
|
Группы мышц |
Названия мышц |
Характер работы |
Функции |
Групи м’язів |
Назви м’язів |
Характер роботи |
Функції |
|
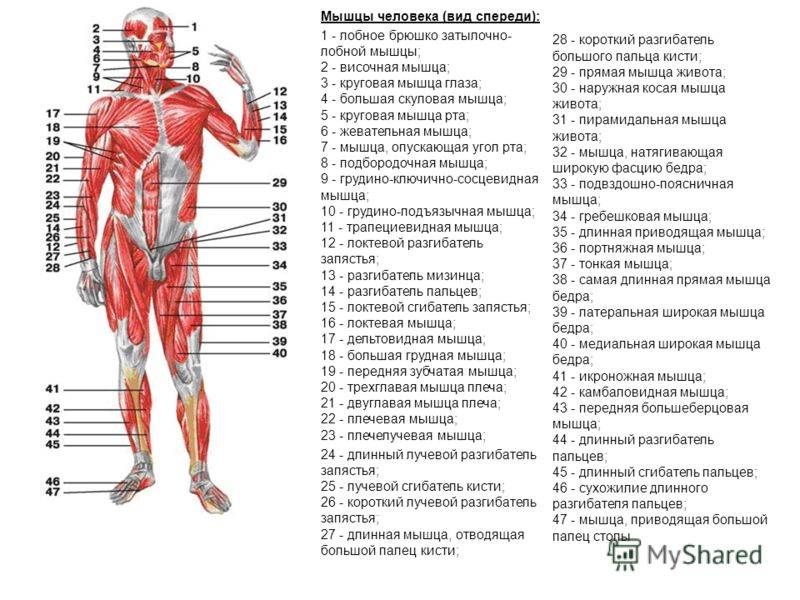
Мышцы головы |
М’язи голови | ||||||
|
Жевательные |
Жевательная, височная, внешняя, крыловидная |
Произвольный |
Движение нижней челюсти |
Жувальна |
Жувальна, скронева, зовнішня, крилоподібна |
Довільний |
Рухи нижньої щелепи |
|
Мимические |
Круговые мышцы рта и глаз, надчерепная |
Изменяют выражение лица, речевая артикуляция |
Мімічні |
Кругові м’язи рота та очей, надчерепна | Змінюють вираз обличчя, мовленнєву артикуляцію | ||
|
Мышцы туловища |
М’язи тулуба | ||||||
|
Затылочные, спинные, грудные, брюшные, диафрагма |
Большая и малая грудные, межреберные. |
Произвольный |
Поддержание туловища в вертикальном положении. |
Потиличні, спинні, грудні, черевні, діафрагма |
Велика та мала грудні, міжреберні. |
Довільний |
Підтримка тулуба у вертикальному положенні. |
|
Поверхностные и глубокие |
Мышцы – сгибатели и разгибатели. |
Поверхневі та глибокі. |
М’язи – згиначі і розгиначі. | ||||
|
Прямые и косые |
Движения тела. |
Прямі та косі |
Рухи тіла. | ||||
| Дыхательные движения. |
Дихальні рухи. | ||||||
|
Мышцы конечностей |
М’язи кінцівок | ||||||
|
Верхняя конечность |
Двуглавая и трехглавая мышцы рук, мышцы кисти и руки. |
Произвольный |
Мышцы – сгибатели и разгибатели рук, ног осуществляющие движения конечностей |
Верхня кінцівка |
Двоголовий та триголовий м’язи кисті і рук |
|
М’язи – згиначі і розгиначі рук, ніг, що здійснюють рухи кінцівок |
|
Нижняя конечность |
Двуглавая и четырехглавая икроножная мышцы ног; мышцы стоп |
Нижня кінцівка |
Двоголовий та чотирьохголовий литковий м’язи ніг, м’язи стоп | ||||
|
Внутренние органы |
Внутрішні органи | ||||||
|
Сердечная мышца. |
Непроизвольный |
Сокращение сердца. |
Серцевий м’яз. |
Мимовільний |
Скорочення серця. | ||
|
Мышцы стенок сосудов, кишечника, желудка, мышцы кожи |
Сокращение стенок полых органов, передвижение крови, пищевой массы |
М’язи стінок судин, кишечника, шлунка, м’язи шкіри |
Скорочення стінок порожнистих органів, пересування крові, харчової маси | ||||



Раскройте взаимосвязь строения и функций основных групп мышц человека.
 Сокращение мышц обеспечивает движение тела и удержание его в вертикальном положении. Вместе со скелетом мышцы придают телу форму. С деятельностью мышц связана функция отдельных органов: дыхания, пищеварения, кровообращения; мышцы гортани и языка участвуют в воспроизведении членораздельной речи.
Сокращение мышц обеспечивает движение тела и удержание его в вертикальном положении. Вместе со скелетом мышцы придают телу форму. С деятельностью мышц связана функция отдельных органов: дыхания, пищеварения, кровообращения; мышцы гортани и языка участвуют в воспроизведении членораздельной речи.Сокращение поперечнополосатой ткани подчинено сознанию. В теле человека насчитывается около 600 скелетных мышц, что составляет 2/5 общей массы тела.
Скелетная мышца покрыта плотной соединительнотканной оболочкой, плотно соединенной с мышечной тканью и препятствующей ее чрезмерному растяжению. Между пучками волокон в мышце расположены кровеносные сосуды и нервы. На концах мышца переходит в сухожилие, обладающее большой прочностью, но в отличие от мышц не обладающее сокращением.
Особый вид мышечной ткани — сердечная мышца, образованная поперечнополосатыми мышечными волокнами, но сокращается она непроизвольно. Следовательно, функциональные особенности, строение отличают мышцу сердца от других мышц.
Различают мышцы короткие и толстые, находящиеся преимущественно в глубоких слоях около позвоночного столба; длинные и тонкие, расположенные на конечностях; широкие и плоские, сосредоточенные в основном на туловище.
По функциям мышцы делятся на сгибатели, разгибатели, приводящие, отводящие, вращатели. При сокращении мышц сгибателей одновременно расслабляются разгибатели, что обеспечивает согласованность движений.
Мышцы, сокращение которых вызывает движение конечности от тела, называются отводящими, а приближающие конечность к телу, — приводящими. Мышцы вращатели при своем сокращении вращают ту или иную часть тела (голову, плечо, предплечье).
В организме человека различают мышцы туловища, головы, верхних и нижних конечностей.
Мышцы живота вызывают сгибание позвоночника вперед, в сторону и поворот его вокруг продольной оси, Образуют брюшной пресс, сокращение которого способствует глубокому выдоху, выведению кала, мочи, а также родовому акту у женщин.
Поверхностные мышцы спины (трапециевидная и широчайшая) обеспечивают укрепление и движение плечевого пояса и рук. Глубокие мышцы спины фиксируют позвоночник, вызывают его разгибание, сгибшие, наклоны в сторону и вращение, разгибание и вращение головы, участвуют в дыхательных движениях. Самая крупная мышца шеи — грудино-ключично-сосцевидная.
Мышцы головы подразделяются на две группы: жевательные и мимические. Собственно жевательная мышца начинается от нижнего края скуловой кости и прикрепляется к нижней челюсти. Сокращаясь, она поднимает нижнюю челюсть, участвуя в пережевывании пищи.
Сокращаясь, она поднимает нижнюю челюсть, участвуя в пережевывании пищи.
Мимические мышцы прикрепляются одним концом к костям черепа, другим — к коже лица. Благодаря им лицо человека выражает те или иные эмоции: гнев, горе, радость. Кроме того, они участвуют в акте речи, дыхания. На лбу расположены лобные мышцы, вокруг глазницы — круговая мышца (способствует закрыванию век). Вокруг ротового отверстия находится круговая мышца рта.
Мускулатура верхних конечностей подразделяется на мышцы плечевого пояса (дельтовидная, большая и малая грудная), которые обеспечивают его подвижность, и мышцы свободной конечности. Важнейшие мышцы свободной конечности — двуглавая мышца (сгибает предплечье) и трехглавая (на задней поверхности плечевой кости), разгибающая плечо и предплечье. На передней поверхности предплечья находятся мышцы — сгибатели предплечья, кисти и пальцев, на задней — мышцы — разгибатели предплечья, кисти и пальцев.
Мышцы нижних конечностей подразделяются на мышцы тазового пояса и свободной конечности. К мышцам таза относятся подвздошно-поясничная мышца и три ягодичные, самая крупная, разгибающая бедро, — большая ягодичная. На задней поверхности бедра выделяются полусухожильная, полуперепончатая и двуглавая мышцы, при сокращении которых происходит сгибание голени в коленном суставе и разгибание бедра.
К мышцам таза относятся подвздошно-поясничная мышца и три ягодичные, самая крупная, разгибающая бедро, — большая ягодичная. На задней поверхности бедра выделяются полусухожильная, полуперепончатая и двуглавая мышцы, при сокращении которых происходит сгибание голени в коленном суставе и разгибание бедра.
На передней поверхности бедра лежит четырехглавая мышца, при сокращении которой разгибается голень. На передней поверхности голени находятся мышцы — разгибатели стопы и пальцев, на задней — их сгибатели. Важнейшие из них — икроножная и камбалообразная. Обе мышцы заканчиваются ахилловым сухожилием, которое прикрепляется к пяточному бугру. Икроножная мышца поднимает пятку при ходьбе и принимает участие в поддержании тела в вертикальном положении.
Мышцы верхней конечности осуществляют разнообразные и многочисленные движения руки. Так как нижние конечности человека выдерживают всю тяжесть тела и целиком принимают на себя функцию его передвижения, то их мышцы значительно массивнее, и, следовательно, сильнее, чем мышцы рук, но вместе с тем обладают более ограниченным размахом движений.
Статическая и динамическая работа мышц
Даже если человек находится в неподвижном положении, его мышцы все равно производят работу, поддерживая корпус и координируя тело в пространстве. В теле человека огромное количество мышц, объединенных в группы, которые работают слаженно, обеспечивая нормальную двигательную активность. Давайте узнаем, что такое статические и динамические мышцы, а также как использовать эти знания для грамотного тренинга. Много полезной информации по тренировкам и питанию вы узнаете в фитнес-клубе «Мультиспорт», где работают опытные специалисты, которые помогут вам достичь желаемых целей в фитнесе и спорте.
Виды работы мышц
Существует два вида работы мышц: статическая и динамическая. Если при работе мышц происходят движения в суставах, то ее называют динамической. Если суставы неподвижны, то работа мышц заключается в поддержании тела в определенном положении. В таком случае мышечную работу называют статической.
Легко понять, чем отличается динамическая и статическая работа мышц: в первом случае суставы совершают движение, во втором случае – остаются неподвижно. Теперь разберемся, чем характеризуется каждый вид работы.
Теперь разберемся, чем характеризуется каждый вид работы.
Ключевой характеристикой динамичной работы мышц является энергозатратность. Несмотря на то, что мышцы тратят энергию в статике, во время активного движения ее затрачивается в разы больше.
Динамическая работа мышц
Под динамической работой мышц подразумевается двигательная активность, при которой происходит попеременное расслабление и сокращение мышц для перемещения тела в пространстве или выполнения определенного движения.
При выполнении динамической работы происходят физиологические реакции организма, которых не возникает во время статической мышечной работы. Примером таких реакций служит увеличение пульса и артериального давления во время активности. Интенсивность проявления реакций зависит от разных факторов: тренированности человека, силы и частоты мышечного сокращения, и даже от того, в каком положении находилось тело до начала активности.
Динамическую работу классифицируют по количеству работающих мышц:
- Глобальная – если в движении принимают участие более двух третей от всех мышц тела;
- Региональная – если в движении задействовано менее двух третей от общего количества мышц;
- Локальная – если в движение участвует менее трети от всех мышц.

Например, базовые упражнения, вроде приседаний, становой тяги, прыжков задействуют огромное количество мышц, в результате чего происходит глобальная или региональная динамическая работа. Изолированные упражнения, например, подъем штанги на бицепс, разгибания на трицепс подключают в работу не слишком много мышц, а потому происходит локальная динамическая работа.
Динамическая работа мышц может быть преодолевающей и уступающей, что значит преодоление сопротивления и непротиводействие. Рассмотрим на примере мышц рук: при отведении выполняется преодолевающая динамическая работа, при приведении – уступающая. А при удержании руки в определенном положении выполняется статическая или удерживающая работа мышц.
Статическая работа мышц
Если вам интересно, какая работа мышц называется динамической и статической, то с первой уже разобрались. Динамическая работа возникает во время любого движения или физической активности. Теперь узнаем, какое отличие у статической работы.
При статической работе мышцы постоянно сокращаются, чтобы удерживать тело в определенном положении или обеспечивать выполнение простых бытовых действий.
При статической работе не происходит чрезмерного потребления кислорода и активации кровотока, но проявляются различные физиологические реакции и происходят энергетические затраты. Например, при выполнении статических упражнений, планки или стульчика тело тратит энергию на удержание определенного положения. Поэтому нагрузку мышц можно получить в статике, хотя энергозатраты, конечно, не сравнятся с динамической работой. Физиологические реакции организма в виде учащения пульса и повышения давления зависит от продолжительности работы и силы сокращений.
Между статической и динамической работой мышц есть различия, например, динамическая работа обеспечивается сокращающимися и расслабляющимися мышцами, а статическая – непрерывно сокращающимися. Но эти виды работы последовательно сменяют друг друга в нашей повседневной деятельности и не могут существовать друг без друга.
Что еще важно знать
В реальной жизни мышцы не работают изолированно, поэтому таблиц о конкретно динамической или статической работе вы не найдете. Важно помнить, что в статике всегда есть элементы динамики и наоборот.
Планируя тренировки на увеличение силы мышц, следует включать плиометрические и статические упражнения чтобы развивать медленные и быстрые мышечные волокна, что сделает вас сильнее и выносливее.
Многих интересует быстрое утомление мышц при статической нагрузке. Дело в том, что перманентное сокращение определенных мышц затрудняет насыщение клеток кислородом и выведение продуктов распада, что приводит к усталости. Во время динамических движений такого не происходит, поэтому поднимать и опускать руку вы можете дольше, чем удерживать ее в неподвижном положении.
Если вас интересует, что такое статические и динамические мышцы, а также как применить знания на практике, то приходите в клуб «Мультиспорт», где вас ждет множество интересных активностей, профессиональные тренеры, современно оборудованные залы и многое другое.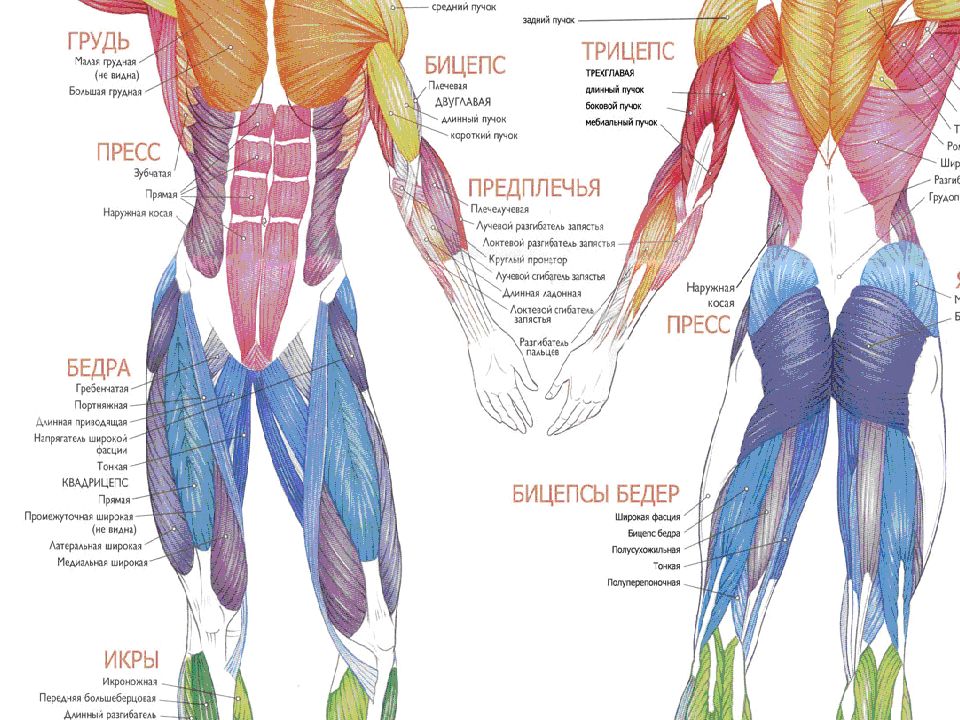 Звоните прямо сейчас, чтобы получить подробную консультацию!
Звоните прямо сейчас, чтобы получить подробную консультацию!
Поделиться:
Транскрипционное профилирование старения в мышцах человека выявляет общий признак старения
Abstract
Мы проанализировали экспрессию 81 образца нормальной мышцы человека разного возраста и определили молекулярный профиль старения, состоящий из 250 генов, регулируемых возрастом. Этот молекулярный профиль коррелирует не только с хронологическим возрастом, но и с мерой физиологического возраста. Мы сравнили транскрипционный профиль мышечного старения с предыдущими транскрипционными профилями старения в почках и головном мозге и обнаружили общие признаки старения в этих различных тканях человека.Общий признак старения состоит из шести генетических путей; четыре пути увеличивают экспрессию с возрастом (гены внеклеточного матрикса, гены, участвующие в росте клеток, гены, кодирующие факторы, участвующие в активации комплемента, и гены, кодирующие компоненты цитозольной рибосомы), а два пути снижают экспрессию с возрастом (гены, участвующие в транспорте хлоридов). и гены, кодирующие субъединицы митохондриальной электрон-транспортной цепи). Мы также сравнили транскрипционные профили старения у людей с таковыми у мышей и мух и обнаружили, что путь цепи переноса электронов снижает экспрессию с возрастом у всех трех организмов, предполагая, что это может быть общедоступным маркером старения у разных видов.
и гены, кодирующие субъединицы митохондриальной электрон-транспортной цепи). Мы также сравнили транскрипционные профили старения у людей с таковыми у мышей и мух и обнаружили, что путь цепи переноса электронов снижает экспрессию с возрастом у всех трех организмов, предполагая, что это может быть общедоступным маркером старения у разных видов.
Сводка
Старение — сложное явление, характеризующееся угасанием биологических функций с течением времени, что в конечном итоге приводит к смерти. Высокопроизводительные методы исследования изменений в экспрессии генов, такие как ДНК-микрочипы, оказались успешными в выяснении некоторых полногеномных изменений, происходящих с возрастом в некоторых тканях человека. Авторы профилировали изменения экспрессии генов в мышцах 81 человека в возрасте восьмидесяти лет. Они обнаружили 250 генов и 3 генетических пути, которые демонстрируют измененные уровни экспрессии у пожилых людей.Профиль транскрипции генов, регулируемых возрастом, позволил отличить пожилых пациентов с тяжелым мышечным старением от тех, у кого сохранился высокий уровень мышечной функции; то есть профили экспрессии генов отражали как физиологический, так и хронологический возраст. Чтобы найти генетические изменения, которые могут затронуть большинство или все ткани во время старения, авторы сравнили полногеномные профили старения в мышцах с таковыми в почках и мозге и обнаружили общие признаки старения, характерные для этих трех тканей, состоящие из из шести генетических путей.Один из этих путей старения (цепь переноса электронов) регулируется возрастом не только у людей, но и у двух модельных организмов (мышей и мух), что дает представление об общих возрастных изменениях у животных с совершенно разной продолжительностью жизни.
Чтобы найти генетические изменения, которые могут затронуть большинство или все ткани во время старения, авторы сравнили полногеномные профили старения в мышцах с таковыми в почках и мозге и обнаружили общие признаки старения, характерные для этих трех тканей, состоящие из из шести генетических путей.Один из этих путей старения (цепь переноса электронов) регулируется возрастом не только у людей, но и у двух модельных организмов (мышей и мух), что дает представление об общих возрастных изменениях у животных с совершенно разной продолжительностью жизни.
Образец цитирования: Zahn JM, Sonu R, Vogel H, Crane E, Mazan-Mamczarz K, Rabkin R, et al. (2006) Транскрипционное профилирование старения в мышцах человека выявляет общий признак старения. Генетика PLoS 2(7): е115. https://дои.org/10.1371/journal.pgen.0020115
Редактор: Wayne Frankel, The Jackson Laboratory, United States of America
Получено: 14 марта 2006 г. ; Принято: 9 июня 2006 г.; Опубликовано: 21 июля 2006 г.
; Принято: 9 июня 2006 г.; Опубликовано: 21 июля 2006 г.
Это статья с открытым доступом, распространяемая в соответствии с условиями декларации Creative Commons Public Domain, которая предусматривает, что после помещения в общественное достояние эта работа может свободно воспроизводиться, распространяться, передаваться, изменены, построены или иным образом использованы кем-либо в любых законных целях.
Финансирование: Эта работа была поддержана стипендией для выпускников Национального научного фонда, Стэнфордским грантом на обучение геному, финансированием оценки заслуг Департамента по делам ветеранов, Национальных институтов здравоохранения и Медицинского фонда Эллисона.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Сокращения: ГСЭА, анализ обогащения набора генов
Введение
Старение характеризуется постепенным снижением множества физиологических функций, что приводит к увеличению вероятности смерти. Некоторые изменения, связанные со старением, влияют на внешний вид человека, например, морщинистая кожа, в то время как другие влияют на функцию органов, например снижение скорости почечной фильтрации и снижение мышечной силы. На молекулярном уровне мы только начинаем собирать белки и изменения экспрессии генов, которые можно использовать в качестве маркеров старения. Вместо того, чтобы искать молекулярные маркеры старения, сосредотачиваясь только на одном гене или пути за раз, привлекательным подходом является параллельный скрининг всех генетических путей на предмет возрастных изменений с использованием полногеномных олигонуклеотидных чипов для поиска изменений экспрессии генов в организме. пожилой.Полногеномный транскрипционный профиль старения может идентифицировать молекулярные маркеры процесса старения и даст представление о молекулярных механизмах, которые в конечном итоге ограничивают продолжительность жизни человека.
Некоторые изменения, связанные со старением, влияют на внешний вид человека, например, морщинистая кожа, в то время как другие влияют на функцию органов, например снижение скорости почечной фильтрации и снижение мышечной силы. На молекулярном уровне мы только начинаем собирать белки и изменения экспрессии генов, которые можно использовать в качестве маркеров старения. Вместо того, чтобы искать молекулярные маркеры старения, сосредотачиваясь только на одном гене или пути за раз, привлекательным подходом является параллельный скрининг всех генетических путей на предмет возрастных изменений с использованием полногеномных олигонуклеотидных чипов для поиска изменений экспрессии генов в организме. пожилой.Полногеномный транскрипционный профиль старения может идентифицировать молекулярные маркеры процесса старения и даст представление о молекулярных механизмах, которые в конечном итоге ограничивают продолжительность жизни человека.
Молекулярные маркеры старения должны отражать физиологические функции, а не просто хронологический возраст, поскольку люди стареют с разной скоростью [1]. У мышей изменения уровней CD4-иммуноцитов и изменения экспрессии генов клеточного цикла, таких как p16 INK4a , являются молекулярными маркерами старения, поскольку они предсказывают как оставшуюся продолжительность жизни, так и физиологический возраст мыши. 2–4].У человека эксперименты по профилированию экспрессии генов выявили 447 генов, регулируемых возрастом, которые могут предсказывать физиологический возраст почек [5]. Полногеномное профилирование экспрессии также использовалось для идентификации генов, которые изменяют экспрессию с возрастом в головном мозге [6], скелетных мышцах [7,8] и дермальных фибробластах [9], но изменения в экспрессии этих маркерных генов не были выявлены. однако было показано, что они коррелируют с физиологическим старением.
У мышей изменения уровней CD4-иммуноцитов и изменения экспрессии генов клеточного цикла, таких как p16 INK4a , являются молекулярными маркерами старения, поскольку они предсказывают как оставшуюся продолжительность жизни, так и физиологический возраст мыши. 2–4].У человека эксперименты по профилированию экспрессии генов выявили 447 генов, регулируемых возрастом, которые могут предсказывать физиологический возраст почек [5]. Полногеномное профилирование экспрессии также использовалось для идентификации генов, которые изменяют экспрессию с возрастом в головном мозге [6], скелетных мышцах [7,8] и дермальных фибробластах [9], но изменения в экспрессии этих маркерных генов не были выявлены. однако было показано, что они коррелируют с физиологическим старением.
В этой статье мы провели полногеномный анализ изменений экспрессии генов в скелетных мышцах человека.С возрастом скелетные мышцы дегенерируют, теряют массу, теряют общую аэробную способность и становятся заметно слабее [10]. Одним из показателей физиологии мышц является соотношение диаметров мышечных волокон I и II типов. Было обнаружено, что уменьшение размера мышечных волокон II типа (быстрых сокращений) коррелирует со снижением мышечной функции как у человека [11], так и у крысы [12]. Известно, что мышечные волокна типа II атрофируются и становятся меньше с возрастом у человека, что частично объясняет снижение мышечной силы и гибкости в пожилом возрасте.Поскольку мышечные волокна II типа с возрастом становятся меньше, отношение диаметров волокон II типа к волокнам I типа уменьшается.
Одним из показателей физиологии мышц является соотношение диаметров мышечных волокон I и II типов. Было обнаружено, что уменьшение размера мышечных волокон II типа (быстрых сокращений) коррелирует со снижением мышечной функции как у человека [11], так и у крысы [12]. Известно, что мышечные волокна типа II атрофируются и становятся меньше с возрастом у человека, что частично объясняет снижение мышечной силы и гибкости в пожилом возрасте.Поскольку мышечные волокна II типа с возрастом становятся меньше, отношение диаметров волокон II типа к волокнам I типа уменьшается.
Степень, в которой возрастная регуляция генетических путей является специфичной для конкретной ткани или общей для многих тканей, неизвестна. Установлено, что возрастная регуляция экспрессии генов между корковым и мозговым веществами почек человека сильно коррелирует [5]. Выявлена высокая корреляция изменений экспрессии генов с возрастом в разных областях коры головного мозга, но сходства между корой и мозжечком обнаружено не было [13].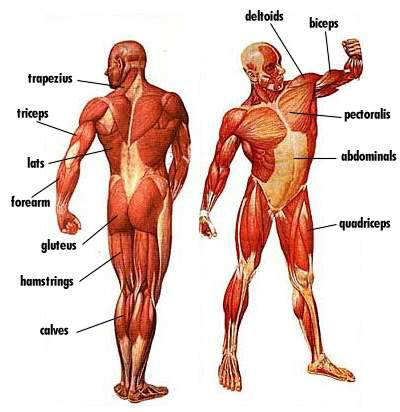 Таким образом, существуют сходства в паттернах возрастной регуляции между различными областями почек и между различными областями коры головного мозга, но общих признаков старения во многих различных тканях обнаружено не было.
Таким образом, существуют сходства в паттернах возрастной регуляции между различными областями почек и между различными областями коры головного мозга, но общих признаков старения во многих различных тканях обнаружено не было.
Другой ключевой вопрос заключается в том, существуют ли генетические пути, которые обычно регулируются возрастом у разных видов с очень разной продолжительностью жизни, таких как человек, мышь, муха и червь. Транскрипционные профили старения были выполнены как для скелетных мышц, так и для мозга у мышей [14,15], у Drosophila melanogaster [16,17] и у Caenorhabditis elegans [18].Сравнение паттернов изменения экспрессии генов при старении у мухи и червя позволило сделать вывод, что гены, кодирующие митохондриальные компоненты, снижали экспрессию с возрастом у обоих видов [19].
В этой работе мы представляем профиль транскрипционной экспрессии 81 образца скелетных мышц человека в зависимости от возраста. Активность симпортера, активность сиалилтрансферазы и пути транспорта хлоридов снижают экспрессию с возрастом в мышцах человека. Было обнаружено, что гены, регулирующие возраст, являются маркерами физиологического возраста, а не только хронологического возраста.Сравнивая наши результаты по старению в мышцах с предыдущими профилями транскрипции старения в почках и головном мозге, мы обнаружили общую характеристику старения в различных тканях человека, состоящую из шести генетических путей, которые показали общие закономерности возрастной регуляции во всех трех тканях. Наконец, сравнив сигнатуру старения у людей с транскрипционными профилями старения у мышей, мух и червей, мы обнаружили, что экспрессия электрон-транспортной цепи снижается с возрастом у людей, мышей и мух, что представляет собой общедоступную сигнатуру старения во всем мире. виды с очень разной продолжительностью жизни.
Было обнаружено, что гены, регулирующие возраст, являются маркерами физиологического возраста, а не только хронологического возраста.Сравнивая наши результаты по старению в мышцах с предыдущими профилями транскрипции старения в почках и головном мозге, мы обнаружили общую характеристику старения в различных тканях человека, состоящую из шести генетических путей, которые показали общие закономерности возрастной регуляции во всех трех тканях. Наконец, сравнив сигнатуру старения у людей с транскрипционными профилями старения у мышей, мух и червей, мы обнаружили, что экспрессия электрон-транспортной цепи снижается с возрастом у людей, мышей и мух, что представляет собой общедоступную сигнатуру старения во всем мире. виды с очень разной продолжительностью жизни.
Результаты
Глобальный профиль экспрессии генов старения в мышцах человека
Чтобы изучить эффекты старения мышц человека, мы получили 81 образец скелетных мышц человека в возрасте от 16 до 89 лет (таблица 1). Шестьдесят три образца были получены из брюшной полости, 5 — из руки, 2 — из дельтовидной мышцы, 2 — из внутренней поверхности бедра и 9 — из четырехглавой мышцы (таблица S1). Мы использовали массивы ДНК Affymetrix для создания транскрипционного профиля старения мышц человека.Мы выделили общую РНК из каждого образца мышц и синтезировали биотинилированную кРНК из общей РНК. Затем мы гибридизовали кРНК с массивами олигонуклеотидов Affymetrix 133 2.0 Plus, представляющими почти весь геном человека (54 675 отдельных наборов зондов, соответствующих 31 948 отдельным генам человека). Мы построили график экспрессии каждого гена в зависимости от возраста, в результате чего был получен набор данных, который показывает экспрессию почти каждого гена в геноме в зависимости от возраста в мышцах человека (данные общедоступны на сайте Gene Expression Omnibus по адресу http:/ /www.ncbi.nlm.nih.gov/geo).
Шестьдесят три образца были получены из брюшной полости, 5 — из руки, 2 — из дельтовидной мышцы, 2 — из внутренней поверхности бедра и 9 — из четырехглавой мышцы (таблица S1). Мы использовали массивы ДНК Affymetrix для создания транскрипционного профиля старения мышц человека.Мы выделили общую РНК из каждого образца мышц и синтезировали биотинилированную кРНК из общей РНК. Затем мы гибридизовали кРНК с массивами олигонуклеотидов Affymetrix 133 2.0 Plus, представляющими почти весь геном человека (54 675 отдельных наборов зондов, соответствующих 31 948 отдельным генам человека). Мы построили график экспрессии каждого гена в зависимости от возраста, в результате чего был получен набор данных, который показывает экспрессию почти каждого гена в геноме в зависимости от возраста в мышцах человека (данные общедоступны на сайте Gene Expression Omnibus по адресу http:/ /www.ncbi.nlm.nih.gov/geo).
Мы использовали метод множественной регрессии для каждого гена, чтобы определить, как его экспрессия меняется с возрастом, как это было сделано ранее для возрастной регуляции в почках (Материалы и методы) [5].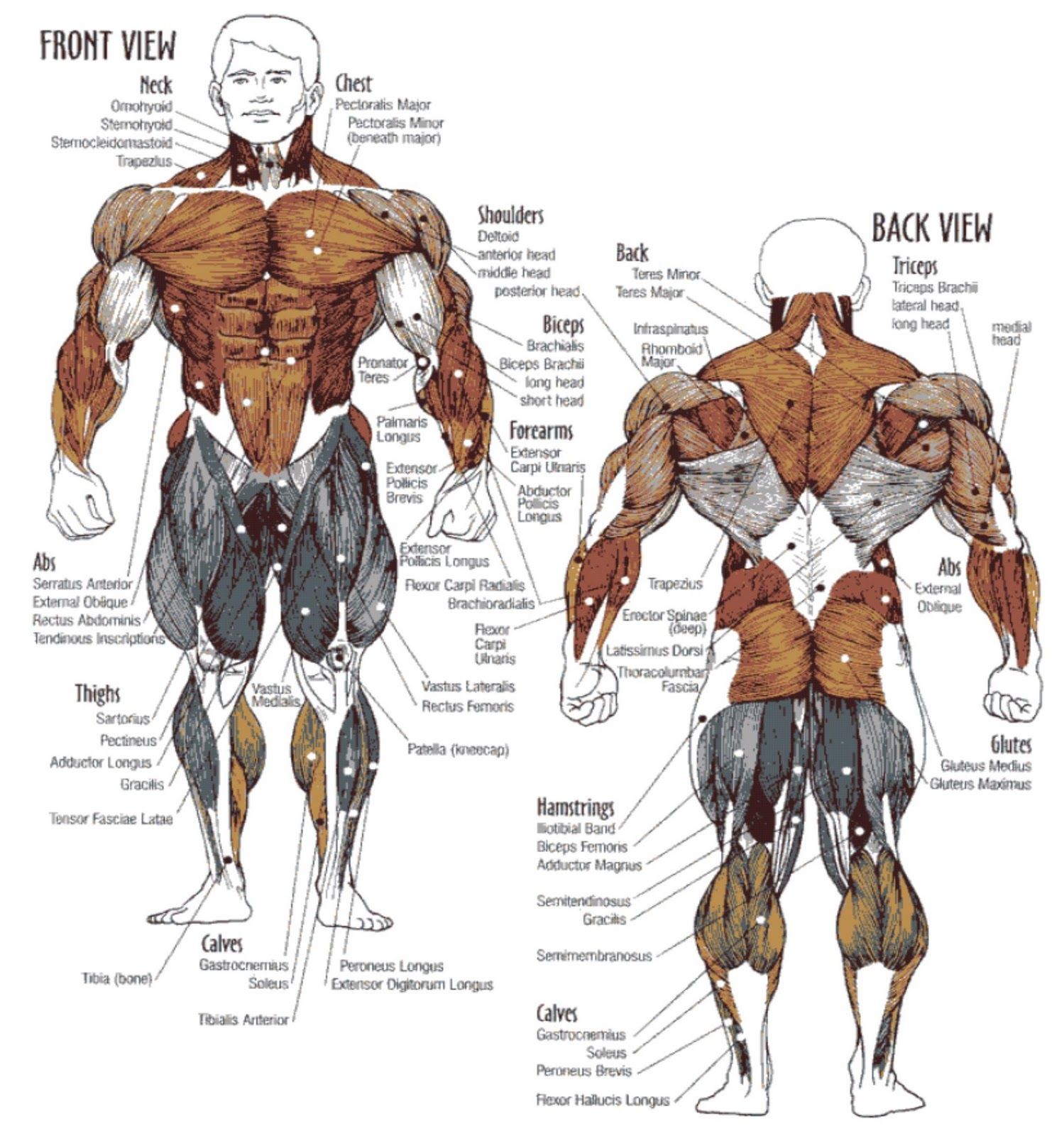 Мы проанализировали возрастную регуляцию скелетных мышц двумя способами. В первом случае мы нашли отдельные гены, соответствующие строгому порогу статистической значимости корреляции с возрастом. Во-вторых, мы обнаружили группы генов (определенные консорциумом Gene Ontology), в которых существует тонкая, но последовательная возрастная регуляция.
Мы проанализировали возрастную регуляцию скелетных мышц двумя способами. В первом случае мы нашли отдельные гены, соответствующие строгому порогу статистической значимости корреляции с возрастом. Во-вторых, мы обнаружили группы генов (определенные консорциумом Gene Ontology), в которых существует тонкая, но последовательная возрастная регуляция.
