Упражнения на мышцы груди
Мышцы груди
Мышцы груди делятся на три части: вехняя, средняя и нижняя части. Upper, Middle, Lower. В нашем случае мы будем рассматривать верхнюю и среднюю части груди, поскольку волокна нижней части самые маленькие по размеру и выделить их обособленно нет возможности. Грудные мышцы всегда работают только в Поперечной (горизонтальной) плоскости выполнения упражнений.
Мышцы вехней части груди работают в межплоскостном пространстве: между сгибанием плеча, которое происходит в Сагиттальной плоскости и горизонтальным приведением плеча в горизонтальной плоскости. Упражнения выполняются на скамье под углом 40-45 градусов. Мышцы нижней части груди работают в плоскостях горизонтального приведения и коронального или фронтального приведения. Выполняют упражнения на скамье под отрицательным углом 30 градусов.
В каждом упражнении на мышцы груди участвует лопаточный сустав, в котором происходит отведение лопатки с небольшим поворотом наружу.
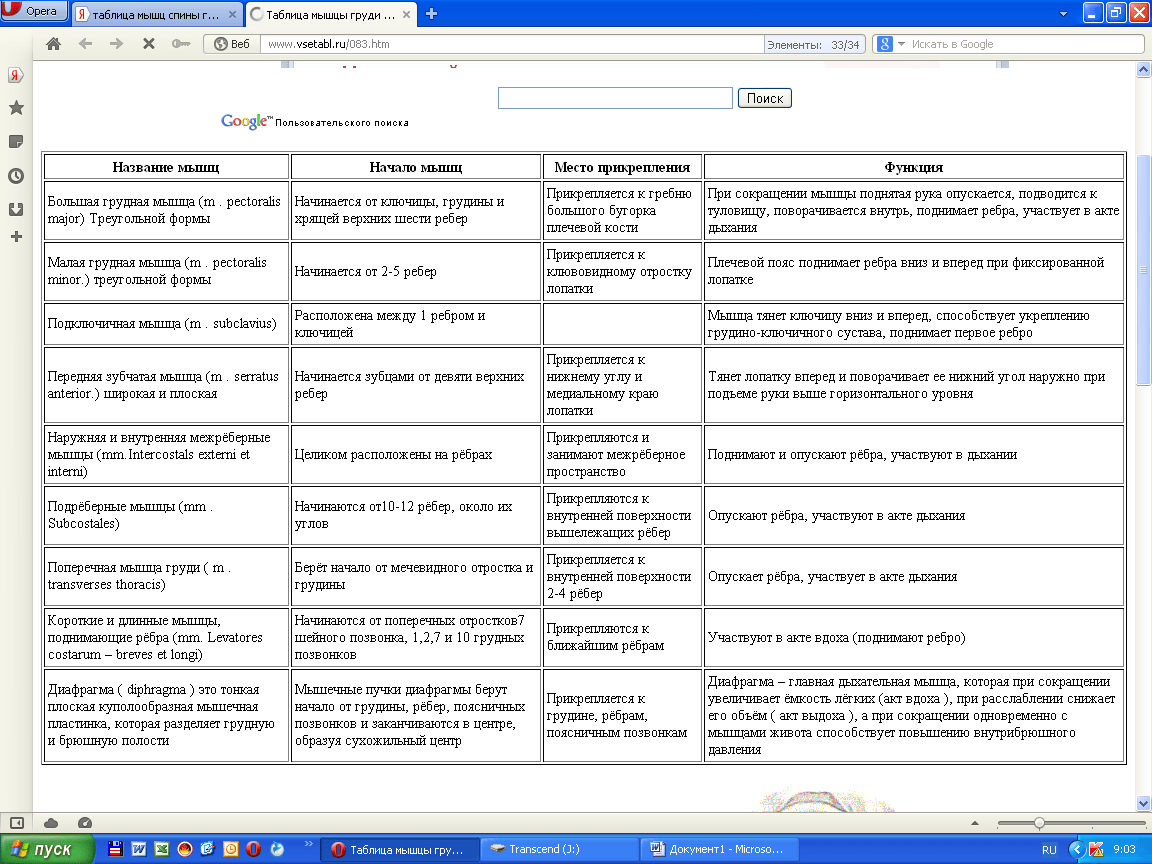
В каждом упражнении на мышцы груди антагонистом всегда является Posterior Deltoid (задний пучок плечевой мышцы). Упражнения на мышцы груди всегда многосуставные. В локтевом суставе всегда работает Triceps в рычаге 1. В упражнениях на верхнюю и среднюю часть груди синергисты одни и те же. В упражнениях на нижнюю часть груди подключается синергист Latissimus Dorsi (широчайшая мышца спины). Поэтому силовые показатели упражнений под отрицательным углом выше показателей на ровной скамье. Выполняя упражнение узким хватом, движение идет в плоскостях сгибания плеча и разгибания локтя. Агонистом здесь является
 Различия при выполнении упражнений на мышцы груди с помощью штанги, гантелей и машины Смитта различаются только по работе синергистов в момент движения.
Различия при выполнении упражнений на мышцы груди с помощью штанги, гантелей и машины Смитта различаются только по работе синергистов в момент движения.
1.Flat Bench Press. В положении лежа, жим штанги от груди.
2.Bench Press + Smith Machine. В положении лежа, жим штанги на машине Смитта.
3.Dumbbell Bench Press. В положении лежа, жим гантелей от груди.
4.Жим от груди на специальном тренажере (жим вертикальный, жим под углом + 45 градусов, жим под углом -30 градусов).
Упражнение главным образом воздействует и укрепляет мышцы груди Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо | Горизонтальное приведение | Концентрическое на подъеме | Pectoralis Major Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 1 |
Posterior Deltoid Teres Minor |
| Локоть | Разгибание | Концентрическое | Triceps Рычаг 1 |
————— |
Biceps,Brahialis, Brahioradialis |
| Лопатка | Отведение с поворотом наружу | Концентрическое | Seratus Anterior | ————— | Rhomboids Middle Trapezius |
Положение для выполнения: лежа на скамье, расстояние захвата грифа позволяет держать руки под углом 90 градусов в средней позиции. При работе на машине Смитта необходимо правильно и удобно расположить скамью. Упражнение на машине Смитта менее травмоопасно и подходит начинающим. Движение грифа строго вверх, что обеспечивает мышцам получение реально подобранной нагрузки. Наиболее безопасны упражнения на тренажере с горизонтальным жимом, подходит наименее опытным спортсменам. При изменении захвата на узкий, или при развороте кистей в “Mid Position”, цель упражнения становится Triceps, работа груди облегчается.
При работе на машине Смитта необходимо правильно и удобно расположить скамью. Упражнение на машине Смитта менее травмоопасно и подходит начинающим. Движение грифа строго вверх, что обеспечивает мышцам получение реально подобранной нагрузки. Наиболее безопасны упражнения на тренажере с горизонтальным жимом, подходит наименее опытным спортсменам. При изменении захвата на узкий, или при развороте кистей в “Mid Position”, цель упражнения становится Triceps, работа груди облегчается.
5.Incline Bench Press. В положении лежа под углом 40-45 градусов, жим штанги от груди.
6.Incline Bench Press + Smith Machine. В положении лежа под углом 40-45 градусов, жим на машине Смитта.
7.Incline Dumbbell Bench Press. В положении лежа под углом 40-45 градусов, жим гантелей от груди.
Упражнение главным образом воздействует и укрепляет мышцы груди Upper Pectoralis Major. Мышцы работают в различных суставах следующим образом:
Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Сгибание плеча и горизонтальное приведение |
Концентрическое на подъеме |
Upper Pectoralis Major Рычаг 1 |
Anterior Deltoid Coracobrachialis Рычаг 1 |
Posterior Deltoid |
| Локоть |
Разгибание |
Концентрическое | Triceps Рычаг 1 |
—————- |
Biceps,Brahialis, Brahioradialis |
| Лопатка |
Отведение и разворот наружу |
Концентрическое | Seratus Anterior | Upper + Lower Trapezius | Rhomboids & Middle Trapezius |
Положение для выполнения: положение лежа, выполняется на специальной скамье под углом 40-45 градусов.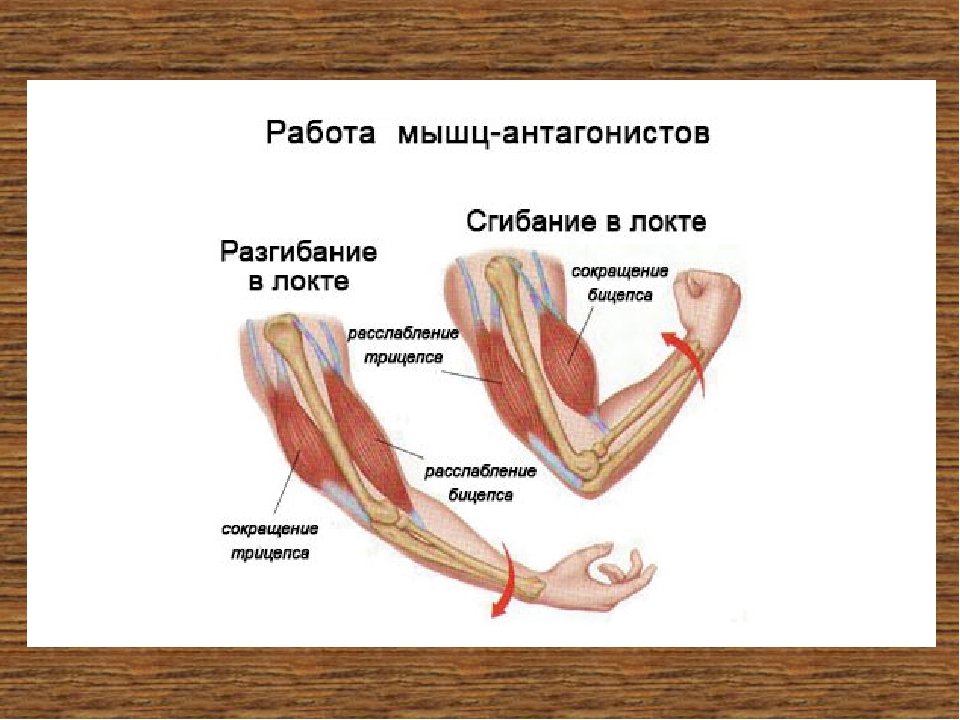 Со свободной штангой больше работа синергистов, с гантелями синергисты работают в максимальной степени, техника спортсмена должна быть достаточно высокой. Опускание грифа при выпонении упажнения производится почти до уровня ключицы.
Со свободной штангой больше работа синергистов, с гантелями синергисты работают в максимальной степени, техника спортсмена должна быть достаточно высокой. Опускание грифа при выпонении упажнения производится почти до уровня ключицы.
8.Decline Bench Press. В положении лежа под отрицательнаым углом 30 градусов, жим штанги от груди
9.Decline Bench Press + Smith Machine. В положении лежа под отрицательнаым углом 30 градусов, жим штанги на машине Смитта.
10.Decline Dumbbell Bench Press. В положении лежа под отрицательнаым углом 30 градусов, жим гантелей от груди.
Упражнение главным образом воздействует и укрепляет мышцы груди Lower Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Приведение плеча и горизонтальное приведение |
Концентрическое на подъеме |
Lower Pectoralis Major Рычаг 3 |
Latissimus Dorsi Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Локоть |
Разгибание |
Концентрическое | Triceps Рычаг 1 |
—————— |
Biceps,Brahialis, Brahioradialis |
| Лопатка |
Отведение и разворот наружу |
Концентрическое | Seratus Anterior | Upper + Lower Trapezius | Rhomboids & Middle Trapezius |
Замечания: рабочий вес при выполнении данной категории упражнений будет больше за счет вступления в работу синергиста
 Опускание грифа производится до ближайшей к центру точки груди.
Опускание грифа производится до ближайшей к центру точки груди.
11.Flat Dumbbell Flys. «Бабочка» лежа: горизонтальное приведение плеча с гантелями.
Упражнение главным образом воздействует и укрепляет мышцы груди Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Горизонтальное приведение |
Концентрическое на подъеме |
Pectoralis Major Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Локоть | Нет движения | Изометрическое |
Biceps,Brahialis, Brahioradialis |
—————- | ————- |
| Лопатка |
Отведение и поворот наружу |
Концентрическое | Seratus Anterior | Upper + Lower Trapezius |
Rhomboids & Middle Trapezius |
12. «Бабочка» сидя, горизонтальное приведение плеча на тренажере.
«Бабочка» сидя, горизонтальное приведение плеча на тренажере.
Упражнение главным образом воздействует и укрепляет мышцы груди Pectoralis Major
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Горизонтальное приведение |
Концентрическое на подъеме |
Pectoralis Major Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Лопатка |
Отведение и поворот наружу |
концентрическое | Seratus Anterior | Upper + Lower Trapezius | Rhomboids & Middle Trapezius |
Спинку тренажера надо установить таким образом, чтобы образовать угол 90 градусов между плечом и предплечием. Движение равномерно делается всей рукой, а не только локтями или кистями рук.
Движение равномерно делается всей рукой, а не только локтями или кистями рук.
13.. «Бабочка» лежа на кроссовере. Горизонтальное приведение плеча на кабеле кросса.
Упражнение главным образом воздействует и укрепляет мышцы груди Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Горизонтальное приведение |
Концентрическое на подъеме |
Pectoralis Major Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Локоть | Нет движения | Изометрическое |
Biceps,Brahialis, Brahioradialis |
————— | —————- |
| Лопатка |
Отведение и поворот наружу |
Концентрическое | Seratus Anterior | Upper + Lower Trapezius | Rhomboids & Middle Trapezius |
Положение для выполнения: С нижнего блока лечь на скамью и опустить руки так же, как и при выполнении упражнения с гантелями.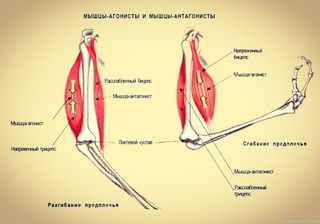 Нагрузка идет от нижних блоков кросса, хорошо растягивает грудную мышцу, но менее трвавмоопасно, чем свободные веса. Следить, чтобы руки не были сильно согнуты. Максимальная нагрузка в момент угла 90 градусов между кабелем и конечностью.
Нагрузка идет от нижних блоков кросса, хорошо растягивает грудную мышцу, но менее трвавмоопасно, чем свободные веса. Следить, чтобы руки не были сильно согнуты. Максимальная нагрузка в момент угла 90 градусов между кабелем и конечностью.
14. «Бабочка» сидя на скамье 40-45 градусов. Горизонтальное приведение плеча с гантелями.
Угол наклона скамьи 40-45 градусов. Требует значительного умения и опыта спортсмена. Движение в локтевом суставе отсутствует.
15. Cable Crossovers «Бабочка» сидя на скамье 40-45 градусов. Горизонтальное приведение плеча на кроссовере.
Упражнения 14 и 15 главным образом воздействует, и укрепляют мышцы груди Upper Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Приведение плеча и горизонтальное приведение |
Концентрическое на подъеме |
Upper Pectoralis Major Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Локоть | Нет движения | Изометрическое |
Biceps,Brahialis, Brahioradialis |
———— | ————- |
| Лопатка |
Отведение и поворот наружу |
Концентрическое | Seratus Anterior | Upper + Lower Trapezius |
|
16. Cable Crossovers «Бабочка». В положении стоя приведение плеча с верхнего блока кроссовера.
Cable Crossovers «Бабочка». В положении стоя приведение плеча с верхнего блока кроссовера.
Упражнение главным образом воздействует и укрепляет мышцы груди Lower Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Приведение плеча и горизонтальное приведение |
Концентрическое на подъеме |
Lower Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Локоть | Нет движения | Изометрическое |
Biceps,Brahialis, Brahioradialis |
————— | —————- |
| Лопатка |
Отведение и поворот наружу |
Концентрическое | Seratus Anterior | Upper + Lower Trapezius |
|
17.
 Parallel Bar Dips. Отжимания на брусьях широким хватом.
Parallel Bar Dips. Отжимания на брусьях широким хватом.Упражнение главным образом воздействует и укрепляет мышцы груди Lower Pectoralis Major. Мышцы работают в различных суставах следующим образом:
| Сустав | Движение | Сокращение | Агонист | Синергист | Антагонист |
| Плечо |
Сгибание плеча и горизонтальное приведение |
Концентрическое на подъеме |
Lower Рычаг 3 |
Anterior Deltoid Coracobrachialis Рычаг 3 |
Posterior Deltoid |
| Локоть | Разгибание |
Концентрическое | Triceps | ————— |
Biceps,Brahialis, Brahioradialis |
Положение для выполнения: если в упоре на брусьях локти прижаты к туловищу, основную нагрузку принимает Triceps, в плечевом суставе работает вся грудная мышца.
 Если брусья широкие, и упор тоже широкий, агонистом является Lower Pectoralis Major.
Если брусья широкие, и упор тоже широкий, агонистом является Lower Pectoralis Major.Внимание! У Вас нет прав для просмотра скрытого текста.
Ключевые теги: мышцы груди, машина Смитта, плечо, мышцы
Что такое нижняя прямая мышца?
Есть шесть экстраокулярных мышц, которые вместе перемещают зрачок вверх, вниз, из стороны в сторону и в комбинации этих взглядов. Как видно из названия, нижняя прямая мышца прикрепляется к склере — белому «шарику» глазного яблока — в нижней части структуры и в основном работает для перемещения взгляда вниз в процессе, известном как депрессия. Другими экстраокулярными мышцами являются верхняя прямая, медиальная прямая, латеральная прямая, верхняя косая и нижняя косая. Все эти мышцы, кроме одной, образуют коническую форму на каждой глазной орбите с устьем конуса, где мышцы соединяются с глазным яблоком, и острой точкой конуса, где мышцы сходятся вместе в кольцевидном сухожилии. Зрительный нерв, черепной нерв II, проходит через это кольцевидное сухожилие через конус к зрачку.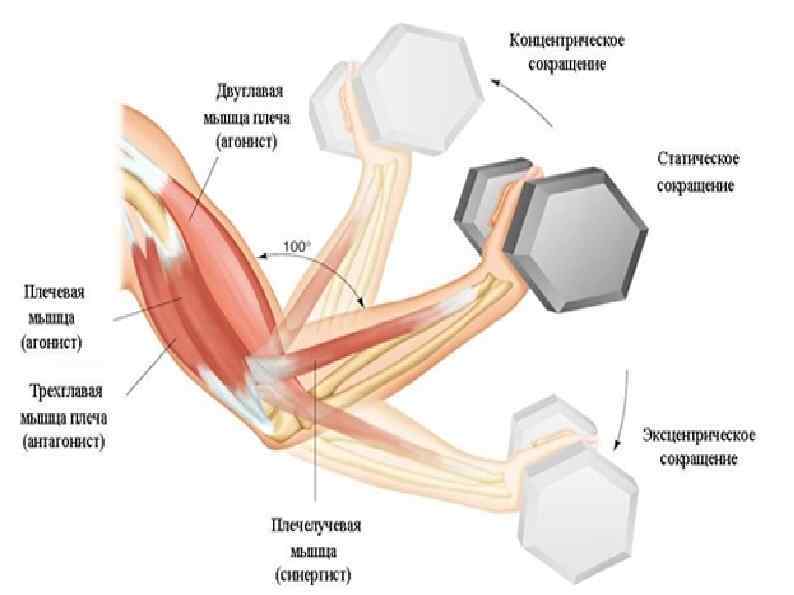
Каждая из четырех нижних и верхних экстраокулярных мышц имеет три функции различной ответственности. Основная функция нижней прямой мышцы — нисходящее движение или депрессия зрачка. Вторичной функцией нижней прямой мышцы является вращение верхушки глазного яблока вдали от носа в процессе, известном как выгибание. Наконец, третичная функция этой мышцы состоит в том, чтобы перемещать зрачок внутрь или медиально при отведении.
