
Гистология.mp3 — Мышечные ткани
Слушать (13 464 Кб):
Общая гистология — скелетная, сердечная и гладкая мышечные ткани
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве всего организма в целом или его частей (пример – скелетная мускулатура) и движение органов внутри организма (пример – сердце, язык, кишечник).
Свойством изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
Общая характеристика и классификация
Основные морфологические признаки элементов мышечных тканей — удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы — миофиламенты обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков —
В основу классификации мышечных тканей положены два принципа — морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы: исчерченные мышечные ткани и гладкие мышечные ткани.
Поперечнополосатые (исчерченные) мышечные ткани. В цитоплазме их элементов миозиновые филаменты постоянно полимеризованы, образуют с актиновыми нитями постоянно существующие миофибриллы. Последние организованы в характерные комплексы — саркомеры. В соседних миофибриллах структурные субъединицы саркомеров расположены на одинаковом уровне и создают поперечную исчерченность. Исчерченные мышечные ткани сокращаются быстрее, чем гладкие.
Гладкие (неисчерченные) мышечные ткани. Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине нитями.
В соответствии с гистогенетическим принципом в зависимости от источников развития (т.е. эмбриональных зачатков) мышечные ткани подразделяются на 5 типов:
- мезенхимные (из десмального зачатка в составе мезенхимы)
- эпидермальные (из кожной эктодермы и из прехордальной пластинки)
- нейральные (из нервной трубки)
- целомические (из миоэпикардиальной пластинки висцерального листка спланхнотома)
- соматические (миотомные)
Первые три типа относятся к подгруппе гладких мышечных тканей, четвертый и пятый — к подгруппе поперечнополосатых.
Поперечнополосатые мышечные ткани
Имеется две основные разновидности поперечнополосатых (исчерченных) тканей — скелетная мышечная ткань и сердечная мышечная ткань.
Скелетная мышечная ткань
Гистогенез
Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов — миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела.
В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (или миосателлиты). Эти клетки располагаются на поверхности миосимпластов.
Строение
Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной.
Длина всего волокна может измеряться сантиметрами при толщине всего 50—100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Саркомер — это структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски, имеющие неодинаковое лучепреломление (анизотропные A-диски и изотропные I-диски). Каждая миофибрилла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети, или саркоплазматического ретикулума. Соседние саркомеры имеют общую пограничную структуру — Z-линию (или телофрагму). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы тонких, актиновых, филаментов. От соседних Z-линий актиновые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию, или мезофрагму. В узлах этой М-линии закреплены концы толстых, миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в кортикальном слое цитоплазмы, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна.
Источником ионов кальция служат цистерны агранулярной эндоплазматической сети. Они вытянуты вдоль миофибрилл около каждого саркомера и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или (латеральные) L-цистерны.
С поверхности миосимпласта плазмолемма образует длинные трубочки, идущие поперечно в глубину клетки (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда клетка получает сигнал о начале сокращения, этот сигнал перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в цистернах саркоплазматического ретикулума и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет АТФ- АДФ-превращений. Роль АТФазы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается из миоглобина и участвует в биохимических реакциях.
Миосателлитоциты — это малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосателлитоциты одноядерны, их ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов — гликолитических и окислительных.
По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные. Наиболее заметно мышечные волокна различаются особенностями молекулярной организации миозина. Среди различных его изоформ существуют две основных — «быстрая» и «медленная». При постановке гистохимических реакций их различают по АТФазной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они более богаты гликогеном, в них меньше миоглобина, поэтому их называют также белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными.
Свойства мышечных волокон меняются при изменении нагрузок — спортивных, профессиональных, а также в экстремальных условиях (таких как невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
Регенерация скелетной мышечной ткани
Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами.
Восстановление любых тканей организма может осуществляется за счет двух механизмов: гипертрофии и гиперплазии. Под гипертрофией подразумевают компенсаторное увеличение объема самого симпласта, в т.ч. за счет увеличения количества миофибрилл. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмолеммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Под гиперплазией понимают пролиферацию миосателлитоцитов. Сохранившиеся рядом с повреждением миосателлитоциты делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Скелетная мышца как орган
Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани — эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Васкуляризация. Артерии вступают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви 5—6-го порядка образуют в перимизии артериолы. В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тканевых базофилов, принимающих участие в регуляции проницаемости сосудистой стенки.
Иннервация. В мышцах выявлены миелинизированные эфферентные (двигательные), афферентные (чувствительные), а также немиелинизированные вегетативные нервные волокна. Отросток нервной клетки, приносящий импульс от моторного нейрона спинного мозга, ветвится в перимизии. Каждая его ветвь проникает сквозь базальную мембрану, и у поверхности симпласта на плазмолемме образует терминали, участвуя в организации так называемой моторной бляшки, или нервно-мышечного соединения. При поступлении нервного импульса из терминали выделяется ацетилхолин — медиатор, который вызывает возбуждающий потенциал действия, распространяющееся отсюда по плазмолемме миосимпласта.
Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется нервно-мышечной единицей. Характерно, что мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Чувствительные нервные окончания располагаются не на рабочих мышечных волокнах, а связаны со специализированными мышечными волокнами в так называемых мышечных веретенах, которые расположены в перимизии. Волокна в таких чувствительных мышечных веретенах именуются интрафузальными волокнами, а обычные рабочие мышечные волокна – экстрафузальными.
Интрафузальные мышечные волокна веретен значительно тоньше рабочих. Существует два их вида — волокна с ядерной сумкой и волокна с ядерной цепочкой. Каждое мышечное волокно веретена спирально обвито терминалью чувствительного нервного волокна. В результате сокращения или расслабления рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена, соответственно изменяется тонус интрафузальных мышечных волокон. Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом.
Сердечная мышечная ткань
Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани — симметричные участки висцерального листка спланхнотома в шейной части зародыша — так называемые миоэпикардиалъные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 3 вида кардиомиоцитов:
- рабочие, или типичные, или же сократительные, кардиомиоциты,
- атипичные кардиомиоциты (сюда входят пейсмекерные, проводящие и переходные кардиомиоциты, а также
- секреторные кардиомиоциты.
Рабочие (сократительные) кардиомиоциты образуют свои цепочки. Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим.
Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают гормон — натрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах.
Сократительные кардиомиоциты имеют удлиненную (100—150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски. Кардиомиоциты могут ветвиться и образуют трехмерную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Миофибриллы слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены, контактируют с мембранами гладкой эндоплазматической (т.е. саркоплазматической) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта.
Кардиомиоциты соединяются друг с другом своими торцевыми концами. Здесь образуются так называемые вставочные диски: эти участки выглядят как тонкие пластинки при увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (или щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани.
Гладкие мышечные ткани
По происхождению различают три группы гладких (или неисчерченных) мышечных тканей — мезенхимные, эпидермальные и нейральные.
Мышечная ткань мезенхимного происхождения
Гистогенез. Стволовые клетки и клетки-предшественники гладкой мышечной ткани, будучи уже детерминированными, мигрируют к местам закладки органов. Дифференцируясь, они синтезируют компоненты матрикса и коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Структурно-функциональной единицей гладкой, или неисчерченной, мышечной ткани является гладко-мышечная клетка, или гладкий миоцит — это веретеновидная клетка длиной 20—500 мкм, шириной 5—8 мкм. Ядро клетки палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены в цитоплазме около полюсов ядра. Аппарат Гольджи и гранулярная эндо плазматическая сеть развиты слабо, что свидетельствует о малой активности синтетических функций. Рибосомы в большинстве своем расположены свободно.
Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно, точнее косо-продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца.
Миозиновые филаменты находятся в деполимеризованном состоянии. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует впячивания — кавеолы, в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков (здесь из пузырьков освобождается кальций). Это влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и «миофибриллы» распадаются. Таким образом, актино-миозиновые комплексы существуют в гладких миоцитах только в период сокращения.
