Напротив, отсутствие MyD88 привело к гораздо более сильному росту бактерий в почках и коленях, что позволяет предположить, что эффекты, вызванные MyD88, защищают хозяина в почках, но не в печени.
Однако количество бактерий SA113 wt в печени и других органах мышей MyD88 — / — было выше, чем у Δ lgt , что указывает на Lpp-опосредованное преимущество роста SA113 wt у хозяина с ослабленным воспалением. . Печень является основным органом хранения железа и переработки гема. S. aureus предпочтительно использует железо из гема переносчиками Hts и Isd, которые содержат Lpp (7). В нашем мутанте SA113 Δ lgt эти переносчики состоят из незрелых Lpp HtsA и IsdE, функция которых может быть нарушена. Следовательно, можно ожидать, что незрелые HtsA и IsdE способствуют снижению бактериальной нагрузки в печени мышей MyD88 — / — . Соответственно, ранее было показано, что делеционный мутант в гене htsB связан с уменьшением образования абсцесса и количества бактерий в печени (7).В соответствии с этими результатами мы экспериментально подтвердили, что дефицит Lpp замедляет рост мутантов S. aureus Δ lgt в условиях ограничения железа in vitro, а также снижает чувствительность к стрептонигрину in vitro и ex vivo из-за пониженного содержания железа ( 55 ). Fe) приобретение. Наш вывод подкрепляется тем фактом, что созревание Lpp улучшает поглощение железа в S. pneumoniae , о чем свидетельствует более высокая чувствительность к стрептонигрину Δ lspA , чем у wt (26).Снижение поглощения железа, вероятно, также объясняет повышенную чувствительность к стрептонигрину, которую мы наблюдали с Newman Δ lgt , и может объяснять дефект роста мутанта Newman Δ lgt в цельной крови и макрофагах, описанный ранее (23).
На основании этих результатов мы заключаем, что дефицит роста S. aureus Δ lgt в органах C57BL / 6, TLR2 — / — , и особенно у мышей MyD88 — / — с нарушенным воспалением, был из-за пониженного поглощения и хранения железа.Подтверждение нашей гипотезы было предоставлено демонстрацией того, что перегрузка железом у мышей MyD88 — / — обращала вспять дефект роста Δ lgt . И SA113 wt, и Δ lgt стали более вирулентными, но бактериальная нагрузка увеличивалась только при инфицировании Δ lgt выше уровня, достигнутого у мышей MyD88 — / — при нормальной диете. Однако мы не можем исключить, что иммунокомпетентный хозяин может влиять на рост Δ lgt за счет снижения доступности других питательных веществ, таких как олигопептиды, Cu или Zn (42).Кроме того, связанное с Lpp поглощение других питательных веществ и металлов (1) может способствовать росту Δ lgt у мышей MyD88 — / — in vivo.
Ранее нами был обнаружен предшественник Lpp, накопленный в супернатанте SA113 Δ lgt in vitro (1), что указывает на менее сильное закрепление или LspA-независимый процессинг предшественника Lpp в SA113 Δ lgt . Высокие концентрации железа, вероятно, насыщали системы захвата железа Δ lgt перед высвобождением про-Lpp и, таким образом, обеспечивали лучший рост Δ lgt в хозяине с дефицитом MyD88.