Некоторые движения глаз являются результатом только одного сокращения глазной мышцы, в то время как другие требуют, чтобы две мышцы работали вместе. Мышца, в первую очередь ответственная за движение зрачка, называется агонистом. Другие экстраокулярные мышцы, которые помогают в движении, известны как синергисты. Те экстраокулярные мышцы, которые двигают зрачок в противоположном направлении, являются антагонистами. Таким образом, нижняя прямая мышца является агонистом движения зрачка вниз; синергист для экструзии или аддукции; и антагонист движения верхней прямой мышцы глаза вверх во время подъема.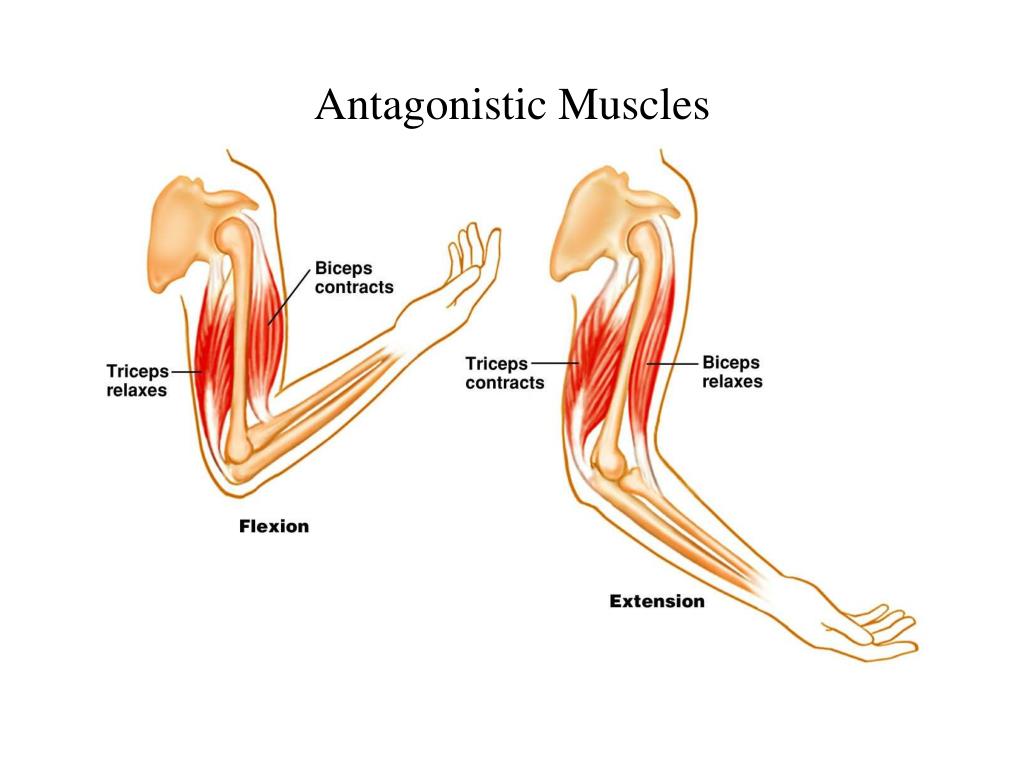
Эти описания относятся только к движению одного глаза, которое называется протоком. Для взгляда одного ученика вниз, это нисходящее движение депрессии также известно как инфрадукция. Однако, когда два глаза работают вместе, движение глаз называется версией. Чтобы смотреть вниз обоими глазами, выполняя либо инфраверсию, либо деорусверсию, обе правые и левые нижние прямые мышцы — или прямые — должны сжиматься, чтобы форсировать движение. Кроме того, правая и левая верхняя прямая косая прямая синергист действуют на нижнюю прямую мышцу, двигая каждый глаз в одном направлении.
ДРУГИЕ ЯЗЫКИ
muscle- – phrases – Multitran dictionary
| English | Russian |
| abdominal muscle deficiency syndrome | синдром «подрезанного живота» (врождённое частичное или полное отсутствие мышц брюшной стенки в сочетании с аномалиями других органов) |
| abdominal muscles | брюшной пресс |
| adductor muscle | аддуктор (у моллюсков) |
| adductor muscle | замыкающий мускул (у моллюсков) |
| adductor muscle | приводящая мышца |
| adductor muscle | мускула-замыкатель |
| adductor muscle | аддуктор |
| anterior tibial muscle | передняя мышца голени |
| Ca 2+ transport by the sarcoplasmic reticulum Ca 2+ -ATPase in sea cucumber Ludwigothurea grisea muscle | транспорт Ca 2+ посредством Ca 2+ -АТФазы саркоплазматического ретикулума мускулатуры голотурии (Ludwigothurea grisea) |
| centrally acting muscle relaxant | миорелаксант центрального действия |
| chicken muscles | мышцы цыплят |
| ciliary muscle | цилиарная мышца (глаза) |
| closing muscle | замыкающий мускул (у моллюсков) |
| closing muscle | аддуктор (у моллюсков) |
| congenerous muscles | группа мышц, производящих одинаковое действие |
| constrict muscle | сокращать мышцу |
| contract a muscle | сокращать мускул |
| dark-muscle fish | рыба с тёмным мясом |
| degeneration of muscles | мышечная дегенерация |
| depolarizing muscle relaxant | деполяризующий миорелаксант |
| depolarizing muscle relaxant | курареподобное деполяризующее средство |
| depressor muscle of angle of mouth | мышца, опускающая угол рта |
| develop one’s muscles | развивать мускулы |
| develop one’s muscles | развивать мускулатуру |
| develop muscles | развить мускулатуру |
| do exercises to develop muscles | делать упражнения для развития мышц |
| do exercises to muscles | делать упражнения для развития мышц |
| elevator muscle of angle of mouth | мышца, поднимающая угол рта |
| elevator muscle of upper lip and wing of nose | мышца, поднимающая верхнюю губу и крыло носа |
| extraocular muscles | мышцы глазного яблока (только глазодвигательные мышцы, они же в просторечии наружные мышцы глаза doc090) |
| fibrillar flight muscle | фибриллярная летательная мышца |
| firm muscles | упругие мускулы |
| fixation muscle | мускул-фиксатор |
| flex a muscle | напрягать мускул |
| flex one’s muscle | играть мускулами |
| flex one’s muscles | напрячь мускулы |
| flex one’s muscles | напрягать мускулы |
| flex one’s muscles | демонстрировать силу |
| flight muscle | летательная мышца |
| he tensed his muscles | он напряг мускулы |
| heat application to relieve muscle spasm | применение тепла для устранения мышечного спазма |
| his muscles are like jelly | у него не мускулы, а кисель |
| his muscles are soft | у него слабые мышцы |
| his muscles relaxed | его мышцы расслабились |
| hold one’s stomach muscles in | втянуть мышцы живота |
| insufficiency of ocular muscles | недостаточность динамического равновесия глазных мышц |
| insufficiency of ocular muscles | недостаточность глазных мышц |
| intercostal muscles | межрёберные мышцы |
| intrinsic muscle | мышца, у которой обе точки прикрепления расположены на конечности |
| Ivanov’s muscle | радиальный пучок |
| Ivanov’s muscle | мышца Иванова |
| long abductor muscle of thumb | длинная мышца, отводящая больший палец кисти |
| longitudinal muscle | продольная мышца |
| mastication muscle | жевательная мышца |
| move a muscle | двигать мускулом |
| muscle activity | мышечная активность |
| muscle contraction | мышечное сокращение |
| muscle contracts when it is irritated by electricity | под воздействием электричества мышца сокращается |
| muscle electrostimulator | электростимулятор мышц |
| muscle of mastication | жевательный мускул |
| muscle of tragus | козелковая мышца колена |
| muscle pill | анаболик (увеличивающий мышечную массу у спортсменов) |
| muscle sugar | инозит |
| muscle track | Brach. мускульный след мускульный след |
| muscles | мышцы |
| muscles of auditory ossicles | мышцы слуховых косточек мышцы |
| muscles of iron | стальные мускулы |
| neurobiology of Duchenne muscular dystrophy: learning lessons from muscle? | нейробиологические аспекты миодистрофии Дюшенна: получают хороший урок от мускул? |
| non-depolarizing muscle relaxant | курареподобное недеполяризующее средство |
| orbicular muscle | круговая мышца |
| packing with muscle | тампонада мышцей |
| palpebral muscle | мышца века |
| papillary muscle | сосочковая мышца |
| papillary muscle | папиллярная мышца |
| phasic muscles | быстрые мышцы |
| plantar muscle reflex | тыльно-стопный рефлекс |
| postural muscles | тонические мышцы |
| postural muscles | медленные мышцы |
| presence of a fatty acid-binding protein and lipid stores in flight muscles of Dipetalogaster maximus | Hemiptera: Reduviidae Наличие белков, связывающихся с жирными кислотами, и депо липидов в летательных мышцах Dipetalogaster maximus |
| pull a muscle | натягивать мускул |
| pull a muscle | растянуть себе мускул |
| relax a muscle | расслаблять мускул |
| relax muscles | расслаблять мышцы |
| sardine ordinary and dark muscles | светлое и тёмное мясо сардины |
| sever a muscle | пересечь мышцу |
| skew muscle | косая мышца |
| somatic muscle | скелетная мышца |
| sphincter muscle | сфинктер |
| split the muscles fibers bluntly in a longitudinal direction | тупо разводить мышцы по ходу волокон |
| strain a muscle | напрягать мускул |
| such muscles are termed rotators | такие мышцы называются мускулами-вращателями |
| synergistic muscle | мышца-синергист |
| tense a muscle | напрягать мускул |
| the degree of stretch in the muscle fibres | степень растяжения мышечной ткани |
| the muscle contracts | мышца сокращается |
| the neurobiology of Duchenne muscular dystrophy: learning lessons from muscle? | нейробиологические аспекты миодистрофии Дюшенна: получают хороший урок от мускул? |
| thyrotropin-releasing hormone activates K Ca currents in stomach smooth muscle cells | активация тиролиберином K Ca + -токов в клетках стенки желудка |
| to muscle in | вторгаться силой |
| to muscle in | вторгаться |
| to muscle in | врываться силой |
to muscle in on smth. | примазаться к (чему-л.) |
| to muscle in on smth. | ввязаться во (что-л.) |
| try not to sit with your shoulders hunched up like that, it’s bad for your neck muscles | не сиди сутулясь, это плохо сказывается на мышцах шеи |
| twist of muscle | растяжение мышцы |
| unbrace the muscles | расслабить мышцы |
| uterine muscle | мышца матки |
| uterine muscle | миометрий |
| voluntary muscle | поперечно-полосатая мышца |
| voluntary muscle contraction | произвольное мышечное сокращение |
| weak muscles | слабые мускулы |
| whipcord muscles | крепкие мускулы |
| wing muscle | крыловидная мышца |
| wrench a muscle | растягивать мускул |
Проксимальные и дистальные спинальные нейроны, иннервирующие несколько моторных пулов синергистов и антагонистов
1) Вы должны смягчить часть идентификации происхождения V0v или лучше сделать соответствующую характеристику, как предложено ниже;
Мы тщательно отредактировали разделы, связанные с идентификацией интернейронов V0v. Мы согласны с рецензентами, что у нас нет достаточных доказательств, чтобы точно определить генетическое происхождение дальних нисходящих интернейронов, и мы значительно смягчили это.Как было предложено рецензентом №1, мы провели дальнейшие эксперименты с использованием антитела Evx1 для выборочной метки интернейронов V0v. Однако, хотя хорошее мечение было получено в неонатальной (P0) ткани, никакого мечения не было обнаружено в ткани того возраста, который мы использовали в настоящем исследовании (P9-P10), в соответствии с подавлением факторов транскрипции во время развития — особенность, которая также уменьшала наша чувствительность в анализе Lhx1, хотя и в меньшей степени. Чтобы усугубить нашу неопределенность, недавнее исследование было опубликовано, когда мы пересматривали нашу рукопись, определяя класс интернейронов, производных dI2, с морфологическими особенностями, подобными LDPN, описанным в нашем исследовании.Заметим, что входы dI2 также выражают Lhx1. Учитывая эту дополнительную неопределенность и сниженный уровень обнаружения Lhx1 в более зрелой ткани, мы смягчили любой вывод, который может указывать на то, что мы идентифицировали кардинальный класс дивергентных LDPN.
Мы согласны с рецензентами, что у нас нет достаточных доказательств, чтобы точно определить генетическое происхождение дальних нисходящих интернейронов, и мы значительно смягчили это.Как было предложено рецензентом №1, мы провели дальнейшие эксперименты с использованием антитела Evx1 для выборочной метки интернейронов V0v. Однако, хотя хорошее мечение было получено в неонатальной (P0) ткани, никакого мечения не было обнаружено в ткани того возраста, который мы использовали в настоящем исследовании (P9-P10), в соответствии с подавлением факторов транскрипции во время развития — особенность, которая также уменьшала наша чувствительность в анализе Lhx1, хотя и в меньшей степени. Чтобы усугубить нашу неопределенность, недавнее исследование было опубликовано, когда мы пересматривали нашу рукопись, определяя класс интернейронов, производных dI2, с морфологическими особенностями, подобными LDPN, описанным в нашем исследовании.Заметим, что входы dI2 также выражают Lhx1. Учитывая эту дополнительную неопределенность и сниженный уровень обнаружения Lhx1 в более зрелой ткани, мы смягчили любой вывод, который может указывать на то, что мы идентифицировали кардинальный класс дивергентных LDPN. Но мы предпочитаем хранить данные в рукописи, поскольку мы приводим доказательства того, что по крайней мере некоторые LDPN экспрессируют Lhx1 (и, следовательно, вероятно, V0v или dI2, как обсуждалось). Конкретные изменения в тексте подробно описаны в ответах рецензентам №1 и №3.
Но мы предпочитаем хранить данные в рукописи, поскольку мы приводим доказательства того, что по крайней мере некоторые LDPN экспрессируют Lhx1 (и, следовательно, вероятно, V0v или dI2, как обсуждалось). Конкретные изменения в тексте подробно описаны в ответах рецензентам №1 и №3.
2) Вам следует дополнительно обсудить характеристики нейротрансмиттера и, в конечном итоге, решить этот вопрос экспериментально, если это возможно;
Идентификация фенотипа передатчика всех дивергентных интернейронов потребовала бы другого протокола, например.г. с генетически (non-cre) мышью. В то время как возбуждающая природа дивергентных LDPN достаточно установлена, расходящиеся премоторные IN в поясничной и грудной областях определенно являются тормозящими и возбуждающими. Теперь мы продемонстрируем этот момент дальше, добавив дополнительные рисунки, показывающие терминалы с двойной меткой, которые являются положительными или отрицательными для VGlut2 и локализованы по всему грудному и поясничному отделам спинного мозга (Рисунок 1 — рисунок в приложении 6). Необходимы дальнейшие исследования, чтобы понять, разделены ли расходящиеся возбуждающие и тормозные премоторные интернейроны и принадлежат ли они к однородным кардинальным классам или нет.
Необходимы дальнейшие исследования, чтобы понять, разделены ли расходящиеся возбуждающие и тормозные премоторные интернейроны и принадлежат ли они к однородным кардинальным классам или нет.
3) Вам следует дополнительно описать ростро-каудальное расширение помеченных пре-мотонейронов.
Мы добавили новые рисунки, где распределение премоторных интернейронов показано вдоль рострокаудальной оси для поясничного, грудного и крестцового отдела спинного мозга (см. Ответ рецензенту № 3 и рисунок 1 — дополнительный рисунок 4, рисунок 2 — дополнительный рисунок 4 и рисунок 3- дополнительный рисунок 4).
4) Кроме того, явно отсутствует функциональный аспект, чем больше вы сможете решить этот вопрос в своей пересмотренной версии, тем больше он укрепит документ, но мы понимаем, что полное исследование этого потребует слишком много времени и представляет собой значительный объем работы. если данные еще не доступны.
Вопрос о функциях этих LDPN, несомненно, является одним из наших главных приоритетов. Наша рукопись сосредоточена на открытии этих своеобразных клеток, но определение их функции является необходимым следующим шагом. Обнаружение точного анатомического местоположения LDPN и доступность методов отслеживания теперь позволит нам получить доступ к этим клеткам и управлять ими, но это явно отдельный проект, и, по нашему мнению, его нельзя сводить к придатку к и без того плотной рукописи.
Наша рукопись сосредоточена на открытии этих своеобразных клеток, но определение их функции является необходимым следующим шагом. Обнаружение точного анатомического местоположения LDPN и доступность методов отслеживания теперь позволит нам получить доступ к этим клеткам и управлять ими, но это явно отдельный проект, и, по нашему мнению, его нельзя сводить к придатку к и без того плотной рукописи.
Рецензент №1 (Рекомендации авторам):
1. Во всех случаях следует давать более подробную информацию о рострально-каудальном расположении спинного мозга. Сначала я думаю, что следует включить рисунок, описывающий расположение моторных пулов для 4 инъецированных мышц — это может быть в дополнительных данных или это может быть панель на рисунке 1. Это необходимо для того, чтобы у нас было четкое представление о том, как далеки от аксонов проекта PMI. По моему опыту, моторный бассейн может охватывать несколько поясничных сегментов.Также должна быть предоставлена более конкретная информация относительно расположения разделов, содержащих описываемые PMI.
Я понимаю, что после разрезания ткани это не точная наука, но, учитывая вашу осторожность при подсчете каждой третьей части, вы должны иметь довольно хорошее представление о том, в каком сегменте находятся нейроны. Это предпочтительнее нынешнего описания «рострально-шейный» или его эквивалента.
