
Врожденные заболевания обмена веществ | Сант Жоан де Деу
Что такое наследственные болезни обмена веществ (врожденные метаболические заболевания)
Патологии, также известные как врожденные нарушения метаболизма, — это заболевания, причина которых кроется в генетическом изменении белка или фермента, в результате чего блокируется определенный процесс метаболизма. Такая блокировка влияет на нормальное функционирование некоторых клеток и органов и проявляется рядом симптомов, различных у каждого пациента. Среди таких симптомов могут встречаться разные виды неврологических синдромов.
Эта группа патологий очень обширна, однако ее можно систематизировать с помощью действующей классификации, которая в данный момент претерпевает значительные изменения ввиду того, что сегодня мы располагаем гораздо большими знаниями о базовых механизмах развития таких патологий. Ниже приведены основные группы патологий, составленные на основании типа поражения организма при каждой из них.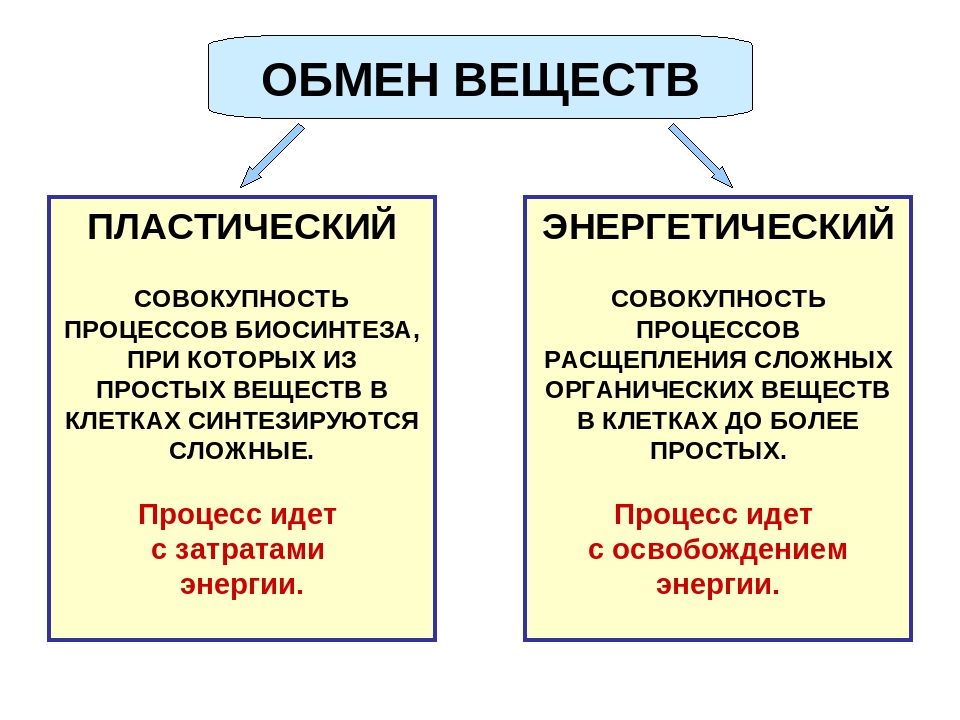
Врожденное нарушение метаболизма малых молекул
Влияют на промежуточный метаболизм. Сюда входят аминоацидопатии (фенилкетонурия и пропионовая ацидурия). Также сюда входит нарушения обмена углеводов или нейромедиаторов и нейромодуляторов.
Врожденное нарушение энергетического обмена
Характеризуется недостаточной выработкой и использованием энергии. Сюда входят митохондриальные заболевания, недостаточная выработка пирувата или глюкозы (в мышцах или печени) и т. д.
Врожденное нарушение метаболизма сложных молекул
Группа заболеваний, которые препятствуют синтезу больших молекул. Они проявляются в виде постоянных симптомов, не связанных с питанием. Сюда входят лизосомные (мукополисахаридоз, олигосахаридоз, сфинголипидоз и т. д.), пероксисомальные (синдром Цельвегера, адренолейкодистрофия, сцепленная с хромосомой Х) заболевания и врожденные нарушения гликозилирования, а также другие врожденные нарушения метаболизма.
Энергетический метаболизм прокариот: брожение и дыхание
В «Сириусе» участники проектной образовательной программы «Большие вызовы» продолжают работать в лабораториях Парка науки и искусства над проектами, предложенными ребятам в рамках профильных направлений. С лекциями перед юными исследователями ежедневно выступают ведущие ученые страны.
С лекциями перед юными исследователями ежедневно выступают ведущие ученые страны.
Накануне участники направления «Агропромышленные и биотехнологии» встречались с кандидатом биологических наук, доцентом кафедры генетики, микробиологии и биотехнологии биологического факультета КубГУ Андреем Александровичем Самковым. Педагог разбирал тему «Энергетический метаболизм прокариот».
– На лекции мы рассмотрели способы извлечения, трансформации и накопления энергии в живых клетках на примере прокариот, а также разобрались, как клетки используют эту энергию в процессах своего метаболизма. Рассмотрены типы фосфорилирования (окислительного и субстратного), отвечающих за брожение и дыхание, – рассказывает педагог.
Брожение – это внутренний окислительно-восстановительный процесс, при котором акцептором электронов служит органическая молекула, и суммарная степень окисления образующихся продуктов отличается от степени окисления сбраживаемого вещества. Дыхание – это окислительный распад органических веществ при участии кислорода с образованием воды, углекислого газа и макроэргических соединений, которые используются клетками.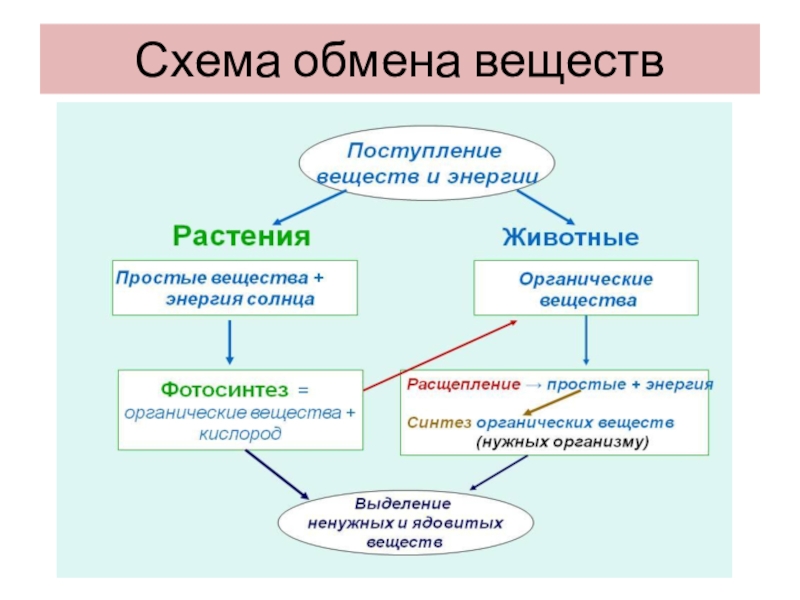
– Моему проекту информация о получении энергии на примерах и прокариот (доядерных организмов), и эукариот (ядерных организмов), очень пригодится, – говорит Иван Венчиков из Ижевска. В завершении образовательной программы школьник вместе с остальными участниками проектной группы представит экспертам биоплато из растений и микроорганизмов для очистки воды от поступающих в нее загрязняющих веществ. Одновременно оно будет обладать и функциями источника возобновляемой энергии, используемой для электропитания датчиков агро- и экомониторинга среды, уточняют ребята.
(PDF) Оценка наследуемости и повторяемости уровня метаболизма покоя птиц на примере свободноживущих мухоловок-пеструшек Ficedula hypoleuca (Aves: Passeriformes) [Estimation of heritability and repeatability of resting metabolic rate in birds by the example of free-living pied flycatchers Ficedula hypoleuca (Aves: Passeriformes)] In Russian
ЖУРНАЛ ОБЩЕЙ БИОЛОГИИ том 71 № 5 2010
404 БУШУЕВ и др.
популяциях ввиду средовых и/или генетических
отличий, поэтому величина повторяемости и на-
следуемости является популяционной характери-
стикой (Versteegh et al., 2008).
Несмотря на то что BMR и RMR являются наи-
более исследованными энергетическими характе-
ристиками гомойотермных животных, число ра-
бот, посвященных изучению их наследуемости,
довольно ограничено. В основном эти исследова-
ния проведены на человеке (Fontaine et al., 1985;
Bogardus et al., 1986; Ravussin, 1993; Rice et al.,
1996; Luke et al., 2000; Pérusse et al., 2001 и др.) и
на лабораторных линиях грызунов (Lacy, Lynch,
1979; Lynch, Sulzbach, 1984; Johnson, Speakman,
2000, 2007; Dohm et al., 2001; Nespolo et al., 2003a,
2005; Bacigalupe et al., 2004; Książek et al., 2004;
Labocha et al., 2004; Sadowska et al., 2005, 2007;
Wone et al., 2009). В большинстве работ по гры-
зунам значимый уровень наследуемости метабо-
лизма покоя выявлен не был. Нам удалось обна-
Нам удалось обна-
ружить только 4 работы на млекопитающих из
природных популяций (1 – на рыжей полевке, 2 –
на пашенной полевке и 1 – на ласке), в которых
подтверждалось существование значимой насле-
дуемости RMR (Johnson, Speakman, 2000, 2007;
Sadowska et al., 2007; Szafrańska et al., 2007b).
Следует отметить, что только наследуемость
RMR ласки была оценена в популяции свободно-
живущих животных. В случае же с мышевидны-
ми грызунами показатель h3 (lim 0.40–0.44) был
получен в череде поколений, выращенных в лабо-
раторных условиях, хотя их предки были взяты из
диких популяций. В условиях лаборатории можно
легко прослеживать родословную каждой особи и
контролировать множество факторов среды, кото-
рые сильно влияют на энергетику животных в ес-
тественных условиях (температура, доступность
еды, фотопериод, брачная активность и другие
социальные взаимодействия). Стабильность сре-
довых факторов в лаборатории приводит к тому,
что оценки наследуемости признаков, сделанные
у животных в лабораторных условиях и в дикой
природе, обычно очень различаются (Riska et al. ,
,
1989; Simons, Roff, 1994; Weigensberg, Roff, 1996;
Hoffman, Merilä, 1999; Blanckenhorn, 2002).
К числу немногих работ, в которых была оце-
нена наследуемость BMR птиц, относятся ис-
следование на 3 поколениях размножавшихся в
вольерах зебровых амадин (h3 = 0.041; Rønning
et al., 2007), а также недавнее исследование груп-
пы ученых из разных стран, проведенное на раз-
множавшихся в неволе черноголовых чеканах
трех подвидов из трех диких популяций. В зави-
симости от размерности BMR, популяционной
принадлежности и метода расчета h3 = 0.20–0.69
(Tieleman et al., 2009a,b). Публикации по насле-
дуемости энергетических параметров свободно-
живущих птиц исчерпываются нашими работа-
ми по мухоловке-пеструшке (Bushuev et al., 2005;
Бушуев, 2009) и исследованием лазоревок швед-
скими орнитологами (Nilsson et al., 2009). Для
обоих видов в качестве энергетической характе-
ристики потомков использовали RMR птенцов на
асимптотической стадии роста, т. е. тогда, когда
е. тогда, когда
у птенцов завершался рост массы и скелета, но
еще продолжало формироваться оперение. Соот-
ветственно RMR таких птенцов помимо расхода
энергии на BMR включал затраты на продуктив-
ные процессы (PE), в основном связанные с ро-
стом пера (RMR = BMR + PE). Для лазоревок был
получен довольно высокий показатель аддитив-
ной генетической изменчивости RMR (h3 = 0.59).
Работы по строгой оценке наследуемости уровня
метаболизма ночного покоя свободноживущих
птиц, когда родителей и потомков сравнивают по
дефинитивному состоянию признака, а именно
по BMR (в идеале с учетом сезона, пола и воз-
раста, которые у некоторых видов являются фак-
тором вариации BMR, – см., например, Керимов,
Иванкина, 1999; Broggi, 2006; Broggi et al., 2007),
нам не известны.
Основные работы по исследованию повторяе-
мости и наследуемости энергетических харак-
теристик животных приведены в Приложении
(табл. 1–2).
1–2).
Объектом нашего исследования послужили сво-
бодноживущие мухоловки-пеструшки (Ficedula
hypoleuca Pallas) из двух естественных популя-
ций. Выбор данного вида во многом обусловлен
его редкой особенностью: выраженным полимор-
физмом в брачной окраске самцов (Drost, 1936).
Степень меланизации оперения самцов имеет на-
следственную природу (Lundberg, Alatalo, 1992;
Высоцкий, 1993; Гриньков, 2000), и вариация этого
признака связана с рядом репродуктивных, физио-
логических, морфологических и поведенческих
характеристик (Lundberg, Alatalo, 1992; Ivankina
et al., 2001; Ilyina, Ivankina, 2001; Иванкина и др.,
2007), в том числе и c параметрами энергетики
(Røskaft et al., 1986; Gavrilov et al., 1993; Гаври-
лов и др., 1993, 1996; Ильина, 2004а,б). Оказа-
лось, что ранее обнаруженная зависимость BMR
взрослых самцов от типа окраски практически
идентична зависимости RMR птенцов на стадии
асимптотического роста от типа окраски их отцов
(см. рис. 1; Kerimov, Ivankina, 1999; Bushuev et al.,
рис. 1; Kerimov, Ivankina, 1999; Bushuev et al.,
2003; Бушуев, 2009). Этот результат ставит целый
ряд вопросов: обусловлено ли сходство зависимо-
Страница не найдена |
Страница не найдена |404. Страница не найдена
Архив за месяц
ПнВтСрЧтПтСбВс
3456789
10111213141516
17181920212223
24252627282930
31
12
12
1
3031
12
15161718192021
25262728293031
123
45678910
12
17181920212223
31
2728293031
1
1234
567891011
12
891011121314
11121314151617
28293031
1234
12
12345
6789101112
567891011
12131415161718
19202122232425
3456789
17181920212223
24252627282930
12345
13141516171819
20212223242526
2728293031
15161718192021
22232425262728
2930
Архивы
Фев
Мар
Апр
Май
Июн
Июл
Авг
Сен
Окт
Ноя
Дек
Метки
Настройки
для слабовидящих
Неинфекционные заболевания
Общая информация
Неинфекционные заболевания (НИЗ), также известные как хронические заболевания, как правило, имеют продолжительное течение и являются результатом воздействия комбинации генетических, физиологических, экологических и поведенческих факторов.
К основным типам НИЗ относятся сердечно-сосудистые заболевания (такие, как инфаркт и инсульт), раковые заболевания, хронические респираторные заболевания (такие, как хроническая обструктивная болезнь легких и астма) и диабет.
Бремя НИЗ диспропорционально велико в странах с низким и средним уровнем доходов, на которые приходится более трех четвертей случаев смерти от НИЗ во всем мире (32 миллиона).
Кто подвергается риску таких заболеваний?
НИЗ распространены во всех возрастных группах, всех регионах и всех странах. Эти заболевания часто связывают с пожилыми возрастными группами, но фактические данные свидетельствуют о том, что среди всех случаев смерти, связываемых с НИЗ, 17 миллионов регистрируется в возрастной группе от 30 до 69 лет. При этом более 85% этих случаев «преждевременной» смерти имеет место в странах с низким и средним уровнем доходов. Дети, взрослые и пожилые люди — все уязвимы перед факторами риска, способствующими развитию НИЗ, такими как нездоровое питание, недостаточная физическая активность, воздействие табачного дыма или злоупотребление алкоголем.
Развитию этих заболеваний способствуют такие факторы, как быстрая и неорганизованная урбанизация, глобализация нездорового образа жизни и старение населения. Последствия нездорового питания и недостаточной физической активности могут проявляться у отдельных людей в виде повышенного кровяного давления, повышенного содержания глюкозы в крови, повышенного уровня липидов в крови и ожирения.
Факторы риска
Поддающиеся изменению поведенческие факторы риска
Поддающиеся изменению формы поведения, такие как употребление табака, отсутствие физической активности, нездоровое питание и вредное употребление алкоголя, — повышают риск развития НИЗ.
- От последствий употребления табака каждый год умирает более 7,2 млн. человек (в том числе от последствий воздействия вторичного табачного дыма), и по прогнозам в предстоящие годы эта цифра заметно вырастет . (1)
- Каждый год от последствий чрезмерного потребления соли/натрия умирает 4,1 млн. человек. (1)
- Из 3,3 млн.
 случаев смерти в год в результате употребления алкоголя более половины приходятся на долю НИЗ, в том числе раковых заболеваний . (2)
случаев смерти в год в результате употребления алкоголя более половины приходятся на долю НИЗ, в том числе раковых заболеваний . (2) - 1,6 млн. случаев смерти в год можно признать связанными с недостаточным уровнем физической активности . (1)
 случаев смерти в год в результате употребления алкоголя более половины приходятся на долю НИЗ, в том числе раковых заболеваний . (2)
случаев смерти в год в результате употребления алкоголя более половины приходятся на долю НИЗ, в том числе раковых заболеваний . (2)Метаболические факторы риска
Метаболические факторы риска способствуют развитию четырех основных изменений метаболизма, приводящих к повышению риска НИЗ:
- повышенное кровяное давление
- излишний вес/ожирение
- гипергликемия (высокое содержание глюкозы в крови)
- гиперлипидемия (высокое содержание липидов в крови)
Первым метаболическим фактором риска смерти от НИЗ во всем мире является повышенное кровяное давление (с которым связано 19% всех случаев смерти в мире) (1), за которым следуют избыточный вес, ожирение и повышенное содержание глюкозы в крови.
Каковы социально-экономические последствия НИЗ?
НИЗ ставят под угрозу прогресс на пути осуществления Повестки дня в области развития на период до 2030 г.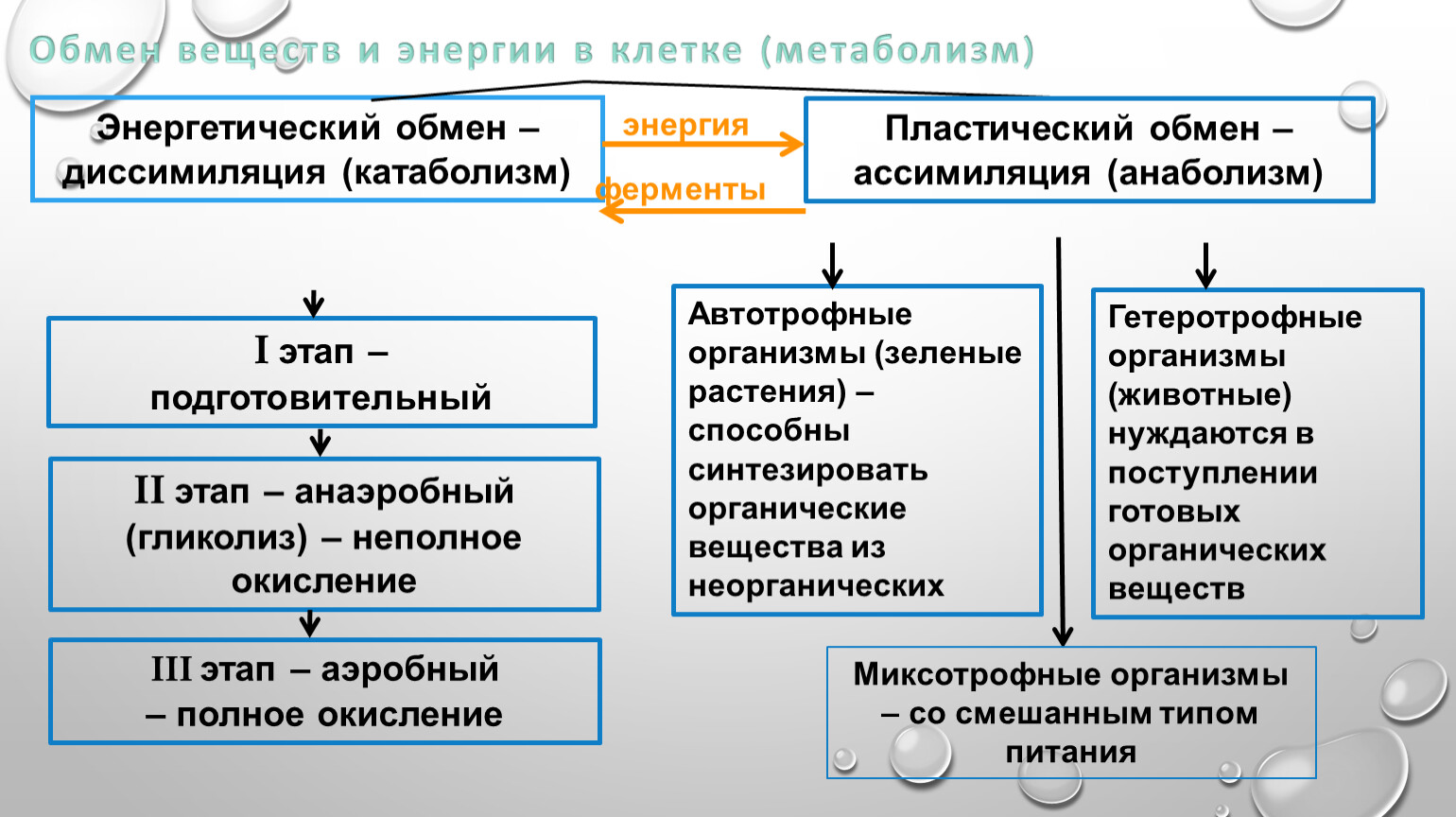 , где предусмотрена цель по снижению числа случаев преждевременной смерти от НИЗ на одну треть к 2030 году.
