
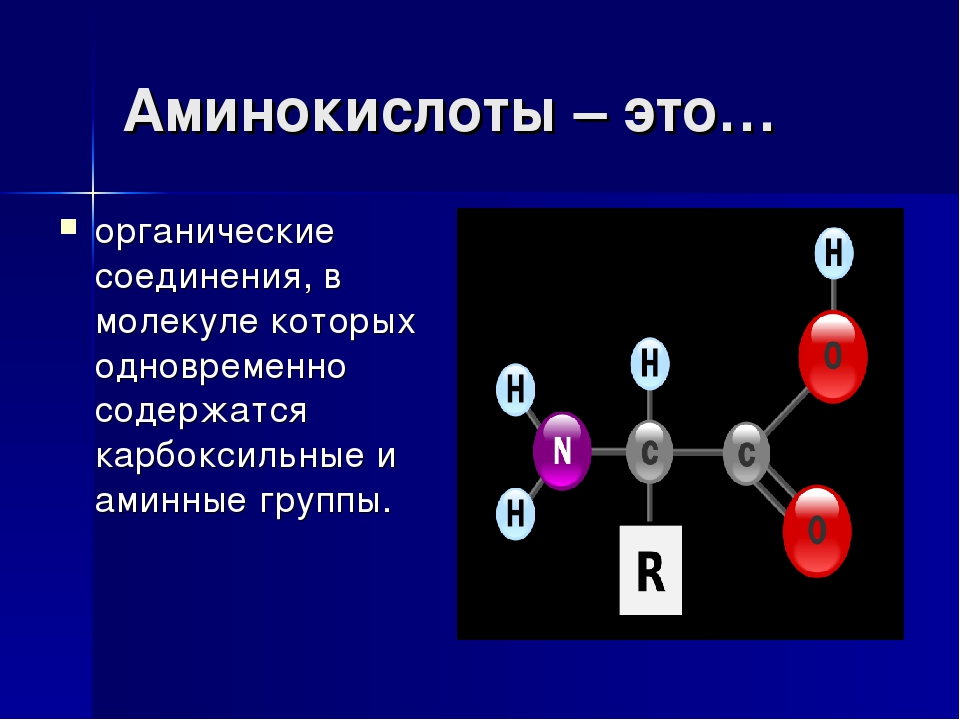
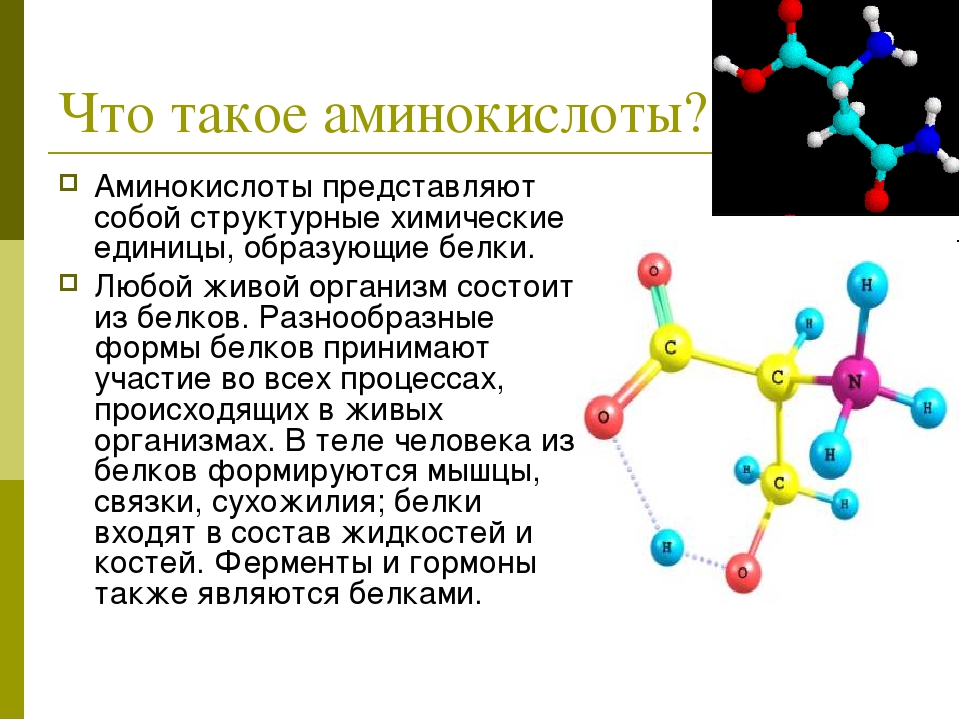
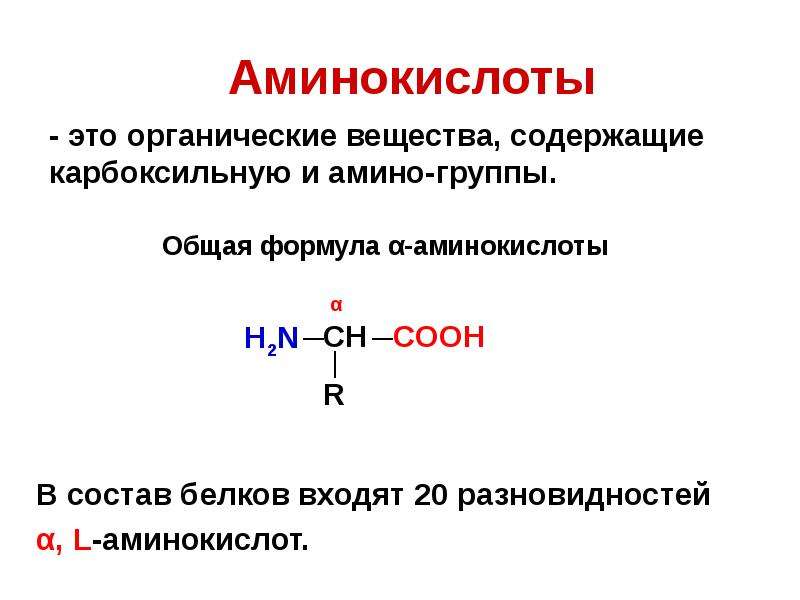
Аминокислоты I Что это? Для чего? Как принимать?
Как принимать аминокислоты?
Аминоксилоты принимают как при наборе массы, так и при «сушке». Если вы пьете протеин, то в дополнительной подпитке нет необходимости.
- Дозировка аминокислот должна определяться вашим весом и рекомендациями на упаковке. Это обусловлено тем, что в продуктах может содержаться различный процент «чистых» аминокислот. В основном принимают от 10 до 20 г в сутки.
- Дозу аминокислот лучше делить на несколько приемов. Их можно пить с утра, во время тренировки и вечером. Это не постулат, поэтому каждый, исходя из целей тренинга, может сам определить для себя время приема.
- Аминокислоты выпускаются в разных формах. Усвояемость каждой из форм индивидуальна, поэтому рекомендую попробовать разные варианты. Для кого-то более эффективными оказываются порошковые формы, кому-то больше подходят капсулы, а кто-то рад только жидким аминокислотам.
Какие аминокислоты выбрать?
Существуют комплексные и изолированные типы аминокислот.
Заключение
Употребление аминокислот позволяет значительно улучшить тренировочный процесс, насытить органы и мышцы питательными веществами и сократить периоды восстановления. При этом необходимо придерживаться правильного питания, ведь аминокислоты не являются полноценной заменой пище. Кроме того, эта добавка безопасна для употребления и не вызывает привыкания.
Исходя из ваших целей, вы можете приобрести определенную аминокислоту, которая поможет вам достичь их. Например, аргинин атлеты выбирают с целью пампинга во время тренировки, ВСАА — для восстановления после интенсивных нагрузок, а глютамин активно используется при наборе мышечной массы.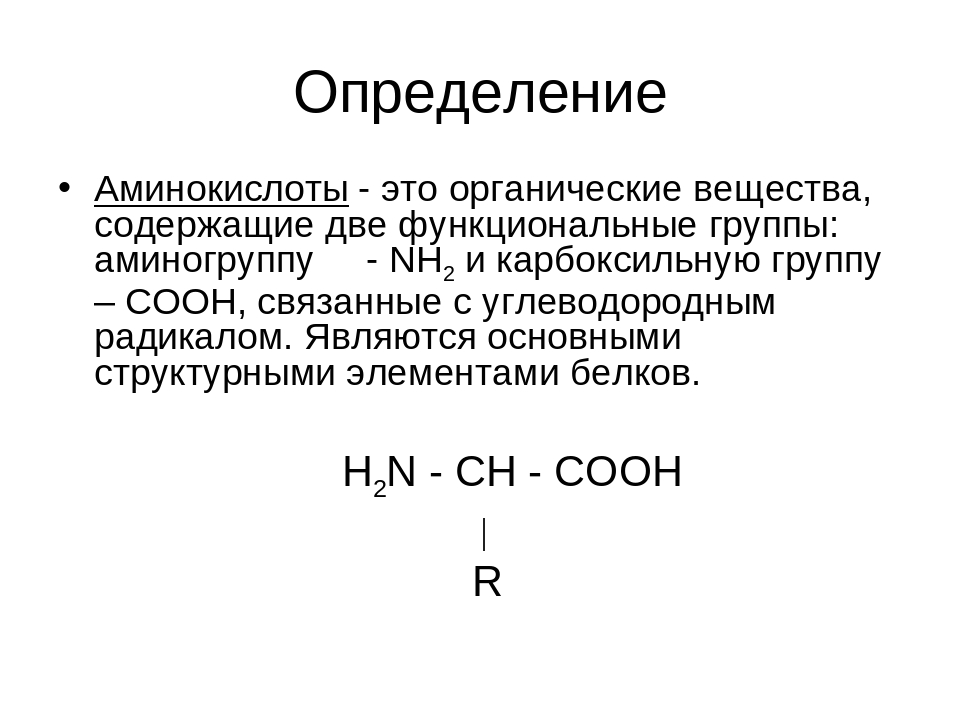
Будьте здоровы! Выбирайте проверенные продукты!
описание, свойства, способ применения, суточная норма и действие лизина
L-Лизин (L-Lysine)
Полезные свойства:
-
Формирование карнитина, отвечающего за состояние волос и кожи;
-
Синтез коллагена, помогающего избежать морщин;
-
Снижение уровня холестерина;
-
Улучшение усвоения кальция.
Лизин – незаменимая аминокислота, которая не может быть синтезирована организмом и поступает в организм только с пищей и добавками. Лизин обладает широким спектром биологических эффектов и прежде всего лизин жизненно необходим как составляющая белков организма.
Некоторые исследования показали то, что l-lysine может сократить частоту проявления вируса герпеса.
Способ воздействия вещества на вирус герпеса неизвестен, но есть версия о том, что лизин влияет на другую аминокислоту — аргинин. В свою очередь, было установлено влияние аргинина на ускорение деления клеток зараженных вирусом герпеса. Чем меньше аргинина, тем медленнее развивается герпес, а l-лизин снижает активность аргинина.
В 2007 году, учетые из Университета Флориды (FSU) установили эффект «конъюгатов лизина» (lysine conjugates), позволяющий идентифицировать отдельные повреждения ДНК. Исследованием занимался профессор биохимии FSU Игорь Алабугин. Для лечения рака было предложено использовать конъюгаты лизина под воздействием фототерапии. Такое комплексное воздействие позволяет уничтожить 90%+ больных клеток, при этом не повреждая здоровые. Изобретение профессора Алабугина уже запатентовано и проходит испытания.
Использование лизина для лечения рака было протестировано на колоректальном раке у мышей в 2014 году. В ходе исследования 0 животных погибло, в том числе животных находившихся на поздних стадиях заболевания.
Исследования показали эффективное влияние л-лизина даже на такие виды заболевания как лейкимия, что вселяет дополнительную надежду на скорое избавление человечества от такой страшной болезни.