Иммунокомпетентные мыши C57BL / 6 могут использовать множество путей секвестрации железа в макрофагах после инфицирования. Цитокины увеличивают запасы железа, индуцируя синтез ферритина и эритрофагоцитоз за счет усиления регуляции импортера железа и восстановления молекул экспортера железа (43). Дополнительное зависимое от воспаления ограничение железа достигается увеличением насыщения трансферрина, а также высвобождением липокалина и лактоферрина из гранул PMN (43, 44). Все эти механизмы могут объяснить, что только S.aureus wt, но не Δ lgt , смог извлечь выгоду из перегрузки железом у мышей C57BL / 6. Конкуренция за железо между хозяином и патогеном была дополнительно продемонстрирована инфекцией ограниченным железом S. aureus . SA113 wt без внутриклеточного запасенного железа не получил преимущества от Lpp для роста в органах после инфицирования иммунокомпетентного хозяина. Это говорит о том, что S. aureus должен накапливать железо до заражения, чтобы расти в органах и противостоять уничтожению PMN in vivo.Однако в отсутствие MyD88 лишенный железа SA113 wt выигрывал в гонке за железом над хозяином, а Lpp давал преимущество в росте во время инфекции. В отсутствие воспаления задержка клеточного железа, вероятно, была недостаточной; например, гепсидин, молекула, удерживающая железо в печени, был низким, как опубликовано для S. pneumoniae (45). В этих условиях Lpp разрешил рост S. aureus во время инфицирования.
В заключение мы продемонстрировали, что, с одной стороны, распознавание Lpp с помощью передачи сигналов TLR2-MyD88 контролирует системную инфекцию и органоспецифический ранний иммунитет у хозяина.С другой стороны, как обобщено в таблице I⇑ по общим рангам заболевания, мы показали, что Lpp SA113 wt усиливают рост 1) у необработанных мышей C57BL / 6, TLR2 — / — и MyD88 — / — , 2) обедненных железом стафилококков в отсутствие MyD88-опосредованного воспаления и 3) у перегруженных железом иммунокомпетентных мышей C57BL / 6. Следовательно, эволюция может отобрать S. aureus с липидированным Lpp.
Благодарности
Мы благодарим доктора Вольфа-Дитриха Хардта (ETH Zürich) за предоставление мышей MyD88 — / — и W.J. Rieflin (Tularik) для мышей TLR2 — / — . Мы благодарим доктора Луиджи Террачано за помощь с патологическим анализом и доктора Клауса Хантке за помощь в экспериментах по поглощению 55 Fe. Мы также благодарим доктора Дженнаро Де Либеро и доктора Терезу Ресинк за обсуждение и критическое прочтение рукописи, а также за полезные обсуждения.
Раскрытие информации
У авторов нет финансового конфликта интересов.
Сноски
Затраты на публикацию этой статьи были частично покрыты за счет оплаты страницы.Таким образом, данная статья должна быть помечена как реклама в соответствии с 18 U.S.C. Раздел 1734 исключительно для указания этого факта.
№1 Работа поддержана Швейцарским национальным научным фондом, гранты №№. 3100A0-104259 / 1 и / 2 и 3100A0-120617, а также Deutsche Forschungsgemeinschaft Sonderforschungsbereich 766.
↵2 Адрес для переписки и перепечатки запросов на перепечатку д-ру Регине Ландманн, Отдел биомедицины, Отделение университетской инфекционной биологии , Hebelstrasse 20, CH-4031 Базель, Швейцария.Адрес электронной почты: regine.landmann {at} unibas.ch
№3 В данной статье используются сокращения: Lpp, липопротеин; ДИП, дипридил; lgt , ген пролипопротеиндиацилглицерилтрансферазы; Δ lgt , lgt мутант с делецией гена; LTA, липотейхоевая кислота; MHA, агар Мюллера-Хинтона; вес, дикий тип.
№4 Онлайн-версия этой статьи содержит дополнительные материалы.
- Получено 22 декабря 2008 г.
- Принято 23 марта 2009 г.
- Авторские права © 2009 Американской ассоциации иммунологов, Inc.
Ссылки
- ↵
Столл, Х., Дж. Денгель, К. Нерц, Ф. Гётц. 2005. Staphylococcus aureus , дефицитный по липидированию прелипопротеинов, ослаблен в росте и активации иммунной системы. Заразить. Иммун. 73: 2411-2423.