, где предусмотрена цель по снижению числа случаев преждевременной смерти от НИЗ на одну треть к 2030 году.
Существует тесная связь между бедностью и НИЗ. По прогнозам, стремительный рост заболеваемости НИЗ будет препятствовать реализации инициатив в области сокращения бедности в странах с низким уровнем доходов, в частности, приводя к росту медицинских расходов домохозяйств. Представители уязвимых и незащищенных категорий населения болеют чаще и умирают в более молодом возрасте по сравнению с представителями более благополучных социальных групп, главным образом, в силу своей подверженности более высокому риску воздействия со стороны вредной для здоровья продукции, такой как табак, или склонности к нездоровому питанию, а также ввиду ограниченного доступа к услугам здравоохранения.
В условиях низкой обеспеченности ресурсами медицинские расходы, связанные с НИЗ, приводят к быстрому истощению ресурсов домохозяйств. Каждый год непомерные издержки, связанные с НИЗ, включая расходы, связанные с нередко продолжительным и дорогостоящим лечением, и последствия потери кормильца, ввергают людей в нищету и мешают развитию.
Профилактика НИЗ и борьба с ними
Важным способом борьбы с НИЗ является целенаправленная деятельность по сокращению воздействия факторов риска развития этих болезней. Есть недорогостоящие решения, которыми правительства и другие заинтересованные стороны могли бы воспользоваться для сокращения воздействия поддающихся изменению факторов риска. При выработке мер политики и расстановке приоритетов важное значение имеет мониторинг прогресса и тенденций в области НИЗ.
Для снижения негативного воздействия НИЗ на отдельных людей и общество в целом требуется комплексный подход, в реализации которого должны участвовать все сектора, включая здравоохранение, финансы, транспорт, образование, сельское хозяйство, планирование и другие, работая в сотрудничестве друг с другом над сокращением рисков, связанных с НИЗ, и содействуя осуществлению мероприятий по профилактике этих болезней и борьбе с ними.
Инвестиции в более эффективное ведение НИЗ имеют важнейшее значение. Ведение НИЗ включает выявление, скрининг и лечение этих болезней и предоставление доступа к паллиативной помощи для всех нуждающихся. Высокоэффективные основные мероприятия в отношении НИЗ могут осуществляться на базе первичной медико-санитарной помощи, что позволит укрепить раннее выявление и своевременное лечение. Фактические данные показывают, что при условии их своевременного проведения такие мероприятия с экономической точки зрения являются прекрасной инвестицией, поскольку они могут снизить потребность в более дорогостоящем лечении.
Высокоэффективные основные мероприятия в отношении НИЗ могут осуществляться на базе первичной медико-санитарной помощи, что позволит укрепить раннее выявление и своевременное лечение. Фактические данные показывают, что при условии их своевременного проведения такие мероприятия с экономической точки зрения являются прекрасной инвестицией, поскольку они могут снизить потребность в более дорогостоящем лечении.
Страны с недостаточным охватом медицинским обслуживанием едва ли будут способны обеспечить всеобщий охват основными мероприятиями по лечению и профилактике НИЗ. Такие мероприятия имеют важнейшее значение для достижения поставленной на глобальном уровне цели относительного сокращения риска преждевременной смертности от НИЗ на 25% к 2025 г. и задачи ЦУР по снижению числа случаев преждевременной смерти от НИЗ на одну треть к 2030 году.
Ответные действия ВОЗ
Роль ВОЗ в области общего руководства и координации
В повестке дня в области устойчивого развития на период до 2030 г. НИЗ признаны одним из главных препятствий на пути к устойчивому развитию. В рамках Повестки дня главы государств и правительств взяли на себя обязательство принять на национальном уровне решительные меры с тем, чтобы к 2030 г. сократить на одну треть преждевременную смертность от НИЗ посредством лечебно-профилактической работы (задача 3.4 ЦУР). Эта задача была поставлена по итогам проведения Совещаний высокого уровня Генеральной Ассамблеи ООН по НИЗ в 2011 и 2014 гг., на которых вновь подтверждалась роль ВОЗ в области общего руководства и координации в сфере мониторинга и содействия глобальной работе по борьбе с НИЗ. Генеральная Ассамблея ООН проведет третье Совещание высокого уровня в 2018 г. для анализа хода работы и выработки консенсуса относительно дальнейших действий на период 2018-2030 гг.
НИЗ признаны одним из главных препятствий на пути к устойчивому развитию. В рамках Повестки дня главы государств и правительств взяли на себя обязательство принять на национальном уровне решительные меры с тем, чтобы к 2030 г. сократить на одну треть преждевременную смертность от НИЗ посредством лечебно-профилактической работы (задача 3.4 ЦУР). Эта задача была поставлена по итогам проведения Совещаний высокого уровня Генеральной Ассамблеи ООН по НИЗ в 2011 и 2014 гг., на которых вновь подтверждалась роль ВОЗ в области общего руководства и координации в сфере мониторинга и содействия глобальной работе по борьбе с НИЗ. Генеральная Ассамблея ООН проведет третье Совещание высокого уровня в 2018 г. для анализа хода работы и выработки консенсуса относительно дальнейших действий на период 2018-2030 гг.
Для оказания странам поддержки в их работе на национальном уровне ВОЗ подготовила Глобальный план действий в области профилактики неинфекционных заболеваний и борьбы с ними на 2013-2020 гг. , включающий девять глобальных целей, достижение которых окажет наибольшее влияние на глобальные показатели смертности от НИЗ. Эти цели касаются вопросов профилактики и ведения НИЗ.
, включающий девять глобальных целей, достижение которых окажет наибольшее влияние на глобальные показатели смертности от НИЗ. Эти цели касаются вопросов профилактики и ведения НИЗ.
Справочная литература
(1) GBD 2015 Risk Factors Collaborators. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet, 2016; 388(10053):1659-1724
Сибирский углозуб противостоит полному замораживанию, гипоксии и свободным радикалам
Российские ученые исследовали, как меняется метаболизм хвостатой амфибии сибирского углозуба в ответ на замораживание. В отличие от других земноводных, животное использует в качестве «антифриза» глицерин, а не глюкозу; добывает энергию в ходе метаболических процессов, не требующих кислорода, а также не накапливает вещество, способное привести к появлению свободных радикалов при выходе из условий гипоксии.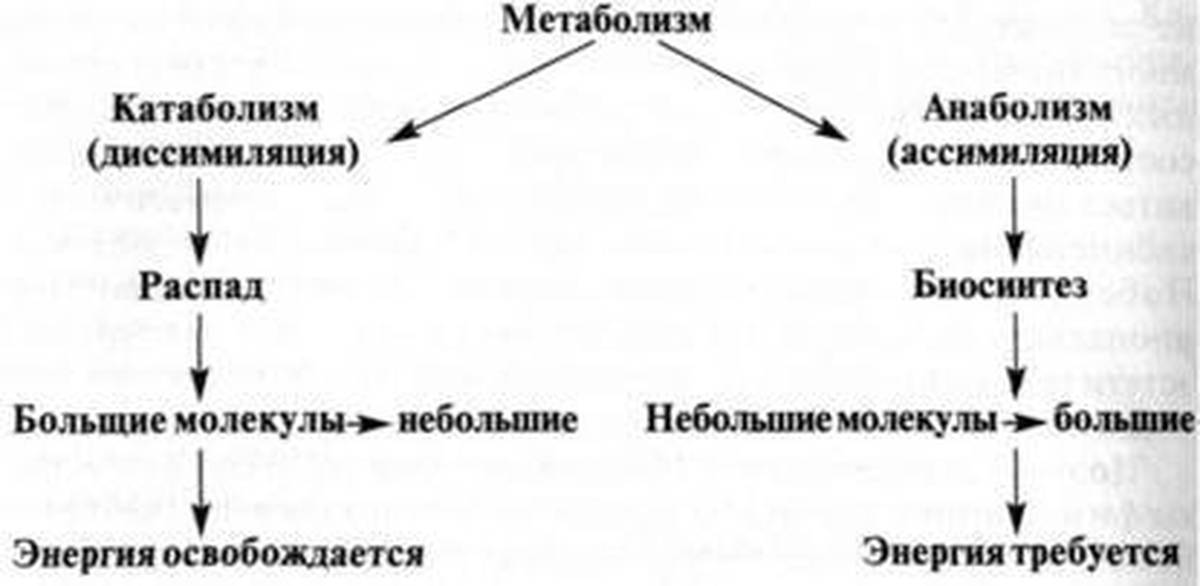 Результаты проекта могут в перспективе дать возможность создания новых методов криоконсервации органов. Исследование выполнено при поддержке гранта Российского научного фонда (РНФ) и опубликовано в журнале Biology.
Результаты проекта могут в перспективе дать возможность создания новых методов криоконсервации органов. Исследование выполнено при поддержке гранта Российского научного фонда (РНФ) и опубликовано в журнале Biology.
Холодные зимы — суровое испытание для животных. Замерзание воды в клетках чревато образованием кристалликов льда, способных разорвать мембраны. Кроме того, заморозка тканей ведет к остановке кровотока и кислородному голоданию — гипоксии. Большинство животных не способно перенести такие состояния и потому выбрало другие стратегии выживания при отрицательных температурах — отрастило мех или мощный жировой слой, изменило размер частей тела, впадает в спячку и прочее. Однако некоторые организмы смогли адаптироваться и к заморозке. Выдающийся пример устойчивости среди них — сибирский углозуб Salamandrella keyserlingii.
«Это уникальная амфибия, способная выдерживать длительное замораживание при температурах до –55°C и оставаться жизнеспособной. Мы проанализировали изменения метаболизма, происходящие при этом в тканях сибирского углозуба», — рассказал руководитель проекта по гранту РНФ Сергей Шеховцов, кандидат биологических наук, старший научный сотрудник ФИЦ «Институт цитологии и генетики» СО РАН.
С 80-х годов прошлого века известно, что защитой от полного замораживания углозуба служит глицерин. Он синтезируется из запасаемого на зиму в печени и мышцах вещества — гликогена — и разносится током крови по всему организму. Глицерин вытесняет из клеток воду, которая кристаллизуется в межклеточном пространстве без ущерба для животного. Сами же клетки не замерзают, что и спасает жизнь всему организму.
Коллектив авторов, включающий в себя ученых из ФИЦ ИЦиГ СО РАН (Новосибирск), Института биологических проблем Севера ДВО РАН (Магадан) и Института «Международный томографический центр» СО РАН (Новосибирск), сравнил метаболомы — наборы малых молекул — углозубов при положительных температурах (3–4 °C) и после продолжительного замораживания (до –8 °C).
Анализ показал, что глицерин — единственный криопротектор среди низкомолекулярных соединений у этого вида. В этом особенность сибирского углозуба, отличающая его от других амфибий, у которых в качестве «антифриза» используется глюкоза.
Также авторы выявили при замораживании усиление гликолиза — процесса окисления веществ для получения энергии без кислорода, — что, очевидно, было следствием гипоксии, возникающей при замораживании. Еще одной уникальной чертой сибирского углозуба оказалось то, что его организм не накапливает сукцинат. Концентрация этого вещества всегда растет при гипоксии у всех изученных на данный момент позвоночных. Сукцинат, в свою очередь, служит причиной синтеза свободных радикалов при возобновлении поступления кислорода, что вызывает различные патологические состояния.
«Все это говорит о том, что метаболизм сибирского углозуба имеет много отличий от метаболизма других животных, что и обеспечивает его выдающуюся устойчивость к замораживанию. В перспективе результаты подобных исследований помогут понять механизмы этой устойчивости и, возможно, дадут ключ к созданию новых способов замораживания органов и тканей», — отметил Сергей Шеховцов.
Демиелинизирующие заболевания нервной системы
Неврологическое отделениеДемиелинизирующие заболевания – аутоиммунные болезни, при которых разрушается миелин белого вещества центральной или периферической нервной системы.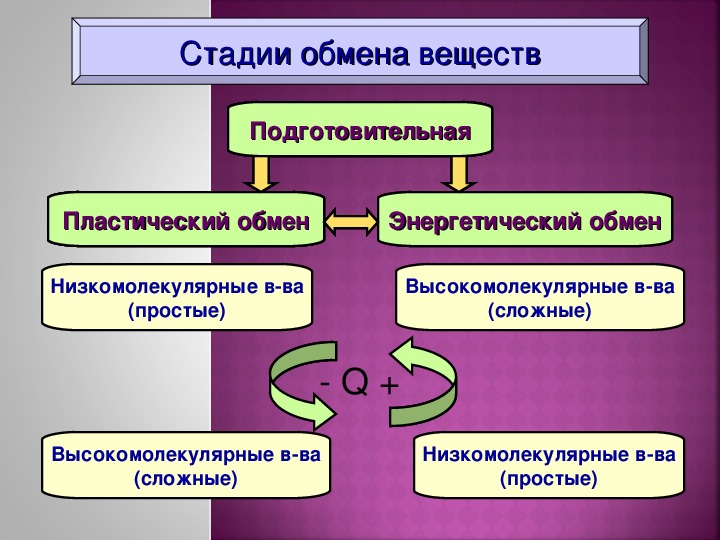 Заболевание вызвано взаимодействием внешних (вирусы, инфекции, интоксикации, особенности диеты, стресс, плохая экология) и наследственных факторов.
Заболевание вызвано взаимодействием внешних (вирусы, инфекции, интоксикации, особенности диеты, стресс, плохая экология) и наследственных факторов.
Наиболее распространенным демиелинизирующим заболеванием является
К демиелинизирующим заболеваниям относятся:
- острый рассеянный энцефаломиелит;
- диффузно-диссеминированный склероз;
- острый оптиконевромиелит, концентрический склероз.
Причины возниконовения:
- Иммунная реакция на белки, которые входят в состав миелина. Эти белки начинают восприниматься иммунной системой как чужеродные и подвергаются ее атаке, в результате чего происходит их разрушение. Это самая опасная причина возникновения заболевания. Толчком к запуску такого механизма может быть инфекция либо врожденные особенности иммунитета: рассеянный энцефаломиелит, рассеянный склероз, синдром Гийена-Барре, ревматические болезни и инфекции в хронической форме.
- Нейроинфекция: некоторые вирусы могут поражать миелин, в результате чего происходит демиелинизация головного мозга.
- Сбой в механизме обмена веществ. Данный процесс может сопровождаться нарушением питания миелина и последующей его гибелью. Это характерно для таких патологий, как заболевания щитовидной железы, сахарный диабет.
- Интоксикация химическими веществами различного характера: алкогольными, наркотическими, психотропными сильного действия, отравляющими веществами, продуктами лакокрасочного производства, ацетоном, олифой, либо отравление продуктами жизнедеятельности собственного организма: перекиси, свободные радикалы.
- Паранеопластические процессы – патологии, которые являются осложнением опухолевых процессов. Последние исследования подтверждают, что в запуске механизма данного заболевания играют важную роль взаимодействия факторов окружающей среды и предрасположенность наследственного характера.


Диагностика демиелинизирующих заболеваний:
Диагноз ставится в случае обнаружения при магнитно-резонансной томографии головного и спинного мозга очагов повышенной интенсивности круглой или овальной формы. Посредством МРТ можно диагностировать развитие атрофии мозга, нарушение проведения импульсов в зрительных путях, стволе мозга и спинном мозге. Деструкцию миелина и аксональную дегенерацию можно выявить при помощи электронейромиографии.Также проводятся иммунологические исследования (высокая концентрация IgG свидетельствует о демиелинизирующем процессе).
Лечение:
Мероприятия по лечению демиелинизирующих заболеваний бывают специфические и симптоматические. Новые исследования в медицине позволили добиться хороших успехов в специфических методах лечения. Бета-интерфероны считаются одними из самых эффективных препаратов: к ним относятся ребиф, бетаферон, аванекс. Клинические исследования бетаферона показали, что его применение снижает на 30% темпы прогрессирования заболевания, предотвращает развитие инвалидности и уменьшает частоту обострений.
Бета-интерфероны считаются одними из самых эффективных препаратов: к ним относятся ребиф, бетаферон, аванекс. Клинические исследования бетаферона показали, что его применение снижает на 30% темпы прогрессирования заболевания, предотвращает развитие инвалидности и уменьшает частоту обострений.Специалисты все чаще отдают предпочтение методу внутривенного введения иммуноглобулинов (биовен, сандоглобулин, веноглобулин). Таким образом проводится лечение обострений данного недуга. Более 20 лет назад была разработана новая, достаточно эффективная методика лечения демиелинизирующих заболеваний – иммунофильтрация ликвора. В качестве средств специфического лечения применяются кортикостероиды, плазмаферез, цитостатики. Также широко применяются ноотропы, нейропротекторы, аминокислоты, миорелаксанты.
Правила госпитализации в стационар
Услуги и цены отделения
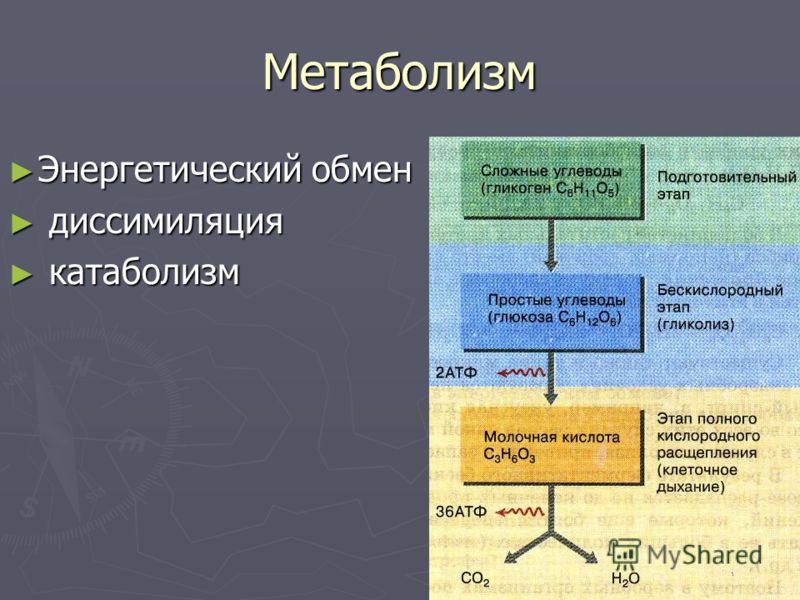
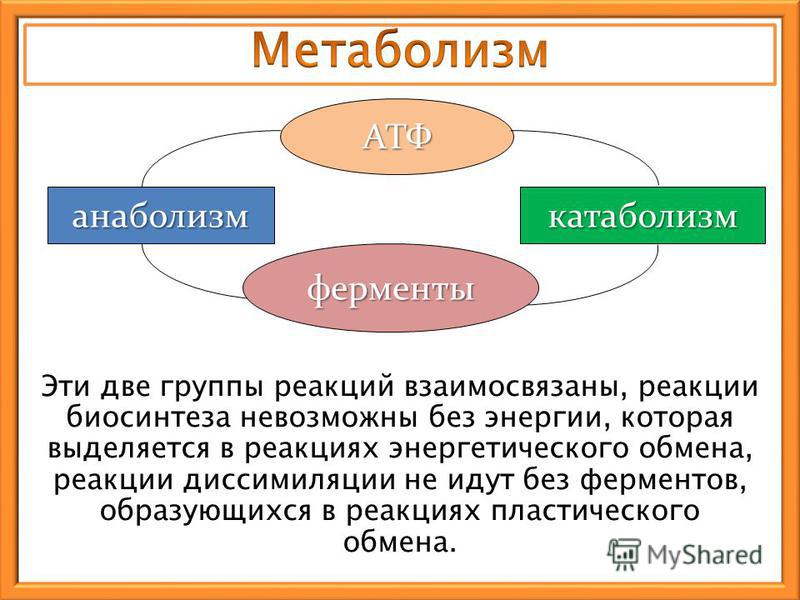
Метаболизм — Принципы биологии
Метаболизм организма — это сумма всех химических реакций, происходящих в организме. Эти химические реакции делятся на две основные категории:
Эти химические реакции делятся на две основные категории:
- Анаболизм: строительные полимеры (большие молекулы, которые нужны клетке).
- Катаболизм: разрушение полимеров с выделением энергии.
Это означает, что метаболизм состоит из синтеза (анаболизма) и деградации (катаболизма) (, рисунок 1, ).