 Я понимаю, что после разрезания ткани это не точная наука, но, учитывая вашу осторожность при подсчете каждой третьей части, вы должны иметь довольно хорошее представление о том, в каком сегменте находятся нейроны. Это предпочтительнее нынешнего описания «рострально-шейный» или его эквивалента.
Я понимаю, что после разрезания ткани это не точная наука, но, учитывая вашу осторожность при подсчете каждой третьей части, вы должны иметь довольно хорошее представление о том, в каком сегменте находятся нейроны. Это предпочтительнее нынешнего описания «рострально-шейный» или его эквивалента.Мы добавили в качестве дополнительных рисунков ростро-каудальное распределение дивергентных премоторных ИН поясничных сегментов и рострально-каудальное распределение всех премоторных ИН грудного и шейного отделов (Рисунок 1 — добавление к рисунку 4, Рисунок 2 — дополнение к рисунку 4 и рисунок 3 — приложение к рисунку 4).Мы также предоставили количество инфицированных мотонейронов от каждой пары инъецированных мышц, число, которое, как мы отмечаем, занижено из-за токсичности вируса для стартовых клеток (дополнительный файл 1).
На рисунках количество MNs, инфицированных любым из вирусов, обычно довольно низкое, поэтому я полагаю, что секции не очень близки к середине инфицированных моторных пулов?
На Рисунке 1 представлен разрез близок к середине инфицированного моторного пула. В самом деле, 2 MN, инфицированные на одном участке, как в примере на Рисунке 1, представляют собой типичное количество MN в одном участке помеченного моторного столбца. Частично это связано с редкостью или заражением бешенством, но, что более важно, с тем фактом, что каждый срез имеет толщину всего 30 мкм. Мы также добавили дополнительные рисунки (рисунки 1,2,3 к рисункам 1,2,3), показывающие больше примеров поясничных отделов после инъекций в различные моторные бассейны. Среди показанных примеров на одном участке инфицировано до 8 MN, но это редкое явление (рисунок 1 — приложение к рисунку 1A).
В самом деле, 2 MN, инфицированные на одном участке, как в примере на Рисунке 1, представляют собой типичное количество MN в одном участке помеченного моторного столбца. Частично это связано с редкостью или заражением бешенством, но, что более важно, с тем фактом, что каждый срез имеет толщину всего 30 мкм. Мы также добавили дополнительные рисунки (рисунки 1,2,3 к рисункам 1,2,3), показывающие больше примеров поясничных отделов после инъекций в различные моторные бассейны. Среди показанных примеров на одном участке инфицировано до 8 MN, но это редкое явление (рисунок 1 — приложение к рисунку 1A).
2. Показано небольшое количество необработанных данных. На рисунке показана одна иммунная панель. Я понимаю, что главная идея исходит из схем, но было бы неплохо увидеть изображения отделов позвоночника с большим количеством одиночных и двойных инфицированных клеток. Каждое иммуно-изображение содержит только один расходящийся PMI, и кроме рисунков 1 и 6 нет других клеток, которые кажутся инфицированными красным или зеленым экспрессирующим вирусом на иммуно-изображении.

На Рисунке 1 мы выделили 3 расходящихся интернейрона на изображении.Из-за относительно низкой плотности дивергентных премоторных нейронов и тонкости секций невозможно иметь больше расходящихся нейронов в одном отделе, особенно в грудном и шейном отделах спинного мозга. Чтобы обеспечить более широкую перспективу дивергентных премоторных нейронов, мы добавили больше примеров окрашенных срезов поясничного, грудного и шейного отделов спинного мозга, показывающих расходящиеся нейроны. Они представлены в приложениях к рисункам 1,2,3, связанным с рисунками 1, 2 и 3.
3. Мне немного неудобно интерпретировать данные в конце результатов — особенно с разделом, озаглавленным «Цервикальные LDPN» возбуждают — это предположение, и его не должно быть в заголовке.Данные просто показывают, что 95% этих клеток не являются глицинергическими.
Мы изменили заголовок этого раздела: «Цервикальные LDPN не являются ни ингибирующими, ни холинергическими» (строка 201), поскольку только 1/21 цервикальные LDPN были положительными по eGFP, и ни один из них не был положительным по ChAT.
4. Относится к пункту 3 — следующий раздел озаглавлен «Цервикальные премоторные LDPN, вероятно, возникают из домена V0v». Помимо клеток V0 и V1, Lhx1 также постмитотически экспрессируется в популяциях dI2, dI4, dI6 и V2 и даже в некоторых клетках V3 (Alanyick et al., 2012, Delile… .Бюджетная статья Сагнера https://doi.org/10.1101/472415). Таким образом, крайне спекулятивно предполагать, что именно клетки V0v — это именно те клетки, которые мы здесь рассматриваем. Даже в большей степени, если принять во внимание открытие, что Lhx1 играет ключевую роль в установлении идентичности ГАМКергических нейронов (Pillai et al., 2007).
Между прочим, текущее мнение о клетках V0v заключается не в том, что они контактируют с MN напрямую, а скорее дисинаптически через ингибирующий IN (см. Kiehn 2016 Nat Rev Nsci review). Было показано, что некоторые нейроны V0 и V3 проецируются на MN (как указано в строке 200, показанной в Lanuza et al., 2004 и Zhang et al., 2008), но в последнее время также было показано, что клетки V3 проецируются на мишени, не являющиеся MN (статья Chopek Cell Rep), и клетки V0, вполне возможно, имеют много других нижестоящих мишеней, это просто еще предстоит исследовать.
Я действительно считаю, что эту часть рукописи не следует включать из-за ее предварительного характера. Для включения должно быть какое-то окрашивание Evx1 или функциональный анализ у мышей с дефицитом V0v, использованный в статье Talpalar 2012, в котором проблемы с жесткостью мышц видны в отсутствие клеток V0v.
Причина, по которой у меня возникла такая проблема, заключается в том, что идентичность этих ячеек как принадлежащих к популяции V0v очень косвенно основана на данных здесь. Если бы это было опубликовано как есть, клетки V0v немедленно были бы упомянуты в работах других как ключевые регуляторы жесткости мышц. Даже в этой рукописи — строке 398 обсуждения авторы говорят, что подмножество помеченных LDPN «явно является производным от V0v». Это проблема.
 .
. Мы благодарим рецензента и соглашаемся, что наши доказательства идентификации интернейронов V0 не являются окончательными.При подготовке ревизии мы попытались идентифицировать интернейроны, происходящие от Evx1, с помощью специфических антител (подаренных Колумбийским университетом). Однако, хотя мы получили хорошее окрашивание молодой (P0) ткани, окрашивания не наблюдалось в ткани P9, возраст, в котором ткань анализировалась в этой статье. Попробовав много разных концентраций и времени инкубации, с аналогичными результатами, мы пришли к выводу, что экспрессия Evx1 подавляется даже в большей степени, чем Lhx1 (см. Рисунок 5 — рисунок в приложении 1) и, следовательно, апостериорная идентификация длинных нейронов нисходящей проекции невозможна.Только генетическая маркировка может решить проблему, но это невозможно, поскольку у нас нет соответствующей линии мышей. Чтобы усугубить нашу неопределенность, недавнее исследование было опубликовано, когда мы пересматривали нашу рукопись, определяя класс интернейронов, производных dI2, с морфологическими особенностями, подобными LDPN, описанным в нашем исследовании. Заметим, что входы dI2 также выражают Lhx1. Учитывая эту дополнительную неопределенность и сниженный уровень обнаружения Lhx1 в более зрелой ткани, мы смягчили любой вывод, который может указывать на то, что мы идентифицировали основной класс дивергентных LDPN и переписали соответствующий раздел (теперь он начинается со строки 219).
Однако, хотя мы получили хорошее окрашивание молодой (P0) ткани, окрашивания не наблюдалось в ткани P9, возраст, в котором ткань анализировалась в этой статье. Попробовав много разных концентраций и времени инкубации, с аналогичными результатами, мы пришли к выводу, что экспрессия Evx1 подавляется даже в большей степени, чем Lhx1 (см. Рисунок 5 — рисунок в приложении 1) и, следовательно, апостериорная идентификация длинных нейронов нисходящей проекции невозможна.Только генетическая маркировка может решить проблему, но это невозможно, поскольку у нас нет соответствующей линии мышей. Чтобы усугубить нашу неопределенность, недавнее исследование было опубликовано, когда мы пересматривали нашу рукопись, определяя класс интернейронов, производных dI2, с морфологическими особенностями, подобными LDPN, описанным в нашем исследовании. Заметим, что входы dI2 также выражают Lhx1. Учитывая эту дополнительную неопределенность и сниженный уровень обнаружения Lhx1 в более зрелой ткани, мы смягчили любой вывод, который может указывать на то, что мы идентифицировали основной класс дивергентных LDPN и переписали соответствующий раздел (теперь он начинается со строки 219). .Но мы предпочитаем хранить данные в рукописи, поскольку мы приводим доказательства того, что по крайней мере некоторые LDPN экспрессируют Lhx1 (и, следовательно, вероятно, V0v или dI2, как обсуждалось). См. Также ответ рецензента №2, относящийся к тому же вопросу.
.Но мы предпочитаем хранить данные в рукописи, поскольку мы приводим доказательства того, что по крайней мере некоторые LDPN экспрессируют Lhx1 (и, следовательно, вероятно, V0v или dI2, как обсуждалось). См. Также ответ рецензента №2, относящийся к тому же вопросу.
Рецензент №2 (Рекомендации авторам):
4) Может ли автор представить данные или упомянуть, что они тестировали, что чистые мыши RΦGT, которым вводили инъецированный вирус, не метили какие-либо премоторные нейроны?
Мы проверили утечку гликопротеина бешенства, выполнив инъекцию псевдотипированного вируса бешенства EnvA гетерозиготным мышам RΦGT.Мы решили использовать псевдотип бешенства, чтобы проверить эктопическую экспрессию как гликопротеина бешенства, так и рецептора TVA (поскольку эти мыши содержат флоксированные кассеты, экспрессирующие оба белка, Takatoh et al., 2013). Примеры данных показаны в нашем препринте, в котором мы описываем аналогичный подход (Ronzano et al. , 2021). Таким образом, после инъекций мы наблюдали скудную маркировку мотонейронов (1, 1 и 3 мотонейрона у каждого из тестируемых животных), но отсутствие интернейронов. Поскольку мы использовали псевдотип бешенства, инфицирование некоторых мотонейронов показывает, что существует некоторая эктопическая экспрессия TVA в мотонейронах, которая вызывает инфекцию мотонейронов из-за чрезвычайно высокого сродства EnvA к рецептору TVA.Однако не наблюдалось транссинаптического мечения, что указывает на то, что любой утечки G-белка было недостаточно для поддержания транссинаптических скачков. Теперь мы указываем это в конце раздела «Методы», посвященного внутримышечным инъекциям (строки 504-508).
, 2021). Таким образом, после инъекций мы наблюдали скудную маркировку мотонейронов (1, 1 и 3 мотонейрона у каждого из тестируемых животных), но отсутствие интернейронов. Поскольку мы использовали псевдотип бешенства, инфицирование некоторых мотонейронов показывает, что существует некоторая эктопическая экспрессия TVA в мотонейронах, которая вызывает инфекцию мотонейронов из-за чрезвычайно высокого сродства EnvA к рецептору TVA.Однако не наблюдалось транссинаптического мечения, что указывает на то, что любой утечки G-белка было недостаточно для поддержания транссинаптических скачков. Теперь мы указываем это в конце раздела «Методы», посвященного внутримышечным инъекциям (строки 504-508).
Рецензент №3 (Рекомендации авторам):
— Ранее в разделе «Результаты» следует упомянуть, что это исследование проводилось на новорожденных.
Это было разъяснено и теперь упоминается в строке 95 результатов.
— Зеленый и синий на иммуно-цифрах (т.
 е. сечениях на рис. 1Bi, рис. 2Ai) трудно различить из-за низкого контраста между ними.
е. сечениях на рис. 1Bi, рис. 2Ai) трудно различить из-за низкого контраста между ними. Цвета, к сожалению, выбрать сложно, если принять во внимание то, что люди действительно могут их увидеть по-разному. Мы проводим слишком много времени на лабораторных собраниях, обсуждая цвета. Нам пришлось выбрать два цвета, которые каждый мог бы легко различить для eGFP (зеленый) и mCherry (розовый), сохраняя при этом цвета, близкие к их флуоресцентному спектру излучения, чтобы избежать недоразумений.Кроме того, поскольку мы фокусируемся на нейронах с двойной меткой, совместная локализация двух цветов должна дать совершенно другой оттенок (в нашем случае белый). Синий оставался единственным вариантом в качестве четвертого цвета, так что его можно было легко отличить от трех других. Это правда, что синий цвет труднее визуализировать, однако, поскольку синий цвет используется для окрашивания мотонейронов, что не является предметом исследования, мы подумали, что использованная комбинация была наилучшим вариантом.
— Строка 9-11: Дивергент используется до того, как он определен.Он работает в начальном предложении (строка 9), но немного вводит в заблуждение в следующем (строки 10-11), поскольку, кажется, подразумевает, что дивергент относится только к моторным пулам антагонистов.
Дивергент теперь четко определен в аннотации перед его использованием.
— Строки 37-42: Я предлагаю скорректировать оформление этого раздела. На большинство этих вопросов есть ответы, как указано в последней строке абзаца и в Обсуждении. Вместо этого указание на то, что местоположение и идентичность многих из этих нейронов неизвестны, может лучше подготовить последующие результаты.
Мы включили предложение судьи и, сохраняя список вопросов, подчеркнули, что неизвестно местонахождение и идентичность интернейронов, опосредующих синергию (строки 46-48).
— Строки 146–148: неясно, к чему относятся эти 22%.
 Может быть, «где они составляют 22% помеченных нейронов»?
Может быть, «где они составляют 22% помеченных нейронов»?Это было разъяснено (строка 164).
— Схема на Рисунке S4 A подразумевает, что вентральный расходящийся интернейрон в шейном канатике является двойным прыжком из моторного пула передних конечностей.Вероятно, что на схеме присутствует «неправильная связь» — вентральный нейрон должен синапсировать с двигательными нейронами передних конечностей, а не дорсальный нейрон с вентральным нейроном.
Благодарим рецензента за этот комментарий и приносим свои извинения. Действительно, мы допустили ошибку в схеме того, что сейчас является рис. 6 — дополнением к рисунку 1А. Теперь это исправлено.
https://doi.org/10.7554/eLife.70858.sa2мышц-синергистов в эксцентрической фазе приседаний | Live Healthy
Приседания — обычное упражнение, выполняемое во всем мире спортсменами разного уровня, от соревновательного до развлекательного.Это можно делать со свободными весами, штангой или без веса. Его можно использовать для наращивания мышечной массы или реабилитации колена. Это требует баланса, уравновешенности, силы, сосредоточенности и гибкости. Вы можете практиковать приседания в тренажерном зале или дома.
Его можно использовать для наращивания мышечной массы или реабилитации колена. Это требует баланса, уравновешенности, силы, сосредоточенности и гибкости. Вы можете практиковать приседания в тренажерном зале или дома.
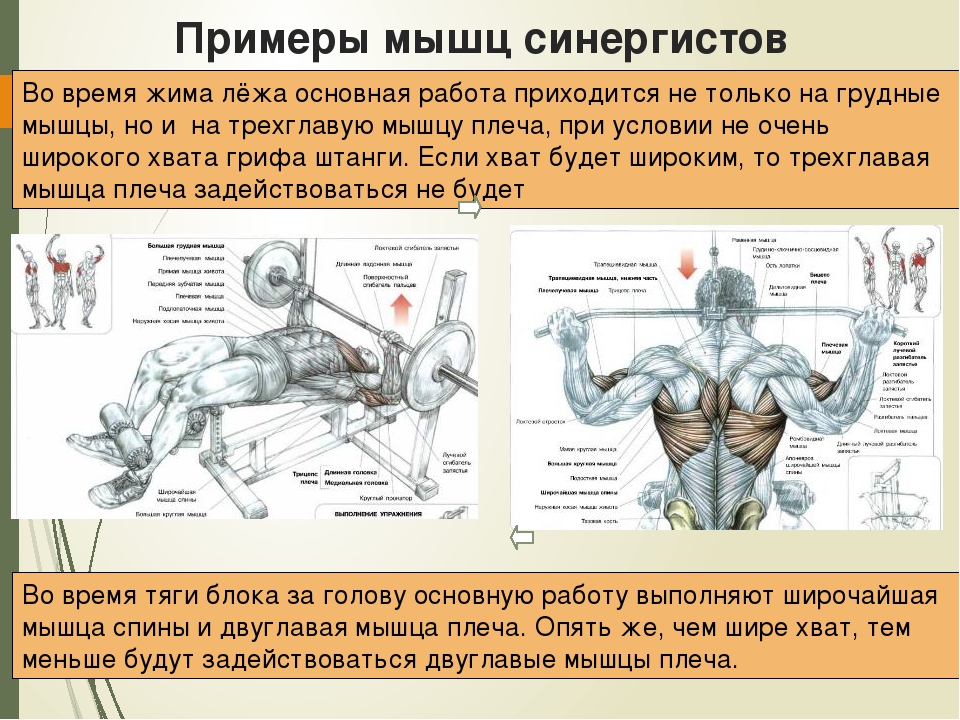
Определение синергиста и эксцентрика
Чтобы движение могло происходить, мышцы должны сокращаться. Согласно «Путеводителю по телу» Эндрю Биля, мышца, которая ведет действие в качестве основного двигателя, называется агонистом. Мышца, поддерживающая агонист, называется синергистом.Мышцы, которые работают вместе для достижения полного движения, называются синергистами. Концентрическое движение — это когда мышца сокращается и укорачивается одновременно. Эксцентрическое движение — это когда мышца одновременно сокращается, но удлиняется.
Эксцентрическая фаза
Приседания можно разделить на две части. Фаза опускания или опускания и фаза подъема или стояния. Когда вы приседаете, ваши бедра и колени сгибаются, а стопа сгибается в тыльном направлении.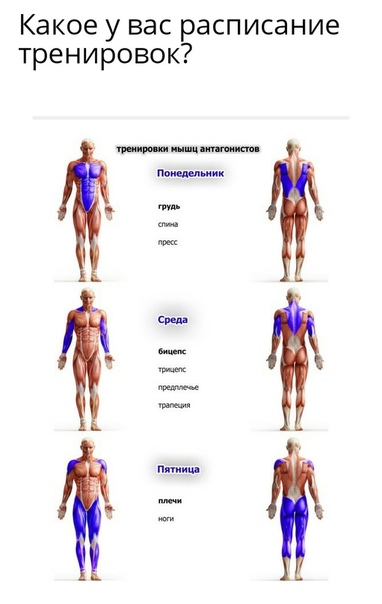 Во время фазы опускания вы удлиняете квадрицепсы, ягодицы и подколенные сухожилия. Таким образом, фаза опускания / опускания классифицируется как эксцентрическая часть приседа. Следовательно, фаза подъема / подъема — это концентрическая секция. Обратите внимание, что сгибание тыла — это когда верхняя часть стопы движется к голени или, в случае приседания, голень движется к верху стопы.
