
тренировка мышц влагалища и лучшие тренажеры для вумбилдинга
Продолжаем разбираться в очень актуальной «женской теме» — имбилдинг и все, что с ним связано. Сегодня Наталия Музыка, наш эксперт, рассказывает о лучших тренажерах для этого особого вида спорта!
Наталия Музыка, сертифицированным тренером по имбилдингу и руководителем студии женского развития «PION»
Одна из главных женских причин заняться интимной гимнастикой — это укрепление собственного здоровья. Что дает нам интимная гимнастика?
Имбилдинг (на регулярной основе) улучшает кровообращение в органах малого таза — клетки снабжаются кислородом, обновляются, усиливается перистальтика (что решает проблему запоров), происходит избавление от застойных явлений и их нежелательных последствий, таких как эрозии, опущения стенок влагалища и матки, недержание мочи, доброкачественные образования, геморрой.
И даже если у женщины всё в порядке со здоровьем, интимной гимнастикой можно заниматься просто так, «для себя» — и получать хорошие результаты.
О пользе интимной гимнастики, упражнениях для начинающих мы говорили, сегодня же разберем тему: тренажеры для интимной гимнастики. Какие выбрать и какие из них – лучшие?
Имбилдинг: какой тренажер выбрать?
Если зайти в Google и написать в поисковике «тренажер для интимных мышц», от количества гаджетов можно просто потеряться! Однако при более детальном изучении понимаешь, что по дизайну и функционалу они – практически одинаковые, только названия – разные. Чтобы помочь вам подобрать идеальный тренажер для имбилдинга, я расскажу об основных группах тренажеров, которые существуют, об их плюсах и минусах.
Тренажер PelvicToner
Представляет собой две пластмассовых пластины, соединённые пружиной, которую женщина должна сжимать вагинальными мышцами.
- К плюсам PelvicToner можно отнести нагрузку в виде пружин и простоту в применении.
- Минусы: отсутствие обратной связи, угловатый дизайн, не очень приятный в ощущениях, многие женщины боятся, что он может защемить стенки влагалища или шейку матки, очень тугие пружины (практически ни одна женщина не может их сжать).
- Эффективность: если у вас получается сжать пружину, то через месяц тренировок будут первые результаты. А если нет, можно заниматься год, и не заметить никакого результата.
Тренажер Муранивского
Оригинальный авторский тренажер для вумбилдинга создан в 1978 году.
Тренажер Муранивского состоит из двух камер (внутренней — латексной и внешней) и манометра, который дает обратную связь.
- К плюсам этого тренажера, я могу отнести, что у него есть обратная связь, и этот тренажер дает возможность приобретения навыка полного вагинального сжатия (применяется в сексе), но этому обучаются далеко не на первом занятии.

- Минусы: он не предназначен для начинающих, так как приступая к занятиям на таком тренажере без предварительно укрепленных мышц тазового дна женщина может получить ряд осложнений и нанести вред своему интимному здоровью (опущение органов малого таза, недержание мочи и геморрой). Дело в том, что занятия с тренажером Муранивского неизбежно сопровождаются повышенным внутрибрюшным давлением, которое не только сжимает влагалище, но и давит на тазовое дно. И если мышцы тазового дна слабые, то вместо их укрепления можно получить обратный эффект.
- Эффективность: очень сомнительна, если рядом нет инструктора, который будет контролировать правильность выполнения упражнений.

Тренажер Vagiton PNEUMO c насадкой Кегеля
- Плюсом Vagiton PNEUMO является то, что это тренажер 2 в 1. У него есть две насадки: Классическая «Кегеля» и насадка «Муранивского». Есть манометр с биологически обратной связью. Сама насадка Кегеля — это идеальная «гантелька». Есть возможность накачать датчик до определенной упругости и постепенно усложнять работу мышцам, по мере их укрепления. С помощью насадки Муранивского можно делать гинекологический массаж вагины, что очень полезно для интимного здоровья женщины.
- Минусы у этого тренажера такие же, как и у предыдущего, без квалифицированной помощи тренера, можно навредить себе и получить обратный от ожидаемого эффект.
- Эффективность: тренировка мышц влагалища с Vagiton PNEUMO дает ощутимые результаты уже через 2-3 недели после начала тренировок.
 Есть возможность накачать датчик до определенной упругости и постепенно усложнять работу мышцам, по мере их укрепления. С помощью насадки Муранивского можно делать гинекологический массаж вагины, что очень полезно для интимного здоровья женщины.
Есть возможность накачать датчик до определенной упругости и постепенно усложнять работу мышцам, по мере их укрепления. С помощью насадки Муранивского можно делать гинекологический массаж вагины, что очень полезно для интимного здоровья женщины.Тренажер «Лазерный» Юрия Корнева
Тренажер для вумбилдинга представляет собой стержень, на верхнем конце которого установлен шар большего или меньшего размера, а снизу – лазер.
- К плюсам можно отнести уникальность тренажера: на сегодня он не имеет аналогов в мире! Это красивый дизайн и простота в использовании, биологическая обратную связь (в виде лазерной точки), которая дает возможность женщине оценивать правильность выполнения упражнений имбилдинга и соответственно повышает уровень личной мотивации в достижении определенного результата.
- Минусы этого тренажера, в том, что один стержень, с большой вероятностью будет пылиться в коробочке за его ненадобностью. К тренажеру требуется инструктор, который поможет понять, как правильно выполнять упражнения для мышц влагалища, чтобы двигаться в верном направлении и получить намеченный результат.
- Эффективность работы — высокая, первые результаты можно уже почувствовать после 2-3 недель регулярного использования по 10-15 мин в день.

Тренажер Magic Kegel Master
«Magic Kegel» — силиконовое устройство в виде объёмной восьмёрки с вибрирующим эффектом. Оснащен специальным приложением для смартфона и наличием датчика давления.
- К плюсам отнесу современный дизайн, привлекательный внешний вид и приложение для мобильного телефона с обратной связью.
- Минусы: Создатели тренажера объединили в нем несовместимые вещи: вагинальные шарики и устройство для контроля силы сжатия мышц. Датчик считывания мышечных сокращений находится в самом узком месте этого тренажера, поэтому практически ни одна женщина не может достичь требуемых показателей. Тренажер для имбилдинга часто теряет связь с приложением, или вообще не подключается к устройству, то зависает, то выдает длинные инструкции, отвлекающие от занятий, дает ложные результаты тренировки. Именно поэтому для многих Magic Kegel остается красивой, но бесполезной игрушкой.
- Эффективность: нулевая, так как нет четкого понимания, как сделать так, чтобы датчик давления сработал правильно.
 Датчик считывания мышечных сокращений находится в самом узком месте этого тренажера, поэтому практически ни одна женщина не может достичь требуемых показателей. Тренажер для имбилдинга часто теряет связь с приложением, или вообще не подключается к устройству, то зависает, то выдает длинные инструкции, отвлекающие от занятий, дает ложные результаты тренировки. Именно поэтому для многих Magic Kegel остается красивой, но бесполезной игрушкой.
Датчик считывания мышечных сокращений находится в самом узком месте этого тренажера, поэтому практически ни одна женщина не может достичь требуемых показателей. Тренажер для имбилдинга часто теряет связь с приложением, или вообще не подключается к устройству, то зависает, то выдает длинные инструкции, отвлекающие от занятий, дает ложные результаты тренировки. Именно поэтому для многих Magic Kegel остается красивой, но бесполезной игрушкой.Тренажер KGoal (Кейгол)
Тренажер для мышц влагалища представляет собой силиконовую «подушку» для ввода во влагалище. Она отслеживает интенсивность сжатия и расслабления мышц и дает обратную связь в виде вибрации.
- Плюсы этого тренажера, такие же как и у предыдущего тренажера современный дизайн, привлекательный внешний вид и приложение для мобильного телефона с обратной связью.
- Минусы: занятия с тренажером требуют постоянного участия: нужно контролировать силу сжатия мышц и нельзя отвлекаться. Для того, чтобы это было не так изнурительно, производитель включил в приложение ряд игр с помощью которых происходит тренировка.

К сожалению, зависимость тренировок от смартфона имеет свои проблемы: соединение постоянно сбрасывается, а приложение зависает и не работает. Приложение часто показывает ложные результаты.
Еще она проблема, с которой вы можете столкнуться – не соответствие размера тренажера и размеров влагалища. Kgoal имеет крупный размер и, несмотря на то, что его «подушка» сдувается и накачивается, он подходит далеко не всем.
- Эффективность: сомнительна.
Тренажер-миостимулятор Elise
Миостимуляция (воздействие низкочастотным током) мышц дошла и до самых потаенных уголков нашего тела. Вагинальный зонд этого тренажера вводится во влагалище и стимулирует мышцы тазового дна с помощью тока.
- Плюсы: как и любая миостимуляция, это тренировка для ленивых, вам совершенно не надо работать мышцами. А это большой плюс для тех, кто по каким-то причинам потерял контроль над мышцами тазового дна, будучи не в состоянии держать их в тонусе.
- Минусы: воздействие тока на внутренние мышцы – не самое приятное ощущение. К тому же, до конца не изучен вопрос влияния низкочастотных импульсов тока на организм. Под воздействием миостимулятора мышцы начинают сокращаться и работать, о чем становится известно мозгу. А ведь мозг такой команды мышцам не давал! И мозг начинает искать причину неповиновения мышц — начинает перенастраивать свою систему управления! Из-за этого могут появиться такие побочные явления как нарушение работы мышц и спазмы.
- Эффективность: примерно за 20 сеансов можно придать мышцам хороший тонус, но, к сожалению, эффект быстро проходит, как только вы перестаете им пользоваться.
Что же выбрать?
Прежде всего, я за грамотный подход к делу. Я не рекомендую подбирать тренажер самостоятельно. Доверьте это дело профессионалу, ведь на кону ваше здоровье. То же самое касается тренировок: лучше не делать этого без инструктора. Практика показывает, что большинство женщин, начав практиковать имбилдинг самостоятельно, приносят себе только вред. Причина? Напрягают совершенно «не те» мышцы или тужатся, а это чревато опущением внутренних органов, матки, геморроем.
Я не рекомендую подбирать тренажер самостоятельно. Доверьте это дело профессионалу, ведь на кону ваше здоровье. То же самое касается тренировок: лучше не делать этого без инструктора. Практика показывает, что большинство женщин, начав практиковать имбилдинг самостоятельно, приносят себе только вред. Причина? Напрягают совершенно «не те» мышцы или тужатся, а это чревато опущением внутренних органов, матки, геморроем.
Но если вы все-таки решили сэкономить на самом дорогом, вот несколько рекомендаций:
- тренажер должен быть изготовлен из медицинского материала и быть гипоаллергенным;
- в комплекте с ним должна идти подробная инструкция;
- вам должно быть комфортно, когда вы помещаете его внутрь;
- и главное: не старайтесь сжимать как можно сильнее то, что помещаете внутрь, сосредоточьтесь на правильности выполнения (старайтесь напрягать «именно те мышцы»).
Перед началом тренировок, обязательно проконсультируйтесь со своим гинекологом на предмет выявления противопоказаний!
Будьте здоровы и тренируйтесь с умом!
Тренажер для развития интимных мышц Тренажер для развития вагинальных мышц Тренажер Муранивского Тренажер Кегеля
Регулярные тренировки с тренажером для развития интимных мышц, показали улучшение состояния 86% женщин, имеющих проблемы, вызванные первоначальным ослаблением мышц малого таза.
Тренажер для развития вагинальных мышц – правильный выбор! В данном случае это выбор регулярных тренировок мышц тазового дна, с тем, чтобы ослабить симптомы или вовсе избавиться от недержания, а также улучшить «вагинальные объятия», вместо согласия на более сложные процедуры вплоть до хирургического вмешательства.
Тренажер Муранивского, является совершенно новым в мировой практике многофункциональным тренажером индивидуального пользования. Наличие видимых показаний манометра значительно повышает эффективность тренировок, позволяет наглядно видеть и оценивать состояние вагинальных мышц и результаты их развития. В основе техники работы с Тренажером Муранивского лежит принцип обратной связи, который очень эффективно используется для обучения спортсменов и космонавтов, в лечении для восстановления двигательных навыков и координации движений после травм. Задача упражнений с Тренажером Муранивского — научить мозг вырабатывать импульс на сокращение соответствующих мышц. Когда женщина делает хоть малейшее правильное мышечное усилие, то с помощью манометра она видит этот момент и естественным образом фиксирует его подсознательно. Таким образом, женщина очень быстро научится правильно работать интимными и связными мышцами.