Чтобы идентифицировать отдельные гены, демонстрирующие сильную возрастную регуляцию, мы исследовали наклон по отношению к возрасту для каждого гена и выявили 250 генов, в которых наклон был значительно положительным или отрицательным ( p <0,001) (рис. 1, таблица S2 и Материалы и методы). При этом статистическом пороге мы ожидаем, что только 32 гена будут случайными, что предполагает уровень ложных открытий 13% или меньше. Кроме того, мы случайным образом переставили возраст образцов мышц, сохраняя фиксированными экспрессию генов, пол и анатомические переменные, и подсчитали количество генов, которые в значительной степени регулируются возрастом, снова при p <0.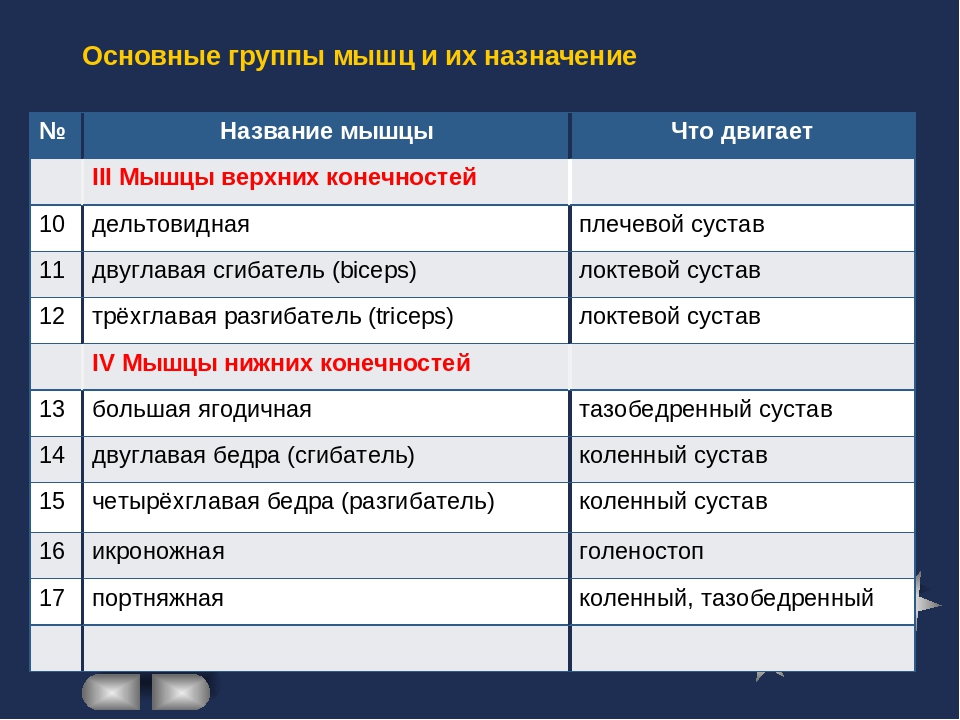 001. В 1000 таких перестановок мы обнаружили менее 107 значимых генов в 95% случаев. Таким образом, мы уверены, что большинство из 250 генов, регулируемых возрастом, не являются артефактами выборки. Из 250 генов, регулируемых возрастом, 125 генов увеличивают экспрессию, а 125 генов уменьшают экспрессию с возрастом.
001. В 1000 таких перестановок мы обнаружили менее 107 значимых генов в 95% случаев. Таким образом, мы уверены, что большинство из 250 генов, регулируемых возрастом, не являются артефактами выборки. Из 250 генов, регулируемых возрастом, 125 генов увеличивают экспрессию, а 125 генов уменьшают экспрессию с возрастом.
Рисунок 1. Экспрессия 250 генов, регулируемых возрастом, в мышцах
Строки соответствуют отдельным генам, расположенным в порядке от наибольшего увеличения экспрессии с возрастом вверху до наибольшего снижения экспрессии с возрастом внизу.Столбцы представляют отдельных пациентов, от самого младшего слева до самого старшего справа. Возраст определенных лиц отмечен для справки. Шкала представляет уровень экспрессии log 2 (Exp). Гены, обсуждаемые в тексте, помечены для справки. Навигационная версия этого рисунка, показывающая идентичность конкретных генов, может быть найдена по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
https://doi.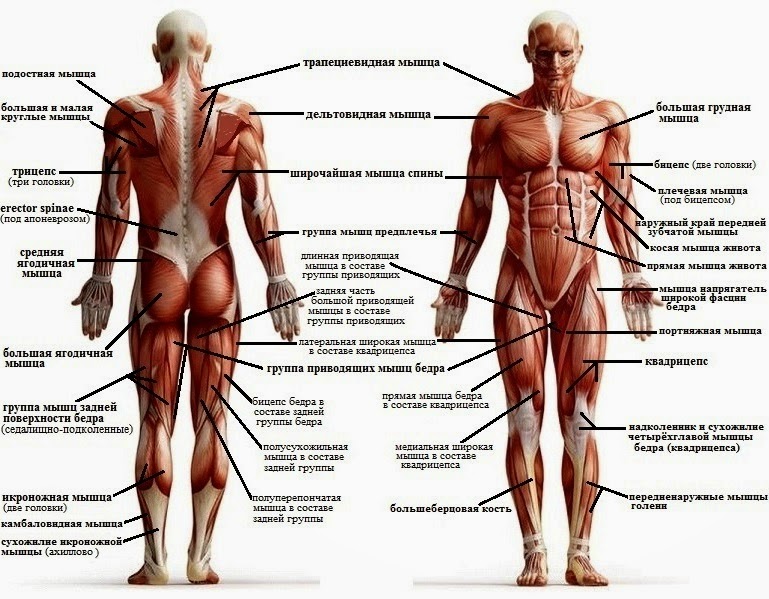 org/10.1371/journal.pgen.0020115.g001
org/10.1371/journal.pgen.0020115.g001
Мы рассмотрели возможность того, что некоторые из 250 генов могут не регулироваться возрастом сами по себе, а могут показаться, что они регулируются возрастом, потому что они связаны с патологическое состояние, усиливающееся с возрастом.Например, известно, что заболеваемость диабетом увеличивается с возрастом в общей популяции людей [20]. Наша выборка пациентов может показать предвзятость диабета у пожилых людей, и в этом случае гены, которые изменяют экспрессию в ответ на диабет, могут оказаться возрастными в нашем исследовании. В дополнение к диабету мы рассмотрели тринадцать других факторов, которые также могут исказить наше исследование старения, например, был ли пациент мужчиной или женщиной, анатомическое происхождение образца мышцы, тип патологии, связанной с пациентом, и типы лекарств. принимается пациентом (таблица S1).
За исключением гипотиреоза, ни один из медицинских факторов не продемонстрировал сильной связи с возрастом, поэтому маловероятно, что эти смешанные факторы могут привести к тому, что гены будут регулироваться возрастом (рис. S1). Гипотиреоз отсутствовал у молодых и присутствовал примерно у половины пожилых людей.
S1). Гипотиреоз отсутствовал у молодых и присутствовал примерно у половины пожилых людей.
Мы использовали два метода, чтобы проверить, влияет ли какой-либо из факторов на наклон экспрессии генов по отношению к возрасту 250 генов, регулируемых возрастом. Во-первых, мы использовали модель множественной регрессии, которая включала четвертый член, представляющий медицинский фактор (например, гипотиреоз) в дополнение к возрасту, полу и анатомии.Затем мы сравнили коэффициент старения, используя эту новую модель, с коэффициентом исходной модели, которая не включала этот термин. Если бы любой из 250 генов регулировался медицинским фактором, а не возрастом как таковым, можно было бы ожидать заметных различий в коэффициентах старения, полученных двумя моделями множественной регрессии. Ни один из четырнадцати медицинских факторов, включая гипотиреоз, не оказал существенного влияния на возрастную регуляцию (рис. S2). Во-вторых, мы провели неконтролируемый иерархический кластерный анализ 250 генов, регулируемых возрастом.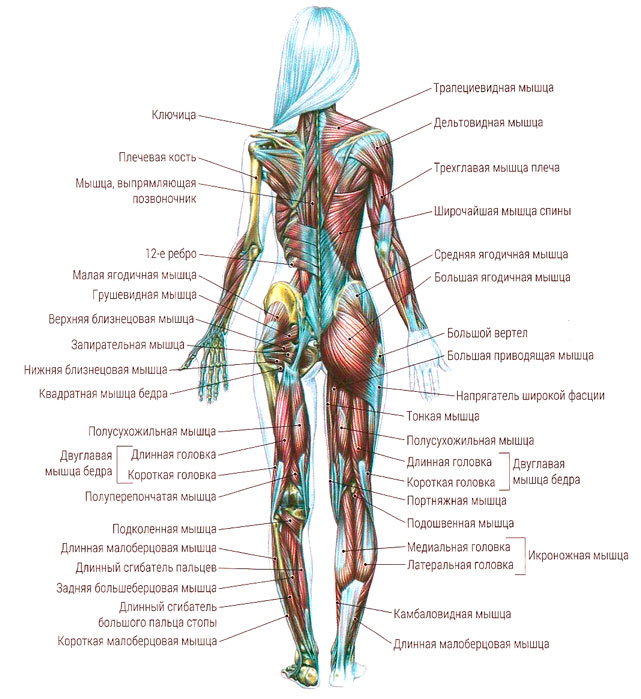 Если бы наш анализ возрастной регуляции был смешан с медицинским фактором, мы могли бы ожидать, что присутствие медицинского фактора будет сгруппировано, когда мы отсортируем 81 пациента в соответствии с их паттернами экспрессии генов. Ни один из патологических или фармацевтических факторов не показал кластеризации (рис. S3). Большинство неабдоминальных образцов были взяты у молодых пациентов, и, как и ожидалось, наблюдалась некоторая группировка образцов мышц в соответствии с их анатомическим происхождением. Эта кластеризация не влияет на наш анализ возрастной регуляции, поскольку анатомическое происхождение было включено в качестве термина в модель множественной регрессии.Таким образом, эти два метода не показали никаких доказательств того, что анатомические, патологические или фармацевтические факторы искажают результаты нашего исследования старения.
Если бы наш анализ возрастной регуляции был смешан с медицинским фактором, мы могли бы ожидать, что присутствие медицинского фактора будет сгруппировано, когда мы отсортируем 81 пациента в соответствии с их паттернами экспрессии генов. Ни один из патологических или фармацевтических факторов не показал кластеризации (рис. S3). Большинство неабдоминальных образцов были взяты у молодых пациентов, и, как и ожидалось, наблюдалась некоторая группировка образцов мышц в соответствии с их анатомическим происхождением. Эта кластеризация не влияет на наш анализ возрастной регуляции, поскольку анатомическое происхождение было включено в качестве термина в модель множественной регрессии.Таким образом, эти два метода не показали никаких доказательств того, что анатомические, патологические или фармацевтические факторы искажают результаты нашего исследования старения.
Таким образом, мы создали глобальный профиль изменений экспрессии генов при старении в мышцах человека (рис. 1). Хорошо известно, что старение оказывает множество эффектов на мышцы, такие как снижение физиологических показателей, изменения в морфологии и повышенная восприимчивость к заболеваниям. Данные на рисунке 1 расширяют наше понимание мышечного старения до уровня конкретных генов и генетических путей, обеспечивая понимание возможных механизмов, лежащих в основе общего снижения мышечной функции в пожилом возрасте.В целом, разница в экспрессии генов между молодой и старой мышечной тканью относительно невелика. В частности, только 250 генов обнаруживают значительные изменения в экспрессии с возрастом ( p < 0,001), и подавляющее большинство этих регулируемых возрастом генов изменяют экспрессию менее чем в два раза за 50 лет. Эти результаты согласуются с моделью, в которой возрастное снижение клеточных функций вызывается накоплением множественных незначительных изменений в регуляции генов и путей.
Данные на рисунке 1 расширяют наше понимание мышечного старения до уровня конкретных генов и генетических путей, обеспечивая понимание возможных механизмов, лежащих в основе общего снижения мышечной функции в пожилом возрасте.В целом, разница в экспрессии генов между молодой и старой мышечной тканью относительно невелика. В частности, только 250 генов обнаруживают значительные изменения в экспрессии с возрастом ( p < 0,001), и подавляющее большинство этих регулируемых возрастом генов изменяют экспрессию менее чем в два раза за 50 лет. Эти результаты согласуются с моделью, в которой возрастное снижение клеточных функций вызывается накоплением множественных незначительных изменений в регуляции генов и путей.
Генетические функции многих из 250 генов, показанных на рис. 1, известны, и некоторые предполагают наличие биологических механизмов, которые могут вызывать возрастное ухудшение физиологии мышц.Например, CYP26B1 демонстрирует среднее увеличение экспрессии на 90% за 50 лет.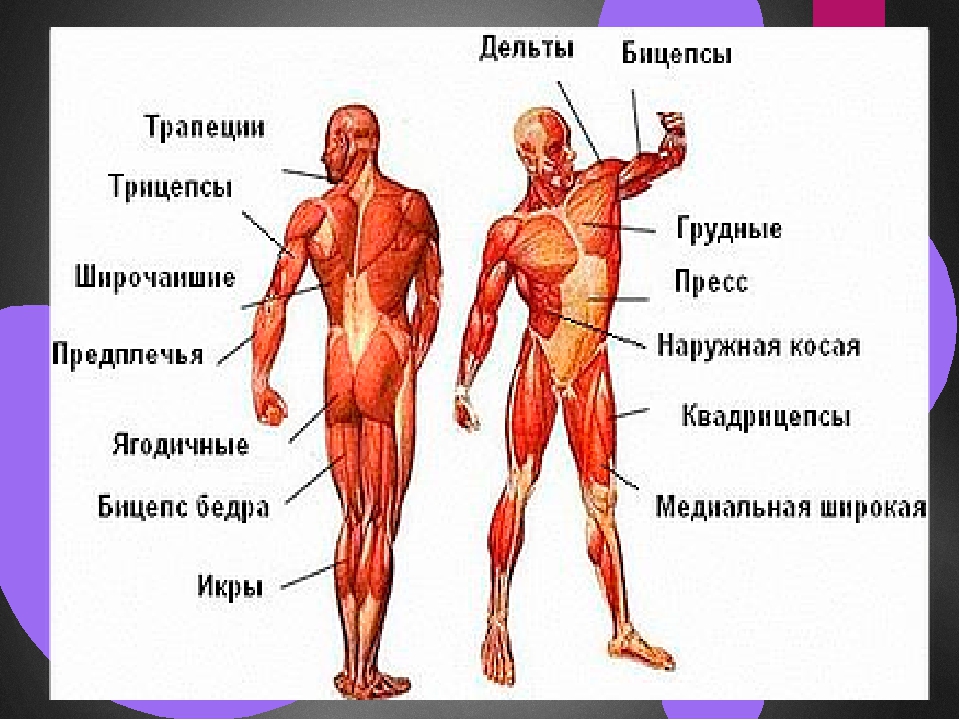 CYP26B1 является членом семейства цитохромов P450, которые представляют собой монооксигеназы, используемые для метаболизма токсичных веществ. Повышенная экспрессия CYP26B1 в пожилом возрасте может способствовать устранению токсинов, которые накапливаются с возрастом.
CYP26B1 является членом семейства цитохромов P450, которые представляют собой монооксигеназы, используемые для метаболизма токсичных веществ. Повышенная экспрессия CYP26B1 в пожилом возрасте может способствовать устранению токсинов, которые накапливаются с возрастом.
LASS5 снижает экспрессию примерно на 25% в течение 50 лет. LASS5 является человеческим ортологом гена обеспечения долголетия дрожжей lag1 .У дрожжей экспрессия lag1 снижается в старых дрожжевых клетках [21], аналогично нашим результатам, показывающим снижение экспрессии в пожилом возрасте в мышцах человека. LASS5 участвует в сигнальном пути церамидов, который играет важную роль в нескольких процессах, связанных с продолжительностью жизни, таких как стрессоустойчивость и апоптоз [22]. Снижение экспрессии LASS5 в пожилом возрасте может нарушать функцию клеток за счет снижения передачи сигналов церамидов.
В дополнение к поиску гена регуляции возраста по одному за раз, мы также провели скрининг известных генетических путей на наличие тех, которые показывают общее изменение с возрастом.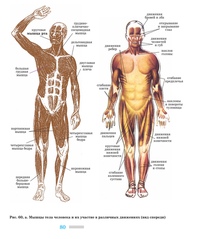 При таком подходе возрастная регуляция для каждого гена в пути комбинируется, чтобы определить, существует ли общая регуляция всего пути. Скрининг скоординированной возрастной регуляции генетических путей повышает чувствительность нашего анализа, поскольку комбинированные эффекты небольшой регуляции многих генов в пути могут быть значительными. Например, в предыдущем исследовании диабета 2 типа скрининг генетических путей на предмет изменений в экспрессии дал ключевую информацию, которая была невозможна при индивидуальном анализе генов [23].
При таком подходе возрастная регуляция для каждого гена в пути комбинируется, чтобы определить, существует ли общая регуляция всего пути. Скрининг скоординированной возрастной регуляции генетических путей повышает чувствительность нашего анализа, поскольку комбинированные эффекты небольшой регуляции многих генов в пути могут быть значительными. Например, в предыдущем исследовании диабета 2 типа скрининг генетических путей на предмет изменений в экспрессии дал ключевую информацию, которая была невозможна при индивидуальном анализе генов [23].
Мы разработали вариант анализа обогащения набора генов (GSEA), чтобы определить, свидетельствует ли генетический путь о возрастной регуляции [23]. Мы проанализировали 624 набора генов, определенных консорциумом Gene Ontology [24] (таблица S3). Мы модифицировали первоначальную парадигму GSEA, поскольку она предназначалась для наборов данных с двумя категориями выборки, и вместо этого подгоняли регрессионные модели к постоянно меняющимся независимым и зависимым переменным.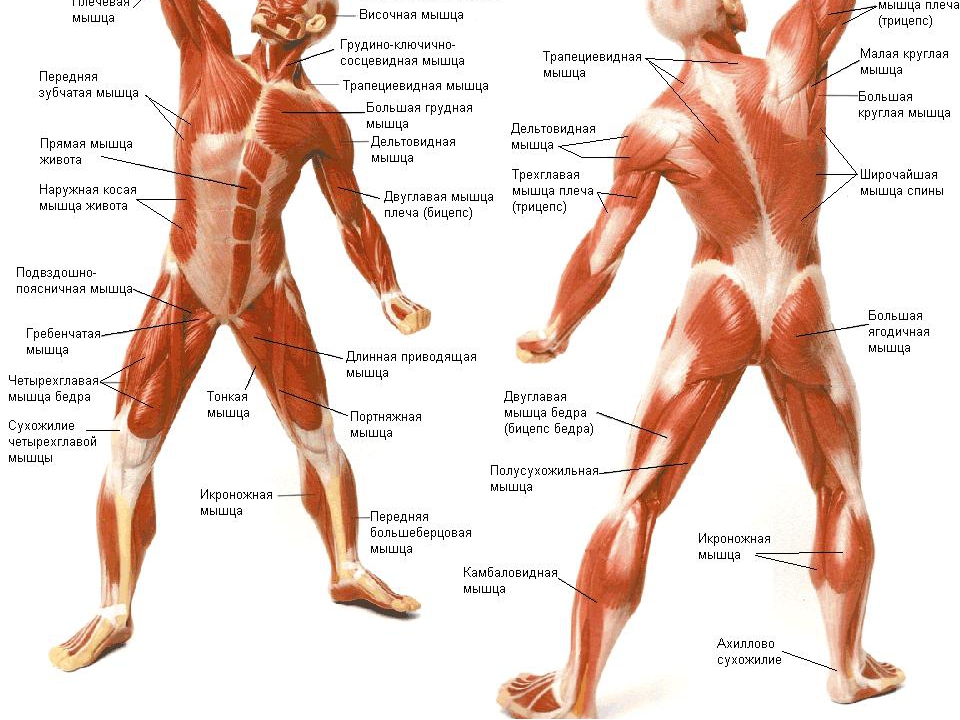 Соответственно, мы заменили двухвыборочную тестовую статистику в GSEA расчетным наклоном регрессии для возраста.Мы также заменили статистику Колмогорова-Смирнова статистикой Ван дер Вардена, потому что предпочитаем тот тип зависимости, который отражает статистика Ван дер Вардена. Наконец, мы заменили стратегию перестановки бутстрапом, чтобы лучше обрабатывать ковариаты (Материалы и методы).
Соответственно, мы заменили двухвыборочную тестовую статистику в GSEA расчетным наклоном регрессии для возраста.Мы также заменили статистику Колмогорова-Смирнова статистикой Ван дер Вардена, потому что предпочитаем тот тип зависимости, который отражает статистика Ван дер Вардена. Наконец, мы заменили стратегию перестановки бутстрапом, чтобы лучше обрабатывать ковариаты (Материалы и методы).
Наша версия алгоритма GSEA оценивает набор генов в соответствии с тем, как гены в нем демонстрируют скоординированное увеличение (или уменьшение) в среднем в ответ на увеличение возраста. Увеличение измеряется статистикой Ван дер Вардена.Чтобы судить о том, значима ли конкретная статистика Ван дер Вардена, мы использовали повторную выборку начальной загрузки. Каждый бутстреп-образец был составлен путем повторной выборки массивов и сохранения измерений экспрессии генов, связанных с возрастом, полом и переменными анатомии. 624 балла Ван дер Вардена для групп генов были пересчитаны для каждой из 1000 бутстрэп-выборок. Было обнаружено, что шесть наборов генов имеют статистически значимые оценки Ван дер Вардена ( p <0,001) в этой повторной выборке. При этом пороге значения p мы ожидаем найти только 0.6 наборов генов случайно. Мы также требовали, чтобы группы генов демонстрировали некоторую практическую значимость, отбрасывая группы с показателем Ван-дер-Вардена менее 3,1 по абсолютной величине. Мы обнаружили три пути, которые соответствовали обоим критериям: симпортерные гены, сиалилтрансферазы и переносчики хлоридов показали снижение экспрессии с возрастом (рис. 2 и табл. 2). Коэффициенты старения для всех генов в этих путях перечислены в таблице S4.
Было обнаружено, что шесть наборов генов имеют статистически значимые оценки Ван дер Вардена ( p <0,001) в этой повторной выборке. При этом пороге значения p мы ожидаем найти только 0.6 наборов генов случайно. Мы также требовали, чтобы группы генов демонстрировали некоторую практическую значимость, отбрасывая группы с показателем Ван-дер-Вардена менее 3,1 по абсолютной величине. Мы обнаружили три пути, которые соответствовали обоим критериям: симпортерные гены, сиалилтрансферазы и переносчики хлоридов показали снижение экспрессии с возрастом (рис. 2 и табл. 2). Коэффициенты старения для всех генов в этих путях перечислены в таблице S4.
Рис. 2. Три набора генов регулируются возрастом в мышцах
Строки представляют наборы генов симпортера, активности сиалилтрансферазы и транспорта хлоридов.Столбцы соответствуют отдельным генам в данном наборе генов. Шкала представляет наклон изменения уровня экспрессии log 2 с возрастом ( β 1j ). Навигационная версия этого рисунка, показывающая идентичность конкретных генов, может быть найдена по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
Навигационная версия этого рисунка, показывающая идентичность конкретных генов, может быть найдена по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
https://doi.org/10.1371/journal.pgen.0020115.g002
Гены Symporter (63 гена) и переносчики хлоридов (35 генов) необходимы для транспорта растворенных веществ при мышечном сокращении [25]; снижение уровня экспрессии этих транспортеров может быть связано со слабостью старых мышц.Гены с сиалилтрансферазной активностью (19 генов) опосредуют гликозилирование путем переноса групп сиаловой кислоты на секретируемые молекулы. Снижение активности сиалилтрансферазы ранее было обнаружено в сыворотке стареющего человека [26], нейронах [27] и лимфоцитах [28].
Молекулярные маркеры физиологического старения
Некоторые люди стареют медленно и остаются сильными и здоровыми в свои 70 лет, в то время как другие стареют быстро, становятся слабыми и восприимчивыми к возрастным заболеваниям. Мы хотели определить, коррелирует ли профиль экспрессии 250 генов, регулирующих старение, с физиологическим в дополнение к хронологическому старению. Например, пациенту V17 был 41 год, но его возраст-регулируемые гены проявлялись так же, как и у пациентов, которые были на 10–20 лет старше, и мы хотели бы определить, была ли у этого пациента плохая мышечная физиология для его возраста (рис. 1). Наоборот, пациенту М73 было 64 года, но его молекулярный профиль был подобен другим пациентам, которые были на 30 лет моложе, и мы хотели определить, имеет ли этот пациент относительно хорошую мышечную физиологию для своего возраста. Наш список состоит из 250 генов, которые значительно коррелируют с хронологическим возрастом.Мы стремились определить, коррелируют ли они также с физиологическим возрастом, измеряемым соотношением диаметров типа II/типа I. Мы подготовили гистологические срезы для всех 81 образцов скелетных мышц и смогли надежно измерить диаметры мышечных волокон типа I и типа II для 32 образцов (рис. 3А и 3В; таблица S5).
Например, пациенту V17 был 41 год, но его возраст-регулируемые гены проявлялись так же, как и у пациентов, которые были на 10–20 лет старше, и мы хотели бы определить, была ли у этого пациента плохая мышечная физиология для его возраста (рис. 1). Наоборот, пациенту М73 было 64 года, но его молекулярный профиль был подобен другим пациентам, которые были на 30 лет моложе, и мы хотели определить, имеет ли этот пациент относительно хорошую мышечную физиологию для своего возраста. Наш список состоит из 250 генов, которые значительно коррелируют с хронологическим возрастом.Мы стремились определить, коррелируют ли они также с физиологическим возрастом, измеряемым соотношением диаметров типа II/типа I. Мы подготовили гистологические срезы для всех 81 образцов скелетных мышц и смогли надежно измерить диаметры мышечных волокон типа I и типа II для 32 образцов (рис. 3А и 3В; таблица S5).
Рис. 3. Экспрессия генов предсказывает физиологию старения
(A) Поперечное сечение гистологически ничем не примечательной дельтовидной мышцы 48-летней женщины, демонстрирующее относительно эквивалентные размеры мышечных волокон I и II типов.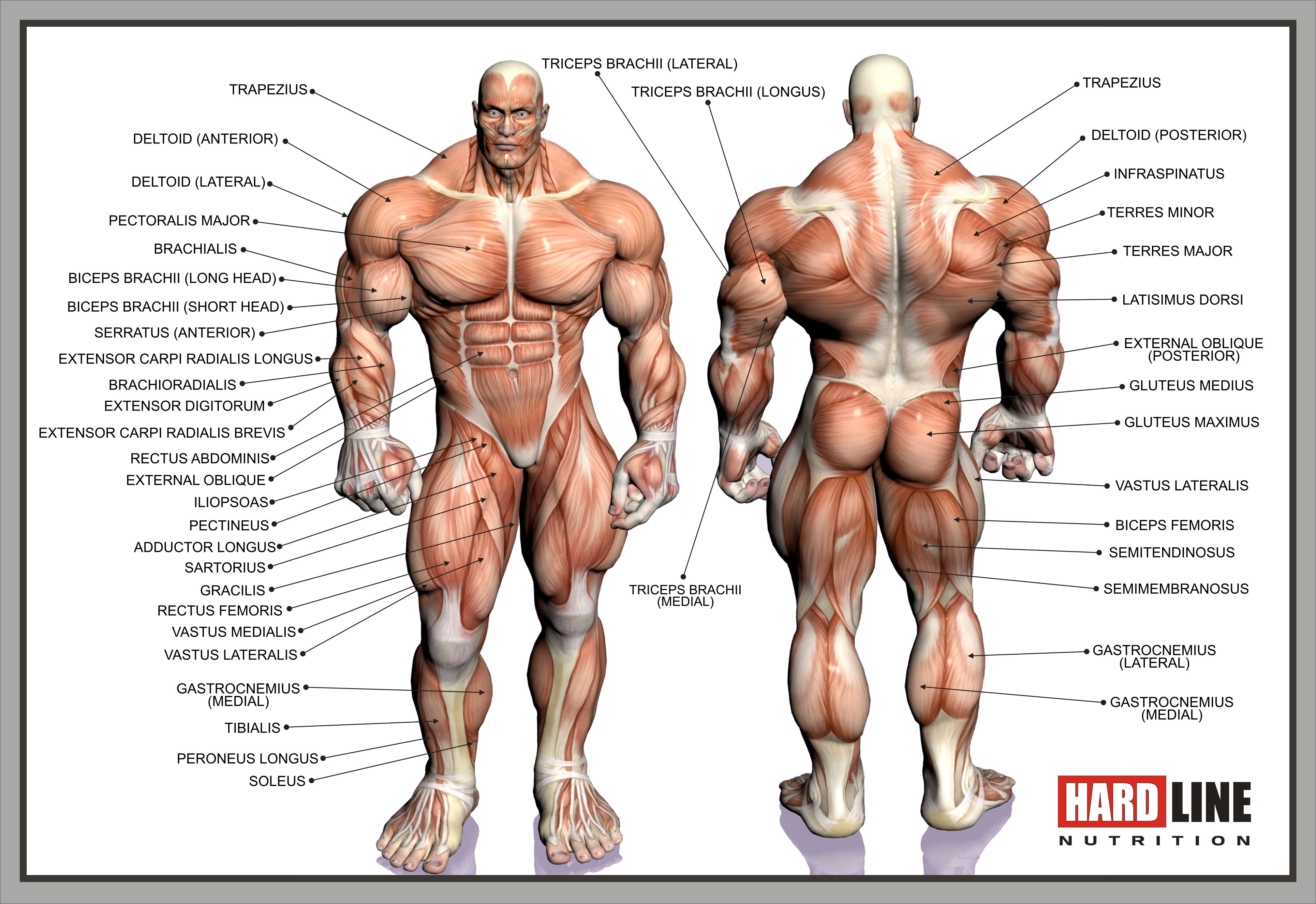 Стрелки обозначают типы волокон, различаемые по ферментативной гистохимии (криосрез, 200×, миозин-АТФаза при рН 9,4).
Стрелки обозначают типы волокон, различаемые по ферментативной гистохимии (криосрез, 200×, миозин-АТФаза при рН 9,4).
(B) Поперечный срез дельтовидной мышцы 88-летней женщины, демонстрирующий селективную атрофию мышечных волокон типа II, которая темнеет при гистохимическом анализе фермента АТФазы (криосрез, 200×, миозин-АТФаза при рН 9,4).
(C) Гистограммы, показывающие корреляцию между физиологией мышц и экспрессией генов, регулируемых возрастом. Верхняя панель: для каждого из 250 генов, регулируемых возрастом, мы рассчитали коэффициенты частичной корреляции между соотношением диаметров мышечных волокон типа II/типа I и экспрессией генов без учета возрастных изменений (ось x ).Нижняя панель: то же, что и верхняя панель, за исключением того, что коэффициенты корреляции были рассчитаны для всех 31 948 генов. Квадрат частичного коэффициента корреляции обозначает степень, на которую изменения в экспрессии генов объясняют вариации в соотношении диаметров мышечных волокон типа II/типа I, исключая влияние возраста.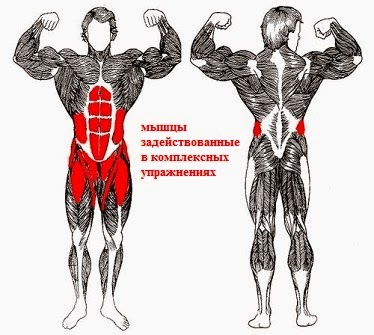
(D) Гистограмма, показывающая вероятность обнаружения 92 генов с | р | > 0,2 из набора случайных генов. Мы провели эксперимент Монте-Карло, случайным образом выбрав наборы из 250 генов из генома и подсчитав, сколько генов в наборе имеют | р | > 0.2 как в (С). Процедура была повторена 1000 раз, и гистограмма показывает количество генов из каждой случайной выборки, которые имеют | р | > 0,2. Стрелка показывает количество генов, превышающих этот порог (92) из набора 250 генов, регулируемых возрастом ( p < 0,001). Мы также определили общее количество генов в геноме с | р | > 0,2, а затем показал, что 92 гена из набора из 250 значимы (гипергеометрическое распределение; p < 1 × 10 -4 ).
https://doi.org/10.1371/journal.pgen.0020115.g003
Для наших целей недостаточно простой корреляции экспрессии генов с соотношением типов мышц. Такая корреляция может возникнуть просто потому, что экспрессия генов и соотношение типов мышц коррелируют с возрастом. Соответственно, мы использовали частичные корреляции экспрессии генов с соотношениями типов мышц после поправки на влияние хронологического возраста. Для этого мы регрессировали соотношение диаметров мышечных волокон типа II/типа I в зависимости от возраста, регрессировали экспрессию генов в зависимости от возраста и, наконец, коррелировали остатки обеих регрессий, чтобы получить коэффициенты частичной корреляции.Частичные корреляции для 250 связанных с возрастом генов показаны на рисунке 3C.
Соответственно, мы использовали частичные корреляции экспрессии генов с соотношениями типов мышц после поправки на влияние хронологического возраста. Для этого мы регрессировали соотношение диаметров мышечных волокон типа II/типа I в зависимости от возраста, регрессировали экспрессию генов в зависимости от возраста и, наконец, коррелировали остатки обеих регрессий, чтобы получить коэффициенты частичной корреляции.Частичные корреляции для 250 связанных с возрастом генов показаны на рисунке 3C.
Если ген коррелирует с коэффициентом диаметра мышц только потому, что и он, и диаметр мышц коррелируют с возрастом, то описанная выше частичная корреляция должна быть близка к нулю. Мы обнаружили, что большое количество генов в нашем списке имело статистически значимую связь с соотношением типа II/типа I после поправки на возраст. Однако многие из генов, не включенных в наш список, также были связаны с соотношением типа II/типа I с поправкой на возраст.Мы смогли показать, что гены с большими частичными корреляциями были значительно перепредставлены в нашем списке из 250 генов, регулируемых возрастом. Мы насчитали 92 из 250 связанных с возрастом генов, для которых (абсолютная) частичная корреляция составила более 0,2 (таблица S6). Только 7 768 из 31 948 генов не попали в список с такой большой частичной корреляцией. Используя гипергеометрическое распределение, мы нашли значение p ниже 0,0001 и пришли к выводу, что возрастные гены с большей вероятностью, чем другие гены, имеют некоторую частичную корреляцию с отношением диаметра мышц.Чтобы проиллюстрировать этот эффект, мы также отобрали 250 генов из генома 1000 раз, каждый раз подсчитывая, сколько из них имеют частичную корреляцию более 0,2 по абсолютной величине. Ни в одном из образцов количество не превышало 92 (рис. 3D).
Мы насчитали 92 из 250 связанных с возрастом генов, для которых (абсолютная) частичная корреляция составила более 0,2 (таблица S6). Только 7 768 из 31 948 генов не попали в список с такой большой частичной корреляцией. Используя гипергеометрическое распределение, мы нашли значение p ниже 0,0001 и пришли к выводу, что возрастные гены с большей вероятностью, чем другие гены, имеют некоторую частичную корреляцию с отношением диаметра мышц.Чтобы проиллюстрировать этот эффект, мы также отобрали 250 генов из генома 1000 раз, каждый раз подсчитывая, сколько из них имеют частичную корреляцию более 0,2 по абсолютной величине. Ни в одном из образцов количество не превышало 92 (рис. 3D).
Наш результат, показывающий, что 250 генов, регулируемых возрастом, обогащены генами, регулируемыми соотношением диаметров мышечных волокон типа II/типа I, действителен, даже когда мы используем другие пороги отбора для физиологии мышц (т. е. отличные от абсолютного значения r > 0. 2). Мы сравнили распределение частичных корреляций 250 генов, регулируемых возрастом, с соотношением типа II/типа I с распределением частичных корреляций остальных генов в геноме, используя непараметрические методы (рис. 3C). Используя критерий согласия Колмогорова-Смирнова, мы обнаружили, что распределение 250 генов, регулируемых возрастом, шире, чем общее распределение в двустороннем тесте ( p < 1 × 10 −15 , с Д = 0,27). Этот результат указывает на то, что кажущаяся физиологическая основа нашего набора генов не является следствием того, что мы выбрали 0.2 в качестве порога.