Во время фазы опускания вы удлиняете квадрицепсы, ягодицы и подколенные сухожилия. Таким образом, фаза опускания / опускания классифицируется как эксцентрическая часть приседа. Следовательно, фаза подъема / подъема — это концентрическая секция. Обратите внимание, что сгибание тыла — это когда верхняя часть стопы движется к голени или, в случае приседания, голень движется к верху стопы.
Мышцы-синергисты
Когда вы опускаетесь в эксцентрическую фазу приседания, все четыре квадрицепса вытягиваются.Четыре квадрицепса — это латеральная широкая мышца бедра, промежуточная широкая мышца бедра, медиальная широкая мышца бедра и прямая мышца бедра. Обратите внимание, что прямая мышца бедра пересекает бедро. Следовательно, даже если нижние волокна прямой мышцы бедра растягиваются из-за сгибания колена, верхние волокна укорачиваются из-за сгибания бедра. Точно так же верхние волокна мышц подколенного сухожилия — двуглавая мышца бедра, полусухожильная и полуперепончатая мышца — удлиняются из-за сгибания бедра, но нижние волокна сгибаются из-за сгибания колена.
Synergist Muscles (продолжение)
Большая ягодичная мышца, называемая большой ягодичной мышцей, также эксцентрично растягивается во время фазы приседания вниз. По мере того, как ваше колено продолжает сгибаться, а тыльная сторона стопы сгибается, вы также удлиняете камбаловидную мышцу, глубокие икроножные мышцы. Нижние волокна более крупной икроножной мышцы, икроножной мышцы, растягиваются из-за сгибания спины, но верхние волокна укорачиваются из-за сгибания колена. В любом случае, ваши квадрицепсы, ягодицы, подколенные сухожилия и икры считаются эксцентрическими в фазе опускания / опускания приседаний, известной как эксцентрическая фаза.
Ссылки
Writer Bio
Таня Сейжи Гершон специализируется на лечении хронической мышечной боли с помощью йоги и миофасциального расслабления. Она имеет степень бакалавра наук в области физиологии упражнений, является опытным преподавателем йоги в Yoga Alliance и сертифицированным специалистом по телу в NCTMB. Она опубликовала множество видео и статей о здоровье и благополучии в AZ Central Living, ModernMom, eHow, Chron, LIVESTRONG и TheNestWomen.
Разделение влияния задачи и синергетического вклада мышц для оценки нейрофизиологических механизмов явления двустороннего дефицита
Бете, Энн (2020) Разделение влияния задачи и синергистического мышечного вклада для оценки нейрофизиологических механизмов феномена двустороннего дефицита. Докторская диссертация, Питтсбургский университет. (Не опубликовано)
Аннотация
Феномен двустороннего дефицита (BLD) — это неспособность максимально сокращать двусторонние сокращения по сравнению с суммированными односторонними сокращениями. Механизм неизвестен, но наличие двустороннего гомологичного (BH) BLD в отличие от двусторонних негомологичных (BNH) сокращений предполагает, что BLD отражает различия в кортикоспинальном контроле. Тем не менее, влиянию методологических факторов, таких как методика расчета BLD, знакомство с задачами и различия в участии мышц-синергистов, уделялось меньше внимания. Цель: изучить кортикоспинальный и методологический вклад в BLD, чтобы определить его механистическую основу. Методы. Одиннадцать здоровых взрослых людей (6 женщин / 5 мужчин, 25,6 ± 3,7 года; 171,81 ± 11,44 см; 74,4 ± 21,2 кг) приняли участие в исследовании с повторными измерениями уравновешивания. Первый и седьмой сеансы, транскаллозальное торможение (TCI) и произвольная активация оценивались с помощью транскраниальной магнитной стимуляции во время максимальной ЧД, НД и доминантного сгибания (ДФ) проксимального отдела локтя. Сеансы со второго по шестой, двусторонние и односторонние изометрические сокращения выполнялись повторно с электромиографическими измерениями агонистов, антагонистов и мышечной активности, стабилизирующей лопатку.Результаты: BH BLD присутствовал на сеансах 2-7, в то время как BNH не отображал BI. Кортикоспинальные измерения не различались между сокращениями или сеансами. Практика выполнения увеличила все показатели максимальной силы, но без перевода на производительность BH и BNH BLD, с низкой надежностью во время сеансов тестирования.
Цель: изучить кортикоспинальный и методологический вклад в BLD, чтобы определить его механистическую основу. Методы. Одиннадцать здоровых взрослых людей (6 женщин / 5 мужчин, 25,6 ± 3,7 года; 171,81 ± 11,44 см; 74,4 ± 21,2 кг) приняли участие в исследовании с повторными измерениями уравновешивания. Первый и седьмой сеансы, транскаллозальное торможение (TCI) и произвольная активация оценивались с помощью транскраниальной магнитной стимуляции во время максимальной ЧД, НД и доминантного сгибания (ДФ) проксимального отдела локтя. Сеансы со второго по шестой, двусторонние и односторонние изометрические сокращения выполнялись повторно с электромиографическими измерениями агонистов, антагонистов и мышечной активности, стабилизирующей лопатку.Результаты: BH BLD присутствовал на сеансах 2-7, в то время как BNH не отображал BI. Кортикоспинальные измерения не различались между сокращениями или сеансами. Практика выполнения увеличила все показатели максимальной силы, но без перевода на производительность BH и BNH BLD, с низкой надежностью во время сеансов тестирования. BLD варьировался в зависимости от конкретной задачи стабилизационной практики. В частности, BNH и DF стабилизировались горизонтальным вращением верхней конечности, что приводило к большим силам по сравнению с BH, которая стабилизировалась в сагиттальной плоскости.Заключение: результаты этого исследования показывают, что BLD отражает тонкие различия в стабилизации мышц во время сокращений BH, BNH и DF по сравнению с различиями в кортикоспинальном контроле или знакомстве с заданием. Синергетическая совместная активация мышц во время максимальной изометрической BNH и односторонних сокращений, вероятно, улучшает стабильность, увеличивая силу. Однако наиболее важным выводом исследования является низкая надежность измерений BH и BNH BLD, что вызывает необходимость учитывать пороговые значения, используемые для определения наличия BLD.
BLD варьировался в зависимости от конкретной задачи стабилизационной практики. В частности, BNH и DF стабилизировались горизонтальным вращением верхней конечности, что приводило к большим силам по сравнению с BH, которая стабилизировалась в сагиттальной плоскости.Заключение: результаты этого исследования показывают, что BLD отражает тонкие различия в стабилизации мышц во время сокращений BH, BNH и DF по сравнению с различиями в кортикоспинальном контроле или знакомстве с заданием. Синергетическая совместная активация мышц во время максимальной изометрической BNH и односторонних сокращений, вероятно, улучшает стабильность, увеличивая силу. Однако наиболее важным выводом исследования является низкая надежность измерений BH и BNH BLD, что вызывает необходимость учитывать пороговые значения, используемые для определения наличия BLD.
Поделиться
| Образец цитирования / Экспорт: | Выберите формат . .. Цитирование — TextCitation — HTMLEndnoteBibTexDublin CoreOpenURLMARC (ISO 2709) METSMODSEP3 XMLReference ManagerRefer .. Цитирование — TextCitation — HTMLEndnoteBibTexDublin CoreOpenURLMARC (ISO 2709) METSMODSEP3 XMLReference ManagerRefer |
|---|---|
| Социальная сеть: |
Детали
| Тип изделия: | Университет Питтсбурга ETD |
|---|---|
| Статус: | Неопубликованные |
| Создатели / Авторы: | |
| Комитет ETD: | |
| Дата: | 19 июня 2020 |
| Тип даты: | Публикация |
| Дата защиты: | 4 февраля 2020 |
| Дата утверждения: | 19 июня 2020 |
| Дата отправки: | 27 марта 2020 |
| Ограничение доступа: | 2 года — Ограничить доступ в Университет Питтсбурга сроком на 2 года. |
| Количество страниц: | 160 |
| Учреждение: | Питтсбургский университет |
| Школы и программы: | Школа наук о здоровье и реабилитации> Спортивная медицина и питание |
| Степень: | PhD — доктор философских наук |
| Тип диссертации: | Докторская диссертация |
| Реферировано: | Есть |
| Неконтролируемые ключевые слова: | Транскраниальная магнитная стимуляция Двусторонний феномен дефицита Добровольная активация Транскаллозальное ингибирование |
| Дата депонирования: | 26 августа 2020 12:46 |
| Последнее изменение: | 26 августа 2020 12:46 |
| URI: | http: // d-scholarship. pitt.edu/id/eprint/38414 pitt.edu/id/eprint/38414 |
Метрики
просмотров по месяцам за последние 3 года
Plum Analytics
Действия (требуется логин)
| Просмотреть товар |
Пулы моторных нейронов синергетических мышц бедра разделяют большую часть своих синаптических входов
Введение
Давно подозревалось, что действия, затрагивающие несколько мышц, управляются с помощью упрощенного набора команд высокого уровня, отпечатки которых можно наблюдать как скоординированная деятельность мышц, т.е.е., «мышечная синергия» (для обзора см. Tresch and Jarc, 2009; Bizzi and Cheung, 2013). Однако существует важный спор относительно того, подразумевает ли такая активность, что несколько мышц управляются вместе с помощью общих нейронных входов, или просто отражает механические ограничения данной задачи (Tresch and Jarc, 2009; Kutch and Valero-Cuevas, 2012; Бицци и Чунг, 2013). У некоторых видов стимуляция кожного (Tresch et al., 1999), спинного (Giszter et al., 1993; Levine et al., 2014) или кортикального (Graziano et al., 2002; Overduin et al., 2012, 2014) нейроны производят скоординированную мышечную активность, что убедительно свидетельствует о создании мышечной синергии нервной системой. Тем не менее, остается много вопросов, связанных с использованием таких схем у людей и, в частности, с тем, в какой степени мышцы синергии управляются общими или независимыми входами. Наиболее распространенные методы, используемые для характеристики мышечной синергии у людей, не могут оценить распределение / доставку нервного импульса к активированным мышцам.
У некоторых видов стимуляция кожного (Tresch et al., 1999), спинного (Giszter et al., 1993; Levine et al., 2014) или кортикального (Graziano et al., 2002; Overduin et al., 2012, 2014) нейроны производят скоординированную мышечную активность, что убедительно свидетельствует о создании мышечной синергии нервной системой. Тем не менее, остается много вопросов, связанных с использованием таких схем у людей и, в частности, с тем, в какой степени мышцы синергии управляются общими или независимыми входами. Наиболее распространенные методы, используемые для характеристики мышечной синергии у людей, не могут оценить распределение / доставку нервного импульса к активированным мышцам.
На протяжении десятилетий нервно-мышечные управляющие сигналы изучались путем изучения взаимосвязанной активности моторных единиц (Sears and Stagg, 1976; De Luca et al., 1982; Rosenberg et al., 1989; Farmer et al., 1993; Halliday et al. ., 1995), потому что это отражает их общий (и «эффективный») нервный драйв (Farina et al. , 2014; Farina and Negro, 2015). Эта стратегия обычно не применялась для анализа групп мышц. В прошлых исследованиях было чрезвычайно редко, чтобы корреляции как внутри мышечных, так и между мышечных двигательных единиц оценивались во время выполнения двигательной задачи.Даже с такими измерениями невозможно определить относительную силу / пропорцию нервного импульса, который является уникальным для данной мышцы по сравнению с другими одновременно активированными мышцами.
, 2014; Farina and Negro, 2015). Эта стратегия обычно не применялась для анализа групп мышц. В прошлых исследованиях было чрезвычайно редко, чтобы корреляции как внутри мышечных, так и между мышечных двигательных единиц оценивались во время выполнения двигательной задачи.Даже с такими измерениями невозможно определить относительную силу / пропорцию нервного импульса, который является уникальным для данной мышцы по сравнению с другими одновременно активированными мышцами.
В этом исследовании мы преодолели эти ограничения, чтобы изучить простую синергию. В частности, мы исследовали две синергистические мышцы четырехглавой мышцы, латеральную широкую мышцу и медиальную широкую мышцу, во время создания двух изометрических сил разгибания колена (10 и 30% от максимального произвольного усилия).Для всесторонней оценки внутримышечного и поперечного нервного возбуждения мы использовали хорошо зарекомендовавший себя метод анализа когерентности двигательных единиц (Rosenberg et al., 1989; Farmer et al. , 1993; Halliday et al., 1995; Amjad et al. , 1997), который описывает частотный спектр нейронного входа. Затем мы применили новый метод, основанный на анализе частичной когерентности, чтобы разделить общий нервный импульс к каждой мышце на общие (кросс-мышечные) и уникальные (специфичные для мышц) компоненты и оценить относительную силу каждой из них.
, 1993; Halliday et al., 1995; Amjad et al. , 1997), который описывает частотный спектр нейронного входа. Затем мы применили новый метод, основанный на анализе частичной когерентности, чтобы разделить общий нервный импульс к каждой мышце на общие (кросс-мышечные) и уникальные (специфичные для мышц) компоненты и оценить относительную силу каждой из них.
Наша общая гипотеза заключалась в том, что две мышцы живота будут контролироваться главным образом общим нервным двигателем, с относительно небольшим уникальным двигателем к каждой мышце. Этот прогноз был мотивирован высокой степенью синхронизированных разрядов двигательных единиц (Mellor and Hodges, 2005) и колебаниями скорости стрельбы (Beck et al., 2012) в двух васти-мышцах, которые достигают величин, аналогичных тем, которые были зарегистрированы для внутренних мышц. корреляции мышечных двигательных единиц (De Luca et al., 2009; Beck et al., 2011).
Идея о том, что несколькими мышцами можно управлять, главным образом, с помощью совместного ввода, имеет большое значение для понимания синергии мышц. Наше исследование представляет собой первую прямую проверку (и прямую поддержку) этой концепции на уровне нервного импульса у людей для простой задачи и синергии. Мы также представляем наиболее полную на сегодняшний день характеристику нервного импульса к vasti-мышцам.
Наше исследование представляет собой первую прямую проверку (и прямую поддержку) этой концепции на уровне нервного импульса у людей для простой задачи и синергии. Мы также представляем наиболее полную на сегодняшний день характеристику нервного импульса к vasti-мышцам.
Результаты
Разложение моторных единиц
После исключения любых испытаний, в которых каждая мышца имела менее трех моторных единиц, разложенных для любого уровня силы, для дополнительного анализа был доступен общий набор данных из 80 испытаний с участием девяти субъектов.Всего было проанализировано девять сокращений на каждый уровень силы для каждого из первых восьми субъектов, тогда как восемь испытаний были проанализированы для субъекта 9. У субъекта 10 было слишком мало единиц, разложенных на уровне 10% MVC, чтобы быть включенным в дополнительный анализ. По записям обширной мышцы латеральной мышцы, среднее ± стандартное отклонение количество разложенных моторных единиц составило 7,5 ± 2,3 для испытаний 10% MVC и 7 ± 3,2 для испытаний 30% MVC. Среднее значение ± стандартное отклонение для этих единиц составляло 8,8 ± 1,0 Гц при 10% MVC и 10,7 ± 1,6 Гц при 30% MVC. От медиальной широкой мышцы бедра — в среднем 7.7 ± 2,8 единиц были разложены при 10% MVC и 7,5 ± 3,5 единиц при 30% MVC. Среднее значение ± стандартное отклонение частоты активации двигательных единиц широкой мышцы бедра составило 9 ± 0,9 и 10,9 ± 1,6 Гц для 10 и 30% MVC, соответственно.
Согласованность моторных единиц в мышцах и между ними
На рисунке 3, A и B показан процент испытаний, которые показали значительную согласованность на каждой частоте для латеральной широкой и медиальной мышц бедра соответственно. Пропорции, превышающие отмеченный 95% CL, указывают на то, что когерентность наблюдалась чаще, чем ожидалось случайно.Цифры показывают, какие частоты были постоянными составляющими общего толчка каждой мышцы для каждого уровня силы. Ниже оси x каждого графика приведены результаты рандомизационного теста, сравнивающего величину согласованности между двумя уровнями силы. Черные полосы представляют частоты, на которых когерентность значительно различалась. Для удобства интерпретации значение p для значимости в этом тесте было установлено на 0,01, что позволяет каждой полосе представлять интервал 1 Гц (пять частотных выборок).Общие профили когерентности сходны между обеими мышцами, при этом основные характеристики нервного возбуждения включают компонент 1–5 Гц, компонент ~ 10 Гц и компонент ~ 20 Гц, причем последние два встречаются только при 30% MVC. При сравнении уровней силы различия в когерентности наблюдались на частотах> 5 Гц и были особенно сильными в медиальной широкой мышце бедра (компоненты ∼10 и ∼20 Гц демонстрируют сильную зависимость от силы).
Черные полосы представляют частоты, на которых когерентность значительно различалась. Для удобства интерпретации значение p для значимости в этом тесте было установлено на 0,01, что позволяет каждой полосе представлять интервал 1 Гц (пять частотных выборок).Общие профили когерентности сходны между обеими мышцами, при этом основные характеристики нервного возбуждения включают компонент 1–5 Гц, компонент ~ 10 Гц и компонент ~ 20 Гц, причем последние два встречаются только при 30% MVC. При сравнении уровней силы различия в когерентности наблюдались на частотах> 5 Гц и были особенно сильными в медиальной широкой мышце бедра (компоненты ∼10 и ∼20 Гц демонстрируют сильную зависимость от силы).