Когда женщина делает хоть малейшее правильное мышечное усилие, то с помощью манометра она видит этот момент и естественным образом фиксирует его подсознательно. Таким образом, женщина очень быстро научится правильно работать интимными и связными мышцами.
В конце 40-х годов 19 века, гинеколог по имени Арнольд Кегель разработал программу упражнений по развитию мышц промежности, предназначенной для лечения женщин от недержания мочи. В наше время эта методика известна как «Упражнения Кегеля» и рекомендуются женщинам и мужчинам для развития мышц тазового дна.
Таким образом , применяя тренажер Кегеля вырабатывается умение сжимать только вагинальный сфинктер (мышцы входа) очень далеко от необходимой совокупной работы всех вагинальных и сопряженных мышц.
Для полноценного выполнения всех необходимых мышечных действий (секс, роды, поддержание органов, выделительные функции и т.д. необходима тренировка всей совокупности вагинальных мышц, а не отдельно взятой, например, как это происходит при чистом исполнении упражнения Кегеля)
Читать «Развитие забытых мышц» — Муранивский Владимир Леонидович — Страница 6
Представляет собой шарик, к которому прикреплен прочный шнур, на конец которого подвешивается груз.
Примечание: ТГр можно изготовить самостоятельно (см. на сайте)
Процесс тренировки заключается в том, что шарик вводится в вагину так, чтобы он находился в зоне тренируемых мышц. Сжатием этих мышц пытаются удержать груз (бутылка с водой) в положении стоя или при ходьбе.
Если этого сделать не удается — груз выскальзывает или опускается до мышц входа, значит в бутылке слишком много воды.
По мере укрепления, развития мышц увеличивается вес груза и уменьшается размер шарика.
Нормально развитые мышцы способны удерживать определенный вес в течение определенного промежутка времени.
Удержание груза на любой глубине вагины свидетельствует о гармоничном развитии мышц.
В зависимости от результатов работы с ТГр женщина принимает решение о необходимости приступить к дальнейшим тренировкам своих мышц.
На это же решение влияют ее коитальные успехи, популярность у партнера. Под этим понимается ее способность работать (сжать — расслабить) мышцами и ощущать эту работу.
При однократном сильном сжатии женщина ощущает, что вагинальные стенки сомкнулись, что она сможет удержать даже такой тонкий предмет, как спица (не вздумайте вводить спицу).
При многократных сжатиях ощущается появление тепла и удовольствия, вплоть до оргазма — приливает кровь к нагруженным мышцам и нервным окончаниям.
!!! Основное внимание нужно уделить развитию не интроитальных (на входе), а вагинальных, расположенных внутри, особенно ближе к сводам, матке. Мышечные сжатия должны быть достаточно сильными[3] и производить неизгладимое впечатление на партнера.
Глава 6–2. Тренажер пневматический (ТП)
ТП является базовым тренажером и предназначен для развития любых групп интимных мышц по пунктам 1–5 (см. гл. 6).
ТП состоит из:
1. внутренняя камера (вводится во влагалище)
2. манометр (для визуального контроля создаваемого вагинального давления)
3. нагнетатель с клапаном (для создания начального давления в тренажере)
4. внешняя камера (для быстрого изменения давления в рабочей камере)
внешняя камера (для быстрого изменения давления в рабочей камере)
Процесс тренировки (одно из упражнений) заключается в том, что рабочая камера вводится в вагину. Насосом подкачивается наружная камера. Сжимая ее, перегоняют воздух в рабочую камеру и она раздувается, воздействуя на нужные мышцы. (Подробнее о работе с ТП смотрите в инструкции, в видеолекции и в разделе «Упражнения с ТП» на сайте).
Для увеличения эффективности тренировок рекомендуется совместная тренировка брюшных и вагинальных мышц. Одевают плотные обтягивающие брюки, джинсы и закладывают между животом и брюками наружную камеру (живот втянут).
Выпячивая (наружная камера сжимается, рабочая раздувается) и втягивая брюшные мышцы живота, воздействуют на наружную камеру, а последняя на мышцы.
Однако, если брюшные мышцы развиты плохо и не поддаются управлению или совместная работа с вагинальными мышцами не получается, наружную камеру нужно сжимать рукой, ногой, можно привлечь партнера.
Сам процесс тренировки происходит следующим образом.
— Сжимая наружную камеру (рукой или животом на вдохе) сопротивляются вагинальными мышцами раздуванию рабочей камеры. Эффективность сопротивления оценивают по манометру.
— Отпуская наружную камеру (рукой или втягивая живот на выдохе), стараются вагинальными мышцами выдавить весь воздух из рабочей камеры.
Можно при сжатии наружной камеры расслаблять вагинальные мышцы, камера помогает мышцам «раскрыться». После освоения можно надутую наружную камеру положить перед глазами: при расширении мышц — камера «опадает», при сжатии — расширяется.
В процессе тренировок, по мере развития мышечной силы, наступит такое положение, при котором женщина будет испытывать оргазм.
Это не повод для беспокойства. Такое явление возникает также при развитии мышц путем выполнения йоговских упражнений, у некоторых — при сжатии бедер и мышц ягодиц.
Первым критерием достаточности тренировок является возникшее умение мышечными действиями ускорять, предотвращать или вызывать оргазм.
Простым сжатием тех или иных мышц можно вызвать в любое время, в различных ситуациях то или другое состояние.
Научившись управлять мышцами, превратив их в ВУМы, можно научиться управлять своими эмоциями, внешними проявлениями.
Овладев этим, тренировки с ТП можно проводить только для поддержания мышц в таком состоянии, а лучше перейти только на коитальные упражнения. Дальнейшее наращивание мышечной силы нецелесообразно, а в ряде случаев — вредно (напоминаeм: не выше 200мм по шкале ТП).
Другим критерием является способность удерживать ВУМами в течение определенного времени груз определенного веса на определенной глубине (см. описание ТГр).
Напоминание: ВУМы являются значительным фактором риска при полигамном поведении в смысле инфицирования венерическими заболеваниями и ВИЧ. При неумелом использовании ВУМов (а особенно мышц входа — упражнения Кегеля) презерватив либо стянется, либо порвется.
ВУМы — только для моногамии и проверенных партнеров.
ТП применяется по всем пяти пунктам назначения, в том числе девственницами (конструкция рабочей камеры исключает дефлорацию).
При слабых вагинальных мышцах (природные, возрастные изменения, послеродовой период и т. п. — мышцы не работают, не ощущаются, в нормальном состоянии канал вагины не сжат) увеличивается суммарная (общая) упругость камеры.
Научившись испытывать оргазм при тренировках, бывшие фригидные женщины могут научиться испытывать это состояние при коитусе.
Глава 6–3. Тренажер гидравлический (ТГ)
ТГ предназначен для:
— выработки автоматизма сокращения мышц входа и вагинальных мышц
— струйного массажа этих зон и связанных с ними органов
— профилактики заболеваний этих зон
— восстановления и совершенствования сексологических способностей, вплоть до оргазмических состояний при фригидности.
ТГ работает от напора горячего и холодного водоснабжения при наличии гибкого шланга душа. Вместо лейки навинчивается корпус рассеивателя ТГ.
Вместо лейки навинчивается корпус рассеивателя ТГ.
ТГ исключает контакт воды со слизистой вагины.
Тренировки в ванне с водой, нужной температуры, создают определенный комфорт, что позволяет пользователю сосредоточиться на выполняемых действиях физически и психологически. Это также повышает эффективность.
Совмещение силовых тренировок с положительно действующими струйным массажным воздействием предопределяет привлекательность пользования ТГ.
Отрегулировав напор и температуру воды, пользователь вводит камеру в вагину. Вода, заполнив камеру через разбрызгиватель, вытекает в ванну.
Корпус имеет быстродействующий регулятор, для экстренного сброса напора воды.
ТГ позволяет осуществлять различные программы мышечных действий:
1) Сжимаются сфинктерные мышцы (вход), при этом уменьшается кольцевой зазор вокруг разбрызгивателя, создается сопротивление вытекающей из камеры воды. Камера начинает расширяться, воздействуя на внутренние вагинальные мышцы.
Камера начинает расширяться, воздействуя на внутренние вагинальные мышцы.
Расслабляя сфинктерные мышцы, увеличивают зазор для вытекания воды, уменьшают сопротивление. Объем камеры сокращается, нагрузка на внутренние мышцы снижается.
Вумбилдинг, Имбилдинг, Система Вагитон – в чем отличие и что выбрать? — Статьи на сайте Женской Академии Яны Чебатаревой
Про ВумбилдингВ России и СНГ тема интимных мышц получила мощный толчок благодаря трудам В.Л. Муранивского. Он внес очень большой вклад в популяризацию идеи тренировки вагинальных умений.Основной его идеей было то, что необходимо тренировать сжатие всего вагинального канала, а не, как он говорил, «мышц входа», которые тренируются упражнениями Кегеля.
Он полагал, что вагинальные мышцы расположены по всей длине вагины, и что они также поддаются развитию, как и «мышцы входа». Более того, он считал, что эти мышцы расположены кольцами, одно над другим, и что можно научиться управлять каждым кольцом отдельно. Именно так, по его мнению, женщина может делать волну сжатий. Совокупность этих колец он назвал ВУМами (Вагинально Управляемыми Мышцами). Отсюда и пошло название его методики – вумбилдинг. Эта методика и сегодня пропагандируется многими школами Вумбилдинга.
Именно так, по его мнению, женщина может делать волну сжатий. Совокупность этих колец он назвал ВУМами (Вагинально Управляемыми Мышцами). Отсюда и пошло название его методики – вумбилдинг. Эта методика и сегодня пропагандируется многими школами Вумбилдинга.
По мнению В.Л. Муранивского, первое кольцо — это мышца входа, второе кольцо сжимает вагину в районе точки G, третье — еще выше, а четвертое он называл мышцами сводов.
Сам В.Л. Муранивский, впервые, наблюдал такие вагинальные действия у своей возлюбленной, с которой они впоследствии разошлись. И тогда у него родилась мысль обучать этому других женщин. Подкрепление своих убеждений насчет ВУМов он нашел в литературе по древним энергетическим сексуальным практикам, таким как Дао, Тантра, в частности, в книгах Мантека Чиа, где также можно встретить термин «кольца». В подобных трактатах содержатся сокровенные знания, но по большей части они описаны в иносказательном и образном виде, через призму энергетики и философии, с очень поверхностным описанием упражнений и методик. В них красочно повествуется о том, что женщине необходимо практиковать тренировки интимных мышц для улучшения физического и духовного здоровья и гармонизации сексуальных отношений. Владимир Леонидович, не углубляясь в анатомию, на основании собственных ощущений и данных из прочитанной даосской литературы, естественно решил, что можно научиться чувствовать и управлять мышцами всего влагалища, а не только «мышцами входа».
В них красочно повествуется о том, что женщине необходимо практиковать тренировки интимных мышц для улучшения физического и духовного здоровья и гармонизации сексуальных отношений. Владимир Леонидович, не углубляясь в анатомию, на основании собственных ощущений и данных из прочитанной даосской литературы, естественно решил, что можно научиться чувствовать и управлять мышцами всего влагалища, а не только «мышцами входа».
Таким образом, В. Муранивский, на базе известного тренажера Кегеля, изобрел и запатентовал тренажер пневматический (ТП), вагинальная камера которого представляет собой эластичный баллончик (на тот момент из презервативов), который вставляется до самого конца вагины, за «мышцы входа» и должен показывать сжатие ВУМов.
ТП и правда очень хорошо показывает любое изменение вагинального объема, и женщина действительно может сжимать всю вагину, вопрос состоит только в том — за счет чего именно, за счет каких усилий и почему это может быть опасно для здоровья женщины?
Согласно анатомическим данным, которые можно почерпнуть в любом анатомическом источнике, мышцы делятся на два типа:
— поперечно-полосатые (управляемые)
К ним относятся все скелетные мышцы, включая дыхательную диафрагму и тазовые мышцы. Главной особенностью поперечно-полосатых мышц является то, что они способны ПРОИЗВОЛЬНО, по желанию человека, СОКРАЩАТЬСЯ и РАССЛАБЛЯТЬСЯ. Посредством тренировки человек может придать им необходимую силу и увеличить их объем.
Главной особенностью поперечно-полосатых мышц является то, что они способны ПРОИЗВОЛЬНО, по желанию человека, СОКРАЩАТЬСЯ и РАССЛАБЛЯТЬСЯ. Посредством тренировки человек может придать им необходимую силу и увеличить их объем.