Рисунок 1 Катаболические пути — это те пути, которые генерируют энергию за счет разрушения более крупных молекул. Анаболические пути — это те, которые требуют энергии для синтеза более крупных молекул. Оба типа путей необходимы для поддержания энергетического баланса клетки. Важно знать, что химические реакции метаболических путей не происходят сами по себе. Каждая стадия реакции ускоряется или катализируется белком, называемым ферментом . Ферменты важны для катализирования всех типов биологических реакций — как тех, которые требуют энергии, так и тех, которые выделяют энергию.Вернитесь к главе о ферментах, если вам нужно напоминание по этой теме.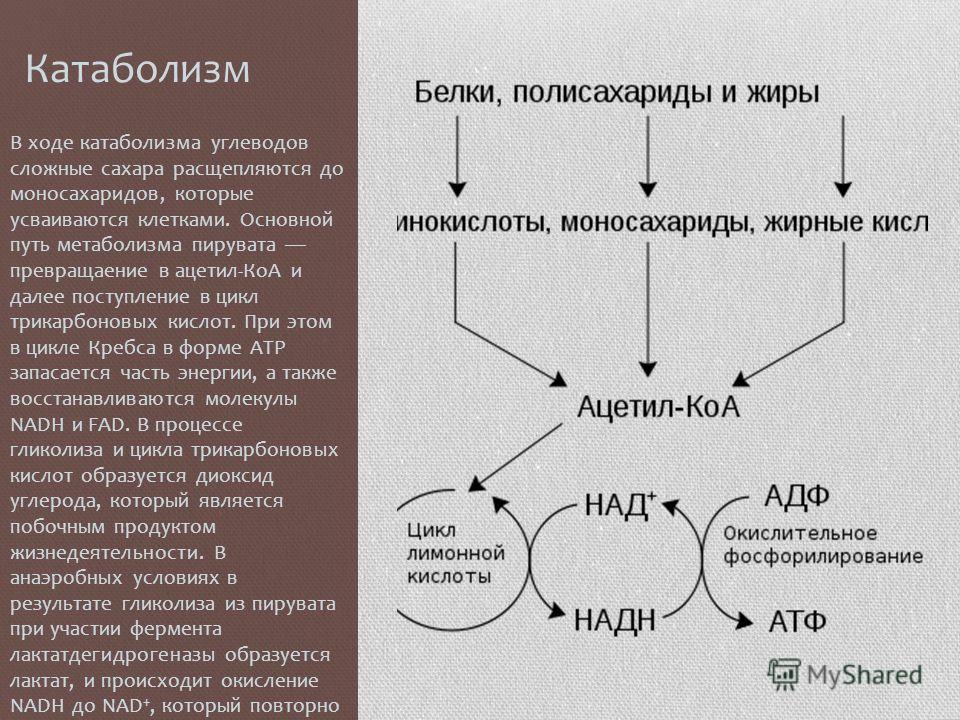
Рассмотрим метаболизм сахара (углевода). Это классический пример одного из многих клеточных процессов, которые используют и производят энергию. Живые существа потребляют сахар в качестве основного источника энергии, потому что молекулы сахара имеют большое количество энергии, хранящейся в их связях. По большей части фотосинтезирующие организмы, такие как растения, производят эти сахара. Во время фотосинтеза растения используют энергию (первоначально солнечного света) для преобразования газообразного углекислого газа (CO
6CO 2 + 6H 2 O–> C 6 H 12 O 6 + 6O 2
Вспомните из химии, что сокращение «CO 2 » означает «один атом углерода, ковалентно связанный с двумя атомами кислорода». Вода, «H 2 O», представляет собой два атома водорода, ковалентно связанные с одним атомом кислорода. А «C 6 H 12 O 6 » имеет 6 атомов углерода, 12 атомов водорода и 6 атомов кислорода, которые ковалентно связаны вместе.
Вода, «H 2 O», представляет собой два атома водорода, ковалентно связанные с одним атомом кислорода. А «C 6 H 12 O 6 » имеет 6 атомов углерода, 12 атомов водорода и 6 атомов кислорода, которые ковалентно связаны вместе.
Процесс производства глюкозы из углекислого газа и воды требует затрат энергии, потому что глюкоза содержит больше энергии в своих молекулярных связях, чем углекислый газ.
Напротив, молекулы-накопители энергии, такие как глюкоза, расходуются на расщепление для использования своей энергии.Реакцию, которая собирает энергию молекулы сахара в клетках, нуждающихся в кислороде для выживания, можно описать обратной реакцией на фотосинтез. В этой реакции расходуется кислород и выделяется углекислый газ в качестве побочного продукта. Реакция резюмируется как:
Реакция резюмируется как:
C 6 H 12 O 6 + 6O 2 -> 6H 2 O + 6CO 2
Обе эти реакции включают много этапов.
Процессы производства и расщепления молекул сахара иллюстрируют два примера метаболических путей.Метаболический путь представляет собой серию химических реакций, в которых исходная молекула изменяется, шаг за шагом, через ряд промежуточных продуктов метаболизма, в конечном итоге давая конечный продукт. В примере метаболизма сахара первый метаболический путь синтезирует сахар из более мелких молекул, а другой путь расщепляет сахар на более мелкие молекулы.
Если не указано иное, изображения на этой странице лицензированы OpenStax по лицензии CC-BY 4.0.
Текст адаптирован из: OpenStax, Концепции биологии.OpenStax CNX. 18 мая 2016 г. http://cnx.org/contents/[email protected]
Метаболизм растений — обзор
4.
 58.5 Другие эндофитические механизмы, влияющие на статус питательных веществ растений
58.5 Другие эндофитические механизмы, влияющие на статус питательных веществ растений Некоторые эндофиты, по-видимому, обладают способностью манипулировать метаболизмом растения-хозяина для увеличения поглощения питательных веществ и изменения гомеостаза питательных веществ. Когда гриб AM G. intraradices заселяет корни кукурузы, одна из реакций хозяина заключается в подавлении собственной нитратредуктазы, позволяя более эффективному грибу сокращать весь ассимилят азота [105].С другой стороны, инфицирование корней томатов G. intraradices Sy167 усиливает экспрессию высокоаффинного транспортера нитратов LeNRT2.3 в корнях, стимулируя большее поглощение нитратов растениями [106]. Трава Broomsedge ( Andropogon virginicus L.), зараженная двумя грибами AM и растущая на песке с низким содержанием P, имеет более высокую эффективность использования фосфора при низких концентрациях Pi и поддерживает постоянный уровень P в тканях не только за счет увеличения поглощения P, но и за счет изменения растительные закономерности распределения и использования фосфора [107]. Неизвестно, как микоризы вызывают эти изменения в гомеостазе питательных веществ растений. Грибковый эндофит P. indica может колонизировать внутреннюю часть корней ряда различных видов растений, где он способствует их росту. В Arabidopsis и корнях табака P. indica стимулирует поглощение / уменьшение / накопление нитратов за счет активации фактора транскрипции хозяина, который активирует P. indica -чувствительную нитратредуктазу и разрушающий крахмал фермент глюкан-водная дикиназа (SEX1). ) [108].Овсяница высокорослая ( Festuca arundinacea ), выращенная с грибковым эндофитом N. coenophialum , способна поглощать большее количество почвенных нитратов и накапливать больше аминокислот в своих побегах, особенно аргинина в листьях [109]. Столкнувшись с конкуренцией за азот между растением и его эндофитом, похоже, что растение увеличивает свою потребность в поглощении азота за счет усиления активности глутаминсинтетазы побегов (на 32%).
Неизвестно, как микоризы вызывают эти изменения в гомеостазе питательных веществ растений. Грибковый эндофит P. indica может колонизировать внутреннюю часть корней ряда различных видов растений, где он способствует их росту. В Arabidopsis и корнях табака P. indica стимулирует поглощение / уменьшение / накопление нитратов за счет активации фактора транскрипции хозяина, который активирует P. indica -чувствительную нитратредуктазу и разрушающий крахмал фермент глюкан-водная дикиназа (SEX1). ) [108].Овсяница высокорослая ( Festuca arundinacea ), выращенная с грибковым эндофитом N. coenophialum , способна поглощать большее количество почвенных нитратов и накапливать больше аминокислот в своих побегах, особенно аргинина в листьях [109]. Столкнувшись с конкуренцией за азот между растением и его эндофитом, похоже, что растение увеличивает свою потребность в поглощении азота за счет усиления активности глутаминсинтетазы побегов (на 32%). Точно так же овсяница высокорослая ( F. arundinacea Schreb.), Инфицированная грибковым эндофитом N.coenophialum (Morgan-Jones and Gams) испытывают повышенный рост растений и повышенное содержание фосфора в тканях, что позволяет предположить, что N. coenophialum является дополнительным поглотителем фосфора и стимулирует растение к увеличению поглощения фосфора [110]. У тополя, инфицированного азотфиксирующим эндофитным штаммом Paenibacillus , метаболическая сигнатура растения была изменена, увеличив уровни аспарагина и мочевины при одновременном снижении сахаров TCA и органических кислот [111]. В отличие от вышеуказанных стратегий, снижение уровней нитратов и аминокислот в тканях растений было зарегистрировано у Lolium perenne , инфицированного N.lolii [112]; Предполагается, что такое сокращение количества питательных веществ делает растение менее привлекательным для травоядных.
Точно так же овсяница высокорослая ( F. arundinacea Schreb.), Инфицированная грибковым эндофитом N.coenophialum (Morgan-Jones and Gams) испытывают повышенный рост растений и повышенное содержание фосфора в тканях, что позволяет предположить, что N. coenophialum является дополнительным поглотителем фосфора и стимулирует растение к увеличению поглощения фосфора [110]. У тополя, инфицированного азотфиксирующим эндофитным штаммом Paenibacillus , метаболическая сигнатура растения была изменена, увеличив уровни аспарагина и мочевины при одновременном снижении сахаров TCA и органических кислот [111]. В отличие от вышеуказанных стратегий, снижение уровней нитратов и аминокислот в тканях растений было зарегистрировано у Lolium perenne , инфицированного N.lolii [112]; Предполагается, что такое сокращение количества питательных веществ делает растение менее привлекательным для травоядных.
Некоторые из сигналов, используемых эндофитами для воздействия на своих хозяев, начинают обнаруживаться: Epichloë festucae выделяет активные формы кислорода (АФК) для связи со своим травяным хозяином, L. perenne ; когда уровни ROS изменяются, отношения переключаются с мутуалистических на антагонистические, что приводит к болезни и гибели растения [113]. Другой элиситор был обнаружен в фильтратах культур стимулирующего рост микроба B.thuringiensis NEB17, которые содержат новый белок бактериоцин, называемый туригеном, который увеличивает биомассу как сои, так и кукурузы [114]. P. fluorescens B16 — это стимулирующие рост ризобактерии, которые продуцируют пирролохинолинхинин в условиях с низким содержанием питательных веществ; Исследования бактериальных мутантов с использованием биоанализа на проростках огурцов показали, что это соединение отвечает за наблюдаемую стимуляцию роста растений [115]. Давно известно, что факторы Nod, секретируемые ризобиями, важны для образования клубеньков, но эти соединения также способны влиять на другие изменения в растении, такие как повышенное поглощение кальция корнями сои, посредством неизвестных механизмов [116].Обработка семян Z.
perenne ; когда уровни ROS изменяются, отношения переключаются с мутуалистических на антагонистические, что приводит к болезни и гибели растения [113]. Другой элиситор был обнаружен в фильтратах культур стимулирующего рост микроба B.thuringiensis NEB17, которые содержат новый белок бактериоцин, называемый туригеном, который увеличивает биомассу как сои, так и кукурузы [114]. P. fluorescens B16 — это стимулирующие рост ризобактерии, которые продуцируют пирролохинолинхинин в условиях с низким содержанием питательных веществ; Исследования бактериальных мутантов с использованием биоанализа на проростках огурцов показали, что это соединение отвечает за наблюдаемую стимуляцию роста растений [115]. Давно известно, что факторы Nod, секретируемые ризобиями, важны для образования клубеньков, но эти соединения также способны влиять на другие изменения в растении, такие как повышенное поглощение кальция корнями сои, посредством неизвестных механизмов [116].Обработка семян Z. mays (кукуруза), O. sativa (рис), Beta vulgaris (сахарная свекла), G. max (соя), P. vulgaris (фасоль) и Gossypium hirsutum (хлопок) с Nod-фактором BjV B. japonicum 532C привел к усиленному прорастанию семян и раннему росту в лабораторных и полевых условиях, которые могут обеспечить развивающимся сеянцам оптимальный доступ к питательным веществам в ризосфере [117]. Трансгенный рис, сверхэкспрессирующий ортолога раннего гена нодулина, OsENOD93-1, имел более высокую сухую биомассу побегов, урожай семян, общее количество аминокислот и общее количество азота в корнях [118].Хотя функция этого гена неизвестна, учитывая его гомологию с генами бобовых, участвующих в формировании клубеньков, интересно предположить, что небобовые растения могут иметь древний бактериально-зависимый путь стимуляции роста растений.
mays (кукуруза), O. sativa (рис), Beta vulgaris (сахарная свекла), G. max (соя), P. vulgaris (фасоль) и Gossypium hirsutum (хлопок) с Nod-фактором BjV B. japonicum 532C привел к усиленному прорастанию семян и раннему росту в лабораторных и полевых условиях, которые могут обеспечить развивающимся сеянцам оптимальный доступ к питательным веществам в ризосфере [117]. Трансгенный рис, сверхэкспрессирующий ортолога раннего гена нодулина, OsENOD93-1, имел более высокую сухую биомассу побегов, урожай семян, общее количество аминокислот и общее количество азота в корнях [118].Хотя функция этого гена неизвестна, учитывая его гомологию с генами бобовых, участвующих в формировании клубеньков, интересно предположить, что небобовые растения могут иметь древний бактериально-зависимый путь стимуляции роста растений.
Модификация почвы через экссудаты — важный способ, которым корни могут увеличить доступность питательных веществ. Было показано, что растения секретируют до 40% связанного углерода через свои корневые системы в виде аминокислот, органических кислот, сахаров, фенольных смол, слизи, белков и множества дополнительных вторичных метаболитов, которые могут помочь в оптимизации их ризосфер химически и микробиологически. [119].Пример измененного экссудата, влияющего на питание растений, был замечен при заражении овсяницы высокорослой ( F. arundinacea Shreb.) Грибком N. coenophialum (Morgan-Jones and Gams), который, как наблюдали, стимулировал поглощение и транспорт большего P, Ca, Zn и Cu в корнях, выращенных в питательном растворе с низким содержанием P [28]. Этот эффект был специфическим для генотипа растения DN2 и, по-видимому, был связан с репрограммированием роста корней и измененным паттерном кислотной экссудации корнями. Инфекция N. coenophialum , как было показано, увеличивает высвобождение органического углерода овсяницей из ее корней, что приводит к более высокой микробной активности и дыханию, стимулируемому изменениями в ризодепозитах [120].
Было показано, что растения секретируют до 40% связанного углерода через свои корневые системы в виде аминокислот, органических кислот, сахаров, фенольных смол, слизи, белков и множества дополнительных вторичных метаболитов, которые могут помочь в оптимизации их ризосфер химически и микробиологически. [119].Пример измененного экссудата, влияющего на питание растений, был замечен при заражении овсяницы высокорослой ( F. arundinacea Shreb.) Грибком N. coenophialum (Morgan-Jones and Gams), который, как наблюдали, стимулировал поглощение и транспорт большего P, Ca, Zn и Cu в корнях, выращенных в питательном растворе с низким содержанием P [28]. Этот эффект был специфическим для генотипа растения DN2 и, по-видимому, был связан с репрограммированием роста корней и измененным паттерном кислотной экссудации корнями. Инфекция N. coenophialum , как было показано, увеличивает высвобождение органического углерода овсяницей из ее корней, что приводит к более высокой микробной активности и дыханию, стимулируемому изменениями в ризодепозитах [120]. Овсяница высокорослая, лишенная фосфора, инфицированная этим эндофитом, также может увеличивать корневую экссудацию фенольных соединений на 7%, что приводит к увеличению на 375% скорости восстановления почвенного Fe 3+ , что является необходимым шагом в поглощении железа (Fe 2+ ) растениями [110]. При определенных условиях запасы углерода и азота в почве могут также увеличиваться эндофитной инфекцией овсяницы высокорослой, вызванной либо снижением микробного дыхания почвы [121], либо уменьшением количества определенных видов ризобактерий, потребляющих углерод [122].Это может быть вызвано изменением характера экссудации корней или накоплением в почве алкалоидов эндофитного происхождения [123]. В дополнение к Neotyphodium было показано, что AM-грибы изменяют экссудаты растений в почву, в том числе снижают уровни общих сахаров, выделяемых из корней, изменяя пропорции выделяемых аминокислот, уменьшая утечку K + и P, и увеличение выделения азота, фенолов и ГА [124].
Овсяница высокорослая, лишенная фосфора, инфицированная этим эндофитом, также может увеличивать корневую экссудацию фенольных соединений на 7%, что приводит к увеличению на 375% скорости восстановления почвенного Fe 3+ , что является необходимым шагом в поглощении железа (Fe 2+ ) растениями [110]. При определенных условиях запасы углерода и азота в почве могут также увеличиваться эндофитной инфекцией овсяницы высокорослой, вызванной либо снижением микробного дыхания почвы [121], либо уменьшением количества определенных видов ризобактерий, потребляющих углерод [122].Это может быть вызвано изменением характера экссудации корней или накоплением в почве алкалоидов эндофитного происхождения [123]. В дополнение к Neotyphodium было показано, что AM-грибы изменяют экссудаты растений в почву, в том числе снижают уровни общих сахаров, выделяемых из корней, изменяя пропорции выделяемых аминокислот, уменьшая утечку K + и P, и увеличение выделения азота, фенолов и ГА [124].
В отличие от эндофитов Neotyphodium , которые модифицируют почвы изнутри своих растений-хозяев, AM-подобные грибы способны расти из корней и напрямую изменять почву.Как упоминалось ранее, по крайней мере один вид (http://www.nature.com/nature/journal/v413/n6853/full/413297a0.html) AM, по-видимому, способен усиливать разложение органического N [16,125], но как это происходит, неизвестно. Некоторые AM способны влиять на поведение других почвенных микробов: прививка G. mosseae на различных видах растений привела к метаболической стимуляции популяций бактериальной ризосферы, включая различные группы некультивируемых бактерий и видов Paenibacillus , вероятно, через изменение структуры экссудатов в почву. [126].Сами грибы AM напрямую выделяют в почву большое количество гликопротеина, называемого гломалином, который может служить для агрегирования частиц почвы, увеличения удержания воды, хелатирования железа или служить источником энергии для почвенных микробов [124]. Другое исследование действия мицелиальных экссудатов грибка AM Glomus spp. MUCL 43205 показал, что он вызывал увеличение популяции в почве нескольких Gammaproteobacteria, включая группу Enterobacteriaceae, хотя какие функциональные изменения привели к ризосфере, неясно [127].Точно так же было показано, что почти повсеместно распространенные корневые колонизирующие грибы, известные как DSE, производят гифы, которые выходят из корня растения и поглощают органически связанные минеральные питательные вещества. Было показано, что эти грибы секретируют целлюлазы, лакказы, амилазы, липазы, пектиназы, ксиланазы, протеолитические ферменты, тирозиназы и полифенолоксидазы, но пока неизвестно, секретируются ли эти ферменты в почву для усвоения питательных веществ [22]. .
Другое исследование действия мицелиальных экссудатов грибка AM Glomus spp. MUCL 43205 показал, что он вызывал увеличение популяции в почве нескольких Gammaproteobacteria, включая группу Enterobacteriaceae, хотя какие функциональные изменения привели к ризосфере, неясно [127].Точно так же было показано, что почти повсеместно распространенные корневые колонизирующие грибы, известные как DSE, производят гифы, которые выходят из корня растения и поглощают органически связанные минеральные питательные вещества. Было показано, что эти грибы секретируют целлюлазы, лакказы, амилазы, липазы, пектиназы, ксиланазы, протеолитические ферменты, тирозиназы и полифенолоксидазы, но пока неизвестно, секретируются ли эти ферменты в почву для усвоения питательных веществ [22]. .
В качестве последнего механизма некоторые почвенные ризосферные бактерии и грибы, которые могут существовать как эндофиты внутри корней, способны минерализовать органические или нерастворимые формы азота и фосфора. Поскольку редко было показано, что эндофиты выходят из корня, чтобы напрямую воздействовать на почву, неясно, как может происходить такая очевидная солюбилизация фосфатов или разложение органических соединений. Например, при исследовании эндофитов сои было обнаружено, что 49% из них были способны растворять минеральный фосфат по сравнению с 52% эпифитных бактерий листьев, хотя не было показано, перемещались ли эндофиты на поверхности растений или в почву, где фосфат солюбилизация была бы важна [128]. В салате-латуке и кукурузе инокуляция семян фосфат-солюбилизирующими штаммами R.leguminosarum bv. phaseoli увеличивает содержание фосфора на 6% и 8%, соответственно, в полевых условиях, хотя не было продемонстрировано, что эти обычно населяющие клубеньки бактерии ведут внутри этих растений эндофитный образ жизни [129]. Есть несколько примеров, когда подтвержденные эндофиты покидают растение, улучшая биодоступность питательных веществ в почве. Например, семена орхидей, которые малы и бедны питательными веществами для развития зародыша, обладают эндофитными грибами Rhizoctonia , которые растут из семян и ферментативно разрушают окружающий субстрат, чтобы обеспечить зародыш питательными веществами для роста [130].