Общая согласованность внутримышечных двигательных единиц и согласованность мышечных двигательных единиц для широких мышц. A и B показывают долю испытаний (всего 80), в которых наблюдалась значительная согласованность внутримышечных двигательных единиц для латеральной широкой мышцы бедра и медиальной широкой мышцы бедра соответственно.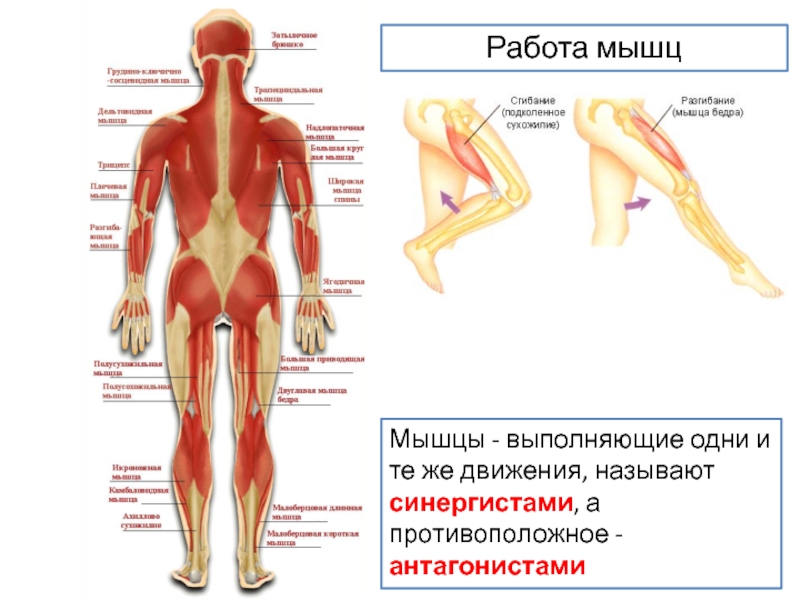 Красные кривые показывают результаты для мышечных сокращений, удерживаемых при 30% MVC, тогда как коричневые кривые показывают результаты для 10% MVC. Пунктирная горизонтальная линия указывает на самый высокий процент, который можно было наблюдать просто случайно. Ниже оси x каждого графика приведены результаты рандомизационного теста, сравнивающего значения когерентности, наблюдаемые при 10% MVC с 30% MVC.Черные полосы указывают частоты, при которых существовала значительная разница между величинами когерентности, измеренными в испытаниях на каждом уровне силы. Уровень значимости был установлен на 0,01 для корректировки множественных сравнений и, таким образом, позволяет каждой полосе представлять интервал частот 1 Гц. C и D представляют тот же анализ, что и в A и B , но после устранения влияния кросс-мышечной активности на внутримышечную когерентность.Следовательно, показанные остатки представляют собой частотную составляющую нервного импульса, специфичного для мышц.
Красные кривые показывают результаты для мышечных сокращений, удерживаемых при 30% MVC, тогда как коричневые кривые показывают результаты для 10% MVC. Пунктирная горизонтальная линия указывает на самый высокий процент, который можно было наблюдать просто случайно. Ниже оси x каждого графика приведены результаты рандомизационного теста, сравнивающего значения когерентности, наблюдаемые при 10% MVC с 30% MVC.Черные полосы указывают частоты, при которых существовала значительная разница между величинами когерентности, измеренными в испытаниях на каждом уровне силы. Уровень значимости был установлен на 0,01 для корректировки множественных сравнений и, таким образом, позволяет каждой полосе представлять интервал частот 1 Гц. C и D представляют тот же анализ, что и в A и B , но после устранения влияния кросс-мышечной активности на внутримышечную когерентность.Следовательно, показанные остатки представляют собой частотную составляющую нервного импульса, специфичного для мышц. E и F показывают результаты рандомизационного теста, сравнивающего величину общей внутримышечной когерентности с остаточной (специфичной для мышц) когерентностью при 30% MVC (вверху) и 10% MVC (внизу) . Обе мышцы демонстрируют схожие профили общей внутримышечной когерентности, при этом основной компонент каждой возникает ниже 5 Гц, но распространяется дальше и включает компоненты около 10 и 20 Гц, когда сила увеличивается с 10% MVC до 30% MVC.Величина когерентности <5 Гц, по-видимому, не изменяется с силой, в отличие от высокочастотных компонентов, которые действительно показывают силовую зависимость. Специфическая для мышц когерентность была слабее, ограничивалась в основном частотами <5 Гц и не сильно зависела от силы.
E и F показывают результаты рандомизационного теста, сравнивающего величину общей внутримышечной когерентности с остаточной (специфичной для мышц) когерентностью при 30% MVC (вверху) и 10% MVC (внизу) . Обе мышцы демонстрируют схожие профили общей внутримышечной когерентности, при этом основной компонент каждой возникает ниже 5 Гц, но распространяется дальше и включает компоненты около 10 и 20 Гц, когда сила увеличивается с 10% MVC до 30% MVC.Величина когерентности <5 Гц, по-видимому, не изменяется с силой, в отличие от высокочастотных компонентов, которые действительно показывают силовую зависимость. Специфическая для мышц когерентность была слабее, ограничивалась в основном частотами <5 Гц и не сильно зависела от силы.
Рисунок 3, C и D , изображает процент испытаний, имеющих значительную когерентность после статистического удаления общего кросс-мышечного сигнала из общей согласованности внутримышечных двигательных единиц. Результирующая остаточная когерентность отражает уникальное, «специфическое для мышцы» движение к данной мышце.В целом, для каждой из мышц, по-видимому, существует только компонент мышечной активности с частотой 1–5 Гц. Небольшой пик на 18 Гц присутствовал для медиальной широкой мышцы бедра (30% MVC), хотя он был слабым с точки зрения силы, ширины полосы и последовательности. Не было различий в специфическом для мышцы толчке на латеральную широкую мышцу бедра при разных уровнях силы, тогда как для медиальной широкой мышцы бедра были небольшие различия, в основном <3 Гц.
Результирующая остаточная когерентность отражает уникальное, «специфическое для мышцы» движение к данной мышце.В целом, для каждой из мышц, по-видимому, существует только компонент мышечной активности с частотой 1–5 Гц. Небольшой пик на 18 Гц присутствовал для медиальной широкой мышцы бедра (30% MVC), хотя он был слабым с точки зрения силы, ширины полосы и последовательности. Не было различий в специфическом для мышцы толчке на латеральную широкую мышцу бедра при разных уровнях силы, тогда как для медиальной широкой мышцы бедра были небольшие различия, в основном <3 Гц.
На рис. 3, E и F показаны результаты рандомизационного теста, сравнивающего величины общей когерентности с остаточной (специфичной для мышц) когерентностью при 30% MVC (вверху) и 10% MVC (внизу).При 30% MVC компонент, специфичный для мышц, был значительно меньше, чем общая когерентность, почти на каждой частоте, где он существовал, и в обеих мышцах. То же самое верно и для 10% MVC.
На рис. 4 показан процент испытаний, показывающих значительную согласованность поперечных мышечных двигательных единиц. В этом случае остаточная когерентность представляет собой оставшуюся когерентность после вычитания любых компонентов, которые коррелируют с силой. Рисунок 4 A показывает, что межмышечная когерентность содержала по существу такое же частотное содержание, что и внутримышечная когерентность, опять же с частотами выше ~ 5 Гц, показывающими значительную зависимость от силы.Рисунок 4 B показывает, что компонент поперечного мышечного движения, не коррелированный с временными колебаниями силы, зависел от общего уровня силы. При 30% MVC частоты выше ~ 8 Гц формируют важные компоненты остаточной когерентности, что означает, что кросс-мышечный драйв на этом уровне силы не был точно переведен в силу. На рис. 4 C показано явное снижение низкочастотной кросс-мышечной когерентности после удаления любых компонентов, синхронизированных с колебаниями силы.При 10% MVC, по существу, каждая частота <6 Гц была уменьшена, тогда как при 30% MVC все частоты <10 Гц были уменьшены.
4 показан процент испытаний, показывающих значительную согласованность поперечных мышечных двигательных единиц. В этом случае остаточная когерентность представляет собой оставшуюся когерентность после вычитания любых компонентов, которые коррелируют с силой. Рисунок 4 A показывает, что межмышечная когерентность содержала по существу такое же частотное содержание, что и внутримышечная когерентность, опять же с частотами выше ~ 5 Гц, показывающими значительную зависимость от силы.Рисунок 4 B показывает, что компонент поперечного мышечного движения, не коррелированный с временными колебаниями силы, зависел от общего уровня силы. При 30% MVC частоты выше ~ 8 Гц формируют важные компоненты остаточной когерентности, что означает, что кросс-мышечный драйв на этом уровне силы не был точно переведен в силу. На рис. 4 C показано явное снижение низкочастотной кросс-мышечной когерентности после удаления любых компонентов, синхронизированных с колебаниями силы.При 10% MVC, по существу, каждая частота <6 Гц была уменьшена, тогда как при 30% MVC все частоты <10 Гц были уменьшены. Эффекты> 10 Гц при 30% MVC были менее стойкими.
Эффекты> 10 Гц при 30% MVC были менее стойкими.
Поперечная когерентность мышц и ее отражение в силе разгибания колена. A показывает долю испытаний, имеющих значительную согласованность поперечных мышечных двигательных единиц. Как и раньше, красные кривые показывают результаты для 30% MVC силы, а коричневые кривые показывают результаты для 10% MVC. Общая частотная составляющая кросс-мышечного драйва была очень похожа на мышечную тягу с основным компонентом от 1 до 5 Гц, который расширялся и включал компоненты около 10 и 20 Гц при 30% MVC.Ниже оси x оси A результаты рандомизационного теста показывают, что нервный драйв выше ∼5 Гц демонстрирует четкую зависимость от силы, тогда как самые низкие частоты (ниже ∼5 Гц) — нет. B показывает тот же анализ, что и A , но после удаления любых компонентов межмышечной связности, которые были синхронизированы с силой разгибания колена. Остаточная когерентность представляет собой кросс-мышечный драйв, который не преобразуется в колебания силы.Опять же, частоты в диапазоне 10–20 Гц зависели от силы, тогда как частотные составляющие <5 Гц, как правило, не зависели. C показывает сравнение общей с остаточной когерентностью на каждом уровне силы. Удаление силы из межмышечной когерентности значительно уменьшило ее величину почти на каждой частоте <10 Гц, при которой когерентность наблюдалась постоянно. Снижение высокочастотной (10–20 Гц) когерентности было менее выраженным.
Остаточная когерентность представляет собой кросс-мышечный драйв, который не преобразуется в колебания силы.Опять же, частоты в диапазоне 10–20 Гц зависели от силы, тогда как частотные составляющие <5 Гц, как правило, не зависели. C показывает сравнение общей с остаточной когерентностью на каждом уровне силы. Удаление силы из межмышечной когерентности значительно уменьшило ее величину почти на каждой частоте <10 Гц, при которой когерентность наблюдалась постоянно. Снижение высокочастотной (10–20 Гц) когерентности было менее выраженным.
Вклад мышечного возбуждения в общую (внутримышечную) когерентность
Для каждой мышцы и уровня силы рассчитывалась область значительной внутримышечной когерентности двигательных единиц до и после устранения эффектов кросс-мышечного возбуждения .Остаточная согласованность представляет собой уникальный, специфичный для мышц драйв. На рисунке 5 соотношение между площадью остаточной когерентности и общей площадью когерентности внутри мышцы показано для каждого пациента для каждой мышцы. Верхняя строка показывает результаты для 30% сокращений MVC, а нижняя показывает результаты для 10% сокращений MVC. Высота столбцов представляет собой среднюю долю, рассчитанную по испытаниям, завершенным каждым участником, а столбцы ошибок представляют собой стандартное отклонение по повторным испытаниям. Эти планки погрешностей показывают приблизительную согласованность этой меры в разных испытаниях и не предназначены для статистических сравнений.Доля общей согласованности двигательных единиц, объясняемая мышечным драйвом, была низкой во всех случаях, в среднем <20% у большинства испытуемых.
Верхняя строка показывает результаты для 30% сокращений MVC, а нижняя показывает результаты для 10% сокращений MVC. Высота столбцов представляет собой среднюю долю, рассчитанную по испытаниям, завершенным каждым участником, а столбцы ошибок представляют собой стандартное отклонение по повторным испытаниям. Эти планки погрешностей показывают приблизительную согласованность этой меры в разных испытаниях и не предназначены для статистических сравнений.Доля общей согласованности двигательных единиц, объясняемая мышечным драйвом, была низкой во всех случаях, в среднем <20% у большинства испытуемых.
Пропорция общей внутримышечной связности, представленная мышечным толчком. Доля общей согласованности внутримышечных двигательных единиц, объясняемая независимым движением к латеральной широкой мышце бедра (слева) или медиальной широкой мышце бедра (справа), показана для каждого пациента. Каждая полоса представляет собой среднюю долю, рассчитанную по всем испытаниям для данного субъекта. Столбики ошибок показывают стандартное отклонение пропорций, рассчитанных для пробных повторений для каждого человека, и указывают на то, что показатель был относительно стабильным при испытаниях и сеансах записи. При силе разгибания колена 30% MVC (вверху) и 10% MVC (внизу) почти все испытания показали пропорции мышечного толчка <20%. Другими словами,> 80% когерентности между единицами, измеренной в любой из мышц, было связано с кросс-мышечным возбуждением.
Столбики ошибок показывают стандартное отклонение пропорций, рассчитанных для пробных повторений для каждого человека, и указывают на то, что показатель был относительно стабильным при испытаниях и сеансах записи. При силе разгибания колена 30% MVC (вверху) и 10% MVC (внизу) почти все испытания показали пропорции мышечного толчка <20%. Другими словами,> 80% когерентности между единицами, измеренной в любой из мышц, было связано с кросс-мышечным возбуждением.
Доля когерентности поперечных мышц, не связанная с силой
На рисунке 6 показана доля когерентности поперечных мышц, остающаяся после удаления любых компонентов, которые были синхронными с колебаниями силы во времени.Как для 30% MVC (вверху), так и для 10% MVC (внизу) доля когерентности, не связанная с силой, была <20% для всех субъектов. Это означает, что межмышечный нервный драйв тесно связан с общей силой.
Рис. 6. Пропорция когерентности поперечных мышц, не связанная с силой разгибания колена. На рисунке изображена пропорция когерентности поперечных мышечных двигательных единиц, которая не коррелировала с силой.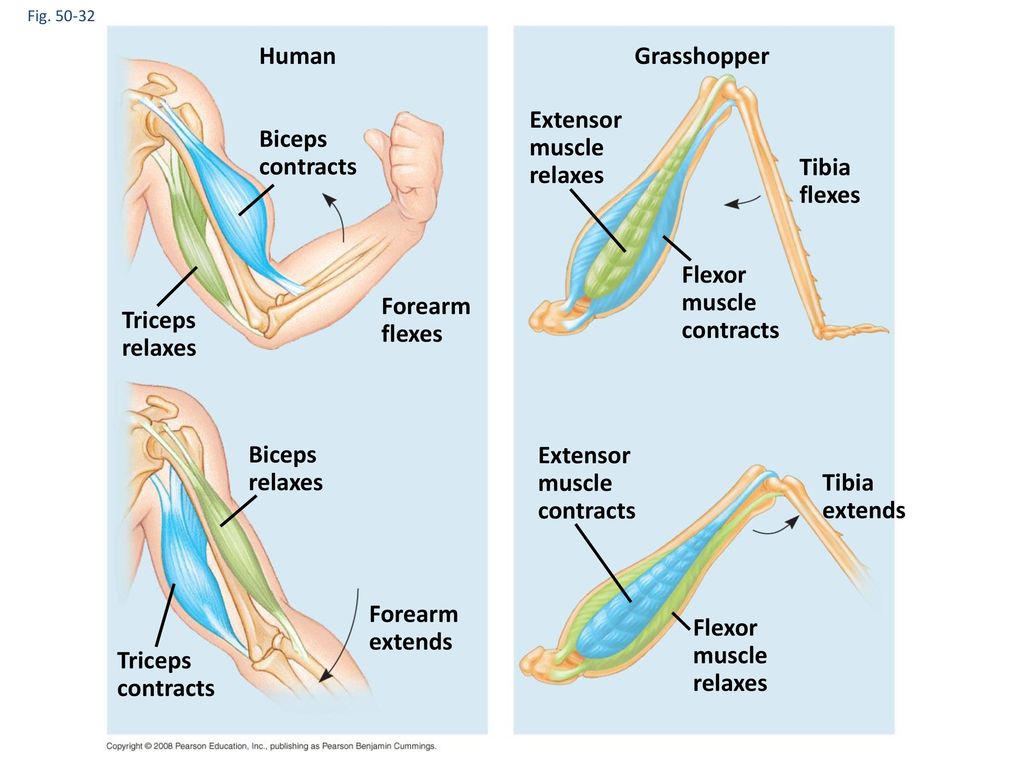 Как при 30% MVC (вверху), так и при 10% MVC (внизу) доля очень мала, что позволяет предположить, что большая часть (> 80%) кросс-мышечного драйва связана с силой.Как и на рисунке 5, столбики ошибок показывают SD пропорций, рассчитанных по пробным повторам для каждого человека.
Как при 30% MVC (вверху), так и при 10% MVC (внизу) доля очень мала, что позволяет предположить, что большая часть (> 80%) кросс-мышечного драйва связана с силой.Как и на рисунке 5, столбики ошибок показывают SD пропорций, рассчитанных по пробным повторам для каждого человека.
Обсуждение
В этом исследовании мы охарактеризовали частотный состав и силовую зависимость как общего (поперечно-мышечного), так и независимого (специфически для мышц) нервного импульса к двигательным единицам мышц-синергистов. Наши результаты предоставляют прямую нейрофизиологическую поддержку теории о том, что синергетически активируемые мышцы контролируются главным образом общим нервным импульсом.Наше исследование также представляет собой наиболее полную на сегодняшний день характеристику нервного импульса к vasti-мышцам.
Частотный состав нервного импульса к латеральной широкой мышце бедра и медиальной широкой мышце бедра был очень похож. При 10% MVC нервный драйв к любой из мышц охватил частоты до ~ 6 Гц со значительной когерентностью <3 Гц в большинстве исследований. В общем, когерентность моторных единиц на частотах <5 Гц отражает общее стремление, то есть одновременные колебания скорости их работы (De Luca et al., 1982; Myers et al., 2004). Общее побуждение неизвестно физиологического происхождения, но самые низкие частоты (<3 Гц) не затрагиваются капсульным инсультом (Farmer et al., 1993) и усиливаются при мозжечковом инсульте (Sauvage et al., 2006). Общее побуждение распространяется на пары мышц-антагонистов, если они функционально связаны (De Luca and Mambrito, 1987) и может зависеть от контекста задачи (Mochizuki et al., 2006; Laine et al., 2013, 2014) и проприоцептивного воздействия (De Luca et al. al., 2009; Laine et al., 2014).
В общем, когерентность моторных единиц на частотах <5 Гц отражает общее стремление, то есть одновременные колебания скорости их работы (De Luca et al., 1982; Myers et al., 2004). Общее побуждение неизвестно физиологического происхождения, но самые низкие частоты (<3 Гц) не затрагиваются капсульным инсультом (Farmer et al., 1993) и усиливаются при мозжечковом инсульте (Sauvage et al., 2006). Общее побуждение распространяется на пары мышц-антагонистов, если они функционально связаны (De Luca and Mambrito, 1987) и может зависеть от контекста задачи (Mochizuki et al., 2006; Laine et al., 2013, 2014) и проприоцептивного воздействия (De Luca et al. al., 2009; Laine et al., 2014).