Вместе с употреблением пищи богатой витаминами группы Б, магнием и омегой-3, можно принимать л-лизин для лечения психологических проблем.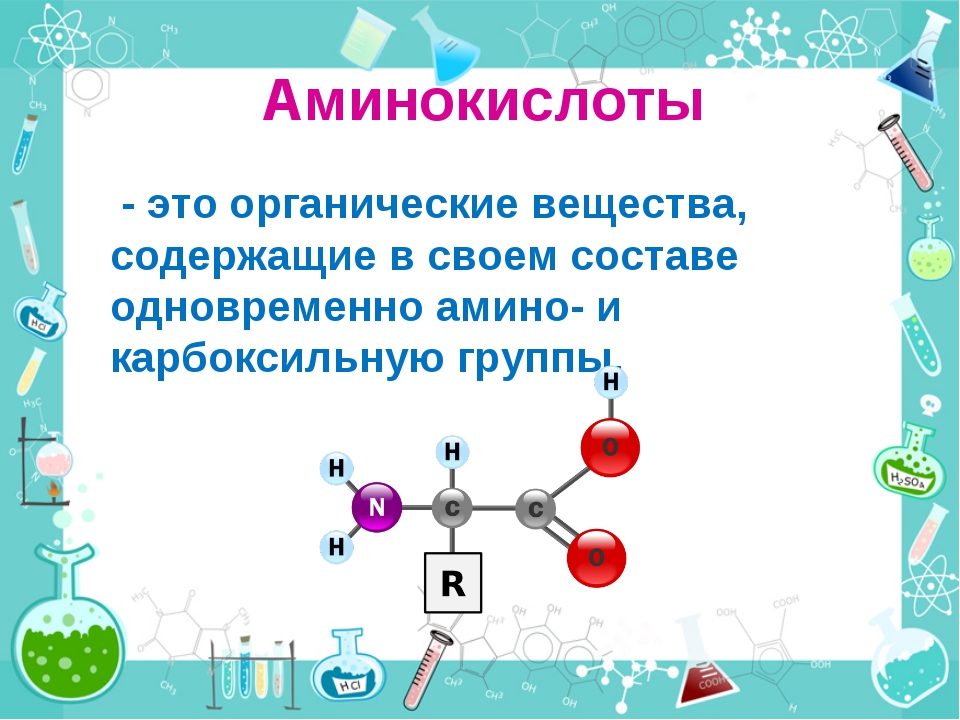 Лизин улучшает усвоение кальция и других веществ, тем самым восстанавливая нормальное состояние организма. Данная аминокислота используется для снятия синдрома беспокойства и может безвредно для организма снимать депрессивное состояние.
Лизин улучшает усвоение кальция и других веществ, тем самым восстанавливая нормальное состояние организма. Данная аминокислота используется для снятия синдрома беспокойства и может безвредно для организма снимать депрессивное состояние.
L-лизин ведет себя как антагонист рецептора серотонина. Он связывается с рецепторами и частично предотвращает ощущение тревоги. Более того, вещество влияет на вызванные стрессом реакции, включая лечение диареи.
Употребление л-лизина связывают с улучшением усвоения кальция, что крайне важно для людей с риском развития остеопороза. На сегодняшний день не существует исследований, которые смогли бы подтвердить связь между лечением остеопороза и употреблением лизина, однако важность влияния кальция на состояние костей дает основание предполагать, что лизин может быть полезен.
Кальций влияет не только на ваши кости, но и воздействует на ваш вес, защищает от серьёзных заболеваний, снижает симптомы ПМС, улучшает состояние зубов, влияет на работу нервной системы и мышц, предотвращает диабет.
Большая часть жителей нашей планеты испытывает проблемы, связанные со здоровьем кишечника. Многие люди даже не знают об этих проблемах, но они оказывают существенное влияние на их здоровье. Речь идет не только о прямых проблемах, таких как кислотный рефлюкс или диарея, а о многих других. Например, нарушение микрофлоры кишечника может влиять на состояние кожи, волос и костей, из-за того, что необходимые вещества не усваиваются организмом.
Одна из форм л-лизина, известная как поли-л-лизин оказывает противовоспалительный эффект и избавляет от многих проблем кишечника.
Американские ученые далеко продвинулись, экспериментируя с лизином, их опыты доказали, что применение лизина способно остановить выпадение волос при андрогенной алопеции и железодефицитной анемии. Существует американский патент на применение Лизина в лечении различных проблем, связанных с выпадением волос, включая облысение. Лизин блокирует 5-альфаредуктазу. Дефицит лизина неблагоприятно сказывается на синтезе белка, что приводит к утомляемости, усталости и слабости, плохому аппетиту.
Дефицит лизина неблагоприятно сказывается на синтезе белка, что приводит к утомляемости, усталости и слабости, плохому аппетиту.
Лизин широко используется в бодибилдинге, добавляется в спортивное питание и БАДы. Эта аминокислота позволяет ускорить восстановление мышц после силового тренинга. Модулирующее влияние лизина поддерживает положительный азотистый обмен и построение мышечных белков.
Защита и питание мышц — это две основные функции лизина в спорте. Кроме того лизин укрепляет сухожильный корсет и костную систему, что снижает риск травмирования, а также ускоряет восстановление.
В среднем человеку необходимо получать от 800 до 3000 мг l-лизина ежедневно. Дозировка при лечении герпеса может увеличиваться от 1 до 3 гр вещества.
Лизин может быть использован в виде крема, который ускоряет снятие симптомов герпеса.
Лизин в рекомендуемых дозах абсолютно безопасен и не вызывает никаких побочных эффектов.
Литература
1. Рональд Клатц, Роберт Голдман «Эра молодости (Anti-aging революция), изд.Москва, Санкт-Петербург, «Ост»,2007 г.
2. Патент США № 5678617.
Чем нас лечат: Глицин — Индикатор
В ответ сайт производителей приводит такие аргументы:
- Не проходит через гематоэнцефалический барьер? Ну и что, от препарата «нейроны мозга увеличивают собственный синтез» (чего, как, где исследования — информации нет), а еще в кишечнике тоже есть нервные клетки.
- Нет выраженного результата при приеме? А его и не должно быть: Глицин — это «стимулятор обмена» и «витамин для мозга».
- Не препарат, а БАД? Неправда, Глицин зарегистрирован как лекарство: «без подтвержденной эффективности данные о препарате не вносят в инструкцию», а значит, он прошел клинические испытания (публикаций результатов этих испытаний не приводится).

Мало того, что эти заявления противоречат друг другу, они еще и не подкреплены ссылками на клинические испытания. К тому же, как знают читатели нашей рубрики, регистрация в списках — хоть регистре лекарственных средств, хоть российском списке жизненно необходимых и важнейших лекарственных средств (который, кстати, был создан не для того, чтобы подтвердить, что лекарства работают, а чтобы ограничить их стоимость в продаже) — еще не гарантия эффективности. К тому же это в России Глицин — лекарство, в США, к примеру, он всего лишь БАД. А к билогически активным добавкам требования для регистрации гораздо ниже, да еще и спрос с них не так строг. А на сайте Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (Food and Drug Administration, или FDA) США он значится как основа для применяющегося в урологических операциях раствора.
Сколько нужно глицина?
Забавно и такое уточнение: «Неправильное мнение о Глицине возникло из-за применения многочисленных БАДов, они содержат аминокислоту глицин и витамины. Разница между ними и медикаментами состоит в том, что для биодобавок не предусмотрены клинические исследования, а значит, эффективность их применения не имеет доказательств». Выходит, что Глицин отдельно — это лекарство, а с витаминами — уже добавка? Или, может, все дело в количестве?
Разница между ними и медикаментами состоит в том, что для биодобавок не предусмотрены клинические исследования, а значит, эффективность их применения не имеет доказательств». Выходит, что Глицин отдельно — это лекарство, а с витаминами — уже добавка? Или, может, все дело в количестве?
Среднестатистическая диета включает около двух граммов глицина. Однако человеческий организм может синтезировать его и самостоятельно, используя другую аминокислоту, серин. Последнюю, кстати, тоже незаменимой не назовешь, так как и ее мы производим внутри себя. Но есть и данные в пользу того, что количество, в котором наш организм синтезирует глицин (три грамма в день) и в котором мы получаем его в с пищей (1,5-3 г), — это две трети от общей потребности. Учитывая «расходы» на синтез коллагена, где глицина используется очень много, человеку массой 70 кг нужно до десяти граммов глицина в день.
Допустим, от этого мы убедимся, что стоило бы есть побольше глицина. Но в таблетке Глицин содержится лишь 100 мг (0,1 г) одноименной аминокислоты. Сильно ли одна таблетка меняет ситуацию — казалось бы, вопрос риторический. На самом деле, при инсульте дневная дозировка составляет один-два грамма, так что это действительно может иметь смысл. Но есть ли от глицина какой-то значимый эффект на организм?
Сильно ли одна таблетка меняет ситуацию — казалось бы, вопрос риторический. На самом деле, при инсульте дневная дозировка составляет один-два грамма, так что это действительно может иметь смысл. Но есть ли от глицина какой-то значимый эффект на организм?
Пудра для полости рта и борьба с алкоголизмом
Ответить на этот вопрос помогут испытания на больших выборках пациентов. Правда, хоть упоминаний глицина в PubMed тысячи, среди них практически нет его клинических испытаний в качестве лекарства. Результаты таких работ разрозненные и чаще всего предварительные, дизайн их несовершенен. Ниже мы обсудим двойные слепые контролируемые исследования, которых среди тысяч экспериментов оказалось не так уж много.
Протеины или аминокислоты — советы от Sportfood
Аминокислоты – это комплексы аминокислот. В таком комплексе имеются все виды аминокислот. Протеин – это белок. А белок – это аминокислоты. Итак, протеин и аминокислоты достаточно схожи друг с другом.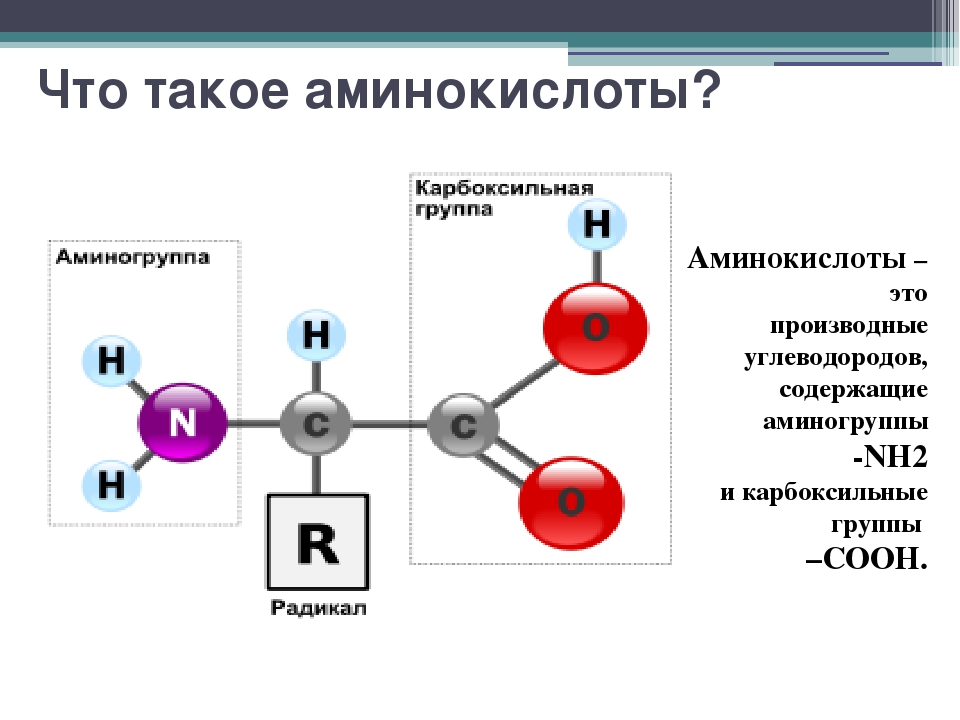 Но все же присутствуют некоторые отличия. Уточним плюсы применения аминокислот и протеинов.
Но все же присутствуют некоторые отличия. Уточним плюсы применения аминокислот и протеинов.
Аминокислоты – это важные составляющие части белка. Комплексы аминокислот нужны для скорого восстановления и роста мышечной ткани. Обычная пища не может удовлетворить потребность организма во время занятий спортом всеми необходимыми аминокислотами. Каждая из существующих аминокислот нужна организму и выполняет свою определенную функцию. Аминокислотные комплексы содержат в составе все аминокислоты. В таких комплексах содержится очень большое количество незаменимых аминокислот, которые в нашем организме не производятся.