Поскольку редко было показано, что эндофиты выходят из корня, чтобы напрямую воздействовать на почву, неясно, как может происходить такая очевидная солюбилизация фосфатов или разложение органических соединений. Например, при исследовании эндофитов сои было обнаружено, что 49% из них были способны растворять минеральный фосфат по сравнению с 52% эпифитных бактерий листьев, хотя не было показано, перемещались ли эндофиты на поверхности растений или в почву, где фосфат солюбилизация была бы важна [128]. В салате-латуке и кукурузе инокуляция семян фосфат-солюбилизирующими штаммами R.leguminosarum bv. phaseoli увеличивает содержание фосфора на 6% и 8%, соответственно, в полевых условиях, хотя не было продемонстрировано, что эти обычно населяющие клубеньки бактерии ведут внутри этих растений эндофитный образ жизни [129]. Есть несколько примеров, когда подтвержденные эндофиты покидают растение, улучшая биодоступность питательных веществ в почве. Например, семена орхидей, которые малы и бедны питательными веществами для развития зародыша, обладают эндофитными грибами Rhizoctonia , которые растут из семян и ферментативно разрушают окружающий субстрат, чтобы обеспечить зародыш питательными веществами для роста [130]. Точно так же кактус кардон ( Pachycereus pringlei ) может расти на голой скале в Северной Мексике с помощью своих эндофитов, передаваемых через семена (в основном Bacillus spp., Klebsiella spp., Staphylococcus spp.), Которые появляются. чтобы покинуть семя для колонизации и выветривания породы, высвобождая жизненно важные минералы для развития сеянца [95]. Бактерии, ассоциированные с кактусом cardon, могут либо солюбилизировать неорганические фосфаты, выделяя органические кислоты, такие как глюконовая кислота и 2-кетоглюконовая кислота, либо минерализовать органические фосфаты, секретируя внеклеточные фосфатазы [131].
Точно так же кактус кардон ( Pachycereus pringlei ) может расти на голой скале в Северной Мексике с помощью своих эндофитов, передаваемых через семена (в основном Bacillus spp., Klebsiella spp., Staphylococcus spp.), Которые появляются. чтобы покинуть семя для колонизации и выветривания породы, высвобождая жизненно важные минералы для развития сеянца [95]. Бактерии, ассоциированные с кактусом cardon, могут либо солюбилизировать неорганические фосфаты, выделяя органические кислоты, такие как глюконовая кислота и 2-кетоглюконовая кислота, либо минерализовать органические фосфаты, секретируя внеклеточные фосфатазы [131].
Вирусный захват клеточного метаболизма | BMC Biology
Павлова Н.Н., Томпсон ЦБ. Новые признаки метаболизма рака. Cell Metab. 2016; 23: 27–47.
CAS PubMed PubMed Central Статья Google Scholar
Тай М., Грэм Н.А., Браас Д., Нехил М. , Комисопулу Э., Курдистанский СК и др. Активация MYC, вызванная аденовирусом E4ORF1, способствует анаболическому метаболизму глюкозы в клетке-хозяине и репликации вируса.Cell Metab. 2014; 19: 694–701.
, Комисопулу Э., Курдистанский СК и др. Активация MYC, вызванная аденовирусом E4ORF1, способствует анаболическому метаболизму глюкозы в клетке-хозяине и репликации вируса.Cell Metab. 2014; 19: 694–701.
CAS PubMed PubMed Central Статья Google Scholar
Vastag L, Koyuncu E, Grady SL, Shenk TE, Rabinowitz JD. Различные эффекты цитомегаловируса человека и вируса простого герпеса-1 на клеточный метаболизм. PLoS Pathog. 2011; 7: e1002124.
CAS PubMed PubMed Central Статья Google Scholar
Хитон Н.С., Рэндалл Г. Аутофагия, вызванная вирусом Денге, регулирует липидный обмен. Клеточный микроб-хозяин. 2010; 8: 422–32.
CAS PubMed PubMed Central Статья Google Scholar
Перера Р., Райли С., Исаак Г., Хопф-Яннаш А.С., Мур Р.Дж., Вейтц К.В. и др. Инфекция, вызванная вирусом денге, нарушает гомеостаз липидов в инфицированных клетках комаров.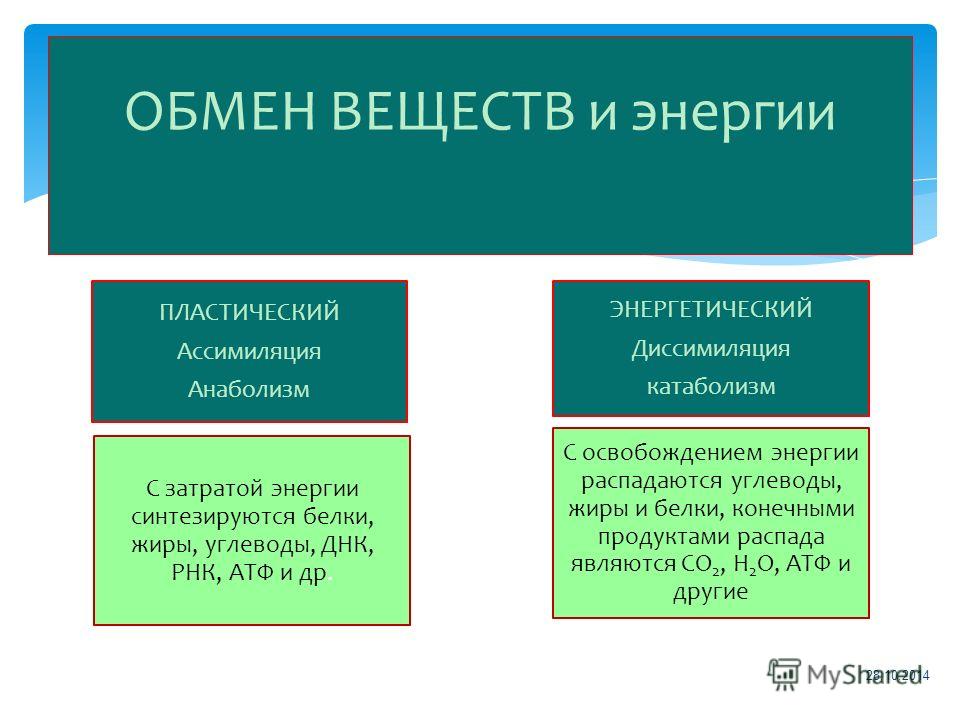 PLoS Pathog. 2012; 8: e1002584.
PLoS Pathog. 2012; 8: e1002584.
CAS PubMed PubMed Central Статья Google Scholar
Ли Х, Чжу В., Чжан Л., Лей Х, Ву Х, Го Л. и др. Метаболические реакции на вирусную инфекцию гепатита В проливают новый свет на патогенез и цели лечения. Научный доклад 2015; 5: 8421.
CAS PubMed PubMed Central Статья Google Scholar
Teng CF, Wu HC, Hsieh WC, Tsai HW, Su IJ. Активация цитратлиазы АТФ сигналом mTOR вызывает нарушение липидного метаболизма в пре-S2 мутантном туморогенезе вируса гепатита B.J Virol. 2015; 89: 605–14.
PubMed Статья CAS PubMed Central Google Scholar
Дельгадо Т., Санчес Э.Л., Камарда Р., Лагунов М. Глобальное метаболическое профилирование инфекции, вызванной онкогенным вирусом: KSHV индуцирует липогенез и требует его для выживания латентной инфекции. PLoS Pathog. 2012; 8: e1002866.
PLoS Pathog. 2012; 8: e1002866.
CAS PubMed PubMed Central Статья Google Scholar
O’Shea CC. Вирусы — ищут и уничтожают опухолевую программу. Онкоген. 2005; 24: 7640–55.
CAS PubMed Статья PubMed Central Google Scholar
Фишер Т.Н., Гинзберг Х.С. Накопление органических кислот клетками HeLa, инфицированными аденовирусом 4 типа. Proc Soc Exp Biol Med. 1957; 95: 47–51.
CAS PubMed Статья PubMed Central Google Scholar
Барделл Д. Поглощение глюкозы и продукция молочной кислоты инфицированными аденовирусом 5-го типа клетками HEp-2, культивированными в условиях экспоненциального роста и стационарной фазы. Microbios. 1977; 20: 139–44.
CAS PubMed PubMed Central Google Scholar

Thai M, Thaker SK, Feng J, Du Y, Hu H, Ting Wu T. и др. MYC-индуцированное перепрограммирование катаболизма глутамина поддерживает оптимальную репликацию вируса. Nat Commun. 2015; 6: 8873.
CAS PubMed PubMed Central Статья Google Scholar
Whitley RJ. Герпесвирусы. В: Барон С., редактор. Медицинская микробиология. Галвестон; 1996. Глава 68.
Льюис В.Дж. младший, Скотт Л.В. Требования к питанию для производства вируса простого герпеса. I. Влияние глюкозы и глутамина на продукцию вируса простого герпеса клетками HeLa. J Bacteriol. 1962; 83: 475–82.
PubMed PubMed Central Google Scholar
Abrantes JL, Alves CM, Costa J, Almeida FC, Sola-Penna M, Fontes CF и др. Простой герпес 1 типа активирует гликолиз за счет задействования фермента 6-фосфофрукто-1-киназы (PFK-1). Biochim Biophys Acta. 1822; 2012: 1198–206.
Google Scholar
Grady SL, Purdy JG, Rabinowitz JD, Shenk T. Истощение запасов аргининосукцинатсинтетазы 1 вызывает метаболическое состояние, способствующее инфицированию вирусом простого герпеса 1. Proc Natl Acad Sci U S A.2013; 110: E5006–15.
CAS PubMed PubMed Central Статья Google Scholar
Oh Y, Zhang F, Wang Y, Lee EM, Choi IY, Lim H, et al. Вирус Зика напрямую поражает периферические нейроны и вызывает гибель клеток. Nat Neurosci. 2017; 20: 1209–12.
CAS PubMed PubMed Central Статья Google Scholar
Смит С., Реувен Н., Мохни К.Н., Шумахер А.Дж., Веллер С.К.Структура генома вируса простого герпеса 1: манипуляции с зазубринами и промежутками могут отменить инфекционность и изменить реакцию на повреждение клеточной ДНК. J Virol. 2014; 88: 10146–56.
PubMed PubMed Central Статья CAS Google Scholar

Boehmer PE, Lehman IR. Репликация ДНК вируса простого герпеса. Анну Рев Биохим. 1997; 66: 347–84.
CAS PubMed Статья PubMed Central Google Scholar
Биркманн А., Циммерманн Х. Противовирусные препараты от HSV — текущие и будущие варианты лечения. Curr Opin Virol. 2016; 18: 9–13.
CAS PubMed Статья PubMed Central Google Scholar
Esquenazi D, Wigg MD, Miranda MM, Rodrigues HM, Tostes JB, Rozental S, et al. Антимикробная и противовирусная активность полифенолов из Cocos nucifera Linn. (Palmae) экстракт волокон шелухи. Res Microbiol. 2002; 153: 647–52.
CAS PubMed Статья PubMed Central Google Scholar
Ноулз Р.В., Персона С. Влияние 2-дезоксиглюкозы, глюкозамина и маннозы на слияние клеток и гликопротеины вируса простого герпеса. J Virol. 1976; 18: 644–51.
CAS PubMed PubMed Central Google Scholar
Кортни Р.Дж., Штайнер С.М., Бенеш-Мельник М. Эффекты 2-дезокси-D-глюкозы на репликацию вируса простого герпеса. Вирусология. 1973; 52: 447–55.
CAS PubMed Статья PubMed Central Google Scholar
McArdle J, Schafer XL, Munger J. Ингибирование кальмодулин-зависимой киназы киназы блокирует гликолитическую активацию, индуцированную цитомегаловирусом человека, и сильно ослабляет продукцию вирусного потомства. J Virol. 2011; 85: 705–14.
CAS PubMed Статья PubMed Central Google Scholar
Blough HA, Giuntoli RL. Успешное лечение инфекций генитального герпеса человека с помощью 2-дезокси-D-глюкозы. ДЖАМА. 1979; 241: 2798–801.
CAS PubMed Статья PubMed Central Google Scholar

Kern ER, Glasgow LA, Klein RJ, Friedman-Kien AE. Неспособность 2-дезокси-D-глюкозы в лечении экспериментальных кожных и генитальных инфекций, вызванных вирусом простого герпеса. J Infect Dis. 1982; 146: 159–66.
CAS PubMed Статья PubMed Central Google Scholar
Гриффитс П., Бараняк И., Ривз М.Патогенез цитомегаловируса человека. J Pathol. 2015; 235: 288–97.
CAS PubMed Статья PubMed Central Google Scholar
Landini MP. Раннее повышенное поглощение глюкозы клетками, инфицированными цитомегаловирусом человека. J General Virol. 1984; 65 (Pt 7): 1229–32.
CAS Статья Google Scholar
Munger J, Bennett BD, Parikh A, Feng XJ, McArdle J, Rabitz HA, et al.Профили метаболического потока на системном уровне определяют синтез жирных кислот как цель противовирусной терапии. Nat Biotechnol. 2008; 26: 1179–86.
Nat Biotechnol. 2008; 26: 1179–86.
CAS PubMed PubMed Central Статья Google Scholar
Yu Y, Maguire TG, Alwine JC. Цитомегаловирус человека активирует экспрессию транспортера глюкозы 4 для увеличения поглощения глюкозы во время инфекции. J Virol. 2011; 85: 1573–80.
CAS PubMed Статья Google Scholar
Чемберс Дж. У., Магуайр Т. Г., Алвин Дж. С.. Метаболизм глутамина важен для цитомегаловирусной инфекции человека. J Virol. 2010; 84: 1867–73.
CAS PubMed Статья PubMed Central Google Scholar
Munger J, Bajad SU, Coller HA, Shenk T., Rabinowitz JD. Динамика клеточного метаболома при цитомегаловирусной инфекции человека. PLoS Pathog. 2006; 2: e132.
PubMed PubMed Central Статья CAS Google Scholar

Yu Y, Maguire TG, Alwine JC. ChREBP, глюкозо-зависимый транскрипционный фактор, усиливает метаболизм глюкозы для поддержки биосинтеза в клетках, инфицированных цитомегаловирусом человека. Proc Natl Acad Sci U S. A. 2014; 111: 1951–6.
CAS PubMed PubMed Central Статья Google Scholar
McArdle J, Moorman NJ, Munger J. HCMV воздействует на метаболический стрессовый ответ посредством активации AMPK, активность которого важна для репликации вируса.PLoS Pathog. 2012; 8: e1002502.
CAS PubMed PubMed Central Статья Google Scholar
Vysochan A, Sengupta A, Weljie AM, Alwine JC, Yu Y. ACSS2-опосредованный синтез ацетил-КоА из ацетата необходим для инфицирования цитомегаловирусом человека. Proc Natl Acad Sci U S. A. 2017; 114: E1528 – E35.
CAS PubMed PubMed Central Статья Google Scholar

Yu Y, Maguire TG, Alwine JC. Инфекция, вызванная цитомегаловирусом человека, индуцирует липогенез, подобный адипоцитам, за счет активации белка 1, связывающего регуляторный элемент стерола. J Virol. 2012; 86: 2942–9.
CAS PubMed PubMed Central Статья Google Scholar
Yu Y, Pierciey FJ Jr, Maguire TG, Alwine JC. PKR-подобная киназа эндоплазматического ретикулума необходима для липогенной активации во время инфекции HCMV. PLoS Pathog.2013; 9: e1003266.
CAS PubMed PubMed Central Статья Google Scholar
Jordan TX, Randall G. Модуляция клеточного метаболизма флавивирусом. Curr Opin Virol. 2016; 19: 7–10.
CAS PubMed PubMed Central Статья Google Scholar
Фонтейн К.А., Санчес Е.Л., Камарда Р., Лагунофф М. Вирус денге индуцирует гликолиз и требует его для оптимальной репликации. J Virol. 2015; 89: 2358–66.
J Virol. 2015; 89: 2358–66.
PubMed Статья CAS Google Scholar
Heaton NS, Perera R, Berger KL, Khadka S, Lacount DJ, Kuhn RJ, et al. Неструктурный белок 3 вируса денге перераспределяет синтазу жирных кислот к участкам репликации вируса и увеличивает синтез жирных кислот в клетках. Proc Natl Acad Sci U S. A. 2010; 107: 17345–50.
CAS PubMed PubMed Central Статья Google Scholar
Jordan TX, Randall G. Вирус денге активирует ось AMP-киназа-mTOR, чтобы стимулировать провирусную липофагию. J Virol. 2017; 91: e02020–16.
CAS PubMed PubMed Central Статья Google Scholar
Мартин-Асебес М.А., Мерино-Рамос Т., Бласкес А.Б., Касас Дж., Эскрибано-Ромеро Е., Собрино Ф. и др. Состав липидной оболочки вируса Западного Нила раскрывает роль метаболизма сфинголипидов в биогенезе флавивирусов. J Virol. 2014; 88: 12041–54.
J Virol. 2014; 88: 12041–54.
PubMed PubMed Central Статья CAS Google Scholar
Макино С., Дженкин Х.М. Влияние жирных кислот на рост вируса японского энцефалита, культивируемого в клетках BHK-21, и метаболизм фосфолипидов в инфицированных клетках. J Virol. 1975. 15: 515–25.
CAS PubMed PubMed Central Google Scholar
Майнер Дж. Дж., Даймонд МС.Патогенез и тканевой тропизм вируса Зика. Клеточный микроб-хозяин. 2017; 21: 134–42.
CAS PubMed PubMed Central Статья Google Scholar
Thaker SK, Chapa T, Garcia G Jr, Gong D, Schmid EW, Arumugaswami V, et al. Дифференциальное метаболическое перепрограммирование вирусом Зика способствует гибели клеток человека по сравнению с клетками комаров. Cell Metab. 2019; 29: 1206–16 e4.
CAS PubMed Статья PubMed Central Google Scholar

Tolonen N, Doglio L, Schleich S, Krijnse LJ. Репликация ДНК вируса осповакцины происходит в мини-ядрах цитоплазмы, заключенных в эндоплазматический ретикулум. Mol Biol Cell. 2001; 12: 2031–46.
CAS PubMed PubMed Central Статья Google Scholar
Фонтейн К.А., Камарда Р., Лагунофф М. Вирусу осповакцины для эффективной репликации требуется глутамин, но не глюкоза. J Virol. 2014; 88: 4366–74.
PubMed PubMed Central Статья CAS Google Scholar
Greseth MD, Traktman P. Биосинтез жирных кислот de novo вносит значительный вклад в создание биоэнергетически благоприятной среды для заражения вирусом осповакцины. PLoS Pathog. 2014; 10: e1004021.
PubMed PubMed Central Статья CAS Google Scholar
Mazzon M, Peters NE, Loenarz C, Krysztofinska EM, Ember SW, Ferguson BJ, et al. Механизм индукции гипоксического ответа вирусом осповакцины.Proc Natl Acad Sci U S. A. 2013; 110: 12444–9.
Механизм индукции гипоксического ответа вирусом осповакцины.Proc Natl Acad Sci U S. A. 2013; 110: 12444–9.
CAS PubMed PubMed Central Статья Google Scholar
Semenza GL. Ориентация на HIF-1 для лечения рака. Нат Рев Рак. 2003; 3: 721–32.
CAS PubMed Статья PubMed Central Google Scholar
Taubenberger JK, Morens DM. Патология вирусных инфекций гриппа. Анну Рев Патол.2008; 3: 499–522.
CAS PubMed PubMed Central Статья Google Scholar
Риттер Дж. Б., Валь А. С., Фройнд С., Гензель Ю., Райхл У. Метаболические эффекты инфекции вируса гриппа в культивируемых клетках животных: профили внутри- и внеклеточных метаболитов. BMC Syst Biol. 2010; 4: 61.
PubMed PubMed Central Статья CAS Google Scholar

Кохио HP, Адамсон А.Л. Гликолитический контроль активности АТФазы вакуолярного типа: механизм регулирования вирусной инфекции гриппа. Вирусология. 2013; 444: 301–9.
CAS PubMed Статья PubMed Central Google Scholar
Smallwood HS, Duan S, Morfouace M, Rezinciuc S, Shulkin BL, Shelat A, et al. Направление метаболического перепрограммирования при гриппозной инфекции для терапевтического вмешательства. Cell Rep. 2017; 19: 1640–53.
CAS PubMed PubMed Central Статья Google Scholar
Су МА, Хуанг Ю.Т., Чен ИТ, Ли Д.Й., Се Ю.С., Ли СиЙ и др. Эффект Варбурга для беспозвоночных: вирус креветок достигает успешной репликации, изменяя метаболом хозяина через путь PI3K-Akt-mTOR. PLoS Pathog. 2014; 10: e1004196.
PubMed PubMed Central Статья CAS Google Scholar

Chen IT, Aoki T, Huang YT, Hirono I, Chen TC, Huang JY, et al. Вирус синдрома белого пятна вызывает метаболические изменения, напоминающие эффект Варбурга, в гемоцитах креветок на ранней стадии инфекции.J Virol. 2011; 85: 12919–28.
CAS PubMed PubMed Central Статья Google Scholar
Ван К., Хосино Ю., Дауделл К., Бош-Марс М., Майерс Т.Г., Сармиенто М. и др. Добавки глутамина подавляют реактивацию вируса простого герпеса. J Clin Invest. 2017; 127: 2626–30.
PubMed PubMed Central Статья Google Scholar
Plummer M, de Martel C, Vignat J, Ferlay J, Bray F, Franceschi S.Глобальное бремя рака, связанное с инфекциями, в 2012 г .: синтетический анализ. Lancet Global Health. 2016; 4: e609–16.