При 10% MVC нервный драйв на частотах> 6 Гц не сильно отражался на когерентности двигательных единиц любой из мышц.Когда сила сокращения была увеличена до 30% MVC, наблюдался входной сигнал 6–12 Гц (пик между 8 и 10 Гц), а также новый компонент в бета-диапазоне (∼15–35 Гц) с пиком на 20 Гц. . Нейронный драйв в диапазоне частот 6–12 Гц связан с физиологическим тремором и может частично возникать из-за колебаний возбуждения вокруг афферентной петли обратной связи Ia (Sutton and Sykes, 1967; Lippold, 1970; Hagbarth and Young, 1979; Young and Hagbarth).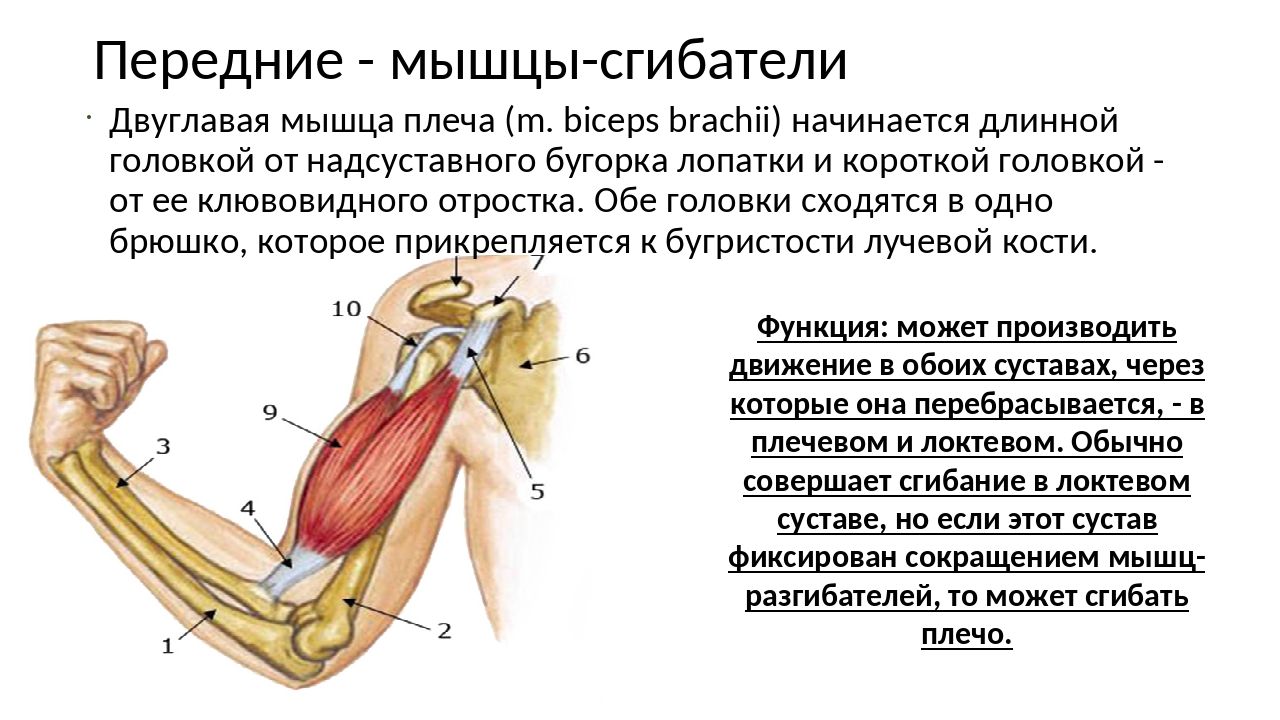 , 1980; Erimaki, Christakos, 1999, 2008; Christakos et al., 2006). Считается, что высокочастотные компоненты нервного импульса (15–35 Гц) имеют корковое происхождение (Farmer et al., 1993, 1997; Conway et al., 1995; Salenius et al., 1997; Brown et al. ., 1998). Если эти ассоциации верны для настоящего сценария, из этого следует, что афферентная обратная связь и корковое возбуждение были достаточно сильными только для того, чтобы вызвать значительную когерентность двигательных единиц при уровнях силы> 10% MVC. Конечно, отсутствие значительной когерентности может не указывать на полное отсутствие нервного импульса на высоких частотах, учитывая множество факторов, которые могут ограничивать чувствительность мер когерентности (Negro and Farina, 2012).
, 1980; Erimaki, Christakos, 1999, 2008; Christakos et al., 2006). Считается, что высокочастотные компоненты нервного импульса (15–35 Гц) имеют корковое происхождение (Farmer et al., 1993, 1997; Conway et al., 1995; Salenius et al., 1997; Brown et al. ., 1998). Если эти ассоциации верны для настоящего сценария, из этого следует, что афферентная обратная связь и корковое возбуждение были достаточно сильными только для того, чтобы вызвать значительную когерентность двигательных единиц при уровнях силы> 10% MVC. Конечно, отсутствие значительной когерентности может не указывать на полное отсутствие нервного импульса на высоких частотах, учитывая множество факторов, которые могут ограничивать чувствительность мер когерентности (Negro and Farina, 2012).
Согласованность поперечных мышечных двигательных единиц была почти идентична когерентности внутримышечных двигательных единиц с точки зрения как частотного содержания, так и зависимости от силы. Пики ∼10 и ∼20 Гц в когерентности двигательных единиц при 30% MVC особенно интересны, потому что они предполагают, что кросс-мышечный драйв содержит как обратную связь Ia, так и нисходящий кортикальный компонент. Удивительно, но наш анализ частичной когерентности показал, что эти более высокие частоты когерентности отражают исключительно кросс-мышечный драйв, потому что уникальный импульс к любой из мышц не содержал значительной когерентности выше ∼6 Гц.Стоит отметить, что если корковое возбуждение — это, по сути, сигнал между мышцами, то наши результаты напрямую подтверждают основной принцип теории мышечной синергии, а именно, что корковые команды имеют более низкую размерность, чем контролируемые мышцы.
Удивительно, но наш анализ частичной когерентности показал, что эти более высокие частоты когерентности отражают исключительно кросс-мышечный драйв, потому что уникальный импульс к любой из мышц не содержал значительной когерентности выше ∼6 Гц.Стоит отметить, что если корковое возбуждение — это, по сути, сигнал между мышцами, то наши результаты напрямую подтверждают основной принцип теории мышечной синергии, а именно, что корковые команды имеют более низкую размерность, чем контролируемые мышцы.
Мы обнаружили, что колебания силы <6 Гц были хорошо синхронизированы с поперечным мышечным толчком. Для обоих уровней силы подавляющее большинство (> 80%) согласованности поперечных мышечных двигательных единиц объяснялось колебаниями силы. Во время визуально управляемого контроля силы низкочастотные колебания силы отражают добровольную коррекцию ошибок и непроизвольную модуляцию общей скорости стрельбы (общий драйв) среди моторных единиц (Sutton and Sykes, 1967; Allum et al., 1978; Де Лука и др. , 1982; Миалл и др., 1993; Слифкин и др., 2000; Squeri et al., 2010). Из-за эффектов фильтрации нижних частот мышечной ткани и остальной части системы фиксации ноги / ноги колебания силы выше ~ 5 Гц чрезвычайно малы и пренебрежимо малы с точки зрения общего контроля силы (исключая сценарий патологического тремора). . Как и ожидалось, кросс-мышечный драйв> 10 Гц не очень хорошо коррелировал с колебаниями силы.
, 1982; Миалл и др., 1993; Слифкин и др., 2000; Squeri et al., 2010). Из-за эффектов фильтрации нижних частот мышечной ткани и остальной части системы фиксации ноги / ноги колебания силы выше ~ 5 Гц чрезвычайно малы и пренебрежимо малы с точки зрения общего контроля силы (исключая сценарий патологического тремора). . Как и ожидалось, кросс-мышечный драйв> 10 Гц не очень хорошо коррелировал с колебаниями силы.
Оказалось, что уникальный нервный импульс к каждой мышце ограничен общим частотным диапазоном возбуждения (1–5 Гц).В этом исследовании специфическая для мышц когерентность отражает либо фактическое уникальное воздействие на одну мышцу, либо поперечный мышечный сигнал, который был скрыт в одной из двух мышц, например, в результате шума. Привод 1–5 Гц к каждой vasti-мышце неизменно демонстрировал специфический для мышцы компонент. Несмотря на то, что он относительно слабый, его последовательность предполагает, что он действительно представляет собой отдельный источник частоты 1–5 Гц.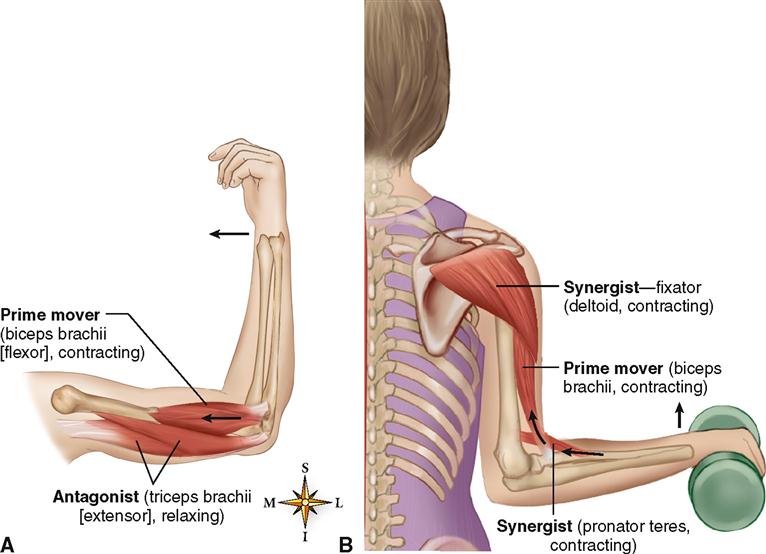 Независимо от источника, величина когерентности 1–5 Гц не сильно зависит от силы. Тем не менее, взаимосвязь между силой когерентности и силой нервного импульса может быть несколько сложной и зависит от многих факторов, включая физическое распределение аксонов, а также скорость активации моторных единиц по отношению к частоте совместного входа (Negro и Фарина, 2012).
Независимо от источника, величина когерентности 1–5 Гц не сильно зависит от силы. Тем не менее, взаимосвязь между силой когерентности и силой нервного импульса может быть несколько сложной и зависит от многих факторов, включая физическое распределение аксонов, а также скорость активации моторных единиц по отношению к частоте совместного входа (Negro и Фарина, 2012).
Мы также наблюдали небольшой компонент специфического для мышц движения к медиальной широкой мышце бедра при 18 Гц во время 30% сокращений MVC. Ограниченная полоса пропускания (1 Гц) и постоянство (на два испытания больше, чем ожидалось случайно) этого входного сигнала предполагают, что поперечное мышечное возбуждение ~ 20 Гц плохо отражается на латеральной широкой мышце бедра, что делает ее уникальной для медиальной широкой мышцы бедра. Сама широкая мышца бедра не показывала никаких сигналов, специфичных для мышц, с частотой более 6 Гц.
Что касается пропорций, мы обнаружили, что внутримышечная когерентность, относящаяся к мышечной активности, составляла от 5 до 25% от общей внутримышечной связности, независимо от уровня силы или мышцы, с которой регистрировались. Это соотношение в целом было постоянным для разных субъектов, испытаний и уровней силы. Хотя обнаруживаемая полоса кросс-мышечного возбуждения была больше при более высоких уровнях силы, общая степень согласованности внутримышечных двигательных единиц, связанная с кросс-мышечным возбуждением, была довольно стабильной. Хотя увеличение силы привело к большей полосе пропускания поперечного мышечного возбуждения, полученный высокочастотный сигнал мало повлиял на силу, и в результате доля когерентной активности, не коррелированной с силой, оказалась больше при 30% MVC. чем при 10% MVC.
Это соотношение в целом было постоянным для разных субъектов, испытаний и уровней силы. Хотя обнаруживаемая полоса кросс-мышечного возбуждения была больше при более высоких уровнях силы, общая степень согласованности внутримышечных двигательных единиц, связанная с кросс-мышечным возбуждением, была довольно стабильной. Хотя увеличение силы привело к большей полосе пропускания поперечного мышечного возбуждения, полученный высокочастотный сигнал мало повлиял на силу, и в результате доля когерентной активности, не коррелированной с силой, оказалась больше при 30% MVC. чем при 10% MVC.
Поскольку наши результаты подтверждают теорию о том, что синергетически активируемые мышцы контролируются главным образом общим нервным импульсом, важно дополнительно обсудить, как этот результат дополняет предыдущую литературу, поддерживающую понятие мышечной синергии. Существует множество доказательств того, что синергия или «моторные примитивы» действительно имеют нейронное происхождение (см. Обзор Bizzi and Cheung, 2013). Например, они могут быть вызваны и изменены посредством афферентной обратной связи в препаратах спинализованных лягушек (Tresch et al., 1999; Kargo and Giszter 2000a, b), они могут быть вызваны посредством стимуляции спинномозговых интернейронов у лягушек (Giszter et al., 1993) и мышей (Levine et al., 2014), а у макак-резусов они могут быть задействованы посредством внутрикортикальной микростимуляции ( Graziano et al., 2002; Overduin et al., 2012, 2014). У лягушек спинальные интернейроны, участвующие в организации моторных примитивов, были широко охарактеризованы (Hart and Giszter, 2010). Тем не менее, многие вопросы остаются без ответа, особенно в контексте произвольного моторного контроля у людей.Во-первых, люди (и некоторые высшие приматы) имеют прямые кортико-мотонейрональные клетки, которые могли развиться специально, чтобы освободить произвольное поведение от ограничений спинномоторных примитивов (Rathelot and Strick, 2009). Конечно, эти прямые кортико-мотонейрональные клетки сосуществуют со «старым» кортикоспинальным треком, в котором клетки M1 проецируются непосредственно на спинномозговые интернейроны, которые могут координировать активацию мышц (Rathelot and Strick, 2009).
Например, они могут быть вызваны и изменены посредством афферентной обратной связи в препаратах спинализованных лягушек (Tresch et al., 1999; Kargo and Giszter 2000a, b), они могут быть вызваны посредством стимуляции спинномозговых интернейронов у лягушек (Giszter et al., 1993) и мышей (Levine et al., 2014), а у макак-резусов они могут быть задействованы посредством внутрикортикальной микростимуляции ( Graziano et al., 2002; Overduin et al., 2012, 2014). У лягушек спинальные интернейроны, участвующие в организации моторных примитивов, были широко охарактеризованы (Hart and Giszter, 2010). Тем не менее, многие вопросы остаются без ответа, особенно в контексте произвольного моторного контроля у людей.Во-первых, люди (и некоторые высшие приматы) имеют прямые кортико-мотонейрональные клетки, которые могли развиться специально, чтобы освободить произвольное поведение от ограничений спинномоторных примитивов (Rathelot and Strick, 2009). Конечно, эти прямые кортико-мотонейрональные клетки сосуществуют со «старым» кортикоспинальным треком, в котором клетки M1 проецируются непосредственно на спинномозговые интернейроны, которые могут координировать активацию мышц (Rathelot and Strick, 2009).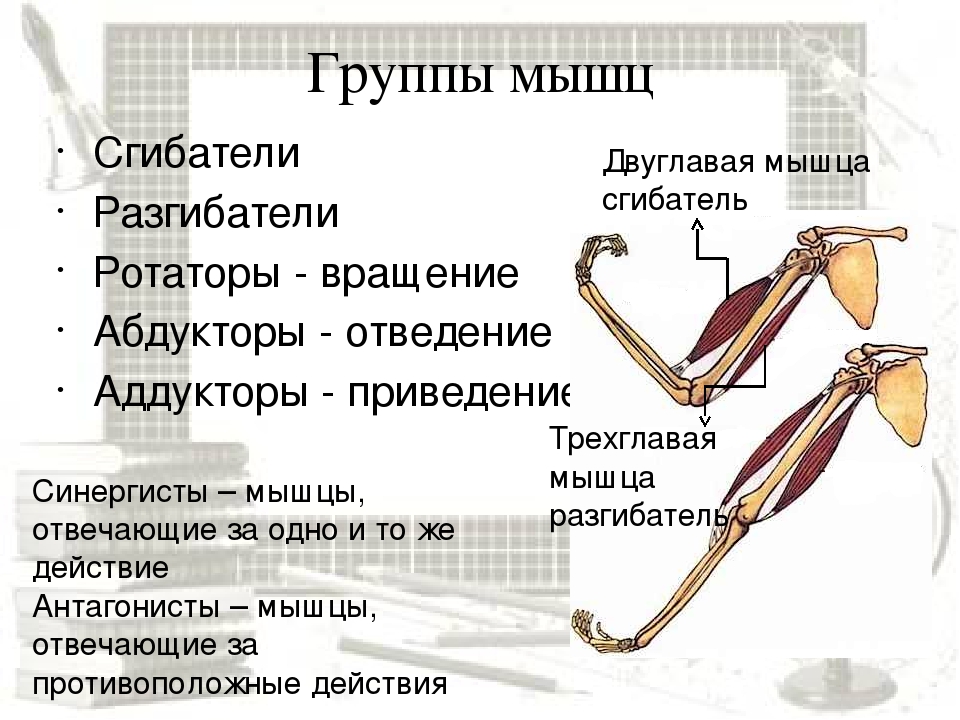 Независимо от того, на каком конкретном уровне могут быть подготовлены двигательные примитивы (спинномозговые или корковые), неясно, как именно этот сигнал доставляется к целевым мышцам.Например, несколько мышц могут показывать фиксированные коэффициенты активации, даже если каждая мышца получает свой собственный уникальный вход и не получает общего / общего толчка. С другой стороны, моторные нейроны мышц-синергистов могут совместно использовать низкочастотный общий импульс, возникающий на уровне позвоночника, но не высокочастотный нервный импульс, исходящий из коры головного мозга. Точная схема формирования синергии не совсем понятна, и наши результаты показывают, что ценную информацию можно получить, подходя к этой проблеме с уровня моторных нейронов.
Независимо от того, на каком конкретном уровне могут быть подготовлены двигательные примитивы (спинномозговые или корковые), неясно, как именно этот сигнал доставляется к целевым мышцам.Например, несколько мышц могут показывать фиксированные коэффициенты активации, даже если каждая мышца получает свой собственный уникальный вход и не получает общего / общего толчка. С другой стороны, моторные нейроны мышц-синергистов могут совместно использовать низкочастотный общий импульс, возникающий на уровне позвоночника, но не высокочастотный нервный импульс, исходящий из коры головного мозга. Точная схема формирования синергии не совсем понятна, и наши результаты показывают, что ценную информацию можно получить, подходя к этой проблеме с уровня моторных нейронов.