Принимать аминокислоты нужно всегда. Независимо от того тренируется спортсмен или временно прекратил свои занятия. Потому что во время тренировок спортсмен наращивает свою мышечную массу, а во время прекращения тренировок аминокислоты не дают разрушаться мышцам и поддерживают их в норме.
Принимать аминокислотные комплексы нужно в соответствии с массой тела и делить прием на 2 раза в день.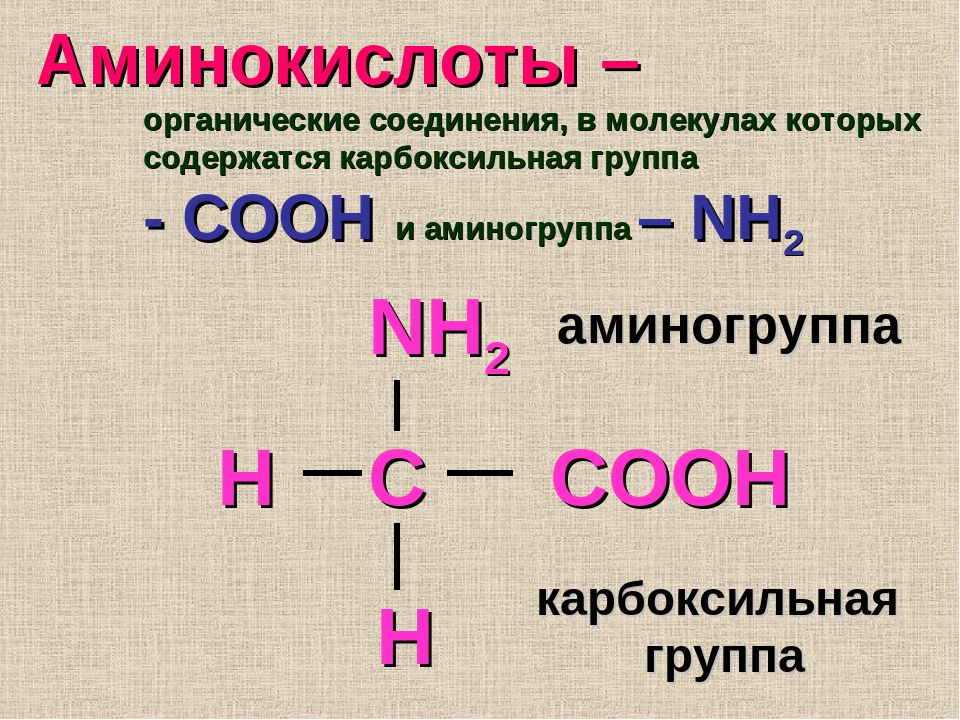
Выпускаются препараты аминокислотных комплексов в форме порошка, капсул или таблеток и в жидкой форме. Жидкая форма содержит около 50% аминокислот.
- Низкая стоимость протеинов. Разница в стоимости протеинов и аминокислотных комплексов довольно ощутима. Протеины не нуждаются в дополнительной переработке. А кроме этого его не нужно изготавливать в форме таблеток или рассыпать по капсулам.
- Достаточно долгий процесс расщепления протеинов. Если применить казеин, то этот протеин будет расщепляться в течение 3-4 часов. В это время организм будет постоянно получать аминокислоты, которые помогут набрать мышечную массу или восстановить ее.
- Протеиновые препараты подходят как для набора мышечной массы, так и для похудения. Протеины лучше и быстрее насыщают. Во время диеты протеины способны заменить часть пищевых продуктов и не дать мышечной массе уменьшиться. Происходит увеличение количества потребляемого белка, а углеводы и жиры, наоборот, уменьшаются. При применении аминокислот такого насыщения не произойдет.
- Удобство в применении. В случае необходимости употребления аминокислотных комплексов не требуется использование специальной посуды (шейкера) и молочных продуктов. Можно взять определенное количество таблеток и запить их водой.
- Комплексы аминокислот не нуждаются в процессе переваривания. Скорость усвоения аминокислот намного быстрее, чем протеинов. Это происходит из-за необходимости изначального расщепления протеинов до получения аминокислоты. Процесс расщепления занимает чуть больше часа.
- Аминокислоты практически не содержат углеводов и жира. Большое количество комплексов аминокислот не содержат в своем составе жир и углеводы. Из этого следует, что такие комплексы являются более подходящими для тех, кто стремиться похудеть. Но есть и протеины, не содержащие большого количества углеводов и жиров.
Специалисты в этой сфере утверждают, что не будет никакого эффекта при одновременном использовании аминокислотных комплексов и протеинов. Рекомендуется выбрать что-то одно.
Если цель спортсмена состоит в быстром наборе мышечной массы, то лучшим вариантом будет применение протеинов. Не стоит забывать о том, что применение протеина обойдется примерно в 1,5 раза дешевле, по сравнению с аминокислотными комплексами.
Следует обращать внимание на состав основы комплекса. От этого зависит качество приобретаемого товара. Если в составе выбранного комплекса аминокислот содержится концентрат сывороточного протеина, то это простой, спрессованный в таблетки, протеин.
Натуральные аминокислотные комплексы обязательно произведены из гидролизованных белков. Такие комплексы очень редко встречаются в продаже и стоят намного дороже всех остальных аминокислот. При выборе жидких аминокислотных комплексов также следует быть очень внимательными. В большинстве из них в составе находится гидролизат желатина. А этот компонент в основном используется для восстановления травмированных связок, а не для быстрого набора мышечной массы.
Из всего этого следует, что протеины использовать лучше, чем комплексы аминокислот.
Различные характеристики экскреции аминокислот с мочой у людей и использование добавок аминокислот для снижения утомляемости и нарушений здоровья у взрослых | Nutrition Journal
Boirie Y. Физиопатологический механизм саркопении. J Nutr Здоровье Старение. 2009; 13: 717–23.
КАС Статья пабмед Google ученый
Jagoe RT, Engelen MPKJ. Атрофия мышц и изменения метаболизма мышечных белков при хронической обструктивной болезни легких.Eur Respir J. 2003; 22:52–63с.
КАС Статья Google ученый
Oehler R, Roth E. Метаболизм глютамина. В: Cynober LA, редактор. Метаболические и терапевтические аспекты аминокислот в лечебном питании. Второе изд. Нью-Йорк: CRC Press; 2004. с. 169–82.
Google ученый
 «>
«>Уденфренд С., Вингаарден Дж.Б. Предшественники надпочечникового адреналина и норадреналина in vivo.Биохим Биофиз Акта. 1956; 20: 48–52.
КАС Статья пабмед Google ученый
de Koning TJ, Snell K, Duran M, Berger R, Poll-The BT, Surtees R. L-серин при заболевании и развитии. Биохим Дж. 2003; 371:653–61.
Артикул пабмед ПабМед Центральный Google ученый
Poortmans JR, Carpentier LO, Pereira-Lancha LO, Lancha Jr A. Белковый обмен, потребности в аминокислотах и рекомендации для спортсменов и активного населения.Braz J Med Biol Res. 2012;45:875–90.
КАС Статья пабмед ПабМед Центральный Google ученый
Филипс СМ. Потребность в белке и добавки в силовых видах спорта. Питание. 2004; 20: 689–95.
КАС Статья пабмед Google ученый
 «>
«>Waterlow JC, Jackson AA. Питание и белковый обмен у человека. Бр Мед Булл. 1981; 37: 5–10.
КАС Статья пабмед Google ученый
Corsetti R, Barassi A, Perego S, Sansoni V, Rossi A, Damele CAL, d’Eril GM, Banfi G, Lombardi G. Изменения экскреции аминокислот с мочой в связи с маркерами мышечной активности на этапе профессионального велоспорта гонка: в поисках маркеров усталости. Аминокислоты. 2016; 48:183–92.
КАС Статья пабмед Google ученый
Данстан Р.Х., Спаркс Д.Л., Даскомб Б.Дж., Эванс К.А., Макдональд М.М., Кромптон М.Дж., Фрэнкс Дж., Мерфи Г., Готфрис Дж., Карлтон Б.Пот способствовал потере аминокислот у лошадей стандартной породы и применению стратегий добавок для поддержания формы во время тренировок. Комп Упражнение Физиол. 2015;11:201–212.
Артикул Google ученый
 «>
«>Данстан Р. Х., Спаркс Д. Л., Даскомб Б. Дж., Макдональд М. М., Эванс К. А., Стивенс К. Дж., Кромптон М. Дж., Готфрис Дж., Фрэнкс Дж., Мерфи Г. и др. Пот способствовал потерям аминокислот у спортсменов-мужчин во время тренировки при температуре 32-34 °C.ПЛОС Один. 2016; 11:1–16.
Артикул Google ученый
Данстан Р.Х. Измененный гомеостаз аминокислот и развитие усталости у пациентов с лучевой терапией рака молочной железы: пилотное исследование. Клин Биохим. 2011;44:208–15.
Ниблетт С.Н. Гематологические и мочевыделительные аномалии у больных с синдромом хронической усталости. Экспер Биол Мед. 2007; 232:1041–9.
КАС Статья Google ученый
Liappis N, Hungerland H.Количественное исследование свободных аминокислот в эккринном поте человека в нормальных условиях. Am J Clin Nutr. 1972; 25: 661–3.
1972; 25: 661–3.
КАС пабмед Google ученый
Armstrong MD, Stave U. Исследование уровней свободных аминокислот в плазме. II. Нормальные значения для детей и взрослых. Метаболизм. 1973; 22: 561–9.
КАС Статья пабмед Google ученый
Вешлер ЛБ.Концентрации электролитов пота, полученные внутри окклюзионных покрытий, ложно завышены, поскольку пот сам выщелачивает кожные электролиты. J Appl Physiol. 2008; 105:1376–7.
Артикул пабмед Google ученый
Скотт И.Р., Хардинг Ч.Р., Барретт Дж.Г. Богатый гистидином белок кератогиалиновых гранул. Источник свободных аминокислот, урокановой кислоты и пирролидонкарбоновой кислоты в роговом слое. Биохим Биофиз Акта.1982; 719: 110–7.
КАС Статья пабмед Google ученый
 «>
«>Kingsbury KJ, Kay L, Hjelm M. Сравнение свободных аминокислот в плазме у элитных спортсменов: связь с усталостью и инфекцией. Бр Дж Спорт Мед. 1998; 32:25–32.
КАС Статья пабмед ПабМед Центральный Google ученый
Paddon-Jones D. Атрофия и нарушение синтеза мышечного белка при длительном бездействии и стрессе.J Clin Метабол эндокринолов. 2006;91:4836–41.
КАС Статья Google ученый
Данстан Р.Х., Спаркс Д.Л., Робертс Т.К., Кромптон М.Дж., Готфрис Дж., Даскомб Б.Дж. Разработка комплексной аминокислотной добавки, усталость reviva™, для перорального приема: первоначальная оценка концепции продукта и влияние на симптомы недомогания в группе мужчин. Нутр Дж. 2013;12:115.
КАС Статья пабмед ПабМед Центральный Google ученый
 «>
«>Данстан Р.Х., Спаркс Д.Л., Робертс Т.К., Даскомб Б.Дж.Предварительные оценки комплексной аминокислотной добавки Reviva TM для снижения усталости у группы профессиональных спортсменов-мужчин и группы мужчин, набранных из числа населения. Food Nutr Sci. 2014;5:231–5.
Артикул Google ученый
Dunstan RH, McGregor NR, Butt HL, Roberts TK, Klineberg IJ, Niblett SN, Rothkirch TB, Buttfield I. Характеристика дифференциального гомеостаза аминокислот среди подгрупп населения: основа для определения конкретных потребностей в аминокислотах.J Nutr Environ Med. 2000; 10: 211–23.
КАС Статья Google ученый
Ватанабе Н., Стюарт Р., Дженкинс Р., Бхугра Д.К., Фурукава Т.А. Эпидемиология хронической усталости, соматических заболеваний и симптомов распространенных психических расстройств: поперечное исследование второго британского национального исследования психиатрической заболеваемости. Дж. Психосом Рез. 2008; 64: 357–62.