— гладкие (неуправляемые).
К ним относятся мышцы внутренних органов, например, матки, вагины, мочевого пузыря, кишечника, стенок сосудов и т.д. Главной особенностью гладких мышц, является то, что человек не может ими сознательно работать, сокращать и расслаблять. Эти мышцы сокращаются рефлекторно, непроизвольно, независимо от желания человека.
Вагинальный канал — это тоже гладко-мышечная трубка, не управляемая волевым приказом, также как и мышцы других внутренних органов.
Вы не можете почувствовать и научиться управлять гладкомышечными волокнами вагинальной трубки, как и не можете сокращать гладкую мускулатуру каких-либо других внутренних органов.
В результате, основные положения, на которых основывается теория В.Л.Муранивского, не соответствуют анатомическим сведениям, т.е. Вагинальных Управляемых Мышц (ВУМов) нет!!! И мышцы, которые принимают участие в сжатии вагинального канала – это не влагалищные!!!
Мышцы, которые охватывают стенки вагинального канала и при сокращении его сдавливают – называются мышцами тазового дна. Помимо мышц тазового дна, в сжатии и расширении вагины участвуют также мышцы пресса и дыхательная диафрагма. МТД, пресс и диафрагма – это скелетные мышцы, которые могут управляться волей нашего сознания.
Способность управлять разными участками мышц живота совместно с диафрагмой и тазовыми мышцами позволяет создавать различные усилия, которые будут поднимать, опускать, либо сжимать внутренние органы. Важно понимать, что брюшная полость в своей массе сравнима с неким замкнутым «пузырем» с жидким содержимым, как, например, полиэтиленовый пакет, наполненный водой. Его возможно деформировать, но, невозможно сжать, т. е. уменьшить объем. Деформация любой части этого «пузыря» вызовет ответную реакцию в другой части «пузыря» за счет перемещения его содержимого.
е. уменьшить объем. Деформация любой части этого «пузыря» вызовет ответную реакцию в другой части «пузыря» за счет перемещения его содержимого.
Когда мы говорим, что дыхательная диафрагма давит вниз и уменьшает объем брюшной полости, мы имеем в виду, что вместо фактического уменьшения объема происходит его деформация сверху, что приводит к смещению брюшных оранов вниз. Если при этом живот и тазовые мышцы напряжены, то смещения органов нет, но происходит ПОВЫШЕНИЕ ВНУТРИБРЮШНОГО ДАВЛЕНИЯ. Повышенное внутрибрюшное давление не только сжимает все брюшные и тазовые органы (вагинальный канал в том числе), но и оказывает давление на мышцы тазового дна. СИЛА ИХ ВТЯГИВАНИЯ В ДАННЫЙ МОМЕНТ ИГРАЕТ РЕШАЮЩУЮ РОЛЬ В ПРЕДОТВРАЩЕНИИ ОПУЩЕНИЯ ОРГАНОВ. Но если тазовые мышцы или живот в этот момент расслаблены или не способны втягиваться, то органы получают возможность смещения вниз и давление растягивает слабую область, т.е. тазовое дно и низ живота.
НЕСПОСОБНОСТЬ ТАЗОВЫХ МЫШЦ И МЫШЦ НИЗА ЖИВОТА СПРАВЛЯТЬСЯ С ПОВЫШЕННЫМ ВНУТРИБРЮШНЫМ ДАВЛЕНИЕМ ЧАСТО ПРИВОДИТ К ОПУЩЕНИЮ ОРГАНОВ.
Большинство школ вумбилдинга с первого же занятие предлагают своим клиенткам использовать пневматический тренажер (ТП). ТП показывает сжатие всего вагинального канала, а это значит, что он показывает повышение внутрибрюшного давления, которое создается при работе МТД, пресса и дыхательной диафрагмы, а не за счет работы вагинальных мышц. Зная, что упражнения с «ТП» сопровождает повышенное внутрибрюшное давление, необходимо учитывать, что помимо сжатия внутренних органов (в том числе и вагины), оно с такой же силой давит и на тазовое дно. А если мышцы тазового дна слабые (что встречается у большинства женщин в независимости от возраста), то они не справятся с этой нагрузкой. Практика показывает, что результатами регулярных тренировок с тренажерами «ТП» при слабых интимных мышцах являются опущение органов малого таза, недержание мочи и геморрой.
Опасность для потенциальных и уже действующих клиенток многих школ вумбилдинга заключается в том, что уже на первом занятии их учат выполнять полное вагинальное сжатие, которое возможно только при повышении ВБД вне зависимости от состояния их тазовых мышц.
Дело в том, что специального тренажера, оценивающего состояние тазовых мышц в школах, работающих строго по В. Муранивскому, нет. Предлагаемое тестирование с помощью металлических грузов не выдерживает никакой критики, то же самое можно сказать и об удержании груза к подвешенному шарику или яйцу.
Дело в том, что прежде чем начинать подключать к работе интимных мышц внутрибрюшное давление, необходимо сначала укрепить мышцы тазового дна. Т.к. внутрибрюшное давление способствует смещению органов вниз!!!
Если МТД слабые (как показывает практика — так и есть у большинства представительниц женского пола) и мы на них начинаем регулярно оказывать давление, вместо успехов в личной жизни мы можем приобрести проблемы со здоровьем — это опущение органов, геморрой и недержание мочи. Только если поддерживающий аппарат предварительно натренирован специальными упражнениями, и способен справиться с повышенным давлением сверху, то органы не будут смещаться вниз, а наоборот, будут получать мощный и полезный массаж.
В. Муранивский обучил не один десяток учениц для преподавания Вумбилдинга по его методике, и сегодня тренажер ТП применяется абсолютно во всех школах Имбилдинга и Вумбилдинга (в большинстве — с самого начала, без предварительной подготовки поддерживающего аппарата).
Про систему ВагитонРуководитель школы Юрий Корнев более чем за 10 лет работы, основываясь на опыте обучения Вумбилдингу, тех проблемах, с которыми он сталкивался, изучив анатомическое строение малого таза, в частности анатомо-физиологические особенности интимных мышц, проконсультировавшись с гинекологами и акушерами, изобрел свою методику тренировки интимных мышц, которая признана и пользуется большой популярностью среди женщин в мире. Эта методика называется – система Вагитон.
Согласно этой методике сначала укрепляются Мышцы Тазового Дна, и только потом осваивается методика полного сжатия вагины, путем создания внутрибрюшного давления с помощью мышц пресса и диафрагмы на пневматическом тренажере.
Юрием Корневым был изобретен и запантетован специальный тренажер, который показывает силу втягивания мышц тазового дна и который позволяет укрепить тот поддерживающий аппарат, о котором говорилось выше. Этот тренажер называется Лазерный.
Лазерный тренажер позволяет отслеживать силу втягивания мышц тазового дна, и только когда тренажер показывает хорошие результаты, разрешается постепенно переходить на полное вагинальное сжатие, используя пневматический тренажер Муранивского. Только при таком подходе, когда мы уверены в том, что мышцы тазового дна крепкие, использование тренажера Муранивского является безопасным и полезным для здоровья.
В основе системы Вагитон — только обоснованные методики и самые эффективные техники, давшие нужный результат и многократно проверенные на практике. Мы рекомендуем только то, что полезно абсолютно всем женщинам, независимо от возраста, комплекции, рода занятий или семейного положения.
Система Вагитон основана на древних учениях в сочетании с современными анатомическими знаниями, а также новейшими технологиями обратной связи и создания нагрузки на интимные мышцы.
СИСТЕМА ВАГИТОН — ЭТО СПЛАВ ДРЕВНИХ ПРАКТИК И СОВРЕМЕННЫХ ТЕХНОЛОГИЙ.
Про имбилдингПосле смерти В.Л. Муранивского образовалось две школы: — Международная Школа Имбилдинга Ю. Корнева — Студия Вумбилдинга им. Муранивского (с участием сына Муранивского, которому по наследству перешли все права). Совсем недавно, в связи с тем, что Муранивский В.В. на основании полученных по наследству прав от Муранивского В.Л. зарегистрировал товарный знак «IMBUILDING», а также в связи с тем, что слово Имбилдинг» стало применяться всеми подряд и перестало иметь различительную способность, Юрием Корневым было принять решение переименовать свою методику, которая получила название «Система Вагитон».
P.S. Милые женщины!!! Каждый раз, когда Вы принимаете решение о том, в какую школу идти учиться и какого инструктора выбрать, пожалуйста, уделите немного внимание, почитайте о том, какая методика используется инструктором, какие мнения существуют по поводу нее. И только поняв отличие, принимайте решение!!!
И только поняв отличие, принимайте решение!!!
Как Вы уже, наверное, поняли, наша Академия работает по методике «Система Вагитон» Юрия Корнева, потому что эта система безопасная, эффективная и проверена временем!!!
© Copyright:Чебатарева Яна, 2014
Пневматический тренажер для интимных мышц
1. Пневматический тренажер для интимных мышц
12. Тренажер для интимных мышц разработан в Великобритании, производится на заводе, выпускающем медицинскую технику, и успешно продается в СШ
Пневматический тренажер для интимныхмышц — это уникальный тренажер с биологической
обратной связью.
Биологическая обратная связь позволяет объективно
оценивать правильность выполнения упражнений и
результат тренировок.
Принцип действия тренажера:
в вагинальный датчик с помощью пульта
накачивается воздух. Когда женщина сжимает
мышцы тазового дна часть воздуха по трубке
поступает в пульт управления, который фиксирует и
отображает силу сжатия на экране.

Тренажер для интимных мышц разработан в Великобритании, производится на
заводе, выпускающем медицинскую технику, и успешно продается в США, Канаде,
Великобритании, Франции, ЮАР, Дании и др. странах.
2
3. В комплект тренажера входят: пульт управления с цветным экраном, отображающим секундомер, силу сжатия, степень наполнения вагинального да
В комплект тренажера входят:пульт управления с цветным экраном, отображающим секундомер,
силу сжатия, степень наполнения вагинального датчика воздухом.
вагинальный датчик, покрытый 100% медицинским силиконом
иллюстрированная инструкция на русском языке
кейс для хранения
3
4. Преимущества тренажера
•Биологическая обратная связь•Оснащен информационным дисплеем
•Голосовой гид на английском языке
•Покрытие датчика- 100% медицинский силикон
•Надежное крепление датчика к пульту управления предупреждает его нежелательное
отсоединение
•Уникальная конструкция датчика позволяет выполнять упражнения Кегеля в любом
положении тела
• 2 режима работы: «Тест» и «Тренинг».
 Режим «Тест» помогает оценить эффективность
Режим «Тест» помогает оценить эффективностьтренировки
•Автоматическое сохранение результатов последней тренировки
•Встроенный секундомер
•Информирование о недостаточном/избыточном количестве воздуха в датчике
•Низкое энергопотребление (работает от 3-х батареек ААА)
•Современный привлекательный дизайн
4
5. Порядок тренировки с тренажером
•Вставьте 3 батарейки ААА в пульт управления•Тщательно промойте теплой мыльной водой вагинальный датчик, ополосните кипяченой
водой комнатной температурой
•Подсоедините датчик к пульту управления
•Введите датчик во влагалище
•Включите пульт управления и выберите необходимый режим
•Далее следуйте голосовому гиду и/или информации, отображающейся на экране
•После окончания тренировки отсоедините датчик и промойте его теплой мыльной водой.
Затем уберите датчик в футляр.
Часто задаваемые вопросы
-Какие есть противопоказания и ограничения использования тренажера?
Первые 6 недель после родов или хирургического вмешательства в области таза, наличие
вагинальной инфекции и/или инфекции мочевыводящих путей.
 Наличие барьерных
Наличие барьерныхконтрацептивов (диафрагма) или пессария.
-Как часто надо заниматься с тренажером?
Мы рекомендуем тренироваться дважды в день: утром и вечером.
-Когда я увижу первые результаты?
Обычно ощутимое улучшение наступает через 6-8 недель регулярных тренировок.
5
Будем рады сотрудничеству с Вами!
ООО «ДжиЭс Медикал»
197101, Санкт-Петербург,
Ул. Дивенская, д.3 лит. Е
8 (812) 332 41 47
6
«Школа гейши» и развитие интимных (вагинальных) мышц.
ОТВЕТ
автора методики ВУМБИЛДИНГ по развитию вагинальных мышц и
изобретателя тренажеров
МУРАНИВСКОГО В.Л.