2). Мы сравнили распределение частичных корреляций 250 генов, регулируемых возрастом, с соотношением типа II/типа I с распределением частичных корреляций остальных генов в геноме, используя непараметрические методы (рис. 3C). Используя критерий согласия Колмогорова-Смирнова, мы обнаружили, что распределение 250 генов, регулируемых возрастом, шире, чем общее распределение в двустороннем тесте ( p < 1 × 10 −15 , с Д = 0,27). Этот результат указывает на то, что кажущаяся физиологическая основа нашего набора генов не является следствием того, что мы выбрали 0.2 в качестве порога.
Таким образом, эти статистические тесты показывают, что набор генов, регулируемых возрастом, является маркером относительного уровня мышечной функции даже среди пациентов одного возраста. Наши выводы дополнительно подтверждаются двумя дополнительными статистическими тестами, описанными в разделе «Материалы и методы» (таблицы S7 и S8). Таким образом, возраст-регулируемые гены обогащены теми, которые предсказывают физиологический, а не только хронологический возраст. Корреляцию между профилем экспрессии генов и физиологическим возрастом можно увидеть у пациентов V17 и M73 на рисунке 1.Хотя пациент V17 относительно молод (41 год), профиль экспрессии 250 генов, регулируемых возрастом, наиболее похож на пожилых людей, а соотношение диаметров мышечных волокон типа II и типа I низкое для его возраста. И наоборот, хотя пациент М73 относительно стар (64 года), картина экспрессии генов аналогична таковой у более молодых людей, а соотношение диаметров мышечных волокон типа II/типа I высокое для его возраста (рис. 1).
Корреляцию между профилем экспрессии генов и физиологическим возрастом можно увидеть у пациентов V17 и M73 на рисунке 1.Хотя пациент V17 относительно молод (41 год), профиль экспрессии 250 генов, регулируемых возрастом, наиболее похож на пожилых людей, а соотношение диаметров мышечных волокон типа II и типа I низкое для его возраста. И наоборот, хотя пациент М73 относительно стар (64 года), картина экспрессии генов аналогична таковой у более молодых людей, а соотношение диаметров мышечных волокон типа II/типа I высокое для его возраста (рис. 1).
Общий признак старения мышц, почек и мозга
Некоторые аспекты старения влияют только на определенные ткани; примеры включают прогрессирующую слабость мышц, снижение синаптической функции в головном мозге или снижение скорости фильтрации в почках.Другие аспекты старения проявляются во всех клетках, независимо от типа их ткани, например, накопление окислительного повреждения митохондрий, повреждение ДНК и повреждение белка. Наш полногеномный поиск изменений экспрессии генов во время старения будет включать оба типа изменений экспрессии, и было бы интересно определить, какие изменения экспрессии специфичны для мышц, а какие являются общими для всех тканей. Профили экспрессии, которые являются общими для старения во всех тканях, могут обеспечить понимание основных механизмов, лежащих в основе клеточного старения.Поэтому мы сравнили данные экспрессии ДНК-чипов из наших исследований старения мышц с предыдущими исследованиями экспрессии ДНК-чипов при старении в головном мозге и почках. Родуэлл и др. охарактеризовали изменения экспрессии генов с возрастом в корковом и мозговом веществе почек у 74 пациентов, а Lu et al. исследовали изменения экспрессии генов в лобной коре головного мозга у 30 пациентов [5,6].
Наш полногеномный поиск изменений экспрессии генов во время старения будет включать оба типа изменений экспрессии, и было бы интересно определить, какие изменения экспрессии специфичны для мышц, а какие являются общими для всех тканей. Профили экспрессии, которые являются общими для старения во всех тканях, могут обеспечить понимание основных механизмов, лежащих в основе клеточного старения.Поэтому мы сравнили данные экспрессии ДНК-чипов из наших исследований старения мышц с предыдущими исследованиями экспрессии ДНК-чипов при старении в головном мозге и почках. Родуэлл и др. охарактеризовали изменения экспрессии генов с возрастом в корковом и мозговом веществе почек у 74 пациентов, а Lu et al. исследовали изменения экспрессии генов в лобной коре головного мозга у 30 пациентов [5,6].
Наша первоначальная попытка сравнить транскрипционные изменения между тканями основывалась на анализе Венна, в котором мы напрямую сравнивали перекрытия в списках генов, регулируемых возрастом, из трех тканей. Затем мы искали общий признак старения, сравнивая корреляцию Пирсона возрастной регуляции между двумя тканями. Оба этих простых метода продемонстрировали лишь пограничные статистические данные о сходстве старения между тремя тканями (Материалы и методы), но ни один из них не может быть мощным. В конечном счете, мы сравнили ткани, используя групповой анализ генов. Группировка генов может быть более эффективной, если в каждом из нескольких генов есть небольшие, но устойчивые эффекты. Кроме того, конкретные биологические процессы, связанные с каждым генетическим путем, позволяют лучше понять механизмы старения.Мы использовали описанный выше модифицированный GSEA для анализа ранее опубликованных данных о возрастной регуляции в почках и головном мозге [5,6]. Как и раньше, мы рассмотрели возможность того, что наблюдаемые корреляции могут быть связаны с тем, что могут быть случайные различия выборки в разных тканях, которые скоординировано влияют на уровни экспрессии генов во всем наборе генов.
Затем мы искали общий признак старения, сравнивая корреляцию Пирсона возрастной регуляции между двумя тканями. Оба этих простых метода продемонстрировали лишь пограничные статистические данные о сходстве старения между тремя тканями (Материалы и методы), но ни один из них не может быть мощным. В конечном счете, мы сравнили ткани, используя групповой анализ генов. Группировка генов может быть более эффективной, если в каждом из нескольких генов есть небольшие, но устойчивые эффекты. Кроме того, конкретные биологические процессы, связанные с каждым генетическим путем, позволяют лучше понять механизмы старения.Мы использовали описанный выше модифицированный GSEA для анализа ранее опубликованных данных о возрастной регуляции в почках и головном мозге [5,6]. Как и раньше, мы рассмотрели возможность того, что наблюдаемые корреляции могут быть связаны с тем, что могут быть случайные различия выборки в разных тканях, которые скоординировано влияют на уровни экспрессии генов во всем наборе генов. Чтобы контролировать эту возможность, мы провели повторную выборку данных микрочипа 1000 раз (с заменой) и повторили анализ 624 наборов генов при каждой повторной выборке.Мы выбрали только те наборы генов, которые были статистически значимы во всех трех тканях при p < 0,05. Затем мы удалили любой набор генов, который не достиг абсолютного значения по шкале Ван-дер-Вардена 1,65 или более во всех трех тканях. Из 624 наборов генов мы обнаружили, что гены внеклеточного матрикса, гены клеточного роста и гены активации комплемента значительно увеличивают экспрессию с возрастом в среднем во всех трех тканях человека, тогда как гены транспорта хлоридов и гены транспорта электронов значительно снижают экспрессию в среднем. с возрастом в тех же тканях (табл. 3).Цитозольный рибосомный путь показал повышенную экспрессию, которая была значительной в мышцах и почках и почти значительной в головном мозге (bootstrap p <0,06). Коэффициенты старения для всех генов в каждом из этих путей перечислены в таблице S9.
Чтобы контролировать эту возможность, мы провели повторную выборку данных микрочипа 1000 раз (с заменой) и повторили анализ 624 наборов генов при каждой повторной выборке.Мы выбрали только те наборы генов, которые были статистически значимы во всех трех тканях при p < 0,05. Затем мы удалили любой набор генов, который не достиг абсолютного значения по шкале Ван-дер-Вардена 1,65 или более во всех трех тканях. Из 624 наборов генов мы обнаружили, что гены внеклеточного матрикса, гены клеточного роста и гены активации комплемента значительно увеличивают экспрессию с возрастом в среднем во всех трех тканях человека, тогда как гены транспорта хлоридов и гены транспорта электронов значительно снижают экспрессию в среднем. с возрастом в тех же тканях (табл. 3).Цитозольный рибосомный путь показал повышенную экспрессию, которая была значительной в мышцах и почках и почти значительной в головном мозге (bootstrap p <0,06). Коэффициенты старения для всех генов в каждом из этих путей перечислены в таблице S9. Мы ожидаем, что 0,08 (практически ни один) из 624 путей обычно не регулируются возрастом случайно (90 045 p 90 048 < 0,05 во всех трех тканях, и, следовательно, в сумме 90 045 p 90 048 < 1,25 × 10 90 046 -4 90 047).
Мы ожидаем, что 0,08 (практически ни один) из 624 путей обычно не регулируются возрастом случайно (90 045 p 90 048 < 0,05 во всех трех тканях, и, следовательно, в сумме 90 045 p 90 048 < 1,25 × 10 90 046 -4 90 047).
Увеличение общей экспрессии набора генов внеклеточного матрикса (152 гена) с возрастом может способствовать распространению фиброза у пожилых людей (рис. 4).Фиброз — это процесс, при котором волокнистая соединительная ткань разрастается по органам и нарушает функцию многих тканей. TIMP1, , который кодирует тканевой ингибитор металлопротеиназы 1, показывает наибольшее увеличение экспрессии с возрастом (в среднем 236% в 50 лет).
Рис. 4. Общие признаки старения мышц, почек и мозга цепные гены.Ряды – ткани человека (М – мышцы, К – почки, Б – головной мозг). Столбцы соответствуют отдельным генам в каждом наборе генов. Шкала представляет наклон изменения уровня экспрессии log 2 с возрастом (β 1j ). Серый цвет означает, что в наборе данных отсутствуют гены. Навигационную версию, показывающую идентичность конкретных генов, можно найти по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
Навигационную версию, показывающую идентичность конкретных генов, можно найти по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
https://doi.org/10.1371/journal.pgen.0020115.g004
Набор генов роста клеток (29 генов) включает гены, кодирующие факторы роста, такие как TGFB1, и FGFR1. Индукция генов в этом наборе генов может отражать попытку восстановить повреждения тканей, которые накапливаются в течение жизни.
Хотя гены активации комплемента (22 гена) индуцируются в мышцах, почках и головном мозге, они преимущественно экспрессируются в печени [29]. Следовательно, если гены комплемента также не регулируются возрастом в печени, физиологическое значение возрастной регуляции генов комплемента в мышцах, почках и головном мозге в настоящее время неясно.
Цитозольные рибосомные гены включают 85 генов, экспрессия которых с возрастом увеличивается во всех трех тканях.Этот результат интересен тем, что известно, что скорость синтеза белка снижается в пожилом возрасте [30], и все же наши результаты экспрессии показывают увеличение экспрессии рибосомных генов.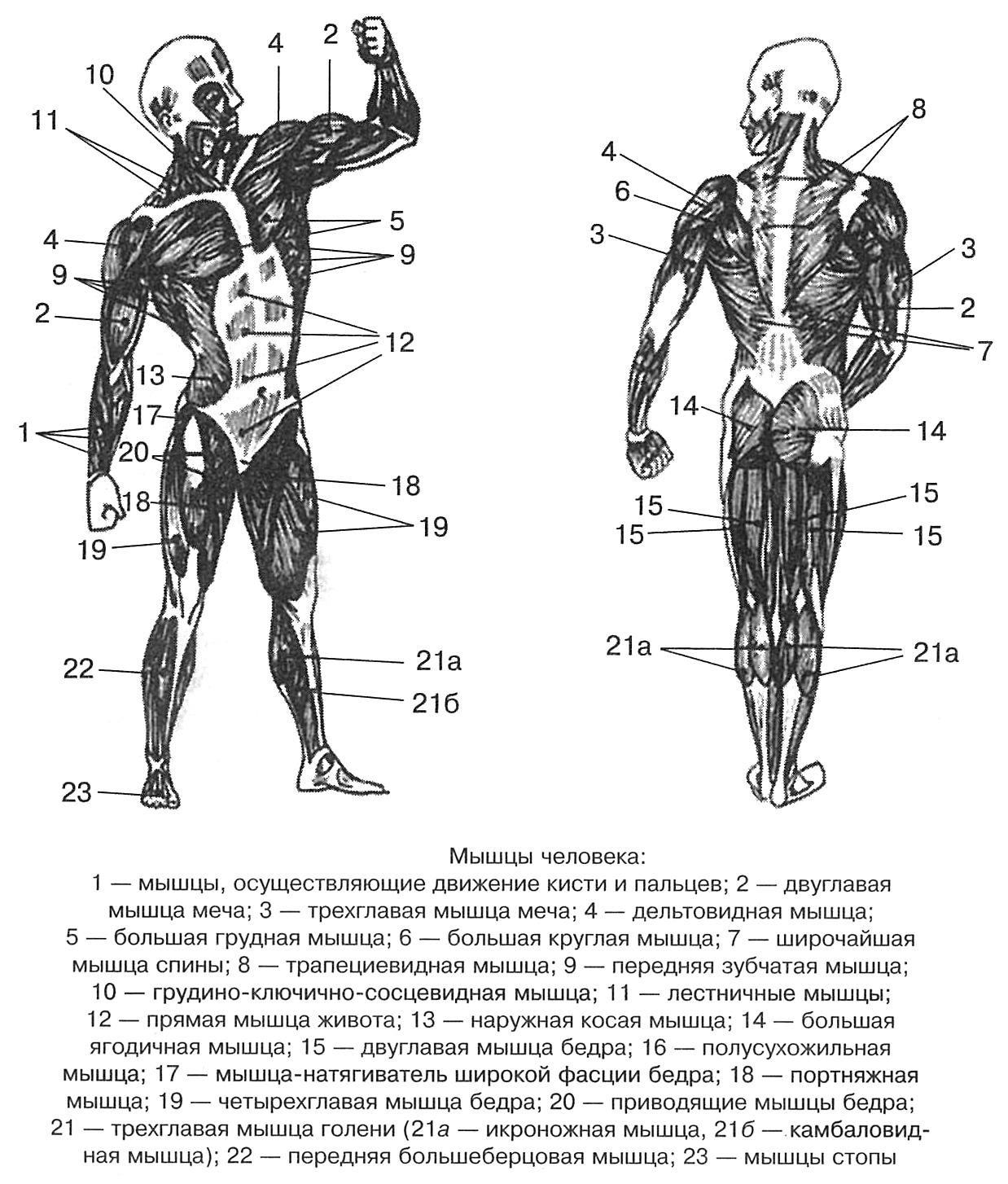 Одна возможность заключается в том, что сниженный синтез белка в старых клетках индуцирует экспрессию рибосомных генов как часть гомеостатической петли обратной связи, чтобы частично компенсировать потерю эффективности трансляции.
Одна возможность заключается в том, что сниженный синтез белка в старых клетках индуцирует экспрессию рибосомных генов как часть гомеостатической петли обратной связи, чтобы частично компенсировать потерю эффективности трансляции.
Путь транспорта хлоридов состоит из 35 генов, экспрессия которых с возрастом снижается во всех трех тканях.Ионный транспорт многих типов важен не только для сокращения мышц [25], но и для поддержания солевого баланса в почках [31] и функции нейронов в головном мозге через ГАМК-опосредованные рецепторы [32]. Снижение транспорта хлоридов с возрастом может привести ко многим типам физиологических нарушений, связанных с дефицитом транспорта ионов.
Было обнаружено, что митохондриальная цепь переноса электронов демонстрирует общее снижение экспрессии с возрастом. В эту группу входят 95 генов, в том числе гены, ассоциированные с семейством НАДН-дегидрогеназ (комплекс I), сукцинат-кофермент Q-редуктазы (комплекс II), убихинон-цитохром-с-редуктазы (комплекс III), цитохром-с-оксидазы (комплекс IV), H + -АТФ-синтаза (комплекс V) и разобщающие белки. Обнаружение того, что экспрессия генов, участвующих в цепи переноса электронов, снижается в пожилом возрасте, подтверждает митохондриальную свободнорадикальную теорию старения [33], поскольку образование свободных радикалов митохондриями предпочтительно повреждает белковый комплекс цепи переноса электронов. Снижение экспрессии генов переноса электронов (закодированных в ядре) может быть вызвано регуляцией обратной связи из-за повреждения белкового комплекса цепи переноса электронов. Другие белковые комплексы в митохондриях (например, митохондриальные рибосомные гены) не снижают экспрессию с возрастом.Таким образом, старение не оказывает общего влияния на гены, кодирующие митохондриальные компоненты, а скорее специфически влияет на экспрессию генов, являющихся частью цепи переноса электронов.
Обнаружение того, что экспрессия генов, участвующих в цепи переноса электронов, снижается в пожилом возрасте, подтверждает митохондриальную свободнорадикальную теорию старения [33], поскольку образование свободных радикалов митохондриями предпочтительно повреждает белковый комплекс цепи переноса электронов. Снижение экспрессии генов переноса электронов (закодированных в ядре) может быть вызвано регуляцией обратной связи из-за повреждения белкового комплекса цепи переноса электронов. Другие белковые комплексы в митохондриях (например, митохондриальные рибосомные гены) не снижают экспрессию с возрастом.Таким образом, старение не оказывает общего влияния на гены, кодирующие митохондриальные компоненты, а скорее специфически влияет на экспрессию генов, являющихся частью цепи переноса электронов.
Приведенные выше результаты показывают, что существует общая возрастная регуляция этих шести генетических путей в почках, мышцах и головном мозге. Затем мы определили, что имеется мало статистических доказательств корреляции возрастной регуляции отдельных генов в пути в одной ткани с их возрастной регуляцией в другой ткани (Материалы и методы). Таким образом, неясно, проявляют ли одни и те же гены или разные гены в пределах пути возрастную регуляцию между разными тканями. Например, некоторые гены в пути транспорта электронов могут регулироваться возрастом в почках, тогда как другие гены транспорта электронов могут регулироваться возрастом в мышцах.
Таким образом, неясно, проявляют ли одни и те же гены или разные гены в пределах пути возрастную регуляцию между разными тканями. Например, некоторые гены в пути транспорта электронов могут регулироваться возрастом в почках, тогда как другие гены транспорта электронов могут регулироваться возрастом в мышцах.
Публичный возрастной путь у людей, мышей и мух
Идентифицировав генетические пути, которые обычно регулируются возрастом в различных тканях человека, мы затем определили, является ли их регуляция возраста специфичной для человека (частная) или эти группы также регулируются возрастом у других видов (общедоступная).Генетические пути, которые регулируются возрастом у разных видов, будут представлять особый интерес, поскольку они позволят выявить механизмы, неразрывно связанные со старением, даже у животных с совершенно разной продолжительностью жизни.
Мы сравнили регуляцию возраста у людей с ранее опубликованными исследованиями регуляции возраста у D. melanogaster [16] и C. elegans [18] . Чтобы изучить возрастную регуляцию в почках стареющих мышей, мы взяли образцы почек десяти мышей C57BL/6 в возрасте 1, 6, 16 и 24 месяцев, всего 40 образцов почек мышей.РНК из каждой почки экстрагировали, метили P 33 -dCTP и гибридизовали с кДНК фильтрующими мембранами, содержащими 16 896 клонов кДНК, соответствующих 11 512 уникальным генам. Мы нормализовали значения экспрессии с помощью метода Z-score [34] и проанализировали возрастную регуляцию каждого гена с помощью модели множественной регрессии с учетом возраста и пола каждой мыши-донора. Таблица S10 показывает наклон экспрессии по отношению к возрасту для каждого гена.
elegans [18] . Чтобы изучить возрастную регуляцию в почках стареющих мышей, мы взяли образцы почек десяти мышей C57BL/6 в возрасте 1, 6, 16 и 24 месяцев, всего 40 образцов почек мышей.РНК из каждой почки экстрагировали, метили P 33 -dCTP и гибридизовали с кДНК фильтрующими мембранами, содержащими 16 896 клонов кДНК, соответствующих 11 512 уникальным генам. Мы нормализовали значения экспрессии с помощью метода Z-score [34] и проанализировали возрастную регуляцию каждого гена с помощью модели множественной регрессии с учетом возраста и пола каждой мыши-донора. Таблица S10 показывает наклон экспрессии по отношению к возрасту для каждого гена.
Сначала мы идентифицировали ортологи человеческих генов у каждого из трех других видов.Затем мы определили изменение экспрессии по отношению к возрасту для каждого гена у каждого вида, используя методы множественной регрессии, аналогичные тем, которые использовались для наших исследований старения в мышцах человека (Материалы и методы).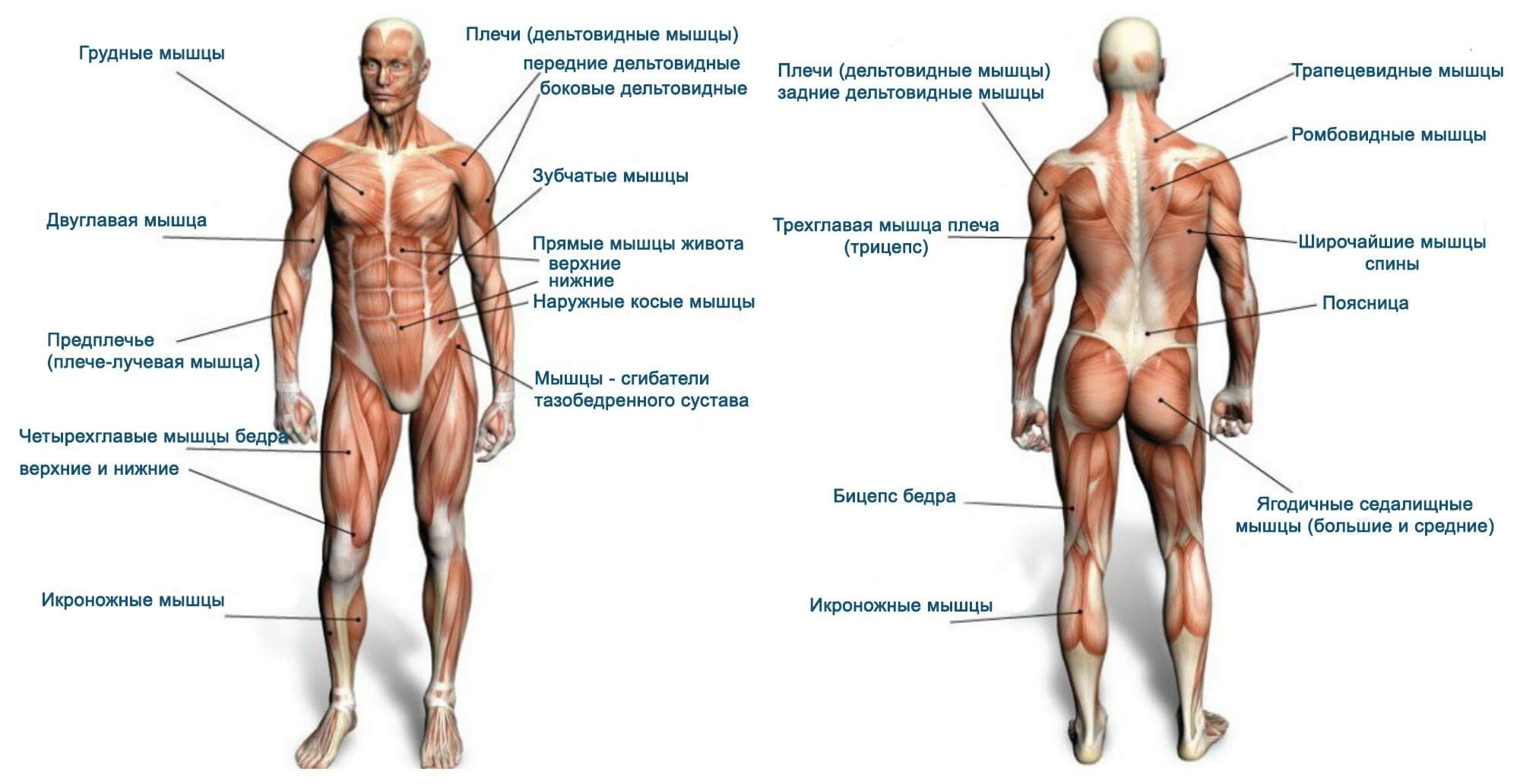 Мы взяли шесть наборов генов, которые, как было показано, регулируют старение в различных тканях человека, а затем спросили, проявляют ли они также регуляцию старения у любого из трех других видов. Мы проанализировали экспрессию каждого из наборов генов с использованием модифицированного GSEA, чтобы определить, демонстрируют ли они общее смещение в экспрессии с возрастом у каждого вида.Гены внеклеточного матрикса, гены клеточного роста, гены активации комплемента, гены цитозольных рибосом и гены транспорта хлора не проявляли возрастной регуляции у других видов.
Мы взяли шесть наборов генов, которые, как было показано, регулируют старение в различных тканях человека, а затем спросили, проявляют ли они также регуляцию старения у любого из трех других видов. Мы проанализировали экспрессию каждого из наборов генов с использованием модифицированного GSEA, чтобы определить, демонстрируют ли они общее смещение в экспрессии с возрастом у каждого вида.Гены внеклеточного матрикса, гены клеточного роста, гены активации комплемента, гены цитозольных рибосом и гены транспорта хлора не проявляли возрастной регуляции у других видов.
Гены транспортной цепи электронов показали постоянное общее снижение экспрессии с возрастом у людей, мышей и Drosophila, , но не показали значительной возрастной регуляции у C. elegans (рис. 5 и табл. 4). Чтобы показать, что возрастная регуляция вряд ли связана со случайной ошибкой биологической выборки, мы повторили выборку набора данных транспорта электронов у каждого вида с заменой и обнаружили, что гены цепи транспорта электронов показали значительную возрастную регуляцию у мышей ( p < 0. 02) и мух ( p < 0,001), но не C. elegans. Набор генов цепи переноса электронов также демонстрирует высокий показатель Ван-дер-Вардена у мышей и мух (менее -3,7). Таким образом, у людей, мышей и мух наблюдается снижение экспрессии цепи переноса электронов во время старения, что определяет общедоступный путь старения у видов с очень разной продолжительностью жизни. В C. elegans, неясно, связано ли отсутствие поддержки возрастной регуляции пути цепи переноса электронов с тем, что этот путь не регулируется возрастом, или с тем, что экспериментам с микрочипами ДНК не хватает статистической мощности для обнаружения возрастной регуляции.
02) и мух ( p < 0,001), но не C. elegans. Набор генов цепи переноса электронов также демонстрирует высокий показатель Ван-дер-Вардена у мышей и мух (менее -3,7). Таким образом, у людей, мышей и мух наблюдается снижение экспрессии цепи переноса электронов во время старения, что определяет общедоступный путь старения у видов с очень разной продолжительностью жизни. В C. elegans, неясно, связано ли отсутствие поддержки возрастной регуляции пути цепи переноса электронов с тем, что этот путь не регулируется возрастом, или с тем, что экспериментам с микрочипами ДНК не хватает статистической мощности для обнаружения возрастной регуляции.
Рис. 5. Уменьшение экспрессии электрон-транспортной цепи с возрастом у людей, мышей и мух
Строки представляют либо ткани человека, либо модельные организмы. Столбцы соответствуют отдельным генам человека и гомологам генов человека, определяемым взаимными лучшими совпадениями BLAST у других видов. Шкала представляет собой нормированный наклон изменения уровня экспрессии log 2 с возрастом ( β 1j ). Данные по разным видам были нормализованы путем деления наклона экспрессии с возрастом на стандартное отклонение всех подобных наклонов в наборе данных.Серый цвет указывает на отсутствие генов у этого вида. Навигационная версия этого рисунка, показывающая идентичность конкретных генов, может быть найдена по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
Данные по разным видам были нормализованы путем деления наклона экспрессии с возрастом на стандартное отклонение всех подобных наклонов в наборе данных.Серый цвет указывает на отсутствие генов у этого вида. Навигационная версия этого рисунка, показывающая идентичность конкретных генов, может быть найдена по адресу http://cmgm.stanford.edu/~kimlab/aging_muscle.
https://doi.org/10.1371/journal.pgen.0020115.g005
Обсуждение
В этом исследовании мы создали транскрипционный профиль старения мышц человека с высоким разрешением. Велле и др. ранее использовали ДНК-чипы для профилирования изменений экспрессии мышц человека при старении [7,8], и эта работа расширяет их предыдущие исследования, поскольку мы использовали значительно больший размер выборки, что обеспечило гораздо более высокое статистическое разрешение.
Люди стареют с разной скоростью, особенно это касается мышечного старения. Некоторые остаются здоровыми и сильными, тогда как другие становятся хрупкими и слабыми, когда стареют.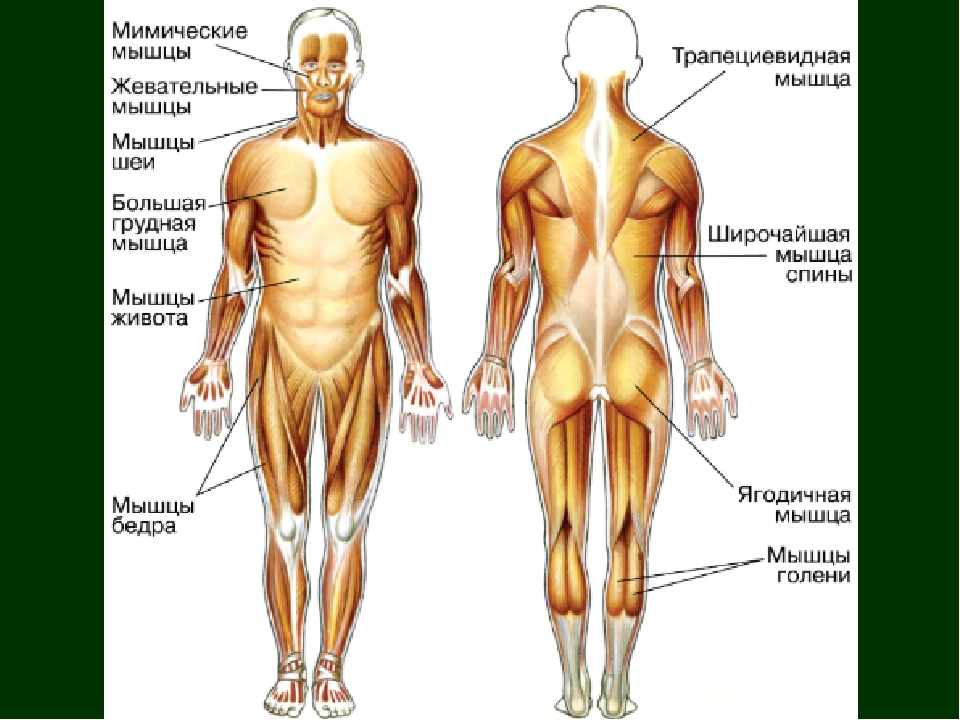 Профиль транскрипции старения в этом исследовании отражает физиологический возраст субъектов, измеренный по соотношению диаметров мышц после внесения поправки на их хронологический возраст. Предыдущая работа по регуляции возраста в почках также выявила молекулярные маркеры, которые могли предсказать физиологический возраст почки [5].
Профиль транскрипции старения в этом исследовании отражает физиологический возраст субъектов, измеренный по соотношению диаметров мышц после внесения поправки на их хронологический возраст. Предыдущая работа по регуляции возраста в почках также выявила молекулярные маркеры, которые могли предсказать физиологический возраст почки [5].
Наши результаты являются одними из первых доказательств общих признаков изменений экспрессии генов в различных тканях человека. В частности, мы обнаружили сходные закономерности возрастной регуляции для шести биологических путей в мышцах, почках и мозге. Предыдущие исследования выявили сходные закономерности старения между разными частями одной и той же ткани, но не между совершенно разными органами (т. е. было обнаружено, что возрастная регуляция одинакова между корой и мозговым веществом почки [5], а также между лобным полюсом и префронтальной долей). кора головного мозга [13]).
За исключением набора генов активации комплемента, пути, которые показывают общую регуляцию возраста в различных тканях, также функционируют во всех клетках. Изменения экспрессии этих путей в пожилом возрасте могут привести к дегенерации не только основных клеточных функций (таких как транспорт ионов и выработка энергии), но и к дегенерации тканеспецифических функций (таких как почечная фильтрация и синаптическая передача сигналов), которые зависят от «домашнего хозяйства». пути. Определив общий признак старения в тканях, мы теперь можем сосредоточиться на путях старения, которые являются общими, а не тканеспецифичными.Общий признак старения отражает возраст различных органов, тогда как гены, которые регулируются возрастом только в одной ткани, отражают возраст этой ткани. Наконец, можно ожидать, что лечение или терапия, которая изменяет экспрессию четырех общих возрастно-регулируемых путей, повлияет на различные ткани, а не на конкретную ткань, и, следовательно, может иметь общий эффект на продолжительность жизни.
Изменения экспрессии этих путей в пожилом возрасте могут привести к дегенерации не только основных клеточных функций (таких как транспорт ионов и выработка энергии), но и к дегенерации тканеспецифических функций (таких как почечная фильтрация и синаптическая передача сигналов), которые зависят от «домашнего хозяйства». пути. Определив общий признак старения в тканях, мы теперь можем сосредоточиться на путях старения, которые являются общими, а не тканеспецифичными.Общий признак старения отражает возраст различных органов, тогда как гены, которые регулируются возрастом только в одной ткани, отражают возраст этой ткани. Наконец, можно ожидать, что лечение или терапия, которая изменяет экспрессию четырех общих возрастно-регулируемых путей, повлияет на различные ткани, а не на конкретную ткань, и, следовательно, может иметь общий эффект на продолжительность жизни.
Несмотря на то, что некоторые закономерности старения в разных тканях человека сходны, большая часть старения является тканеспецифичной. Снижение экспрессии генов сиалилтрансфераз и симпортеров является изменениями, характерными для мышц, и, по-видимому, не происходит ни в почках, ни в головном мозге.
Снижение экспрессии генов сиалилтрансфераз и симпортеров является изменениями, характерными для мышц, и, по-видимому, не происходит ни в почках, ни в головном мозге.
Почти все обнаруженные нами возрастные регуляции специфичны для человека и, по-видимому, не встречаются у старых мышей, мух или червей. Таким образом, большая часть возрастной регуляции у людей является видоспецифичной (частной), а не универсальной для всех животных (общественной). Этот результат подчеркивает важность изучения старения у людей, а не у модельных организмов с короткой продолжительностью жизни, чтобы понять, как люди стареют.