Гладкие миоциты располагаются без заметных межклеточных пространств и разделены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Ретикулярные волокна проникают в щели на концах миоцитов, закрепляются там и передают усилие сокращения клетки всему их объединению.
Регенерация. Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается (рабочая гипертрофия клеток). Не исключена, однако, и пролиферация клеток (т.е. гиперплазия).
В составе органов миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова.
Гладкая мышечная ткань эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с железистыми секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках — сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Гладкая мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
Некоторые термины из практической медицины:
- лейомиома — доброкачественная опухоль, развивающаяся из гладкой мышечной ткани;
- миогелез — образование в мышцах болезненных очагов уплотнения, обусловленное переходом коллоидов миофибрилл в фазу геля, их гомогенизацией и восковидным некрозом; наблюдается, напр., при охлаждении тела, травмах;
- миоциты Аничкова — клетки с характерным расположением ядерного хроматина в виде зубчатой полоски, проявляющие фагоцитарную активность; встречаются в миокарде, напр. при миокардитах;
www.morphology.dp.ua
мышечные ткани
Профессор Ролдугина Н.П.
ЛЕКЦИЯ: МЫШЕЧНЫЕ ТКАНИ
Мышечная ткань— важнейшая из всех гистологических тканей, составляющих мясную продукцию животных.
В живом организме мышечные ткани специализированы для сократительных процессов и выполняют сложную работу: приводят в движение рычаги скелета, осуществляя тем самым передвижение животных, обеспечивают ритмичную деятельность сердца и циркуляцию крови в сосудах, активно участвуют в перистальтике пищеварительного тракта, в дыхательном процессе, в функционировании сфинктеров, способствуют поддержанию тонуса организма, его формы и позы.
Таким образом, мышечные ткани осуществляют ряд произвольных и непроизвольных движений всевозможных органов и тканей, а также всего организма, расходуя при этом большое количество энергии. Поэтому эти ткани всегда снабжены густой сетью кровеносных сосудов, беспрерывно доставляющих питательные и энергетические ресурсы, необходимые для синтеза и восстановления веществ, участвующих в образовании двигательных реакций. Сокращение мышц вызывается их возбуждением, поэтому к мышцам подходят многочисленные нервы и нервные окончания.
Кроме собственно-мышечных структур в состав мышечной ткани входят соединительно-тканные элементы.
Для мышечных тканей характерны удлиненная форма составляющих их структурных элементов — миоцитов и мышечных волокон и сократительного аппарата, представленного системой тонких нитей (миофиламентов), состоящих из белков актина и миозина. Эти тонкие короткие нити расположены равными чередующимися рядами внутри миофибрилл, специфических органелл в мышечных волокнах и клетках мышечных тканей.
Классификация мышечных тканей
Мышечные ткани делятся на две основные группы: неисчерченные (гладкие) и исчерченные (поперечно-полосатые).
По источнику эмбрионального развития гладкие мышечные ткани подразделяются на 3 подгруппы (схема 1).
Схема 1.
Гладкие мышечные ткани
мезенхимные (в сосудах и внутренних органах) | эктодермальные (миоэпителиальные клетки слюнных, молочных, слезных и потовых желез) | нейральные (гл. мышечная ткань радужной оболочки глаз и ресничного тела) |
Исчерченные (поперечно-полосатые)
скелетные (миотомного) происхождения) | сердечные (целомического происхождения), из клеток, выстилающих полость целома, формируются мышцы сердца. |
Скелетная ткань— это сократительная ткань туловища, головы, конечностей, глотки, гортани, верхней половины пищевода (у жвачных и хищных животных всего пищевода), языка, жевательных мышц, диафрагмы, глазного яблока, уха и др.
Эта ткань является основой мясной продукции животных и составляет у крупного и мелкого рогатого скота от 45 до 65%, а у свиней от 35 до 55% массы туши. На это соотношение влияет возраст, порода, упитанность животных и др. факторы.
Основная функция этой ткани — движение животного в пространстве и движение отдельных частей тела. Кроме того, она является основным источником тепла в организме, т.к. 30-40% энергии мышечного сокращения используется в виде механической работы, остальное превращается в тепловую энергию. Скелетную мышечную ткань относят к произвольной мускулатуре, т.к. ее сокращение контролируется волей животного.
В основе строения поперечно-полосатой мышечной ткани лежат мышечные волокна (симпласты), имеющие вид тонких длинных цилиндров с тупыми или слегка заостренными концами.
Длина волокон колеблется от нескольких мм до 13-15 см диаметр от 10 до 150 мкм. Объединенные с помощью прослоек рыхлой соединительной ткани пучки таких волокон составляют мышцы животных. Волокна в пучках лежат продольно в одном направлении.
В каждом волокне различают наружную оболочку (сарколемму), цитоплазму или саркоплазму, многочисленные овальные ядра и белковые сократимые нити (миофибриллы).
Оболочка волокна (сарколемма) представляет собой тонкую прозрачную пленку, состоящую из двух слоев, которые видны только с помощью электронной микроскопии. Наружный слой оболочки представлен базальной мембраной. Второй слой сарколеммы является собственной оболочкой волокна или плазмолеммой. С помощью этого слоя осуществляется активный процесс обмена веществ между мышечным волокном и окружающей средой.
Каждое волокно одето тонкой соединительно-тканной оболочкой — эндомизием. Волокна эндомизия (коллагеновые и эластические) вплетаются в базальную мембрану. Несколько мышечных волокон объединяются в группы, разделенные более широкими соединительно-тканными прослойками, называемыми перимизием. Мышца снаружи покрыта эпимизием или фасцией.
В эндомизии расположены лимфатические сосуды и питающие кровеносные сосуды, образующие сеть капилляров вокруг каждого мышечного волокна. В перемизии могут откладываться жировые клетки, придавая мясу мраморность.
На поверхности мышечных волокон находятся специальные нервные окончания — моторные бляшки, через которые поступают нервные импульсы. Проведение их к миофибриллам обеспечивается поперечными трубочками, образованными глубокими впячиваниями плазмолеммы внутрь волокна. Эти трубки расположены через равные промежутки по длине мышечного волокна. Под плазмолеммой находятся многочисленные ядра.
В периферической части саркоплазмы у полюсов ядер расположены участки эндоплазматической сети, комплекс Гольджи, митохондрии.
В центральной части симпластов расположены исчерченные миофибриллы, между которыми находятся митохондрии, канальцы агранулярной эндоплазматической сети и включения гликогена, необходимого для обеспечения процесса сокращения энергией.
В саркоплазме мышечных волокон имеется растворимый пигментный белок — миоглобин, способный связывать кислород и отдавать его по мере необходимости. Мясо, содержащее много миоглобина, приобретает коричневый оттенок.
Общее количество миофибрилл в различных волокнах различно. В некоторых волокнах их очень много. Волокна с большим количеством миофибрилл обладают большой силой и вместе с тем быстро утомляются, т.к. в них небольшое количество жидкой саркоплазмы, поставляющей энергетические ресурсы для сократительных реакций миофибрилл. Волокна с большим количеством миофибрилл делают мясо более светлым, т.к. в таких волокнах мало саркоплазмы, содержащей красноватый пигмент миоглобин. Волокна, составляющие мышцу, с небольшим содержанием миофибрилл имеют красноватый оттенок. Эти мышцы меньше утомляются, но обладают меньшей силой.
Мышцы молодых животных окрашены менее интенсивно, чем старых, так как в их волокнах содержится меньше миоглобина.
При длительном воздействии кислорода воздуха на мясо во время хранения мясных продуктов двухвалентное железо молекул миоглобина окисляется в 3-х валентное, что придает мясу коричневый оттенок.