Дж. Психосом Рез. 2008; 64: 357–62.
Артикул пабмед Google ученый
Li G, Xie F, Yan S, Hu X, Jin B, Wang J, Wu J, Yin D, Xie Q: Субздоровье: определение, критерии диагностики и потенциальная распространенность в центральном регионе Китая.Общественное здравоохранение BMC. 2013;13.
Эванс С., Дунстан Р.Х., Роткирх Т., Робертс Т.К., Райхельт К.Л., Косфорд Р., Дид Г., Эллис Э.Б., Спаркс Д.Л. Изменение экскреции аминокислот у детей с аутизмом. Нутр Невроски. 2008; 11: 9–17.
КАС Статья пабмед Google ученый
Чалдер Т., Береловиц Г., Павликовска Т., Уоттс Л., Вессели С., Райт Д., Уоллес Э.П. Разработка шкалы усталости. Дж. Психосом Рез. 1993; 37: 147–53.
КАС Статья пабмед Google ученый
 «>
«>МакГрегор Н.Р., Дунстан Р.Х., Зербес М., Батт Х.Л., Робертс Т.К., Клинеберг И.Дж. Предварительное определение связи между выраженностью симптомов и метаболитами в моче у субъектов с синдромом хронической усталости. Биохим Мол Мед. 1996; 58: 85–92.
КАС Статья пабмед Google ученый
Данстан Р.Х., МакГрегор Н.Р., Батт Х.Л., Робертс Т.К.Биохимические и микробиологические аномалии при синдроме хронической усталости: разработка лабораторных тестов и возможная роль токсичных химических веществ. J Nutr Environ Med. 1999; 9: 97–108.
КАС Статья Google ученый
Ричардс Р.С., Робертс Т.К., МакГрегор Н.Р., Данстан Р.Х., Батт Х.Л. Параметры крови, указывающие на окислительный стресс, связаны с выраженностью симптомов при синдроме хронической усталости. Redox Rep. 2000; 5:35–41.
КАС Статья пабмед Google ученый
 «>
«>Кодексы пищевых стандартов Австралии и Новой Зеландии [http://www.comlaw.gov.au/Search/Australia%20New%20Zealand%20Food%20Standards]. По состоянию на 13 июня 2013 г.
Закон о пищевых продуктах 2003 г. № 43 [http://www.legislation.nsw.gov.au/viewtop/inforce/act+43+2003+FIRST+0+N/]. По состоянию на 13 июня 2013 г.
Регламент о пищевых продуктах 2010 г. [http://www.legislation.nsw.gov.au/viewtop/inforce/subordleg+250+2010+cd+0+N].По состоянию на 13 июня 2013 г.
Клеменс Р.А., Коппле Д.Д., Свендсайд М.Э. Метаболические эффекты диеты с дефицитом гистидина, которую кормили растущим крысам через желудочный зонд. Дж Нутр. 1984; 114: 2138–46.
КАС пабмед Google ученый
Cooperman JM, Lopez R. Роль гистидина в анемии дефицита фолиевой кислоты. Экспер Биол Мед. 2002; 227: 998–1000.
КАС Google ученый
 «>
«>Виру А., Виру А.Биохимический мониторинг спортивной тренировки. 1-е изд. Шампейн: Издательство Human Kinetics; 2001.
Google ученый
Haralambie G, Berg A. Изменения мочевины и аминного азота в сыворотке крови в зависимости от продолжительности упражнений. Eur J Phys Occup Physiol. 1976; 36: 39–48.
КАС Статья Google ученый
Хеджес Р.Э.М., Клемент Дж.Г., Томас Д.Л., О’Коннелл Т.К. Обмен коллагена в средней части диафиза бедренной кости взрослого человека: смоделировано на основе измерений антропогенных радиоуглеродных индикаторов.Am J Phys Антропол. 2007; 133:808–16.
Артикул пабмед Google ученый
Wang W, Wu Z, Dai Z, Yang Y, Wang J, Wu G. Метаболизм глицина у животных и человека: значение для питания и здоровья. Аминокислоты. 2013;45:463–77.
2013;45:463–77.
Артикул пабмед Google ученый
Кори Дж.Г. Метаболизм пуриновых и пиримидиновых нуклеотидов. В: Девлин ТМ, редактор.Учебник биохимии с клиническими корреляциями. 3-е изд. Нью-Йорк: Уайли-Лисс; 1992. с. 529–73.
Google ученый
Fessas PH, Koniavitis A, Zeis A. Экскреция бета-аминоизомасляной кислоты с мочой при талассемии. Джей Клин Патол. 1969; 22: 154–157.
КАС Статья пабмед ПабМед Центральный Google ученый
Энхжаргал Т., Цереннадмид С. Экскреция бета-аминоизомасляной кислоты с мочой при гематологических заболеваниях (аннотация).Риншо Бёри Jpn J Clin Pathol. 2004; 52:17–21.
КАС Google ученый
Мацумото А.М. Андропауза: клинические последствия снижения уровня тестостерона в сыворотке крови с возрастом у мужчин.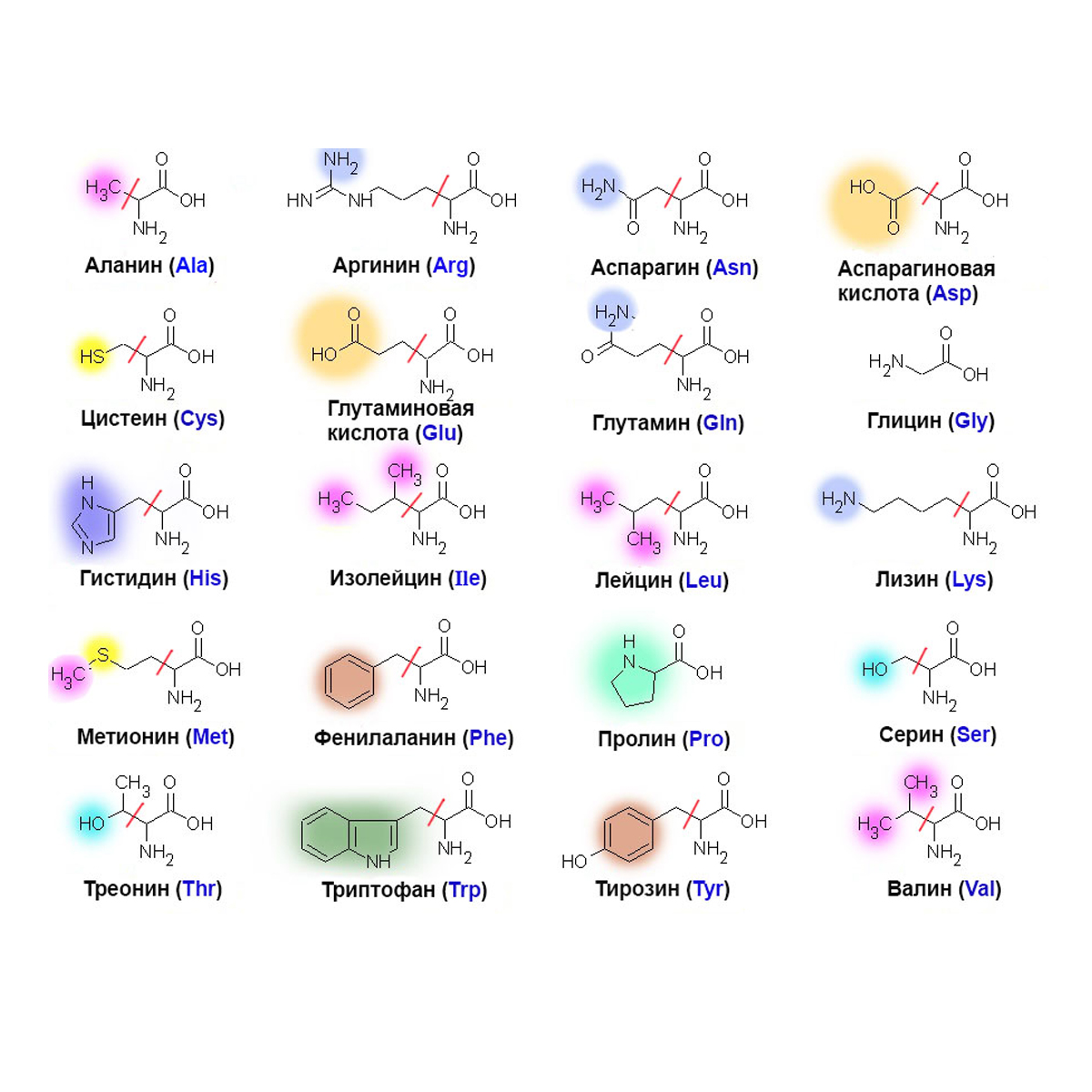 Дж Геронтол. 2002; 57А:М76–99.
Дж Геронтол. 2002; 57А:М76–99.
КАС Статья Google ученый
Новак А., Брод М., Элберс Дж. Андропауза и качество жизни: результаты фокус-групп пациентов и клинических экспертов.Зрелые. 2002;43:231-7.
КАС Статья пабмед Google ученый
Bain J. Заместительная терапия тестостероном для стареющих мужчин. Кан Фам Врач. 2001;47:91–7.
КАС пабмед ПабМед Центральный Google ученый
Nogueira Ade C, Vale RG, Gomes AL, Dantas EH. Влияние мышечных действий на уровень повреждения соединительной ткани. Рес Спорт Мед.2011;19:259–70.
ПабМед Google ученый
Prockop DJ, Sioerdsma D. Значение гидроксипролина в моче у человека. Клин Инвест. 1961; 40: 843–9.
КАС Статья Google ученый
Кьер М., Хансен М. Тайна женской соединительной ткани. J Appl Physiol. 2008; 105:1026–7.
Артикул пабмед Google ученый
Джейсон Л.А., Ричман Дж.А., Радемейкер А.В., Джордан К.М., Плиоплис А.В., Тейлор Р.Р., Маккриди В., Хуанг С-Ф, Плиоплис С.Общественное исследование синдрома хронической усталости. Arch Intern Med. 1999; 159:2129–37.
КАС Статья пабмед Google ученый
Вессели С. Эпидемиология синдрома хронической усталости. Эпидемиологическое издание 1995; 17:139–51.
КАС Статья Google ученый
Boudes P. Соблюдение режима лечения в терапевтических исследованиях: обзор. Контрольные клинические испытания.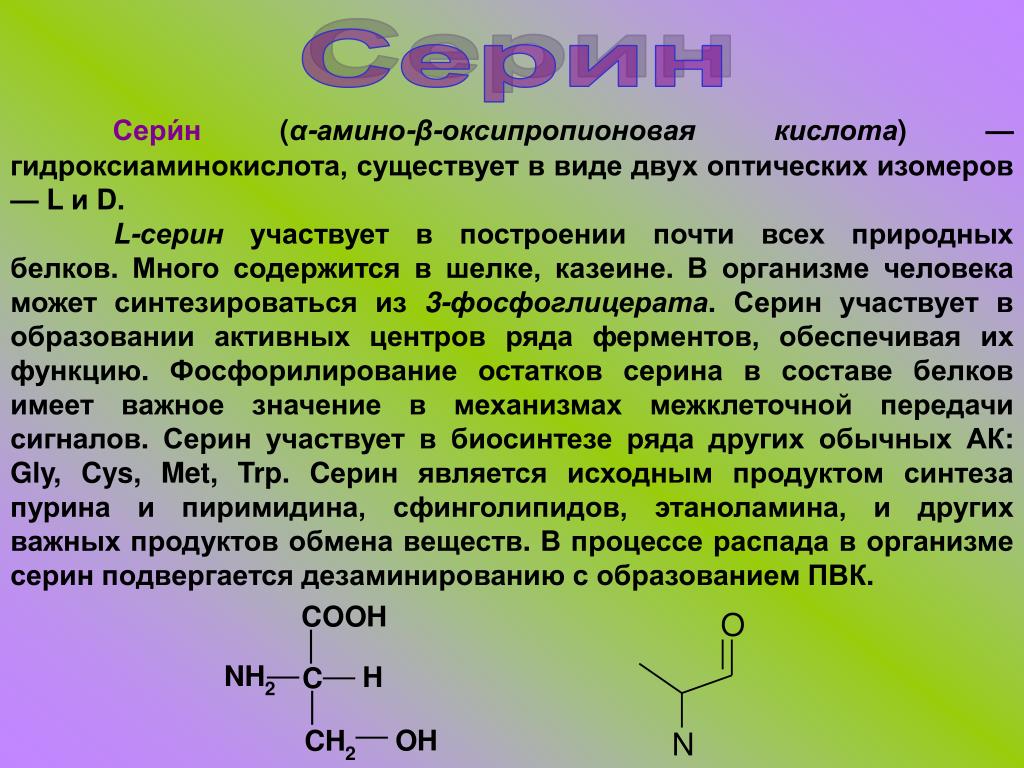 1998; 19: 257–68.