Тем не менее, мы обнаружили один путь, который регулируется возрастом у людей, мышей и мух. Путь гена транспортной цепи электронов снижает экспрессию с возрастом у всех трех видов. Предыдущие исследования обнаружили незначительное сходство или отсутствие сходства в регуляции возраста между людьми и мышами [5] или приматами [13]. Эти исследования могли упустить из виду общедоступные модели возрастной регуляции у разных видов, потому что они искали сходства в возрастной регуляции на уровне отдельных генов, а не на уровне целых генетических путей (слишком низкая чувствительность) или потому, что в экспериментах со старением участвовало лишь несколько особей (слишком низкая чувствительность).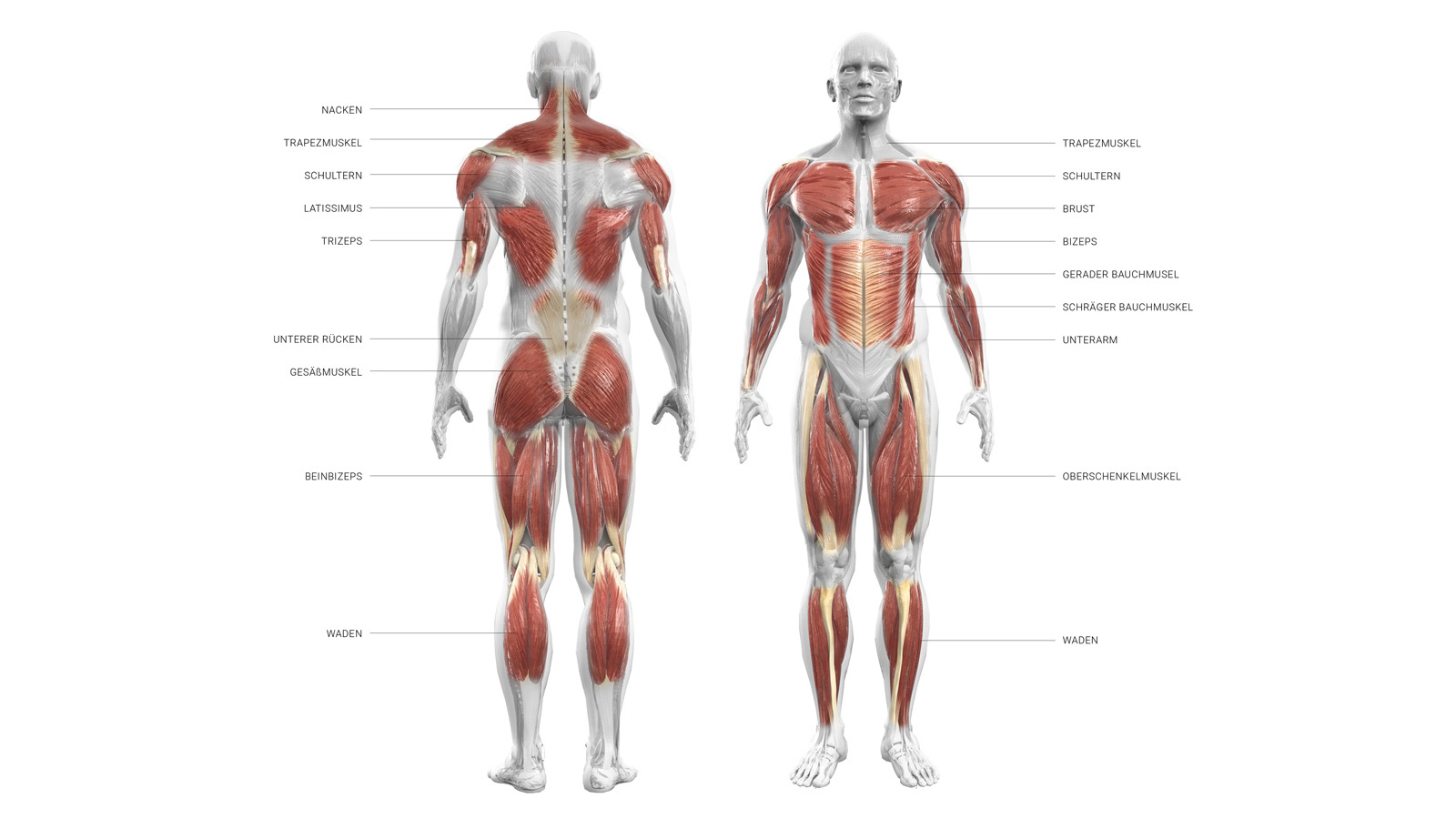 много экспериментального шума).Другое предыдущее исследование сравнило старение у мух и червей и сообщило, что в молодом возрасте наблюдается общее снижение экспрессии генов, кодирующих митохондриальные белки [19].
много экспериментального шума).Другое предыдущее исследование сравнило старение у мух и червей и сообщило, что в молодом возрасте наблюдается общее снижение экспрессии генов, кодирующих митохондриальные белки [19].
У млекопитающих еще не сообщалось о прямых генетических тестах функциональной значимости сниженной экспрессии пути цепи переноса электронов на продолжительность жизни. Однако у C. elegans снижение активности восьми генов, участвующих в цепи переноса электронов, с помощью РНКи значительно увеличило продолжительность жизни [35,36].Ген, кодирующий субъединицу НАДН-дегидрогеназы (NDUFA10) , является одним из генов, демонстрирующих наибольшее снижение экспрессии с возрастом у людей, и было обнаружено, что его ортолог у червей K04G7.4, вызывает одно из самых больших увеличение продолжительности жизни с использованием РНКи у C. elegans [36]. Действительно, в этих глобальных скринингах РНКи путь цепи переноса электронов выделяется как путь, демонстрирующий наибольший и наиболее последовательный эффект на увеличение продолжительности жизни червей [35].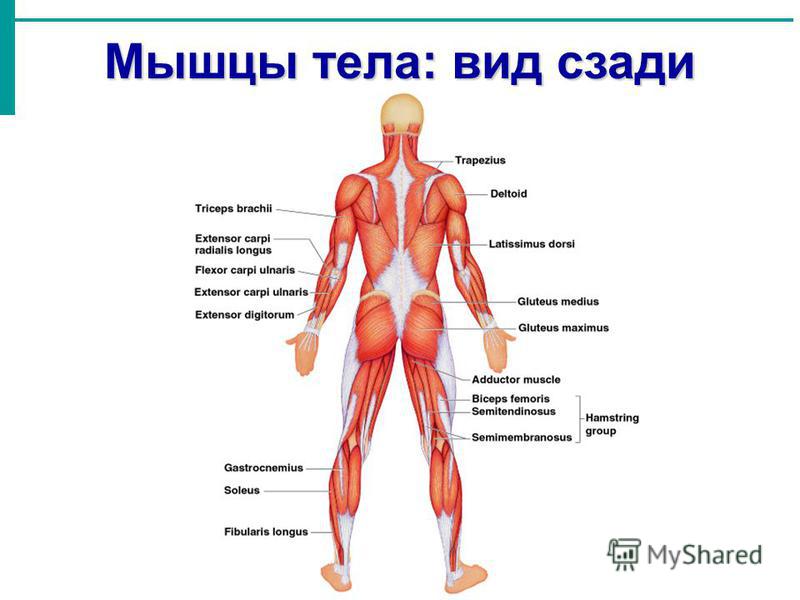 Генетические результаты червей предполагают, что снижение экспрессии пути цепи переноса электронов в пожилом возрасте у людей может быть полезным, способствуя увеличению продолжительности жизни.
Генетические результаты червей предполагают, что снижение экспрессии пути цепи переноса электронов в пожилом возрасте у людей может быть полезным, способствуя увеличению продолжительности жизни.
Какие типы восходящих событий могут вызывать снижение экспрессии электрон-транспортной цепи с возрастом? Другие митохондриальные пути, такие как митохондриальные рибосомы, не проявляют возрастной регуляции, аналогичной пути цепи переноса электронов. Одной из потенциальных причин снижения экспрессии пути цепи переноса электронов является то, что метаболизм может замедляться в пожилом возрасте, что приводит к снижению экспрессии механизма производства энергии в клетке. Другая возможность заключается в том, что окислительное повреждение белков в цепи переноса электронов в митохондриях может привести к снижению экспрессии соответствующих генов в ядре.Цепь переноса электронов создает свободные радикалы в процессе выработки энергии, которые преимущественно повреждают белковые компоненты цепи переноса электронов [33].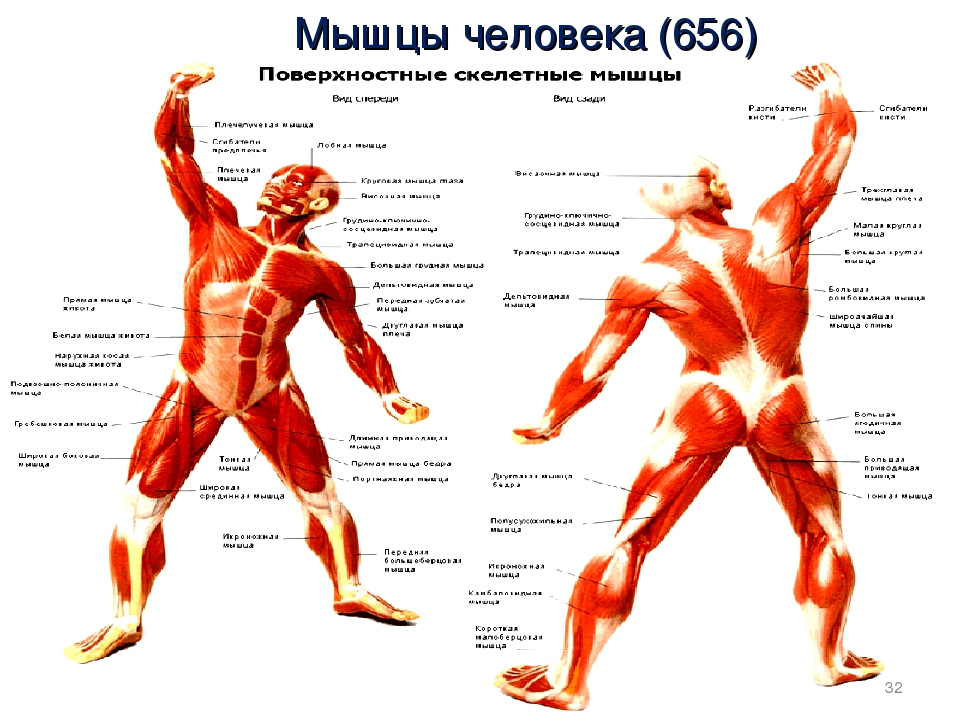
Кажется маловероятным, что обычная возрастная регуляция пути цепи переноса электронов напрямую связана с эволюционной консервацией. События в пожилом возрасте вряд ли окажут существенное влияние на приспособленность популяции, потому что старые животные (такие как 3-летние мыши и 80-летние люди) составляют небольшую часть естественных популяций (за исключением недавней истории человечества). .Возможно, электрон-транспортная цепь регулируется при старении как косвенное следствие регуляции в процессе развития (антагонистическая плейотропия) [37]. С другой стороны, возрастная регуляция этого пути может быть неизбежным следствием старения (например, окислительное повреждение электрон-транспортной цепи в пожилом возрасте может происходить у всех животных) [33].
Интересно, что уровень возрастной регуляции электрон-транспортной цепи у всех видов примерно одинаков, тогда как продолжительность жизни сильно различается.По сравнению с людьми мыши стареют в 20–30 раз, а мухи — в 400 раз быстрее.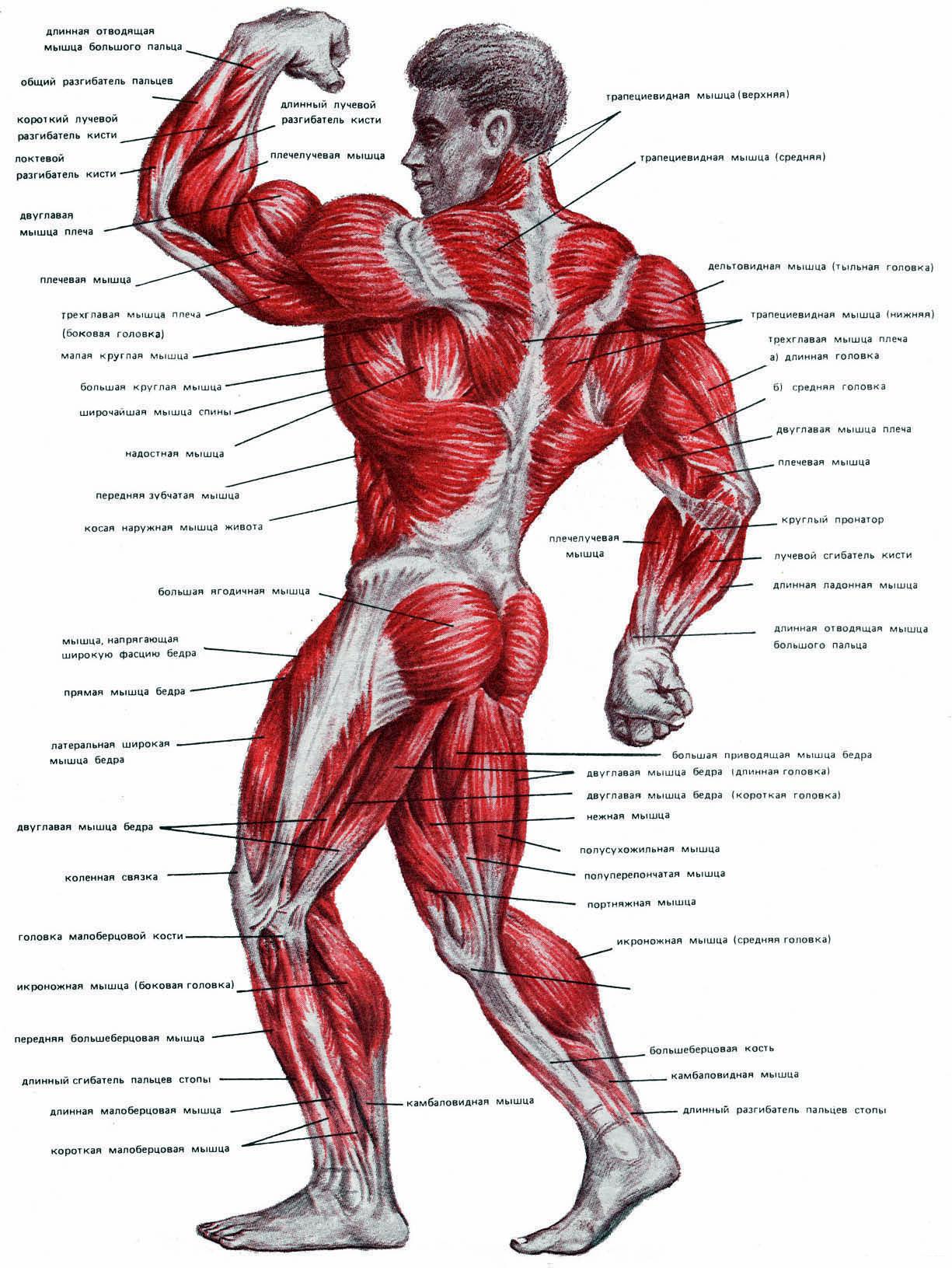 Таким образом, кинетика изменения экспрессии генов электрон-транспортной цепи точно соответствует разнице в продолжительности жизни между видами. Это говорит о том, что снижение экспрессии электрон-транспортной цепи с возрастом может быть особенно информативным в качестве маркера физиологического старения.
Таким образом, кинетика изменения экспрессии генов электрон-транспортной цепи точно соответствует разнице в продолжительности жизни между видами. Это говорит о том, что снижение экспрессии электрон-транспортной цепи с возрастом может быть особенно информативным в качестве маркера физиологического старения.
Материалы и методы
Коллекция образцов.
Образцы мышц были получены из биоптатов пациентов, взятых либо во время операции, либо в амбулаторных процедурах, а медицинские состояния, связанные с каждой биопсией, перечислены в таблице S1.Например, образцы мышц живота брали во время операций по лечению желудочно-кишечных патологий. Не было никакой известной патологии, связанной с самими образцами мышц живота, за исключением того, что они были получены от пациентов с различными желудочно-кишечными расстройствами. В случае пациентов с раком желудочно-кишечного тракта образцы мышц живота брали из областей живота, не пораженных раком. Каждый образец мышцы немедленно замораживали в жидком азоте и впоследствии хранили при температуре -80 °C.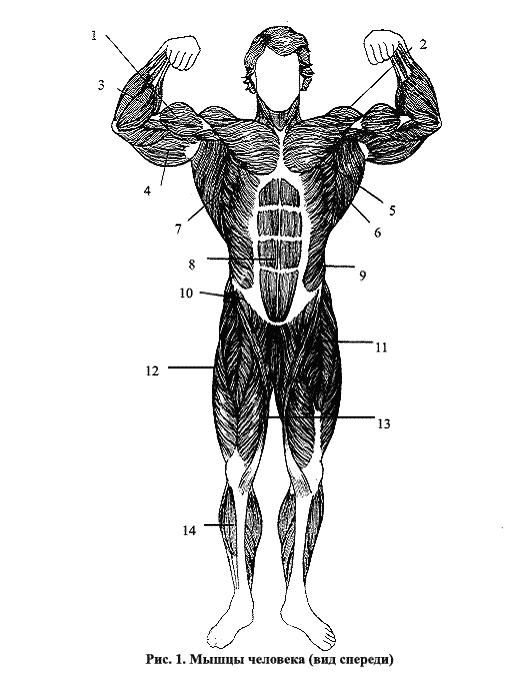 Наконец, мы проверили каждый образец с помощью гистологического окрашивания и исключили любые образцы, которые казались аномальными или больными.
Наконец, мы проверили каждый образец с помощью гистологического окрашивания и исключили любые образцы, которые казались аномальными или больными.
Выделение РНК.
Образцы замороженных мышц взвешивали (50–100 мг), разрезали на мелкие кусочки на сухом льду, а затем помещали в 1 мл реагента TRIzol (Invitrogen, Карлсбад, Калифорния, США). Ткань гомогенизировали с использованием гомогенизатора PowerGen700 (Fisher Scientific, Питтсбург, Пенсильвания, США) и выделяли тотальную РНК в соответствии с протоколом TRIzol Reagent.
Гибридизация генных чипов ДНК.
Стандартный протокол, разработанный Affymetrix (Санта-Клара, Калифорния, США) для их массивов олигонуклеотидов высокой плотности HG-U133 2.0 Plus, был слегка изменен Стэнфордским центром геномных технологий (Стэнфорд, Калифорния, США), и все образцы были обрабатываются на их предприятии (см. протокол S1). Для синтеза кРНК для каждого образца использовали восемь микрограммов тотальной РНК, и 15 мкг кРНК гибридизовали с каждым ДНК-чипом. Образцы обрабатывались в случайном порядке по возрасту.
Образцы обрабатывались в случайном порядке по возрасту.
Нормализация и анализ данных микрочипов.
Мы использовали программу DChip [38] для нормализации данных и создания уровней экспрессии для каждого отдельного зонда, установленного с помощью модели, основанной только на идеальном совпадении. Все данные экспрессии будут общедоступны на веб-сайте Gene Expression Omnibus после принятия. Когда разные наборы зондов соответствовали одному и тому же гену, мы вместе усредняли уровни экспрессии. После усреднения мы использовали log 2 -преобразованных значений выражений для всех последующих анализов.
Измерение диаметра мышечных волокон.
Поперечные срезы криосрезов мышц были сфотографированы с увеличением 200х, а изображения либо измерены в цифровом виде (образцы диагностической биопсии мышц, препараты АТФазы), либо распечатаны (образцы мышц живота, комбинированные препараты СДГ-цитохром c оксидазы) и измерены вручную . Были рассмотрены все диагностические биопсии мышц, и 32 из 81 образца мышц были достаточно интактными для измерения, остальные были неадекватно ориентированы для поперечных сечений или слишком малы для получения значимых данных.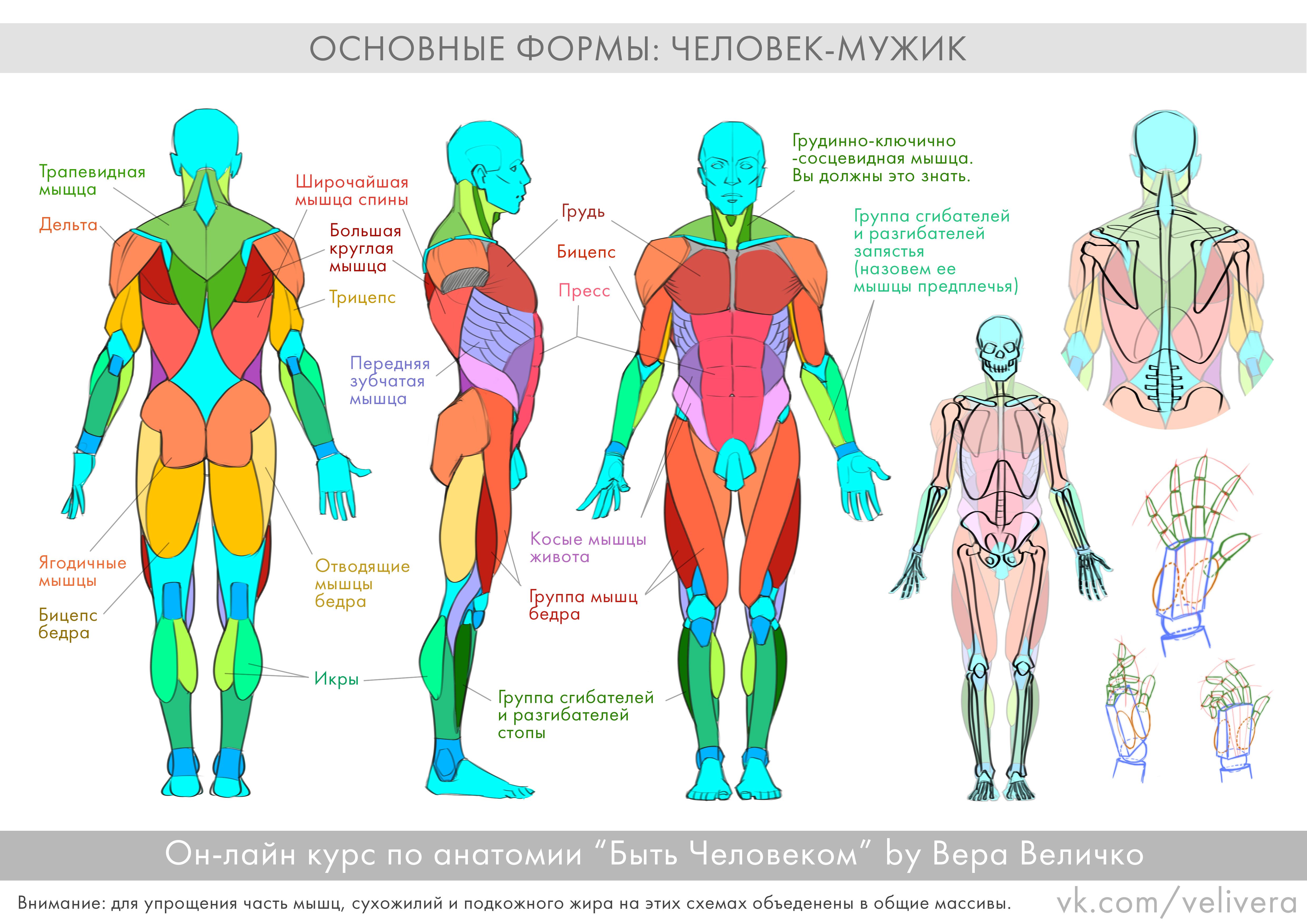 Цифровой анализ заключался в измерении наименьшей ширины приблизительно через центр клетки. После калибровки с известной длиной диаметры измеряли и переводили в микроны с помощью программного обеспечения SigmaScan Pro 5.0 (SPSS Software, Чикаго, Иллинойс, США). Диаметры были сведены в таблицу по типам клеток I и II типа. Количество клеток варьировалось от примерно 30 клеток каждого типа до более 100 в зависимости от размера выборки. Анализ печати проводился по аналогичной методике. Необработанные измерения в миллиметрах использовались для расчета отношения диаметров типа II к диаметру типа I без преобразования в микроны.
Цифровой анализ заключался в измерении наименьшей ширины приблизительно через центр клетки. После калибровки с известной длиной диаметры измеряли и переводили в микроны с помощью программного обеспечения SigmaScan Pro 5.0 (SPSS Software, Чикаго, Иллинойс, США). Диаметры были сведены в таблицу по типам клеток I и II типа. Количество клеток варьировалось от примерно 30 клеток каждого типа до более 100 в зависимости от размера выборки. Анализ печати проводился по аналогичной методике. Необработанные измерения в миллиметрах использовались для расчета отношения диаметров типа II к диаметру типа I без преобразования в микроны.
Множественный регрессионный анализ.
Чтобы определить изменение экспрессии с возрастом, мы использовали модель множественной регрессии, в которой изменение экспрессии с возрастом учитывает возможность того, что уровни экспрессии могут различаться у мужчин и женщин или в мышцах живота по сравнению с периферическими мышцами. В частности, мы использовали следующую модель множественной регрессии:
где Y ij — уровень экспрессии j -го набора зондов для -го образца -го, Пол и соответствует полу образца и (0 для мужчин или 1 для женщин), Анатомия и является анатомическим местом, из которого был взят образец мышц (0 для брюшной полости или 1 для периферических мышц), ɛ ij — ошибка, β 1j — изменение экспрессии с возрастом, β 2j — изменение экспрессии в зависимости от пола, 6 3j 4 β
50 представляет собой изменение выражения в зависимости от анатомического происхождения образца, а β 0j представляет собой точку пересечения регрессии. Для каждого гена j, мы использовали метод наименьших квадратов, чтобы определить все его коэффициенты, при этом нас в первую очередь интересовал коэффициент, относящийся к возрасту (β 1j ). Нас интересовали гены, которые показывают либо положительное, либо отрицательное значение для β 1j , , что указывает либо на увеличение, либо на уменьшение экспрессии в пожилом возрасте, соответственно.
Для каждого гена j, мы использовали метод наименьших квадратов, чтобы определить все его коэффициенты, при этом нас в первую очередь интересовал коэффициент, относящийся к возрасту (β 1j ). Нас интересовали гены, которые показывают либо положительное, либо отрицательное значение для β 1j , , что указывает либо на увеличение, либо на уменьшение экспрессии в пожилом возрасте, соответственно.
Для мозга человека, почки мыши и D. melanogaster, мы определили изменение экспрессии с возрастом для каждого гена, используя следующую модель множественной регрессии:
Для почек человека мы использовали модель множественной регрессии:
В уравнении 3 термин ткани представляет собой бинарный термин, получивший 0 баллов для коры и 1 для мозгового вещества.Для данных C. elegans мы использовали простую линейную регрессию с возрастом:
Рецензенты предложили два дополнительных метода, чтобы показать, что гены, регулирующие возраст, могут служить маркерами физиологического возраста. Во-первых, мы показали, что гены, регулируемые физиологией мышц, также могут предсказывать хронологический возраст. Мы обнаружили гены, которые в значительной степени регулировались соотношением диаметров мышечных волокон типа II и типа I с использованием модели множественной регрессии:
Во-первых, мы показали, что гены, регулируемые физиологией мышц, также могут предсказывать хронологический возраст. Мы обнаружили гены, которые в значительной степени регулировались соотношением диаметров мышечных волокон типа II и типа I с использованием модели множественной регрессии:
Здесь TypeRatio — отношение диаметров мышечных волокон типа II к диаметру мышечных волокон типа I.Мы нашли 585 генов со статистически значимым коэффициентом для TypeRatio с использованием порога p < 0,01. Из этих 585 генов 114 показали частичную корреляцию с возрастом (абсолютное значение r > 0,2), что указывает на значительное перекрытие ( p < 0,02; гипергеометрическое распределение) (таблица S7). 92 гена, обнаруженные в анализе, показанном на рисунке 3, и 114 генов, обнаруженных в этом анализе, имеют общий набор из 7 генов, что указывает на статистически значимое перекрытие ( p < 1 × 10 -8 ; гипергеометрическое распределение).
Во-вторых, мы повторили наш возрастной анализ, принимая во внимание влияние соотношения диаметров мышечных волокон типа II/типа I на возрастную регуляцию. Для этого мы использовали четырехчленную модель множественной регрессии, которая включает условия как для возраста, так и для соотношения типа II/типа I:
Используя уравнение 6, мы обнаружили 543 гена, которые регулируются возрастом ( p < 0,01), и 12 786 генов, регулируемых соотношением типа II/типа I ( p < 0,01; таблица S8). Между этими двумя наборами генов имеется 271 общий ген, что является значительно большим числом, чем можно было бы ожидать случайно (гипергеометрическое p < 1 × 10 -5 ; таблица S8).Мы повторили этот эксперимент, используя порог p <0,001, и обнаружили аналогичное обогащение, подтверждающее наши результаты. Этот анализ показывает, что набор генов, которые регулируются возрастом, обогащен теми, которые отмечают физиологию старения мышц.
Частота ложных открытий, определенная анализом перестановок.
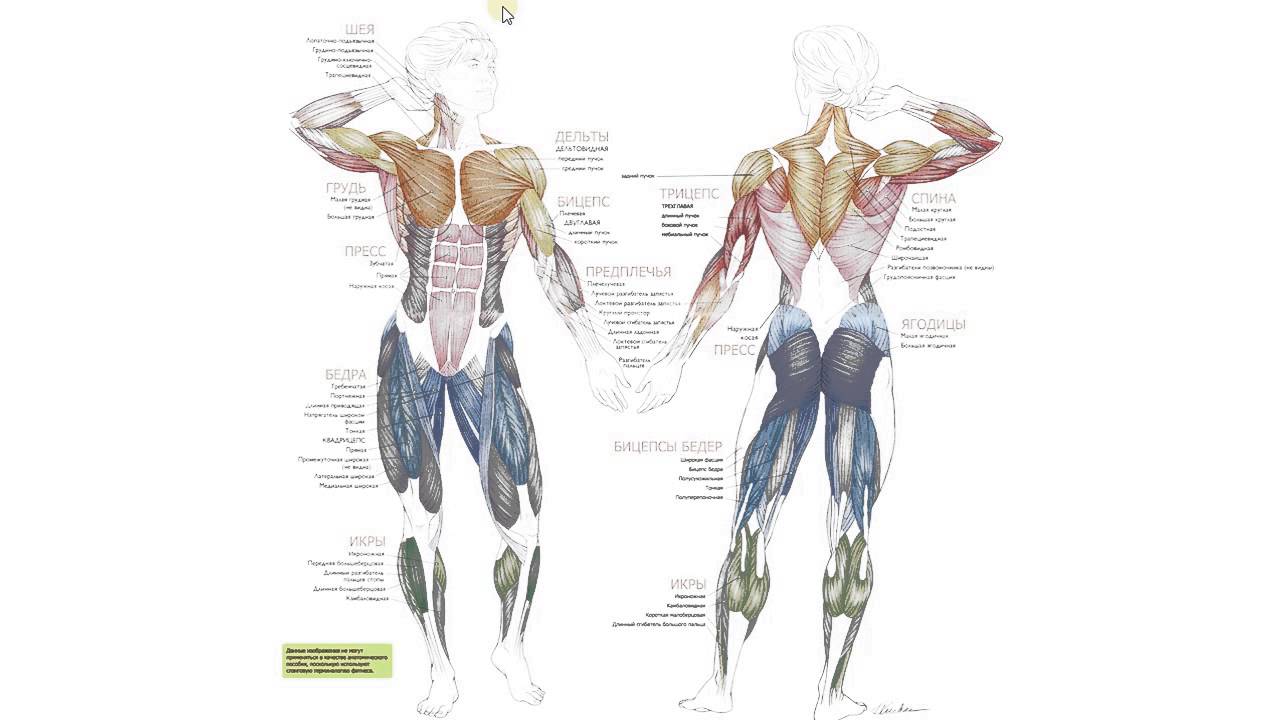
Мы использовали анализ перестановок, чтобы смоделировать количество генов, которые случайно преодолеют пороговое значение ( p < 0,001). Мы рандомизировали возрастные переменные образцов мышц 1000 раз, сохраняя половые и анатомические переменные в образце.Уравнение 1 использовалось для пересчета коэффициентов регрессии и значений p при каждой рандомизации. Теория предсказывает, и наше моделирование подтверждает, что в среднем около 32 генов случайно превышают наш порог ( p < 0,001). Этот результат предполагает, что около 13% ложноположительных результатов в нашем наборе из 250 генов, регулируемых возрастом в мышцах. В 95% пермутированных наборов данных 107 или менее генов были значимы на уровне 0,001.
Кластерный анализ патологических и фармацевтических факторов.
Чтобы проверить, мешают ли патологические или фармацевтические факторы анализу регуляции возраста в мышцах, мы провели неконтролируемую иерархическую кластеризацию по среднему сцеплению 81 образца мышц с использованием программного обеспечения Cluster [39]. 81 образец мышц был сгруппирован на основе 250 генов, которые, как было установлено ранее, регулируются возрастом в мышцах человека.
81 образец мышц был сгруппирован на основе 250 генов, которые, как было установлено ранее, регулируются возрастом в мышцах человека.
Модифицированный анализ обогащения набора генов.
GSEA [40] использует непараметрический тест, чтобы решить, когда n генов в группе G имеют возрастные коэффициенты, которые значительно отличаются от N-n генов, не входящих в G. Модель состоит в том, что n возрастных коэффициентов в G выбираются из распределения G, а Nn коэффициентов, не входящих в G , выбираются из распределения F. Затем мы проверяем нулевую гипотезу о том, что F = G. Тест Смирнова основан на подсчете количества генов из G в первых K генов объединенного списка возрастных коэффициентов и сравнении его с числом, ожидаемым при F = G. Если K изменяется от 1 до N, тест является чувствителен к любой альтернативе F ≠ G.GSEA использует взвешенный критерий Колмогорова-Смирнова, полученный с помощью взвешенного подсчета генов (с большим весом крайних).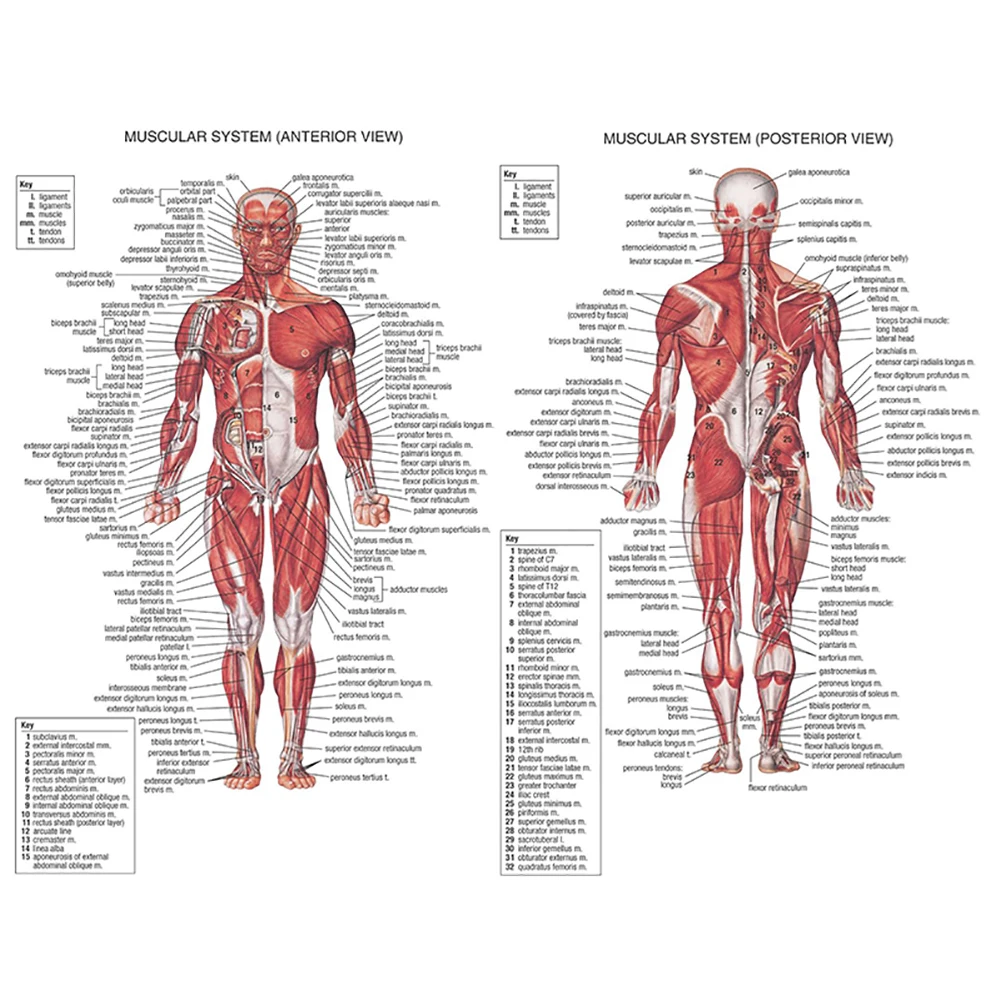 В нашем анализе мы заменили взвешенный критерий Колмогорова-Смирнова взвешенной суммой, критерием нормальных показателей Ван дер Вардена.
В нашем анализе мы заменили взвешенный критерий Колмогорова-Смирнова взвешенной суммой, критерием нормальных показателей Ван дер Вардена.