По содержанию миоглобина и другим признакам в мышцах различают три типа мышечных волокон: белые, красные и промежуточные. Белые бедны миоглобином, имеют меньше митохондрий, содержат большое количество гликогена, значительное число миофибрилл и хорошо развитую саркоплазматическую сеть. Мышцы с преобладанием белых волокон быстро сокращаются и быстро утомляются. Источником АТФ в белых волокнах служит анаэробная фаза клеточного дыхания (гликолиз). Противоположные признаки характерны для красных волокон. В их саркоплазме содержится больше миоглобина, много митохондрий, слабо развита эндоплазматическая сеть, мало гликогена, и сравнительно немного миофибрилл. Источником АТФ служит аэробная фаза клеточного дыхания. Мышцы с преобладанием красных волокон сокращаются в десятки раз медленнее. Сокращение их непрерывное и длительное. Такие мышцы в организме выполняют большую физическую работу.
В разных мышцах количественное соотношение между тремя типами волокон различное, и оно запрограммировано при эмбриональном развитии мышцы. У домашних кур грудные мышцы мало используются (куры почти не летают) и в них преобладают белые мышечные волокна и, наоборот — мышцы ног красные.
Наибольший объем в мышечном волокне занимают миофибриллы. Длина их равна длине волокон. Их количество в разных волокнах различное. В узких пространствах между миофибриллами находятся цистерны эндоплазматической сети и промежуточные микрофиламенты из белка десмина.
По длине миофибрилл чередуются темные и светлые поперечные полоски (диски).
Темные диски всех миофибрилл в волокне расположены против темных, а светлые — против светлых, что и придает всему волокну в целом поперечную исчерченность.
Структурными и функциональными единицами миофибрилл являются саркомеры. Длина их около 4 мкм. В каждый саркомер входят две половинки светлых дисков и целый темный.
Саркомеры состоят из тончайших белковых волоконец -миофиламентов (протофибрилл). Различают два из вида: толстые — из белка миозина, и тонкие — их белка актина. Актиновые миофиламенты образованы двойной спиралью глобулярных белков. Внутри этих спиралей находятся другие белки — тропомиозин и торопомин, которые оказывают регулирующее влияние на актин. Диаметр актиновых нитей ≈7 нм.
Миозиновые нити образованы молекулами белка, также закрученными в виде спирали. В них различают хвостовые части и головки. Головки подвижны и могут подсоединяться к актиновым нитям, втягивая их внутрь. Хвостами молекулы соединяются и образуют толстые нити диаметром 10-12 нм.
Каждая миозиновая нить состоит из 350-500 молекул.
Миозиновые нити составляют темный диск. Каждую толстую миозиновую нить окружают шесть тонких актиновых нитей. Тонкие нити вдвигаются при сокращении между толстыми. И если мышечное волокно расслаблено, то светлые диски состоят только из тонких актиновых протофибрилл, а темные диски состоят только из одних толстых миозиновых протофибрилл. Если мышечное волокно сокращено, то темные диски в ширине не меняются, а светлые сужаются, т.к. тонкие нити вдвигаются между толстыми. Таким образом, мышечное волокно сокращается с помощью протофибрилл (миофиламентов). Процесс сокращения осуществляется не за счет образования складчатых миофибрилл (как предполагали до изобретения электронного микроскопа), а за счет скольжения актиновых и миозиновых нитей одна возле другой. Одну толстую нить окружают 6 тонких.
Для образования акто-миозинового комплекса необходимы ионы Са++, которые поступают из депо внутри клетки (эндоплазматического ретикулума) и извне, через определенные участки плазмолеммы, где расположены специфические белки, восприимчивые к ионам Са+2. Для запуска сокращения необходимы специальные белки (кальмодулин) и ферменты (киназа). Саркомеры разграничены друг от другаZ-полосками, фиксирующими актиновые нити. Миозиновые нити фиксируются М-полоской, от которой с боков находится более светлый участок, куда не доходят актиновые нити (Н-зона). Таким образом, светлые и темные полоски вследствие их фиксированного состояния на одном уровне создают рисунок поперечной исчерченности. Темные диски называют анизатропными, а светлые — изотропными.
В мышечных волокнах млекопитающих поперечные трубочки (Т‑системы) расположены в саркомерах на границе темных и светлых дисков. Концы поперечных трубочек внутри волокна контактируют с терминальными цистернами саркоплазматической сети и, вместе с последними, образуют триады. Когда волокно получает от T-трубочек нервный импульс, то депонированные в цистернах саркоплазматической сети ионы кальция выходят из них, проникают в миофибриллы и запускают процесс сокращения. Во время расслабления мышечных волокон ионы кальция перекачиваются обратно в полость эндоплазматической сети.
Развитие поперечнополосатой скелетной мышечной ткани
Скелетная мышечная ткань развивается из миотомов сегментированного отдела мезодермы. На ранней стадии развития миотомы состоят из плотно расположенных мышечных клеток — миобластов. Цитоплазма этих клеток имеет тонковолокнистое строение, свидетельствующее о развитии сократительных белков. Уже на этой стадии миобласты способны к сокращению. Они интенсивно делятся и перемещаются в участки расположения будущих мышц. Вскоре в цитоплазме миобластов можно различать единичные сократительные нити — миофибриллы, построенные из сократительных белков. Ядра делятся интенсивнее клеток и миобласты становятся многоядерными. Увеличиваясь в длину, они приобретают форму волокон (симпластов). Симпласты могут образовываться и путем слияния миобластов торцевыми концами. Образуются мышечные трубочки, которые разделяются продольно на мышечные волокна.
Часть миобластов не проходят указанный путь, и сохраняются в виде удлиненных одноядерных малодифференцированных клеток, расположенных между плазмолеммой и базальной мембраной. Такие клетки называются миосателлитами. Считают, что они могут быть источником образования новых волокон в течение всей жизни животного. Кроме того, новые волокна могут образовываться путем расщепления толстых волокон и увеличения количества миофибрилл в тонких волокнах.
Возрастные и другие изменения скелетной мышечной ткани
С возрастом в мышечных тканях происходит увеличение размеров волокон. Диаметр волокон увеличивается за счет прибавления протофибрилл (мифиламентов), по бокам миофибрилл. В длину волокно растет за счет присоединения протофибрилл к концам миофибрилл. За счет продольного расщепления увеличивается и количество миофибрилл. Эти процессы происходят и в эмбриональный и постэмбриональный периоды. За это время мышечные волокна утолщаются в 2-4 раза. У молодых животных прослойки рыхлой соединительной ткани не содержат жира.
С ростом животного увеличивается количество пучков волокон путем врастания внутрь соединительно-тканных прослоек. С возрастом внутри мышц рыхлая соединительная ткань уплотняется и мясо становится более жёстким. У животных скороспелых пород мясного типа мышечные волокна более толстые, в межпучковой соединительной ткани (премизии) чаще встречаются жировые клетки. При откорме утолщаются мышечные волокна, между ними появляются жировые клетки. Недостаток кормления приводит к уменьшению диаметра мышечных волокон на 20-35%.
Сердечная мышечная ткань
Сердечная мышечная тканьформирует среднюю оболочку (миокард) предсердий и желудочков сердца и представлена двумя разновидностями рабочей и проводящей.
Рабочая мышечная тканьсостоит из клеток кардиомиоцитов, важнейшей особенностью которых является наличие совершенных контактных зон. Соединяясь друг с другом, торцевыми концами они формируют структуру, сходную с мышечным волокном. На боковых поверхностях кардиомиоциты имеют ответвления. Соединяясь концами с ответвлениями соседних кардиомиоцитов они образуют анастомозы. Границами между торцами соседних кардиомиоцитов являются вставочные диски с прямыми или ступенчатыми контурами. В световом микроскопе они имеют вид поперечных темных полосок. С помощью вставочных дисков и анастомозов сформирована единая структурно-функциональная сократительная система.