ОСНОВАТЕЛЮ «ШКОЛЫ (ГЕЙШИ) ИСКУССТВА ВЗАИМООТНОШЕНИЙ»
ФРОЛОВУ О.В. на его «ОБРАЩЕНИЕ»,
размещенное на сайте www.geishi.ru в апреле-мае 2003г.
Текст
«обращения» можно прочитать здесь.
В подтверждение его существования там же представлен
Протокол осмотра сайта «Школы гейши»
московским нотариусом Амелькиной Е.А.
(Примечание: обращения было изменено, а потом и вовсе удалено после публикации данного «ответа»).
В 2002 году основатель «Школы (гейши) искусства взаимоотношений» О.Фролов обратился ко мне с просьбой, чтобы я не публиковал компрометирующие материалы о его деятельности. Я выполнил эту просьбу, так как ценю вклад основателя «Школы (гейши) искусства взаимоотношений» в популяризацию развития вагинальных управляемых мышц (ВУМбилдинга) в процессе становления «Школы гейши».
Я и сейчас не буду
раскрывать мелочную и грязную подоплеку моего разрыва с основателем «Школы гейши». В подтверждение
привожу несколько цитат из печатных
изданий, которые подтверждают мою
преподавательскую деятельность в «Школе
гейши», и которые можно найти в библиотеках,
архивах и проверить:
В подтверждение
привожу несколько цитат из печатных
изданий, которые подтверждают мою
преподавательскую деятельность в «Школе
гейши», и которые можно найти в библиотеках,
архивах и проверить:
Разве не Вы, г-н Фролов, давали такую рекламу в «Ваш ДОСУГ» (№2, 2000г., стр.103): «Школа гейши» делится тайными знаниями. Одно из них — развитие интимных мышц. У нас в школе работает единственный специалист в России в этой области Муранивский В.Л.» ?
А вот что написано в специальном репортаже Елены Ольшанской «Первый раз в первый класс школы гейши» («Marie Claire», октябрь, 2000г., стр.71) «…Владимир Муранивский, тренер и изобретатель эксклюзивного тренажера, которому нет аналогов в мире, является также автором нескольких книг по управлению влагалищными мышцами…».
В статье Оксаны Семеновой «Слаба на передок
— качайся» («Московский комсомолец» №265 за 25. 11.2000г) говорится: «…Единственное, что показалось нам достойным внимания в школе гейш, — тренажер
для развития интимных мышц. Изобретатель чудо-аппарата Владимир Леонидович Муранивский
разработал целую программу занятий по … интимбилдингу. … Тренажер для развития
интимных мышц — вещь, которая дает результаты. … Все остальное в так называемой
школе гейш — это натуральный развод на деньги…».
11.2000г) говорится: «…Единственное, что показалось нам достойным внимания в школе гейш, — тренажер
для развития интимных мышц. Изобретатель чудо-аппарата Владимир Леонидович Муранивский
разработал целую программу занятий по … интимбилдингу. … Тренажер для развития
интимных мышц — вещь, которая дает результаты. … Все остальное в так называемой
школе гейш — это натуральный развод на деньги…».
«SUPER» №9 (2002г.), стр.5, «Царское дело». Оксана Расторопная качает интимные мышцы. «…После теории началось самое интересное — практические занятия на тренажере для развития интимных мышц, … начал свой рассказ главный мэтр интимных дел в школе гейш…Владимир Леонидович…».
Основатель «Школы (гейши) искусства взаимоотношений» достаточно широко
рекламировал в глянцевых журналах и на телевидении не только пользу развития
вагинальных управляемых мышц (ВУМбилдинга), но и предложенную мною методику, изобретенные и запатентованные мною тренажеры.
Польза рекламы «Школы (гейши) искусства взаимоотношений» перевешивала мой ущерб от попрания моих авторских прав. И вдруг появление «ОБРАЩЕНИЯ».
В нем О. Фролов не только нарушает мои авторские права, но и оскорбляет мою честь, достоинство и деловую репутацию.
Видимо моя просветительская и преподавательская деятельность в интернете в чем-то затронула какие-то интересы основателя «Школы (гейши) искусства взаимоотношений». Мне остается только посочувствовать ему, так как, вдобавок, г-н Фролов О.В. вынужден был сменить «раскрученное» и уже известное название «Школа гейши» (сколько денег стоила эта «раскрутка» !?) на никому неизвестную «Школу искусства взаимоотношений» (может быть в этом также замешано что-то неблаговидное?).
Серьезные голословные обвинения
(«шарлатанство») в мой адрес заставляют меня отбросить скромность и отстаивать
свой вклад в развитие вагинальных управляемых мышц (ВУМбилдинг).
Итак, разберу «Обращение» О. Фролова, основателя «Школы гейши», цитаты из которого выделены курсивом:
«Но, вероятно, наша популярность не даёт кому-то покоя и ряд сограждан, судя по их необузданной одержимости, решили вести подобный предмет».
Г-н Фролов О.В., Вам прекрасно известно, что заниматься развитием вагинальных управляемых мышц (ВУМбилдингом) я начал еще до Вашего появления на Свет (в буквальном смысле).
Но только с 1989
года стала возможной популяризация этих знаний. Доказательством этого являются пять патентов на изобретения по этой теме с приоритетом от 1990-91 годов, два
свидетельства на полезные модели (совершенствование запатентованных ранее тренажеров), четыре книжки («Интимные мышцы» 1993г., «Развитие интимных мышц» 1994г., «Развитие женских мышц» 1996г. , «Счастье и брак» 2002г), две видеокассеты с демонстрацией умений моих учениц (последняя — четырехчасовая
2000г.).
, «Счастье и брак» 2002г), две видеокассеты с демонстрацией умений моих учениц (последняя — четырехчасовая
2000г.).
Смею напомнить, что «Школа гейши» основана почти на десять лет позже патентования мною тренажеров.
Популярность «Школы (гейши) искусства взаимоотношений» никоим образом не влияет на мою деятельность в этом направлении, она меня не тревожит. Мне у Вас учиться нечему. Очень даже хорошо, что появился еще один популяризатор ВУМбилдинга в лице «Школы гейши». Было бы еще лучше, если бы Вы, кроме рекламной деятельности, внесли реальный вклад в популяризацию ВУМбилдинга, хоть что-нибудь дали людям бесплатно.
Моя деятельность является просветительской (книги, кассеты), а Ваша (в виде
рекламы «Школы (гейши) искусства взаимоотношений» — коммерческой.
Но, так как развитие вагинальных управляемых мышц (ВУМбилдинг) является основой
для привлечения посетителей, эксплуатации их интереса и любопытства, «Школа
гейши» явилась мощным средством информирования и популяризации развития
вагинальных управляемых мышц (ВУМбилдинга). За что основателю «Школы (гейши)
искусства взаимоотношений» я всегда буду благодарен, а общество — оценит.
За что основателю «Школы (гейши)
искусства взаимоотношений» я всегда буду благодарен, а общество — оценит.
«…но самый экстравагантный случай с господином В.Л. Муранивским, моряком по специальности, который предлагает всем женщинам по кассетам и якобы придуманному им тренажеру, обучится подобному предмету. Господин Муранивский в своих зазываниях, лукавит трижды и по сему тянет на достойного шарлатана. Последствия его неудачных экспериментов приходится исправлять нам.»
Как известно, моряки всегда были любвеобильным народом, секс их всегда интересовал, а хороший (с ВУМами) — тем более. Кроме этого, г-н Фролов показывает свою некомпетентность даже в вопросах профессий: нет специальности «моряк», а есть «штурман», «радист», «механик» и т.д. Но это — отступление.
Г-н Фролов О.В. решил вершить свой суд и в области патентного права. Выносить
вердикт, что является патентоспособной идеей, а что обычной конструкторской
разработкой. Вот для таких «новаторов» и существует международное
патентное право. Человек с высшим образованием просто обязан знать это, и не
вводить намеренно в заблуждение читателя.
Вот для таких «новаторов» и существует международное
патентное право. Человек с высшим образованием просто обязан знать это, и не
вводить намеренно в заблуждение читателя.
Я являюсь изобретателем тренажера пневматического (ТП), конструкция которого защищена патентом РФ №2012321 с приоритетом от 21.03.1990 и более усовершенствованным типом, защищенным свидетельством РФ на полезную модель №24108 от 15.02.2002.
Примечание: можно найти в базе данных Федерального института промышленной собственности.
Вот тебе и «якобы придуманным», г-н Фролов О.В. Как Вы думаете отмываться от подобной лжи, основатель «Школы (гейши) искусства взаимоотношений»?
«Обучится подобному предмету» действительно можно по моим книжкам,
видеокассете, с помощью ТП и других моих запатентованных тренажеров, а также
описанных в моих книгах упражнений с
подручными средствами.
Это не является «одним из самых потрясающих и эксклюзивных занятий Школы», «тайных знаний тайских Мастеров». В «Школе (гейши) искусства взаимоотношений» обучают «пиратскими» копиями ТП, по методике, описанной в моих книгах и на сайтах.
Причем, от авторской инструкции к ТП в 16 страниц, оставили только 2, а остальные 14 используют в качестве «эксклюзивных» знаний, обучают тому, что было в авторской инструкции — как собирать, подгонять, тренироваться. Этому учатся пользователи Интернета совершенно бесплатно.
Да и вставлять в инструкцию, что Вы являетесь автором ТП беспардонно — это же чистый плагиат. Да и те оставшиеся две страницы можно было бы «передирать» без орфографических ошибок (Ваши бывшие ученицы приносили мне такие инструкции).
«Последствия его неудачных экспериментов приходится исправлять нам. «
Кому, после кого и что приходится исправлять, доказать достаточно сложно. Трудно
получить свидетельские доказательства в таком деликатном деле. Но могу Вас обрадовать:
тем, кто приходит ко мне после «Школы (гейши) искусства взаимоотношений»,
ничего исправлять не приходится, … они просто ничему не научились, ничего не
умеют, ничего не развили. С ними приходится начинать начальный курс с нуля по
полной программе.
«
Кому, после кого и что приходится исправлять, доказать достаточно сложно. Трудно
получить свидетельские доказательства в таком деликатном деле. Но могу Вас обрадовать:
тем, кто приходит ко мне после «Школы (гейши) искусства взаимоотношений»,
ничего исправлять не приходится, … они просто ничему не научились, ничего не
умеют, ничего не развили. С ними приходится начинать начальный курс с нуля по
полной программе.
Впрочем, нет. Приходится исправлять ТП. С этой стороны очень хорошо, что г-н Фролов О.В. приписывает себе авторство на ТП — мне не приходится оправдываться и краснеть за то качество, которое придали ему «умельцы» из «Школы (гейши) искусства взаимоотношений» (даже скопировать с имеющегося образца толком не сумели!).
Это для государственной собственности достаточно
хватательных рефлексов, а интеллектуальная собственность — вещь деликатная,
ума требует, на худой конец, обычной внимательности.
«Во — первых, подобный предмет может преподавать человек обязательно имеющий высшее медицинское образование.»
Подобный предмет не изучается ни в одном медицинском образовательном учреждении. Это физкультура, спорт. Международная патентная классификация (МПК 7) именно к этому относит методики и устройства для развития вагинальных мышц. (можно посмотреть здесь)
Вы слышали когда-нибудь об успехах спортсменки Галины Подольневой? Она развила эти мышцы до такой силы, что (по ее словам) может ими колоть грецкие орехи (без комментариев). А разве «тайские посвященные Мастера», на которых Вы постоянно ссылаетесь, являются врачами? А лично Вы учились сексу по медицинскому пособию или под руководством врача? А врач-стоматолог или хирург с высшим медицинским образованием может преподавать сей предмет?
Если
один из партнеров научил другого
изумительному действию, оба оказались
наверху блаженства, то уместен вопрос: «Где
ты этому научилась (научился)?». Но
дурацким будет: «Ты гинеколог?»
Но
дурацким будет: «Ты гинеколог?»
«Во-вторых, подобный предмет должна вести женщина, хотя бы, для того, чтобы продемонстрировать свой опыт. Как и чем демонстрирует свой опыт этот учитель, для нас загадка?»
Своими рассуждениями основатель «Школы (гейши) искусства взаимоотношений» исключает участие мужчин в обучение женщин и, видимо, наоборот.
А как же великие спортивные тренеры, а тот же Кегель? Даже мировую известность получил благодаря разработанным упражнениям. Да, мужчина показать этого не может, но я могу продемонстрировать на видео достижения своих учениц, согласившихся на съемку. В четырехчасовой видеолекции «Счастье и брак» показываются уникальные умения. Да и продемонстрировать — не значит научить.