Критерий Ван дер Вардена более точно соответствует нашей интерпретации того, что означает возрастная связь группы G генов, чем взвешенный критерий Колмогорова-Смирнова. Когда N велико, то любая небольшая группа, которая содержит единственный ген, наиболее связанный с возрастом, значительно связана с возрастом по взвешенному критерию Колмогорова-Смирнова.Такая группа демонстрирует подлинную статистическую значимость и содержит убедительные доказательства того, что F ≠ G, но не обязательно биологически увеличивает или уменьшает экспрессию как механистическую единицу с возрастом. Например, группа из 30 генов с двумя генами, в наибольшей степени увеличивающими возраст, и двумя генами, в наибольшей степени уменьшающими возраст, может оказаться значимой как группой, увеличивающей возраст, так и группой, уменьшающей возраст, даже если другие 26 генов не особенно связаны с возрастом.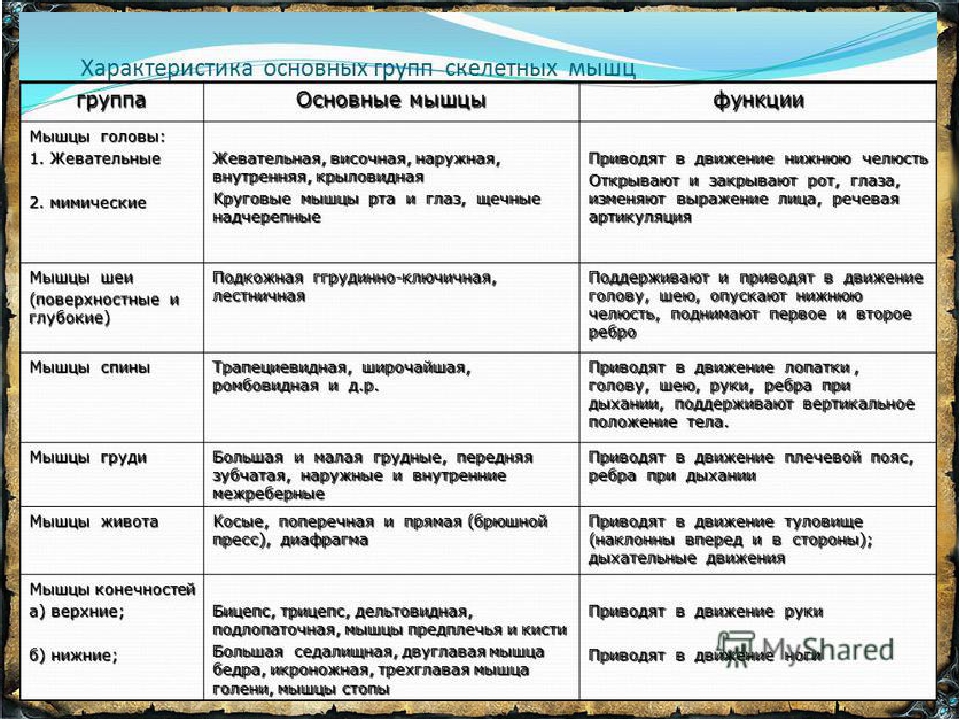 Здесь ясно, что F ≠ G, но, возможно, это просто потому, что G имеет более высокую дисперсию, чем F.
Здесь ясно, что F ≠ G, но, возможно, это просто потому, что G имеет более высокую дисперсию, чем F.
Чтобы вычислить критерий Ван дер Вардена, мы сначала находим ранг r(j) для каждого гена j ∈ G . Этот ранг представляет собой количество исходных N генов с коэффициентом возраста меньше, чем у гена j. Необработанная оценка Ван дер Вардена составляет
., где Φ — стандартная нормальная кумулятивная функция распределения. Когда коэффициенты возраста N независимы с общим непрерывным распределением F = G, тогда распределение Y очень близко к нормальному распределению со средним значением 0 и дисперсией V (Y) , близкой к N-n.Мы заменили показатель обогащения GSEA статистикой Ван дер Вардена, Z = Y/ , что очень близко к N (0,1) при нулевой гипотезе. Когда распределение G смещено влево или вправо относительно F, то значение Z имеет тенденцию увеличиваться сверх того, что мы ожидаем от распределения N (0,1) .
Bootstrap-тест значимости GSEA.

Лучше использовать методы повторной выборки вместо нулевого распределения N (0,1) для оценки значимости показателя обогащения Z.Причина в том, что обычно существуют корреляции между уровнями экспрессии генов в G. . Когда уровни экспрессии двух генов в G коррелируют, возрастные коэффициенты для этих генов также коррелируют [41]. Из этого следует, что их ранги коррелируют, и это обычно увеличивает дисперсию Y, так что в конечном итоге Z больше не равно N (0,1). Значение Z может стать большим либо потому, что гены связаны с возрастом, либо потому, что они коррелируют друг с другом.И то, и другое может быть биологически реальным, но второе не представляет интереса, за исключением, возможно, подтверждения того, что группа G хорошо сконструирована.
Оригинальный GSEA [40] случайным образом переставляет метки двух тестируемых групп, сохраняя при этом данные об экспрессии генов. Это сохраняет корреляции внутри групп, так что любые значимые результаты относятся к нулевому моделированию, которое включает корреляции между генами. Во многих случайных перестановках можно получить гистограмму оценок обогащения для возраста, которая сосредоточена вокруг нуля.Если значение выборки находится далеко за пределами гистограммы, то этот показатель обогащения является статистически значимым.
Во многих случайных перестановках можно получить гистограмму оценок обогащения для возраста, которая сосредоточена вокруг нуля.Если значение выборки находится далеко за пределами гистограммы, то этот показатель обогащения является статистически значимым.
Вместо этого мы приняли подход начальной загрузки. Мы провели повторную выборку данных и повторно вычислили показатели обогащения, получив гистограмму, приблизительно центрированную по наблюдаемому показателю обогащения. Если нулевое значение (ноль) находится далеко за пределами гистограммы с повторной выборкой, то показатель обогащения является статистически значимым. Подход начальной загрузки также сохраняет корреляции между генами, а также корреляции между генами и ковариатами.
Основным мотивом для начальной загрузки является наличие ковариат в наших задачах. Рассмотрим, например, данные с возрастом, полом и переменными выражения. Если мы переставим возрасты по отношению к данным выражения и повторим регрессию, мы должны решить, следует ли привязать переменную пола к возрасту или к выражениям в случайной перестановке. Присоединение пола к возрастным переменным оставит нас с смоделированными наборами данных, в которых женщины экспрессируют гены Y-хромосомы в той же степени, что и мужчины.Из-за таких артефактов это не подходящее нулевое распределение. Присоединение ковариаты к переменным выражения также проблематично. Предположим, что одна из ковариат каким-то образом коррелирует с возрастом. Эффект будет заключаться в увеличении дисперсии первоначально отобранного возрастного коэффициента. В выборках перестановок, где ковариата присоединена к данным выражения, она подвергается повторной выборке независимо от возраста. Такая независимость снижает дисперсию коэффициента возраста в данных перестановки. Следствием этого является то, что гистограмма возрастных коэффициентов, основанная на перестановках, становится слишком узкой, что приводит к ложным открытиям.
Присоединение пола к возрастным переменным оставит нас с смоделированными наборами данных, в которых женщины экспрессируют гены Y-хромосомы в той же степени, что и мужчины.Из-за таких артефактов это не подходящее нулевое распределение. Присоединение ковариаты к переменным выражения также проблематично. Предположим, что одна из ковариат каким-то образом коррелирует с возрастом. Эффект будет заключаться в увеличении дисперсии первоначально отобранного возрастного коэффициента. В выборках перестановок, где ковариата присоединена к данным выражения, она подвергается повторной выборке независимо от возраста. Такая независимость снижает дисперсию коэффициента возраста в данных перестановки. Следствием этого является то, что гистограмма возрастных коэффициентов, основанная на перестановках, становится слишком узкой, что приводит к ложным открытиям.
В рамках бутстрап-подхода мы создали 1000 выборочных наборов данных. В каждом выборочном наборе данных мы имитировали процесс выборки, который привел к получению данных, путем повторной выборки 81 субъекта из 81 субъекта. Повторная выборка сохраняет возраст, выражение и все ковариаты любого данного субъекта вместе. Выборка начальной загрузки имитирует случайный процесс, который генерирует данные.
Повторная выборка сохраняет возраст, выражение и все ковариаты любого данного субъекта вместе. Выборка начальной загрузки имитирует случайный процесс, который генерирует данные.
Мы отмечаем, что как бутстрепная, так и пермутационная выборка оценок Ван дер Вардена привели к Z-оценкам, которые имели почти нормальное распределение, но не обязательно N(0,1) (неопубликованные данные).При перестановочной выборке средние значения гистограммы показателей обогащения, как правило, были близки к нулю, но в некоторых группах дисперсия превышала 1,0. При бутстрап-выборке дисперсии часто отличались от 1, а средние значения обычно находились между нулем и исходной оценкой обогащения.
Венна и корреляционный анализ мышц, почек и головного мозга человека.
Самый прямой способ сравнить старение в мышцах, почках и мозге — использовать анализ Венна: мы находим, какие гены достигают строгого уровня значимости для каждой ткани, и судим, является ли перекрытие статистически значимым, в соответствии с гипергеометрическим распределением. Мы провели попарное сравнение каждой ткани, чтобы найти гены, регулирующие старение, в обоих наборах. Существует шесть генов, регулирующих старение, как в мышцах, так и в почках ( p < 0,09, гипергеометрическое распределение), пять генов, регулирующих старение, как в мышцах, так и в головном мозге ( p < 0,07) и 13 генов, регулирующих старение. общего между почкой и мозгом ( p < 0,29). Во всех трех наборах данных не было генов, строго регулируемых возрастом. Подход анализа Венна легко интерпретируется, но ему не хватает силы, поскольку он заменяет фактические измеренные корреляции менее информативным представлением о том, превышают ли они пороговое значение.
Мы провели попарное сравнение каждой ткани, чтобы найти гены, регулирующие старение, в обоих наборах. Существует шесть генов, регулирующих старение, как в мышцах, так и в почках ( p < 0,09, гипергеометрическое распределение), пять генов, регулирующих старение, как в мышцах, так и в головном мозге ( p < 0,07) и 13 генов, регулирующих старение. общего между почкой и мозгом ( p < 0,29). Во всех трех наборах данных не было генов, строго регулируемых возрастом. Подход анализа Венна легко интерпретируется, но ему не хватает силы, поскольку он заменяет фактические измеренные корреляции менее информативным представлением о том, превышают ли они пороговое значение.
Более чувствительное сравнение может быть основано на корреляции возрастных коэффициентов генов в двух тканях. Мы выбрали все гены, которые регулируются возрастом в любой из двух тканей, нанесли на график коэффициент возраста каждого гена в одной ткани по сравнению с коэффициентом этого гена в другой ткани и вычислили корреляцию Пирсона (r) полученных точек (таблица S11). ). Мы обнаружили самое сильное совпадение старения между почками и мозгом ( r = 0,219) и меньшее, но положительное совпадение старения между мышцами и почками ( r = 0.103) или мышцы и мозг ( r = 0,078).
). Мы обнаружили самое сильное совпадение старения между почками и мозгом ( r = 0,219) и меньшее, но положительное совпадение старения между мышцами и почками ( r = 0.103) или мышцы и мозг ( r = 0,078).
Поскольку гены коррелированы, мы не можем использовать формулы из учебников, чтобы судить о статистической значимости этих показателей Пирсона. Чтобы получить значение p для корреляции Пирсона между почками и мышцами, мы использовали 1000 наборов случайных генов. Количество генов в каждом наборе было таким же, как и число, которое мы использовали для вычисления корреляции в таблице S11. Для каждой случайной группы генов мы рассчитали корреляцию Пирсона между возрастными коэффициентами в почках и мышцах.Из 1000 образцов было шесть, в которых случайная группа генов привела к большей корреляции Пирсона, чем та, которую мы видели в реальных данных. Это соответствует значению p , равному 0,006, для почки-мышцы. Аналогичным образом мы нашли значение p , равное 0,001 для почки-мозг, но только 0,058 для мышцы-мозг. За возможным исключением пары почки-головной мозг, связанные с возрастом гены имеют более стабильные возрастные коэффициенты по тканям, чем случайно выбранные гены.
Мы также провели бутстреп-тест сравнения тканей.В этом тесте мы передискретизировали данные микрочипа с заменой 1000 раз. Каждый раз мы пересчитывали корреляции между возрастными коэффициентами для генов в почках и мышцах. Из 1000 испытаний мы увидели 39, в которых корреляция выборки была меньше или равна нулю. После преобразования в двусторонний критерий это соответствует значению p , равному 0,078, для почки-мышцы. Для экономии вычислений мы использовали один и тот же набор генов в каждом бутстрап-образце вместо того, чтобы заставлять набор генов, связанных с возрастом, варьироваться в зависимости от выборки отдельно.Значение p для мышцы-мозга было 0,07, а для почки-мозга — 0,001. Основываясь на этих индивидуальных анализах на уровне генов, возрастные гены в почках и мозге, как правило, очень похожи. Сравнение мышц-почек и мышц-мозга было слабее.
Сравнение мышц-почек и мышц-мозга было слабее.
Тесты на корреляцию тканей в обычно регулируемых возрастом наборах генов.
Чтобы проверить корреляцию рангов генов между тканями в тех наборах генов, которые обычно регулируются возрастом у человека, мы использовали метод двусторонней корреляции Спирмена, чтобы сначала рассчитать коэффициент корреляции для каждой парной комбинации тканей (т.например, мышца-почка, почка-мозг, мышца-мозг) для этого набора генов, регулируемых возрастом (например, гены внеклеточного матрикса). Чтобы проверить значимость рассчитанных корреляций, мы использовали метод Монте-Карло на основе перестановок, рандомизируя ранги для каждого гена и ткани в наборе генов и пересчитывая корреляции Спирмена 1000 раз. Мы обнаружили, что большинство корреляций между тканями не были значительными (таблица S12).
Дополнительная информация
Рисунок S1.Возрастное распределение анатомических, медицинских и фармацевтических факторов
Каждая строка обозначает медицинский или фармацевтический фактор. Возраст пациентов показан на оси x . В легенде указаны пол, место биопсии и 12 медицинских факторов. Только гипотиреоз демонстрирует явную связь с возрастом.
Возраст пациентов показан на оси x . В легенде указаны пол, место биопсии и 12 медицинских факторов. Только гипотиреоз демонстрирует явную связь с возрастом.
https://doi.org/10.1371/journal.pgen.0020115.sg001
(253 КБ TIF)
Рисунок S2. Медицинские и фармацевтические факторы не влияют на возрастное регулирование
(A) Ишемическая болезнь сердца была включена в уравнение 1 в качестве дополнительного термина, и модель была пересчитана для 250 генов, экспрессия которых значительно меняется с возрастом.Был построен наклон выражения с возрастом (коэффициент возраста) для моделей с (ось y ) и без (ось х ) сроком ишемической болезни сердца. Если бы ишемическая болезнь сердца влияла на экспрессию, можно было бы ожидать большое отклонение возрастного коэффициента. Никаких существенных отклонений не наблюдалось ни для одного из 250 генов, регулирующих возраст, что указывает на то, что ишемическая болезнь сердца не оказывает неблагоприятного влияния на наше исследование регуляции возраста.
(B–L) Аналогично (A) для 11 других медицинских факторов.(Б) Ишемическая болезнь сердца. (С) Колоректальный рак. (D) Терминальная стадия почечной недостаточности. (Е) Гиперлипидемия. (Е) Гипертония. (G) Гипотиреоз. (H) Рак поджелудочной железы. (I) Рак простаты. (J) Лучевая терапия. (К) Статины. (L) Ворсинчатая аденома.
https://doi.org/10.1371/journal.pgen.0020115.sg002
(319 КБ TIF)
Рисунок S3. Кластерный анализ медицинских и фармацевтических факторов
Образцы сгруппированы на основе 250 генов, регулируемых возрастом в мышцах, как показано на верхней дендрограмме.Столбцы представляют собой отдельные образцы мышц, отмеченные по возрасту пациента. Верхние семь рядов соответствуют экспрессии первых семи генов, регулируемых возрастом. На диаграмме показаны анатомические, медицинские и фармацевтические факторы для каждого пациента. Каждая строка соответствует одному медицинскому или фармацевтическому фактору.
https://doi.org/10.1371/journal. pgen.0020115.sg003
pgen.0020115.sg003
(1,2 МБ TIF)
Благодарности
Мы благодарны Drs. Джеффри Нортону, Джозефу Прести, Эндрю Шелтону и Марку Велтону за их помощь в получении образцов мышц человека, а также Мэрилин Масек за ее опыт в области морфометрии мышц.
Вклад авторов
RWD, ABO и SKK задумали и разработали эксперименты. Эксперименты проводили JMZ, RS, HV, EC, KMM, KGB и ABO. JMZ, RS, HV, EC, KMM, KGB и ABO проанализировали данные. HV, RR и RWD предоставили реагенты/материалы/инструменты для анализа. JMZ, RR, ABO и SKK написали статью.
Каталожные номера
- 1. Батлер Р.Н., Спротт Р., Уорнер Х., Бланд Дж., Фейерс Р. и др. (2004) Биомаркеры старения: от примитивных организмов до человека.J Gerontol A Biol Sci Med Sci 59: B560–B567.
- 2. Миллер Р.А., Крисп С., Галецки А. (1997) Уровни Т-клеток памяти CD4 предсказывают продолжительность жизни у генетически гетерогенных мышей. FASEB J 11: 775–783.
- 3. Миллер Р.А. (2001) Биомаркеры старения: прогнозирование продолжительности жизни с помощью определения подмножества чувствительных к возрасту Т-клеток в генетически гетерогенной популяции мышей среднего возраста. J Gerontol A Biol Sci Med Sci 56: B180–B186.
- 4. Krishnamurthy J, Torrice C, Ramsey MR, Kovalev GI, Al-Regaiey K, et al.(2004) Экспрессия Ink4a/Arf является биомаркером старения. Дж. Клин Инвест 114: 1299–1307.
- 5. Rodwell GE, Sonu R, Zahn JM, Lund J, Wilhelmy J, et al. (2004) Транскрипционный профиль старения в почках человека. PLoS Biol 2: e427.. DOI: https://doi.org/10.1371/journal.pbio.0020427.
- 6. Лу Т., Пан И, Као С.И., Ли С., Кохане И. и др. (2004)Генная регуляция и повреждение ДНК в стареющем человеческом мозге. Природа 429: 883–891.
- 7.
Welle S, Brooks AI, Delehanty JM, Needler N, Thornton CA (2003) Профиль экспрессии генов старения в мышцах человека.Physiol Genomics 14: 149–159.
- 8. Уэлле С., Брукс А.И., Делеханти Дж.М., Нидлер Н., Бхатт К. и др. (2004) Профили экспрессии генов скелетных мышц у женщин в возрасте 20–29 и 65–71 лет. Эксперт Геронтол 39: 369–377.
- 9. Ly DH, Lockhart DJ, Lerner RA, Schultz PG (2000) Митотическая неправильная регуляция и старение человека. Наука 287: 2486–2492.
- 10. Наир К.С. (2005)Старение мышц. Ам Дж. Клин Нутр 81: 953–963.
- 11. Ларссон Л., Гримби Г., Карлссон Дж. (1979) Мышечная сила и скорость движения в зависимости от возраста и морфологии мышц.J Appl Physiol 46: 451–456.
- 12. Ларссон Л., Эдстром Л. (1986)Влияние возраста на ферментативно-гистохимические спектры волокон и сократительные свойства быстро- и медленно сокращающихся скелетных мышц у крыс. J Neurol Sci 76: 69–89.
- 13.
Fraser HB, Khaitovich P, Plotkin JB, Paabo S, Eisen MB (2005)Старение и экспрессия генов в мозге приматов. PLoS Biol 3: e274. . DOI: https://doi.org/10.1371/journal.pgen.0030274.
- 14. Lee CK, Klopp RG, Weindruch R, Prolla TA (1999) Профиль экспрессии генов старения и его замедление за счет ограничения калорийности.Наука 285: 1390–1393.
- 15. Lee CK, Weindruch R, Prolla TA (2000) Профиль экспрессии генов стареющего мозга у мышей. Нат Жене 25: 294–297.
- 16. Pletcher SD, Macdonald SJ, Marguerie R, Certa U, Stearns SC, et al. (2002)Полногеномные профили транскриптов у стареющей и ограниченной калорийности Drosophila melanogaster. Курр Биол 12: 712–723.
- 17. Лэндис Г.Н., Абдуева Д., Скворцов Д., Ян Дж., Рабин Б.Е., и соавт. (2004) Сходные паттерны экспрессии генов характеризуют старение и окислительный стресс у Drosophila melanogaster.Proc Natl Acad Sci USA 101: 7663–7668.
- 18.
Лунд Дж., Тедеско П., Дюк К., Ван Дж., Ким С.К. и др. (2002) Транскрипционный профиль старения у C. elegans. Курр Биол 12: 1566–1573.
- 19. McCarroll SA, Murphy CT, Zou S, Pletcher SD, Chin CS, et al. (2004) Сравнение паттернов геномной экспрессии у разных видов выявляет общий профиль транскрипции при старении. Нат Жене 36: 197–204.
- 20. Cowie CC, Eberhardt MS (1995) Социально-демографические характеристики людей с диабетом.В: Harris MI, Cowie CC, Stern MP, Boyko EJ, Reiber GE и др., редакторы. Диабет в Америке. 2-е изд. Bethesda: Национальный информационный центр по диабету. стр. 85–116. стр.
- 21. Д’Мелло Н.П., Чайлдресс А.М., Франклин Д.С., Кале С.П., Пинсвасди С. и др. (1994)Клонирование и характеристика LAG1, гена обеспечения долголетия у дрожжей. J Biol Chem 269: 15451–15459.
- 22. Гульбинс Е. (2003) Регуляция передачи сигналов рецептора смерти и апоптоза с помощью церамида. Фармакол рез. 47: 393–399.
- 23.
Мута В.К., Линдгрен С.М., Эрикссон К.Ф., Субраманиан А., Сихаг С. и др. (2003) PGC-1alpha-чувствительные гены, участвующие в окислительном фосфорилировании, скоординированно подавлены при диабете человека. Нат Жене 34: 267–273.
- 24. Эшбернер М., Болл К.А., Блейк Дж.А., Ботштейн Д., Батлер Х. и др. (2000) Онтология генов: инструмент для объединения биологии. Консорциум генных онтологий. Нат Жене 25: 25–29.
- 25. Перри С.В. (2004)Активация сократительного механизма кальцием.В: Engel AG, Franzini-Armstrong C, редакторы. Миология. 3-е изд. Нью-Йорк: Макгроу-Хилл. стр. 281–306. стр.
- 26. Магуайр ТМ, Джиллиан А.М., О’Махони Д., Кофлан К.М., Деннихан А. и др. (1994) Снижение уровня сиалилтрансферазы в сыворотке крови при болезни Альцгеймера. Нейробиол Старение 15: 99–102.
- 27. Maguire TM, Breen KC (1995) Снижение активности нервной сиалилтрансферазы при болезни Альцгеймера. Деменция 6: 185–190.
- 28. Аллалуф Д., Комлос Л., Нотманн Дж., Хальбрехт И., Левински Х. (1988)Содержание сиаловой кислоты и активность сиалилтрансферазы в лимфоцитах человека с возрастом.Старение меха Dev 44: 45–50.
- 29. Colten HR (1992)Тканеспецифическая регуляция воспаления. J Appl Physiol 72: 1–7.
- 30. Makrides SC (1983)Синтез и расщепление белка при старении и старении. Biol Rev Camb Philos Soc 58: 343–422.
- 31. Дженч Т.Дж., Марицен Т., Здебик А.А. (2005)Заболевания хлоридных каналов, возникающие в результате нарушения трансэпителиального транспорта или везикулярной функции. Дж. Клин Инвест 115: 2039–2046.
- 32.Mihic SJ, Harris RA (1997)ГАМК и рецептор ГАМК A . Нейромедиатор Откр. 21: 127–131.
- 33. Харман Д. (1956) Старение: теория, основанная на свободных радикалах и радиационной химии. Дж Геронтол 11: 298–300.
- 34. Cheadle C, Vawter MP, Freed WJ, Becker KG (2003) Анализ данных микрочипов с использованием преобразования Z-оценки. Дж. Мол Диагн. 5: 73–81.
- 35.
Ли С.С., Ли Р.Ю., Фрейзер А.Г., Камат Р.С., Арингер Дж. и др.(2003) Систематический скрининг РНК-идентифицирует критическую роль митохондрий в долголетии C. elegans. Нат Жене 33: 40–48.
- 36. Гамильтон Б., Донг Ю., Шиндо М., Лю В., Оделл И. и др. (2005)Систематический скрининг генов долголетия у C. elegans с помощью РНК-интерференции. Гены Дев 19: 1544–1555.
- 37. Уильямс Г. (1957)Плейотропия, естественный отбор и эволюция старения. Эволюция 11: 398–411.
- 38. Li C, Wong WH (2001) Модельный анализ массивов олигонуклеотидов: вычисление индекса экспрессии и обнаружение выбросов.Proc Natl Acad Sci U S A 98: 31–36.
- 39. Эйзен М.Б., Спеллман П.Т., Браун П.О., Ботштейн Д. (1998) Кластерный анализ и отображение паттернов экспрессии всего генома. Proc Natl Acad Sci U S A 95: 14863–14868.
- 40. Субраманиан А., Тамайо П., Мутха В.К., Мукерджи С., Эберт Б.Л. и др. (2005) Анализ обогащения набора генов: основанный на знаниях подход к интерпретации профилей экспрессии всего генома. Proc Natl Acad Sci USA 102: 15545–15550.
- 41.Оуэн А.Б. (2005) Дисперсия количества ложных открытий. J Roy Soc Stat B 67: 411–426.
 Миллер Р.А. (2001) Биомаркеры старения: прогнозирование продолжительности жизни с помощью определения подмножества чувствительных к возрасту Т-клеток в генетически гетерогенной популяции мышей среднего возраста. J Gerontol A Biol Sci Med Sci 56: B180–B186.
Миллер Р.А. (2001) Биомаркеры старения: прогнозирование продолжительности жизни с помощью определения подмножества чувствительных к возрасту Т-клеток в генетически гетерогенной популяции мышей среднего возраста. J Gerontol A Biol Sci Med Sci 56: B180–B186.
 . DOI: https://doi.org/10.1371/journal.pgen.0030274.
. DOI: https://doi.org/10.1371/journal.pgen.0030274.
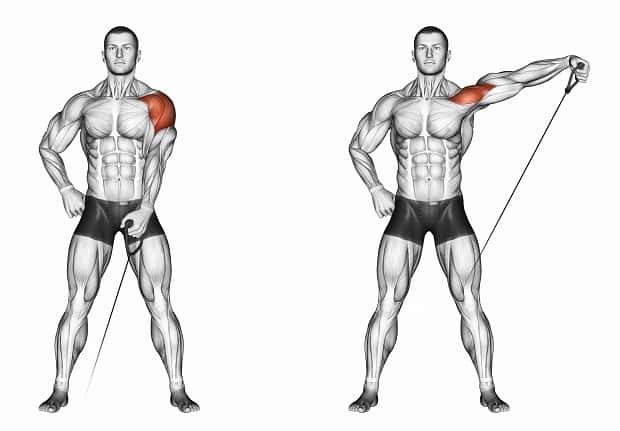 Нат Жене 34: 267–273.
Нат Жене 34: 267–273. Colten HR (1992)Тканеспецифическая регуляция воспаления. J Appl Physiol 72: 1–7.
Colten HR (1992)Тканеспецифическая регуляция воспаления. J Appl Physiol 72: 1–7. elegans. Нат Жене 33: 40–48.
elegans. Нат Жене 33: 40–48. J Roy Soc Stat B 67: 411–426.
J Roy Soc Stat B 67: 411–426.границ | Сравнительный анализ транскрипционных характеристик скелетных мышц, связанных с аэробной нагрузкой или реакцией на тренировку у людей и крыс
Введение
Крупномасштабные клинические исследования показывают сильную связь между низкой переносимостью физических нагрузок и более короткой продолжительностью жизни и более высокой вероятностью развития сложных метаболических заболевания (1, 2).Скелетные мышцы, составляющие до 40% массы тела человека, являются очевидной и важной целью для изучения молекулярных связей между физической нагрузкой и здоровьем человека (3). Адаптация к упражнениям включает временные изменения транскриптома после тренировки и устойчивые изменения в экспрессии генов, вызванные и поддерживаемые тренировками (3). Эти изменения организуются комбинаторной активностью факторов транскрипции, коактиваторов и других сигнальных медиаторов, которые регулируют определенные генные программы, вызывая адаптацию, вызванную физической нагрузкой (3, 4).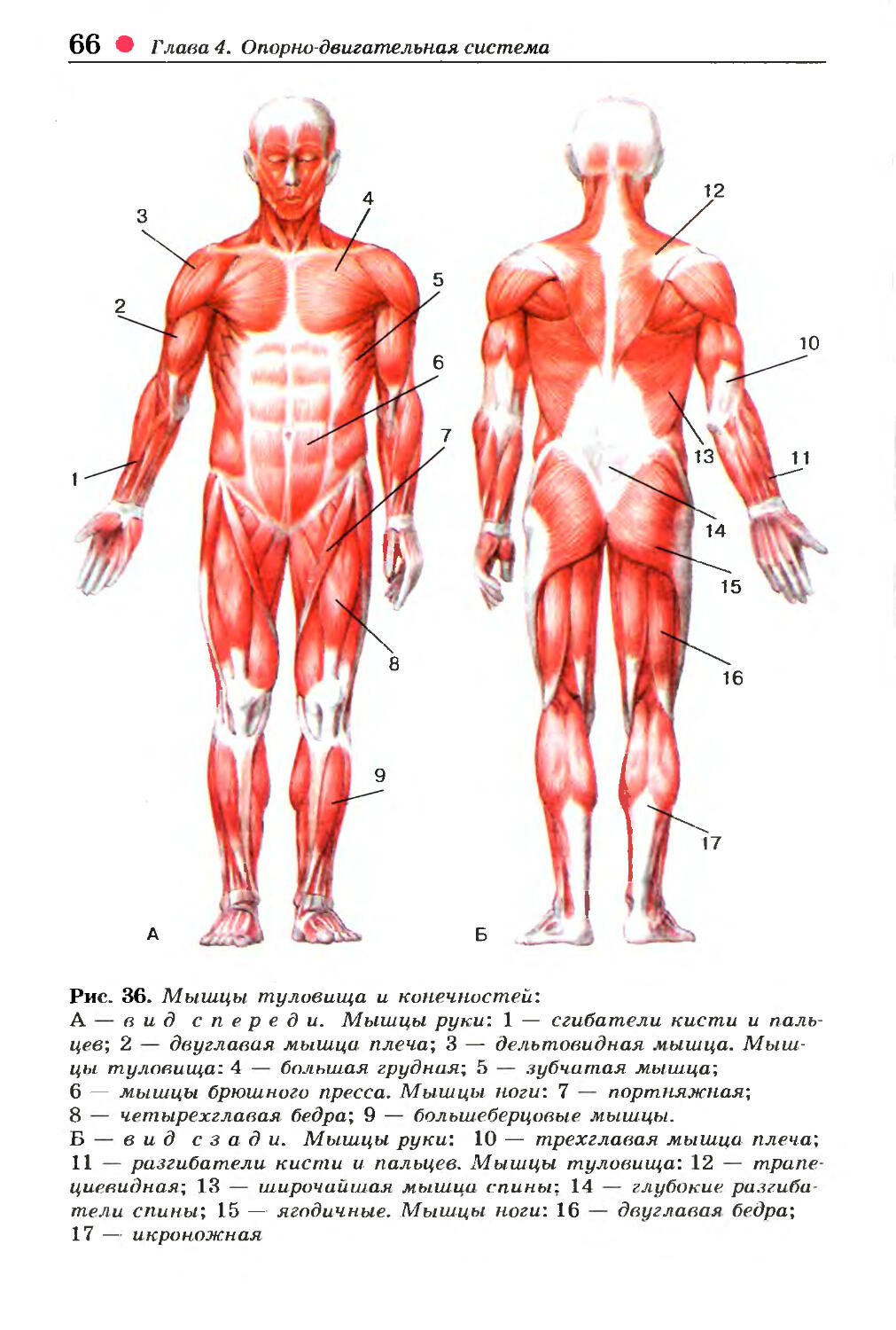
Способность к физической нагрузке — комплексная характеристика, являющаяся результатом суммы генетических факторов и факторов окружающей среды. Первые можно далее разделить на внутренние (уже присутствующие в нетренированном состоянии) и внешние (приобретенные) компоненты, которые способны объяснить до 50% общей фенотипической изменчивости, наблюдаемой у людей (5–7). Модели животных, разработанные для каждого компонента, открыли новые возможности для изучения их связи с физической подготовкой и сложными заболеваниями. Для моделирования внутреннего компонента аэробной физической нагрузки линии бегунов с высокой и низкой производительностью (HCR и LCR соответственно) были получены из гетерогенной популяции крыс-основателей (N:NIH) с отбором заводчиков на основе нетренированной беговой дорожки (8). .Как было показано ранее на людях, межпоколенческие генетические исследования показали, что эти крысы демонстрируют аэробные способности как наследуемый признак (9) и что крысы LCR более восприимчивы к различным заболеваниям, особенно к нарушениям обмена веществ (10). Интересно, что исследования с более ранними поколениями крыс HCR показали, что основной движущей силой фенотипа высокой производительности является улучшение оксигенации скелетных мышц (11–13).
Интересно, что исследования с более ранними поколениями крыс HCR показали, что основной движущей силой фенотипа высокой производительности является улучшение оксигенации скелетных мышц (11–13).
Как у людей, так и у грызунов, некоторые особи демонстрируют четкую реакцию на тренировку на выносливость, в то время как другие демонстрируют меньшее улучшение или отсутствие улучшения определенных физиологических характеристик, таких как максимальное потребление кислорода (10, 14).Хотя ранние исследования на людях показали, что это влечет за собой генетический компонент, составляющий 47% дисперсии в улучшении VO 2 max (7, 15), факторы, лежащие в основе реакции на аэробные упражнения, остаются в значительной степени неизвестными. У крыс доля изменений аэробной способности, объясняемая генетическими факторами, составляет около 45% (9). Для моделирования приобретенного компонента способности к физической нагрузке у крыс была разработана стратегия размножения, аналогичная модели HCR, но основанная на высокой и низкой реакции на физическую нагрузку (HRT и LRT соответственно) (16). Из поколения в поколение этот искусственный отбор по тренируемости дал две линии с одинаковыми исходными способностями, которые отличались по своей реакции на одну и ту же программу тренировок на выносливость.
Из поколения в поколение этот искусственный отбор по тренируемости дал две линии с одинаковыми исходными способностями, которые отличались по своей реакции на одну и ту же программу тренировок на выносливость.
На сегодняшний день транскрипционный анализ мышц HCR был выполнен с помощью целевых подходов или с использованием массивов экспрессии генов на основе зондов, а модель реакции на физическую нагрузку не изучалась так широко (17–19). Сравнение профилей внутренней и приобретенной способности скелетных мышц в масштабе всего транскриптома может предложить новые пути для раскрытия связи между способностью к физической нагрузке и здоровьем.В этом исследовании мы провели сравнительный анализ профилей транскриптов скелетных мышц, связанных с конкретными показателями физической активности у крыс и людей. Наши данные идентифицируют молекулярные пути и специфические гены как потенциальные медиаторы физической способности и тренируемости.
Материалы и методы
Животные
Всех животных содержали, селекционно разводили и проводили экспериментальные процедуры в соответствии с Руководством по уходу и использованию лабораторных животных Института исследований лабораторных животных и в соответствии с рекомендациями Университетского комитета по использованию и Уход за животными в Мичиганском университете.
Селективное разведение для модели переносимости физической нагрузки
Модели для низкой и высокой переносимости нагрузки, LCR (бегуны низкой работоспособности) и HCR (бегуны высокой грузоподъемности) были подробно описаны ранее (8). Вкратце, крыс подвергали двусторонней селективной селекции на нетренированную аэробную способность, измеряемую бегом на выносливость. Начиная с генетически гетерогенной популяции-основателя (количество N:NIH), 13 пар крыс с самой низкой и самой высокой способностью к бегу каждого пола были случайным образом сопоставлены для спаривания.Затем для каждого поколения потомство из каждой семьи тестировали на способность к бегу на беговой дорожке в возрасте 11 недель и подвергали отбору таким же образом. В этом исследовании использовались архивные образцы замороженных икроножных мышц, взятые у 3 самцов крыс поколения 33 HCR и LCR. Фенотип переносимости физической нагрузки был подтвержден протоколом упражнений на беговой дорожке, как описано (8).