При электронной микроскопии выявлено, что в области вставочных дисков одна клетка вдается в другую пальцевидными выступами, на боковых поверхностях которых имеются десмосомы, что обеспечивает высокую прочность сцепления. На концах пальцевидных выступов обнаружены щелевидные контакты, через которые нервные импульсы быстро распространяются от клетки к клетке без участия медиатора синхронизируя сокращение кардиомиоцитов.
Сердечные миоциты – это одноядерные, иногда двухядерные клетки. Ядра расположены в центре в отличие от скелетных мышечных волокон. В околоядерной зоне расположены компоненты аппарата Гольджи, митохондрии, лизосомы, гранулы гликогена.
Сократительный аппарат миоцитов, так же как и в скелетной мышечной ткани, состоит из миофибрилл, которые занимают периферическую часть клетки. Их диаметр от 1 до 3-х мкм.
Миофибриллы сходны с миофибриллами скелетной мышечной ткани. Они также построены из анизотропных и изотропных дисков, что также обуславливает поперечную исчерченность.
Плазмолемма кардиомиоцитов на уровне Z-полосок инвагинирует в глубь цитоплазмы, образуя поперечные трубочки, отличающиеся от скелетной мышечной ткани большим диаметром и наличием базальной мембраны, которая покрывает их снаружи, как и сарколемму. Волны деполяризации, идущие с плазмолеммы внутрь сердечных миоцитов, вызывают скольжение актиновых миофиламентов (протофибрилл) по отношению миозиновым, обуславливая сокращение, как и в скелетной мышечной ткани.
Т-трубочки в сердечных рабочих кардиомиоцитах образуют диады, то есть связаны с цистернами саркоплазматической сети только с одной стороны. Рабочие кардиомиоциты имеют длину 50-120 мкм, ширину 15-20 мкм. Количество миофибрилл в них меньше, чем в мышечных волокнах.
Сердечная мышечная ткань содержит много миоглобина, поэтому темно-красного цвета. В миоцитах много митохондрий и гликогена, т.е.: энергию сердечная мышечная ткань получает и при распаде АТФ, и в результате гликолиза. Таким образом, сердечная мышца работает непрерывно всю жизнь, из-за мощной энергетической оснащенности.
Интенсивность и частота сокращений сердечной мышцы регулируются нервными импульсами.
В эмбриогенезе рабочая мышечная ткань развивается из особых участков висцерального листка несегментированной мезодермы (спланхнотома). В сформировавшейся рабочей мышечной ткани сердца отсутствуют камбиальные клетки (миосателлиты), поэтому при повреждении миокарда в травмированной зоне кардиомиоциты погибают и на месте повреждения развивается волокнистая соединительная ткань.
Проводящая мышечная ткань сердцанаходится в составе комплекса образований синусно-предсердного узла, расположенного в устье краниальной полой вены, предсердно-желудочкового узла, лежащего в межпредсердной перегородке, предсердно-желудочкового ствола (пучка Гиса) и его разветвлений, находящихся под эндокардом межжелудочковой перегородки и в соединительно-тканных прослойках миокарда.
Все компоненты этой системы образованы атипичными клетками, специализированными либо на выработке импульса, распространяющемуся по всему сердцу и вызывающего сокращение его отделов в необходимой последовательности (ритме), либо в проведении импульса к рабочим кардиомиоцитам.
Для атипичных миоцитов характерен значительный объем цитоплазмы, в которой немногочисленные миофибриллы занимают периферическую часть и не имеют параллельной ориентации, вследствие чего этим клеткам не свойственна поперечная исчерченность. Ядра расположены в центре клеток. Цитоплазма богата гликогеном, но в ней мало митохондрий, что свидетельствует об интенсивном гликолизе и низком уровне аэробного окисления. Поэтому клетки проводящей системы более устойчивы к кислородному голоданию, чем сократительные кардиомиоциты.
В составе синусно-предсердного узла атипичные кардиомиоциты более мелкие, округлой формы. В них формируются нервные импульсы и они относятся к главным водителям ритма. Миоциты предсердно-желудочкового узла несколько крупнее, а волокна пучка Гиса (волокна Пуркинье) состоят из крупных округлых и овальных миоцитов с эксцентрично расположенным ядром. Диаметр их в 2-3 раза больше, чем рабочих кардиомиоцитов. Электронно-микроскопически выявлено, что в атипичных миацитах слаборазвита саркоплазматическая сеть, отсутствует система Т-трубочек. Клетки соединяются не только концами, но и боковыми поверхностями. Вставочные диски устроены более просто и не содержат пальцевидных соединений, десмосом и нексусов.
Гладкая мышечная тканьобразует мышечную оболочку трубкообразных органов пищеварения, дыхания, выделения, размножения, находится в стенках кровеносных сосудов, протоков желез, в селезенке, коже и других органах.
Специализированные сократительные гладкомышечные тканивходят в состав потовых, слюнных, молочных желез. Сократительные клетки этих желез в своей цитоплазме содержат миофиламенты, построенные из сократительных белков и развиваются из эпителиальных клеток. Другие разновидности специализированных сократительных тканей имеют нейроглиальное происхождение, суживают зрачок и располагаются в радужной оболочке глаза.
Гладкая мышечная ткань относится к ткани с непроизвольным сокращением, её функцию контролирует вегетативная нервная система. Сокращения гладких мышц могут быть медленными, но достигать большой силы сжатия.
Основной структурной единицей гладкой мышечной ткани являются клетки-миоциты. Они удлиненной веретеновидной формы с заостренными концами. Их длина от 20 до 200 мкм (в беременной матке до 500 мкм), а толщина 8-10 мкм. Ядро палочковидной формы находится в середине клетки. В цитоплазме, около полюсов ядра расположены органеллы: митохондрии, комплекс Гольджи, центросома, рибосомы, эндоплазматическая сеть и включения гликогена (энергетический резерв клетки). В преферической части цитоплазмы расположены миофиламенты. Нити актина и миозина не образуют миофибрилл или постоянных акто-миозиновых комплексов и расположены по-разному. Актиновые нити чаще имеют косое присоединение к плазмолемме с помощью особых плотных телец (прикрепительных дисков). Отдельные пучки актиновых нитей прикрепляются к плотным тельцам, расположенным в цитоплазме. Положение этих телец или дисков с обратной стороны закрепляется промежуточными филаментами.
Миозиновые нити в периоды расслабления миоцитов лежат в цитоплазме продольно или под углом к длинной оси клетки. В процессе сокращения актиновые и миозиновые нити смещаются навстречу друг другу и формируют акто-миозиновые комплексы. В результате клетка сокращается и приобретает неправильную форму. В фазе расслабления комплексы вновь распадаются. Поскольку актиновые и миозиновые нити лежат неупорядоченно, поперечная исчерченность в гладких миоцитах отсутствует.
В процессе сокращения, как было сказано, важную роль играют ионы Са++. Депо для них является гладкая эндоплазматическая сеть миоцита. Кроме того, ионы Са++ поступают извне через кальциевые каналы в цитолемме. В определенных участках плазмолеммы лежат специальные белки, воспринимающие и пропускающие внутрь ионы Са++. Ионы Са++ в комплексе с белком кольмодулином и ферментом киназой запускают процесс сокращения. Головки молекул миозина начинают двигаться и скользить вдоль нитей актина и осуществляется процесс сокращения.
С помощью электронного микроскопа было выявлено, что на концах гладких миоцитов имеются пальцевидные выпячивания, десмосомы и щелевидные контакты-нексусы. Плазмолемма миоцитов впячиваясь в цитоплазму, образует пузырьки (кавеолы), примыкающие к саркоплазматической сети. Предполагают, что эти пузырьки участвуют в проведении нервных импульсов, вызывающих выход ионов Са++ и процесс сокращения.