PubMed Статья PubMed Central Google Scholar
Фенг Х, Шуда М, Чанг И, Мур ПС. Клональная интеграция полиомавируса в карциному из клеток Меркеля человека. Наука (Нью-Йорк). 2008; 319: 1096–100.
CAS Статья Google Scholar
Yabe Y, Trentin JJ, Taylor G. Индукция рака у хомяков аденовирусом человека типа 12. Влияние возраста и дозы вируса. Proc Soc Exp Biol Med. 1962; 111: 343–4.
CAS PubMed Статья PubMed Central Google Scholar
de Martel C, Plummer M, Vignat J, Franceschi S. Мировое бремя рака, связанное с ВПЧ, по месту расположения, стране и типу ВПЧ. Int J Cancer. 2017; 141: 664–70.
PubMed PubMed Central Статья CAS Google Scholar
Го И, Мэн Х, Ма Дж, Чжэн И, Ван Цюй, Ван И и др. Вирус папилломы человека 16 E6 способствует HIF-1альфа-индуцированному эффекту Варбурга, ослабляя взаимодействие VHL-HIF-1alpha. Int J Mol Sci. 2014; 15: 7974–86.
PubMed PubMed Central Статья CAS Google Scholar
Bodily JM, Mehta KP, Laimins LA. Вирус папилломы человека E7 усиливает транскрипцию, опосредованную гипоксией фактора 1, ингибируя связывание гистондеацетилаз.Cancer Res. 2011; 71: 1187–95.
CAS PubMed Статья PubMed Central Google Scholar
Цвершке В., Мазурек С., Массими П., Бэнкс Л., Эйгенбродт Е., Янсен-Дурр П. Модуляция активности пируваткиназы типа M2 онкопротеином E7 вируса папилломы человека 16 типа. Proc Natl Acad Sci U S. A. 1999; 96: 1291–6.
CAS PubMed PubMed Central Статья Google Scholar
Mazurek S, Zwerschke W, Jansen-Durr P, Eigenbrodt E. Влияние онкопротеина E7 вируса папилломы человека HPV-16 на гликолиз и глутаминолиз: роль пируваткиназы типа M2 и комплекса гликолитических ферментов. Биохим Дж. 2001; 356: 247–56.
Биохим Дж. 2001; 356: 247–56.
CAS PubMed PubMed Central Статья Google Scholar
Лай Д., Тан К.Л., Гунаратне Дж., Квек Л.С., Ней В., Тьерри Ф. и др. Локализация Е2 ВПЧ-18 на митохондриальных мембранах вызывает высвобождение АФК и модулирует метаболизм клетки-хозяина.PLoS One. 2013; 8: e75625.
CAS PubMed PubMed Central Статья Google Scholar
ВОЗ. Гепатит B [обновлено 18 июля 2018 г.]. https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-b. По состоянию на 28 июня 2019 г.
Shi YX, Huang CJ, Yang ZG. Влияние инфекции вирусом гепатита B на метаболический сигнальный путь в печени. Мир Дж. Гастроэнтерол. 2016; 22: 8161–7.
CAS PubMed PubMed Central Статья Google Scholar
Дэн И, Чжан И, Ченг Л., Ма Дж, Си И, Ян Л. и др. Нарушения метаболизма нуклеиновых кислот, вызванные белком X вируса гепатита B (HBx), выявленные с помощью (1) метабономики на основе H-ЯМР. Научный отчет 2016; 6: 24430.
и др. Нарушения метаболизма нуклеиновых кислот, вызванные белком X вируса гепатита B (HBx), выявленные с помощью (1) метабономики на основе H-ЯМР. Научный отчет 2016; 6: 24430.
Артикул CAS Google Scholar
Xie Q, Fan F, Wei W, Liu Y, Xu Z, Zhai L, et al. Многокомпонентные анализы выявляют метаболические изменения, регулируемые коровым белком вируса гепатита В в клетках гепатоцеллюлярной карциномы.Научный отчет 2017; 7: 41089.
CAS PubMed PubMed Central Статья Google Scholar
Teng CF, Hsieh WC, Wu HC, Lin YJ, Tsai HW, Huang W и др. Пре-S2 мутант вируса гепатита B индуцирует аэробный гликолиз через млекопитающих, являющихся мишенью сигнального каскада рапамицина. PLoS One. 2015; 10: e0122373.
PubMed PubMed Central Статья CAS Google Scholar
Лю Б. , Фанг М., Хе З, Цуй Д., Цзя С., Линь Х и др. Вирус гепатита B стимулирует экспрессию G6PD посредством HBx-опосредованной активации Nrf2. Cell Death Dis. 2015; 6: e1980.
, Фанг М., Хе З, Цуй Д., Цзя С., Линь Х и др. Вирус гепатита B стимулирует экспрессию G6PD посредством HBx-опосредованной активации Nrf2. Cell Death Dis. 2015; 6: e1980.
CAS PubMed PubMed Central Статья Google Scholar
Шин Х.Дж., Пак Й.Х., Ким СУ, Мун Х.В., Парк Д.С., Хан Й.Х. и др. Белок X вируса гепатита B регулирует гомеостаз глюкозы в печени посредством активации индуцибельной синтазы оксида азота. J Biol Chem.2011; 286: 29872–81.
CAS PubMed PubMed Central Статья Google Scholar
Хаджу М., Норел Р., Карвер Р., Марион П., Каллен Дж., Роглер Л. Е. и др. Анализ микроматрицы кДНК печени трансгенных мышей HBV позволяет идентифицировать гены в путях биосинтеза липидов и путей контроля роста, на которые влияет HBV. J Med Virol. 2005; 77: 57–65.
CAS PubMed Статья PubMed Central Google Scholar
Wu YL, Peng XE, Zhu YB, Yan XL, Chen WN, Lin X. Белок X вируса гепатита B вызывает стеатоз печени, увеличивая экспрессию белка, связывающего жирные кислоты печени. J Virol. 2016; 90: 1729–40.
CAS PubMed PubMed Central Статья Google Scholar
Ким К. Х., Шин Х. Дж., Ким К., Чой Х. М., Ри Ш., Мун Х. Б. и др. Белок X вируса гепатита B вызывает стеатоз печени посредством активации транскрипции SREBP1 и PPARgamma.Гастроэнтерология. 2007; 132: 1955–67.
CAS PubMed Статья PubMed Central Google Scholar
Кан С.К., Чанг Т.В., Ли Дж.Й., Ли Ю.С., Мортон Р.Э., Ким Ч. Белок X вируса гепатита B подавляет секрецию аполипопротеина B за счет увеличения экспрессии N-ацетилглюкозаминилтрансферазы III. J Biol Chem. 2004. 279: 28106–12.
CAS PubMed Статья PubMed Central Google Scholar

Тонг А, Ву Л., Линь Кью, Лау КК, Чжао Х, Ли Дж. И др. Протеомный анализ изменений клеточного белка с использованием клеточной модели, продуцирующей вирус гепатита В. Протеомика. 2008; 8: 2012–23.
CAS PubMed Статья PubMed Central Google Scholar
Хуан B, Сиддики А. Ретиноидный рецептор X RXR альфа связывается с усилителем вируса гепатита B и трансактивирует его. Proc Natl Acad Sci U S. A. 1992; 89: 9059–63.
CAS PubMed PubMed Central Статья Google Scholar
Oehler N, Volz T., Bhadra OD, Kah J, Allweiss L, Giersch K, et al. Связывание вируса гепатита В с его клеточным рецептором изменяет профиль экспрессии генов метаболизма желчных кислот. Гепатология (Балтимор). 2014; 60: 1483–93.
CAS Статья Google Scholar
Эль-Сераг HB, Рудольф KL. Гепатоцеллюлярная карцинома: эпидемиология и молекулярный канцерогенез. Гастроэнтерология. 2007. 132: 2557–76.
Гастроэнтерология. 2007. 132: 2557–76.
CAS PubMed Статья PubMed Central Google Scholar
Эль-Сераг HB. Гепатоцеллюлярная карцинома. N Engl J Med. 2011; 365: 1118–27.
CAS PubMed Статья PubMed Central Google Scholar
Чанг М.Л. Метаболические изменения и гепатит С: от скамьи к постели. Мир Дж. Гастроэнтерол. 2016; 22: 1461–76.
CAS PubMed PubMed Central Статья Google Scholar
Мория К., Фуджи Х., Синтани Й., Йоцуянаги Х., Цуцуми Т., Ишибаши К. и др.Основной белок вируса гепатита С вызывает гепатоцеллюлярную карциному у трансгенных мышей. Nat Med. 1998; 4: 1065–7.
CAS PubMed Статья PubMed Central Google Scholar
Мория К., Тодороки Т., Цуцуми Т., Фуджи Х., Синтани Й. , Миёси Х. и др. Повышение концентрации мононенасыщенных жирных кислот углерода 18 в печени при гепатите С: анализ на трансгенных мышах и людях. Biochem Biophys Res Commun.2001; 281: 1207–12.
, Миёси Х. и др. Повышение концентрации мононенасыщенных жирных кислот углерода 18 в печени при гепатите С: анализ на трансгенных мышах и людях. Biochem Biophys Res Commun.2001; 281: 1207–12.
CAS PubMed Статья PubMed Central Google Scholar
Барба Г., Харпер Ф., Харада Т., Кохара М., Гулине С., Мацуура Ю. и др. Ядровый белок вируса гепатита С имеет цитоплазматическую локализацию и ассоциируется с капельками накопления липидов в клетках. Proc Natl Acad Sci U S. A. 1997; 94: 1200–5.
CAS PubMed PubMed Central Статья Google Scholar
Херкер Э., Харрис Ц., Эрнандес Ц., Карпентье А, Кельке К., Розенберг А.Р. и др. Для эффективного образования частиц вируса гепатита С требуется диацилглицерин ацилтрансфераза-1. Nat Med. 2010; 16: 1295–8.
CAS PubMed PubMed Central Статья Google Scholar

Харрис С., Херкер Э., Фарезе Р.В. мл., Отт М. Корбовой белок вируса гепатита С снижает оборот липидных капель: механизм стеатоза, индуцированного сердцевиной. J Biol Chem.2011; 286: 42615–25.
CAS PubMed PubMed Central Статья Google Scholar
Перлемутер Г., Сабиле А., Леттерон П., Вона Г., Топилко А., Кретьен И. и др. Основной белок вируса гепатита С подавляет активность микросомального белка-переносчика триглицеридов и секрецию липопротеинов очень низкой плотности: модель стеатоза, связанного с вирусом. FASEB J. 2002; 16: 185–94.
CAS PubMed Статья PubMed Central Google Scholar
Морииси К., Мочизуки Р., Мория К., Миямото Х., Мори Й., Абе Т. и др. Критическая роль PA28gamma в связанном с вирусом гепатита C стеатогенезе и гепатоканцерогенезе. Proc Natl Acad Sci U S. A. 2007; 104: 1661–6.
CAS PubMed PubMed Central Статья Google Scholar

Oem JK, Jackel-Cram C, Li YP, Zhou Y, Zhong J, Shimano H, et al. Активация стеринового регулирующего элемента-связывающего белка 1c и транскрипции синтазы жирных кислот неструктурным белком 2 вируса гепатита С.J General Virol. 2008; 89: 1225–30.
CAS Статья Google Scholar
Waris G, Felmlee DJ, Negro F, Siddiqui A. Вирус гепатита C индуцирует протеолитическое расщепление белков, связывающих регуляторные элементы стерола, и стимулирует их фосфорилирование посредством окислительного стресса. J Virol. 2007. 81: 8122–30.
CAS PubMed PubMed Central Статья Google Scholar
Bandiera S, Pernot S, El Saghire H, Durand SC, Thumann C, Crouchet E, et al. Вызванная вирусом гепатита С повышенная регуляция микроРНК miR-146a-5p в гепатоцитах способствует вирусной инфекции и нарушает регуляцию метаболических путей, связанных с патогенезом заболеваний печени. J Virol. 2016; 90: 6387–400.
J Virol. 2016; 90: 6387–400.
CAS PubMed PubMed Central Статья Google Scholar
Мория К., Накагава К., Санта Т., Синтани И., Фуджи Х., Миёси Х. и др.Окислительный стресс в отсутствие воспаления на мышиной модели гепатоканцерогенеза, связанного с вирусом гепатита С. Cancer Res. 2001; 61: 4365–70.
CAS PubMed PubMed Central Google Scholar
Коренага М., Ван Т., Ли И, Шоуолтер Л.А., Чан Т., Сан Дж. И др. Основной белок вируса гепатита С подавляет транспорт электронов в митохондриях и увеличивает производство активных форм кислорода (АФК). J Biol Chem. 2005; 280: 37481–8.
CAS PubMed Статья PubMed Central Google Scholar
Дэн Л., Адачи Т., Китайма К., Бунгёку Ю., Китадзава С., Исидо С. и др. Инфекция вирусом гепатита С вызывает апоптоз через запускаемый Bax, опосредованный митохондриями, зависимый от каспазы 3 путь. J Virol. 2008. 82: 10375–85.
J Virol. 2008. 82: 10375–85.
CAS PubMed PubMed Central Статья Google Scholar
Gong G, Waris G, Tanveer R, Siddiqui A. Белок NS5A вируса гепатита C человека изменяет внутриклеточные уровни кальция, индуцирует окислительный стресс и активирует STAT-3 и NF-kappa B. Proc Natl Acad Sci US A. 2001; 98: 9599–604.
CAS PubMed PubMed Central Статья Google Scholar
Гарсия-Медиавилла М.В., Санчес-Кампос С., Гонсалес-Перес П., Гомес-Гонсало М., Майано П.Л., Лопес-Кабрера М. и др. Дифференциальный вклад NS5A вируса гепатита C и основных белков в индукцию окислительного и нитрозативного стресса в клетках, происходящих из гепатоцитов человека.J Hepatol. 2005; 43: 606–13.
CAS PubMed Статья PubMed Central Google Scholar
Мачида К., Ченг К.Т., Лай К.К. , Дженг К.С., Сун В.М., Лай М.М. Вирус гепатита С вызывает изменение проницаемости митохондрий с образованием активных форм кислорода, что приводит к повреждению ДНК и активации STAT3. J Virol. 2006; 80: 7199–207.
, Дженг К.С., Сун В.М., Лай М.М. Вирус гепатита С вызывает изменение проницаемости митохондрий с образованием активных форм кислорода, что приводит к повреждению ДНК и активации STAT3. J Virol. 2006; 80: 7199–207.
CAS PubMed PubMed Central Статья Google Scholar
Окуда М., Ли К., Борода М.Р., Шоуолтер Л.А., Шолле Ф., Лимон С.М. и др. Повреждение митохондрий, окислительный стресс и экспрессия антиоксидантных генов индуцируются коровым белком вируса гепатита С. Гастроэнтерология. 2002; 122: 366–75.
CAS PubMed Статья PubMed Central Google Scholar
Цуцуми Т., Мацуда М., Айзаки Х., Мория К., Миёси Х., Фуджи Х. и др. Протеомический анализ митохондриальных белков показывает сверхэкспрессию шаперона митохондриального белка, запретитина, в клетках, экспрессирующих коровый белок вируса гепатита С.Гепатология (Балтимор). 2009. 50: 378–86.
CAS Статья Google Scholar
Li Y, Boehning DF, Qian T, Popov VL, Weinman SA. Основной белок вируса гепатита С увеличивает выработку митохондриальных АФК за счет стимуляции активности унипортера Са2 +. FASEB J. 2007; 21: 2474–85.
CAS PubMed Статья PubMed Central Google Scholar
de Mochel NS, Seronello S, Wang SH, Ito C, Zheng JX, Liang TJ, et al.НАД (Ф) Н-оксидазы гепатоцитов как эндогенный источник активных форм кислорода при инфицировании вирусом гепатита С. Гепатология (Балтимор). 2010; 52: 47–59.
Артикул CAS Google Scholar
Boudreau HE, Emerson SU, Korzeniowska A, Jendrysik MA, Leto TL. Белки вируса гепатита С (ВГС) индуцируют экспрессию НАДФН-оксидазы 4 в зависимости от трансформирующего фактора роста бета: новый фактор в индуцированном ВГС окислительном стрессе.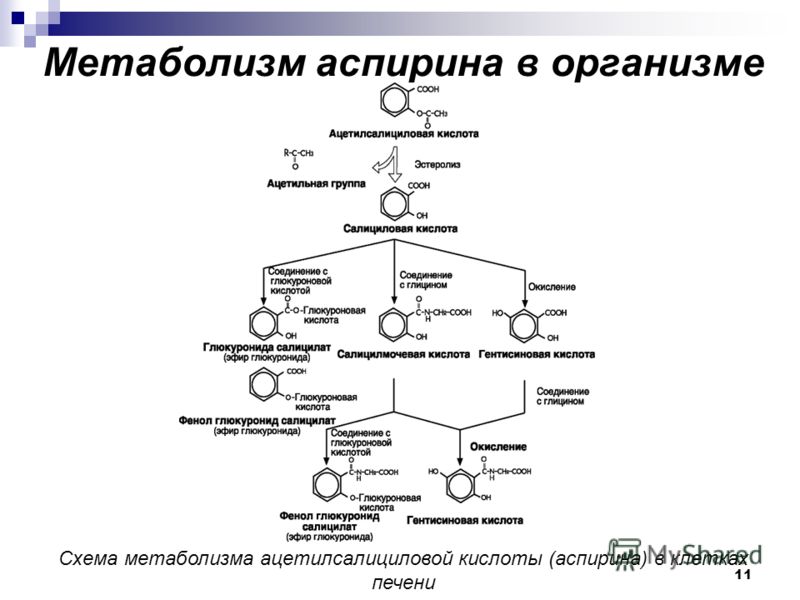 J Virol. 2009; 83: 12934–46.
J Virol. 2009; 83: 12934–46.
CAS PubMed PubMed Central Статья Google Scholar
Кавагути Т., Идэ Т., Танигучи Э., Хирано Э., Итоу М., Суми С. и др. Клиренс HCV улучшает инсулинорезистентность, функцию бета-клеток и печеночную экспрессию субстрата 1 и 2 рецептора инсулина. Am J Gastroenterol. 2007. 102: 570–6.
PubMed Статья PubMed Central Google Scholar
Синтани Й, Фуджи Х, Миёси Х, Цуцуми Т., Цукамото К., Кимура С. и др. Инфекция вируса гепатита С и диабет: прямое участие вируса в развитии инсулинорезистентности. Гастроэнтерология. 2004; 126: 840–8.
CAS PubMed Статья PubMed Central Google Scholar
Banerjee S, Saito K, Ait-Goughoulte M, Meyer K, Ray RB, Ray R. Ядровый белок вируса гепатита C активизирует фосфорилирование серина субстрата-1 рецептора инсулина и нарушает последующую передачу сигналов akt / протеинкиназы B. путь к инсулинорезистентности.J Virol. 2008. 82: 2606–12.
путь к инсулинорезистентности.J Virol. 2008. 82: 2606–12.
CAS PubMed Статья PubMed Central Google Scholar
Кавагути Т., Йошида Т., Харада М., Хисамото Т., Нагао Ю., Иде Т. и др. Вирус гепатита С подавляет субстраты 1 и 2 рецептора инсулина за счет повышения регуляции супрессора передачи сигналов цитокинов 3. Am J Pathol. 2004; 165: 1499–508.
CAS PubMed PubMed Central Статья Google Scholar
Дэн Л., Сёдзи И., Огава В., Канеда С., Сога Т., Цзян Д.П. и др. Инфекция вируса гепатита С способствует глюконеогенезу в печени посредством NS5A-опосредованного, FoxO1-зависимого пути. J Virol. 2011. 85: 8556–68.
CAS PubMed PubMed Central Статья Google Scholar
Леви П.Л., Дюпончель С., Эйшайд Х., Молле Дж., Мишле М., Дисеренс Г. и др. Инфекция вируса гепатита С запускает метаболизм глутамина, подобный опухоли. Гепатология (Балтимор). 2017; 65: 789–803.
Гепатология (Балтимор). 2017; 65: 789–803.
CAS Статья Google Scholar
Rickinson AB. Сопутствующие инфекции, воспаление и онкогенез: будущие направления исследований ВЭБ. Semin Cancer Biol. 2014; 26: 99–115.
CAS PubMed Статья PubMed Central Google Scholar
Lo AK, Dawson CW, Young LS, Ko CW, Hau PM, Lo KW. Активация сигнального пути FGFR1 LMP1, кодируемого вирусом Эпштейна-Барра, способствует аэробному гликолизу и трансформации эпителиальных клеток носоглотки человека.J Pathol. 2015; 237: 238–48.
CAS PubMed Статья PubMed Central Google Scholar
Xiao L, Hu ZY, Dong X, Tan Z, Li W, Tang M, et al. Нацеливание на онкопротеин вируса Эпштейна-Барра, LMP1-опосредованный гликолиз, повышает чувствительность карциномы носоглотки к лучевой терапии. Онкоген. 2014; 33: 4568–78.
2014; 33: 4568–78.
CAS PubMed PubMed Central Статья Google Scholar
Цай Т.Т., Е С.Б., Лю Юн, Хэ Дж, Чен Цюйи, штаб-квартира Май и др. LMP1-опосредованный гликолиз индуцирует рост клеток-супрессоров миелоидного происхождения при карциноме носоглотки. PLoS Pathog. 2017; 13: e1006503.