Искусственный отбор для моделей с приобретенными способностями
Самцов крыс с низкой реакцией (LRT) и с высокой реакцией (HRT) содержали и содержали, как подробно описано ранее (16). Вкратце, 10–11-недельные самцы и самки из генетически гетерогенного поголовья N: NIH были подвергнуты тесту с постепенным бегом на беговой дорожке, определяющему их предтренировочные способности (DIST1). Затем каждая крыса проходила программу тренировок на беговой дорожке (3 дня в неделю в течение 8 недель). Программа постепенно увеличивала скорость с шагом 1 м/мин и продолжительность на 0,5 мин в каждом сеансе, начиная с 10 м/мин в течение 20 мин на 1-й неделе и заканчивая 21 м/мин в течение 31,5 мин на 8-й неделе. В конце тренировочной программы оценивали способность каждой крысы к физической нагрузке после тренировки (DIST2), а реакцию на тренировку измеряли как изменение максимальной дистанции бега (ΔDIST = DIST1 – DIST2).В каждом поколении 10 самцов и 10 самок с наивысшей реакцией были селекционно скрещены для получения линии HRT, и отдельно 10 пар с наименьшей реакцией на обучение были скрещены для получения линии LRT. Почти 100 потомков на линию в поколении оценивали на реакцию на обучение.
Вкратце, 10–11-недельные самцы и самки из генетически гетерогенного поголовья N: NIH были подвергнуты тесту с постепенным бегом на беговой дорожке, определяющему их предтренировочные способности (DIST1). Затем каждая крыса проходила программу тренировок на беговой дорожке (3 дня в неделю в течение 8 недель). Программа постепенно увеличивала скорость с шагом 1 м/мин и продолжительность на 0,5 мин в каждом сеансе, начиная с 10 м/мин в течение 20 мин на 1-й неделе и заканчивая 21 м/мин в течение 31,5 мин на 8-й неделе. В конце тренировочной программы оценивали способность каждой крысы к физической нагрузке после тренировки (DIST2), а реакцию на тренировку измеряли как изменение максимальной дистанции бега (ΔDIST = DIST1 – DIST2).В каждом поколении 10 самцов и 10 самок с наивысшей реакцией были селекционно скрещены для получения линии HRT, и отдельно 10 пар с наименьшей реакцией на обучение были скрещены для получения линии LRT. Почти 100 потомков на линию в поколении оценивали на реакцию на обучение. Все тренировочные занятия и тесты на максимальную выносливость проводились на моторизованной беговой дорожке с наклоном 15°.
Все тренировочные занятия и тесты на максимальную выносливость проводились на моторизованной беговой дорожке с наклоном 15°.
Тренировка выносливости линий с высокой и низкой чувствительностью
Для этого исследования самцов крыс линии 15 поколения респондеров в возрасте 10–12 недель подвергали испытанию на выносливость.Затем подмножество крыс подвергали 8-недельному протоколу тренировок, как описано выше. Отдельная группа крыс выступала в качестве исходного контроля и не подвергалась никакому обучению, представленная как группы НЗТ и ЗГТ (n = 3 в группе). В конце периода обучения всех крыс (как обученных, так и нетренированных) тестировали на переносимость физической нагрузки для измерения реакции на физическую нагрузку, как описано. Обученные крысы представлены как LRTT и HRTT (n = 3 в группе). Икроножные мышцы вырезали у животных через 48 часов после последнего теста с физической нагрузкой, мгновенно замораживали в жидком азоте и хранили при -80°С при ° для последующего анализа.
Секвенирование РНК
Замороженные икроножные мышцы измельчали с помощью пестика-ступки. Тотальную РНК из 50-100 мг тканевого порошка выделяли с использованием реагента Trizol. РНК обрабатывали ДНКазой и очищали с использованием колонок NucleoSpin RNA II (Machery Nagel), целостность подтверждали с помощью биоанализатора Agilent. Секвенирование РНК проводили в GATC Biotech (Констанц, Германия). Набор для подготовки библиотеки Illumina Stranded TruSeq mRNA Library был использован с 1 мкг тотальной РНК для создания библиотек для секвенирования, которые были загружены в проточную кювету Illumina HiSeq 2500 High-output и секвенированы в формате одиночного чтения 1 × 50 п.н.
Исследование обучения человека
План исследования человека подробно описан в Robinson et al. (20). Вкратце, в исследовании молодые и пожилые здоровые участники были случайным образом разделены на три группы: тренировки с отягощениями (R), высокоинтенсивные интервальные тренировки (HIIT) и комбинированные тренировки (C). Участники тренировались в течение 12 недель, после чего следовали повторные дни тестирования. В этом исследовании в анализ были включены только данные молодых участников. Протокол HIIT состоял из 3 дней в неделю езды на велосипеде и 2 дней в неделю ходьбы на моторизованной беговой дорожке.Интервальная сессия была разработана для достижения 16-минутной тренировки с высокой интенсивностью (> 90% VO 2 макс). Ходьба на беговой дорожке выполнялась в самостоятельно выбранном темпе в течение 45 мин при 70 % VO 2 макс. Комбинированная программа тренировок включала 5 дней в неделю, 30 минут езды на велосипеде, за которыми следовали 30 минут тренировки с отягощениями. Циклирование при 70% VO 2 max составляло 20 мин.
Участники тренировались в течение 12 недель, после чего следовали повторные дни тестирования. В этом исследовании в анализ были включены только данные молодых участников. Протокол HIIT состоял из 3 дней в неделю езды на велосипеде и 2 дней в неделю ходьбы на моторизованной беговой дорожке.Интервальная сессия была разработана для достижения 16-минутной тренировки с высокой интенсивностью (> 90% VO 2 макс). Ходьба на беговой дорожке выполнялась в самостоятельно выбранном темпе в течение 45 мин при 70 % VO 2 макс. Комбинированная программа тренировок включала 5 дней в неделю, 30 минут езды на велосипеде, за которыми следовали 30 минут тренировки с отягощениями. Циклирование при 70% VO 2 max составляло 20 мин.
Измерения митохондриального дыхания использовались в качестве косвенного показателя аэробной работоспособности. Как поясняется в Robinson et al., митохондрии были выделены из биоптатов скелетных мышц до и после тренировки и проанализированы с помощью респирометрии высокого разрешения. Митохондрии добавляли в камеру объемом 2 мл (Oxygraph-2K, Oroboros) с последующим последовательным добавлением субстратов и ингибиторов. В этом исследовании в качестве митохондриального дыхания использовалось дыхание в состоянии 3 через комплекс I+II (глутамат-малат-сукцинат). Образцы биопсии из м. латеральную широкую мышцу бедра собирали в состоянии покоя и натощак через 72 часа после тренировки, чтобы избежать острого воздействия упражнений на количество генов.
Митохондрии добавляли в камеру объемом 2 мл (Oxygraph-2K, Oroboros) с последующим последовательным добавлением субстратов и ингибиторов. В этом исследовании в качестве митохондриального дыхания использовалось дыхание в состоянии 3 через комплекс I+II (глутамат-малат-сукцинат). Образцы биопсии из м. латеральную широкую мышцу бедра собирали в состоянии покоя и натощак через 72 часа после тренировки, чтобы избежать острого воздействия упражнений на количество генов.
Биоинформатический анализ
Для анализа секвенирования РНК контроль качества необработанных прочтений определяли с использованием набора инструментов FastQC (http://www.bioinformatics.babraham.ac.uk/). Затем чтения были сопоставлены с эталонным геномом Rattus Norvegicus (Rnor_6.0), загруженным из ENSEMBL, с использованием выравнивателя Tophat2 (21). Выравнивания были собраны в транскрипты с использованием Cufflinks, и список дифференциально экспрессируемых генов (DEG) был получен путем количественного определения транскриптов с использованием программы Cuffdiff (22).
Анализ путей был проведен с помощью инструмента Ingenuity Pathway Analysis (QIAGEN Inc., https://www.qiagenbioinformatics.com/products/ingenuitypathway-analysis, версия 8.6) и классификации PANTHER Gene Ontology (23). Гены с дифференциальной экспрессией были проанализированы на предмет статистической чрезмерной репрезентативности биологических процессов GO Slim с помощью точного критерия Фишера и поправки Бонферрони для множественных сравнений.
Необработанные подсчеты для набора данных RNA-seq от Robinson et al. (20) были загружены из NCBI (GSE97084).
Для анализа аэробных способностей человека мы сопоставили уровни экспрессии генов в нетренированном состоянии с измерениями митохондриального дыхания перед тренировкой. Поскольку этот анализ основан только на нетренированном состоянии (внутренняя аэробная способность), участники из всех трех групп (HIIT, C и R) были включены для статистической достоверности.
Для анализа реакции человека на физическую нагрузку после сортировки показателей реакции участников на основе изменений в митохондриальном дыхании четыре участника, показавшие максимальное улучшение, были определены как участники с высокой реакцией. Четыре участника с наименьшим изменением митохондриального дыхания были обозначены как слабо реагирующие. Затем мы использовали пакет DESeq2 для нормализации данных, а также для идентификации DEG в состояниях до и после тренировки у людей с высоким и низким ответом (24). Наконец, мы исследовали перекрытие DEG с одинаковой направленностью изменений у людей и крыс для дальнейшего анализа.
Четыре участника с наименьшим изменением митохондриального дыхания были обозначены как слабо реагирующие. Затем мы использовали пакет DESeq2 для нормализации данных, а также для идентификации DEG в состояниях до и после тренировки у людей с высоким и низким ответом (24). Наконец, мы исследовали перекрытие DEG с одинаковой направленностью изменений у людей и крыс для дальнейшего анализа.
Чтобы исследовать предполагаемые вышестоящие регуляторы, мы использовали анализ удаленных регуляторных элементов совместно регулируемых генов (DiRE) (25).Вкратце, программа DIRE идентифицирует 3 основных эволюционно консервативных региона (ECR) и ECR промоторов для каждого гена в представленном наборе данных. После определения этих областей и соответствующих последовательностей он передает эту информацию в алгоритм, который ищет известные мотивы связывания факторов транскрипции (TFBM) и то, насколько они обогащены в представленном наборе генов. Параллельно алгоритм использует 5000 случайно выбранных генов и выполняет тот же анализ, что и фон.
Анализ секретома
Для предсказания секретома мы использовали Emanuelsson et al.(26) протокол в качестве основы для предсказания субклеточной локализации на основе последовательностей. Мы применили следующий пакет с настройками по умолчанию для пайплайна (код доступен публично по адресу https://travis-ci.org/github/fmaguire/predict_secretome). Вкратце, после преобразования названий генов в аминокислотные последовательности с помощью Uniprot конвейер начинается с SignalP для идентификации белков, содержащих сигнальные пептиды. После этого для фильтрации белков с трансмембранными доменами используется TMHMM2.0. (Если сигнальный пептид содержит домен ТМ, он все равно пройдет эту стадию).Затем конвейер идентифицирует N-концевые сортирующие пептиды через TargetP. Наконец, с помощью WolfPsort он идентифицирует последовательности, которые, вероятно, находятся во внеклеточном компартменте.
Статистический анализ
Для профилирования транскриптома из выходных данных программы Cuffdiff все гены, демонстрирующие значение p < 0,05 (двустороннее, с поправкой по методу Бенджамини-Хохберга), были дополнительно исследованы на дифференциальную экспрессию и функциональные процессы.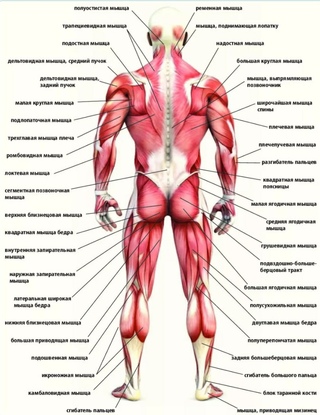
Статистическую значимость приписывали p < 0.05 (двусторонний) и FDR < 0,05, где применимо. Для статистического анализа путей были применены точный критерий Фишера и поправка Бонферрони для множественных сравнений. GraphPad Prism использовали для статистического анализа физиологических данных, которые представлены как среднее ± ± SE.
Для анализа аэробных способностей человека мы провели корреляцию Пирсона для экспрессии генов транскриптома физической нагрузки человека и соответствующего митохондриального дыхания с использованием функции cor.test на языке программирования R.Из полученных результатов гены со значением корреляции p <0,05 (двусторонние) пересекали с DEG в наборе данных аэробной способности крыс (HCR-LCR) для проверки направленности изменений. Сообщалось об изменении генов в одном и том же направлении.
Для анализа реакции человека на физическую нагрузку, после сортировки показателей реакции участников на основе изменений в митохондриальном дыхании, мы определили DEG для состояний до и после тренировки у людей с высокой и низкой реакцией. Для анализа были выбраны перекрывающиеся DEG с одинаковой направленностью изменений у людей и крыс.
Для анализа были выбраны перекрывающиеся DEG с одинаковой направленностью изменений у людей и крыс.
Результаты
Мышечная транскрипционная характеристика способности к аэробным нагрузкам Основные моменты Повышенная васкуляризация и оксигенация
Для исследования молекулярных механизмов, лежащих в основе собственной аэробной способности, мы использовали скелетные мышцы самцов крыс, отобранных на основе их способности к бегу (рис. 1А). Без каких-либо упражнений группа HCR значительно превзошла группу LCR в тесте на максимальную производительность на беговой дорожке (дополнительная фигура 1A). Чтобы определить молекулярные пути, связанные с этой внутренней разницей в переносимости физических упражнений, мы провели глобальный анализ экспрессии генов с помощью массивного параллельного секвенирования РНК на икроножных мышцах этих животных (рис. 1А).Мы наблюдали аналогичный диапазон и распределение прочтений во всех образцах, что подтверждает сопоставимое покрытие транскриптомы.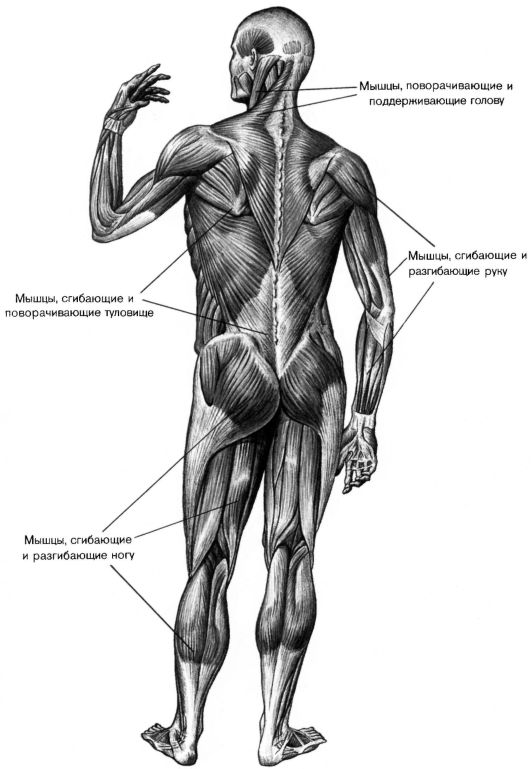 Несмотря на гетерогенный характер этих моделей, отдельные транскриптомы сгруппированы в зависимости от физической нагрузки (рис. 1В). Сравнивая транскриптомы скелетных мышц обеих групп, мы идентифицировали 434 дифференциально экспрессируемых гена (DEG), из которых 257 были повышены в HCR и 177 были повышены в LCR (кратность изменения> 1,5, q <0,05) (рис. 1C). Среди 10 лучших транскриптов с более высокой экспрессией в HCR были гены, связанные с метаболизмом ( Pik3c2g и Mup4 ), пролиферацией и дифференцировкой клеток ( Fam163a , Fosb и Cyp2J4 ) и регуляцией функции кровеносных сосудов. и подача кислорода ( Msln , Cyp2J4 и Hb-b2 ) (рисунок 1C и дополнительный рисунок 1B).Наоборот, несколько из лучших генов с более высокой экспрессией в мышцах LCR (т. е. 10 самых низких в этом анализе) были связаны с миогенезом и регенерацией мышц, включая Tmem8c (также известный как Myomaker), Ranbp3l и Ppm1h . . Igfn1 , негативный регулятор синтеза белка, был высоко экспрессирован в LCR.
Несмотря на гетерогенный характер этих моделей, отдельные транскриптомы сгруппированы в зависимости от физической нагрузки (рис. 1В). Сравнивая транскриптомы скелетных мышц обеих групп, мы идентифицировали 434 дифференциально экспрессируемых гена (DEG), из которых 257 были повышены в HCR и 177 были повышены в LCR (кратность изменения> 1,5, q <0,05) (рис. 1C). Среди 10 лучших транскриптов с более высокой экспрессией в HCR были гены, связанные с метаболизмом ( Pik3c2g и Mup4 ), пролиферацией и дифференцировкой клеток ( Fam163a , Fosb и Cyp2J4 ) и регуляцией функции кровеносных сосудов. и подача кислорода ( Msln , Cyp2J4 и Hb-b2 ) (рисунок 1C и дополнительный рисунок 1B).Наоборот, несколько из лучших генов с более высокой экспрессией в мышцах LCR (т. е. 10 самых низких в этом анализе) были связаны с миогенезом и регенерацией мышц, включая Tmem8c (также известный как Myomaker), Ranbp3l и Ppm1h . . Igfn1 , негативный регулятор синтеза белка, был высоко экспрессирован в LCR. Кроме того, экспрессия Ifit1 была выше в LCR, который кодирует компонент интерферон-индуцированного белкового комплекса, который ранее был связан с воспалением, связанным с ожирением (27).
Кроме того, экспрессия Ifit1 была выше в LCR, который кодирует компонент интерферон-индуцированного белкового комплекса, который ранее был связан с воспалением, связанным с ожирением (27).
Рисунок 1 Транскрипционная характеристика аэробной способности скелетных мышц. (A) Сравнение профилей транскрипции с секвенированием РНК икроножных мышц крыс High Capacity Runner (HCR) и Low Capacity Runner (LCR). (B) Анализ основных компонентов групп HCR и LCR (n = 3). (C) График вулкана, показывающий анализ экспрессии генов, гены с FDR <0,05 показаны синим цветом, а гены выделены красным. (D) Анализ онтологии генов PANTHER, показывающий 5 основных биологических процессов (GO_BP), обогащенных генами с повышенной активностью (красный), и 5 основных терминов, обогащенных генами с отрицательной регуляцией (синий).Иерархически связанные, в значительной степени избыточные термины GO представлены наименьшим набором (кратность обогащения> 5, p <0,05, поправка Бонферрони для множественного сравнения).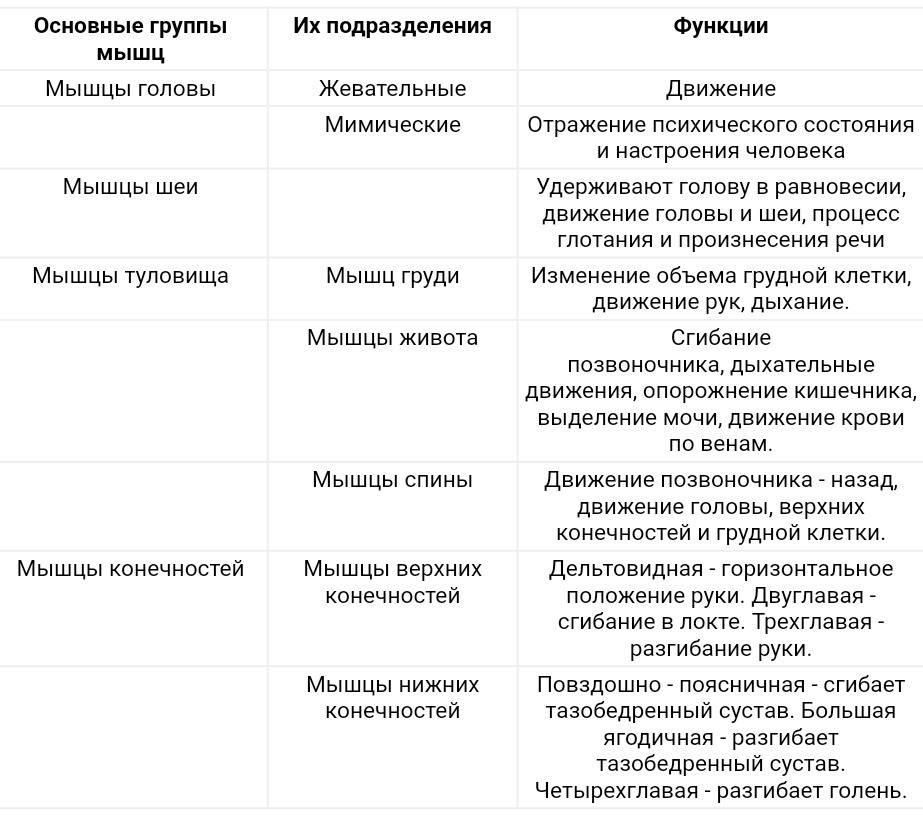 (E, F) Анализ пути изобретательности для биологических функций и вышестоящих регуляторов (входной FDR<0,05, отсечение z-показателя 1,5 с каждой стороны оси, на графике представлены только данные с p <0,05). (G) Дистанционный анализ регуляторных элементов дифференциально регулируемых генов (настроен на 3 верхних ECR + промоторные ECR против 5000 случайных фоновых данных).
(E, F) Анализ пути изобретательности для биологических функций и вышестоящих регуляторов (входной FDR<0,05, отсечение z-показателя 1,5 с каждой стороны оси, на графике представлены только данные с p <0,05). (G) Дистанционный анализ регуляторных элементов дифференциально регулируемых генов (настроен на 3 верхних ECR + промоторные ECR против 5000 случайных фоновых данных).
Чтобы получить общее представление о программах мышечных генов, связанных с повышенной индивидуальной работоспособностью, мы провели анализ генной онтологии (GO) на DEG. Транскрипты, увеличенные в мышцах HCR, показали обогащение генами, связанными со структурой мышц, сократимостью и васкуляризацией (рис. 1D). С другой стороны, гены, связанные с иммунным ответом, были чрезмерно представлены в транскриптоме LCR. Параллельно мы использовали анализ путей изобретательности (IPA) для дальнейшего выявления путей и функций, которые могут способствовать расхождению фенотипов этих моделей (рис.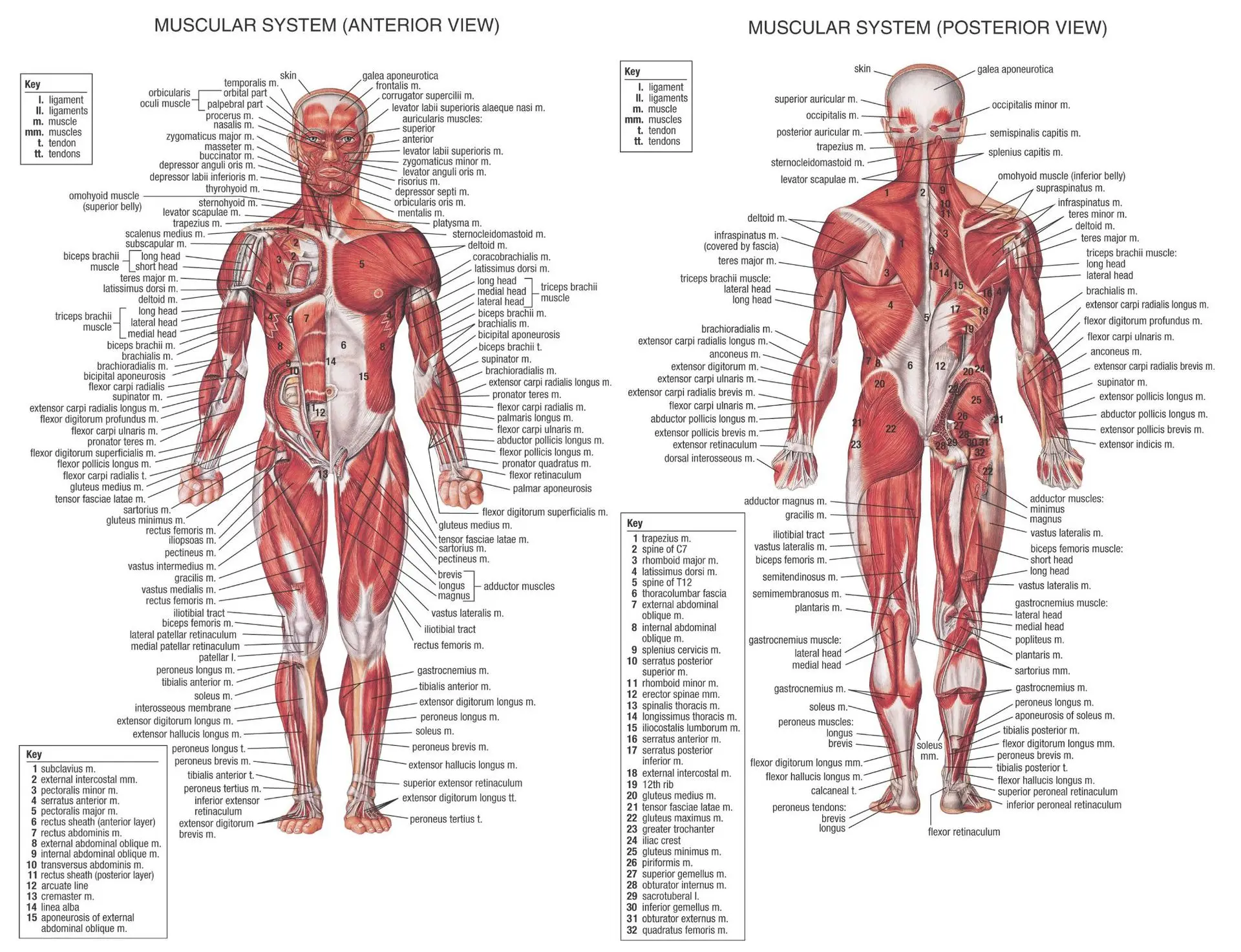 1E).Основываясь на дифференциальной экспрессии генов, IPA предсказывает, активируется ли конкретная биологическая функция (положительный z-показатель) или ингибируется (отрицательный z-показатель). В этом контексте ингибирование функции HCR означает активацию LCR. В соответствии с анализом GO, функции, связанные с ангиогенезом и сократимостью, были широко представлены среди биологических функций, которые, по прогнозам, будут активированы в мышцах HCR. Одной интересной биологической функцией, выделенной в анализе IPA, был фиброгенез, представляющий гены, связанные с ремоделированием внеклеточного матрикса (например,g., Plat , Serpine1 , Thbs1 и F2 ), и которые были повышены в транскриптоме бегуна с высокой емкостью (дополнительная фигура 1C). Биологические функции, активированные в мышечном транскриптоме LCR, включали сроки сердечно-сосудистых заболеваний и смертность, отражающие метаболические проблемы, о которых сообщалось у этих животных (28).
1E).Основываясь на дифференциальной экспрессии генов, IPA предсказывает, активируется ли конкретная биологическая функция (положительный z-показатель) или ингибируется (отрицательный z-показатель). В этом контексте ингибирование функции HCR означает активацию LCR. В соответствии с анализом GO, функции, связанные с ангиогенезом и сократимостью, были широко представлены среди биологических функций, которые, по прогнозам, будут активированы в мышцах HCR. Одной интересной биологической функцией, выделенной в анализе IPA, был фиброгенез, представляющий гены, связанные с ремоделированием внеклеточного матрикса (например,g., Plat , Serpine1 , Thbs1 и F2 ), и которые были повышены в транскриптоме бегуна с высокой емкостью (дополнительная фигура 1C). Биологические функции, активированные в мышечном транскриптоме LCR, включали сроки сердечно-сосудистых заболеваний и смертность, отражающие метаболические проблемы, о которых сообщалось у этих животных (28).
Чтобы изучить, как организованы эти генные программы, мы опросили наборы DEG на наличие предполагаемых вышестоящих регуляторов.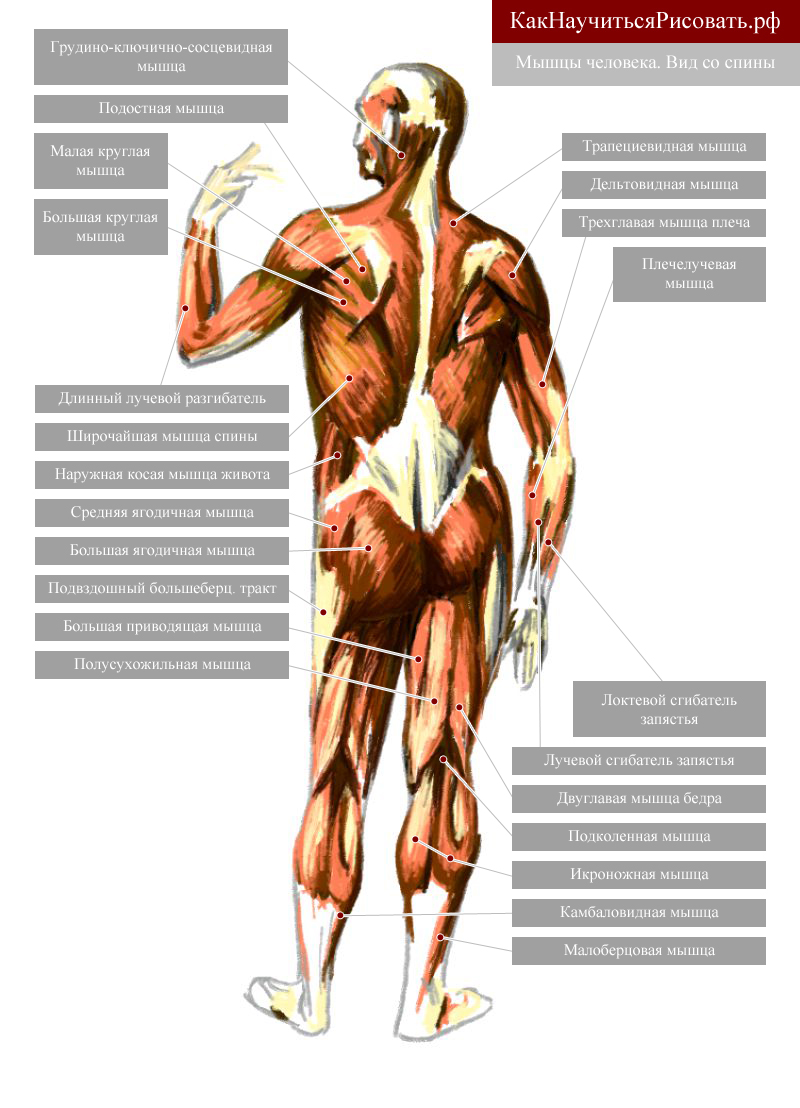 При поиске скоординированных паттернов экспрессии в DEG по сравнению с базами данных коэкспрессии IPA извлек несколько факторов транскрипции, которые, как известно, важны для мышечной функции (Mef2c, PPARγ, SRF и STAT3), активированные в HCR (рис. 1F).Что касается LCR, ДНК-метилазы Dnmt3a и Dnmt3b занимают высокие позиции среди потенциальных регуляторов. Хотя и с меньшим значением, мы также обнаружили регуляторы, которые могут играть роль в состоянии оксигенации и восстановлении после травмы (Nos2 и Egln). Другой способ идентифицировать предполагаемые вышестоящие регуляторы — это сканирование непосредственной близости совместно регулируемых генов на наличие известных мотивов связывания факторов транскрипции (TFBM). С этой целью мы использовали DiRE (дистанционные регуляторные элементы совместно регулируемых генов) (25) для анализа регуляторных областей DEG (рис. 1G).При таком подходе мы снова наблюдали ассоциацию мотивов STAT и PPARγ с генами, связанными с HCR, и идентифицировали дополнительные TF, некоторые из которых, как недавно было показано, модулируют миогенез (Zfp238) (29) и регенерацию мышц (Zfp423) (30).
При поиске скоординированных паттернов экспрессии в DEG по сравнению с базами данных коэкспрессии IPA извлек несколько факторов транскрипции, которые, как известно, важны для мышечной функции (Mef2c, PPARγ, SRF и STAT3), активированные в HCR (рис. 1F).Что касается LCR, ДНК-метилазы Dnmt3a и Dnmt3b занимают высокие позиции среди потенциальных регуляторов. Хотя и с меньшим значением, мы также обнаружили регуляторы, которые могут играть роль в состоянии оксигенации и восстановлении после травмы (Nos2 и Egln). Другой способ идентифицировать предполагаемые вышестоящие регуляторы — это сканирование непосредственной близости совместно регулируемых генов на наличие известных мотивов связывания факторов транскрипции (TFBM). С этой целью мы использовали DiRE (дистанционные регуляторные элементы совместно регулируемых генов) (25) для анализа регуляторных областей DEG (рис. 1G).При таком подходе мы снова наблюдали ассоциацию мотивов STAT и PPARγ с генами, связанными с HCR, и идентифицировали дополнительные TF, некоторые из которых, как недавно было показано, модулируют миогенез (Zfp238) (29) и регенерацию мышц (Zfp423) (30).
Снижение реакции на тренировку на выносливость связано с воспалительной генетической сигнатурой и регуляцией циркадного ритма
Большие межиндивидуальные различия в реакции на тренировку с физическими упражнениями создают серьезные проблемы при разработке стандартизированных вмешательств с физическими упражнениями.Чтобы составить карту транскриптома реакции скелетных мышц на физическую нагрузку, мы использовали выборочно выведенную модельную систему HRT-LRT и тренировали их на беговой дорожке в течение 8 недель, создавая группы HRT-Trained (HRTT) и LRT-Trained (LRTT) (рис. 2A и Дополнительные рисунки 2A – C). Отдельная группа нетренированных животных HRT и LRT прошла тест на максимальную беговую способность до и после 8-недельного тренировочного периода. В то время как производительность HRT и LRT перед тренировкой была одинаковой, HRTT значительно превосходили LRTT в состоянии после тренировки (дополнительные рисунки 2B – C).Отражая это, транскриптомы HRTT и LRTT сгруппированы отдельно (рис. 2B) с 255 дифференциально экспрессируемыми генами (кратность изменения> 1,5, FDR <0,05). Из них 68 были обогащены HRTT, а 187 были понижены в HRTT (т.е. обогащены LRTT). Среди самых повышенных транскриптов в мышцах HRTT были некоторые гены, участвующие в путях, вызванных тренировкой, таких как передача сигналов кальция ( Sln и Adcy1 ), а также некоторые новые гены (Kcna10 и Lgi3 ).(Рисунок 2C и дополнительный рисунок 2D). Интересно, что Lgi3, как сообщается, активирует Akt, хотя и в других типах клеток (31). Верхние транскрипты, обогащенные LRTT, включали негативные регуляторы передачи сигналов кальциневрина ( Rcan3 ), позитивные регуляторы функции NMJ ( Wnt16 и VAT1l ) и медленно сокращающиеся волокна ( Myh6 ). Чтобы исследовать пути и функции, которые потенциально способствуют реакции на обучение, мы исследовали обогащенные наборы генов с анализом GO и биологическими функциями (дополнительные рисунки 2E, F).Транскриптом с высоким ответом выявил сигнатуру, связанную с кровяным давлением, тогда как сигнатура с низким ответом включала циркадную регуляцию (в частности, Per1 , Bhlhe40 , Ciart , Db p, индуцированную экспрессией, и экспрессию Nfil3, репрессированную) и в целом была более разнообразной.