Очень одаренные в этом плане женщины до сих
пор на занятия не приходили. Может быть теперь, после появления программы «Элитарный
секс», они появятся, и кто-нибудь захочет
продемонстрировать свои непревзойденные
возможности.
Несмотря на апломб, Вы, г-н Фролов О.В, как говорится, до сих пор «не въехали в тему». Кто или что может наилучшим образом оценить и скорректировать действия вагинальных мышц? Вы считаете, что женщина, конечно, не имея в виду лесбийский вариант. А я считаю, что мужчина, партнер, вернее, его фаллос. Именно он подсказал мне в свое время, что есть хорошо. Именно он заставил меня почувствовать ущербность безмышечной вагины. Именно он руководил моей «преподавательской» деятельностью. Вот когда его возможности не стали справляться с потоком желающих освоить ВУМбилдинг, на помощь пришел изобретенный тренажер ТП.
Если Вы до сих пор этого не почувствовали, не
ощутили, не можете подсказать (не стали Учителем), то Ваша «очаровательная
женщина, врач по образованию, и реальный Мастер данного предмета» ничего
не умеет. То есть какие-то трюки она может быть и освоила. Но главное применение
своих умений реализовать не может. Вы, как мужчина, этому ее не учите. Если
бы не Ваше «Обращение», то я смог бы ее кое-чему научить.
Вы, как мужчина, этому ее не учите. Если
бы не Ваше «Обращение», то я смог бы ее кое-чему научить.
Я говорил, говорю и буду говорить, что
лучшим тренажером для мужчины является женщина с ВУМами, а для женщины — мужчина с фаллосом, способным оценить ее достижения.
Это самые физиологичные и естественные тренировки, а следовательно, дающие максимальный
эффект в кратчайшее время. К сожалению, большинство мужчин работают фаллосом
только для достижения собственного оргазма или для удовлетворения женщины, но
полностью исключают даже мысль о его использовании (на короткое время для своей
же пользы) в качестве тренажера. Невероятно трудно встретить мужчине женщину
с ВУМами, но еще труднее женщине встретить мужчину, способного помочь в освоении ВУМбилдинга. Да и не всегда такой тренер может быть «под рукой» (вернее,
там, где нужно), когда есть время и желание для тренировки.
» — В третьих, данный господин претендует на авторство тренажера по управлению и развитию интимных мышц — неправда, ибо изобретен он был ученым Кегелем в 50-х годах и подробно описан в любой медицинской энциклопедии. А уже по описанию, для любого конструктора, сделать подобный аппарат не составляет труда. НО вся закавыка в том, что сам по себе тренажер без учителя — НИЧТО, ибо только под руководством Мастера, можно научится — и другого пути нет.»
«Любая медицинская энциклопедия» ни слова не говорит о пневматическом тренажере, речь идет, в основном, только об упражнениях.
Кроме того, порядочный оппонент всегда указывает конкретный источник информации, вплоть до номера страницы и абзаца, а ссылка «любая энциклопедия» неправомочна.
О заблуждении основателя «Школы (гейши) искусства взаимоотношений»
в части патентного права уже говорилось. Ну не дано ему право решений в части
патентоспособности, а как хочется казаться компетентным и мудрым.
Ну не дано ему право решений в части
патентоспособности, а как хочется казаться компетентным и мудрым.
Если тренажер, как и любое другое изделие, требует для освоения специальной подготовки, «руководства Мастера», то права на нормальные потребительские свойства он не имеет. Это же не космический корабль.
А вот, если составить инструкцию (вернее, выбросить все нужное) без правил использования, то пользователь может и не разобраться. Авторская инструкция по ТП содержит все необходимые рекомендации, о чем говорят результаты применения тренажера.
На этом же сайте есть фраза г-на Фролова О.В. «Т.к. мы интересны, нас стало модно показывать по телевидению и освещать в прессе.»
К сожалению, к этому
интересу примешан коммерческий интерес, удовлетворить который мне не под силу.
Если бы СМИ действительно были заинтересованы в оказании помощи людям, я с удовольствием
помог бы миллионам женщин овладеть элементарными навыками развития вагинальных
управляемых мышц (ВУМбилдинга).
Все это давно изложено в моих книжках, но их тираж ограничен. Да и «пиратов» поубавилось бы.
«А так как первое образование Олега — инженер-конструктор, а я по специальности — врач, в процессе определенных совместных усилий, — изобрёлся уникальный тренажер для развития женских интимных мышц, который просто творит чудеса.»
Очень хотелось бы узнать номер патента на это изобретение или хотя бы номер заявки на выдачу патента. Без этого вроде голословной получается реклама, бездоказательной.
Эта публикация касается не только О. Фролова, основателя «Школы (гейши) искусства взаимоотношений», но и других оппонентов, публикующих бездоказательные, оскорбительные и злые комментарии к моим работам на всевозможных форумах и сайтах.
Я с благодарностью
отнесусь к любой критике, способствующей и совершенствующей методику развития
вагинальных управляемых мышц (ВУМбилдинга). Но, не лейте, пожалуйста, грязь в это
нужное, с трудом развивающееся, дело.
Но, не лейте, пожалуйста, грязь в это
нужное, с трудом развивающееся, дело.
Обратите внимание, что показания манометров пиратских ТП не соответсвуют приведенным в авторской методике Вумбилдинг. Поэтому применять их с рекомендуемыми нами давлениями нельзя.
Эйфория, Красноярский центр вумбилдинга им. В.Л. Муранивского в Красноярске
10 важных витаминов для каждой женщиныДля красоты и здоровья каждой женщине необходимо получать витамины и микроэлементы. Какие же полезные вещества должны быть в ежедневном рационе каждой женщины, где они содержатся и чем полезны?
Витамин С
Где содержится:
Ягоды, цитрусовые, сладкий перец, брокколи.
Польза:
Оказывает противовоспалительное действие, повышает сопротивляемость организма инфекциям, регулирует обмен веществ.
Витамин Е
Где содержится:
В растительных маслах, злаках.
Польза:
Приостанавливает процессы увядания организма, предотвращает риск инфарктов и инсультов, укрепляет стенки сосудов.
Фолиевая кислота
Где содержится:
Цитрусовые, зеленые овощи, цельнозерновые злаки
Польза:
Жизненно необходима для будущих мам, чтобы плод развивался правильно. Также нужен для усвоения других витаминов: С и витаминов группы В.
Витамин РР
Где содержится:
Бобовые и цельнозерновые злаки
Польза:
Отвечает за контроль холестерина в крови, помогает получить энергию из жира и глюкозы.
Железо
Где содержится:
Морепродукты, мясо, печенка.
Польза:
Борется с анемией.
Калий
Где содержится:
Апельсины, бананы, изюм, зелень.
Польза:
Необходим для правильной работы сердца.
Бета-каротин
Где содержится:
Морковь, тыква, сладкий перец.
Польза:
Преобразуется в витамин А, необходимый для глаз, кожи и иммунной системы.
Витамин D
Где содержится:
В печени трески, жирной рыбе, яичном желтке.
Польза:
Необходим для усвоения кальция, предотвращает развитие рахита у младенцев.
Кальций
Где содержится:
Во всех молочных продуктах
Польза:
Необходим для укрепления костей, зубов, предотвращает остеопороз. Усваивается с витамином D.
Витамин группы В
Где содержится:
Мясо, рыба, твердый сыр, бобовые, цельные злаки, брокколи, листовые зеленые овощи.
Польза:
Необходим для нормальной работы нервной системы. Дефицит этих витаминов приводит к ухудшению памяти и депрессивным состояниям.
Будьте здоровы! Будьте желанны и любимы!
Моделирование, эксперимент и эволюция: понимание зародышеобразования при сворачивании белка S6
Понимание переходного состояния (TS) является одной из основных технических и интеллектуальных вех на пути к пониманию реакции сворачивания белка (1). В нескольких недавних исследованиях (2–6) были предприняты попытки построить структуры ансамбля TS (TSE) с использованием значений φ в качестве структурных ограничений при моделировании развертывания. Благодаря обширным исследованиям экспериментально и расчетно проверенного белка G (7) мы показали, что экспериментальные значения φ (φ exp ) могут использоваться в моделировании для построения предполагаемого TSE, но это измерение коэффициента передачи конформации (« вероятность сброса», p раз ) является единственным средством, с помощью которого структура может быть классифицирована как член TSE. Однако следует также быть осторожным при выборе того, какой φ exp использовать, потому что точечные мутации, на которых они основаны, могут изменить стабильность белка или структурные особенности TSE, что сделает нормализацию данных дикого типа неоднозначной (8). Учитывая, что наш метод изучения структуры TSE был подтвержден в сложном случае фолдинга белка G (7), мы теперь можем провести такой анализ для других белков на сравнительной основе, чтобы помочь в различении экспериментальных несоответствий. , шум или артефакты, а также для определения общих знаменателей критического ядра при сворачивании белка.
Однако следует также быть осторожным при выборе того, какой φ exp использовать, потому что точечные мутации, на которых они основаны, могут изменить стабильность белка или структурные особенности TSE, что сделает нормализацию данных дикого типа неоднозначной (8). Учитывая, что наш метод изучения структуры TSE был подтвержден в сложном случае фолдинга белка G (7), мы теперь можем провести такой анализ для других белков на сравнительной основе, чтобы помочь в различении экспериментальных несоответствий. , шум или артефакты, а также для определения общих знаменателей критического ядра при сворачивании белка.
Расщепленный β-α-β рибосомный белок S6 из Thermus thermophilus состоит из 97 остатков в четырехцепочечном β-слое, упакованном против двух α-спиралей с гидрофобным ядром (9). Функционально S6 связывается как с РНК, так и со своим белком-партнером S18 кооперативным образом во время промежуточной стадии образования 30S рибосомной субъединицы (10). S6 является идеальным кандидатом для вычислительного исследования из-за большого количества доступных высококачественных экспериментальных данных, включая обширные данные о кинетике и значении φ при различных концентрациях денатуранта (11), исследования круговых пермутантов, которые отражают перестройки в TSE (12 ), а также исследования индуцированных солью промежуточных соединений вне пути (13).![]() Подробная структурная информация также существует для функции S6 (10).
Подробная структурная информация также существует для функции S6 (10).
Мы используем моделирование для структурной интерпретации комбинированного набора значений φ S6. Мы начинаем с создания ансамбля структур, соответствующих структурным ограничениям на основе φ exp . После характеристики каждой конформации ансамбля путем измерения ее p раз , мы строим подробную модель TSE и событий, происходящих до и после зародышеобразования. Эта формализованная обработка φ exp позволяет проводить микроскопический анализ и реконструкцию процесса зародышеобразования складчатости.Наши результаты подтверждают идею о том, что экспериментально наблюдаемые сдвиги значений φ, вызванные денатурантами, сдвигают TSE по профилю свободной энергии и, следовательно, вызывают события раньше или позже на пути сворачивания (как интерпретируется через постулат Хаммонда). Мы также анализируем закономерности сохранения остатков S6, чтобы определить эволюционную историю остатков ядра в расщепленном семействе β-α-β. Благодаря сочетанию экспериментальных данных, информации об эволюции и моделированию всех атомов мы можем расширить интерпретацию экспериментальных данных и создать унифицированное и упорядоченное описание нуклеации, переходного состояния и фолдинга в белке S6 на атомном уровне.
Благодаря сочетанию экспериментальных данных, информации об эволюции и моделированию всех атомов мы можем расширить интерпретацию экспериментальных данных и создать унифицированное и упорядоченное описание нуклеации, переходного состояния и фолдинга в белке S6 на атомном уровне.
Теория и методы
Модель Система. Наш протокол был ранее реализован для реконструкции TSE CI2 (6) и белка G (7) и использовался для моделирования полной укладки белка G (14) и крамбина (15) от случайной катушки до расстояния <1 Å среднеквадратичного значения. (дрмс) из родного состояния. Модель включает представление всех неводородных атомов в основной и боковых цепях в виде твердых сфер, полное представление вращательных степеней свободы основной и боковой цепей, Го-потенциал квадратной ямы (16) с нативными контактами, имеющими притяжение -1 и ненативные контакты, имеющие отталкивание +1 и набор движений MC (с локализованными движениями основной и боковой цепи), который поддерживает связность цепи, плоские пептидные связи и исключенный объем на каждом этапе. В качестве меры структурного сходства с исходным состоянием drms рассчитывается как , где D и D 0 — попарные расстояния C α в выбранной и нативной конформациях.