- ↵
Кокейн, А., П. Дж. Хилл, Н. Б. Пауэлл, К. Бишоп, К. Симс, П. Уильямс. 1998. Молекулярное клонирование 32-килодальтонного липопротеинового компонента нового регулируемого железом транспортера ABC Staphylococcus epidermidis .Заразить. Иммун. 66: 3767-3774.
- ↵
Дейл, С. Э., М. Т. Себульский, Д. Э. Хайнрихс. 2004. Участие SirABC в импорте железо-сидерофоров у Staphylococcus aureus . J. Bacteriol. 186: 8356-8362.
- ↵
Себульский М. Т., Д. Э. Хайнрихс. 2001. Идентификация и характеристика fhuD1 и fhuD2 , двух генов, участвующих в захвате гидроксамата железа в Staphylococcus aureus .J. Bacteriol. 183: 4994-5000.
- ↵
Моррисси, Дж. А., А. Кокейн, П. Дж. Хилл, П. Уильямс. 2000. Молекулярное клонирование и анализ предполагаемого переносчика сидерофоров ABC из Staphylococcus aureus . Заразить. Иммун. 68: 6281-6288.
- ↵
Grigg, J. C., C. L. Vermeiren, D. E. Heinrichs, M. E. Murphy. 2007. Координация гема с помощью Staphylococcus aureus IsdE. J. Biol. Chem. 282: 28815-28822.
- ↵
Skaar, E.П., М. Хумаюн, Т. Бэ, К. Л. ДеБорд, О. Шнеуинд. 2004. Предпочтение источника железа для Staphylococcus aureus инфекций. Наука 305: 1626-1628.
- ↵
Qi, H. Y., K. Sankaran, K. Gan, H. C. Wu. 1995. Взаимосвязь структуры и функции бактериальной пролипопротеиндиацилглицерилтрансферазы: функционально значимые консервативные области. J. Bacteriol. 177: 6820-6824.
- ↵
Таварацумида, К., М. Фуруясики, М.Кацумото, Ю. Фудзимото, К. Фукасе, Ю. Суда, М. Хашимото. 2009. Характеристика N-концевой структуры TLR2-активирующего липопротеина в Staphylococcus aureus . J. Biol. Chem. 284: 9147-9152.
- ↵
Курокава, К., Х. Ли, К. Б. Ро, М. Асанума, Ю. С. Ким, Х. Накяма, А. Сирацучи, Ю. Чой, О. Такеучи, Х. Дж. Канг и др. 2009. Триацилированный АТФ-связывающий кластерный транспортер, связывающий субстрат липопротеин из Staphylococcus aureus , функционирует как нативный лиганд для Toll-подобного рецептора 2.J. Biol. Chem. 284: 8406-8411.
- ↵
Алипрантис, А.О., Р. Б. Янг, М. Р. Марк, С. Суггетт, Б. Дево, Дж. Д. Радольф, Г. Р. Климпель, П. Годовски, А. Зихлинский. 1999. Активация клеток и апоптоз бактериальными липопротеинами через Toll-подобный рецептор-2. Наука 285: 736-739.
- ↵
Такеда, К., О. Такеучи, С. Акира. 2002. Распознавание липопептидов Toll-подобными рецепторами. J Endotoxin Res. 8: 459-463.
- ↵
Хашимото, М., К. Таварацумида, Х. Кария, К. Аояма, Т. Тамура, Ю. Суда. 2006. Липопротеин является преобладающим лигандом Toll-подобного рецептора 2 в компонентах клеточной стенки Staphylococcus aureus . Int. Иммунол. 18: 355-362.
- ↵
Brightbill, HD, DH Libraty, SR Krutzik, RB Yang, JT Belisle, JR Bleharski, M. Maitland, MV Norgard, SE Plevy, ST Smale и др. 1999. Механизмы защиты хозяина, запускаемые микробами липопротеины через Toll-подобные рецепторы.Наука 285: 732-736.