2B) с 255 дифференциально экспрессируемыми генами (кратность изменения> 1,5, FDR <0,05). Из них 68 были обогащены HRTT, а 187 были понижены в HRTT (т.е. обогащены LRTT). Среди самых повышенных транскриптов в мышцах HRTT были некоторые гены, участвующие в путях, вызванных тренировкой, таких как передача сигналов кальция ( Sln и Adcy1 ), а также некоторые новые гены (Kcna10 и Lgi3 ).(Рисунок 2C и дополнительный рисунок 2D). Интересно, что Lgi3, как сообщается, активирует Akt, хотя и в других типах клеток (31). Верхние транскрипты, обогащенные LRTT, включали негативные регуляторы передачи сигналов кальциневрина ( Rcan3 ), позитивные регуляторы функции NMJ ( Wnt16 и VAT1l ) и медленно сокращающиеся волокна ( Myh6 ). Чтобы исследовать пути и функции, которые потенциально способствуют реакции на обучение, мы исследовали обогащенные наборы генов с анализом GO и биологическими функциями (дополнительные рисунки 2E, F).Транскриптом с высоким ответом выявил сигнатуру, связанную с кровяным давлением, тогда как сигнатура с низким ответом включала циркадную регуляцию (в частности, Per1 , Bhlhe40 , Ciart , Db p, индуцированную экспрессией, и экспрессию Nfil3, репрессированную) и в целом была более разнообразной. Когда мы рассмотрели потенциальные вышестоящие регуляторы, факторы транскрипции Foxo4 и Atf4, известные своей ролью в атрофии скелетных мышц (32–35), оказались среди предсказанных факторов, чьи мишени были обогащены транскриптомом LRTT (рис. 2D).Это согласуется с отчетами, показывающими отсутствие реакции на силовые тренировки у LRTT (36). Кроме того, факторы, связанные с иммунным ответом (Ccr2, TNF, IL10ra и p53), также были среди факторов, активирующих мышцу LRTT, что указывает на ответ на повреждение. В подтверждение этого отдельный поиск потенциальных регуляторов выявил обогащение TFBM NFkB50 (гомодимер p50-p50), известным репрессором транскрипции и регулятором иммунных и воспалительных реакций (рис. 2Е).
Когда мы рассмотрели потенциальные вышестоящие регуляторы, факторы транскрипции Foxo4 и Atf4, известные своей ролью в атрофии скелетных мышц (32–35), оказались среди предсказанных факторов, чьи мишени были обогащены транскриптомом LRTT (рис. 2D).Это согласуется с отчетами, показывающими отсутствие реакции на силовые тренировки у LRTT (36). Кроме того, факторы, связанные с иммунным ответом (Ccr2, TNF, IL10ra и p53), также были среди факторов, активирующих мышцу LRTT, что указывает на ответ на повреждение. В подтверждение этого отдельный поиск потенциальных регуляторов выявил обогащение TFBM NFkB50 (гомодимер p50-p50), известным репрессором транскрипции и регулятором иммунных и воспалительных реакций (рис. 2Е).
Рисунок 2 Транскрипционный профиль адаптивного ответа на физическую нагрузку в скелетных мышцах. (A) Крыс с высокой и низкой реакцией тренировали на беговой дорожке в течение 8 недель, получая крыс с низкой и высокой реакцией на тренированных крыс (LRTT и HRTT). (B) Анализ основных компонентов транскриптомов скелетных мышц LRTT и HRTT (n = 3) (C) График вулкана, показывающий анализ экспрессии генов, гены с FDR <0,05 показаны синим цветом, а выделенные гены - красным. (D) Анализ пути изобретательности, показывающий прогнозируемые вышестоящие регуляторы (входной FDR < 0.05, пороговое значение z-показателя равно 1,5 с каждой стороны оси, на график нанесены только данные с p <0,05). (E) Факторы транскрипции для дифференциально регулируемых генов, предсказанные с помощью анализа отдаленных регуляторных элементов (настроены на 3 верхних ECR + промоторные ECR против 5000 случайных фоновых значений). (F) Сравнение диаграмм Венна профилей транскрипции с высокой емкостью и высокой реакцией и анализ онтологии генов Panther для биологических процессов для соответствующих сигнатур генов. Иерархически отсортированные термины GO (кратное обогащение> 10, p <0.05, поправка Бонферони для множественного тестирования) были ранжированы по наименьшему p-значению, и на график были нанесены первые 5.
(B) Анализ основных компонентов транскриптомов скелетных мышц LRTT и HRTT (n = 3) (C) График вулкана, показывающий анализ экспрессии генов, гены с FDR <0,05 показаны синим цветом, а выделенные гены - красным. (D) Анализ пути изобретательности, показывающий прогнозируемые вышестоящие регуляторы (входной FDR < 0.05, пороговое значение z-показателя равно 1,5 с каждой стороны оси, на график нанесены только данные с p <0,05). (E) Факторы транскрипции для дифференциально регулируемых генов, предсказанные с помощью анализа отдаленных регуляторных элементов (настроены на 3 верхних ECR + промоторные ECR против 5000 случайных фоновых значений). (F) Сравнение диаграмм Венна профилей транскрипции с высокой емкостью и высокой реакцией и анализ онтологии генов Panther для биологических процессов для соответствующих сигнатур генов. Иерархически отсортированные термины GO (кратное обогащение> 10, p <0.05, поправка Бонферони для множественного тестирования) были ранжированы по наименьшему p-значению, и на график были нанесены первые 5.
Сравнение индивидуальных эффектов внутренних и приобретенных компонентов способности к физической нагрузке может оказаться полезным для исследования общих и уникальных механизмов, лежащих в основе. Диаграмма Венна профилей транскрипции выявила уникальные наборы генов для каждого компонента (рис. 2F). Уникальные транскрипты внутренней емкости (n = 333) отражают преобладающую сигнатуру мышечной функции, обогащенную организацией саркомеров, сокращением скелетных мышц и регуляцией биологических процессов переноса ионов Ca ++ .С другой стороны, сигнатура для приобретенной способности (n = 163) демонстрирует более фрагментированную сигнатуру с ассоциациями с циркадной регуляцией, трансмембранным транспортом протонов и окислительно-восстановительным процессом. 63 общих транскрипта были в основном связаны с ангиогенезом, метаболическим процессом АФК и передачей сигналов Wnt, все из которых известны как процессы, индуцированные физической нагрузкой.
Поскольку реакция на тренировку HRT и LRT различается, мы затем рассмотрели, могут ли различия в изменениях их транскрипционных профилей, вызванных физическими упражнениями, выявить гены-кандидаты, связанные с активацией или ингибированием таких адаптаций к упражнениям. Чтобы изучить этот угол, мы сравнили каждую группу в состоянии после обучения с их соответствующим состоянием до обучения (дополнительная фигура 2G). Большинство индуцированных тренировкой генов, общих для людей с низкой и высокой реакцией на физическую нагрузку, регулировались в одном и том же направлении. Уникальные гены низкой реакции на тренировку (n = 193) были обогащены терминами GO, связанными с иммунным ответом (воспаление и интерфероновые ответы и интернализация рецепторов) и эффектами упражнений (ангиогенез, транспорт синаптических везикул и метаболический процесс АФК).Наоборот, генная сигнатура, уникальная для высокого ответа, была обогащена процессами, касающимися транспорта ионов, передачи сигналов Ca ++ и окислительного дыхания.
Чтобы изучить этот угол, мы сравнили каждую группу в состоянии после обучения с их соответствующим состоянием до обучения (дополнительная фигура 2G). Большинство индуцированных тренировкой генов, общих для людей с низкой и высокой реакцией на физическую нагрузку, регулировались в одном и том же направлении. Уникальные гены низкой реакции на тренировку (n = 193) были обогащены терминами GO, связанными с иммунным ответом (воспаление и интерфероновые ответы и интернализация рецепторов) и эффектами упражнений (ангиогенез, транспорт синаптических везикул и метаболический процесс АФК).Наоборот, генная сигнатура, уникальная для высокого ответа, была обогащена процессами, касающимися транспорта ионов, передачи сигналов Ca ++ и окислительного дыхания.
Предсказанные мышечные секретомы способности к нагрузке и ответной реакции выдвигают на первый план метаболизм липидов и ремоделирование внеклеточного матрикса
Секретируемые факторы мышц играют важную роль в опосредовании локальной и системной адаптации, связанной с физической нагрузкой (37). Мы использовали данные транскриптомики, чтобы идентифицировать известные или новые миокины, участвующие в фенотипах способности к физической нагрузке или реакции (рис. 3).С этой целью профили транскриптома были обработаны с помощью хорошо зарекомендовавшего себя конвейера для предсказания предполагаемых секретируемых факторов на основе белковых последовательностей (26). Ресурс секретома и мембранного протеома человека, составленный в рамках инициативы Human Protein Atlas (HPA) (38), позволяет нам определить местоположение каждой молекулы-кандидата (рис. 3A–C). Основываясь на транскриптомах емкости и ответа, мы смогли предсказать некоторые известные секретируемые факторы (Lpl, Plat, Adipoq, Lyz2, Igfbp6 и Tcn2) и некоторые предполагаемые.Интересно, что значительная часть прогнозируемой способности секретома классифицируется как потенциально секретируемая в кровь. Почти все соответствующие транскрипты были значительно увеличены в транскриптоме HCR (рис. 3А). Среди них выделяются представители плазминовой системы ( F2/Thrombin, Plat, Serping1 и Serpinf), метаболизма липидов (ApoD, Adipoq, Ces1, Lpl и Pla2g12a) и иммунной функции (Ccl21, Cfi и Slpi).
Мы использовали данные транскриптомики, чтобы идентифицировать известные или новые миокины, участвующие в фенотипах способности к физической нагрузке или реакции (рис. 3).С этой целью профили транскриптома были обработаны с помощью хорошо зарекомендовавшего себя конвейера для предсказания предполагаемых секретируемых факторов на основе белковых последовательностей (26). Ресурс секретома и мембранного протеома человека, составленный в рамках инициативы Human Protein Atlas (HPA) (38), позволяет нам определить местоположение каждой молекулы-кандидата (рис. 3A–C). Основываясь на транскриптомах емкости и ответа, мы смогли предсказать некоторые известные секретируемые факторы (Lpl, Plat, Adipoq, Lyz2, Igfbp6 и Tcn2) и некоторые предполагаемые.Интересно, что значительная часть прогнозируемой способности секретома классифицируется как потенциально секретируемая в кровь. Почти все соответствующие транскрипты были значительно увеличены в транскриптоме HCR (рис. 3А). Среди них выделяются представители плазминовой системы ( F2/Thrombin, Plat, Serping1 и Serpinf), метаболизма липидов (ApoD, Adipoq, Ces1, Lpl и Pla2g12a) и иммунной функции (Ccl21, Cfi и Slpi). . Примечательно, что почти все прогнозируемые факторы профиля ответа были обогащены LRTT (уменьшены HRTT).Самая большая часть прогнозируемого профиля секретома для реакции на физическую нагрузку была отнесена к ECM как локализация секретома HPA и включала факторы, связанные с передачей сигналов Wnt (Wisp2, Wnt16 и Sostdc) (рис. 3B). Интересно, что общие факторы для двух профилей включают белки, связанные с ремоделированием ВКМ (Col1a1, Col14a1, Ecm1, Thbs2, Cilp2 и Serpinf1), чувствительностью к инсулину и метаболизмом (Igfbp6 и Pla2g2a) (39) (рис. 3C).
. Примечательно, что почти все прогнозируемые факторы профиля ответа были обогащены LRTT (уменьшены HRTT).Самая большая часть прогнозируемого профиля секретома для реакции на физическую нагрузку была отнесена к ECM как локализация секретома HPA и включала факторы, связанные с передачей сигналов Wnt (Wisp2, Wnt16 и Sostdc) (рис. 3B). Интересно, что общие факторы для двух профилей включают белки, связанные с ремоделированием ВКМ (Col1a1, Col14a1, Ecm1, Thbs2, Cilp2 и Serpinf1), чувствительностью к инсулину и метаболизмом (Igfbp6 и Pla2g2a) (39) (рис. 3C).
Рисунок 3 Секретомный прогноз высокой переносимости физических нагрузок и высокой реакции на физические нагрузки.Дифференциально экспрессируемые гены высокой способности и высокой реакции использовали для поиска предсказанных секретируемых факторов. Факторы, уникальные для (A) с высокой емкостью и (B) с высокой реакцией мышечных секретомов, показаны разделенными по местоположению на основе данных из Атласа белков человека: кровь (розовый), внеклеточный матрикс (синий), внутриклеточный и мембранный (желтый). и другие ткани/неизвестные местоположения (серые). Активированные транскрипты отмечены красной стрелкой, а синей стрелкой отмечены транскрипты с пониженной экспрессией. (C) Факторы, которые являются общими для мышечных профилей с высокой производительностью и высокой реакцией, показаны в прямоугольниках с одинаковым цветовым кодом (кровь — красным, внеклеточный матрикс — синим, а другие — серым).
и другие ткани/неизвестные местоположения (серые). Активированные транскрипты отмечены красной стрелкой, а синей стрелкой отмечены транскрипты с пониженной экспрессией. (C) Факторы, которые являются общими для мышечных профилей с высокой производительностью и высокой реакцией, показаны в прямоугольниках с одинаковым цветовым кодом (кровь — красным, внеклеточный матрикс — синим, а другие — серым).
Генные сети работоспособности и реакции у людей
Затем мы стремились определить молекулярные факторы, влияющие на аэробные способности человека, и их потенциал для повышения выносливости аналогично моделям крыс, которые мы использовали для этого исследования. Для этого мы использовали наборы данных RNA-seq от Robinson et al.(20), чтобы найти корреляции между относительными уровнями экспрессии генов, физической нагрузкой и реакцией (рис. 4А). В этом исследовании здоровые молодые и пожилые участники подвергались различным модальностям упражнений, и подход множественной омики был направлен на поиск молекулярных преобразователей преимуществ упражнений в скелетных мышцах.
Рисунок 4 Молекулярные медиаторы внутренней работоспособности и реакции на физическую нагрузку у человека. (A) Схема, показывающая набор данных и рабочий процесс, использованный для анализа с использованием данных Robinson et.др. (B) Корреляционный анализ Пирсона измерения митохондриального дыхания скелетных мышц перед тренировкой и уровней экспрессии генов у здоровых молодых людей (n = 29, p < 0,05, на график были нанесены только образцы с нормальным распределением). (C) Субъекты ранжированы на основе изменений в митохондриальном дыхании после аэробных упражнений. Первые 4 человека обозначены как высоко реагирующие (красные), а нижние 4 субъекта сгруппированы как слабо реагирующие (синие). Транскриптомы скелетных мышц субъектов с высоким и низким ответом сравнивали путем повторного анализа данных секвенирования РНК. (D) Сравнение профилей транскриптома до обучения показано на графике вулкана (n = 4), транскрипты с FDR <0,05 показаны синим цветом. (E) Сравнение диаграмм Венна предварительно обученных мышечных транскриптомов крыс с высокой реакцией и человека. (F) Вулканический график для дифференциально экспрессируемых генов между транскриптомами скелетных мышц после тренировки у людей с высоким и низким ответом. (G) Сравнение транскриптома после обучения для крыс с высоким ответом и человека.
(E) Сравнение диаграмм Венна предварительно обученных мышечных транскриптомов крыс с высокой реакцией и человека. (F) Вулканический график для дифференциально экспрессируемых генов между транскриптомами скелетных мышц после тренировки у людей с высоким и низким ответом. (G) Сравнение транскриптома после обучения для крыс с высоким ответом и человека.
Используя измерения митохондриального дыхания мышц перед тренировкой в качестве функционального индикатора внутренней аэробной способности и РНК-последовательности у тех же добровольцев (дополнительная таблица 1), мы искали корреляции между этими двумя показателями. В результате этого анализа мы идентифицировали 574 гена, которые положительно коррелируют со значениями митохондриального дыхания мышц. Внутри этой группы гены попали в биологические процессы, связанные с регенерацией мышц (в первую очередь, с передачей сигналов Notch), метаболизмом и гомеостазом кальция.Меньшее количество генов отрицательно коррелировало с митохондриальной функцией и, среди прочего, сгруппировано под процессингом митоходриальной РНК и регуляцией аутофагии (рис. 4B и дополнительная рис. 3A). Из набора генов, которые показали корреляцию с митохондриальным дыханием (положительную или отрицательную), мы смогли найти 16 среди DEG HCR/LCR (дополнительная таблица 2). Восемь из этих генов регулировались в одном направлении как у людей, так и у крыс (рис. 4В и дополнительная таблица 2). MUTYH , кодирующий фермент, называемый гликозилазой MYH, участвующий в восстановлении окислительного повреждения ДНК, показал наивысшую оценку корреляции. Кроме того, уровни транскрипции транскрипционного фактора Круппеля, подобного фактору 2 ( KLF2 ) и протеинкиназы 1D ( PRKD1/ PKD1), также показали положительную корреляцию с функцией митохондрий и способностью к аэробным упражнениям. Примечательно, что кластер коррелированных генов ( TRIO , COL1A1 и THBS2 ) указывает на роль реорганизации внеклеточного матрикса и цитоскелета в определении переносимости физической нагрузки.
4B и дополнительная рис. 3A). Из набора генов, которые показали корреляцию с митохондриальным дыханием (положительную или отрицательную), мы смогли найти 16 среди DEG HCR/LCR (дополнительная таблица 2). Восемь из этих генов регулировались в одном направлении как у людей, так и у крыс (рис. 4В и дополнительная таблица 2). MUTYH , кодирующий фермент, называемый гликозилазой MYH, участвующий в восстановлении окислительного повреждения ДНК, показал наивысшую оценку корреляции. Кроме того, уровни транскрипции транскрипционного фактора Круппеля, подобного фактору 2 ( KLF2 ) и протеинкиназы 1D ( PRKD1/ PKD1), также показали положительную корреляцию с функцией митохондрий и способностью к аэробным упражнениям. Примечательно, что кластер коррелированных генов ( TRIO , COL1A1 и THBS2 ) указывает на роль реорганизации внеклеточного матрикса и цитоскелета в определении переносимости физической нагрузки.
Чтобы исследовать гены и сети, связанные с приспособляемостью к упражнениям, мы использовали изменение митохондриального дыхания мышц после 12-недельной программы тренировок в качестве индикатора реакции на упражнения.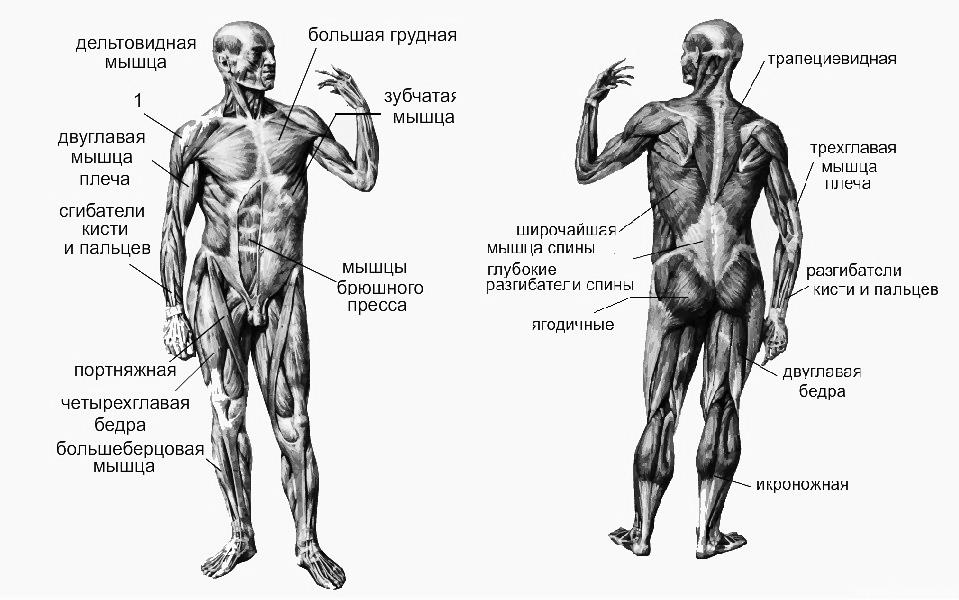 Участники были ранжированы в соответствии с изменением митохондриального дыхания и сгруппированы как с высоким и низким ответом (рисунок 4C и дополнительный рисунок 3B). Сравнивая профили транскрипции этих групп в их состояниях перед тренировкой, мы определили четыре гена, дифференциально экспрессируемые у людей, хорошо реагирующих на физические нагрузки ( DYNC1I1 , HSPA4L , AC008014.1 и FOXO1 ) (рис. 4D). При перекрестном сравнении с транскрипционными сигнатурами крыс с высоким ответом в их предтренировочном состоянии FOXO1 становится общим фактором в соответствии с этой моделью адаптивности к упражнениям (рис. 4Е). В посттренировочном состоянии анализ дифференциальной экспрессии индивидуумов с высоким и низким ответом выявил восемь генов, четыре из которых имели увеличенные транскрипты ( NEU4 , HSPA4L , ND2 и SETD9 ), а остальные четыре уменьшилось ( PIWIL2 , MYZAP , VWA7 и GCOM1 ) (рис.
Участники были ранжированы в соответствии с изменением митохондриального дыхания и сгруппированы как с высоким и низким ответом (рисунок 4C и дополнительный рисунок 3B). Сравнивая профили транскрипции этих групп в их состояниях перед тренировкой, мы определили четыре гена, дифференциально экспрессируемые у людей, хорошо реагирующих на физические нагрузки ( DYNC1I1 , HSPA4L , AC008014.1 и FOXO1 ) (рис. 4D). При перекрестном сравнении с транскрипционными сигнатурами крыс с высоким ответом в их предтренировочном состоянии FOXO1 становится общим фактором в соответствии с этой моделью адаптивности к упражнениям (рис. 4Е). В посттренировочном состоянии анализ дифференциальной экспрессии индивидуумов с высоким и низким ответом выявил восемь генов, четыре из которых имели увеличенные транскрипты ( NEU4 , HSPA4L , ND2 и SETD9 ), а остальные четыре уменьшилось ( PIWIL2 , MYZAP , VWA7 и GCOM1 ) (рис. 4F).Путем сравнения DEG после обучения с профилем транскрипции обученных крыс с высокой реакцией и определения Vwa7 как общего фактора (рис. 4G). Vwa7 представляет собой повсеместно экспрессируемый ген, кодирующий предположительно секретируемый белок с неизвестной функцией.
4F).Путем сравнения DEG после обучения с профилем транскрипции обученных крыс с высокой реакцией и определения Vwa7 как общего фактора (рис. 4G). Vwa7 представляет собой повсеместно экспрессируемый ген, кодирующий предположительно секретируемый белок с неизвестной функцией.
Обсуждение
Взаимодействие между врожденной и приобретенной способностью к физической нагрузке, обе из которых имеют высокую индивидуальную вариабельность (7, 40), представляет собой препятствие для определения точных механизмов, связывающих аэробную работоспособность и здоровье человека.Используя данные об экспрессии генов у крыс и человека, связанные с определенными функциональными показателями, связанными с физическими упражнениями, мы обнаружили транскрипционные сигнатуры мышц, лежащие в основе врожденной аэробной способности и реакции на тренировку у обоих видов.
Транскрипционный профиль мышц HCR тесно связан с путями, которые регулируют васкуляризацию и транспорт кислорода. Эти молекулярные признаки перекликаются с ранее опубликованными функциональными данными той же модели, демонстрирующей повышенное поглощение кислорода и более высокую плотность капилляров в мышцах HCR (11–13, 41), и оба являются хорошо известными адаптациями к аэробным упражнениям как у грызунов, так и у людей (42). Напротив, паттерны экспрессии генов в мышцах LCR показали обогащенный профиль функций, связанных с иммунным ответом. Важно отметить, что существует установленная связь между отсутствием физической активности и хроническим вялотекущим воспалением. Фактически бездействие связано с повышенной заболеваемостью и смертностью от хронических патологий (2, 43). Интересно, что мышечный транскриптом LCR связан с биологическими функциями, связанными с сердечно-сосудистыми заболеваниями, что дополнительно отражает их плохое состояние здоровья, как было задокументировано ранее (28, 44-48).Примечательно, что тот же анализ также выявил любопытную связь между заболеваемостью и смертностью. Взятые вместе, все это предполагает, что определенные нами транскрипционные сигнатуры отражают некоторые подтвержденные функциональные результаты и, следовательно, могут быть использованы для получения нового понимания механизмов, имеющих решающее значение для способности к аэробным упражнениям.
Напротив, паттерны экспрессии генов в мышцах LCR показали обогащенный профиль функций, связанных с иммунным ответом. Важно отметить, что существует установленная связь между отсутствием физической активности и хроническим вялотекущим воспалением. Фактически бездействие связано с повышенной заболеваемостью и смертностью от хронических патологий (2, 43). Интересно, что мышечный транскриптом LCR связан с биологическими функциями, связанными с сердечно-сосудистыми заболеваниями, что дополнительно отражает их плохое состояние здоровья, как было задокументировано ранее (28, 44-48).Примечательно, что тот же анализ также выявил любопытную связь между заболеваемостью и смертностью. Взятые вместе, все это предполагает, что определенные нами транскрипционные сигнатуры отражают некоторые подтвержденные функциональные результаты и, следовательно, могут быть использованы для получения нового понимания механизмов, имеющих решающее значение для способности к аэробным упражнениям.
Профили транскрипции могут быть полезны для изучения того, как определенные генные программы регулируются согласованным образом.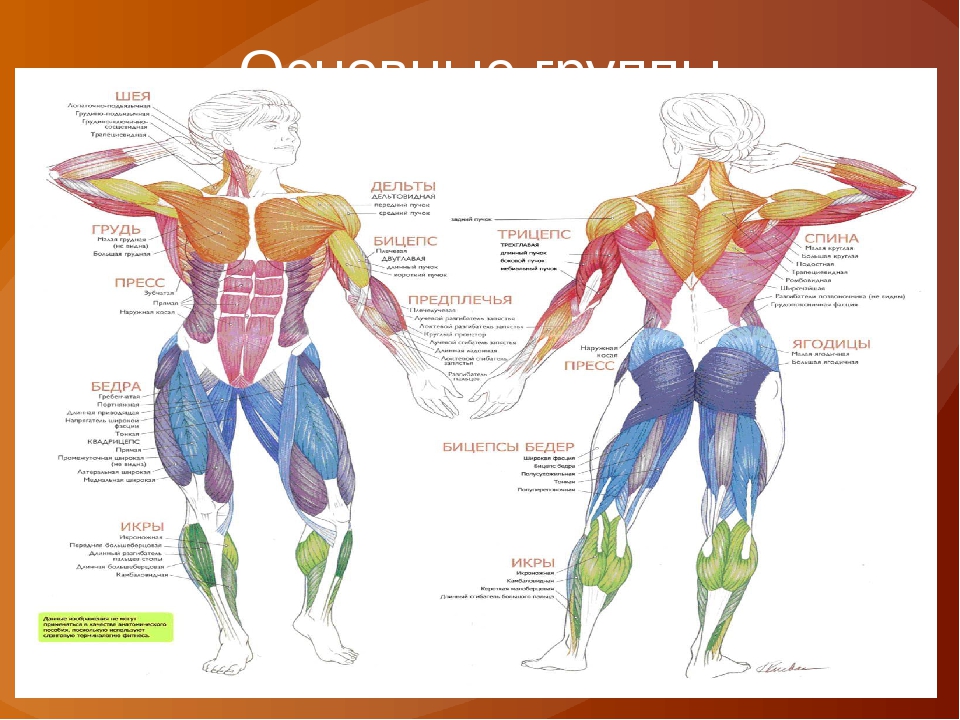 Идентификация потенциальных факторов транскрипции и корегуляторов, связанных с экстремальными фенотипами способности и ответа, может дать нам ключ к пониманию движущих сил дивергентных механизмов.Одним из таких примеров были ДНК-метилазы Dnmt3a и Dnmt3b , активность которых, как было предсказано, снижена в HCR (активирована в LCR). Интересно, что ранее было показано, что активность Dnmt3b ограничивает биоэнергетическую адаптацию к упражнениям в мышцах за счет усиления промоторного метилирования специфических генов (49). Метилирование ДНК в значительной степени связано с трансгенерационным наследованием различных признаков, включая реакцию на физическую нагрузку (50), и может быть частью возможного регуляторного механизма для передачи способности к физической нагрузке следующим поколениям.
Идентификация потенциальных факторов транскрипции и корегуляторов, связанных с экстремальными фенотипами способности и ответа, может дать нам ключ к пониманию движущих сил дивергентных механизмов.Одним из таких примеров были ДНК-метилазы Dnmt3a и Dnmt3b , активность которых, как было предсказано, снижена в HCR (активирована в LCR). Интересно, что ранее было показано, что активность Dnmt3b ограничивает биоэнергетическую адаптацию к упражнениям в мышцах за счет усиления промоторного метилирования специфических генов (49). Метилирование ДНК в значительной степени связано с трансгенерационным наследованием различных признаков, включая реакцию на физическую нагрузку (50), и может быть частью возможного регуляторного механизма для передачи способности к физической нагрузке следующим поколениям.
Наш анализ также показал, что некоторые ТФ являются потенциальными медиаторами аэробной способности и реакции. Пространственно-временная экспрессия TFs играет критическую роль в развитии мышц, метаболизме и их адаптации к физическим нагрузкам (3, 4).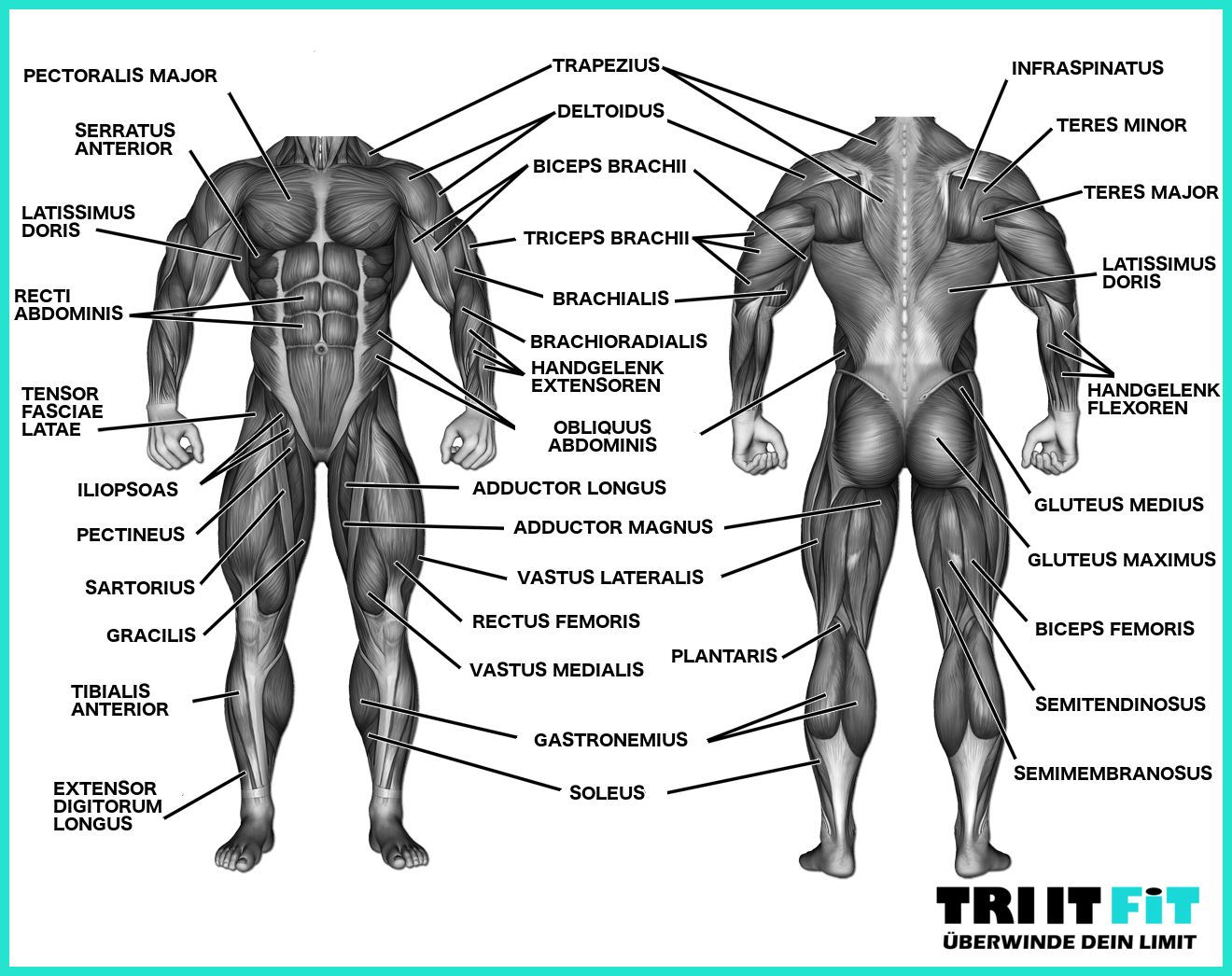 Основываясь на наших данных, одним кандидатом на роль регулятора физической нагрузки является RP58 (также известный как Zfp238), который, как было установлено, модулирует миогенез развития (29). Другим является ROAZ (Zfp423), роль которого недавно была установлена в связи с его ролью в регенерации мышц путем стимулирования роста сателлитных клеток (30), которые важны для восстановления/ремоделирования мышечных волокон в ответ на нагрузку (51).
Основываясь на наших данных, одним кандидатом на роль регулятора физической нагрузки является RP58 (также известный как Zfp238), который, как было установлено, модулирует миогенез развития (29). Другим является ROAZ (Zfp423), роль которого недавно была установлена в связи с его ролью в регенерации мышц путем стимулирования роста сателлитных клеток (30), которые важны для восстановления/ремоделирования мышечных волокон в ответ на нагрузку (51).
Выявление детерминант и посредников приобретенной способности открывает большие перспективы для настройки программ упражнений на основе индивидуальной тренируемости. Транскрипционная сигнатура реакции скелетных мышц на физическую нагрузку может показаться менее очевидной по сравнению с сигнатурой внутренней емкости. Хотя имеются убедительные доказательства того, что фенотип HCR в основном определяется скелетными мышцами (11, 13), другие органы или системы могут играть большую роль в фенотипе тренируемости.Регулярные физические упражнения требуют постоянной нагрузки на организм, включая сердечно-сосудистую систему и несократительные механические компоненты, такие как суставы и сухожилия (42). Кроме того, широко известно, что центральная нервная система участвует в регуляции обмена веществ и может играть роль в физической нагрузке и способности к обучению, поскольку она регулирует двигательную функцию и поведенческие компоненты (например, мотивацию). Любой из этих или других факторов также может ограничивать адаптацию к упражнениям у людей с низким ответом.На самом деле обучение активировало большую генную программу у людей с низкой реакцией, чем у людей с высокой реакцией. Заманчиво предположить, что у людей с низкой реакцией может существовать ингибирующая сигнальная сеть, препятствующая адаптации к тренировкам. Новые данные указывают на повышенную воспалительную сигнализацию и повышенную метаболическую дисфункцию у крыс LRTT (52). Интересно, что циркадианная регуляция экспрессии генов выделяется при анализе путей сигнатуры ответа по сравнению с транскриптомом емкости. Известно, что упражнения синхронизируются с биологическими часами, и есть доказательства того, что время дня влияет на эффективность упражнений (53).
Кроме того, широко известно, что центральная нервная система участвует в регуляции обмена веществ и может играть роль в физической нагрузке и способности к обучению, поскольку она регулирует двигательную функцию и поведенческие компоненты (например, мотивацию). Любой из этих или других факторов также может ограничивать адаптацию к упражнениям у людей с низким ответом.На самом деле обучение активировало большую генную программу у людей с низкой реакцией, чем у людей с высокой реакцией. Заманчиво предположить, что у людей с низкой реакцией может существовать ингибирующая сигнальная сеть, препятствующая адаптации к тренировкам. Новые данные указывают на повышенную воспалительную сигнализацию и повышенную метаболическую дисфункцию у крыс LRTT (52). Интересно, что циркадианная регуляция экспрессии генов выделяется при анализе путей сигнатуры ответа по сравнению с транскриптомом емкости. Известно, что упражнения синхронизируются с биологическими часами, и есть доказательства того, что время дня влияет на эффективность упражнений (53).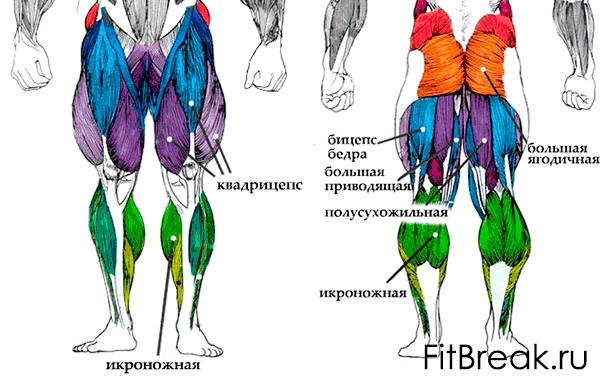 Наконец, было высказано предположение, что основным фактором реакции у людей является интенсивность физических упражнений (54). Чтобы уменьшить этот возможный смешанный эффект, мы использовали данные вмешательства человека, включающего высокоинтенсивные интервальные тренировки (20), которые действительно привели к улучшению VO 2 max и митохондриального дыхания почти у всех участников.