В качестве меры структурного сходства с исходным состоянием drms рассчитывается как , где D и D 0 — попарные расстояния C α в выбранной и нативной конформациях.
Преимущество этой модели заключается в том, что она позволяет собирать статистически значимое количество траекторий, включая детали с атомарным разрешением, такие как упаковка боковых цепей. Потенциалы Go адекватно представляют термодинамику и кинетику белков с минимальными энергетическими нарушениями и позволяют изучить весь процесс сворачивания (17).Их использование также мотивировано теоретическими и экспериментальными данными о том, что переходное состояние является устойчивым в отношении выбора определенных последовательностей, которые сворачиваются до заданной структуры, и наборов потенциалов, используемых для проектирования и фолдинга последовательностей (18–20). Модели Go использовались для предложения механизмов сворачивания (21), прогнозирования скорости сворачивания и интерпретации значений φ (22–24). Они также успешно предсказали значения φ для нескольких белков (25–27). В настоящее время не существует общего потенциала, способного сворачивать α/β-белки, поэтому Go-потенциалы представляют собой лучший вариант для изучения сворачивания малых белков (28).
Они также успешно предсказали значения φ для нескольких белков (25–27). В настоящее время не существует общего потенциала, способного сворачивать α/β-белки, поэтому Go-потенциалы представляют собой лучший вариант для изучения сворачивания малых белков (28).
Конструирование предполагаемых TS-конформаций. Структуры были построены с помощью моделирования развертывания с ограничениями (7) из исходной структуры Protein Data Bank (идентификационный код PDB 1RIS). Обычная интерпретация φ exp — это доля нативных контактов, созданных конкретным остатком в TSE. Мы определяем симуляционное значение φ (φ sim ), которое может быть рассчитано для любого остатка, как долю нативных контактов, созданных остатком i в конформации k .Гармонические ограничения были введены путем использования φ sim для создания структур, отвечающих экспериментальным ограничениям (т. е. имеющих φ sim = φ exp ). Эти ограничения были наложены добавлением энергии φ, E φ , энергии Go, E Перейти .
где Λ — весовой коэффициент, i — остаток, подлежащий защемлению, N — общее количество защемлений.Экспериментальные значения φ, использованные в качестве ограничений, приведены в таблице 1. Моделирование развертывания с использованием функции энергии в уравнениях. 2 и 3 , были инициированы из нативного состояния и распространялись на 10 7 шагов MC. Λ = 10 4 обеспечил быстрое и полное выполнение ограничений. Среднее значение было <10 -3 в конце минимизации. Хотя уравнение 1 не является точным, в нескольких исследованиях было показано, что он является разумным показателем для φ exp (2–6).
Эти ограничения были наложены добавлением энергии φ, E φ , энергии Go, E Перейти .
где Λ — весовой коэффициент, i — остаток, подлежащий защемлению, N — общее количество защемлений.Экспериментальные значения φ, использованные в качестве ограничений, приведены в таблице 1. Моделирование развертывания с использованием функции энергии в уравнениях. 2 и 3 , были инициированы из нативного состояния и распространялись на 10 7 шагов MC. Λ = 10 4 обеспечил быстрое и полное выполнение ограничений. Среднее значение было <10 -3 в конце минимизации. Хотя уравнение 1 не является точным, в нескольких исследованиях было показано, что он является разумным показателем для φ exp (2–6). Более того, было показано, что значения φ, предсказанные с использованием нашей модели и уравнения 1 хорошо коррелируют с достоверными экспериментальными значениями ( R = 0,70) (7).
Более того, было показано, что значения φ, предсказанные с использованием нашей модели и уравнения 1 хорошо коррелируют с достоверными экспериментальными значениями ( R = 0,70) (7).
Значения φ S6 из эксперимента ( 11 ) и моделирование, а также значение CoC и среднее количество нелокальных контактов ( N нелокальный ) в TSE
Проверка ТС. структуры не может быть априорными предполагается членом TSE просто потому, что она соответствует набору ф ограничений. Для того, чтобы определить, является ли структура членом TSE в теоретически обоснованным образом, необходимо измерить его р раза. Коэффициент передачи или р раза 0,5 определяет TS. Для того, чтобы определить эту вероятность, 20 пробных прогонов раскладных были выполнены для каждой структуры. Поскольку расчет р раза суммы в испытании Бернулли, относительная погрешность в результате с помощью N , как бежит весы Н -1/2 .Таким образом, 20 пробных циклов складывания оцениваются в p . раз с точностью до 22% от среднего. Как было показано ранее для фолдинга белков (7) и многих других сложных систем (29), необходимо, чтобы каждый пробный прогон был достаточно длинным, чтобы подтвердить коммитацию. Другими словами, конкретная структура склонна к складыванию, если она прошла барьер TS и если вероятность повторного пересечения барьера (т. е. развертывания) незначительна. Как и в более ранних исследованиях (14), мы определяем τ commit («время фиксации» и продолжительность p раз имитация) как 10 7 шагов MC.
Коэффициент передачи или р раза 0,5 определяет TS. Для того, чтобы определить эту вероятность, 20 пробных прогонов раскладных были выполнены для каждой структуры. Поскольку расчет р раза суммы в испытании Бернулли, относительная погрешность в результате с помощью N , как бежит весы Н -1/2 .Таким образом, 20 пробных циклов складывания оцениваются в p . раз с точностью до 22% от среднего. Как было показано ранее для фолдинга белков (7) и многих других сложных систем (29), необходимо, чтобы каждый пробный прогон был достаточно длинным, чтобы подтвердить коммитацию. Другими словами, конкретная структура склонна к складыванию, если она прошла барьер TS и если вероятность повторного пересечения барьера (т. е. развертывания) незначительна. Как и в более ранних исследованиях (14), мы определяем τ commit («время фиксации» и продолжительность p раз имитация) как 10 7 шагов MC. Приверженность складыванию была определена как встреча структуры ( i ) E Go <-1100 (родной E Go = -1,569), ( ii ) костяк drms < 2,5 Å и ( iii ) доля нативных контактов ( Q ) > 0,75. Р Затем рассчитывали раз как долю (всего 20) прогонов, которые закончились зафиксированной конформацией. Коммитирование происходит в результате специфического коллапса, который в нашей модели эквивалентен полному образованию ядра.После коллапса цепи в условиях складывания ( T = 1) остов никогда не принимает растянутой конформации. Этот результат подтверждается обширными эмпирическими наблюдениями за моделированием фолдинга крамбина (15), белка G (14) и S6, которые демонстрируют, что коммитированные структуры всегда переходят в нативное состояние. Таким образом, при коллапсе белок коммитируется, и нужно только контролировать ( i ) E , чтобы исключить ловушки упаковки боковых цепей, ( ii ) компактность с помощью структурных параметров, таких как drms, и ( iii ) формирование вторичной структуры ( Q )раздвижные складные ловушки.
Приверженность складыванию была определена как встреча структуры ( i ) E Go <-1100 (родной E Go = -1,569), ( ii ) костяк drms < 2,5 Å и ( iii ) доля нативных контактов ( Q ) > 0,75. Р Затем рассчитывали раз как долю (всего 20) прогонов, которые закончились зафиксированной конформацией. Коммитирование происходит в результате специфического коллапса, который в нашей модели эквивалентен полному образованию ядра.После коллапса цепи в условиях складывания ( T = 1) остов никогда не принимает растянутой конформации. Этот результат подтверждается обширными эмпирическими наблюдениями за моделированием фолдинга крамбина (15), белка G (14) и S6, которые демонстрируют, что коммитированные структуры всегда переходят в нативное состояние. Таким образом, при коллапсе белок коммитируется, и нужно только контролировать ( i ) E , чтобы исключить ловушки упаковки боковых цепей, ( ii ) компактность с помощью структурных параметров, таких как drms, и ( iii ) формирование вторичной структуры ( Q )раздвижные складные ловушки.
Результаты и обсуждение
Улучшенное изображение S6 TSE. Свойства нашего предполагаемого TSE приведены в таблице 2. Большинство конформаций имеют промежуточные p раз значений, что указывает на то, что TSE был эффективно отобран и идентифицирован. Однако, как отмечалось ранее (7), не все структуры, удовлетворяющие ограничениям φ, являются истинными TS-конформациями.Этот вывод не удивителен, особенно принимая во внимание информативную аналогию с определением структуры белка с помощью ЯМР, которая сильно зависит от количества и качества ограничений NOE и результаты которых часто требуют дополнительного уточнения (30).
Таблица 2. Свойства ансамбля На рис. 1 представлено параллельное сравнение нативного S6 и конформаций в его TSE (т. е., с р раз = 0,5). В TSE спираль 1 близка к нативной, а вторая и третья β-цепи имеют хорошо сформированную вторичную структуру. Спираль 2 создает небольшое количество нативных контактов, в то время как цепь 1 создает очень мало, а цепь 4 практически денатурирована. В том смысле, что TSE демонстрирует общую расширенную нативную топологию, можно интерпретировать преобладание промежуточных значений φ как обозначение диффузного TSE. Однако, как будет обсуждаться далее, исследование на уровне остатков выявляет четко определенную, полностью установленную сеть контактов, составляющих специфическое ядро.
е., с р раз = 0,5). В TSE спираль 1 близка к нативной, а вторая и третья β-цепи имеют хорошо сформированную вторичную структуру. Спираль 2 создает небольшое количество нативных контактов, в то время как цепь 1 создает очень мало, а цепь 4 практически денатурирована. В том смысле, что TSE демонстрирует общую расширенную нативную топологию, можно интерпретировать преобладание промежуточных значений φ как обозначение диффузного TSE. Однако, как будет обсуждаться далее, исследование на уровне остатков выявляет четко определенную, полностью установленную сеть контактов, составляющих специфическое ядро.
Два представления исходной ( a ) структуры S6 и репрезентативной структуры переходного состояния ( b ). Остатки ядра (V6, I8, I26, L30, P60, V65 и L75) показаны явно.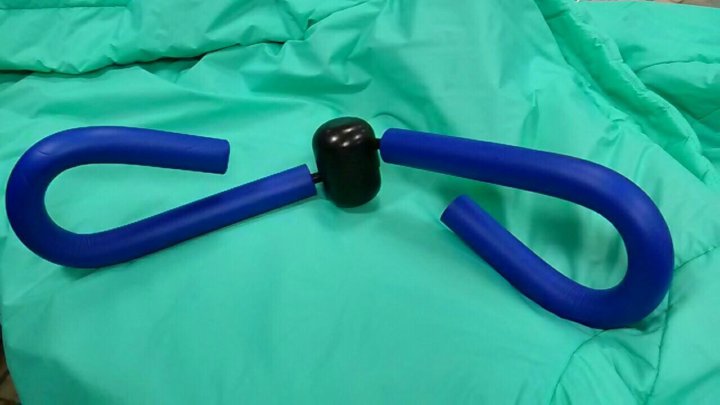 Продвижение по цепи обозначено цветом от синего (N-конец) до красного (C-конец).Изображения были созданы с использованием imol.
Продвижение по цепи обозначено цветом от синего (N-конец) до красного (C-конец).Изображения были созданы с использованием imol.
Зародышевые контакты. С макроскопической точки зрения эксперимент дает картину диффузной, нативной ТГЭ в S6, аналогичной таковой при CI2 (31). Хотя эта точка зрения согласуется с поведением зародышеобразования-конденсации, требуется более глубокое изучение, чтобы понять формирование и роль специфического ядра нелокальных контактов, которые определяют топологию TSE. Здесь мы определяем полностью сформированное (во всех конформациях TSE) подмножество дальних контактов как специфическое ядро.Идентификация специфического ядра важна, потому что после прохождения энергетического барьера для нуклеации структура подвергается быстрому сворачиванию основной цепи, за которым следует более медленный процесс релаксации боковой цепи (32). Для определения роли отдельных остатков в процессе нуклеации были созданы карты контактов для нативной и TSE конформаций (рис. 2 а и б ).
). Из этих карт контактов видно, что небольшое количество остатков составляет большую часть дальних контактов.Эти нелокальные контакты образуются между нитями 1 и 3, нитями 2 и 3, спиралью 2 и нитью 1 и, в меньшей степени, между спиралью 1, петлей 2 и второй спиралью.
2 а и б ).
). Из этих карт контактов видно, что небольшое количество остатков составляет большую часть дальних контактов.Эти нелокальные контакты образуются между нитями 1 и 3, нитями 2 и 3, спиралью 2 и нитью 1 и, в меньшей степени, между спиралью 1, петлей 2 и второй спиралью.