- ↵
Hirschfeld, M., C.J. Kirschning, R. Schwandner, H. Wesche, J. H. Weis, R.M. Wooten, J. J. Weis. 1999. Передний край: воспалительная передача сигналов с помощью Borrelia burgdorferi липопротеинов опосредуется Toll-подобным рецептором 2. J. Immunol. 163: 2382-2386.
- ↵
Лиен, Э., Т. Дж. Селлати, А. Йошимура, Т. Х. Фло, Г. Равади, Р. В. Финберг, Дж. Д. Кэрролл, Т. Эспевик, Р. Р. Ингаллс, Дж.Д. Радольф, Д. Т. Голенбок. 1999. Toll-подобный рецептор 2 функционирует как рецептор распознавания образов для различных бактериальных продуктов. J. Biol. Chem. 274: 33419-33425.
- ↵
Krutzik, SR, MT Ochoa, PA Sieling, S. Uematsu, YW Ng, A. Legaspi, PT Liu, ST Cole, PJ Godowski, Y. Maeda, et al. 2003. Активация и регулирование Toll-подобные рецепторы 2 и 1 при лепре человека. Nat. Med. 9: 525-532.
- ↵
Хашимото, М., К. Таварацумида, Х. Кария, А. Киёхара, Ю. Суда, Ф. Крикаэ, Т. Кирикаэ, Ф. Гётц. 2006. Не липотейхоевая кислота, а липопротеины, по-видимому, являются доминирующими иммунобиологически активными соединениями в Staphylococcus aureus . J. Immunol. 177: 3162-3169.
- ↵
Джин, М. С., С. Е. Ким, Дж. Ю. Хео, М. Е. Ли, Х. М. Ким, С. Г. Пайк, Х. Ли, Дж. О. Ли. 2007. Кристаллическая структура гетеродимера TLR1-TLR2, индуцированная связыванием триацилированного липопептида.Ячейка 130: 1071-1082.
- ↵
Schwandner, R., R. Dziarski, H. Wesche, M. Rothe, C. J. Kirschning. 1999. Активация клеток, индуцированная пептидогликанами и липотейхоевой кислотой, опосредуется Toll-подобным рецептором 2. J. Biol. Chem. 274: 17406-17409.
- ↵
Hoebe, K., P. Georgel, S. Rutschmann, X. Du, S. Mudd, K. Crozat, S. Sovath, L. Shamel, T. Hartung, U. Zahringer, B. Beutler. 2005. CD36 — сенсор диацилглицеридов. Nature 433: 523-527.
- ↵
Schroder, N. W., S. Morath, C. Alexander, L. Hamann, T. Hartung, U. Zahringer, U. B. Gobel, J. R. Weber, R. R. Schumann. 2003. Липотейхоевая кислота (LTA) Streptococcus pneumoniae и Staphylococcus aureus активирует иммунные клетки через Toll-подобный рецептор (TLR) -2, липополисахарид-связывающий белок (LBP) и CD14, тогда как TLR-4 и MD-2 не участвуют. J. Biol. Chem. 278: 15587-15594.
- ↵
Bubeck Wardenburg, J., У. А. Уильямс, Д. Миссиакас. 2006. Защита хозяина от инфекции Staphylococcus aureus требует распознавания бактериальных липопротеинов. Proc. Natl. Акад. Sci. США 103: 13831-13836.
- ↵
Tjalsma, H., V. P. Kontinen, Z. Pragai, H. Wu, R. Meima, G. Venema, S. Bron, M. Sarvas, J. M. van Dijl. 1999. Роль процессинга липопротеинов сигнальной пептидазой II в грамположительной эубактерии Bacillus subtilis : сигнальная пептидаза II необходима для эффективной секреции α-амилазы, нелипопротеина.J. Biol. Chem. 274: 1698-1707.