Функциональной единицей гладкой мышечной ткани является пучок из 10-15 миоцитов, связанных с одним нервным волокном. Благодаря тесной связи клеток с помощью десмосом и щелевых контактов все клетки пучка быстро реагируют на нервное раздражение, несмотря на то, что нервное окончание входит только в одну клетку.
Коллагеновые волокна, соединительно-тканных капсул (эндомизий) оплетают миоциты, вплетаются в базальную пластинку (мембрану) в наружный слой над сарколеммой и тем самым удерживают клетки от чрезмерного сжатия и растяжения.
Пучки отделены друг от друга прослойками соединительной ткани (перемизий), в которой проходят сосуды и нервы.
Гладкая мышечная ткань, иннервируется вегетативной нервной системой. Ее деятельность регулируется корой полушарий, но без участия сознания. Сокращения осуществляются непроизвольно и происходят медленно и ритмично (период сокращения от 3 до 5 минут).
Такой характер сокращения называют тоническим.
В стенках полых органов и сосудов пучки гладких миоцитов объединяются в пласты (продольные и циркулярные).
Гладкая мышечная ткань обладает большой силой, передвигая в кишечнике большие массы пищи, и обладает слабой утомляемостью. В стенке кишечника сокращение происходит 12 раз в минуту.
Отдельные пучки гладких миоцитов находятся в коже животных в виде мышц, поднимающих волос.
Происходит гладкая мышечная ткань внутренних органов и сосудов из мезенхимы, клетки которой дифференцируются в миобласты, а миобласты в миоциты, сохраняющие способность к делению на протяжении всего онтогенеза. Кроме того, гладкие миоциты могут образовываться из недеффиренцированных клеток соединительной ткани (адвентициальных), находящихся около кровеносных сосудов.
studfiles.net
| вид мышечной ткани | ПОПЕРЕЧНОПОЛОСАТАЯ | ГЛАДКАЯ | |
| СКЕЛЕТНАЯ | СЕРДЕЧНАЯ (МИОКАРД) | ||
| образована | образована скелтными поперечнополосатыми мышечными волокнами, которые представляют собой длинные лентовидные клетки (как шланги) — симпласты с большим количеством ядер | образована клетками — кардиомиоцитами, имеющими цилиндрическую ветвящуюся форму, клетки соединяются конец в конец, образуя клеточные цепочки, места соединения кардиомиоцитов называются вставочными дисками, в них много десмосом и нексусов; кардиомиоциты имеют от одного до нескольких ядер | образована клетками — гладкими миоцитами, они веретенообразной формы с одним вытянутым ядром |
| поперечная исчерченность | есть, она обусловлена наличием строго ориентированных миофибрилл | нет; хотя миофибриллы и есть, но они не имеют упорядоченного расположения | |
| расположение ядер | по периферии клеток | в центре клетки | в центре клетки |
| митохондрии | активные | очень активные, их очень много | не очень активные |
| особенности | в цитоплазме хорошо развит гладкий эндоплазматический ретикулум, который: оплетает каждую миофибриллу, подходит близко к Т-трубочкам, является хранилищем ионов кальция; в цитозоле имеются включения глкогена, содержится белок миоглобин, способный связывать кислород; | ||
| Т-трубочки | есть | есть, в них проникает базальная мембрана | нет |
| базальная мембрана | снаружи каждое мышечное волокно, кардиомиоцит, и гладкомышечная клетка окружены базальной мембраной | ||
| тропонин- тропомио- зиновый комплекс | есть | есть | нет |
| прослойки соедини- тельной ткани | есть, в скелетных мышцах мышечные волокна собраны в пучки, между которыми имеются прослойки рыхлой соединительной ткани: эндомизий образует пучки 1-го порядка, перимизий отграничивает пучки 2-го порядка, эпимизий — пучки 3-го порядка | есть | есть |
| малодиф- ференциро- ванные клетки | есть — миосателллитные клетки, располагаются под базальной мембраной мышечного волокна, обеспечивают регенерацию мышечного волокна | нет | есть, ими являются малодифференцированные клетки мезенхимы, их них могут образовываться новые гладкомышечные клетки |
| регенерация | могут образовываться новые мышечные волокна за счет малодифференцированных миосателлитных клеток, внутриклеточная регенерация | только внутриклеточная регенерация, новых кардиомиоцитов не образуется, в случае гибели кардиомиоцитов дефект миокарда замещается соединительной тканью | могут образовываться новые гладкомышечные клетки путем деления и из малодифференцированых клеток мезенхимы, внутриклеточная регенерация |
| источник развития | миотом сомитов | миоэпикардиальные пластинки висцерального листка спланхнотома | мезенхима |
histol.ru
| СКЕЛЕТНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ продольный срез Окраска гематоксилин-эозином Показаны продольно-срезанные мышечные
волокна; |  |
| СКЕЛЕТНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ продольный срез Окраска железным гематоксилином Показаны продольно-срезанные мышечные
волокна; |  |
| СКЕЛЕТНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ
ТКАНЬ продольный срез Окраска железным гематоксилином Показаны продольно-срезанные мышечные
волокна; | 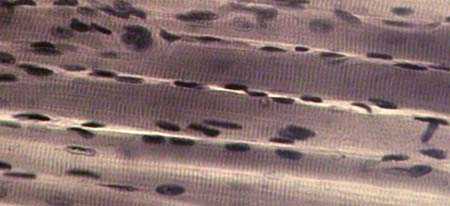 |
| СКЕЛЕТНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ поперечный срез Окраска железным гематоксилином 1 — продольно-срезанные мышечные волокна | 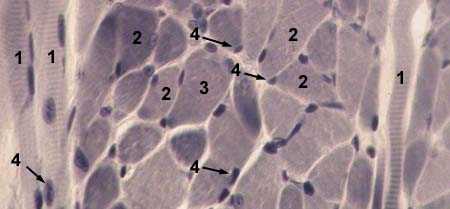 |
| СЕРДЕЧНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ (МИОКАРД) продольный срез Окраска железным гематоксилином 1 — ядра кардиомиоцитов |  |
| ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ продольный срез Окраска гематоксилин-эозином | 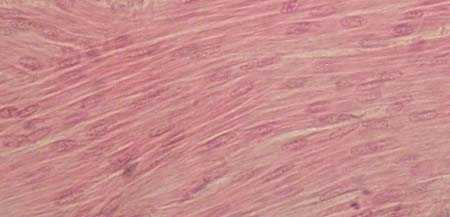 |
| ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ продольный срез Окраска гематоксилин-эозином | 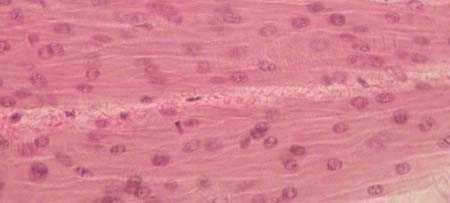 |
histol.ru
Лекция 09. Мышечные ткани
Лекция 09. Мышечные тканиОфициальный план лекции:
Тема 2.6. Мышечные ткани.
- Место ибиологическое значение мышечных тканей в организации тела человека и животных.
- Морфологические критерии и принципы организации мышечных тканей: виды. Классификации мышечных тканей.
- Особенности морфологического строения скелетных мышечных тканей. Строение поперечно-полосатого мышечного волокна. Сократительный аппарат. Иннервация мышечного волокна и особенности взаимодействия сократительных белков. Теория «скользящих нитей» Хаксли. Гистогенез. Регенерация.
- Строение сердечной мышечной ткани. Особенности регенерации. Гистогенез.
- Строение гладкой мышечной ткани. Особенности организации сократительного аппарата. Иннервация. Регенерация. Гистогенез.