Потребуются дополнительные исследования, чтобы полностью понять, как разделяемое и специфическое для мышц влечение к синергетически активируемым мышцам изменяется в зависимости от контекста задачи или заболевания. Например, вполне вероятно, что характеристика синергической активации мышц может дать важную информацию о пластичности и адаптации ЦНС после травмы, например, инсульта (Gizzi et al. , 2011). Кроме того, теория когерентности и частичной когерентности содержит множество вариаций и расширений, которые были описаны ранее (Rosenberg et al., 1989, 1998), и это может быть особенно полезно при изучении координации между большими группами мышц. Важно отметить, что наши результаты обеспечивают первую нейронную поддержку у людей для предположения, что мускулами можно управлять главным образом с помощью общего нервного импульса (Tresch and Jarc, 2009; Kutch and Valero-Cuevas, 2012; Bizzi and Cheung, 2013). В целом, наше исследование расширило текущее понимание активации мышц живота и представило новый подход к исследованию нервного импульса к многомышечным системам.
, 2011). Кроме того, теория когерентности и частичной когерентности содержит множество вариаций и расширений, которые были описаны ранее (Rosenberg et al., 1989, 1998), и это может быть особенно полезно при изучении координации между большими группами мышц. Важно отметить, что наши результаты обеспечивают первую нейронную поддержку у людей для предположения, что мускулами можно управлять главным образом с помощью общего нервного импульса (Tresch and Jarc, 2009; Kutch and Valero-Cuevas, 2012; Bizzi and Cheung, 2013). В целом, наше исследование расширило текущее понимание активации мышц живота и представило новый подход к исследованию нервного импульса к многомышечным системам.
Синергетическая абляция как модель на грызунах для изучения динамики сателлитных клеток в скелетных мышцах взрослых — Профили экспертов по исследованиям в области здравоохранения UTMB
@article {d66d1a5d2adb41f99a5c562e0fe6a429,
title = «Синергетическая абляция как модель сателлитных мышц у взрослых грызунов для изучения спутниковой динамики клеток грызунов. «,
«,
abstract =» В скелетных мышцах взрослого человека сателлитные клетки являются первичными миогенными стволовыми клетками, участвующими в миогенезе. Обычно они остаются в состоянии покоя до активации стимулом, после чего они размножаются, дифференцируются и сливаются в существующий миофибра или формируют миофибер de novo.Для изучения динамики сателлитных клеток на моделях взрослых мышей в большинстве исследований используются модели регенерации, в которых мышца серьезно повреждена и требует участия сателлитных клеток для восстановления. Здесь мы описываем модель для изучения поведения сателлитных клеток при мышечной гипертрофии, которая не зависит от регенерации мышц. Операция синергистической абляции включает хирургическое удаление икроножной и камбаловидной мышц, что приводит к функциональной перегрузке оставшейся подошвенной мышцы. Эта функциональная перегрузка приводит к гипертрофии миофибрилл, а также к активации, пролиферации и слиянию сателлитных клеток с миофибриллами.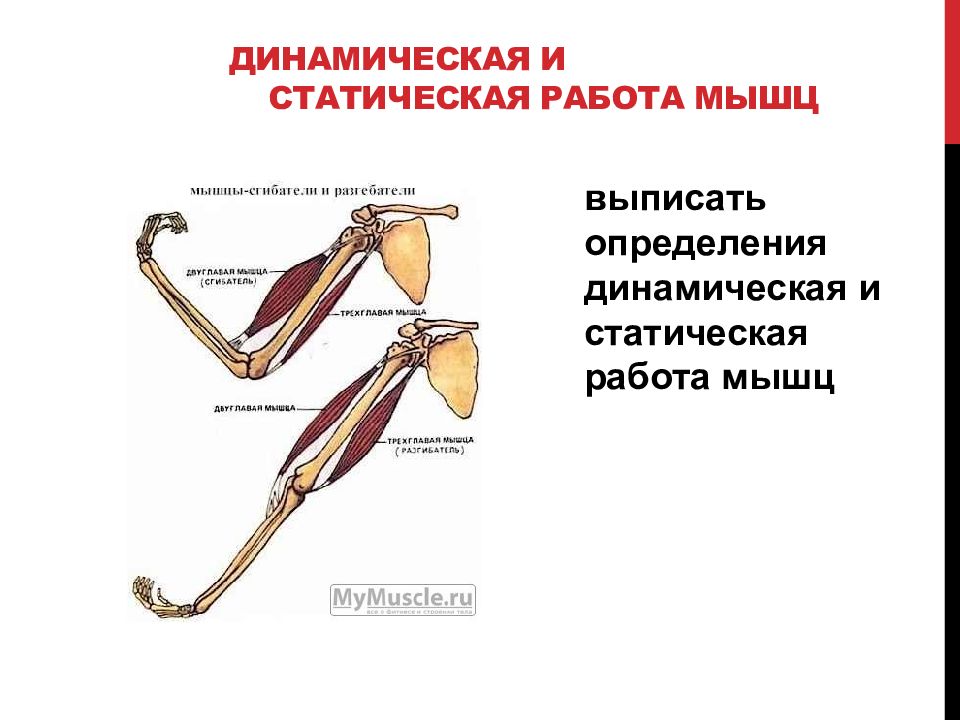 В течение 2 недель функциональной перегрузки содержание сателлитных клеток увеличивается примерно на 275%, что сопровождается увеличением количества миоядер примерно на 60%. Следовательно, это может быть использовано в качестве альтернативной модели для изучения поведения сателлитных клеток в зрелом возрасте, которое отличается от регенерации и способно выявить новые функции сателлитных клеток в регулировании адаптации мышц. »,
В течение 2 недель функциональной перегрузки содержание сателлитных клеток увеличивается примерно на 275%, что сопровождается увеличением количества миоядер примерно на 60%. Следовательно, это может быть использовано в качестве альтернативной модели для изучения поведения сателлитных клеток в зрелом возрасте, которое отличается от регенерации и способно выявить новые функции сателлитных клеток в регулировании адаптации мышц. »,
keywords =« Функциональная перегрузка, Гипертрофия, Сателлитные клетки. , Скелетная мышца, Синергетическая абляция «,
author =» Кирби, {Тайлер Дж.} и Маккарти, {Джон Дж.} и Петерсон, {Шарлотта А.} и Фрай, {Кристофер С.} «,
год =» 2016 «,
месяц = янв,
день =» 1 «,
doi = «10.1007 / 978-1-4939-3810-0_4»,
language = «English (US)»,
volume = «1460»,
pages = «43-52»,
journal = «Методы молекулярной биологии»,
issn = «1064-3745»,
publisher = «Humana Press»,
}
Дифференциальная потребность в клетках-сателлитах во время гипертрофии мышц, вызванной перегрузкой, у растущих мышей по сравнению со зрелыми мышами | Скелет Мышца
Кац Б. Окончания афферентного нервного волокна в мышечном веретене лягушки. Фил Trans Royal Soc Lond Ser B, Biol Sci. 1961; 243: 221-40.
Мауро А. Сателлитная клетка волокон скелетных мышц. J Biophy Biochem Cyt. 1961; 9: 493–5.
CAS Статья Google Scholar
Lepper C, Partridge TA, Fan C-M. Абсолютная потребность в Pax7-положительных сателлитных клетках при регенерации скелетных мышц, вызванной острым повреждением.Разработка. 2011. 138 (17): 3639–46.
CAS Статья PubMed PubMed Central Google Scholar
Маккарти Дж., Мула Дж., Миядзаки М., Эрфани Р., Гаррисон К., Фаруки А.Б., Срикуа Р., Лоусон Б.А., Граймс Б., Келлер С. и др. Эффективная гипертрофия волокон в скелетных мышцах, лишенных сателлитных клеток. Разработка. 2011. 138 (17): 3657–66.
CAS Статья PubMed PubMed Central Google Scholar

Мерфи М.М., Лоусон Д.А., Мэтью С.Дж., Хатчесон Д.А., Кардон Г. Клетки-сателлиты, фибробласты соединительной ткани и их взаимодействия имеют решающее значение для регенерации мышц. Разработка. 2011. 138 (17): 3625–37.
CAS Статья PubMed PubMed Central Google Scholar
Sambasivan R, Yao R, Kissenpfennig A, Van Wittenberghe L, Paldi A, Gayraud-Morel B, Guenou H, Malissen B, Tajbakhsh S, Galy A. Экспрессирующие Pax7 сателлитные клетки незаменимы для скелетных мышц взрослых регенерация.Разработка. 2011. 138 (17): 3647–56.
CAS Статья PubMed Google Scholar
Стокдейл Ф., Хольцер Х. Синтез ДНК и миогенез. Exp Cell Res. 1961; 24: 508–20.
CAS Статья PubMed Google Scholar
Мосс Ф., Леблон С. Природа делящихся ядер в скелетных мышцах растущих крыс. J Cell Biol. 1970. 44 (2): 459–61.
1970. 44 (2): 459–61.
CAS Статья PubMed PubMed Central Google Scholar
Мох FP, Леблон CP. Клетки-сателлиты как источник ядер в мышцах растущих крыс. Анат Рек. 1971. 170 (4): 421–35.
CAS Статья PubMed Google Scholar
Pullman WE, Yeoh GC. Роль миоядер в регенерации мышц: исследование in vitro. J. Cell Phys. 1978. 96 (2): 245–51.
CAS Статья Google Scholar
Скьяффино С., Бормиоли С.П., Алоизи М.Разрастание клеток в скелетных мышцах крыс на ранних стадиях компенсаторной гипертрофии. Virchow Archiv B Cell Patho. 1972. 11 (3): 268–73.
CAS Google Scholar
Кайсар Р., Ларссон Л. Что определяет размер миоядерного домена? Ind J Physiol Pharmacol. 2013; 58 (1): 1–12.
Google Scholar

Teixeira CE, Duarte JA. Миоядерный домен в волокнах скелетных мышц.Критический обзор. Archiv Exerc Health Dis. 2011. 2 (2): 92–101.
Артикул Google Scholar
Ван дер Меер С., Ясперс Р., Дегенс Х. Фиксирован ли размер миоядерного домена? J Musculo Neuro взаимодействовать. 2011; 11 (4): 286–97.
Google Scholar
Cheek D, Holt A, Hill D, Talbert J. Масса и рост скелетных мышц: концепция дезоксирибонуклеиновой единицы. Ped Res.1971; 5: 312–28.
Артикул Google Scholar
Холл З.В., Ральстон Э. Ядерные домены в мышечных клетках. Клетка. 1989; 59: 771–2.
CAS Статья PubMed Google Scholar
Lowe DA, Alway SE. Вызванная растяжением экспрессия миогенина, MyoD и MRF4 и острая гипертрофия в медленных тонических мышцах перепелов не зависят от пролиферации сателлитных клеток. Cell Tiss Res. 1999. 296 (3): 531–9.
Cell Tiss Res. 1999. 296 (3): 531–9.
CAS Статья Google Scholar
Амтор Х., Отто А., Вулин А., Роша А., Дюмонсо Дж., Гарсия Л., Муизель Е., Уорд С., Мачария Р., Фридрихс М. и др. Гипертрофия мышц, вызванная блокадой миостатина, не требует активности стволовых клеток / клеток-предшественников. Proc Nat Acad Sci U S. A. 2009; 106 (18): 7479–84.
CAS Статья Google Scholar
Ли С.Дж., Хьюнь ТВ, Ли Ю.С., Себальд С.М., Уилкокс-Адельман С.А., Ивамори Н., Леппер К., Мацук М.М., Фан С.М. Роль сателлитных клеток по сравнению с миофибриллами в мышечной гипертрофии, вызванной ингибированием сигнального пути миостатин / активин. Proc Nat Acad Sci U S. A. 2012; 109 (35): E2353–60.
CAS Статья Google Scholar
Ван К., Макферрон А.С. Ингибирование миостатина вызывает гипертрофию мышечных волокон до активации сателлитных клеток. J Physiol. 2012. 590 (9): 2151–65.
J Physiol. 2012. 590 (9): 2151–65.
CAS Статья PubMed PubMed Central Google Scholar
Welle S, Bhatt K, Pinkert CA, Tawil R, Thornton CA. Рост мышц после нокаута гена миостатина после развития. Am J Physiol Endo Metab. 2007; 292 (4): E985–991.
CAS Статья Google Scholar
Блаау Б., Канато М., Агатея Л., Тониоло Л., Маммукари К., Масьеро Е., Абрахам Р., Сандри М., Скьяффино С., Реджиани К.Индуцируемая активация Akt увеличивает массу и силу скелетных мышц без активации сателлитных клеток. FASEB J. 2009; 23 (11): 3896–905.
CAS Статья PubMed Google Scholar
Рафаэлло А., Милан Дж., Мазиеро Э., Карнио С., Ли Д., Ланфранки Дж., Голдберг А. Л., Сандри М. Фактор транскрипции JunB поддерживает массу скелетных мышц и способствует гипертрофии. J Cell Biol. 2010.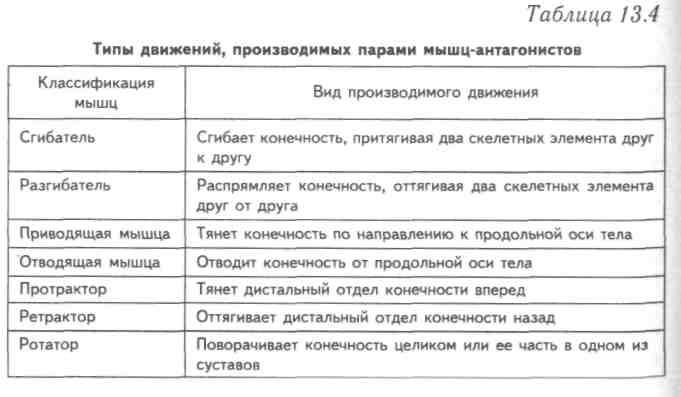 191 (1): 101–13.
191 (1): 101–13.
CAS Статья PubMed PubMed Central Google Scholar
Fry CS, Kosmac KC, McCarthy JJ, Peterson CA. Клетки-сателлиты регулируют ремоделирование внеклеточного матрикса во время адаптации скелетных мышц. Стволовая клетка. 2016; 20 (1): 56–69.
Артикул PubMed Google Scholar
Фрай С.С., Ли Дж. Д., Джексон Дж. Р., Кирби Т. Дж., Стаско С. А., Лю Х., Дюпон-Верстегден Е. Е., Маккарти Дж. Дж., Петерсон, Калифорния. Регулирование микросреды мышечных волокон активированными сателлитными клетками во время гипертрофии.FASEB J. 2014; 28 (4): 1654–65.
CAS Статья PubMed PubMed Central Google Scholar
Кирби Т.Дж., Патель Р.М., Макклинток Т.С., Дюпон-Верстегден Э., Петерсон, Калифорния, Маккарти Дж. Дж. Миоядерная транскрипция реагирует на механическую нагрузку и содержание ДНК, но не зависит от размера клетки во время гипертрофии. Molec Biol Cell. 2016; 27 (5): 788–98.
Molec Biol Cell. 2016; 27 (5): 788–98.
CAS Статья PubMed PubMed Central Google Scholar
McCarthy JJ, Dupont-Versteegden EE, Fry CS, Murach KA, Peterson CA. Методологические вопросы ограничивают интерпретацию негативных эффектов истощения сателлитных клеток на мышечную гипертрофию взрослых. Разработка. 2017; 144 (8): 1363–5.
Артикул PubMed Google Scholar
Фрай К.С., Ли Дж. Д., Мула Дж., Кирби Т. Дж., Джексон Дж. Р., Лю Ф., Ян Л., Мендиас К. Л., Дюпон-Верстегден Е. Э., Маккарти Дж. Дж. И др. Индуцируемое истощение сателлитных клеток у взрослых малоподвижных мышей ухудшает способность к регенерации мышц, не влияя на саркопению.Nat Med. 2015; 21 (1): 76–80.
CAS Статья PubMed Google Scholar
Лю В., Вей-Лапьер Л., Клозе А., Дирксен Р. Т., Чаккалакал СП. Индуцируемое истощение стволовых клеток взрослых скелетных мышц нарушает регенерацию нервно-мышечных соединений. Элиф. 2015; 4: e09221.
Индуцируемое истощение стволовых клеток взрослых скелетных мышц нарушает регенерацию нервно-мышечных соединений. Элиф. 2015; 4: e09221.
PubMed Central Google Scholar
White RB, Biérinx A-S, Gnocchi VF, Zammit PS.Динамика роста мышечных волокон в постнатальном развитии мышей. BMC Dev Biol. 2010; 10 (1): 21.
Артикул PubMed PubMed Central Google Scholar
Bruusgaard J, Liestøl K, Gundersen K. Распределение миоядер и микротрубочек в живых мышечных волокнах молодых, средних и старых мышей. J Appl Physiol. 2006; 100 (6): 2024–2030.
CAS Статья PubMed Google Scholar
Павликовски Б., Пуллиам С., Бетта Н.Д., Кардон Г., Олвин Б.Б. Повсеместный вклад сателлитных клеток в неповрежденные мышечные волокна взрослого человека. Skeletal Musc. 2015; 5: 42.
Артикул Google Scholar

Goodman CA, Frey JW, Mabrey DM, Jacobs BL, Lincoln HC, You JS, Hornberger TA. Роль mTOR скелетных мышц в регуляции роста, вызванного механической нагрузкой. J Physiol. 2011. 589 (22): 5485–501.
CAS Статья PubMed PubMed Central Google Scholar
Brack AS, Bildsoe H, Hughes SM. Доказательства того, что декремент сателлитных клеток способствует преимущественному снижению числа ядер из крупных волокон во время возрастной мышечной атрофии у мышей. J Cell Sci. 2005. 118 (20): 4813–21.
CAS Статья PubMed Google Scholar
Olsen S, Aagaard P, Kadi F, Tufekovic G, Verney J, Olesen JL, Suetta C, Kjr M. Добавка креатина увеличивает увеличение количества сателлитных клеток и миоядер в скелетных мышцах человека, вызванное силовыми тренировками.J Physiol. 2006; 573 (2): 525-534.
Кади Ф., Шерлинг П., Андерсен Л.Л. , Чарифи Н., Мэдсен Дж. Л., Кристенсен Л. Р., Андерсен Дж. Л.: Влияние тренировок с отягощениями и детренированности на сателлитные клетки в скелетных мышцах человека. J Physiol. 2004; 558 (3): 1005-012.
, Чарифи Н., Мэдсен Дж. Л., Кристенсен Л. Р., Андерсен Дж. Л.: Влияние тренировок с отягощениями и детренированности на сателлитные клетки в скелетных мышцах человека. J Physiol. 2004; 558 (3): 1005-012.