Контактные матрицы 1RIS для нативной ( a ), TSE ( b ) и разностной матрицы ( c ) (Post- и Pre-TS), где верхний левый соответствует положительным значениям, а нижний левый соответствует отрицательные значения.
На рис. 3 показано среднее общее количество нелокальных контактов, производимых каждым остатком в TSE (ограничения развертывания определяют количество контактов, производимых удерживаемыми остатками, и косвенно влияют на соседние контакты), что проясняет роль отдельных остатков в нуклеации. Некоторые остатки с высоким φ exp , такие как I26, L30 и M67, не образуют большого количества дальних контактов. Тем не менее, I26 и L30 образуют единственные дальние контакты в спирали 1, которая, по-видимому, формируется на ранней стадии сворачивания и которая контактирует с I8, V65 и L75 в TSE.
Некоторые остатки с высоким φ exp , такие как I26, L30 и M67, не образуют большого количества дальних контактов. Тем не менее, I26 и L30 образуют единственные дальние контакты в спирали 1, которая, по-видимому, формируется на ранней стадии сворачивания и которая контактирует с I8, V65 и L75 в TSE.
Статистика контактов S6 TSE. ( a ) Средняя доля местных контактов, установленных каждым жителем. Кружки обозначают φ-ограничения. ( b ) Среднее количество нелокальных (за исключением n до n ± 5) контактов, сделанных каждым резидентом. ( c ) CoC в S6.Остатки, участвующие в зародышеобразовании (как определено с помощью анализа p кратного и m ) обозначены значком x, а остатки с φ exp ≥ 0,3 обведены кружком, подчеркивая соответствие между CoC и φ exp . Кроме того, CoC (вместе со значениями m и моделированием) указывает на некоторые остатки ядра с явно низким значением φ exp , такие как L79. CoC может также идентифицировать остатки, имеющие функциональное значение, такие как I52.
Кроме того, CoC (вместе со значениями m и моделированием) указывает на некоторые остатки ядра с явно низким значением φ exp , такие как L79. CoC может также идентифицировать остатки, имеющие функциональное значение, такие как I52.
Все остальные остатки с высоким значением φ exp сделал большое количество нелокальных контактов (Y4, V6, I8, F60, V65 и L75).Кроме того, каждый остаток с φ exp < 0,20 установили несколько (<5) нелокальных контактов. Три остатка, которые выделяются на рис. 3 б из-за большого количества нелокальных контактов не могут быть надежно охарактеризованы экспериментами по белковой инженерии из-за недостаточной стабильности изменений. Y33 (φ sim = 0,40), который расположен на конце спирали 1 и взаимодействует с остатками с высоким φ L30 и L75, чтобы объединить спирали 1 и 2 и сформировать часть гидрофобного ядра, не мог дать φ exp , потому что мутация Y33A дестабилизировала белок только на 0. 38 ккал/моль. L48 (φ sim = 0,74) закрепляет область длинной петли между листами 2 и 3, образуя погребенный гидрофобный кластер с I52 и F60 (φ эксп = 0,36). Интересно, что укорочение остатков L48 и I52 до А, которое демонстрирует сильную консервативность в расщепленном семействе β-α-β (11), не приводит к значительной дестабилизации белка.
38 ккал/моль. L48 (φ sim = 0,74) закрепляет область длинной петли между листами 2 и 3, образуя погребенный гидрофобный кластер с I52 и F60 (φ эксп = 0,36). Интересно, что укорочение остатков L48 и I52 до А, которое демонстрирует сильную консервативность в расщепленном семействе β-α-β (11), не приводит к значительной дестабилизации белка.
Важная роль I26 и L30 в определении топологической ориентации спирали 1 в ядре сворачивания также показана экспериментально через кинетические значения m (рис.4). Укорочение боковых цепей I26 и L30 дает увеличение разворачивания м — значение, м и (11), что свидетельствует о смещении пути вдоль параметров вторичного порядка (33). Если спираль 1 не закреплена дальнодействующими контактами в начале сворачивания, S6 складывается по альтернативной «мутантной» траектории. Последовательно, эффект усиливается у двойных мутантов I26A/L30A ( m u = 0,90), I8A/I26A ( м и = 0. 75) и L30A/L75A ( м u = 0,85), которые показывают разворачивание м -значения почти в два раза выше, чем для белка дикого типа ( м u = 0,50) (рис. 4). Способность S6 свертываться через такое альтернативное ядро была независимо продемонстрирована в экспериментах по круговой перестановке (12). Интересно, что у I26A/L79A тоже высокая м u (0,75), что выше, чем у одного I26A ( м и = 0.65), т. е. для L79 изменение является аддитивным. Однако двойные мутанты в других положениях S6 показывают уровень дикого типа или ниже м u значений (т. е. показывают различную степень сдвига Хаммонда). Более того, мы видим, что двойные мутанты 26/79, 8/26, 30/75 и 26/30 и, возможно, одиночные мутанты L30, W62, Y63 и V65 показывают более высокие значения рефолдинга м f , что указывает на то, что сеть контактов, определяемая этими позициями, в некоторой степени присутствует в денатурированном ансамбле.
75) и L30A/L75A ( м u = 0,85), которые показывают разворачивание м -значения почти в два раза выше, чем для белка дикого типа ( м u = 0,50) (рис. 4). Способность S6 свертываться через такое альтернативное ядро была независимо продемонстрирована в экспериментах по круговой перестановке (12). Интересно, что у I26A/L79A тоже высокая м u (0,75), что выше, чем у одного I26A ( м и = 0.65), т. е. для L79 изменение является аддитивным. Однако двойные мутанты в других положениях S6 показывают уровень дикого типа или ниже м u значений (т. е. показывают различную степень сдвига Хаммонда). Более того, мы видим, что двойные мутанты 26/79, 8/26, 30/75 и 26/30 и, возможно, одиночные мутанты L30, W62, Y63 и V65 показывают более высокие значения рефолдинга м f , что указывает на то, что сеть контактов, определяемая этими позициями, в некоторой степени присутствует в денатурированном ансамбле.
Экспериментальный м и ( и ) и м f ( b ) значения для дикого типа и мутантного S6. Значения м были определены стандартными процедурами из линейных областей шевронных графиков, за исключением областей кривизны вниз, которые иногда наблюдаются при высоких концентрациях денатуранта (11).Остатки в конкретном ядре демонстрируют самые высокие м -значения, указывающие на изменение траектории складывания и потерю остаточной структуры в денатурированном ансамбле для м и и м и соответственно.
Эволюционные сигналы нуклеации. Эволюционные сигналы универсально консервативных положений остатков ранее были связаны с информацией о стабильности, кинетике и функции белков.С этой целью мы проанализировали белки, которые имеют общую укладку S6, но не ее последовательность, посредством «консервации сохранения» (CoC) (34). Этот метод также показал свою эффективность при идентификации ядер свертывания белка (35). Для S6 были рассмотрены 42 семейства из базы данных Families of Structurally Similar Proteins с показателем Dali Z > 4 и ID последовательности < 25% (36). Результаты этого анализа представлены на рис. 3 c , где низкие значения указывают на высокую консервативность позиции в исследованных семьях.
Эволюционные сигналы универсально консервативных положений остатков ранее были связаны с информацией о стабильности, кинетике и функции белков.С этой целью мы проанализировали белки, которые имеют общую укладку S6, но не ее последовательность, посредством «консервации сохранения» (CoC) (34). Этот метод также показал свою эффективность при идентификации ядер свертывания белка (35). Для S6 были рассмотрены 42 семейства из базы данных Families of Structurally Similar Proteins с показателем Dali Z > 4 и ID последовательности < 25% (36). Результаты этого анализа представлены на рис. 3 c , где низкие значения указывают на высокую консервативность позиции в исследованных семьях.
Первый кластер низких остатков CoC расположен на листе 1 и включает V6, I8, L10 и P12. V6 и I8 имеют высокие измеренные значения φ exp и образуют большое количество нелокальных контактов в TSE, как и P12. Напротив, L10, который является частью этого гидрофобного кластера, делает очень мало нелокальных контактов, что может быть артефактом медленного (φ exp = 0,07) соседнего V9.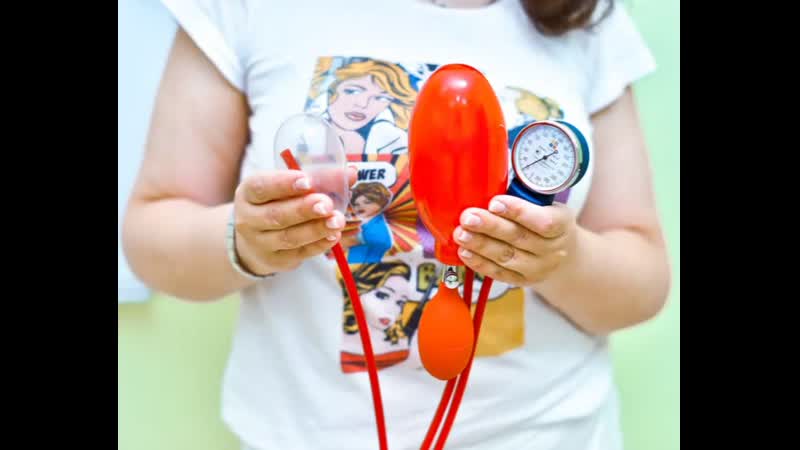 Следующие два низких остатка CoC, I26 и L30 в спирали 1, демонстрируют относительно высокие значения φ exp .Хотя эти остатки образуют сравнительно немного дальних контактов, они играют однозначную важную роль как единственные нелокальные контакты, осуществляемые рано формирующейся первой спиралью. V37, который демонстрирует низкий CoC, участвует в третичных взаимодействиях во второй цепи. Несмотря на умеренные значения φ exp = 0,24, предполагалось, что V37 играет роль на поздних стадиях TSE (11). Мутация низкого остатка CoC I52, который расположен в гидрофобном кластере в области длинной петли, соединяющей нити 2 и 3, и который образует большое количество нелокальных контактов в TSE, не привела к значительной дестабилизации, поэтому значение φ могло бы не измерить.Остатки L48, I52 и F60 находятся в тесном контакте с R87, который может играть функциональную роль в связывании РНК (9). Фактически, большинство остатков в этой области петли демонстрируют высокую степень упорядоченности в TSE (рис.
Следующие два низких остатка CoC, I26 и L30 в спирали 1, демонстрируют относительно высокие значения φ exp .Хотя эти остатки образуют сравнительно немного дальних контактов, они играют однозначную важную роль как единственные нелокальные контакты, осуществляемые рано формирующейся первой спиралью. V37, который демонстрирует низкий CoC, участвует в третичных взаимодействиях во второй цепи. Несмотря на умеренные значения φ exp = 0,24, предполагалось, что V37 играет роль на поздних стадиях TSE (11). Мутация низкого остатка CoC I52, который расположен в гидрофобном кластере в области длинной петли, соединяющей нити 2 и 3, и который образует большое количество нелокальных контактов в TSE, не привела к значительной дестабилизации, поэтому значение φ могло бы не измерить.Остатки L48, I52 и F60 находятся в тесном контакте с R87, который может играть функциональную роль в связывании РНК (9). Фактически, большинство остатков в этой области петли демонстрируют высокую степень упорядоченности в TSE (рис. 3 a ).
), несмотря на несколько дальних контактов. Сворачивание петель, которое, по-видимому, происходит полуавтономно от зародышеобразования, может быть результатом функциональной роли связывания белка, необходимой для образования рибосом (11). Низкие остатки CoC Y63 и V65 в цепи 3 расположены в центре кластера с высоким φ, который включает F60 и M67.Хотя Y63 имеет более низкий φ exp , он обеспечивает гораздо более дальние контакты в TSE, чем более высокий φ exp V65 или M67. В этом смысле эксперимент может опровергнуть важность роли Y63 в нуклеации. Другая группа низких остатков CoC — это V72, L75 и L79. L75 имеет высокое значение φ exp , а V72 демонстрирует самое высокое значение φ sim в спирали 2. ТСЭ или, что более вероятно, роль, которая была смягчена искусственно заниженным сдерживанием.
3 a ).