- ↵
Сандер, П., М. Резван, Б. Уокер, С. К. Рампини, Р. М. Кроппенштедт, С. Элерс, К. Келлер, Дж. Р. Кибл, М. Хагемайер, М. Дж. Колстон, Б. Спрингер, Э. К. Боттгер . 2004. Обработка липопротеинов требуется для вирулентности Mycobacterium tuberculosis . Мол. Microbiol. 52: 1543-1552.
- ↵
Khandavilli, S., K. A. Homer, J. Yuste, S. Basavanna, T. Mitchell, J. S.Коричневый. 2008. Созревание Streptococcus pneumoniae липопротеинов сигнальной пептидазой типа II необходимо для функции переносчика ABC и полной вирулентности. Мол. Microbiol. 67: 541-557.
- ↵
Baumgartner, M., U. Karst, B. Gerstel, M. Loessner, J. Wehland, L. Jansch. 2007. Инактивация Lgt позволяет систематически характеризовать липопротеины из Listeria monocytogenes . J. Bacteriol. 189: 313-324.18.
- ↵
Хеннеке, П., S. Dramsi, G. Mancuso, K. Chraibi, E. Pellegrini, C. Theilacker, J. Hubner, S. Santos-Sierra, G. Teti, DT Golenbock, et al. 2008. Липопротеины являются критически важными токсинами, активирующими TLR2 в стрептококковый сепсис группы B. J. Immunol. 180: 6149-6158.
- ↵
Пети, К. М., Дж. Р. Браун, К. Ингрэхэм, А. П. Брайант, Д. Дж. Холмс. 2001. Липидная модификация прелипопротеинов незаменима для роста in vitro, но необходима для вирулентности Streptococcus pneumoniae .FEMS Microb. Lett. 200: 229-233.
- ↵
Такеучи, О., К. Хосино, С. Акира. 2000. Передний край: мыши с дефицитом TLR2 и MyD88 очень восприимчивы к инфекции Staphylococcus aureus . J. Immunol. 165: 5392-5396.
- ↵
Duthie, E. S., L. L. Lorenz. 1952. Стафилококковая коагулаза; механизм действия и антигенность. J. Gen. Microbiol. 6: 95-107.
- ↵
Фридман Д.Б., Д. Л. Штауфф, Г. Пищаны, К. В. Уитвелл, В. Дж. Торрес, Э. П. Скаар. 2006. Золотистый стафилококк перенаправляет центральный метаболизм для увеличения доступности железа. PLoS Pathog. 2: e87
- ↵
Echchannaoui, H., K. Frei, M. Letiembre, R.M Strieter, Y. Adachi, R. Landmann. 2005. Дефицит CD14 приводит к увеличению продукции MIP-2, экспрессии CXCR2, трансмиграции нейтрофилов и ранней смерти при пневмококковой инфекции. J. Leukocyte Biol. 78: 705-715.
- ↵
Браун, Дж. С., С. М. Гиллиланд, Д. В. Холден. 2001. Остров патогенности Streptococcus pneumoniae , кодирующий транспортер ABC, участвующий в захвате железа и вирулентности. Мол. Microbiol. 40: 572-585.
- ↵
Коэн Дж .. 2002. Иммунопатогенез сепсиса. Природа 420: 885-891.
- ↵
Sibbald, M. J., A. K. Ziebandt, S. Engelmann, M. Hecker, A. de Jong, H. J. Harmsen, G.К. Раангс, И. Стокроос, Дж. П. Арендс, Дж. Й. Дюбуа, Дж. М. ван Дейл. 2006. Картирование путей к патогенезу стафилококков с помощью сравнительной секретомики. Microbiol. Мол. Биол. Rev. 70: 755-788.
- ↵
Andrews, N. C., P. J. Schmidt. 2007. Гомеостаз железа. Анну. Rev. Physiol. 69: 69-85.