Материалы по теме лекции:
Содержание:
Функции:
- Сокращение
- Участие в движении
Способности
- Сокращение и расслабление
- Генерация силы
Классификации
Морфологическая классификация
- Поперечно-полосатая (поперечно-исчерченная)
- Гладкая (неисчерченная)
Классификация по локализации
- Скелетная
- Внутренностная
- Сердечная
Гистогенетическая классификация (по Н.Г. Хлопину)
- Гладкие мышечные ткани
- Висцерального типа
- Мионейрального типа
- Миоэпителиального типа (часть морфологов не выделяют данный вид мышечных тканей, считая миоэпителиальные клетки специфическими клетками эпителия – миоидными клетками эпителия)
- Поперечно-полосатая мышечная ткань соматического типа
- Поперечно-полосатая мышечная ткань целомического типа
|
||||||||||||||||||||
Миоидные клетки
Миоидные клетки – сократимые элементы немышечных тканей
- Миоэпителиальные клетки
- Миофибробласты
- Миоидные клетки стенки извитых семенных канальцев
Структурная организация мышечных тканей
-
Гладкая мышечная ткань висцерального (внутренностного) типа
- Гладкий миоцит – одноядерная клетка вытянутой веретеновидной формы
- Поперечно-полосатая мышечная ткань соматического (скелетного) типа
- Мышечные волокна, основу которых составлчют миосимпласты – многоядерные структуры, которые образуются при слиянии одноядерных клеток миобластов. Кроме миосимпластов в состав мышечных волокон входят и клетки – миосателлитоциты, они образуют камбиальный резерв мышечной скелетной ткани.
-
Поперечно-полосатая мышечная ткань целомического (сердечного) типа
- Кардиомиоцит – сердечная мышечная клетка, одноядерная или двуядерная. Кардиомиоциты соединяются торцевыми участками в функциональные сердечные мышечные волокна, однако эти волокна имеют клеточную структуру. Функциональные волокна анастомозируют боковыми поверхностями
Классификация по типу контроля мышечной актичности
- Поперечно-полосатая мышечная ткань скелетного типа – соматическая нервная система, возможна сознательная регуляция.
- Гладкая мышечная ткань внутренних органов – вегетативная (автомномная) нервная система, в большинстве случаев не контролируется сознательно.
- Поперечно-полосатая мышечная ткань сердечного типа – вегетативная иннервация до 4 нд внутриутробного развития. Затем регуляция собственной проводящей системой сердца. Водители ритма (пейсмекеры). Вегетативная нервная система контролирует деятельность водителей ритма.
Гистологическая структура скелетной мышцы как органа
- Пучки мышечных волокон
- Соединительнотканный футляр – Эпимизий
- Прослойки соединительной ткани
- Эндомизий – прослойки между волокнами
- Перимизий – прослойки между пучками волокон
- Сухожилие
- Иннервация
- Нервномышечные синапсы – моторные бляшки
Поперечно-полосатая мышечная ткань скелетного типа
- Структурная единица – мышечное волокно
- Мышечное волокно
- Миосимпласт
- Оболочка – сарколемма
- плазмолемма
- базальная мембрана
- Ядра
- Саркоплазма
- Исчерченность саркоплазмы – проявление упорядоченной структуры миофибрилл
- Органеллы
- Миофибриллы – специальные сократительные органеллы мышечных волокон
- Митохондрии
- Саркоплазматический ретикулум
- Включения
- Гранулы гликогена
- Миоглобин – железосодержащий белок, акцептирующий кислород
- Оболочка – сарколемма
-
Миосателлитоциты – камбиальные, малодифференцированные клетки, принимающие участие в репаративной регенерации мышечных волокон
- Миосимпласт
- Мышечное волокно
- Миофибриллы (см. схему организации миофибриллы)
- Саркомеры
- Телофрагмы – границы саркомеров
- Миофиламенты (нити)
- Цитоскелетные белки
- Упорядоченное расположение миофиламентов в саркомере
- Сопоставление ультроструктурных и светооптическими данных
- В составе саркомера:
- две половины двух I – дисков, только актин
- Z -линии – телофрагмы (а-актинин, винкулин, десмин)
- две половины двух I – дисков, только актин
- A-диски
– зона перекрытия актиновых и миозиновых филаментов
- H-полоска – только миозин
- M-линия – мезофрагма (зона фиксации толстых миозиновых филаментов
- В составе саркомера:
- Саркомеры
- Механизм мышечного сокращения
- Передача сигнала к сокращению
- Волна деполяризации плазмолеммы
- Триады
и передача сигнала на саркоплазматическую сеть
- Т-канальцы
- Саркоплазматический ретикулум – депо Са++
- Цистерны
- Канальцы
- Выброс Са++ в саркоплазму
- Взаимодействие тонких и толстых миофиламентов
- Сократимые белки и взаимодействие между ними
- Гипотеза скользящих нитей
- Потребности в: АТФ и Са++
- Сократимые белки и взаимодействие между ними
- Укорочение саркомеров и сокращение мышечного волокна
- Расслабление
- Классификация мышечных волокон
- Мышечная сила
- Зависит от степени развития мышцы.
- Мышечные волокна закладываются до рождения
- Гипертрофия (увеличение объема мышцы)
- Увеличение объема существующих мышечных волокон
- Увеличение количества миофибрилл
- Увеличение объема существующих мышечных волокон
- Наследственная мышечная дистрофия. Болезнь Вердинга-Гофмана
- Методы дифференциального окрашивания
- Метод Маллори
- Метод Ван-Гизона
- Метод гистохимического выявления Сукцинатдегидрогеназы
- Эмбриональный гистогенез поперечно-полосатой мышечной ткани соматического типа
- Миобластический период.
Направление дифференцировки:- Симпластическое
- Миосаттелитоциты
- Симпластическая стадия
- Образование мышечных трубочек
- Формирование мышечных волокон
- Миобластический период.
!!! Строение гладкой мышечной ткани и сердечной поперечно-полосатой мышечной ткани на лекции не рассматривалось. Изучить самостоятельно.
Кардиогенез. Генетический контроль
Дополнительные источники информации по теме Мышечные ткани
- Трифонов Е.В. Психофизиология человека Толковый русско-английский словарь
- Трифонов Е.В. МИОЛОГИЯ: РЕСУРСЫ ИНТЕРНЕТ [myology: resources of the Internet]
- John W. Kimball. Muscles. In: Kimball’s Biology Pages.
Мышцы.
В руководстве: «Страницы биологии д-ра Кимбалла» - On-Line Biology Book: Muscular and Skeletal Systems. In: M.J. Farabee. On-Line Biology Book.
Мышечная и скелетная система. В руководстве «Биология». - Michael W. King, Ph.D.: Muscle Biochemistry. In: Michael W. King, Ph.D. Medical Biochemistry. Terre Haute Center for Medical Education.
Биохимия мышцы. В руководстве «Медицинская биохимия».
histology.narod.ru
3 вида мышечной ткани, симпласты, строение и белки миофибриллы
Вы моргнули, повернули голову, вздохнули, посмотрели вдаль, что-то сказали. Каждую минуту в вашем организме сокращается множество мышц тела. Добавьте к этому то, что сердце бьется, в животе урчит, мочеточник мягко препровождает мочу от почки к мочевому пузырю, а сосуды постоянно поддерживают определенное артериальное давление. Древние говорили: «In motu vita est», что значит «В движении жизнь».
Гистологи выделяют 3 вида мышечной ткани: поперечно-полосатую скелетную, поперечно-полосатую сердечную и гладкую. В основных своих чертах они похожи, но именно нюансы, именно тонкости их разнят до неузнаваемости. Поперечно-полосатая скелетная мышечная ткань образует те самые мышцы, которые переставляют ваши ноги, протягивают вашу руку за чашечкой кофе, сгибают и выпрямляют ваше тело. Если заглянуть в окуляр светового микроскопа, то вы не увидите клеток (картинка I). Действительно, мы на 40% состоим не из клеток (ведь приблизительно столько приходится на массу скелетной мускулатуры). Когда-то на этом месте находились клетки, но в те времена мы еще были эмбрионами. А по мере роста и развития цитоплазма их сливалась (рис. 24), обтягиваясь единой мембраной — сарколеммой (4), ядра (3) становились общими, образовывались длинные многоядерные трубчатые волокна — симпласты (1), из которых и состоит поперечно-полосатая скелетная ткань в конечном варианте.