PubMed PubMed Central Статья CAS Google Scholar
Zhang J, Jia L, Lin W, Yip YL, Lo KW, Lau VM, et al. Латентный мембранный белок 1, кодируемый вирусом Эпштейна-Барра, активирует транскрипцию переносчика глюкозы 1 через сигнальные пути mTORC1 / NF-kappaB.J Virol. 2017; 91: e02168–16.
CAS PubMed PubMed Central Google Scholar
Jiang Y, Yan B, Lai W., Shi Y, Xiao D, Jia J, et al. Репрессия генов Hox с помощью LMP1 при карциноме носоглотки и модуляция генов гликолитического пути с помощью HoxC8. Онкоген. 2015; 34: 6079–91.
Онкоген. 2015; 34: 6079–91.
CAS PubMed PubMed Central Статья Google Scholar
Sung WW, Chu YC, Chen PR, Liao MH, Lee JW. Положительная регуляция экспрессии HIF-1A онкобелком EBV LMP1 в клетках карциномы носоглотки. Cancer Lett. 2016; 382: 21–31.
CAS PubMed Статья PubMed Central Google Scholar
Sung WW, Chen PR, Liao MH, Lee JW. Усиление аэробного гликолиза клеток карциномы носоглотки латентным мембранным белком вируса Эпштейна-Барра 1. Exp Cell Res. 2017; 359: 94–100.
CAS PubMed Статья PubMed Central Google Scholar
Кондо С., Сео С.Ю., Йошизаки Т., Вакисака Н., Фурукава М., Йоаб И. и др. Латентный мембранный белок 1 EBV активирует индуцируемый гипоксией фактор 1альфа посредством Siah2-опосредованного подавления пролилгидроксилаз 1 и 3 в эпителиальных клетках носоглотки. Cancer Res. 2006; 66: 9870–7.
Cancer Res. 2006; 66: 9870–7.
CAS PubMed Статья PubMed Central Google Scholar
Дарекар С., Георгиу К., Юрченко М., Йенамандра С.П., Чачами Г., Симос Г. и др. Иммортализация человеческих В-клеток вирусом Эпштейна-Барра приводит к стабилизации индуцированного гипоксией фактора 1 альфа, что соответствует эффекту Варбурга. PLoS One. 2012; 7: e42072.
CAS PubMed PubMed Central Статья Google Scholar
Лю Х, Ван Дж, Гуо Х, Ву Дж, Цзяо Й, Фалети О.Д. и др. EBV-miR-BART1-5P активирует путь AMPK / mTOR / HIF1 через независимый от PTEN способ, способствуя гликолизу и ангиогенезу при карциноме носоглотки.PLoS Pathog. 2018; 14: e1007484.
PubMed PubMed Central Статья CAS Google Scholar
Daker M, Bhuvanendran S, Ahmad M, Takada K, Khoo AS. Нарушение регуляции генов пути метаболизма липидов в клетках карциномы носоглотки.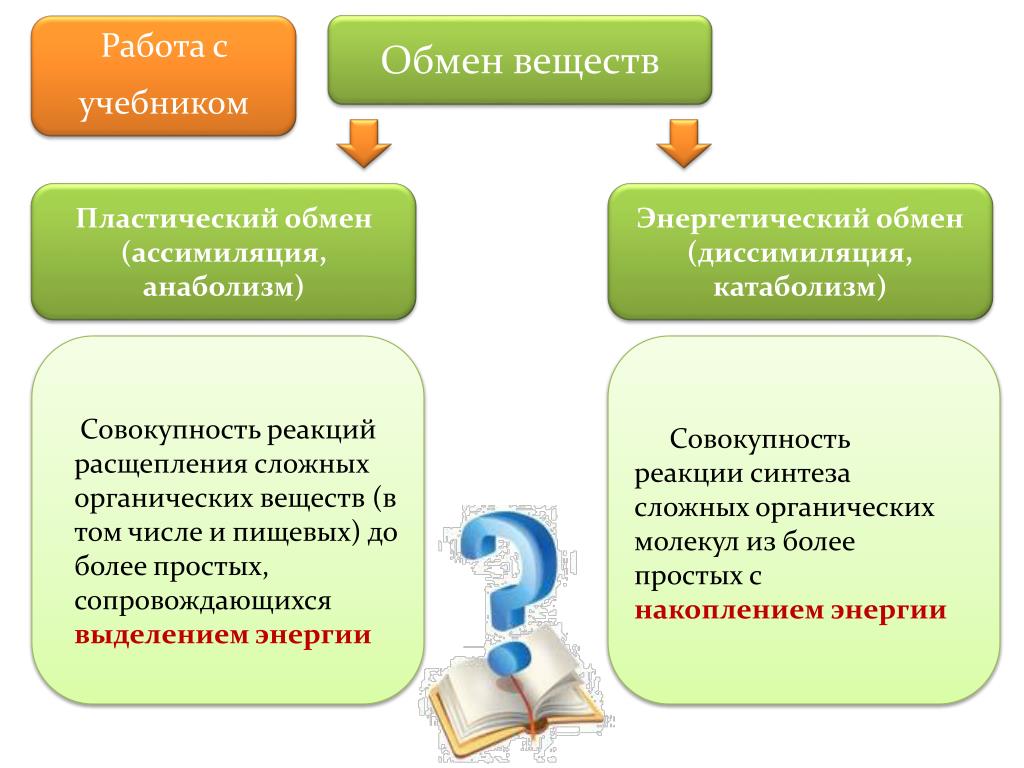 Мол Мед Реп. 2013; 7: 731–41.
Мол Мед Реп. 2013; 7: 731–41.
CAS PubMed Статья PubMed Central Google Scholar
Li Y, Webster-Cyriaque J, Tomlinson CC, Yohe M, Kenney S.Экспрессия синтазы жирных кислот индуцируется немедленным ранним белком вируса Эпштейна-Барра BRLF1 и необходима для литической экспрессии вирусного гена. J Virol. 2004. 78: 4197–206.
CAS PubMed PubMed Central Статья Google Scholar
Месри Э.А., Цезарман Э., Бошофф К. Саркома Капоши и связанный с ней герпесвирус. Нат Рев Рак. 2010; 10: 707–19.
CAS PubMed PubMed Central Статья Google Scholar
Ли С., Бай Л., Донг Дж., Сан Р., Лан К. Вирус герпеса, связанный с саркомой Капоши: эпидемиология и молекулярная биология. Adv Exp Med Biol. 2017; 1018: 91–127.
CAS PubMed Статья PubMed Central Google Scholar

Йогев О., Лагос Д., Энвер Т., Бошофф С. МикроРНК герпесвируса саркомы Капоши индуцируют метаболическую трансформацию инфицированных клеток. PLoS Pathog. 2014; 10: e1004400.
PubMed PubMed Central Статья CAS Google Scholar
Ма Т., Патель Х, Бабапур-Фарроохран С., Франклин Р., Семенца Г. Л., Содхи А. и др. KSHV индуцирует аэробный гликолиз и ангиогенез за счет HIF-1-зависимой активации пируваткиназы 2 в саркоме Капоши. Ангиогенез. 2015; 18: 477–88.
CAS PubMed PubMed Central Статья Google Scholar
Кэрролл П.А., Кенерсон Х.Л., Йунг Р.С., Лагунофф М. Скрытая герпесвирусная инфекция эндотелиальных клеток, ассоциированная с саркомой Капоши, активирует факторы, индуцированные гипоксией.J Virol. 2006; 80: 10802–12.
CAS PubMed PubMed Central Статья Google Scholar
Йогев О., Хендерсон С., Хейс М.Дж., Марелли С.С., Офир-Бирин Ю., Регев-Рудски Н. и др. Герпесвирусы формируют микросреду опухоли посредством экзосомального переноса вирусных микроРНК. PLoS Pathog. 2017; 13: e1006524.
PubMed PubMed Central Статья CAS Google Scholar
Сычев З.Е., Ху А., ДиМайо Т.А., Гиттер А., Кэмп Н.Д., Ноубл В.С. и др. Комплексный системно-биологический анализ латентной инфекции KSHV показывает вирусную индукцию и зависимость от липидного метаболизма, опосредованного пероксисомами. PLoS Pathog. 2017; 13: e1006256.
PubMed PubMed Central Статья CAS Google Scholar
Sharma-Walia N, Chandran K, Patel K, Veettil MV, Marginean A. Каскад 5-липоксигеназы-лейкотриена B4, индуцированный 5-липоксигеназой-лейкотриеном B4, вызванный саркомой Капоши, играет ключевую роль в рекрутинге моноцитов KSHV. и липогенез.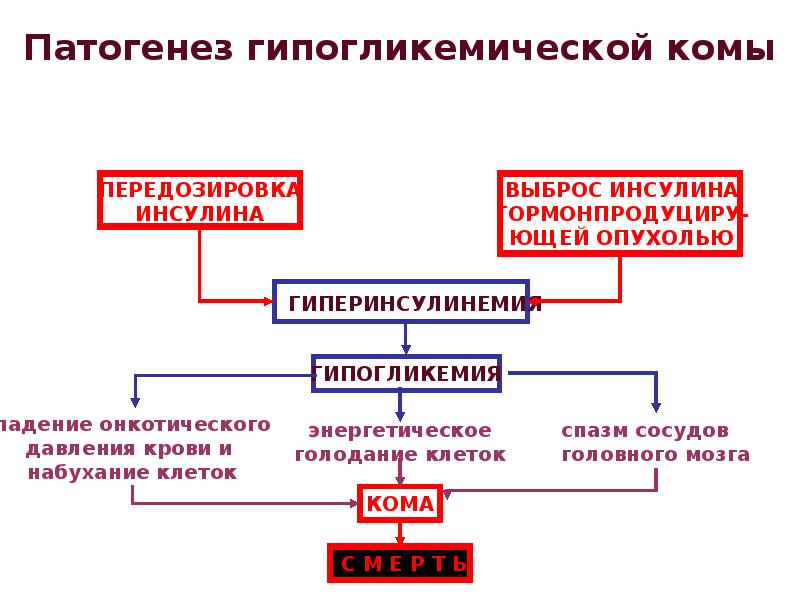 J Virol. 2014; 88: 2131–56.
J Virol. 2014; 88: 2131–56.
PubMed PubMed Central Статья CAS Google Scholar
Serquina AKP, Kambach DM, Sarker O, Ziegelbauer JM. Вирусные микроРНК подавляют путь холестерина, а 25-гидроксихолестерин подавляет инфекцию. mBio. 2017; 8: e00576–17.
PubMed PubMed Central Статья Google Scholar
Sanchez EL, Carroll PA, Thalhofer AB, Lagunoff M.Эндотелиальные клетки, инфицированные латентным KSHV, зависимы от глутамина и требуют глутаминолиза для выживания. PLoS Pathog. 2015; 11: e1005052.
PubMed PubMed Central Статья CAS Google Scholar
Мацуока М., Джанг К.Т. Инфекционность и клеточная трансформация вируса Т-клеточного лейкоза человека типа 1 (HTLV-1). Нат Рев Рак. 2007; 7: 270–80.
CAS PubMed Статья PubMed Central Google Scholar
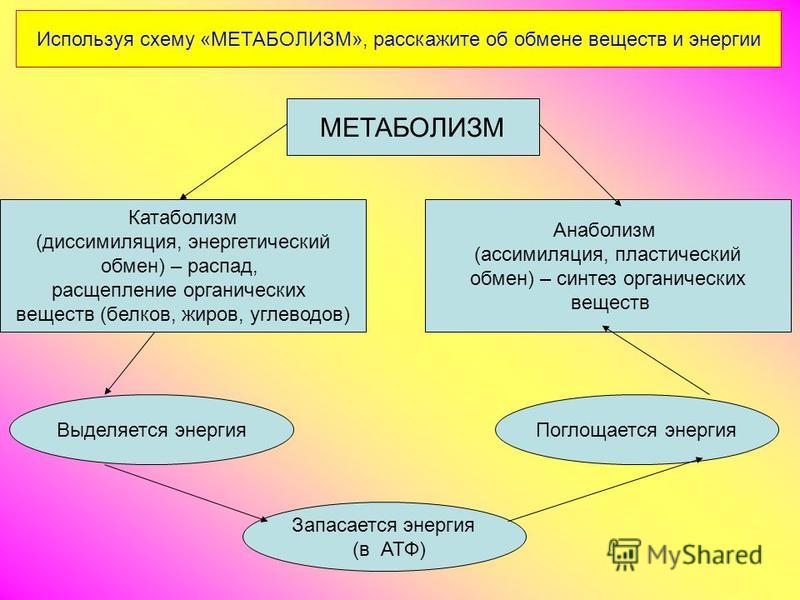
Гессейн А., Кассар О. Эпидемиологические аспекты и мировое распространение инфекции HTLV-1. Front Microbiol. 2012; 3: 388.
PubMed PubMed Central Статья Google Scholar
Кулкарни А., Матеус М., Тиннес С.К., Дж.С. МС, Скофилд С.Дж., Тейлор Г.П. и др. Метаболизм глюкозы и доступность кислорода регулируют реактивацию латентного человеческого ретровируса HTLV-1. Cell Chem Biol. 2017; 24: 1377–87.e3.
CAS PubMed PubMed Central Статья Google Scholar
Манель Н., Ким Ф.Дж., Кинет С., Тейлор Н., Ситбон М., Баттини Дж.Л. Вездесущий переносчик глюкозы GLUT-1 является рецептором для HTLV. Клетка. 2003. 115: 449–59.
CAS PubMed Статья PubMed Central Google Scholar
Coskun AK, Sutton RE. Экспрессия транспортера глюкозы 1 придает чувствительность к слиянию, опосредованному оболочкой вируса Т-клеточного лейкоза человека. J Virol. 2005. 79: 4150–8.
J Virol. 2005. 79: 4150–8.
CAS PubMed PubMed Central Статья Google Scholar
Толстов Ю.Л., Пастрана Д.В., Фен Х., Беккер Дж. К., Дженкинс Ф. Дж., Мошос С. и др. Человеческая полиомавирусная инфекция клеток Меркеля II. MCV — это обычная человеческая инфекция, которая может быть обнаружена с помощью иммуноанализа конформационного эпитопа капсида. Int J Cancer. 2009; 125: 1250–6.
CAS PubMed PubMed Central Статья Google Scholar
Берриос К., Пади М., Кейблер М.А., Парк ДЭ, Молла В., Ченг Дж. И др. Малый Т-антиген полиомавируса клеток Меркеля способствует прогликолитическим метаболическим нарушениям, необходимым для трансформации.PLoS Pathog. 2016; 12: e1006020.
PubMed PubMed Central Статья CAS Google Scholar
Макфадден К., Хафез А.Ю., Киштон Р. , Мессинджер Дж. Э., Никитин П.А., Ратмелл Дж. К. и др. Метаболический стресс является препятствием для иммортализации B-клеток, опосредованной вирусом Эпштейна-Барра. Proc Natl Acad Sci U S. A. 2016; 113: E782–90.
, Мессинджер Дж. Э., Никитин П.А., Ратмелл Дж. К. и др. Метаболический стресс является препятствием для иммортализации B-клеток, опосредованной вирусом Эпштейна-Барра. Proc Natl Acad Sci U S. A. 2016; 113: E782–90.
CAS PubMed PubMed Central Статья Google Scholar
Kraus RJ, Yu X, Cordes BA, Sathiamoorthi S, Iempridee T, Nawandar DM, et al. Индуцируемый гипоксией фактор-1альфа играет роль в естественном жизненном цикле и туморогенезе вируса Эпштейна-Барра, индуцируя литическую инфекцию посредством прямого связывания с промотором немедленного раннего гена BZLF1. PLoS Pathog. 2017; 13: e1006404.
PubMed PubMed Central Статья CAS Google Scholar
Дэвис Д.А., Риндеркнехт А.С., Зетевей Дж. П., Аоки Й., Рид-Коннол Е.Л., Тосато Г. и др.Гипоксия вызывает литическую репликацию герпесвируса, связанного с саркомой Капоши. Кровь. 2001; 97: 3244–50.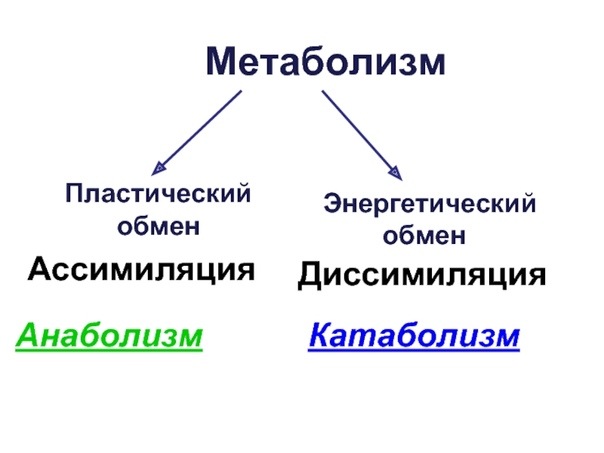
CAS PubMed Статья PubMed Central Google Scholar
Кай Кью, Лан К, Верма СК, Си Х, Лин Д., Робертсон Э.С. Латентный белок LANA герпесвируса, ассоциированный с саркомой Капоши, взаимодействует с HIF-1 альфа для усиления экспрессии RTA во время гипоксии: контроль латентности в условиях низкого содержания кислорода. J Virol. 2006; 80: 7965–75.
CAS PubMed PubMed Central Статья Google Scholar
Sanchez EL, Pulliam TH, Dimaio TA, Thalhofer AB, Delgado T, Lagunoff M. Гликолиз, глутаминолиз и синтез жирных кислот необходимы для различных стадий литической репликации герпесвируса, ассоциированного с саркомой Капоши. J Virol. 2017; 91: e02237–16.
PubMed PubMed Central Статья Google Scholar
Delgado T, Carroll PA, Punjabi AS, Margineantu D, Hockenbery DM, Lagunoff M. Индукция эффекта Варбурга вирусом герпеса саркомы Капоши необходима для поддержания латентно инфицированных эндотелиальных клеток. Proc Natl Acad Sci U S. A. 2010; 107: 10696–701.
Индукция эффекта Варбурга вирусом герпеса саркомы Капоши необходима для поддержания латентно инфицированных эндотелиальных клеток. Proc Natl Acad Sci U S. A. 2010; 107: 10696–701.
CAS PubMed PubMed Central Статья Google Scholar
Ло А.К., Доусон К.В., Янг Л.С., Ло К.В. Роль метаболического репрограммирования в онкогенезе, связанном с гамма-герпесвирусом.Int J Cancer. 2017; 141: 1512–21.
CAS PubMed Статья PubMed Central Google Scholar
Bagga S, Rawat S, Ajenjo M, Bouchard MJ. Опосредованная вирусом гепатита B (HBV) X регуляция метаболических путей гепатоцитов влияет на репликацию вируса. Вирусология. 2016; 498: 9–22.
CAS PubMed Статья PubMed Central Google Scholar
Mushtaq M, Darekar S, Klein G, Kashuba E. Различные механизмы регуляции эффекта Варбурга в лимфобластоидных клетках и клетках лимфомы Беркитта. PLoS One. 2015; 10: e0136142.
PLoS One. 2015; 10: e0136142.
PubMed PubMed Central Статья CAS Google Scholar
O’Shea CC. Вирусы: инструменты для обнаружения опухолевых мишеней и агенты для онколитической терапии — введение. Онкоген. 2005; 24: 7636–9.
CAS PubMed Статья PubMed Central Google Scholar
Krall AS, Xu S, Graeber TG, Braas D, Christofk HR. Аспарагин способствует пролиферации раковых клеток за счет использования в качестве фактора обмена аминокислот. Nat Commun. 2016; 7: 11457.
CAS PubMed PubMed Central Статья Google Scholar
Кауфман Х.Л., Колхапп Ф.Дж., Злоза А. Онколитические вирусы: новый класс иммунотерапевтических препаратов. Nat Rev Drug Discov. 2015; 14: 642–62.
CAS PubMed Статья PubMed Central Google Scholar

Сеймур Л.В., Фишер К.Д. Онколитические вирусы: наконец-то появились. Британский Дж. Рак. 2016; 114: 357–61.
CAS PubMed Статья PubMed Central Google Scholar
Ирвин ЧР, Хитт М.М., Эванс Д.Х. Нацеливание на биосинтез нуклеотидов: стратегия улучшения онколитического потенциала ДНК-вирусов. Фасад Онкол. 2017; 7: 229.
PubMed PubMed Central Статья Google Scholar
МЕТАБОЛИЗМ
МЕТАБОЛИЗМОбмен веществ
Метаболизм — это слово, которое мы используем для управления материальными и энергетическими ресурсами.Энергия может быть получена путем разрушения сложных молекул (катаболизм) или энергия может быть использована для создания сложных молекул (анаболизм). Анаболические и катаболические процессы часто сочетаются таким образом, что энергия, полученная в результате анаболизма, может быть использована для катаболизма.
Несколько слов об энергии:
Энергию можно передавать. Это не может быть создано. Передача энергии всегда неэффективна — некоторая часть энергии всегда теряется в виде тепла. Бесплатная энергия — это энергия, доступная для работы. Есть два типа химических реакций.Экзергонические реакции протекают с чистым высвобождением свободной энергии. Эндергонические реакции поглощают свободную энергию.
Я знаю, что говорил вам, что вы не будете нести ответственности за термины экзэргонический и эндергонический, но вам все равно полезно знать их, чтобы вы могли прочитать остальную часть этих примечаний.