- ↵
Schmidt, K. A., N. P. Donegan, W. A. Kwan, Jr, A. Cheung. 2004. Влияние ςB и agr на экспрессию стафилококкового энтеротоксина B (кожного сала) в Staphylococcus aureus .Жестяная банка. J. Microbiol. 50: 351-360.
- ↵
Миллер, Л. С., Э. М. Пьетрас, Л. Х. Уриккио, К. Хирано, С. Рао, Х. Лин, Р. М. О’Коннелл, Ю. Ивакура, А. Л. Чунг, Г. Ченг, Р. Л. Модлин. 2007. Опосредованная инфламмасомами продукция IL-1β необходима для рекрутирования нейтрофилов против Staphylococcus aureus in vivo. J. Immunol. 179: 6933-6942.
- ↵
Миллер, Л. С., Р. М. О’Коннелл, М. А. Гутьеррес, Э. М. Пьетрас, А.Шахангян, К. Э. Гросс, А. Тирумала, А. Л. Чунг, Г. Ченг, Р. Л. Модлин. 2006. MyD88 опосредует рекрутирование нейтрофилов, инициированное IL-1R, но не активацию TLR2 в иммунитете против Staphylococcus aureus . Иммунитет 24: 79-91.
- ↵
Капетанович Р., М. А. Нахори, В. Баллой, К. Фиттинг, Д. Дж. Филпотт, Дж. М. Кавайон, М. Адиб-Конки. 2007. Вклад фагоцитоза и внутриклеточного зондирования в продукцию цитокинов макрофагами, активированными Staphylococcus aureus .Заразить. Иммун. 75: 830-837.
- ↵
Corbin, BD, EH Seeley, A. Raab, J. Feldmann, MR Miller, VJ Torres, KL Anderson, BM Dattilo, PM Dunman, R. Gerads, et al. 2008. Хелатирование металлов и ингибирование бактериального роста в тканевых абсцессах. Наука 319: 962-965.
- ↵
Theurl, I., G. Fritsche, S. Ludwiczek, K. Garimorth, R. Bellmann-Weiler, G. Weiss. 2005. Макрофаг: клеточная фабрика на границе между железом и иммунитетом для борьбы с инфекциями.Biometals 18: 359-367.
- ↵
Фло, Т. Х., К. Д. Смит, С. Сато, Д. Дж. Родригес, М. А. Холмс, Р. К. Стронг, С. Акира, А. Адерем. 2004. Липокалин 2 опосредует врожденный иммунный ответ на бактериальную инфекцию путем связывания железа. Nature 432: 917-921.
- ↵
Альбигер, Б., А. Сандгрен, Х. Кацураги, У. Мейер-Хофферт, К. Бейтер, Ф. Варта, М. Хорнеф, С. Нормарк, Б. Х. Нормарк. 2005. Миелоидный фактор дифференцировки 88-зависимая передача сигналов контролирует рост бактерий во время колонизации и системного пневмококкового заболевания у мышей.Клетка. Microbiol. 7: 1603-1615.