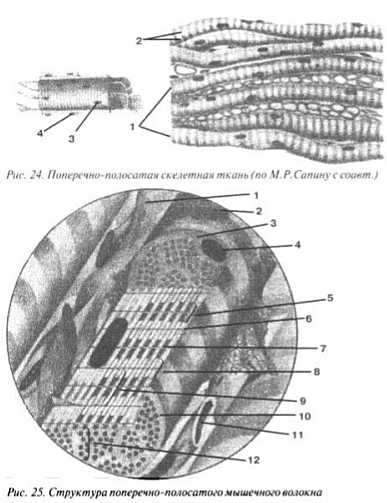
Картинка I. Поперечно-полосатая мышца
Кроме того, под световым микроскопом совершенно отчетливо видно, что название себя прекрасно оправдывает: поперек волокна, чередуя друг друга, располагаются темные и светлые полосы (2). Чтобы лучше рассмотреть, стоит увеличим сильнее симпласт. Схематично он изображен на рис 25. В цитоплазме (3) непосредственно под тонкой сарколеммой (2) расположены вытянутые ядра (4). Соседние мышечные волокна «переслоены» соединительной тканью — эндомизием (1) и многочисленными сосудиками (11). Оказывается, расчерчена не вся цитоплазма. В нее погружены протянутые вдоль всего симпласта многочисленные белковые полоски — миофибриллы (10). Между ними никакой «полосатости» нет: их параллельные пучки (12) окружены митохондриями, эндоплазматической сетью и некоторыми другими органеллами.
Теперь внимательнее рассмотрим строение миофибриллы, например, нижнюю на схеме: куча всяких полосок. Как-нибудь их обозначим для ясности. Толстый светлый промежуток, поделенный пополам тонкой линией, называется I-диском (8), а линия обозначается буквой Z (так называемая Z-линия —6). Два расположенных рядом темных столбика объединяют в А-диск (5), а между ними хорошо видно светлую Н-полоску (7). Участок между рядом расположенными Z-линиями носит название — саркомер (9), который и стоит изучать под электронным микроскопом, чтобы наконец разобраться во всех этих дисках, полосках и линиях (рис. 26, а).
Белки миофибриллы представлены двумя сократительными белками. Более тонкие нити белка актина держатся параллельно друг другу, скрепляясь вместе плоской пластинкой, которая и была названа гистологами Z-линией. Актин не способен преломлять свет дважды, и это качество гистологи решили назвать изотропностью. Стоит запомнить, что изотропность и делает участок около Z-линии светлым, а это не что иное, как I-диск. Другой белок называется миозином. Он толще, представительнее и, что привело в восторг мировую физическую общественность, преломляет дважды пучок проходящего через него света, становясь темнее. Это свойство называется анизотропностью, а отсюда и название — А-диск. Нити белка актина и белка миозина взаимно проникают друг в друга. Средняя часть миозиновой «стопки» свободна от контакта с двумя актиновыми, что делает ее чуть более светлой, чем обе зоны взаимопроникновения — это Н-полоса.
Ну и, наконец, как же это все действует? Всё начинается с поступления сигнала, который говорит о необходимости сокращения определённого симпласта, при этом митохондрии выбрасывают необходимое количество энергии, а на миофибриллы из эндоплазматической сети «высыпаются» ионы кальция. Высвобождение ионов запускает биохимическую реакцию, результатом которой становится то, что нити актина проникают глубже между нитями миозина (рис. 26, б). Z-линии как бы сдвигаются из-за сужения Н-полосы. Подобное укорочение всех саркомеров и приводит собственно к укорочению всей мышцы, то есть ее сокращению. Эту мышечную ткань называют еще поперечно-полосатой произвольной, так как мы сами решаем, какую мышцу «побеспокоить» на этот раз. Этого нельзя сказать о поперечно-полосатой сердечной (или непроизвольной) мышечной ткани, строящей миокард.
Последний вид мышц заложен во внутренних органах и сосудах. Гладкая мышечная ткань представлена клетками — миоцитами (рис. 27). Они имеют вытянутую веретенообразную форму. В каждой клетке расположено одно (редко два) ядро. Оно было создано приспособленным к назойливому желанию мышечной клетки почему-то все время сокращаться. В результате ядра миоцитов научились не отставать от хозяев и вместе с ними сжимаются, укорачиваются и даже пружинисто скручиваются вокруг своей оси. В цитоплазме также находятся миозиновые и актиновые нити, однако они не уложены в стройные миофибриллы. Достаточно беспорядочные, они образуют как бы паутину, заполняющую клетку изнутри, однако в целом принцип работы остается прежним (картинка II).
Картинка II. Гладкая мышечная ткань
Сокращение гладкого миоцита происходит относительно медленно и непроизвольно от нас. Кишечник, сосуды, мочеточник, как бы не спеша, помогают своими движениями прохождению по ним различных образований, будь то кровь или пищевая кашица. Но есть в организме гладкие миоциты «быстрого реагирования»: они складывают мышцы радужки глаза. Именно благодаря этим мышцам зрачок столь стремительно проявляет реакцию на свет (расширяясь или сужаясь).
tardokanatomy.ru
Виды мышечных тканей и их особенности
В статье мы рассмотрим виды мышечных тканей. Это очень важная тема в биологии, ведь каждый должен знать, как функционируют наши мышцы. Они представляют собой сложную систему, изучение которой, надеемся, вам будет интересно. А помогут лучше представить себе виды мышечной ткани картинки, которые вы найдете в этой статье. Прежде всего, дадим определение, которое необходимо при изучении данной темы.
Мышечная ткань — это особая группа тканей человека и животных, основной функцией которой является ее сокращение, обусловливающее перемещение организма или составляющих его частей в пространстве. Данной функции соответствует строение основных элементов, из которых состоят различные виды мышечных тканей. Элементы эти имеют продольную и удлиненную ориентацию миофибрилл, включающих в свой состав сократительные белки — миозин и актин. Мышечная ткань, как и эпителиальная, это сборная тканевая группа, так как основные ее элементы развиваются из эмбриональных зачатков.
Сокращение мышечной ткани
Клетки ее, так же как и нервные, при воздействии электрических и химических импульсов могут возбуждаться. Способность их сокращаться (укорачиваться) в ответ на действие того или иного стимула связана с наличием миофибрилл, особых белковых структур, каждая из которых состоит из микрофиламентов, коротких белковых волокон. В свою очередь, они подразделяются на миозиновые (более толстые) и актиновые (тонкие) волокна. В ответ на нервное раздражение сокращаются различные виды мышечных тканей. Сокращение к мышце передается по нервному отростку через нейромедиатор, которым является ацетилхолин. Мышечные клетки в организме осуществляют энергосберегающие функции, так как расходуемая при сокращении различных мышц энергия выделяется затем в виде тепла. Именно поэтому, когда организм подвержен охлаждению, возникает дрожь. Это не что иное, как частые сокращения мышц.
Можно выделить следующие виды мышечных тканей, в зависимости от того, какое строение имеет сократительный аппарат: гладкую и поперечнополосатую. Они состоят из отличающихся по строению гистогенетических типов.