1998; 19: 257–68.
КАС Статья пабмед Google ученый
Селла М., Чалдер Т. Измерение усталости в клинических и общественных условиях. Дж. Психосом Рез. 2010;69:17–22.
Артикул пабмед Google ученый
Границы | От редакции: Аминокислоты в растениях: регуляция и функции в развитии и защите от стресса
В последние десятилетия важность аминокислот в развитии растений и защите от стресса становится все более очевидной, что вызывает растущий интерес в фундаментальной и прикладной науке о растениях.Здесь мы представляем новые результаты исследований аминокислот и предлагаем максимально актуальную картину современных знаний об этом увлекательном аспекте физиологии растений. Оказалось, что помимо того, что они являются строительными блоками для синтеза белка, многие аминокислоты, в том числе не участвующие в синтезе белка, играют активную роль в развитии растений и участвуют в реакции растений на стрессы окружающей среды.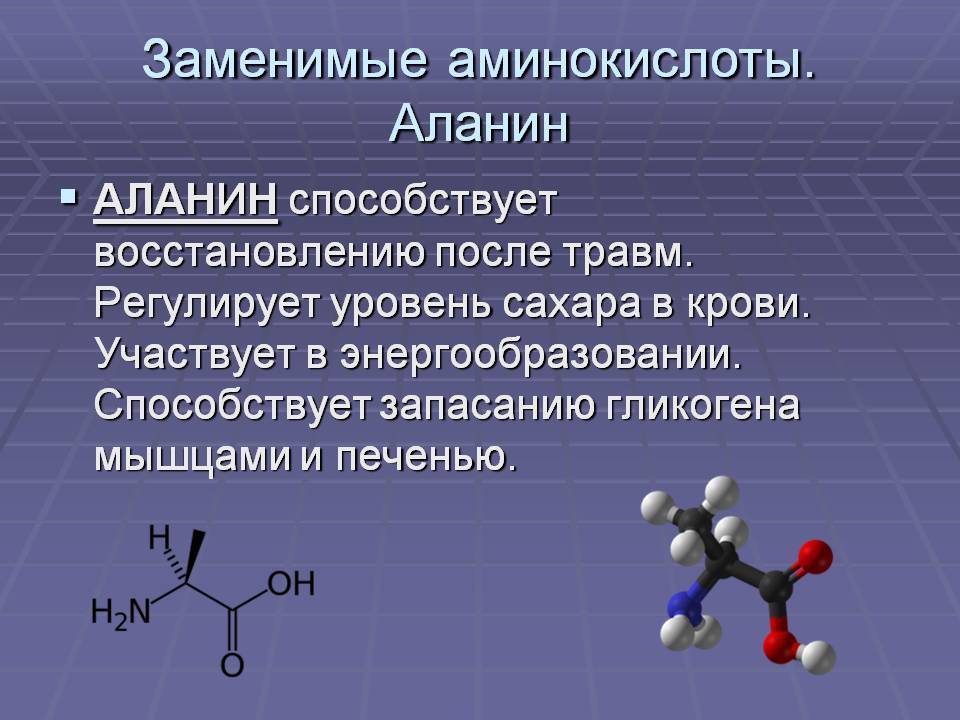 Кроме того, аминокислоты служат предшественниками многих первичных и вторичных метаболитов и играют ключевую роль в питании человека либо в качестве источника нутрицевтических соединений, либо в качестве основных пищевых компонентов.Действительно, девять из двадцати одной протеиногенной аминокислоты не могут быть синтезированы у животных, включая человека, а три или более других не синтезируются в количествах, достаточных для удовлетворения метаболических потребностей (рис. 1). Эти питательно незаменимые аминокислоты должны поступать из рациона, и, безусловно, наибольшая их доля поступает из растений (Galili et al., 2016, Hou and Wu, 2018). В отличие от человека и животных, растения сами синтезируют всю двадцать одну протеиногенную аминокислоту (рис. 1).
Кроме того, аминокислоты служат предшественниками многих первичных и вторичных метаболитов и играют ключевую роль в питании человека либо в качестве источника нутрицевтических соединений, либо в качестве основных пищевых компонентов.Действительно, девять из двадцати одной протеиногенной аминокислоты не могут быть синтезированы у животных, включая человека, а три или более других не синтезируются в количествах, достаточных для удовлетворения метаболических потребностей (рис. 1). Эти питательно незаменимые аминокислоты должны поступать из рациона, и, безусловно, наибольшая их доля поступает из растений (Galili et al., 2016, Hou and Wu, 2018). В отличие от человека и животных, растения сами синтезируют всю двадцать одну протеиногенную аминокислоту (рис. 1).
Рисунок 1 . Биосинтез аминокислот в растениях. Углеродные скелеты аминокислот образуются из различных промежуточных продуктов центрального углеродного метаболизма (обведены синим цветом). В соответствии с их предшественниками аминокислоты сгруппированы в пять семейств, производных от глутамата, серина, пирувата, аспартата или хоризмата.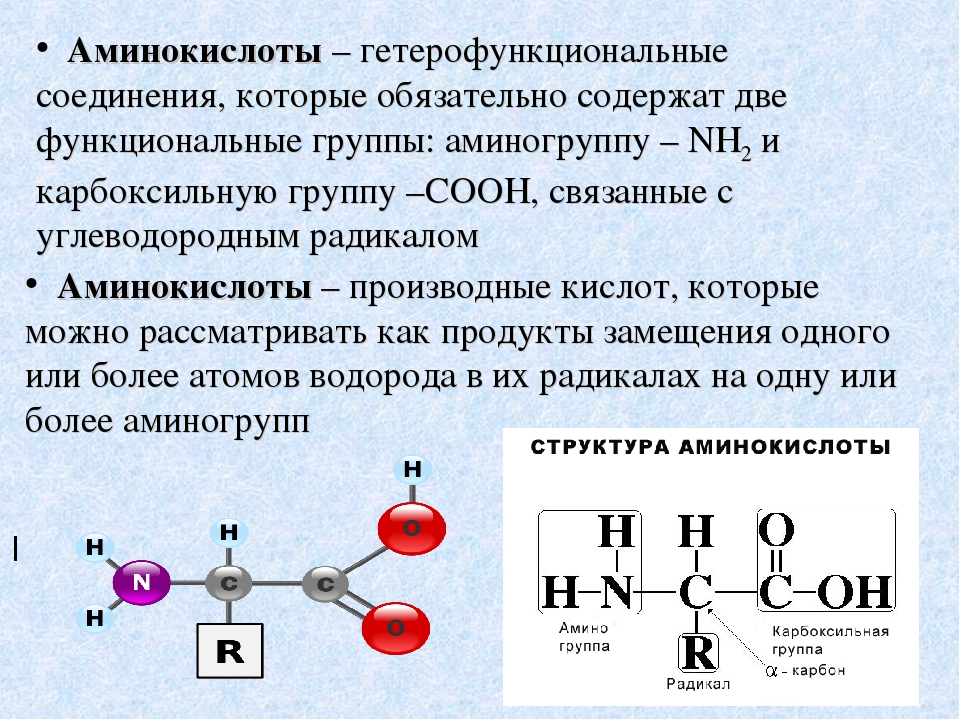 Девять аминокислот, которые не могут быть синтезированы у животных, показаны темно-зелеными прямоугольниками, а те, которые могут быть синтезированы, но дополнительно должны поступать с пищей, отмечены более яркими прямоугольниками.Протеиногенные аминокислоты, которые могут быть в достаточной степени синтезированы у животных, выделены бледно-зелеными рамками, а непротеиногенные аминокислоты и другие важные промежуточные продукты выделены белыми рамками. DAHP, 3-дезокси-D- арабино гептулозонат-7-фосфат.
Девять аминокислот, которые не могут быть синтезированы у животных, показаны темно-зелеными прямоугольниками, а те, которые могут быть синтезированы, но дополнительно должны поступать с пищей, отмечены более яркими прямоугольниками.Протеиногенные аминокислоты, которые могут быть в достаточной степени синтезированы у животных, выделены бледно-зелеными рамками, а непротеиногенные аминокислоты и другие важные промежуточные продукты выделены белыми рамками. DAHP, 3-дезокси-D- арабино гептулозонат-7-фосфат.
Лизин, как одна из незаменимых аминокислот, часто присутствует в растениях в небольших количествах, что ограничивает их пищевую ценность. Обзор Янга и соавт. фокусируется на катаболизме лизина и описывает связи между промежуточными продуктами деградации этой аминокислоты и другими метаболическими путями, такими как метаболизм триптофана, цикл трикарбоновых кислот, реакции на абиотический и биотический стресс, метаболизм крахмала и ответ развернутого белка.Более пристальный взгляд на отношения между лизином и серином представлен Кави Кишор и др. , которые обобщают современные знания о путях биосинтеза, регуляторных механизмах и биологических эффектах обеих аминокислот, уделяя особое внимание сложности их взаимодействий. Авторы сообщают о сложных механизмах транскрипционной и посттранскрипционной регуляции и подчеркивают важность белков, богатых лизином и серином, для развития растений и устойчивости к стрессу. Кроме того, обсуждается роль некодирующих РНК в регуляции белков, богатых лизином и серином, и возможность использования полногеномных стратегий для выявления новых взаимодействий.В то время как интерес к биосинтезу лизина в основном направлен на повышение питательной ценности сельскохозяйственных культур, исследования катаболизма этой аминокислоты больше сосредоточены на устойчивости к биотическим и абиотическим стрессам. Арруда и Баррето изучили известные пути деградации лизина, уделив особое внимание пути сахаропина. Как описано в обзоре, влияние катаболизма лизина на устойчивость к стрессу, вероятно, связано с продукцией пролина и пипеколата из глутамата и α-аминоадипат-δ-полуальдегида, соответственно, опосредованным сахаропиновым путем.
, которые обобщают современные знания о путях биосинтеза, регуляторных механизмах и биологических эффектах обеих аминокислот, уделяя особое внимание сложности их взаимодействий. Авторы сообщают о сложных механизмах транскрипционной и посттранскрипционной регуляции и подчеркивают важность белков, богатых лизином и серином, для развития растений и устойчивости к стрессу. Кроме того, обсуждается роль некодирующих РНК в регуляции белков, богатых лизином и серином, и возможность использования полногеномных стратегий для выявления новых взаимодействий.В то время как интерес к биосинтезу лизина в основном направлен на повышение питательной ценности сельскохозяйственных культур, исследования катаболизма этой аминокислоты больше сосредоточены на устойчивости к биотическим и абиотическим стрессам. Арруда и Баррето изучили известные пути деградации лизина, уделив особое внимание пути сахаропина. Как описано в обзоре, влияние катаболизма лизина на устойчивость к стрессу, вероятно, связано с продукцией пролина и пипеколата из глутамата и α-аминоадипат-δ-полуальдегида, соответственно, опосредованным сахаропиновым путем.