| Имя: | CTGF (182-250 аминокислотных остатков) Человек | |
| , N, N HCS24, IGFBP8, MGC102839, CTGF, фактор роста соединительной ткани. | ||
| Введение: | Фактор роста соединительной ткани принадлежит к семейству белков CCN.Семейство CCN в настоящее время состоит из шести членов у человека, также известных как: Cyr61 (Cystein rich 61), CTGF (фактор роста соединительной ткани), Nov (сверхэкспрессированный ген нефробластомы), WISP-1, 2 и 3 (индуцированные Wnt-1 секретируемые белки). ). Гены CCN кодируют секретируемые белки, связанные с внеклеточным матриксом (ECM) и клеточной мембраной. Белки CCN — это белки матрицеклеток, которые участвуют в регуляции различных клеточных функций, включая пролиферацию, дифференцировку, выживание, адгезию и миграцию.Они экспрессируются в производных трех эмбриональных листков и участвуют в развитии почек, нервной системы, мышц, костного мозга, хрящей и костей. В зрелом возрасте они участвуют в заживлении ран, восстановлении переломов костей и патологиях, таких как фиброз, сосудистые заболевания и онкогенез. Полноразмерные секретируемые белки CCN могут проявлять антипролиферативную активность, тогда как укороченные изоформы могут стимулировать пролиферацию и вести себя как онкогены. Полноразмерный белок состоит из четырех модулей: модуль I частично идентичен N-концевой части белков связывания инсулиноподобного фактора роста (IGFBPs).Модуль II включает в себя участок из 70 аминокислотных остатков, который разделяет идентичность последовательности с повтором фактора фон Виллебранда типа C (VWC). Модуль III содержит последовательности, идентичные повторению с повтором тромбоспондина типа 1 (TSP1) (WSXCSXXCG), который, как предполагается, имеет отношение к в связывании сульфатированных гликоконъюгатов и важен для клеточной адгезии. Модуль IV, также обозначаемый CT, кодируется exon5. Это наименее консервативный один из четырех доменов на уровне нуклеотидной последовательности, но, по-видимому, он имеет решающее значение для некоторых биологических функций, приписываемых белкам CCN.Модуль IV напоминает CT-домен нескольких внеклеточных белков, включая фактор фон Виллебранда и муцины. Сходство последовательностей с гепарин-связывающими мотивами также обнаруживается в этом домене. Протеолиз секретируемых полноразмерных белков CCN, о котором сообщалось в случае CCN2 и CCN3, может приводить к продукции CCN-производных пептидов с высоким сродством к лигандам, которые полноразмерные белки CNN связываются только плохо. Амино-усеченные изоформы CCN2 были биологически активными, тогда как усеченному CCN3 не приписывалось никакой специфической биологической активности.Хотя молекулярные процессы, лежащие в основе продукции этих секретируемых изоформ, в настоящее время неизвестны, важно отметить, что протеолиз происходит в одних и тех же аминокислотных остатках как в CCN2, так и в CCN3. Повышенная экспрессия CCN2 также была обнаружена с помощью Нозерн-блоттинга в инвазивных карциномах протоков молочной железы человека, дерматофибромах, пиогенных гранулемах, эндотелиальных клетках ангиолипом и ангиолейомиом, а также в опухолях поджелудочной железы. Исследование, проведенное с хондросаркомами различных гистологических степеней, показало, что экспрессия CCN2 тесно коррелировала с увеличением уровней злокачественности.В соответствии с тем, что CCN2 играет роль в ангиогенезе опухолей головного мозга, иммуноцитохимические исследования показали, что как опухолевые клетки глиобластомы, так и пролиферирующие эндотелиальные клетки окрашиваются положительно на CCN2. В астроцитомах экспрессия CCN2 была особенно повышена в опухолях высокой степени злокачественности с заметным влиянием CCN2 на пролиферацию клеток. Подавление экспрессии CCN2 в этих клетках было связано с остановкой роста при переходе G1 / S, тогда как сверхэкспрессия CCN2 индуцировала двукратное увеличение количества клеток в фазе G1.Анализ профилей генов позволил идентифицировать набор из примерно 50 генов, экспрессия которых может объяснять пролиферативную активность CCN2 в этих клетках. CCN2 обнаружен в большей доле мононуклеарных клеток пациентов с острым лимфобластным лейкозом. | |