Петрелла Дж. К., Ким Дж. С., Кросс Дж. М., Косек Д. Д., Бамман М. М.. Эффективность миоядерного добавления может объяснить дифференцированный рост миофибрилл у молодых и пожилых мужчин и женщин, тренирующихся с отягощениями. Am J Physiol Endo Metab.2006; 291 (5): E937–946.
CAS Статья Google Scholar
Фрай С.С., Ноэрен Б., Мула Дж., Убеле М.Ф., Вестгейт ПМ, Керн, Пенсильвания, Петерсон, Калифорния. Специфический для типа волокна ответ сателлитных клеток на аэробную тренировку у взрослых, ведущих малоподвижный образ жизни. J Physiol. 2014; 592: 2625-635.
Мурач К.А., Уолтон Р.Г., Фрай С.С., Михаэлис С.Л., Грошонг Дж.С., Финлин Б.С., Керн PA, Петерсон, Калифорния. Циклическая тренировка модулирует сателлитные клетки и реакцию транскрипции на упражнения с отягощениями. Phys Rep.2016; 4 (18): e12973.
Phys Rep.2016; 4 (18): e12973.
Артикул Google Scholar
Петрелла Дж. К., Ким Дж. С., Мэйхью Д. Л., Кросс Дж. М., Бамман М. М.. Сильная гипертрофия миофибрилл во время тренировок с отягощениями у людей связана с миоядерным добавлением, опосредованным сателлитными клетками: кластерный анализ. J Appl Physiol. 2008. 104 (6): 1736–42.
Артикул PubMed Google Scholar
Ли Дж. Д., Фрай С. С., Мула Дж., Кирби Т. Дж., Джексон Дж. Р., Лю Ф., Ян Л., Дюпон-Верстегден Е. Э., Маккарти Дж. Дж., Петерсон, Калифорния.Старые мышцы демонстрируют адаптацию по типу волокон в ответ на механическую перегрузку в отсутствие гипертрофии миофибрилл, независимо от количества сателлитных клеток. J Gerentol Ser A Bio Sci Med Sci. 2016; 71 (4): 461–7.
Google Scholar
Куанг С., Поверенный С.Б., Сил П., Хух М. , Рудницки М.А. Различная роль Pax7 и Pax3 в регенеративном миогенезе взрослых. J Cell Biol. 2006. 172 (1): 103–13.
, Рудницки М.А. Различная роль Pax7 и Pax3 в регенеративном миогенезе взрослых. J Cell Biol. 2006. 172 (1): 103–13.
CAS Статья PubMed PubMed Central Google Scholar
Relaix F, Rocancourt D, Mansouri A, Buckingham M. Pax3 / Pax7-зависимая популяция клеток-предшественников скелетных мышц. Природа. 2005. 435 (7044): 948–53.
CAS Статья PubMed Google Scholar
Сил П., Сабурин Л.А., Гиргис-Габардо А, Мансури А., Грусс П., Рудницки М.А. Pax7 необходим для спецификации миогенных сателлитных клеток. Клетка. 2000. 102 (6): 777–86.
CAS Статья PubMed Google Scholar
Фергюсон В.Л., Айерс Р.А., Бейтман Т.А., Симске С.Дж. Развитие костной ткани и возрастная потеря костной массы у мышей-самцов C57BL / 6J. Кость. 2003. 33 (3): 387–98.
Артикул PubMed Google Scholar
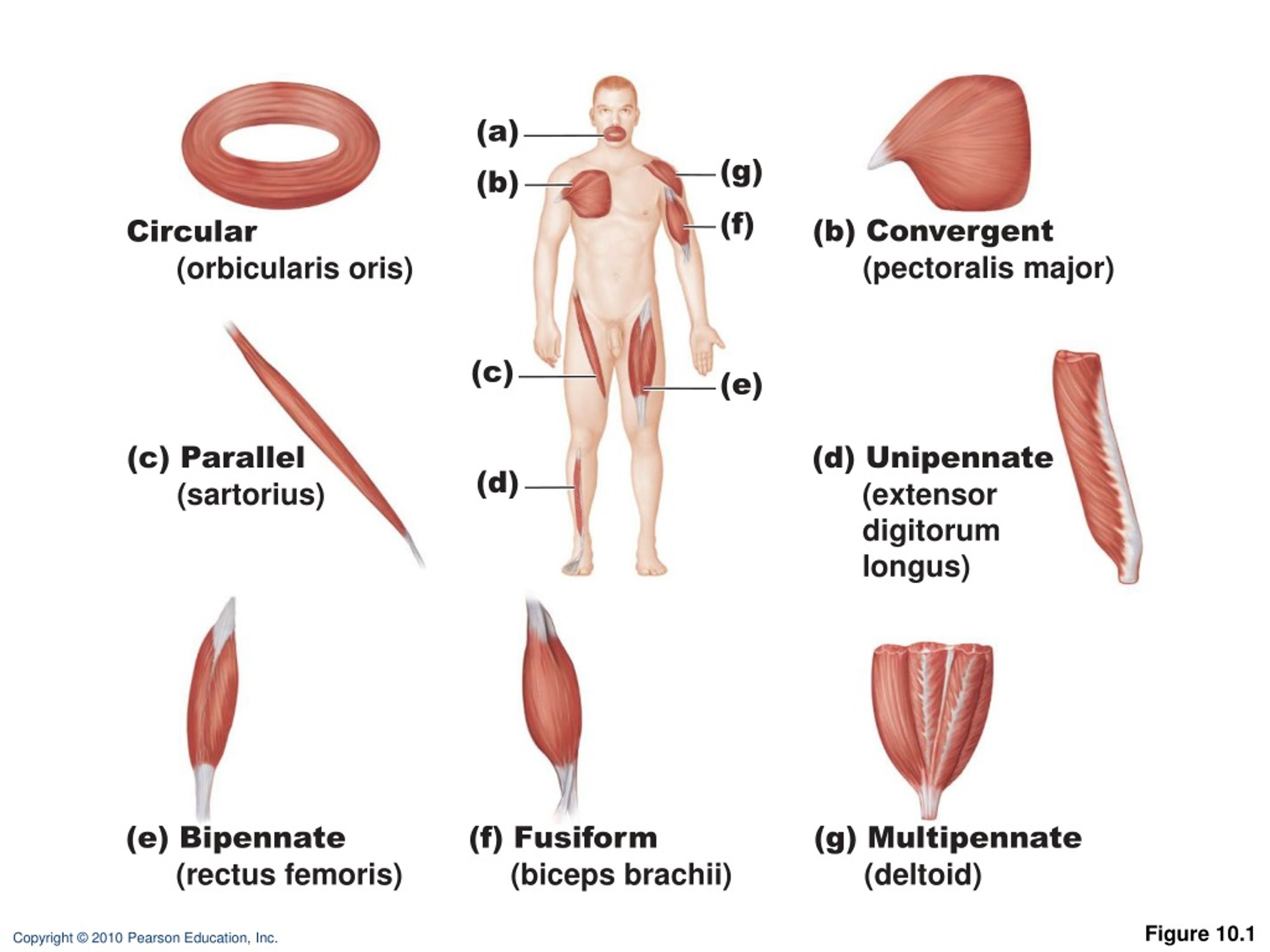
Goh Q, Millay DP. Необходимость опосредованного миомакером слияния стволовых клеток для гипертрофии скелетных мышц. Элиф. 2017; 6: e20007.
Артикул PubMed PubMed Central Google Scholar
Egner IM, Bruusgaard JC, Gundersen K. Истощение сателлитных клеток предотвращает гипертрофию волокон в скелетных мышцах. Разработка. 2016. 143 (16): 2898–906.
CAS Статья PubMed Google Scholar
Gonyea W, Ericson G, Bonde-Petersen F. Расщепление волокон скелетных мышц, вызванное упражнениями с поднятием тяжестей у кошек. Acta Physiol Scand. 1977; 99 (1): 105–9.
CAS Статья PubMed Google Scholar
Gonyea WJ, Sale DG, Gonyea FB, Mikesky A. Увеличение количества мышечных волокон в результате физических упражнений. Eur J Appl Physiol Occupat Physiol. 1986. 55 (2): 137–41.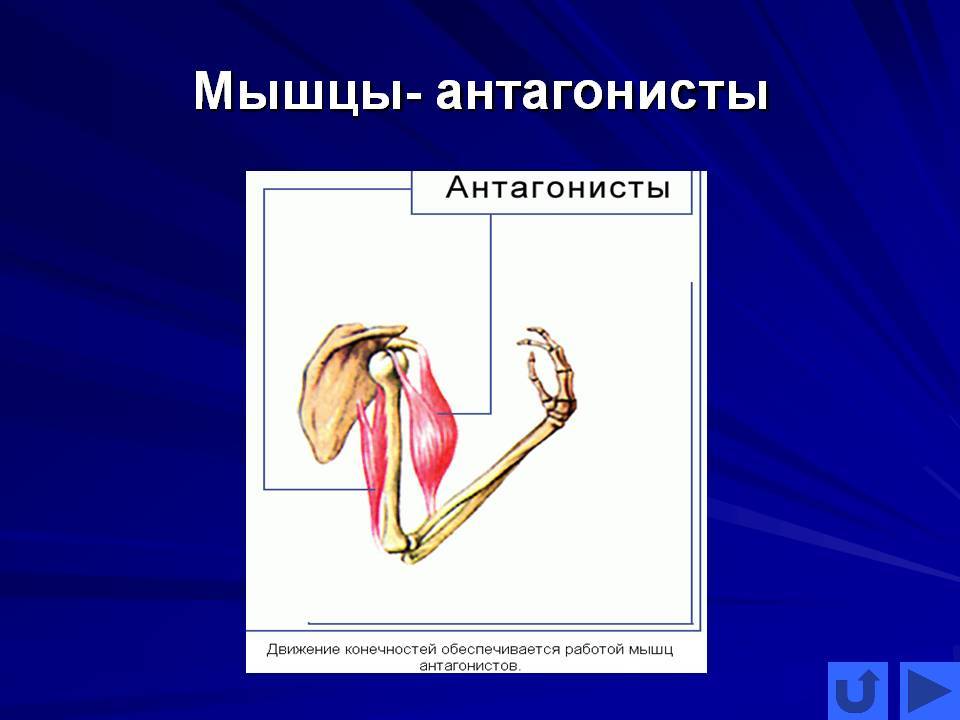
CAS Статья Google Scholar
Холл-Крэггс Э. Продольное деление волокон в перегруженных скелетных мышцах крыс. J Anat. 1970; 107 (Часть 3): 459.
CAS PubMed PubMed Central Google Scholar
Hall-Craggs E, Lawrence CA. Продольное деление волокна в скелетных мышцах: свето- и электронно-микроскопическое исследование. Zeits für Zellfor und Mikroskop Anat. 1970. 109 (4): 481–94.
CAS Статья Google Scholar
Хо К.В., Рой Р., Твидл С., Хойснер В., Ван Хасс В., Кэрроу Р. Расщепление волокон скелетных мышц при выполнении упражнений по поднятию тяжестей на крысах. Am J Anat. 1980. 157 (4): 433–40.
CAS Статья PubMed Google Scholar
Келли Г. Механическая перегрузка и гиперплазия волокон скелетных мышц: метаанализ. J Appl Physiol. 1996. 81 (4): 1584–8.
J Appl Physiol. 1996. 81 (4): 1584–8.
CAS PubMed Google Scholar
Tamaki T, Akatsuka A, Tokunaga M, Uchiyama S, Shiraishi T. Характеристики компенсаторной гипертрофированной мышцы у крысы: I. Электронно-микроскопические и иммуногистохимические исследования. Анат Рек. 1996. 246 (3): 325–34.
CAS Статья PubMed Google Scholar
Vaughan HS, Goldspink G. Число волокон и размер волокон в мышце, подвергшейся хирургической перегрузке. J Anat. 1979; 129: 293.
CAS PubMed PubMed Central Google Scholar
Edgerton VR. Морфология и гистохимия камбаловидной мышцы нормальных и тренированных крыс. Am J Anat. 1970; 127 (1): 81–7.
CAS Статья PubMed Google Scholar
Soffe Z, Radley-Crabb HG, McMahon C, Grounds MD, Shavlakadze T. Влияние произвольных колесных упражнений с нагрузкой на производительность и мышечную гипертрофию у молодых и старых самцов мышей C57Bl / 6J. Scand J Med Sci Sports. 2016; 26 (2): 172–88.
Влияние произвольных колесных упражнений с нагрузкой на производительность и мышечную гипертрофию у молодых и старых самцов мышей C57Bl / 6J. Scand J Med Sci Sports. 2016; 26 (2): 172–88.
CAS Статья PubMed Google Scholar
Egner IM, Bruusgaard JC, Gundersen K. Очевидное отсутствие эффекта истощения сателлитных клеток на гипертрофию могло быть связано с методологическими ограничениями.Ответ на «Методологические вопросы ограничивают интерпретацию негативных эффектов истощения сателлитных клеток на гипертрофию мышц у взрослых». Разработка. 2017; 144 (8): 1365–7.
Артикул PubMed Google Scholar
Эрикссон А., Линдстрём М., Карлссон Л., Торнелл Л. Е.. Гипертрофические мышечные волокна с трещинами у пауэрлифтеров; расщепление волокна или регенерация дефекта? Histochem Cell Biol. 2006. 126 (4): 409–17.
CAS Статья PubMed Google Scholar

Eriksson A, Kadi F, Malm C, Thornell L.E. Морфология скелетных мышц у пауэрлифтеров с анаболическими стероидами и без них. Histo Cell Biol. 2006. 126 (4): 409–17.
CAS Статья Google Scholar
MacDougall J, Sale D, Elder G, Sutton J. Ультраструктурные характеристики мышц у элитных пауэрлифтеров и бодибилдеров. Eur J Appl Physiol Occupat Physiol. 1982. 48 (1): 117–26.
CAS Статья Google Scholar
Кади Ф, Эрикссон А., Холмнер С., Торнелл Л. Е.. Влияние анаболических стероидов на мышечные клетки силовых спортсменов. Med Sci Sport Exerc. 1999. 31 (11): 1528–34.
CAS Статья Google Scholar
Ю. Дж. Г., Боннеруд П., Эрикссон А., Стол П. С., Тегнер Ю., Мальм К. Влияние длительного приема анаболических стероидов андрогенов на скелетные мышцы человека. PLoS ONE. 2014; 9 (9): e105330.
Артикул PubMed PubMed Central Google Scholar
Swash M, Schwartz MS. Последствия продольного расщепления мышечных волокон при нейрогенных и миопатических расстройствах. J Neurol Neurosurg Psych. 1977; 40 (12): 1152–9.
CAS Статья Google Scholar
Wanagat J, Cao Z, Pathare P, Aiken JM. Мутации с делециями митохондриальной ДНК сочетаются с аномалиями сегментарной системы транспорта электронов, атрофией мышечных волокон, расщеплением волокон и окислительным повреждением при саркопении.FASEB J. 2001; 15 (2): 322–32.
CAS Статья PubMed Google Scholar
Голлник П., Тимсон Б., Мур Р., Риди М. Мышечное увеличение и количество волокон в скелетных мышцах крыс. J Appl Physiol. 1981; 50 (5): 936–43.
CAS PubMed Google Scholar

Максвелл Л., Фолкнер Дж., Хаятт Г. Оценка количества волокон в скелетных мышцах морской свинки.J Appl Physiol. 1974. 37 (2): 259–64.
CAS PubMed Google Scholar
Приседания со спиной или приседания со спиной — что выбрать?
Брайан Саттон MS, Массачусетс, NASM-CPT, CES, PES
Что лучше: приседания со штангой или сзади? Ответ на этот непрекращающийся спор во многом зависит от клиента.
Как передние, так и задние приседания требуют сгибания бедра, сгибания колена и тыльного сгибания во время эксцентрической фазы и разгибания бедра, разгибания колена и подошвенного сгибания во время концентрической фазы.Таким образом, как приседания со штангой, так и приседания со спиной задействуют одни и те же группы мышц; основные двигатели включают ягодичные мышцы и квадрицепсы; синергисты включают подколенные сухожилия; и стабилизаторы включают глубокие мышцы живота (поперечный живот).
ЭМГ-активность в каждой группе мышц (квадрицепсы, подколенные сухожилия, ягодичные мышцы) между двумя подъемами практически идентична (1). Из-за естественного изменения положения центра тяжести штанги между приседаниями со штангой спереди и сзади приседания со спиной приводят к снижению активности мышц-разгибателей спины (erector spinae) (1).Приседания со спиной также создают большие сжимающие (нисходящие) силы в коленях. Однако поперечные силы (из стороны в сторону) в колене одинаковы в обоих упражнениях (очень минимальные) (1).
Из-за естественного изменения положения центра тяжести штанги между приседаниями со штангой спереди и сзади приседания со спиной приводят к снижению активности мышц-разгибателей спины (erector spinae) (1).Приседания со спиной также создают большие сжимающие (нисходящие) силы в коленях. Однако поперечные силы (из стороны в сторону) в колене одинаковы в обоих упражнениях (очень минимальные) (1).
Основное различие между двумя упражнениями — это механика верхней части тела. Людям, у которых отсутствует внешнее вращение плеча, могут возникнуть трудности при выполнении приседаний на спине. Этим людям сложно поставить штангу и удерживать ее в правильном положении в верхней части спины из-за плохой подвижности плеч и грудного отдела позвоночника.Эти люди обычно демонстрируют синдром верхнего скрещивания (округленные плечи и голова вперед), обычно наблюдаемый у тех, кто много времени проводит за рулем или работает за компьютером.
Приседания спереди не требуют внешней ротации в плечевом суставе. Однако некоторые люди находят положение штанги на плечах неудобным; особенно если у них дисфункция акромиально-ключичного сустава (АК).
Однако некоторые люди находят положение штанги на плечах неудобным; особенно если у них дисфункция акромиально-ключичного сустава (АК).
Существуют неофициальные данные о том, что приседания со штангой на груди безопаснее для поясницы клиента, но это не было окончательно доказано исследованиями.И наоборот, клиенты, как правило, могут поднимать более тяжелые грузы с помощью приседаний.
Суть в том, что оба упражнения помогают воздействовать на мускулатуру нижних конечностей и могут быть полезны для развития мускулатуры, силы и мощности. Выбранное вами упражнение должно зависеть от позы / техники и предпочтений клиента. Сомнительно, что вы заметите значительную работоспособность, если предпочтете одно упражнение другому. Помните, что все тренировки кумулятивны.
Артикул:
- Гуллетт Дж. К., Тиллман Мэриленд, Гутьеррес Дж. М., Чоу Дж. У.Биомеханическое сравнение приседаний со спиной и спереди у здоровых тренированных людей. Журнал исследований силы и кондиционирования.