Метионин — еще одна незаменимая аминокислота, интенсивно изучаемая для повышения ее содержания, которое обычно очень мало в сельскохозяйственных культурах и овощах. Цистеин можно синтезировать из метионина, но он по-прежнему считается незаменимым в питании из-за низкого содержания метионина во многих растительных тканях. Уиткомб и др. использовали подход метаболической инженерии для увеличения содержания как метионина, так и цистеина в семенах риса путем создания пары трансгенных линий. Каждая трансгенная линия содержала богатый метионином запасной белок семян и экзогенный фермент для биосинтеза либо метионина, либо цистеина.Эта стратегия успешно увеличила содержание метионина в семенах примерно на 50%, но выявила неожиданный молекулярный фенотип, подобный дефициту серы, и изменение профиля белков семян, возможно, вызванное накоплением развернутых белков в эндоплазматическом ретикулуме. Аналогичный подход использовали Girija et al. исследовать, является ли общая способность синтезировать метионин или плотность остатков метионина в белках семян ограничивающим фактором для содержания метионина в семенах.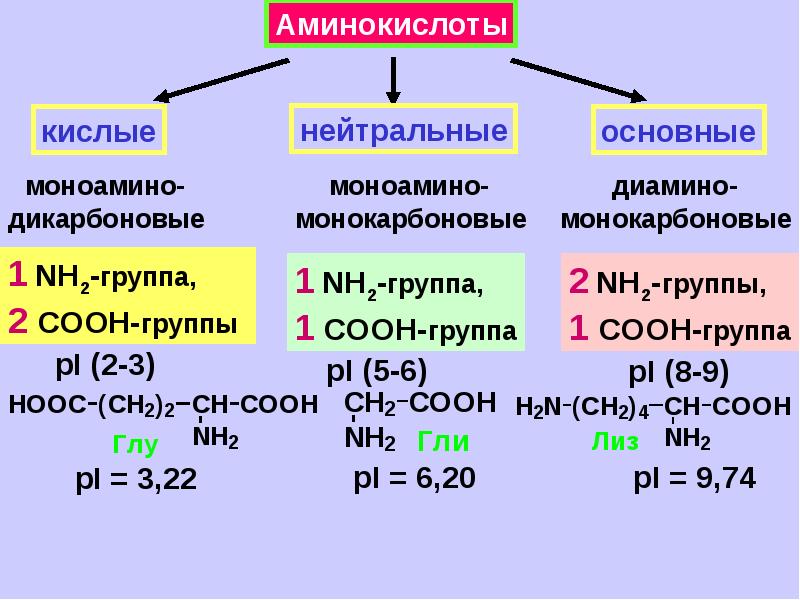 Выводы авторов позволяют предположить, что обилие остатков метионина в запасных белках, вероятно, является основным фактором, ограничивающим накопление метионина в семенах арабидопсиса. Более того, они подтвердили связь между увеличением содержания метионина и накоплением в семенах связанных со стрессом метаболитов, хотя причины этой связи остаются неизвестными. Сложность и важность серосодержащих аминокислот также рассматриваются Watanabe et al. которые рассмотрели метаболизм и регуляторные функции O-ацетилсерина, S-аденозилметионина, гомоцистеина и серина как основных предшественников синтеза цистеина и метионина.
Выводы авторов позволяют предположить, что обилие остатков метионина в запасных белках, вероятно, является основным фактором, ограничивающим накопление метионина в семенах арабидопсиса. Более того, они подтвердили связь между увеличением содержания метионина и накоплением в семенах связанных со стрессом метаболитов, хотя причины этой связи остаются неизвестными. Сложность и важность серосодержащих аминокислот также рассматриваются Watanabe et al. которые рассмотрели метаболизм и регуляторные функции O-ацетилсерина, S-аденозилметионина, гомоцистеина и серина как основных предшественников синтеза цистеина и метионина.
Помимо того, что аминокислоты или их производные являются важными компонентами рациона животных и человека, они могут быть богатым источником нутрицевтических метаболитов. Примером полезного для здоровья функционального соединения является γ-аминомасляная кислота (ГАМК), непротеиногенная аминокислота, замечательные свойства которой были рассмотрены Gramazio et al. Кроме того, в этом обзоре обобщены новейшие стратегии селекции для увеличения содержания ГАМК в сельскохозяйственных культурах с упором на новые подходы на основе CRISPR/Cas9, которые недавно использовались для успешного повышения концентрации ГАМК в спелых плодах томатов без неблагоприятных побочных эффектов.
Метионин, как и другие аминокислоты, также используются для синтеза глюкозинолатов, большого класса серосодержащих метаболитов с признанными антиоксидантными и противоопухолевыми свойствами. Лехлер и др. исследовали функцию изопропилмалатизомеразы, фермента, необходимого для синтеза лейцина и, возможно, участвующего в удлинении цепи метионина. Активный фермент представляет собой гетеродимер, состоящий из большой субъединицы и одной из трех возможных малых субъединиц. У арабидопсиса большой белок кодируется одним геном, а три разных гена кодируют малую субъединицу.Изучая субстратную специфичность и паттерны экспрессии субъединиц, авт. обнаружили, что большая субъединица участвует в метаболизме как лейцина, так и глюкозинолата, а малые субъединицы специфичны для каждого пути. В частности, малая субъединица 1 участвует в биосинтезе лейцина, а малые субъединицы 2 и 3 функционируют в синтезе производных метионина глюкозинолатов.
Помимо своей роли нутрицевтических молекул, многие непротеиногенные аминокислоты участвуют в реакциях растений на стрессы окружающей среды, как было подтверждено Song et al. для цитруллина, промежуточного продукта в синтезе аргинина из орнитина. С помощью транскриптомного и метаболомного анализа авторы продемонстрировали, что быстрое накопление цитруллина и родственных метаболитов в арбузе, подвергнутом водному стрессу, опосредовано синхронизированной активацией биосинтеза и подавлением катаболизма. Кроме того, они обнаружили, что азотный статус растения регулирует синтез цитруллина.
для цитруллина, промежуточного продукта в синтезе аргинина из орнитина. С помощью транскриптомного и метаболомного анализа авторы продемонстрировали, что быстрое накопление цитруллина и родственных метаболитов в арбузе, подвергнутом водному стрессу, опосредовано синхронизированной активацией биосинтеза и подавлением катаболизма. Кроме того, они обнаружили, что азотный статус растения регулирует синтез цитруллина.
Полиамины в растениях образуются либо из аргинина, либо из орнитина.В первом случае аргинин декарбоксилируется до агматина с помощью аргининдекарбоксилазы, а затем превращается в путресцин с помощью агматиниминогидролазы и N-карбамоилпутресцинамидогидролазы. В последнем случае аргинин гидролизуется до орнитина с помощью аргиназы, а затем декарбоксилируется с помощью орнитиндекарбоксилазы до путресцина. Однако недавно было показано, что аргиназы арабидопсиса и сои могут действовать также как агматиназы, обеспечивая третий путь синтеза путресцина (Patel et al., 2017).Чтобы пролить свет на механизм этой реакции, Секула проанализировал с помощью рентгеновских лучей и малоуглового рентгеновского рассеяния кристаллические структуры двух аргиназ из Arabidopsis thaliana и Medicago truncatula и предложил модель, объясняющую двойное связывающие свойства растительных аргиназ.
Среди аминокислот, участвующих в защите от стресса, пролин особенно важен, поскольку он накапливается у большинства видов растений в ответ на различные стрессы и, как полагают, способствует устойчивости к стрессу.Накопление пролина в значительной степени зависит от транскрипционной активации δ 1 -пирролин-5-карбоксилатсинтетазы (P5CS), фермента, катализирующего лимитирующую скорость стадию биосинтеза пролина, которая у большинства видов растений кодируется двумя паралогичными генами. Как сообщают Sabbioni et al., активность P5CS2 в рисе дополнительно регулируется посттрансляционными механизмами, регулирующими синтез пролина в соответствии с окислительно-восстановительным и азотным статусом растительной клетки. У арабидопсиса разные паттерны экспрессии двух изоформ P5CS указывают на функции P5CS1 в индуцированном стрессом накоплении пролина и устойчивости к стрессу и P5CS2 в синтезе пролина для роста и развития.Кроме того, сообщалось о различной локализации двух изоформ как в цитозоле, так и в пластидах (Székely et al.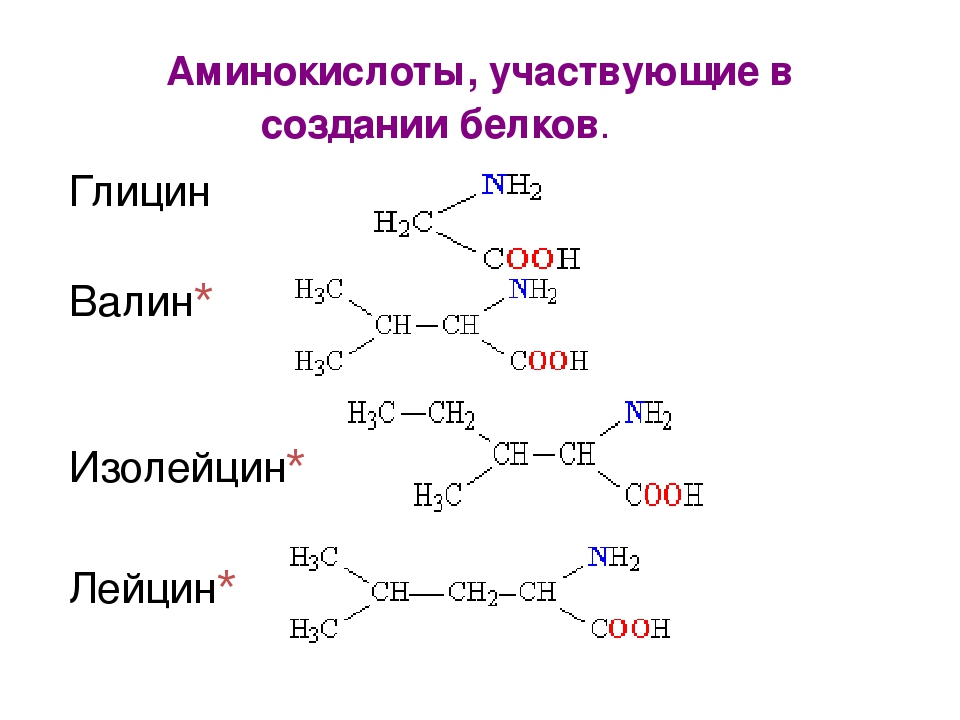 , 2008). Funck et al., однако, частично поставили под сомнение эти представления, обнаружив, что слитые белки P5CS1- и P5CS2-GFP присутствовали только в цитозоле и что вклад обеих изоформ в устойчивость к стрессу был очень низким. Неожиданно эти авторы обнаружили, что мутанты p5cs2 более солеустойчивы, чем мутанты p5cs1 или дикие типы, несмотря на более низкое содержание пролина. Эти результаты предполагают новую функцию P5CS2 в отношении устойчивости к соли и подтверждают гипотезу о том, что метаболизм пролина, а не сам пролин, отвечает за устойчивость к стрессу.Forlani и Funck сообщили о новом методе количественного определения L-пролина. Этот анализ более специфичен, чем широко используемый нингидриновый метод (Bates et al., 1973). По мнению авторов, методы на основе нингидрина ошибочно обнаруживают родственные молекулы, такие как орнитин, гидроксипролин и D-пролин, и теряют линейность в присутствии высоких концентраций аминокислот, что приводит к переоценке содержания пролина. Метод, предложенный Форлани и Функ, основан на обратной реакции редуктазы Р5С (Р5CR) при нефизиологическом рН 10.