| Описание: | Аминокислоты 182–250 фактора роста соединительной ткани, продуцируемые в E.Coli, представляют собой гибридный белок с His Tag (4 кДа), имеющий общую молекулярную массу 15 кДа. | |
| Источник: | Escherichia Coli. | |
| Размер упаковки: | 20 мкг | |
| Внешний вид: | Стерильный отфильтрованный белый лиофилизированный порошок. | |
| Состав: | Лиофилизированный без каких-либо добавок. | |
| Растворимость: | Рекомендуется восстановить лиофилизированный CTGF в стерильном 18 МОм – см вод.ст. не менее 100 мкг / мл, который затем можно разбавить другими водными растворами. | |
| Стабильность: | Лиофилизированный CTGF, хотя и стабилен при комнатной температуре в течение 3 недель, должен храниться осушенным при температуре ниже –18 ° C. После восстановления CTGF следует хранить при 4 ° C в течение 2–7 дней, а для будущего использования при температуре ниже –18 ° C. Для длительного хранения рекомендуется добавить белок-носитель (0,1% HSA или BSA). Пожалуйста, не допускайте замораживания– циклы оттаивания. | |
| Чистота: | Более 90,0% согласно данным SDS – PAGE. | |
| Аминокислотная последовательность: | Н / Д | |
| Биологическая активность: | НЕТ | |
| Использование: | Продукты LASATORY предназначены для анализа в США.Продукт нельзя использовать в качестве лекарств, сельскохозяйственных или пестицидных продуктов, пищевых добавок или бытовой химии. | |
| Uniprot: | P29279 | |
| Функция белка UniProt: | CTGF: основной митоаттрактант соединительной ткани, секретируемый эндотелиальными клетками сосудов. Способствует пролиферации и дифференцировке хондроцитов. Опосредует гепарин- и двухвалентную катион-зависимую клеточную адгезию во многих типах клеток, включая фибробласты, миофибробласты, эндотелиальные и эпителиальные клетки.Усиливает синтез ДНК, индуцированный фактором роста фибробластов. Принадлежит к семейству CCN. Альтернативным сплайсингом получают 2 изоформы человеческого белка. | |
| Белок UniProt Подробные сведения: | Тип белка: Адгезия клеток; Секретный; Секретируемый сигнальный пептид Хромосомное местоположение ортолога человека: 6q23.2 Клеточный компонент: внеклеточная область; Аппарат Гольджи; внутриклеточная мембраносвязанная органелла; плазматическая мембрана Молекулярная функция: связывание с белками ; связывание с С-концом белка Биологический процесс: развитие эпидермиса; положительная регуляция дифференцировки клеток; положительное регулирование каскада JNK; положительная регуляция образования стрессовых волокон; реакция на ранение; инициация транскрипции с промотора РНК-полимеразы II | |
| NCBI Резюме: | Белок, кодируемый этим геном, представляет собой митоген, который секретируется эндотелиальными клетками сосудов.Кодируемый белок играет роль в пролиферации и дифференцировке хондроцитов, клеточной адгезии во многих типах клеток и связан с фактором роста тромбоцитов. Определенные полиморфизмы этого гена связаны с более высокой заболеваемостью системным склерозом. [предоставлено RefSeq, ноябрь 2009 г.] | |
| Код UniProt: | P29279 | |
| NCBI Идентификатор GenInfo: | 116241320 | |
| NCBI Gene NCBI Присоединение: | P29279.2 | |
| Вторичный регистрационный номер UniProt: | P29279, Q6LCY0, Q96A79, Q96QX2, Q9UDL6, E1P578, | |
| UniProt | Вес, связанный с Plecular Plecular: 2||
| NCBI Полное имя: | Фактор роста соединительной ткани | |
| Синоним NCBI Полное имя: | Фактор роста соединительной ткани | |
| CTBI | | Официальный символG Официальные синонимы NCBI: CCN2; NOV2; HCS24; IGFBP8 |
| Информация о белке NCBI: | Фактор роста соединительной ткани | |
| Имя белка UniProt: | Фактор роста соединительной ткани | |
| член семейства CCBN Протеин UniP ; Гипертрофический хондроцит-специфический белок 24; Белок 8, связывающий инсулиноподобный фактор роста; ИБП-8; IGF-связывающий белок 8; IGFBP-8 | ||
| Семейство белков: | Фактор роста соединительной ткани | |
| Имя гена UniProt: | CTGF |