Мышечная ткань поперечнополосатая
Клетки миотомов, которые образуются из дорсальной мезодермы, являются источником ее развития. Эта ткань состоит из удлиненных мышечных волокон, имеющих вид цилиндров, концы которых заострены. 12 см в длину и 80 мкм в диаметре достигают эти образования. Симпласты (многоядерные образования) содержатся в центре мышечных волокон. Снаружи к ним прилегают клетки под названием «миосателлиты». Сарколеммой ограничены волокна. Она образуется плазмолеммой симпласт и базальной мембраной. Под базальной мембраной волокна располагаются миосателлиотоциты — так, что плазмолеммы симпласт касается их плазмолемма. Данные клетки являются камбиальным резервом мышечной скелетной ткани, и именно за счет него осуществляется регенерация волокон. Миосимпласты, кроме плазмолеммы, включают в себя также саркоплазму (цитоплазму) и расположенные по периферии многочисленные ядра.
Значение поперечнополосатой мышечной ткани
Описывая виды мышечной ткани, следует отметить, что поперечнополосатая является исполнительным аппаратом всей двигательной системы. Она формирует скелетные мышцы. Кроме того, этот вид ткани входит в структуру внутренних органов, таких как глотка, язык, сердце, верхний отдел пищевода и др. Общая масса ее у взрослого человека составляет до 40% от массы тела, а у пожилых людей, а также новорожденных, ее доля — 20-30%.
Особенности поперечнополосатой мышечной ткани
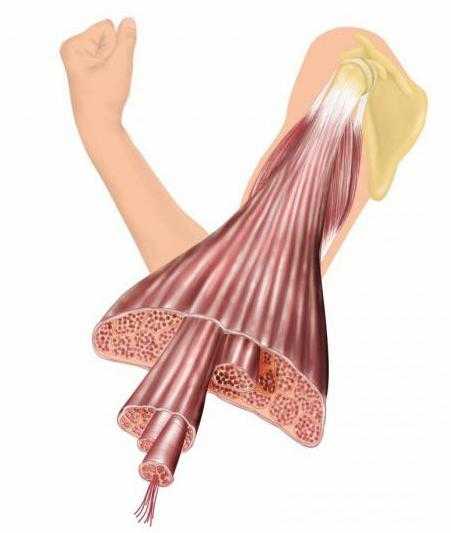
Сокращение данного вида мышечной ткани, как правило, можно производить с участием сознания. Она обладает несколько большим быстродействием по сравнению с гладкой. Как вы видите, виды мышечной ткани отличаются (о гладкой мы поговорим совсем скоро и отметим некоторые другие различия между ними). В поперечнополосатых мышцах нервные окончания воспринимают информацию о текущем состоянии мышечной ткани, а затем передают ее по афферентным волокнам в нервные центры, ответственные за регуляцию двигательных систем. Управляющие функциями мышц сигналы поступают от регуляторов в виде нервных импульсов по двигательным или вегетативным эфферентным нервным волокнам.
Гладкая мышечная ткань
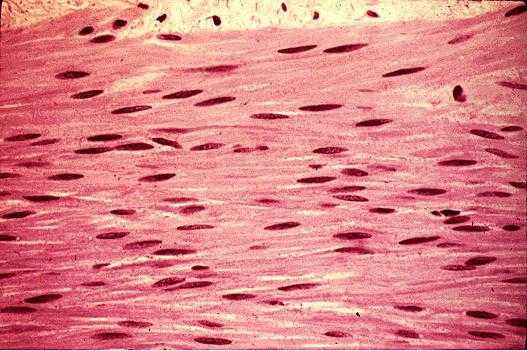
Продолжая описывать виды мышечных тканей человека, переходим к гладкой. Она формируется веретенообразными клетками, длина которых составляет от 15 до 500 мкм, а диаметр находится в промежутке от 2 до 10 мкм. В отличие от волокон мышцы поперечнополосатой, эти клетки имеют одно ядро. Кроме того, у них нет поперечной исчерченности.
Значение гладкой мышечной ткани
От сократительной функции этого вида мышечной ткани зависит функционирование всех систем организма, поскольку она входит в структуру каждой из них. Так, например, гладкая мышечная ткань участвует в управлении диаметром дыхательных путей, кровеносных сосудов, в сокращении матки, мочевого пузыря, в реализации двигательных функций нашего пищеварительного тракта. Она управляет диаметром зрачка глаз, а также участвует во множестве других функций различных систем организма.
Мышечные слои
Мышечные слои образует этот вид ткани в стенках лимфатических и кровеносных сосудов, а также всех полых органов. Обыкновенно это два или три слоя. Толстый циркулярный — наружный слой, средний присутствует не обязательно, тонкий продольный — внутренний. Питающие мышечную ткань кровеносные сосуды, а также нервы проходят параллельно оси мышечных клеток между их пучками. Гладкомышечные клетки можно разделить на 2 типа: унитарные (объединенные, сгруппированные) и автономные миоциты.
Автономные миоциты
Автономные функционируют довольно независимо друг от друга, так как нервным окончанием иннервируется каждая такая клетка. Они были обнаружены в мышечных слоях крупных кровеносных сосудов, а также в ресничной мышце глаза. Также к данному типу относятся клетки, из которых состоят мышцы, поднимающие волосы.
Унитарные миоциты
Унитарные мышечные клетки, напротив, тесно между собой переплетаются, так что мембраны их могут не просто примыкать плотно друг к другу, образуя десмосомы, но также и сливаться, формируя нексусы (щелевые контакты). Пучки образуются в результате данного объединения. Диаметр их составляет около 100 мкм, а длина достигает нескольких мм. Они формируют сеть, и в ее ячейки вплетаются коллагеновые волокна. Волокнами вегетативных нейронов иннервируются пучки, и они становятся функциональными единицами гладкой мышечной ткани. Деполяризация при возбуждении одной клетки пучка распространяется очень быстро на соседние, поскольку мало сопротивление щелевых контактов. Состоящие из унитарных клеток ткани есть в большинстве органов. К ним относятся мочеточники, матка, пищеварительный тракт.
Сокращение миоцитов
Сокращение миоцитов обусловлено в гладкой ткани, как и в поперечнополосатой, взаимодействием миозиновых и актиновых нитей. В этом схожи различные виды мышечной ткани у человека. Данные нити распределены внутри миоплазмы менее упорядоченно, чем в мышце поперечнополосатой. С этим связано отсутствие поперечной исчерченности в гладкой мышечной ткани. Внутриклеточный кальций является конечным исполнительным звеном, управляющим взаимодействием миозиновых и актиновых нитей (то есть сокращением миоцитов). Это же относится и к поперечнополосатой мышце. Однако детали механизма управления существенно отличаются от последней.
Проходящие в самой толще мышечной гладкой ткани вегетативные аксоны формируют не синапсы, что характерно для ткани поперечнополосатой, а многочисленные утолщения, имеющиеся по всей длине, которые и играют роль синапсов. Утолщения выделяют медиатор, который диффундирует к расположенным рядом миоцитам. Рецепторные молекулы находятся на поверхности этих миоцитов. С ними медиатор и взаимодействует. Он вызывает деполяризацию у миоцита внешней мембраны.
Особенности гладкой мышечной ткани
Нервная система, ее вегетативный отдел, управляется без участия сознания работой гладких мышц. Мышцы мочевого пузыря являются единственным исключением. Управляющие сигналы либо непосредственно реализуются, либо опосредованно — через гормональные (химические, гуморальные) воздействия.
Энергетические и механические свойства данного вида мышечной ткани обеспечивают поддержание тонуса (управляемого) стенок полых органов и сосудов. Связано это с тем, что гладкая ткань функционирует эффективно, не требуется больших затрат АТФ. У нее меньшее быстродействие, чем у мышечной ткани поперечнополосатой, однако она способна сокращаться более продолжительное время, кроме того, может развивать существенное напряжение и изменять в широких пределах свою длину.
Итак, мы рассмотрели виды мышечных тканей и особенности их структурной организации. Конечно, это лишь основная информация. Можно долго описывать виды мышечных тканей. Рисунки помогут вам наглядно их представить.
fb.ru