, 2008). Funck et al., однако, частично поставили под сомнение эти представления, обнаружив, что слитые белки P5CS1- и P5CS2-GFP присутствовали только в цитозоле и что вклад обеих изоформ в устойчивость к стрессу был очень низким. Неожиданно эти авторы обнаружили, что мутанты p5cs2 более солеустойчивы, чем мутанты p5cs1 или дикие типы, несмотря на более низкое содержание пролина. Эти результаты предполагают новую функцию P5CS2 в отношении устойчивости к соли и подтверждают гипотезу о том, что метаболизм пролина, а не сам пролин, отвечает за устойчивость к стрессу.Forlani и Funck сообщили о новом методе количественного определения L-пролина. Этот анализ более специфичен, чем широко используемый нингидриновый метод (Bates et al., 1973). По мнению авторов, методы на основе нингидрина ошибочно обнаруживают родственные молекулы, такие как орнитин, гидроксипролин и D-пролин, и теряют линейность в присутствии высоких концентраций аминокислот, что приводит к переоценке содержания пролина. Метод, предложенный Форлани и Функ, основан на обратной реакции редуктазы Р5С (Р5CR) при нефизиологическом рН 10.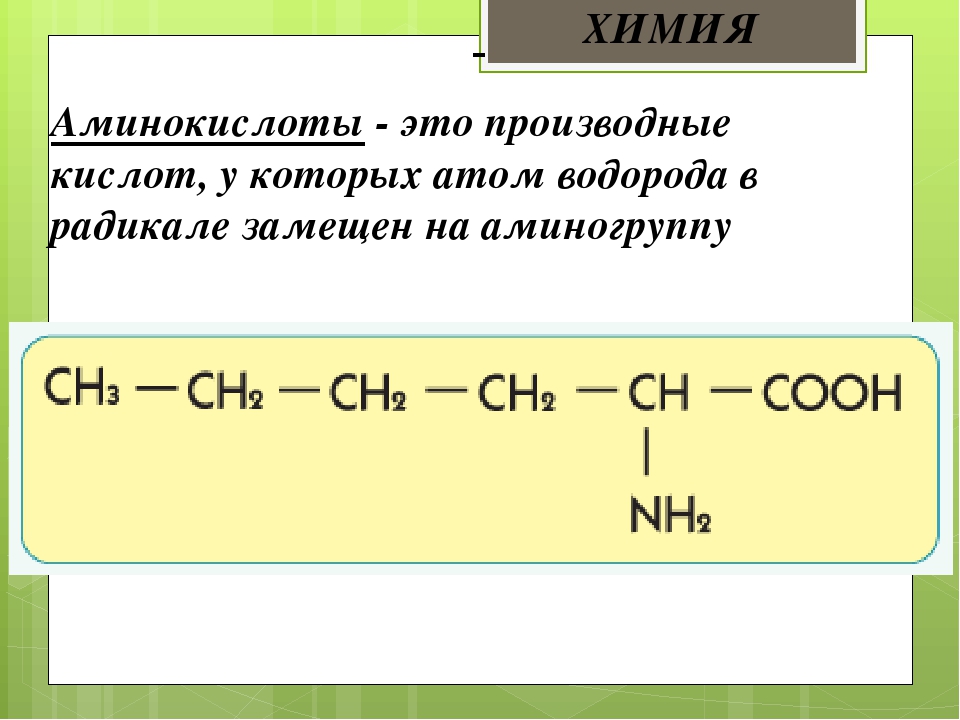 5, может преодолеть пределы специфичности традиционных колориметрических методов, сохраняя при этом аналогичную чувствительность.
5, может преодолеть пределы специфичности традиционных колориметрических методов, сохраняя при этом аналогичную чувствительность.
Независимо от того, обеспечивает ли метаболизм или накопление пролина устойчивость к стрессу, косвенные данные указывают на важность этой аминокислоты на репродуктивной стадии и предполагают, что ее накопление может поддерживать продуктивность в условиях стресса, как сообщает Mattioli et al. и в ячмене Frimpong et al. На основании предыдущей работы (Mattioli et al., 2018), Mattioli et al. подтвердили важность накопления пролина в пыльцевых зернах для поддержания производства семян в условиях солевого стресса, хотя возможность дальнейшего повышения урожайности зерна за счет форсирования синтеза пролина в пыльцевых зернах остается недоказанной. Используя совершенно другой подход, Frimpong et al. проанализировали пять генотипов ярового ячменя с противоположной реакцией на засуху, включая две линии, несущие аллель P5CS1, интрогрессированный от образца дикого ячменя.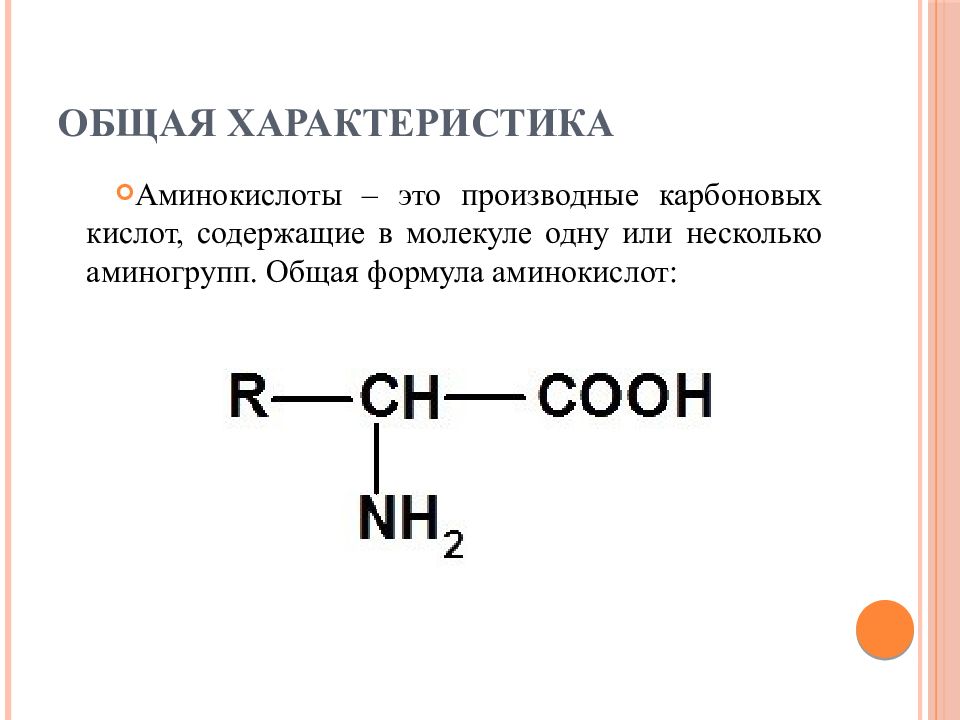 Они обнаружили корреляцию между накоплением пролина и устойчивостью к водному стрессу, особенно у шипов.Линии, несущие дикий аллель P5CS1, оказались более засухоустойчивыми на репродуктивной стадии, что привело к повышению урожайности зерна в условиях водного стресса. Любопытно, что благотворное влияние пролина при стрессе может проявляться и тогда, когда пролин поступает извне, согласно обзору El Moukhtari et al., который обновил современные знания по этой теме и предложил возможные механизмы действия. Хотя мы до сих пор не знаем, как экзогенный пролин может улучшить устойчивость сельскохозяйственных культур к солевому стрессу, эта процедура признана эффективным методом повышения устойчивости сельскохозяйственных культур к стрессу и представляет огромный интерес с точки зрения биотехнологии.Об аналогичном подходе сообщают Alfosea-Simón et al., которые исследовали использование составов для наружного применения, состоящих из различных аминокислот или обогащенных ими, для повышения устойчивости растений к изменениям климата.
Они обнаружили корреляцию между накоплением пролина и устойчивостью к водному стрессу, особенно у шипов.Линии, несущие дикий аллель P5CS1, оказались более засухоустойчивыми на репродуктивной стадии, что привело к повышению урожайности зерна в условиях водного стресса. Любопытно, что благотворное влияние пролина при стрессе может проявляться и тогда, когда пролин поступает извне, согласно обзору El Moukhtari et al., который обновил современные знания по этой теме и предложил возможные механизмы действия. Хотя мы до сих пор не знаем, как экзогенный пролин может улучшить устойчивость сельскохозяйственных культур к солевому стрессу, эта процедура признана эффективным методом повышения устойчивости сельскохозяйственных культур к стрессу и представляет огромный интерес с точки зрения биотехнологии.Об аналогичном подходе сообщают Alfosea-Simón et al., которые исследовали использование составов для наружного применения, состоящих из различных аминокислот или обогащенных ими, для повышения устойчивости растений к изменениям климата.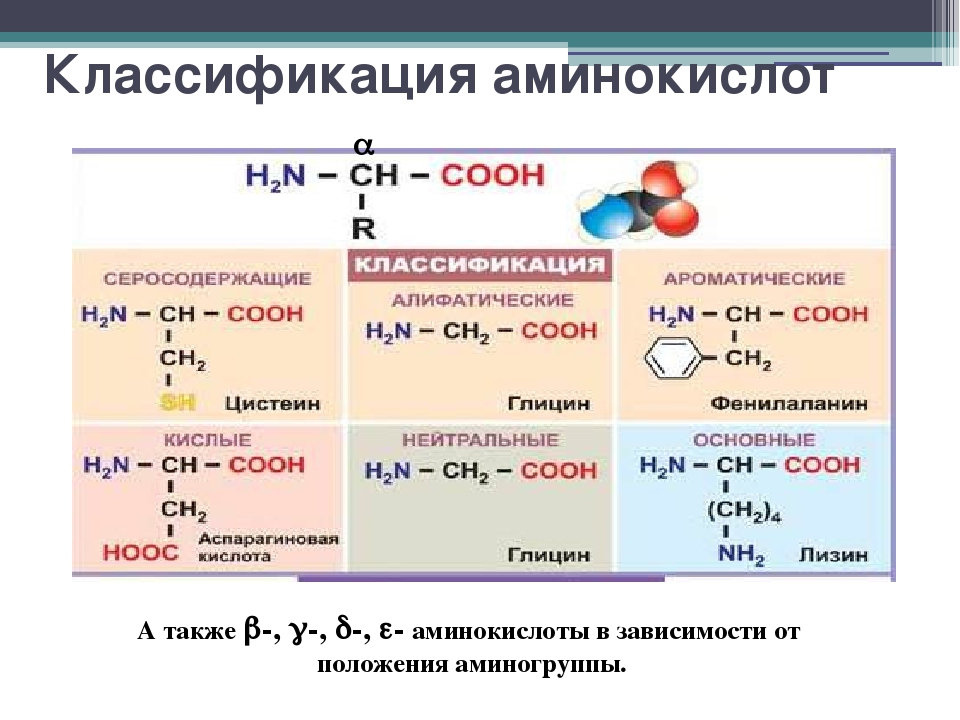 С помощью морфологического, физиологического и метаболического анализов авторы изучили влияние экзогенных аппликаций глутамата, аспартата и аланина на рост томатов и обнаружили синергический положительный эффект аспарагиновой и глутаминовой кислоты и отрицательный эффект аланина.
С помощью морфологического, физиологического и метаболического анализов авторы изучили влияние экзогенных аппликаций глутамата, аспартата и аланина на рост томатов и обнаружили синергический положительный эффект аспарагиновой и глутаминовой кислоты и отрицательный эффект аланина.
Накопление пролина во время стресса зависит как от стимуляции синтеза пролина, так и от ингибирования деградации пролина.Первый процесс катализируется в цитозоле последовательным действием P5CS и P5CR, тогда как второй катализируется в митохондриях последовательным действием пролиндегидрогеназы (ProDH) и пирролин-5-карбоксилатдегидрогеназы (P5CDH). Поскольку P5CS и ProDH катализируют лимитирующие этапы синтеза пролина и окисления из глутамата и в глутамат, соответственно, тщательное определение уровней их активности часто используется в качестве маркера накопления пролина. Распространенная ошибка в определении активности ProDH была обнаружена Lebreton et al., которые показали, что при pH 10 ProDH не катализирует пролин-зависимое восстановление NAD + . Наоборот, эта активность была приписана P5CR, который при высоком, нефизиологическом pH также способен работать в обратном направлении.
Наоборот, эта активность была приписана P5CR, который при высоком, нефизиологическом pH также способен работать в обратном направлении.
Помимо пролина, различные другие аминокислоты участвуют в стрессоустойчивости, среди которых недавно были предложены аминокислоты с разветвленной цепью (BCAA). Буффаньи и др. исследовали роль BCAA в двух сортах твердой пшеницы с контрастной чувствительностью к засухе, выполняя сравнительный биоинформатический анализ и анализ экспрессии генов, кодирующих трансферазы BCAA (BCAAT), и исследуя с помощью ЯМР-анализа метаболический профиль BCAA.В целом они показали, что гены BCAAT индуцируются транскрипцией на ранних фазах реакции на стресс, а накопление BCAA отражает засухоустойчивость сортов, подтверждая участие BCAA в защитной реакции на засуху.