Другой способ представить себе эти реакции — рассмотреть относительную потенциальную энергию продуктов и реагентов (5,3 стр. 74). Эндергонические реакции требуют затрат энергии для получения простых реагентов с низким энергопотреблением и создания сложных высокоэнергетических продуктов.Экзергонические реакции высвобождают энергию, связанную с реагентами, и дают более простые низкоэнергетические продукты. Ключевой стратегией в управлении эндергоническими реакциями является их соединение с экзэргоническими реакциями через энергетический челнок, называемый АТФ.
Ключевой стратегией в управлении эндергоническими реакциями является их соединение с экзэргоническими реакциями через энергетический челнок, называемый АТФ.
Зачем нам эта энергия? Ячейка выполняет три вида работы:
1. Механические работы. Примеры: биение ресничек, мышечные сокращения, внутриклеточные движения.
2. Транспортные работы. Примеры: перекачка веществ через градиенты.
3. Химические работы. Примеры: синтез полимеров из мономеров.
Непосредственным источником энергии для этой работы, этого так называемого энергетического челнока, является АТФ, аденозинтрифосфат.
ATP
АТФ представляет собой нуклеозидтрифосфат, состоящий из аденина, связанного с рибозой, который связан с тремя фосфатными группами (5.4a, стр. 75).
Когда фосфатная группа отрывается от хвоста молекулы АТФ (путем гидролиза), молекула становится АДФ (аденозиндифосфат).Этот гидролиз является экзергонической реакцией и дает энергию. Связи, удерживающие фосфат на АТФ, слабые. Они известны как высокоэнергетические связи, но не потому, что они прочные (если бы они были прочными, потребовалось бы много энергии, чтобы их разорвать. Думайте об АТФ как о подпружиненной молекуле с последним фосфатом, только что застрявшим на конце).
Они известны как высокоэнергетические связи, но не потому, что они прочные (если бы они были прочными, потребовалось бы много энергии, чтобы их разорвать. Думайте об АТФ как о подпружиненной молекуле с последним фосфатом, только что застрявшим на конце).
Когда фосфат удаляется из АТФ, он добавляется к молекуле, которая является частью эндергонической реакции, в которой мы заинтересованы.Теперь эта молекула нестабильна (то есть более реактивна), поэтому некоторая энергия стала доступной для эндергонической реакции. Молекула, к которой добавлена фосфатная группа, называется фосфорилированным промежуточным продуктом.
АТФ регенерируется посредством клеточного дыхания, при котором энергия глюкозы используется для фосфорилирования АДФ с образованием АТФ. Растения также могут использовать энергию света для производства АТФ.
Клеточное дыхание
Жизнь — это работа. Клетки всегда работают: строят молекулы, накачивают ионы, движутся и т. Д.Для этой работы клеткам нужна энергия из внешних источников. Энергия поступает в (большинство) экосистем в виде солнечной энергии, а затем растения превращают ее в химическую энергию. Все остальные организмы получают энергию из пищи, которая восходит к этим растениям.
Энергия поступает в (большинство) экосистем в виде солнечной энергии, а затем растения превращают ее в химическую энергию. Все остальные организмы получают энергию из пищи, которая восходит к этим растениям.
Клетки высвобождают энергию, связанную с пищей, в результате систематического разложения молекул пищи на простые низкоэнергетические отходы. Часть химической энергии используется для работы, часть теряется в виде тепла. Пути распада называются катаболическими путями.Одним из таких катаболических путей является ферментация (без кислорода). Более распространенный и более эффективный путь называется клеточным дыханием. Кислород соединяется с органическими молекулами, выделяя энергию.
Органические соединения + кислород
CO2 + вода + энергияВсе типы макромолекул могут быть разрушены и использованы в качестве топлива. Обычно мы изучаем разложение глюкозы:
C 6 H 12 O 6 +6 O 2
6 CO 2 + 6 H 2 O + энергия (ATP + тепло) Мы склонны потреблять белки, углеводы и жиры с пищей. Если мы используем разные типы молекул для определения их энергетического содержания (в отличие от их
Если мы используем разные типы молекул для определения их энергетического содержания (в отличие от их
Клеточное дыхание не перемещает непосредственно жгутики, не перекачивает растворенные вещества и не выполняет какую-либо клеточную работу. Клеточное дыхание генерирует АТФ, который, в свою очередь, расходуется клеткой для выполнения работы. Помните, что ATP подобен нагруженной пружине.Фосфаты застревают на конце АДФ с образованием АТФ. Этот процесс называется фосфорилированием.
Дыхание делится на три стадии: гликолиз, цикл Кребса и система переноса электронов. Вы должны прочитать модули 6.1 и 6.2 в своем учебнике. Они в значительной степени охватывают то, что я сказал в классе о клеточном дыхании. Обратите внимание, что даже несмотря на то, что мы считаем клеточное дыхание эффективным способом преобразования энергии глюкозы в АТФ (полезную форму), мы все равно не получаем
= t получаем ее полностью.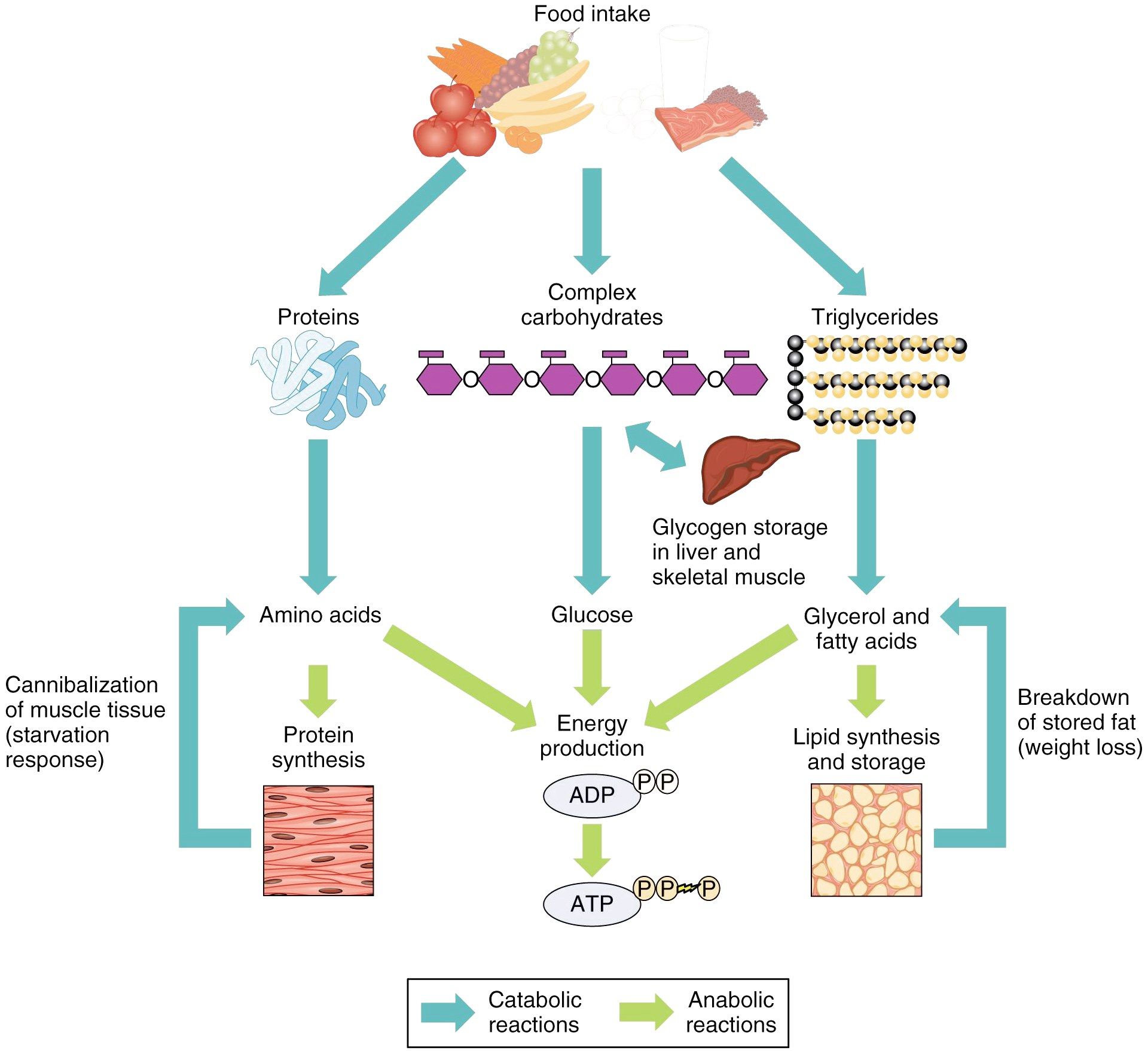 (Куда уходит остальная энергия?)
(Куда уходит остальная энергия?)6.3 говорит о том, сколько энергии мы используем для различных видов деятельности. Это может вас заинтересовать. Обратите внимание, что они используют термин
AK-cal @ или Akilocalorie @. Это эквивалент того, что мы в повседневной жизни называем калорийностью.Ферменты
Ферменты — катализаторы. Катализаторы — это химические вещества, которые изменяют скорость реакции, но не расходуются на нее. Это то, что регулирует различные реакции обмена веществ.
Несмотря на то, что многие реакции являются экзергоническими, они все же требуют некоторой энергии для их запуска.Этот дополнительный импульс называется энергией активации. Ферменты действуют, чтобы снизить энергию активации, необходимую для запуска реакций. Они не влияют на изменение чистой энергии уравнения. (5.5a и b, стр. 76)
Реагент, над которым работает фермент, называется его субстратом. Ферменты очень специфичны для субстрата. Обычно это белки. Напомним, что белки имеют определенную форму. Есть область фермента, называемая активным центром, куда подходят молекулы субстрата. Катализатор (фермент) может выполнять ряд функций, облегчающих реакцию.Он может скручивать молекулы, способствуя разрыву связи, он может служить шаблоном для размещения молекул субстрата в правильном положении, он может обеспечивать «микросреду», способствующую ускорению реакции (например, pH). (5,6, стр 77)
Есть область фермента, называемая активным центром, куда подходят молекулы субстрата. Катализатор (фермент) может выполнять ряд функций, облегчающих реакцию.Он может скручивать молекулы, способствуя разрыву связи, он может служить шаблоном для размещения молекул субстрата в правильном положении, он может обеспечивать «микросреду», способствующую ускорению реакции (например, pH). (5,6, стр 77)
На действие ферментов влияют многие вещи. Большинство ферментов функционируют в определенном оптимальном диапазоне температуры и pH. Многие ферменты требуют помощи кофакторов. Ингибиторы ферментов могут мешать действию фермента, связываясь (постоянно или временно) с участком на ферменте.Ингибитор может прикрепляться к активному сайту, блокируя субстраты (так называемый конкурентный ингибитор), или он может прикрепляться в другом месте, вызывая изменение формы (следовательно, функции) фермента (неконкурентный ингибитор) (5.8, стр. 78). Производство ферментов, кофакторов, ингибиторов и т. Д.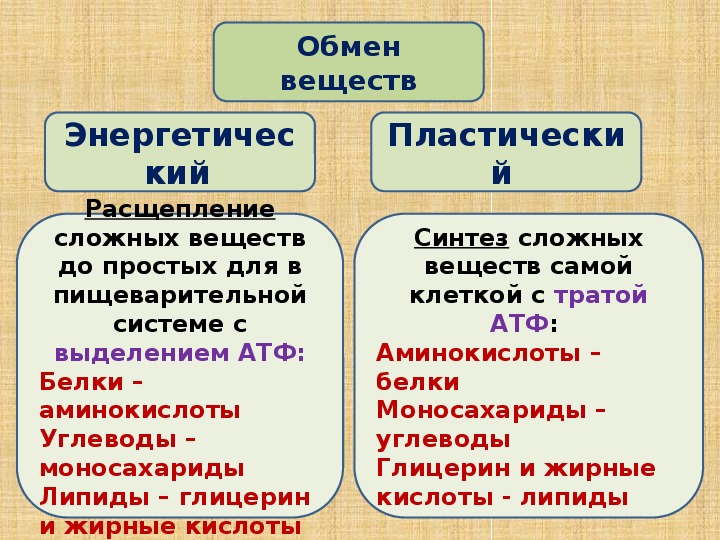 — это средства, с помощью которых организм контролирует метаболизм.
— это средства, с помощью которых организм контролирует метаболизм.
Новое исследование показывает, как сложный метаболизм мог образоваться из простых предшественников — ScienceDaily
Вся современная жизнь использует энергию для воспроизводства.Во время этого процесса организмы строят и расщепляют более крупные молекулы, такие как жиры и сахара, используя замечательно общий набор реактивных промежуточных молекул-носителей энергии. Эти промежуточные носители энергии (например, АТФ) сами по себе часто не являются строительными блоками, но они позволяют объединить энергию между отдельными реакциями, необходимыми для ускорения клеточного воспроизводства. Одним из классов этих соединений являются тиоэфиры, химические соединения, содержащие высокоэнергетическую связь углерод-сера. В течение некоторого времени предполагалось, что тиоэфиры могут быть одними из самых древних версий таких метаболических реактивных промежуточных продуктов, отчасти потому, что современные организмы все еще используют тиоэфиры, чтобы помочь расщепить сахара и сделать белки из аминокислот. То, как реакционноспособные промежуточные соединения, такие как тиоэфиры, могли образоваться до того, как появилась жизнь или когда ранняя жизнь делала свои первые шаги, остается тайной. Новая работа исследователей из Института наук о Земле (ELSI) при Токийском технологическом институте показывает, что простое соединение, которое можно найти в некоторых современных вулканических газах, — тиокислота (соединение, образованное из органической кислоты и сероводорода, это соединение, в значительной степени ответственное за запах «тухлых яиц» многих горячих источников) — легко вступает в реакцию с простыми серосодержащими тиоловыми соединениями, подобными тем, которые имеют фундаментальное значение для современного метаболизма, с образованием реактивных тиоэфиров, подобных тем, которые обнаруживаются в современной биологии.Эта реакция легко происходит в воде и, возможно, послужила отправной точкой для эволюции более сложной биохимии. В ходе исследования они обнаружили, что, когда они включали в свои реакции железо (которое является наиболее распространенным элементом на Земле и составляет около 5% поверхностного слоя Земли, известного как кора), выход продуктов реакции увеличивается.
То, как реакционноспособные промежуточные соединения, такие как тиоэфиры, могли образоваться до того, как появилась жизнь или когда ранняя жизнь делала свои первые шаги, остается тайной. Новая работа исследователей из Института наук о Земле (ELSI) при Токийском технологическом институте показывает, что простое соединение, которое можно найти в некоторых современных вулканических газах, — тиокислота (соединение, образованное из органической кислоты и сероводорода, это соединение, в значительной степени ответственное за запах «тухлых яиц» многих горячих источников) — легко вступает в реакцию с простыми серосодержащими тиоловыми соединениями, подобными тем, которые имеют фундаментальное значение для современного метаболизма, с образованием реактивных тиоэфиров, подобных тем, которые обнаруживаются в современной биологии.Эта реакция легко происходит в воде и, возможно, послужила отправной точкой для эволюции более сложной биохимии. В ходе исследования они обнаружили, что, когда они включали в свои реакции железо (которое является наиболее распространенным элементом на Земле и составляет около 5% поверхностного слоя Земли, известного как кора), выход продуктов реакции увеличивается.
Команда предполагает, что это может означать, что энергетическая связь между множественными реакциями, в которых одна реакция движет вперед другой, может иметь свои корни в химии неживой окружающей среды.Примечательно, что они также обнаружили, что побочный продукт реакции, которую они обнаружили, можно использовать для создания второго типа универсального соединения, связывающего энергию, которое требуется всем живым существам: кластеров FeS (сокращение от железо-сера). Это небольшие агрегаты, состоящие всего из нескольких атомов железа и серы, которые помогают организмам метаболизировать, перемещая электроны от одной молекулы к другой. Одним из важных примеров такого пути использования кластеров FeS является фотосинтез, который переносит электроны от воды на CO 2 для производства сахаров и кислорода.Таким образом, эта работа дает новое понимание того, как молекулы с высокой энергией и реакции переноса электронов могли возникать естественным образом в процессе раннего развития метаболизма.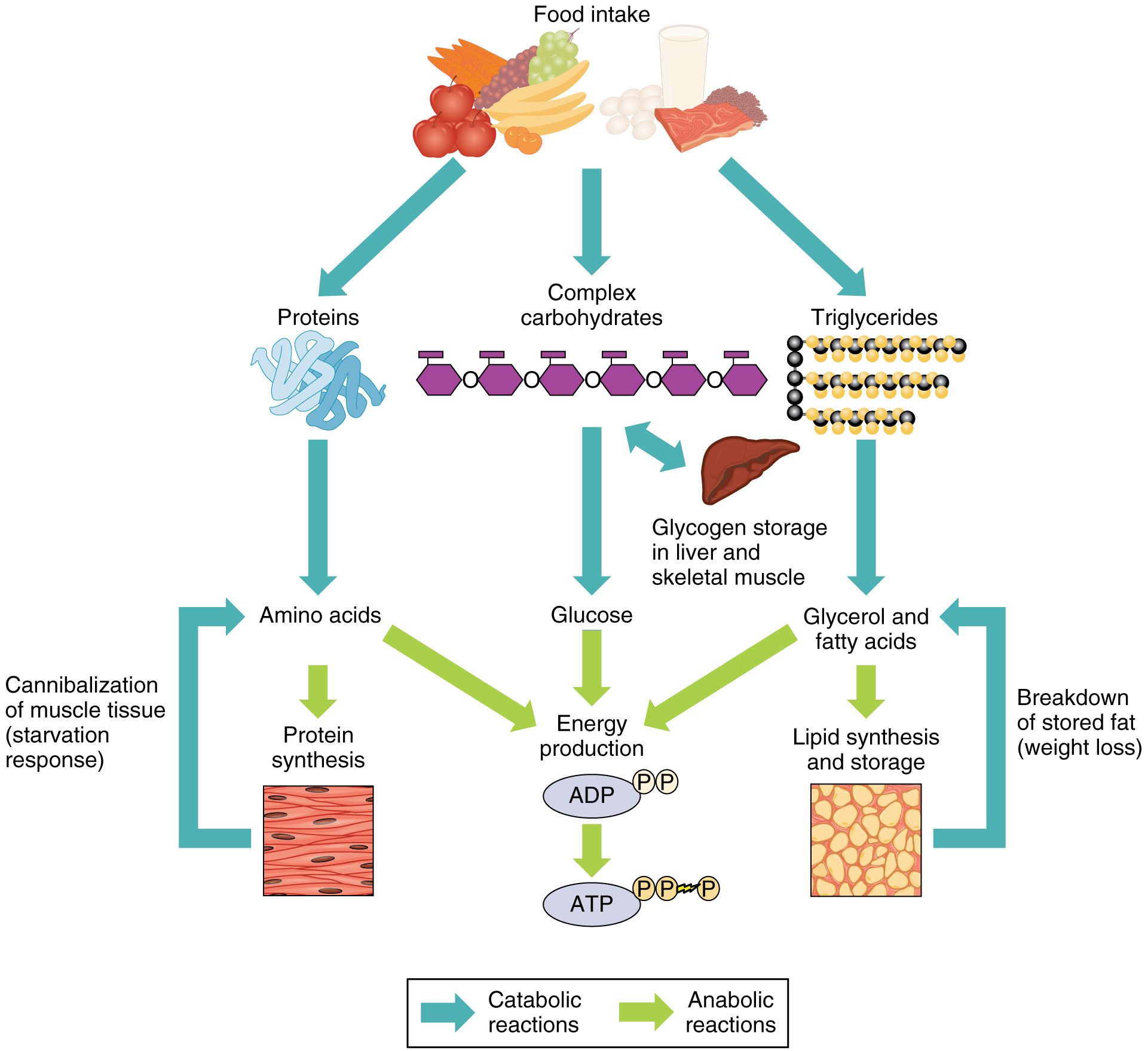
Хотя ученые пытались понять происхождение строительных блоков жизни, таких как аминокислоты, нуклеотиды, пептиды и т. Д., В течение некоторого времени мало усилий было приложено для понимания того, как передача энергии возникла в химии пребиотиков. Понимание этого обмена энергией может быть столь же важным, как и понимание происхождения строительных блоков, поэтому команда ELSI решила поискать реакции, которые могли бы быть связаны друг с другом энергетически.Как отмечает ведущий автор, Себастьян Санден: «Мы уже изучали минералы FeS и знали, насколько легко их образование, поэтому мы хотели посмотреть, сможем ли мы связать эту избыточную потерянную энергию с другой реакцией». Тиокислота, с которой они начали свое исследование, содержит серу, которая, как они знали, должна была реагировать только с железом, чтобы образовались кластеры FeS, которые они уже изучали.