Наконец, было высказано предположение, что основным фактором реакции у людей является интенсивность физических упражнений (54). Чтобы уменьшить этот возможный смешанный эффект, мы использовали данные вмешательства человека, включающего высокоинтенсивные интервальные тренировки (20), которые действительно привели к улучшению VO 2 max и митохондриального дыхания почти у всех участников.
Выявление новых секретируемых мышцами факторов может предложить новые способы использования некоторых преимуществ физической активности. На основании мышечного транскриптома соответствующий секретомный профиль HCR указывает на более высокое присутствие факторов, секретируемых в кровь.Заманчиво предположить, что мышцы HCR легко оснащены для отправки необходимых циркулирующих факторов, способствующих высокой аэробной производительности, таких как те, которые участвуют в метаболизме липидов и улучшенном кровообращении. Основываясь на предсказаниях мышечного секретома, ApoD вместе с Lpl и Ces1d может способствовать поглощению жирных кислот мышцами. Это может способствовать ранее описанному эффективному использованию топлива и энергетическому метаболизму в мышцах HCR (55). С другой стороны, профиль респондеров предполагает увеличение секретируемых факторов, прежде всего во внеклеточный матрикс.LRT могут сигнализировать о ремоделировании ECM из-за неспособности справиться с длительным стрессом от тренировок. Это согласуется с предыдущими исследованиями сетей, регулируемых в ответ на упражнения как у крыс, так и у людей (52, 56). Расширение наших знаний о секретируемых мышцами факторах, регулируемых внутренней или адаптивной реакцией на физическую нагрузку, может проложить путь к индивидуальной терапии, основанной на физических упражнениях.
Это может способствовать ранее описанному эффективному использованию топлива и энергетическому метаболизму в мышцах HCR (55). С другой стороны, профиль респондеров предполагает увеличение секретируемых факторов, прежде всего во внеклеточный матрикс.LRT могут сигнализировать о ремоделировании ECM из-за неспособности справиться с длительным стрессом от тренировок. Это согласуется с предыдущими исследованиями сетей, регулируемых в ответ на упражнения как у крыс, так и у людей (52, 56). Расширение наших знаний о секретируемых мышцами факторах, регулируемых внутренней или адаптивной реакцией на физическую нагрузку, может проложить путь к индивидуальной терапии, основанной на физических упражнениях.
Предпосылка беспородных, генетически гетерогенных крысиных моделей физической способности частично отражает сложность метаболических заболеваний и разнообразие среди людей.Мы использовали набор данных о людях от Robinson et al. (20) для поиска моста между профилями транскриптома аэробной способности и обучаемости у крыс и людей. Несмотря на четкие межвидовые различия и небольшую когорту, мы смогли идентифицировать 8 генов, экспрессия которых коррелирует с внутренней работоспособностью у людей и имеет общий транскриптом физической способности крыс. Среди них экспрессия Prkd1 (PKD1) положительно коррелирует с аэробными физическими нагрузками у людей. PKD1 представляет собой киназу HDAC класса II, которая, как было показано, способствует фенотипу устойчивости к медленному сокращению в скелетных мышцах (57).Это интригует, поскольку наследственные факторы, влияющие на способность к аэробным нагрузкам, могут включать эпигенетические модификации. Фактически, накопленные данные свидетельствуют о том, что эпигенетические метки могут опосредовать передачу эффектов упражнений и диеты следующему поколению (50). Сравнение сигнатур ответа показало, что Foxo1 является общим фактором как у крыс, так и у людей. Foxo1 является транскрипционным фактором, который, как известно, индуцируется в условиях атрофии скелетных мышц, способствуя аутофагии и убиквитин-протеосомной системе (32).
Несмотря на четкие межвидовые различия и небольшую когорту, мы смогли идентифицировать 8 генов, экспрессия которых коррелирует с внутренней работоспособностью у людей и имеет общий транскриптом физической способности крыс. Среди них экспрессия Prkd1 (PKD1) положительно коррелирует с аэробными физическими нагрузками у людей. PKD1 представляет собой киназу HDAC класса II, которая, как было показано, способствует фенотипу устойчивости к медленному сокращению в скелетных мышцах (57).Это интригует, поскольку наследственные факторы, влияющие на способность к аэробным нагрузкам, могут включать эпигенетические модификации. Фактически, накопленные данные свидетельствуют о том, что эпигенетические метки могут опосредовать передачу эффектов упражнений и диеты следующему поколению (50). Сравнение сигнатур ответа показало, что Foxo1 является общим фактором как у крыс, так и у людей. Foxo1 является транскрипционным фактором, который, как известно, индуцируется в условиях атрофии скелетных мышц, способствуя аутофагии и убиквитин-протеосомной системе (32). Что касается реакции на физическую нагрузку, повышенные уровни Foxo1 у людей с низкой реакцией могут указывать на повышенную деградацию белка для очистки поврежденных белков в результате длительного тренировочного стресса.
Что касается реакции на физическую нагрузку, повышенные уровни Foxo1 у людей с низкой реакцией могут указывать на повышенную деградацию белка для очистки поврежденных белков в результате длительного тренировочного стресса.
Это исследование заложило основу для изучения молекулярной карты переносимости физической нагрузки и реакции. Среди различных транскрипционных сигнатур, связанных с проанализированными нами показателями, связанными с физическими нагрузками, ангиогенез и доставка кислорода выступают в качестве основных факторов внутренней работоспособности. И наоборот, провоспалительная передача сигналов связана с более низкой адаптацией к тренировкам.Лучшее понимание лежащих в основе механизмов может в конечном итоге проложить путь к персонализированным программам обучения, которые оптимизируют результаты для здоровья в зависимости от индивидуальных потребностей.
Ограничения
В этом исследовании мы использовали уникальные модели грызунов для определения способности к физической нагрузке, чтобы определить наборы генов/транскриптов, которые способствуют внутренним и адаптивным компонентам способности к физической нагрузке. Следует отметить, что стратегия разведения, используемая для создания этих моделей крыс, может привести к фиксации генетических мутаций с усилением или потерей функции, которые мы не исследовали.При анализе мы отметили высокую степень изменчивости, что можно объяснить внепородным характером этих моделей. Дальнейшие исследования с более крупными когортами или целевыми подходами могли бы изучить потенциальные связи, отмеченные в этом исследовании, и пролить свет на лежащие в их основе механизмы. Наш исследовательский подход, сравнивающий наборы данных о людях и грызунах, не указывает на причинно-следственную связь и служит ключом для дальнейшей оценки. Наконец, масса тела может влиять на аэробную работоспособность, а также играет существенную роль в возникновении метаболических и сердечно-сосудистых заболеваний.Многие поколения селекции по аэробным способностям привели к значительной корреляции с живой массой у линий HCR и LCR. Однако увеличение массы тела в LCR и снижение HCR больше не расходились после поколения 13 и, как сообщалось, стабилизировались на уровне 0,7–0,8 раза к поколению 28 (9).
Следует отметить, что стратегия разведения, используемая для создания этих моделей крыс, может привести к фиксации генетических мутаций с усилением или потерей функции, которые мы не исследовали.При анализе мы отметили высокую степень изменчивости, что можно объяснить внепородным характером этих моделей. Дальнейшие исследования с более крупными когортами или целевыми подходами могли бы изучить потенциальные связи, отмеченные в этом исследовании, и пролить свет на лежащие в их основе механизмы. Наш исследовательский подход, сравнивающий наборы данных о людях и грызунах, не указывает на причинно-следственную связь и служит ключом для дальнейшей оценки. Наконец, масса тела может влиять на аэробную работоспособность, а также играет существенную роль в возникновении метаболических и сердечно-сосудистых заболеваний.Многие поколения селекции по аэробным способностям привели к значительной корреляции с живой массой у линий HCR и LCR. Однако увеличение массы тела в LCR и снижение HCR больше не расходились после поколения 13 и, как сообщалось, стабилизировались на уровне 0,7–0,8 раза к поколению 28 (9). Кроме того, взяв массу тела и генерацию в качестве предикторов аэробной способности в множественном регрессионном анализе, Wisloff et al. (28) показали, что, несмотря на то, что масса тела объясняет некоторую вариацию дистанции бега (7% для женщин, 14–20% для мужчин), большая часть вариации связана с другими факторами.
Кроме того, взяв массу тела и генерацию в качестве предикторов аэробной способности в множественном регрессионном анализе, Wisloff et al. (28) показали, что, несмотря на то, что масса тела объясняет некоторую вариацию дистанции бега (7% для женщин, 14–20% для мужчин), большая часть вариации связана с другими факторами.
Заявление о доступности данных
Наборы данных RNAseq, созданные для этого исследования, можно найти в NCBI Gene Expression Omnibus (GEO) (https://www.ncbi.nlm.nih.gov/geo/) под регистрационным номером GSE155230. Робинсон и др. Доступ к наборам данных RNAseq можно получить в GEO с регистрационным номером GSE97084. Модели крыс с низкой и высокой внутренней способностью к физической нагрузке и низкой и высокой реакцией на физическую нагрузку на тренировку поддерживаются в качестве международного ресурса при поддержке Департамента физиологии и фармакологии Медицинского колледжа Университета Толедо, Толедо, Огайо.Свяжитесь с LK ([email protected]) или SB (brittons@umich. edu) для получения информации о моделях крыс.
edu) для получения информации о моделях крыс.
Заявление об этике
Исследования с участием людей были рассмотрены и одобрены Институциональным наблюдательным советом клиники Мэйо и зарегистрированы как клинические испытания № NCT01477164 и № NCT01738568 (clinicaltrials.gov). Пациенты/участники предоставили письменное информированное согласие на участие в этом исследовании. Исследование на животных было рассмотрено и одобрено университетским комитетом по использованию и уходу за животными Мичиганского университета.
Вклад авторов
JLR, PRJ и YK разработали исследование. LGK, SLB, HW, KSN, MMR и JLR предоставили ресурсы. IC, JZ, PRJ и YK провели анализ данных. YK и JLR написали первоначальный вариант рукописи. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Работа выполнена при поддержке Шведского исследовательского совета (2016-00785) и Фонда Ново Нордиск (NNF19OC0054132) JR., Фондов Веннера-Грена (Швеция) (UPD2017-0175), FAPESP (Бразилия) (14 /26797-5) в PJ, Шведское общество медицинских исследований в IC, Национальный центр развития трансляционных наук (UL1TR000135) в KSN и Национальные институты здравоохранения (США) (T32DK7352 в MR и R01AG09531 в KSN и P40ODO21331 в ЛК и СБ).
Конфликт интересов
JR является консультантом Bayer AG.
Остальные авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы хотели бы поблагодарить д-ра Хорхе Коррейа и д-ра Хосе Морейра за проницательные обсуждения и д-ра Хорхе Коррейа за конструктивную критику рукописи.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fendo.2020.591476/full#supplementary-material
Ссылки
1. Myers J, Prakash M, Froelicher V, Do D, Partington S, Atwood JE. Толерантность к физической нагрузке и смертность среди мужчин, направленных на тестирование с физической нагрузкой. N Engl J Med (2002) 346:793–801. doi: 10.1056/NEJMoa011858
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
2.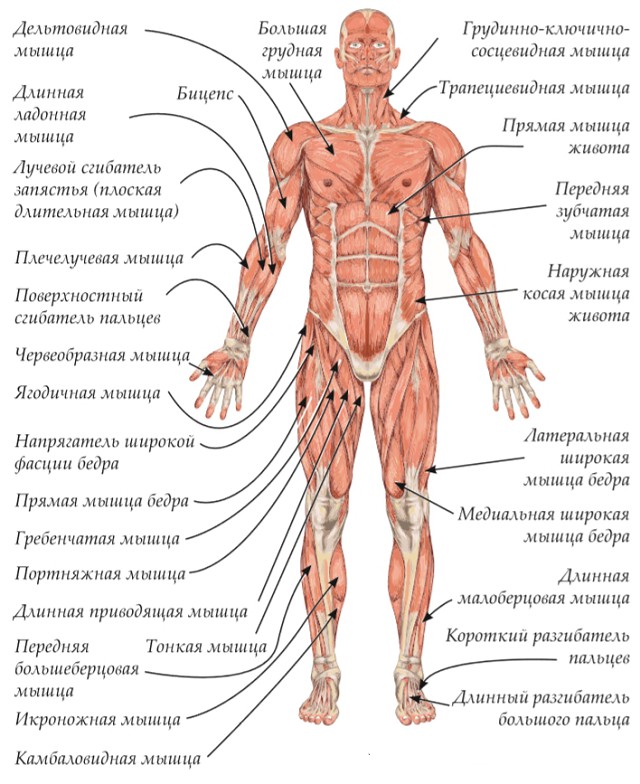 Мандсагер К., Харб С., Кремер П., Фелан Д., Ниссен С.Е., Джабер В. Ассоциация кардиореспираторной подготовки с долгосрочной смертностью среди взрослых, проходящих тестирование на беговой дорожке. JAMA Netw Open (2018) 1:e183605. doi: 10.1001/jamanetworkopen.2018.3605
Мандсагер К., Харб С., Кремер П., Фелан Д., Ниссен С.Е., Джабер В. Ассоциация кардиореспираторной подготовки с долгосрочной смертностью среди взрослых, проходящих тестирование на беговой дорожке. JAMA Netw Open (2018) 1:e183605. doi: 10.1001/jamanetworkopen.2018.3605
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
5. Bouchard C, Daw EW, Rice T, Perusse L, Gagnon J, Province MA, et al. Семейное сходство VO2max в сидячем положении: семейное исследование HERITAGE. Med Sci Sports Exerc (1998) 30: 252–8. doi: 10.1097/00005768-199802000-00013
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
6. Bouchard C, Rankinen T, Chagnon YC, Rice T, Perusse L, Gagnon J, et al.Геномное сканирование для максимального поглощения кислорода и его реакции на тренировку в семейном исследовании HERITAGE. J Appl Physiol (1985) (2000) 88:551–9. doi: 10.1152/jappl.2000.88.2.551
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
7. Bouchard C, An P, Rice T, Skinner JS, Wilmore JH, Gagnon J, et al. Семейная агрегация отклика VO(2max) на тренировку: результаты семейного исследования HERITAGE. J Appl Physiol (1985) (1999) 87:1003–8. дои: 10.1152/jappl.1999.87.3.1003
Bouchard C, An P, Rice T, Skinner JS, Wilmore JH, Gagnon J, et al. Семейная агрегация отклика VO(2max) на тренировку: результаты семейного исследования HERITAGE. J Appl Physiol (1985) (1999) 87:1003–8. дои: 10.1152/jappl.1999.87.3.1003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
9. Ren YY, Overmyer KA, Qi NR, Treutelaar MK, Heckenkamp L, Kalahar M, et al. Генетический анализ крысиной модели аэробной способности и метаболической приспособленности. PLoS One (2013) 8:e77588. doi: 10.1371/journal.pone.0077588
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
10. Кох Л.Г., Бриттон С.Л. Теоретическая и биологическая оценка связи между низкой физической нагрузкой и риском заболевания. Cold Spring Harb Perspect Med (2018) 8:1–15. doi: 10.1101/cshperspect.a029868
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
11. Howlett RA, Gonzalez NC, Wagner HE, Fu Z, Britton SL, Koch LG, et al.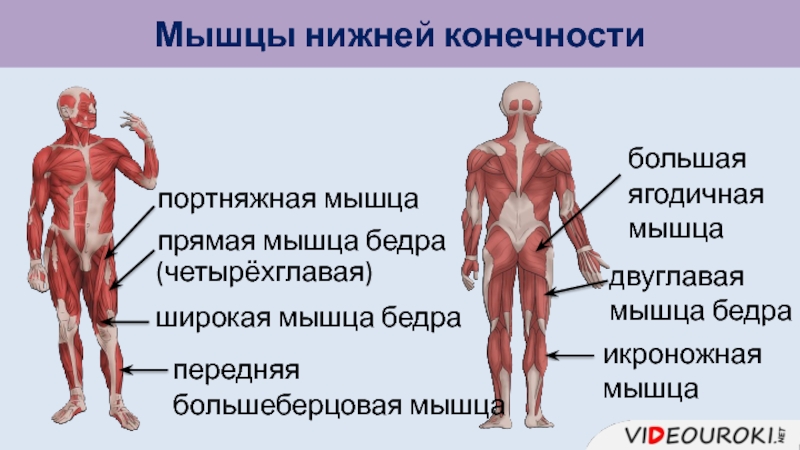 Избранный вклад: капиллярность скелетных мышц и активность ферментов у крыс, отобранных для беговой выносливости. J Appl Physiol (1985) (2003) 94:1682–8. doi: 10.1152/japplphysiol.00556.2002
Избранный вклад: капиллярность скелетных мышц и активность ферментов у крыс, отобранных для беговой выносливости. J Appl Physiol (1985) (2003) 94:1682–8. doi: 10.1152/japplphysiol.00556.2002
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
12.Howlett RA, Kirkton SD, Gonzalez NC, Wagner HE, Britton SL, Koch LG, et al. Периферический транспорт и утилизация кислорода у крыс после продолжительной селекции на выносливость при беге. J Appl Physiol (1985) (2009) 106:1819–25. doi: 10.1152/japplphysiol.00914.2007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
13. Gonzalez NC, Howlett RA, Henderson KK, Koch LG, Britton SL, Wagner HE, et al. Системный транспорт кислорода у крыс, искусственно отобранных на беговую выносливость. Respir Physiol Neurobiol (2006) 151:141–50. doi: 10.1016/j.resp.2005.09.012
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
14. Bray MS, Hagberg JM, Perusse L, Rankinen T, Roth SM, Wolfarth B, et al.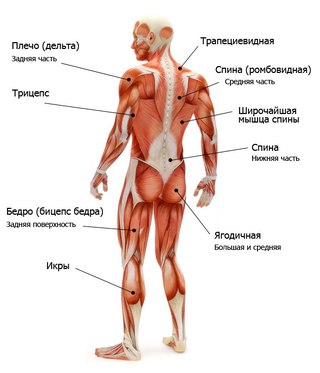 Карта генов человека для фенотипов работоспособности и физической подготовки, связанных со здоровьем: обновление 2006–2007 гг. Med Sci Sports Exerc (2009) 41:35–73. doi: 10.1249/MSS.0b013e3181844179
Карта генов человека для фенотипов работоспособности и физической подготовки, связанных со здоровьем: обновление 2006–2007 гг. Med Sci Sports Exerc (2009) 41:35–73. doi: 10.1249/MSS.0b013e3181844179
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
16.Кох Л.Г., Поллотт Г.Э., Бриттон С.Л. Селекционно выведенная модельная система крыс для низкой и высокой реакции на тренировки. Physiol Genomics (2013) 45:606–14. doi: 10.1152/physiolgenomics.00021.2013
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
17. Kivela R, Silvennoinen M, Lehti M, Rinnankoski-Tuikka R, Purhonen T, Ketola T, et al. Центроиды экспрессии генов, которые связаны с низкой внутренней способностью к аэробным нагрузкам и риском комплексных заболеваний. FASEB J (2010) 24:4565–74.doi: 10.1096/fj.10-157313
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
18. Bye A, Hoydal MA, Catalucci D, Langaas M, Kemi OJ, Beisvag V, et al. Профилирование экспрессии генов скелетных мышц у тренированных и малоподвижных крыс с врожденным высоким и низким VO2max. Physiol Genomics (2008) 35:213–21. doi: 10.1152/physiolgenomics.
Профилирование экспрессии генов скелетных мышц у тренированных и малоподвижных крыс с врожденным высоким и низким VO2max. Physiol Genomics (2008) 35:213–21. doi: 10.1152/physiolgenomics.
.2008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
19. Pinto SK, Lamon S, Stephenson EJ, Kalanon M, Mikovic J, Koch LG, et al.Экспрессия микроРНК и белков-мишеней в скелетных мышцах крыс, селективно выведенных на высокую и низкую беговую способность. Am J Physiol Endocrinol Metab (2017) 313:E335–43. doi: 10.1152/ajpendo.00043.2017
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
20. Robinson MM, Dasari S, Konopka AR, Johnson ML, Manjunatha S, Esponda RR, et al. Улучшенная трансляция белка лежит в основе улучшенной метаболической и физической адаптации к различным режимам тренировок у молодых и пожилых людей. Cell Metab (2017) 25:581–92. doi: 10.1016/j.cmet.2017.02.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
21. Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: точное выравнивание транскриптомов при наличии вставок, делеций и слияний генов. Genome Biol (2013) 14:R36. doi: 10.1186/gb-2013-14-4-r36
Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: точное выравнивание транскриптомов при наличии вставок, делеций и слияний генов. Genome Biol (2013) 14:R36. doi: 10.1186/gb-2013-14-4-r36
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
22. Trapnell C, Roberts A, Goff L, Pertea G, Kim D, Kelley DR, et al.Дифференциальный анализ экспрессии генов и транскриптов в экспериментах по секвенированию РНК с помощью TopHat и Cufflinks. Nat Protoc (2012) 7:562–78. doi: 10.1038/nprot.2012.016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
23. Mi H, Muruganujan A, Ebert D, Huang X, Thomas PD. PANTHER версии 14: больше геномов, новый PANTHER GO-slim и улучшения в инструментах анализа обогащения. Nucleic Acids Res (2019) 47:D419–26. doi: 10.1093/nar/gky1038
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
26.Эмануэльссон О., Брунак С., фон Хейне Г. , Нильсен Х. Поиск белков в клетке с использованием TargetP, SignalP и связанных с ними инструментов. Nat Protoc (2007) 2:953–71. doi: 10.1038/nprot.2007.131
, Нильсен Х. Поиск белков в клетке с использованием TargetP, SignalP и связанных с ними инструментов. Nat Protoc (2007) 2:953–71. doi: 10.1038/nprot.2007.131
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
27. McManus DD, Beaulieu LM, Mick E, Tanriverdi K, Larson MG, Keaney JF Jr., et al. Взаимосвязь между циркулирующими воспалительными белками, экспрессией генов тромбоцитов и сердечно-сосудистым риском. Arterioscler Thromb Vasc Biol (2013) 33:2666–73.doi: 10.1161/ATVBAHA.112.301112
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
28. Wisloff U, Najjar SM, Ellingsen O, Haram PM, Swoap S, Al-Share Q, et al. Факторы сердечно-сосудистого риска возникают после искусственного отбора на низкую аэробную способность. Наука (2005) 307:418–20. doi: 10.1126/science.1108177
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
29. Yokoyama S, Ito Y, Ueno-Kudoh H, Shimizu H, Uchibe K, Albini S, et al.Системный подход показывает, что сеть генома миогенеза регулируется репрессором транскрипции RP58. Dev Cell (2009) 17:836–48. doi: 10.1016/j.devcel.2009.10.011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
30. Addison WN, Hall KC, Kokabu S, Matsubara T, Fu MM, Gori F, et al. Zfp423 регулирует регенерацию и пролиферацию скелетных мышц. Мол Селл Биол (2019) 39 (8): e00447-18. doi: 10.1128/MCB.00447-18
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
31.Kim IW, Jeong HS, Kwon NS, Baek KJ, Yun HY, Kim DS. LGI3 способствует дифференцировке кератиноцитов человека по пути Akt. Exp Dermatol (2018) 27:1224–9. doi: 10.1111/exd.13766
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
32. Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, Picard A, et al. Факторы транскрипции Foxo индуцируют связанную с атрофией убиквитинлигазу атрогин-1 и вызывают атрофию скелетных мышц. Cell (2004) 117:399–412. дои: 10.1016/S0092-8674(04)00400-3
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
33. Mandai S, Mori T, Nomura N, Furusho T, Arai Y, Kikuchi H, et al. WNK1 регулирует гипертрофию клеток скелетных мышц, модулируя ядерную локализацию и транскрипционную активность FOXO4. Научный представитель (2018) 8:9101. doi: 10.1038/s41598-018-27414-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
34. Ebert SM, Monteys AM, Fox DK, Bongers KS, Shields BE, Malmberg SE, et al.Фактор транскрипции ATF4 способствует атрофии скелетных миофибрилл во время голодания. Мол Эндокринол (2010) 24:790–9. doi: 10.1210/me.2009-0345
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
35. Ebert SM, Bullard SA, Basisty N, Marcotte GR, Skopec ZP, Dierdorff JM, et al. Активирующий фактор транскрипции 4 (ATF4) способствует атрофии скелетных мышц за счет образования гетеродимера с регулятором транскрипции C/EBPbeta. J Biol Chem (2020) 295:2787–803.doi: 10.1074/jbc.RA119.012095
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
36. Ахтиайнен Дж. П., Ленсу С., Руотсалайнен И., Шуманн М., Ихалайнен Дж. К., Фачада В. и др. Физиологическая адаптация к тренировке с отягощениями у крыс, отобранных для низкой и высокой реакции на тренировку с аэробными упражнениями. Exp Physiol (2018) 103:1513–23. doi: 10.1113/EP087144
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
39. Kuefner MS, Pham K, Redd JR, Stephenson EJ, Harvey I, Deng X, et al.Секреторная фосфолипаза А2 группы IIA модулирует чувствительность к инсулину и метаболизм. J Lipid Res (2017) 58:1822–33. doi: 10.1194/jlr.M076141
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
40. Bouchard C, Rankinen T. Индивидуальные различия в реакции на регулярную физическую активность. Med Sci Sports Exerc (2001) Дополнение 4S446-51; обсуждение S452-3. doi: 10.1097/00005768-200106001-00013
Полный текст CrossRef | Академия Google
41.Хендерсон К.К., Вагнер Х., Фаврет Ф., Бриттон С.Л., Кох Л.Г., Вагнер П.Д. и др. Детерминанты максимального поглощения O(2) у крыс, отобранных для выносливости при беге. J Appl Physiol (1985) (2002) 93:1265–74. doi: 10.1152/japplphysiol.00809.2001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
43. Erikssen G, Liestol K, Bjornholt J, Thaulow E, Sandvik L, Erikssen J. Изменения в физической подготовке и изменения в смертности. Ланцет (1998) 352:759–62.doi: 10.1016/S0140-6736(98)02268-5
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
44. Hoydal MA, Stolen TO, Johnsen AB, Alvez M, Catalucci D, Condorelli G, et al. Сниженная аэробная способность вызывает негерметичность рианодиновых рецепторов, которые вызывают аритмию у крыс, искусственно отобранных и выведенных для низкой аэробной беговой способности. Acta Physiol (Oxf) (2014) 210:854–64. doi: 10.1111/apha.12238
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
45.Моррис Э.М., Джекман М.Р., Джонсон Г.К., Лю Т.В., Лопес Д.Л., Кирни М.Л. и др. Собственная аэробная способность влияет на восприимчивость к острому стеатозу печени, вызванному диетой с высоким содержанием жиров. Am J Physiol Endocrinol Metab (2014) 307:E355–364. doi: 10.1152/ajpendo.00093.2014
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
46. Кох Л.Г., Бриттон С.Л., Вислофф У. Модельная система крыс для изучения комплексных рисков заболеваний, физической формы, старения и долголетия. Trends Cardiovasc Med (2012) 22:29–34.doi: 10.1016/j.tcm.2012.06.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
47. Koch LG, Kemi OJ, Qi N, Leng SX, Bijma P, Gilligan LJ, et al. Собственная аэробная способность устанавливает границу между старением и долголетием. Circ Res (2011) 109:1162–72. doi: 10.1161/CIRCRESAHA.111.253807
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
48. Morris EM, Meers GME, Ruegsegger GN, Wankhade UD, Robinson T, Koch LG, et al. Внутренняя высокая аэробная способность у самцов крыс защищает от резистентности к инсулину, вызванной диетой. Эндокринология (2019) 160:1179–92. doi: 10.1210/en.2019-00118
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
49. Barres R, Osler ME, Yan J, Rune A, Fritz T, Caidahl K, et al. Не-CpG метилирование промотора PGC-1alpha через DNMT3B контролирует плотность митохондрий. Cell Metab (2009) 10:189–98. doi: 10.1016/j.cmet.2009.07.011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
51. Snijders T, Nederveen JP, McKay BR, Joanisse S, Verdijk LB, van Loon LJ, et al.Клетки-сателлиты в пластичности скелетных мышц человека. Фронт Физиол (2015) 6:283. doi: 10.3389/fphys.2015.00283
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
52. Lessard SJ, Rivas DA, Alves-Wagner AB, Hirshman MF, Gallagher IJ, Constantin-Teodosiu D, et al. Сопротивление аэробным тренировкам вызывает метаболическую дисфункцию и выявляет новые сигнальные сети, регулируемые физическими упражнениями. Диабет (2013) 62:2717–27. doi: 10.2337/db13-0062
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
54.Сиссон С.Б., Кацмарзик П.Т., Эрнест С.П., Бушар С., Блэр С.Н., Черч Т.С. Объем физических упражнений и отсутствие реакции на физическую форму у женщин в постменопаузе, ведущих малоподвижный образ жизни. Med Sci Sports Exerc (2009) 41: 539–45. doi: 10.1249/MSS.0b013e3181896c4e
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
55. Overmyer KA, Evans CR, Qi NR, Minogue CE, Carson JJ, Chermside-Scabbo CJ, et al. Максимальная окислительная способность во время упражнений связана с выбором топлива скелетных мышц и динамическими изменениями в ацетилировании митохондриальных белков. Cell Metab (2015) 21:468–78. doi: 10.1016/j.cmet.2015.02.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
56. Kjaer M, Magnusson P, Krogsgaard M, Boysen Moller J, Olesen J, Heinemeier K, et al. Адаптация внеклеточного матрикса сухожилий и скелетных мышц к физической нагрузке. J Anat (2006) 208:445–50. doi: 10.1111/j.1469-7580.2006.00549.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
57. Kim MS, Fielitz J, McAnally J, Shelton JM, Lemon DD, McKinsey TA, et al.Протеинкиназа D1 стимулирует активность MEF2 в скелетных мышцах и повышает мышечную активность. Mol Cell Biol (2008) 28:3600–9. doi: 10.1128/MCB.00189-08
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Анатомия, функции и общие состояния
Обзор
Что такое мышцы бедра?
Бедра содержат одни из самых крупных мышц тела. Мышцы бедра позволяют нижней части тела сгибаться, сгибаться и вращаться. Они также несут большую часть веса тела и удерживают бедра и ноги на одном уровне, а также обеспечивают равновесие и помогают ему.
Мышцы бедра можно сгруппировать в зависимости от их функции и расположения:
- Аддукторы.
- Подколенные сухожилия.
- Пектиней.
- Квадрицепсы, часто называемые квадрицепсами.
- Сарториус.
Функция
Каково назначение мышц бедра?
Каждая группа мышц бедра выполняет свою работу:
- Аддукторы: Позволяет сводить бедра друг к другу, что называется аддукцией.Они помогают сохранять равновесие, удерживают ноги и бедра на одной линии и позволяют вращать бедра и ноги.
- Подколенные сухожилия: Позволяет разгибать (наклонять) бедро, чтобы отвести ногу назад, например, когда вы идете, поставив одну ногу позади себя. Они также позволяют сгибать (сгибать) колено, например, когда вы приседаете.
- Pectineus: Позволяет сгибать и вращать бедро в тазобедренном суставе. Это также помогает стабилизировать таз.
- Quadriceps: Позволяет сгибать бедро (например, приседая или сидя) или разгибать колено (стоя или вытягивая ногу прямо перед собой, чтобы сделать шаг).
- Sartorius: Помогает сгибать и вращать бедро в тазобедренном суставе. Вы используете его, когда скрещиваете ноги, чтобы положить одну лодыжку на противоположную ногу. Другие примеры включают сидение со скрещенными ногами на полу или сгибание и вращение ноги, чтобы посмотреть на нижнюю часть стопы.
Анатомия
Где расположены приводящие, подколенные сухожилия, гребенчатые, четырехглавые и портняжные мышцы?
- Приводящие мышцы включают пять мышц: тонкую, наружную запирательную, короткую приводящую, длинную приводящую и большую приводящую.Они находятся на внутренней стороне бедра, начиная с таза и заканчивая бедренной костью (бедренной костью).
- Подколенные сухожилия представляют собой группу из трех мышц: полусухожильной, полуперепончатой и двуглавой бедра. Они спускаются по задней части бедра, от бедра чуть ниже колена.
- Pectineus — плоская мышца, простирающаяся от передней части таза до верхушки бедренной кости.
- Четырехглавая мышца включает четыре крупные мышцы, расположенные в передней части бедра: латеральную широкую, медиальную широкую, промежуточную широкую и прямую мышцу бедра.Они начинаются от таза (тазовая кость) и бедренной кости (бедренная кость) и распространяются вниз к надколеннику (коленная чашечка) и большеберцовой кости (голень).
- Портняжная мышца — длинная тонкая мышца — самая длинная в человеческом теле. Она начинается от бедра и огибает переднюю часть бедра, а затем спускается к колену.
Из чего состоят мышцы бедра?
Мышцы бедра относятся к скелетным мышцам. Они прикрепляются к костям с помощью сухожилий и помогают двигать части скелета.
Они состоят из пучков мышечных волокон, содержащих кровеносные сосуды и нервные волокна.Все компоненты скреплены соединительной тканью.
Эти мышцы очень эластичны, поэтому они могут сильно растягиваться.
Условия и расстройства
Какие состояния могут повлиять на мышцы бедра?
Наиболее распространенными проблемами мышц бедра являются мышечные напряжения, растяжения и разрывы. Этот тип травмы возникает, когда мышца растягивается за ее пределы, и мышечные волокна расходятся.
Растяжения часто встречаются в подколенных сухожилиях и четырехглавых мышцах.Они часто возникают во время занятий спортом или других упражнений, когда человеку приходится быстро менять направление или он сталкивается с чем-то или кем-то. Примеры включают футбол и футбол.
Симптомы растяжения мышц бедра включают:
- Синяк, который развивается быстро или в течение следующих нескольких дней после травмы.
- Боль, обычно внезапная и сильная, может усиливаться при сгибании или разгибании бедра или колена.
- Хлопающий или щелкающий звук или ощущение в бедре.
- Отек.
уход
Как сохранить здоровье и безопасность мышц бедра?
Вы можете предпринять шаги, чтобы сохранить мышцы бедер более безопасными и здоровыми, особенно во время упражнений:
- Избегайте видов спорта, требующих быстрой смены направления движения или захвата мяча.
- Не «играй сквозь боль». Если что-то болит, остановитесь и отдохните.
- Соблюдайте здоровую диету с высоким содержанием белка.
- Часто занимайтесь спортом. Чем сильнее ваши мышцы, тем меньше вероятность их травм.
- Поддерживайте здоровый вес, чтобы уменьшить нагрузку на мышцы.
- Избегайте обезвоживания, пейте много воды или воды.
- Разомнитесь перед тренировкой, постепенно увеличивайте интенсивность, а затем выполняйте растяжку.
- Носите подходящую обувь и все необходимое защитное снаряжение для любого вида спорта, которым вы занимаетесь.
Часто задаваемые вопросы
Когда мне следует позвонить поставщику медицинских услуг по поводу проблем с мышцами бедра?
Повреждение мышцы бедра может повысить вероятность того, что вы еще больше поранитесь.Поговорите с поставщиком медицинских услуг, если у вас есть:
- Неспособность согнуть или разогнуть бедро или колено.
- Потеря чувствительности или постоянная слабость в бедре или тазобедренном суставе.
- Боль, которая не проходит или усиливается при сгибании или разгибании бедра или колена.
- Хлопающий или щелкающий шум в ноге.
- Повторные травмы.
- Отек или синяк.
- Проблемы с опорой на ногу.
Записка из клиники Кливленда
Бедра содержат множество мышц, которые контролируют движения бедер и ног.Основными типами являются приводящие, подколенные сухожилия, гребенчатые мышцы, четырехглавые мышцы и портняжные мышцы. Часто встречаются травмы мышц бедра, особенно растяжения, растяжения и разрывы у спортсменов. Позвоните своему поставщику медицинских услуг, если у вас есть боль, онемение или слабость или какие-либо проблемы с переносом веса на ногу.
.