В растениях ароматические аминокислоты (ААА) синтезируются из хоризмата, конечного продукта шикиматного пути, и являются предшественниками широкого спектра вторичных метаболитов. Чтобы исследовать возможную роль ААА в устойчивости к биотическому и абиотическому стрессу, Oliva et al. сгенерировали трансгенные растения табака, сверхэкспрессирующие нечувствительную к обратной связи версию AroG, ген 3-дезокси-D- арабино -гептулозонат-7-фосфатсинтазы, кодирующий первый фермент шикиматного пути. Метаболический анализ подтвердил, что листья трансгенных растений содержат более высокие уровни фенилаланина, тирозина и триптофана, а также родственных метаболитов по сравнению с контрольными растениями. Трансгенные растения приобрели некоторую устойчивость к солевому стрессу, но не к окислительному или засушливому стрессу, а также сильную устойчивость к инфекциям, вызванным паразитом растений Phelipanche aegyptica , что свидетельствует о том, что повышение уровня ААА в растениях может быть эффективной стратегией борьбы с паразитами растений.Путь шикимата и, следовательно, синтез ААА является целью глифосата, гербицида, используемого во всем мире. В частности, глифосат является конкурентным ингибитором фермента 5- енол пирувилшикимат-3-фосфатсинтазы (ЭПСФС).
сгенерировали трансгенные растения табака, сверхэкспрессирующие нечувствительную к обратной связи версию AroG, ген 3-дезокси-D- арабино -гептулозонат-7-фосфатсинтазы, кодирующий первый фермент шикиматного пути. Метаболический анализ подтвердил, что листья трансгенных растений содержат более высокие уровни фенилаланина, тирозина и триптофана, а также родственных метаболитов по сравнению с контрольными растениями. Трансгенные растения приобрели некоторую устойчивость к солевому стрессу, но не к окислительному или засушливому стрессу, а также сильную устойчивость к инфекциям, вызванным паразитом растений Phelipanche aegyptica , что свидетельствует о том, что повышение уровня ААА в растениях может быть эффективной стратегией борьбы с паразитами растений.Путь шикимата и, следовательно, синтез ААА является целью глифосата, гербицида, используемого во всем мире. В частности, глифосат является конкурентным ингибитором фермента 5- енол пирувилшикимат-3-фосфатсинтазы (ЭПСФС). Зулет-Гонсалес и др. проанализировали быстрорастущий сорняк Amaranthus palmeri , некоторые популяции которого устойчивы к глифосату, потому что они сверхэкспрессируют EPSPS, чтобы исследовать роль AAA в регуляции пути шикимата и устойчивости к глифосату.Они обнаружили сложное взаимодействие глифосата и ААА в регуляции обратной связи шикиматного пути, который был изменен сверхэкспрессией EPSPS. Однако механизмы, лежащие в основе этого эффекта, остаются неизвестными.
Зулет-Гонсалес и др. проанализировали быстрорастущий сорняк Amaranthus palmeri , некоторые популяции которого устойчивы к глифосату, потому что они сверхэкспрессируют EPSPS, чтобы исследовать роль AAA в регуляции пути шикимата и устойчивости к глифосату.Они обнаружили сложное взаимодействие глифосата и ААА в регуляции обратной связи шикиматного пути, который был изменен сверхэкспрессией EPSPS. Однако механизмы, лежащие в основе этого эффекта, остаются неизвестными.
Несмотря на множественные функции аминокислот в развитии растений и защите от стресса, аминокислоты нуждаются в азоте для своего биосинтеза, и понимание того, как азот поглощается, хранится и транспортируется в растениях, представляет огромный интерес для биологии аминокислот. О’Нил и Ли описывают метод определения количества и локализации свободных аминокислот в тканях растений, который может оказать большую помощь в решении этих вопросов.Авторы успешно использовали матричную лазерную десорбционную ионизацию (MALDI) и масс-спектрометрическую визуализацию (MSI) в сочетании с дериватизацией конифериловым альдегидом для изучения поглощения и распределения аминокислот в корне кукурузы, предлагая использовать MALDI-MSI в качестве действенный метод изучения усвоения, хранения и транспорта азота в растениях.
Поскольку аргинин имеет высокое отношение азота к углероду, растения, как правило, запасают азот в виде аргинина, когда азота в избытке. Накопление аргинина достигается за счет снятия ингибирования обратной связи N-ацетилглутаматкиназы гена биосинтеза аргинина (NAGK).Этот механизм зависит от регуляторного белка PII, который способен определять азотный и углеродный статус клетки для оптимизации активности биосинтеза аргинина. В исследовании Llebrés et al. представлены структурно-функциональные характеристики белка PII сосны приморской, дополненные новыми сведениями о механизмах регуляции метаболизма аргинина.
Аммоний является основным источником неорганического азота, используемого для синтеза аминокислот. Ассимиляция и рециркуляция аммония требуют согласованной активности глутаминсинтетазы (GS), глутаматсинтазы (GOGAT) и глутаматдегидрогеназы (GDH).В то время как GS и GOGAT являются наиболее важными ферментами для ассимиляции органических молекул в растениях, GDH участвует в гомеостазе глутамата и обеспечивает цикл TCA с 2-оксоглутаратом (2OG), когда доступность углерода ограничена. Структурное исследование GDh2 из Arabidopsis thaliana представлено Grzechowiak et al. которые проанализировали кристаллическую структуру AtGDh2 как в его апо-форме, так и в связанном с ним кофакторе NAD + и продукте реакции 2OG. Исследование показало, что фермент подвергается открытому/закрытому конформационному изменению, которое требует связывания как NAD + , так и 2OG для его полной активации.Большая часть почвенного аммония поглощается растениями семейства высокоаффинных переносчиков аммония (АМТ), особенно АМТ1;1 и АМТ1;2. Недавние данные (Xuan et al., 2016) показали, что лечение брассиностероидами (BR) усиливает экспрессию AMT1;1 и AMT1;2 в рисе, механизм которого еще предстоит понять. Чтобы исследовать взаимосвязь между AMT и BR, Yang et al. изучали уровни экспрессии АМТ и зависящую от АМТ эффективность поглощения аммония линиями риса с измененной экспрессией BES1 и BZR1, двух главных регуляторов передачи сигналов BR у растений.Выводы авторов предполагают, что BR-регуляция поглощения Nh5 + в рисе включает транскрипционную регуляцию переносчиков аммония и что BR-зависимое поглощение аммония частично контролируется BZR1.
Структурное исследование GDh2 из Arabidopsis thaliana представлено Grzechowiak et al. которые проанализировали кристаллическую структуру AtGDh2 как в его апо-форме, так и в связанном с ним кофакторе NAD + и продукте реакции 2OG. Исследование показало, что фермент подвергается открытому/закрытому конформационному изменению, которое требует связывания как NAD + , так и 2OG для его полной активации.Большая часть почвенного аммония поглощается растениями семейства высокоаффинных переносчиков аммония (АМТ), особенно АМТ1;1 и АМТ1;2. Недавние данные (Xuan et al., 2016) показали, что лечение брассиностероидами (BR) усиливает экспрессию AMT1;1 и AMT1;2 в рисе, механизм которого еще предстоит понять. Чтобы исследовать взаимосвязь между AMT и BR, Yang et al. изучали уровни экспрессии АМТ и зависящую от АМТ эффективность поглощения аммония линиями риса с измененной экспрессией BES1 и BZR1, двух главных регуляторов передачи сигналов BR у растений.Выводы авторов предполагают, что BR-регуляция поглощения Nh5 + в рисе включает транскрипционную регуляцию переносчиков аммония и что BR-зависимое поглощение аммония частично контролируется BZR1.
Класс транспортеров, а именно ОБЫЧНО НЕСКОЛЬКО КИСЛОТ, ПЕРЕМЕЩАЮЩИХСЯ В И ИЗ ТРАНСПОРТЕРОВ (UMAMIT), недавно был идентифицирован как переносчики аминокислот (Ladwig et al., 2012). Besnard et al. рассматривали специфическую функцию аминокислот по определению доступности азота в клетке и обмену этой информацией между различными путями для запуска соответствующих метаболических реакций.Они изучили сверхэкспрессию некоторых генов UMAMIT у арабидопсиса, чтобы исследовать возможные связи между транспортом аминокислот и реакцией на стресс, и обнаружили убедительные доказательства того, что активность экспорта аминокислот положительно коррелирует с фенотипами стресса и устойчивостью к патогенам, скорее всего, из-за установления конститутивной салициловой кислоты. кислотно-опосредованная реакция на стресс. Взаимосвязь между аминокислотами и гормональными путями, обнаруженная Besnard et al. и Ян и др. и связи с ключевыми метаболическими путями, о которых сообщают разные авторы этой коллекции, подчеркивают центральную роль аминокислот в стрессе и развитии и прокладывают путь к новым и захватывающим открытиям в фундаментальной и прикладной науке о растениях.
Вклад авторов
Все перечисленные авторы внесли существенный, непосредственный и интеллектуальный вклад в работу и одобрили ее для публикации.
Финансирование
MT подтверждает поддержку Университета Сапиенца (Progetti Ateneo 2020), GF подтверждает поддержку Университета Феррары (FAR 2021), SO подтверждает поддержку Национального научного фонда (MCB 1052048, IOS 135336), RA подтверждает поддержку Колледжа Тель Хай и Израильский научный фонд (1857/20).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все претензии, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Приглашенные редакторы рады поблагодарить всех авторов, которые предоставили свои статьи для этой темы исследования. Мы также выражаем благодарность рецензентам, редакторам и редакции за техническую и научную поддержку.
Ссылки
Бейтс, Л.С., Уолдрен, Р.П., и Тир, И.Д. (1973). Быстрое определение свободного пролина для исследований водного стресса. Растительная почва 39, 205–207. дои: 10.1007/BF00018060
Полнотекстовая перекрестная ссылка | Академия Google
Галили, Г., Амир, А., и Ферни, А. (2016). Регуляция синтеза и накопления незаменимых аминокислот в растениях. год. Rev. Plant Biol . 67, 53–78. doi: 10.1146/annurev-arplant-043015-112213
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
Ladwig, F., Stahl, M., Ludewig, U., Hirner, A.A., Hammes, U.Z., Stadler, R., et al. (2012). Siliques Are Red1 из арабидопсиса действует как двунаправленный переносчик аминокислот, который имеет решающее значение для гомеостаза аминокислот в стручках. Завод Физиол . 158, 1643–1655. doi: 10.1104/стр.111.192583
Завод Физиол . 158, 1643–1655. doi: 10.1104/стр.111.192583
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
Mattioli, R., Biancucci, M., El Shall, A., Mosca, L., Costantino, P., Funck, D., et al. (2018). Синтез пролина в развивающихся микроспорах необходим для развития и фертильности пыльцы. BMC Растительный Биол . 18:356. doi: 10.1186/s12870-018-1571-3
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
Патель, Дж., Ariyaratne, M., Ahmed, S., Ge, L., Phuntumart, V., Kalinoski, A., et al. (2017). Двойное функционирование растительных аргиназ обеспечивает третий путь синтеза путресцина. Растениеводство . 262, 62–73. doi: 10.1016/j.plantsci.2017.05.011
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
Секели, Г., Абрахам, Э., Чепло, А., Риго, Г., Жигмонд, Л., и Чизар, Дж. (2008), Дублированные гены P5CS арабидопсиса играют различные роли в регуляции стресса, контроле развития пролина. биосинтез. Завод J . 53, 11–28. doi: 10.1111/j.1365-313X.2007.03318.x
биосинтез. Завод J . 53, 11–28. doi: 10.1111/j.1365-313X.2007.03318.x
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
Xuan, H.X., Duan, F.Y., Je, B.I., Kim, C.M., Li, T.Y., Liu, J.M., et al. (2016). Родственный ABI3/VP1-подобному 1 (RAVL1) регулирует опосредованную брассиностероидами активацию AMT1;2 в рисе ( Oryza sativa ). Дж. Экспл. Бот . 68, 727–737. дои: 10.1093/jxb/erw442
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера для приема файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie.
- Ваш браузер не поддерживает файлы cookie. Попробуйте другой браузер, если вы подозреваете это.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
 Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie
потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Предоставить доступ без файлов cookie
потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только та информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
.