Эксперименты и анализы, которые проводили исследователи ELSI, должны были проводиться в быстрой последовательности, чтобы отслеживать ход реакции.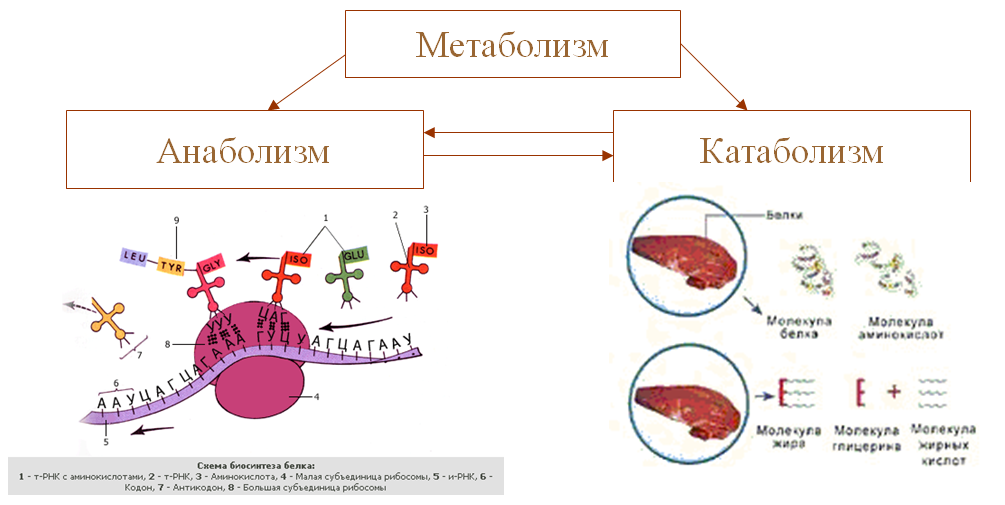 Они разработали методы для этого и, таким образом, смогли определить, насколько быстро эти реакции произошли. Их предварительные эксперименты по получению тиоэфира не прошли так быстро, как они первоначально надеялись, но, добавив катализатор и повысив температуру, они обнаружили, что максимальный выход тиоэфира был получен менее чем за час, а не за несколько дней до внесения этих изменений. .
Они разработали методы для этого и, таким образом, смогли определить, насколько быстро эти реакции произошли. Их предварительные эксперименты по получению тиоэфира не прошли так быстро, как они первоначально надеялись, но, добавив катализатор и повысив температуру, они обнаружили, что максимальный выход тиоэфира был получен менее чем за час, а не за несколько дней до внесения этих изменений. .
Команда считает особенно интересным то, что такие реакции могут создавать «каскадные реакции», в результате которых образуются все более и более сложные молекулы: пируват разлагается, помогая образовывать тиоэфир, который затем позволяет пептидам (меньшим родственникам белков) образовываться через новые открыл путь тиоэфира.Команда надеется протестировать это экспериментально и создать систему, которая сама по себе может увеличить количество содержащихся в ней компонентов и их сложность, возможно, вплоть до самовоспроизведения. Некоторые современные микробы фактически используют разложение пирувата и образование тиоэфиров с помощью кластеров FeS в своем метаболизме, и возможно, что реакции, обнаруженные командой, повторяют то, как их открыла ранняя пребиологическая или биологическая эволюция. Как сказал ведущий исследователь этой работы, доцент ELSI Шон МакГлинн: «Эта работа обеспечивает новые связи между множественными компонентами пребиотической реакции, которые могли иметь решающее значение для установления раннего энергетического метаболизма на Земле.«
Как сказал ведущий исследователь этой работы, доцент ELSI Шон МакГлинн: «Эта работа обеспечивает новые связи между множественными компонентами пребиотической реакции, которые могли иметь решающее значение для установления раннего энергетического метаболизма на Земле.«
Несмотря на то, что эта работа может пролить новый свет на то, как естественные реакции обмена энергии, возможно, помогли «запустить» метаболизм, она также может быть важна для области «зеленой химии», которая занимается поиском наиболее энергетически эффективных и экологически безопасных дружественные методы для создания химических соединений. В то время как токсичные тяжелые металлы, такие как кадмий и ртуть, и растворители, такие как хлороформ, часто используются в промышленной органической химии, реакции, обнаруженные этой исследовательской группой, очень эффективны и работают в воде с использованием нетоксичного железа в качестве катализатора.
История Источник:
Материалы предоставлены Токийским технологическим институтом . Примечание. Содержимое можно редактировать по стилю и длине.
Примечание. Содержимое можно редактировать по стилю и длине.
Типы, причины, симптомы и методы лечения
Унаследованные метаболические нарушения — это генетические состояния, которые приводят к нарушениям обмена веществ. У большинства людей с наследственными метаболическими нарушениями дефектный ген приводит к дефициту ферментов. Существуют сотни различных генетических нарушений обмена веществ, симптомы, методы лечения и прогнозы которых сильно различаются.
Что такое метаболизм?
Метаболизм — это все химические реакции, происходящие в организме для преобразования или использования энергии. Вот несколько основных примеров метаболизма:
- Расщепление углеводов, белков и жиров в пище для высвобождения энергии.
- Преобразование избыточного азота в продукты жизнедеятельности, выделяемые с мочой.
- Разрушение или преобразование химических веществ в другие вещества и транспортировка их внутрь клеток.
Метаболизм — это организованная, но хаотическая линия сборки химических веществ. Сырье, полуфабрикаты и отходы постоянно используются, производятся, транспортируются и выбрасываются. «Рабочие» на конвейере — это ферменты и другие белки, которые вызывают химические реакции.
Сырье, полуфабрикаты и отходы постоянно используются, производятся, транспортируются и выбрасываются. «Рабочие» на конвейере — это ферменты и другие белки, которые вызывают химические реакции.
Причины наследственных нарушений обмена веществ
При большинстве наследственных метаболических нарушений один фермент либо вообще не вырабатывается организмом, либо вырабатывается в неработающей форме. Недостающий фермент подобен отсутствующему работнику на конвейере.В зависимости от функции этого фермента его отсутствие означает, что могут накапливаться токсичные химические вещества или может не производиться необходимый продукт.
Код или план производства фермента обычно содержится в паре генов. Большинство людей с наследственными нарушениями обмена веществ наследуют две дефектные копии гена — по одной от каждого родителя. Оба родителя являются «носителями» плохого гена, что означает, что они несут одну дефектную копию и одну нормальную копию.
У родителей нормальная копия гена компенсирует плохую копию. Уровень их ферментов обычно адекватен, поэтому у них может не быть симптомов генетического нарушения обмена веществ. Однако ребенок, унаследовавший две дефектные копии гена, не может производить достаточно эффективных ферментов, и у него развивается генетическое нарушение обмена веществ. Эта форма генетической передачи называется аутосомно-рецессивным наследованием.
Уровень их ферментов обычно адекватен, поэтому у них может не быть симптомов генетического нарушения обмена веществ. Однако ребенок, унаследовавший две дефектные копии гена, не может производить достаточно эффективных ферментов, и у него развивается генетическое нарушение обмена веществ. Эта форма генетической передачи называется аутосомно-рецессивным наследованием.
Первопричиной большинства генетических нарушений обмена веществ является мутация гена, произошедшая много-много поколений назад. Мутация гена передается из поколения в поколение, обеспечивая ее сохранение.
Каждое наследственное нарушение обмена веществ довольно редко встречается в общей популяции. В совокупности наследственные нарушения обмена веществ могут поражать примерно 1 из 1000–2 500 новорожденных. У некоторых этнических групп населения, таких как евреи-ашкенази (евреи центрально-восточноевропейского происхождения), частота наследственных нарушений обмена веществ выше.
Типы наследственных нарушений обмена веществ
Выявлены сотни наследственных нарушений обмена веществ, и продолжают обнаруживаться новые. Некоторые из наиболее распространенных и важных генетических нарушений метаболизма включают:
Некоторые из наиболее распространенных и важных генетических нарушений метаболизма включают:
Лизосомные нарушения накопления : Лизосомы — это пространства внутри клеток, которые расщепляют отходы метаболизма. Дефицит различных ферментов внутри лизосом может привести к накоплению токсичных веществ, вызывая нарушения обмена веществ, в том числе:
- Синдром Гурлера (аномальная структура костей и задержка развития)
- Болезнь Ниманна-Пика (у детей развивается увеличение печени, затруднения при кормлении и повреждение нервов)
- Болезнь Тея-Сакса (прогрессирующая слабость у месячного ребенка, прогрессирующая до тяжелого повреждения нервов; ребенок обычно доживает до 4 или 5 лет)
- Болезнь Гоше (боль в костях, увеличение печени и низкое количество тромбоцитов, часто легкая форма, у детей или взрослых)
- Болезнь Фабри (боль в конечностях в детстве, при заболеваниях почек и сердца и инсультах во взрослом возрасте; поражаются только мужчины)
- Болезнь Краббе (прогрессирующее поражение нервов, задержка развития у маленьких детей; иногда взрослые)
Галактоземия: Нарушение расщепления сахарной галактозы приводит к желтухе, рвоте и поражению печени увеличение новорожденного после грудного или искусственного вскармливания.
Болезнь мочи кленовым сиропом: Дефицит фермента BCKD вызывает накопление аминокислот в организме. В результате повреждаются нервы, и моча пахнет сиропом.
Фенилкетонурия (PKU): Дефицит фермента PAH приводит к высокому уровню фенилаланина в крови. Если заболевание не распознается, наступает умственная отсталость.
Болезни накопления гликогена: Проблемы с накоплением сахара приводят к низкому уровню сахара в крови, болям в мышцах и слабости.
Митохондриальные нарушения: Проблемы внутри митохондрий, электростанций клеток, приводят к повреждению мышц.
Атаксия Фридрейха: Проблемы, связанные с белком под названием фратаксин, вызывают повреждение нервов и часто проблемы с сердцем. Неспособность ходить обычно наступает в молодом возрасте.
Пероксисомные расстройства: Как и лизосомы, пероксисомы представляют собой крошечные пространства, заполненные ферментами внутри клеток. Плохая ферментативная функция пероксисом может привести к накоплению токсичных продуктов метаболизма. К пероксисомальным расстройствам относятся:
К пероксисомальным расстройствам относятся:
- Синдром Зеллвегера (аномальные черты лица, увеличенная печень и повреждение нервов у младенцев)
- Адренолейкодистрофия (симптомы повреждения нервов могут развиться в детстве или в раннем взрослом возрасте в зависимости от формы.)
Метаболизм металлов нарушения: Уровни микроэлементов в крови контролируются специальными белками. Унаследованные метаболические нарушения могут привести к нарушению функции белков и токсическому накоплению металлов в организме:
Органические ацидемии: метилмалоновая ацидемия и пропионовая ацидемия.
Нарушения цикла мочевины: Дефицит орнитин-транскарбамилазы и цитруллинемия
Симптомы наследственных нарушений обмена веществ
Симптомы генетических нарушений обмена веществ широко варьируются в зависимости от имеющихся проблем метаболизма. Некоторые симптомы наследственных нарушений обмена веществ включают:
Симптомы могут возникать внезапно или медленно прогрессировать. Симптомы могут быть вызваны продуктами питания, лекарствами, обезвоживанием, незначительными заболеваниями или другими факторами. Симптомы появляются в течение нескольких недель после рождения при многих заболеваниях.Симптомы других наследственных метаболических нарушений могут проявиться через годы.
Диагностика наследственных нарушений обмена веществ
Наследственные нарушения обмена веществ присутствуют при рождении, а некоторые выявляются при плановом обследовании. Все 50 штатов проводят скрининг новорожденных на фенилкетонурию (ФКУ). Большинство штатов также проверяют новорожденных на галактоземию. Однако государственных тестов на все известные наследственные нарушения обмена веществ у младенцев не проводилось.
Усовершенствованная технология тестирования побуждает многие штаты расширять скрининг новорожденных на генетические нарушения обмена веществ.Национальный центр скрининга новорожденных и генетических ресурсов предоставляет информацию о методах скрининга в каждом штате.
Если наследственное нарушение обмена веществ не обнаруживается при рождении, его часто не диагностируют до появления симптомов. При появлении симптомов доступны специальные анализы крови или ДНК для диагностики большинства генетических нарушений обмена веществ. Направление в специализированный центр (обычно при университете) увеличивает шансы на постановку правильного диагноза.
Лечение наследственных нарушений обмена веществ
Доступны ограниченные методы лечения наследственных нарушений обмена веществ.Существенный генетический дефект, вызывающий это состояние, не может быть исправлен с помощью современных технологий. Вместо этого лечение пытается обойти проблему с метаболизмом.
Лечение генетических нарушений метаболизма основывается на нескольких общих принципах:
- Уменьшите или исключите потребление любых продуктов питания или лекарств, которые не могут метаболизироваться должным образом.
- Замените фермент или другое химическое вещество, которое отсутствует или неактивно, чтобы восстановить метаболизм, максимально приближенный к нормальному.
- Удаляет токсичные продукты метаболизма, которые накапливаются из-за нарушения обмена веществ.
Лечение может включать такие меры, как:
- Специальные диеты, исключающие определенные питательные вещества
- Прием заменителей ферментов или других добавок, поддерживающих метаболизм
- Обработка крови химическими веществами для детоксикации опасных побочных продуктов метаболизма
По возможности , человек с наследственным нарушением обмена веществ должен получить помощь в медицинском центре, имеющем опыт лечения этих редких состояний.
Дети и взрослые с наследственными нарушениями обмена веществ могут серьезно заболеть, требуя госпитализации, а иногда и жизнеобеспечения.Лечение во время этих эпизодов направлено на оказание неотложной помощи и улучшение функции органов.
Определение, типы, биология и метаболическая активность »
Метаболизм — это химический процесс в нашем организме, и посредством этого процесса наше тело получает энергию для выполнения различных действий. Проще говоря, мы можем сказать, что мы получаем энергию от то, что мы едим и пьем, а также химический процесс внутри нашего тела, который помогает преобразовывать пищу в энергию, известен как метаболизм.
(Изображение скоро будет обновлено)
Метаболизм происходит во всех живых организмах, и это необходимо для поддержания жизни.Мы все знаем, что всем живым организмам требуется энергия для выполнения различных процессов и действий, а метаболические процессы помогают поддерживать различные структуры живых организмов.
Типы метаболизма
Метаболизм состоит из двух разных процессов:
Катаболизм: расщепление веществ для получения энергии или структуры, чтобы вернуться к строительным блокам.
Анаболизм: восстанавливайте вещества так, как мы считаем их полезными.
Катаболические и анаболические пути
Метаболический путь — это серия связанных химических реакций, которые включают образование и расщепление молекул глюкозы и высвобождение или использование энергии. Путь наращивания — это анаболический путь, тогда как разрушение путей — это катаболический путь.
Давайте обсудим эти два пути подробно:
Анаболический путь
Он помогает в построении сложных молекул из более простых молекул, и здесь расходуется энергия i.е. это требует ввода энергии. Несколько примеров анаболического пути включают:
Синтез белков из аминокислот
цепей ДНК из строительных блоков нуклеиновых кислот, называемых нуклеотидами.
Превращение аминокислот в кератин
Превращение гликогена или крахмала из молекулы глюкозы
Энергия, используемая в этих важных процессах биосинтеза, находится в форме АТФ и других молекул краткосрочного хранения энергии.
Катаболический путь
Он включает в себя расщепление сложных молекул на более простые и представляет собой путь, высвобождающий энергию в форме АТФ. Примеры метаболизма катаболического пути включают:
Распад глюкозы на пируват
Превращение белков в аминокислоты
Превращение липидов или жиров в жирные кислоты
Все эти разложения происходят в путь и энергия (АТФ) высвобождаются для обеспечения работы клеток.
Когда мы едим, молекулы превращаются в аминокислоты, сахара и жирные кислоты с высвобождением АТФ (энергии) и таких отходов, как азот, CO2, фосфор и сера, которые выводятся из организма. Здесь мы можем видеть, что все, что мы помещаем в свое тело в виде пищи, превращается в другую форму.
Результаты катаболического пути абсорбируются нашим кишечником, попадают в кровоток и затем распределяются по клеткам организма. Его функции включают в себя:
Энергетическая клетка формирует запасные части для своих собственных молекул, которые изношены, а в других случаях она выстраивает молекулы энергии в теле.
Они действуют как топливо для будущей деятельности клетки. Пример. Мышечные клетки используют молекулы энергии для сокращения, а клетки мозга используют их для формирования мыслительных процессов.
Что такое метаболизм в биологии и метаболическая активность?
С биологической точки зрения метаболизм — это цепь биохимических реакций, включающая все биохимические реакции, которые помогают выживанию организмов. Метаболизм также необходим организмам для роста, воспроизводства и поддержания различных жизненных условий.Даже когда организмы реагируют на окружающую среду, когда они получают стимул, это также происходит из-за метаболизма. Таким образом, метаболизм играет важную роль в общем росте организмов, и от переваривания до выделения, все процессы в живых организмах происходят в результате биохимической реакции, известной как метаболизм. В организмах есть два типа метаболических процессов: —
Катаболизм: когда мы что-то едим или пьем, молекулы, которые попадают внутрь нашего тела, становятся больше по размеру, и их необходимо расщепить на более мелкие молекулы, чтобы дать нам энергию. .Следовательно, в процессе катаболизма более крупные молекулы внутри организма превращаются в более мелкие и выделяют энергию.
Анаболизм: этот процесс помогает создавать и синтезировать сложные вещества из более простых веществ, необходимых для различных клеток, а также накапливает энергию.
Как работает метаболизм?
Когда мы едим, наша пищеварительная система начинает работать. Пищеварительная система использует различные типы ферментов для выполнения следующих процессов:
Первоначальный процесс — это расщепление белков на аминокислоты.
Ферменты также превращают жиры в жирные кислоты.
Углеводы были преобразованы в сахара (глюкозу) с помощью ферментов.
Когда ферменты превращают белки, жиры и углеводы в аминокислоты, жирные кислоты и сахара, организм может использовать их в любое время в качестве источника энергии, и эти соединения поглощаются кровью, что помогает переносить их в разные клетки. для их функционирования.
После того, как кровь переносит их к клеткам, и они попадают в клетки, другие ферменты начинают свою работу и действуют на них, а также происходит множество химических реакций, которые включают «метаболизм» различных соединений.Энергия, вырабатываемая этими соединениями, может использоваться для разных целей, в том числе использоваться организмом для выполнения различных задач, а также может сохраняться внутри тела в тканях, таких как мышцы, печень и жировые отложения.
Ферменты
Белок, действующий как катализатор, облегчающий реакции в биологических системах, называется ферментами. Каждая стадия реакции метаболических путей катализируется ферментом и контролирует биохимические реакции.
Аденозинтрифосфат (АТФ)
Это органическое соединение, которое обеспечивает энергией многие процессы в живых клетках, например сокращение мышц, химический синтез и распространение нервных импульсов.Он встречается во всех формах жизни и часто упоминается как «молекулярная денежная единица». При потреблении в метаболических процессах он превращается либо в аденозиндифосфат (АДФ), либо в аденозинмонофосфат (АМФ).
[Изображение будет скоро загружено]
Питание, обмен веществ и энергия
Питание — важнейший ключ к метаболизму. В процессе метаболизма различная пища, потребляемая живыми организмами, расщепляется на более простые вещества для высвобождения энергии, которая помогает выполнять различные процессы внутри тела, такие как пищеварение, транспортировка пищи в разные клетки и т. Д., а также помогает организму выполнять другие действия для поддержания жизнедеятельности. Кроме того, энергия, производимая в организме в процессе метаболизма, также используется для производства новых белков и нуклеиновых кислот, которые также известны как ДНК и РНК.
Основное питание, необходимое человеческому организму, включает водород, кислород, углерод, фосфор, азот, серу и еще 20 элементов. Кроме того, углеводы, белки и липиды являются основными элементами, поступающими в наш организм, а дополнительными элементами являются вода, витамины и минералы.
Углеводный обмен, метаболизм липидов и метаболизм белков.
Расщепление углеводов посредством различных биохимических процессов известно как углеводный обмен. Один из важнейших путей метаболического процесса — это углеводы. В процессе фотосинтеза в растениях происходит синтез углеводов из углекислого газа и воды. Процесс клеточного дыхания используется животными и грибами, потребляющими растения, для расщепления углеводов с целью производства энергии, которая используется разными клетками для выполнения различных действий.
Метаболизм липидов — это расщепление и хранение жира в живых организмах. Процесс синтеза липидов известен как липогенез. Холестерин и триглицериды являются основными жирами в организме человека, а жирные кислоты и мембранные липиды являются дополнительными липидами, обнаруженными в организме человека.
Метаболизм белка — это цепь различных биохимических процессов, которые необходимы для синтеза или расщепления белка и преобразования его в аминокислоты. Транскрипция, трансляция и посттрансляционные модификации — это этапы синтеза белка.Расщепление белка с помощью различных ферментов известно как полипептид.
Как увеличить метаболизм?
Есть несколько предложений по увеличению метаболизма:
Мы все знаем, что для того, чтобы вести здоровый и подтянутый образ жизни, нам нужно избегать всех тех веществ, которые потребляют больше калорий и становятся причиной лишних килограммов в нашем организме. тело. Хотя, если вы уменьшите количество еды, это также негативно скажется на вашем метаболизме.Поэтому важно придерживаться правильно сбалансированной диеты, которая обеспечивает адекватное качество питания вашего тела и ускоряет метаболизм.
Завтрак играет важную роль в поддержании обмена веществ. Нам нужна энергия в течение дня для выполнения различных повседневных дел. Поэтому вам не следует пропускать утренний завтрак, потому что он придает энергию и активность вашему телу в течение дня, что помогает вам стать продуктивным и активным, а вы всегда оставаться в форме и здоровым.