), несмотря на несколько дальних контактов. Сворачивание петель, которое, по-видимому, происходит полуавтономно от зародышеобразования, может быть результатом функциональной роли связывания белка, необходимой для образования рибосом (11). Низкие остатки CoC Y63 и V65 в цепи 3 расположены в центре кластера с высоким φ, который включает F60 и M67.Хотя Y63 имеет более низкий φ exp , он обеспечивает гораздо более дальние контакты в TSE, чем более высокий φ exp V65 или M67. В этом смысле эксперимент может опровергнуть важность роли Y63 в нуклеации. Другая группа низких остатков CoC — это V72, L75 и L79. L75 имеет высокое значение φ exp , а V72 демонстрирует самое высокое значение φ sim в спирали 2. ТСЭ или, что более вероятно, роль, которая была смягчена искусственно заниженным сдерживанием.
Рис. 3 c указывает на то, что CoC тесно связан с нуклеацией: например, из 14 остатков, участвующих в нуклеации по значению φ, анализу p кратно и m , 12 имеют низкий CoC <0,5. Вероятность того, что это произойдет случайно, составляет ≈10 -7 . Ценность CoC явно заключается в том, как он дополняет φ exp и моделирование. Чтобы идентифицировать и понять складчатое ядро, простого изучения φ exp недостаточно.Один такой пример, L79, имеет низкое значение φ exp , но играет роль в зародышеобразовании, что подтверждается моделированием и анализом значений m . Однако роль L79 может быть недооценена при анализе значений φ из-за контактов в денатурированном состоянии. Из приведенных выше экспериментальных, симуляционных и эволюционных данных мы получаем четкую и взаимно непротиворечивую картину TSE, которая позволяет нам исследовать упорядоченную последовательность событий до и после нуклеации и, таким образом, полностью описать процесс складывания S6.
Съемка подъемов и спусков в фолдинге. Р Анализ кратности позволяет классифицировать каждую конформацию как структуру до-TS, после-TS или TS, из чего мы можем сделать вывод о порядке событий при складывании. Для этого была отдельно собрана и нормализована контактная статистика для p кратность < 0,4 и > 0,6 (до и после ТГЭ) структуры.Для определения контактов, преобладающих в каждом состоянии, и исключения влияния контактов, общих для всех состояний, была рассчитана и построена разностная матрица контактов (пост- и до-ТСЭ) (рис. 2 c ).
). Результаты этого анализа можно сравнить с результатами непрерывных экспериментов по значению φ, которые исследуют порядок событий сворачивания путем смещения TSE путем нарушения стабильности белка (37). Эти изменения значений φ строго объясняются постулатом Хаммонда (38), который легко применим к белкам (39, 40).Такой анализ был впервые проведен на U1A (37), а затем проведен на CI2 (35) и S6 (11) путем измерения сдвигов кинетических м — и φ-значений в ответ на изменения денатурирующего вещества [хлорида гуанидиния ( GdmCl)] концентрации.
Для этого была отдельно собрана и нормализована контактная статистика для p кратность < 0,4 и > 0,6 (до и после ТГЭ) структуры.Для определения контактов, преобладающих в каждом состоянии, и исключения влияния контактов, общих для всех состояний, была рассчитана и построена разностная матрица контактов (пост- и до-ТСЭ) (рис. 2 c ).
). Результаты этого анализа можно сравнить с результатами непрерывных экспериментов по значению φ, которые исследуют порядок событий сворачивания путем смещения TSE путем нарушения стабильности белка (37). Эти изменения значений φ строго объясняются постулатом Хаммонда (38), который легко применим к белкам (39, 40).Такой анализ был впервые проведен на U1A (37), а затем проведен на CI2 (35) и S6 (11) путем измерения сдвигов кинетических м — и φ-значений в ответ на изменения денатурирующего вещества [хлорида гуанидиния ( GdmCl)] концентрации.
Было высказано предположение, что мутации, уменьшающие m u и, следовательно, демонстрируют более высокие значения φ при 7 M GdmCl, вносят свой основной энергетический вклад после складчатого барьера (11). В S6 такие остатки (Y4, V37, V88 и V90) указывают на увеличение структуры спирали 2 и С-концевой цепи после зародышеобразования.Эта же тенденция отражена на рис. 2 c . Эксперимент также предполагает, что формирование интерфейса между N- и C-концами (высокие значения φ при 7 M GdmCl для Y4, V88 и V90) происходит поздно при складывании. Наши результаты показывают, что N- и C-концы и контакты между нитями 1 и 3 консолидируются в основном после зародышеобразования. Эксперимент предполагает, что в средней точке перехода нити 2 и 3 соединяются, что также наблюдается при моделировании. Фактически разные контакты в цепях 2 и 3 наблюдаются в структурах до и после TSE, что указывает на то, что этот процесс охватывает переходную область.На этот результат указывает рис. 2, на котором показана структурированная область нити 2–3. Рис. 3 б показывает критическую роль остатков, образующих нелокальные контакты, включая те, которые образуют L48, F60, Q57 и W62, которые не были идентифицированы в эксперименте.
В S6 такие остатки (Y4, V37, V88 и V90) указывают на увеличение структуры спирали 2 и С-концевой цепи после зародышеобразования.Эта же тенденция отражена на рис. 2 c . Эксперимент также предполагает, что формирование интерфейса между N- и C-концами (высокие значения φ при 7 M GdmCl для Y4, V88 и V90) происходит поздно при складывании. Наши результаты показывают, что N- и C-концы и контакты между нитями 1 и 3 консолидируются в основном после зародышеобразования. Эксперимент предполагает, что в средней точке перехода нити 2 и 3 соединяются, что также наблюдается при моделировании. Фактически разные контакты в цепях 2 и 3 наблюдаются в структурах до и после TSE, что указывает на то, что этот процесс охватывает переходную область.На этот результат указывает рис. 2, на котором показана структурированная область нити 2–3. Рис. 3 б показывает критическую роль остатков, образующих нелокальные контакты, включая те, которые образуют L48, F60, Q57 и W62, которые не были идентифицированы в эксперименте. Стыковка нити 3 со спиралью 2, по-видимому, важна для нуклеации как из-за высоких φ exp (V65, M67 и L75), так и из-за большого количества нелокальных контактов в TS (особенно L75). Как предполагалось ранее (11), остатки с увеличением м Значения f , по-видимому, участвуют в структуре перед переходным барьером в наших симуляциях.Эти положения (V6, L19, L30, L61 и L65) находятся в спирали 1 и цепи 3.
Стыковка нити 3 со спиралью 2, по-видимому, важна для нуклеации как из-за высоких φ exp (V65, M67 и L75), так и из-за большого количества нелокальных контактов в TS (особенно L75). Как предполагалось ранее (11), остатки с увеличением м Значения f , по-видимому, участвуют в структуре перед переходным барьером в наших симуляциях.Эти положения (V6, L19, L30, L61 и L65) находятся в спирали 1 и цепи 3.
Поскольку конформации до и после TS подвергаются тем же ограничениям φ exp , что и TSE, они заметно не различаются по количеству контактов, Rg или drms (таблица 2), как можно было бы ожидать для структуры раньше или позже складываются. Хотя φ exp предполагает в целом более расширенный TS при 0M, чем при 7M GdmCl, по вышеуказанным причинам наше моделирование этого не делает.Когда в моделировании использовались φ 0M и φ 7M , они давали ансамбли с p раз ≈ 0 и ≈ 1 соответственно. Хотя значения φ обеспечивают меру структуры TS, из-за различий в экспериментальных условиях складывания (0 или 7 M) не следует ожидать одинакового TSE. Действительно, из-за различий в значениях φ мы знаем, что TSE, исследуемые в разных условиях, различаются. Таким образом, логично, что эти ТГЭ проявляют разные p кратно при повторном складывании в разных условиях.Тем не менее, поскольку p кратность является хорошей координатой реакции для фолдинга белка (29), мы можем узнать упорядоченные детали фолдинга из структур, полученных из средней точки φ . Это открытие демонстрирует доминирующую роль и надежность зародышеобразующих контактов и общую пластичность других контактов. Сравнение интерпретаций с м ф , м u , φ 0M и φ 7M с нашими конформациями до и после TS подтверждают интерпретацию манипуляций с растворителем как эффективного способа исследовать сдвиги в TSE и, следовательно, порядок событий при складывании.
Хотя значения φ обеспечивают меру структуры TS, из-за различий в экспериментальных условиях складывания (0 или 7 M) не следует ожидать одинакового TSE. Действительно, из-за различий в значениях φ мы знаем, что TSE, исследуемые в разных условиях, различаются. Таким образом, логично, что эти ТГЭ проявляют разные p кратно при повторном складывании в разных условиях.Тем не менее, поскольку p кратность является хорошей координатой реакции для фолдинга белка (29), мы можем узнать упорядоченные детали фолдинга из структур, полученных из средней точки φ . Это открытие демонстрирует доминирующую роль и надежность зародышеобразующих контактов и общую пластичность других контактов. Сравнение интерпретаций с м ф , м u , φ 0M и φ 7M с нашими конформациями до и после TS подтверждают интерпретацию манипуляций с растворителем как эффективного способа исследовать сдвиги в TSE и, следовательно, порядок событий при складывании.
Синергия моделирования, эксперимента и эволюционных данных дает более глубокое понимание свертывания S6. Путем моделирования мы построили на атомном уровне картину зарождающихся контактов и порядок событий складывания в S6, которая согласуется с обширным массивом эволюционных и экспериментальных данных и расширяет их интерпретацию. S6 сворачивается по механизму зародышеобразования-конденсации, при этом нить 1 и спираль 1 соединяются вместе посредством контактов дальнего действия внутри кластера V6-I8-I26-L30.Это специфическое ядро стабилизировано V65 и L75 в цепи 3 и спирали 2. Большинство других остатков, которые кажутся важными только в одном или двух из вышеперечисленных методов, расположены в гидрофобном ядре (L10, P12, Y33, V37, Y63, M67, и V72). Также имеется интересная группа из трех остатков в петле большой нити 2–3. Остаток F60, который демонстрирует высокое значение φ exp и образует большое количество нелокальных контактов, граничит с L48 и I52, которые, по-видимому, закрепляют петлевую область посредством поздно формирующихся контактов с V9. Эта петля играет важную роль в формировании рибосомы, где она связывает белок S18 (10) и, таким образом, может иметь функциональные преимущества, будучи независимой от единицы кооперативного фолдинга. Белок S6 представляет собой пример классического поведения нуклеации-конденсации (18). При образовании специфического ядра (энтропийного барьера) сворачивание идет вниз энергетически за счет конденсации пластичного, гидрофобного облака опорных контактов.
Эта петля играет важную роль в формировании рибосомы, где она связывает белок S18 (10) и, таким образом, может иметь функциональные преимущества, будучи независимой от единицы кооперативного фолдинга. Белок S6 представляет собой пример классического поведения нуклеации-конденсации (18). При образовании специфического ядра (энтропийного барьера) сворачивание идет вниз энергетически за счет конденсации пластичного, гидрофобного облака опорных контактов.
Порядок складывания событий, предложенный экспериментом, проясняется с помощью моделирования.Сначала образуются контакты между нитью 1 и спиралью 1, при этом зародышеобразование сосредоточено на образовании кластера V6-I8-I26-L30. V65 и L75, которые отделены от этого кластера по последовательности, но близки по структуре, также, по-видимому, вовлечены в ядро. Хотя эксперимент еще не определил степень вторичной структуры в спирали 1, моделирование показывает, что она хорошо сформирована до зародышеобразования. Большая петля, разделяющая нити 2 и 3, замыкается вокруг F60, L48 и I52, стыкуясь с V9 после зародышеобразования. Спираль 2 образует вторичную структуру и стыкуется с нитью 1 в основном после зародышеобразования. Нити 1 и 4 также объединяются после зародышеобразования. В дополнение к тому, что эти позднеформирующиеся контакты неблагоприятны с точки зрения энтропии, они взаимодействуют как с РНК, так и с белком S18 в рибосоме (10) и могут получить функциональное преимущество благодаря такой конформационной гибкости.
Спираль 2 образует вторичную структуру и стыкуется с нитью 1 в основном после зародышеобразования. Нити 1 и 4 также объединяются после зародышеобразования. В дополнение к тому, что эти позднеформирующиеся контакты неблагоприятны с точки зрения энтропии, они взаимодействуют как с РНК, так и с белком S18 в рибосоме (10) и могут получить функциональное преимущество благодаря такой конформационной гибкости.
Как мы наблюдаем в р кратность симуляций, при зарождении, если структура выходит за пределы переходного барьера, она подвергается полному и быстрому складыванию.Хотя большинство φ exp имеют промежуточные значения в S6, предполагая, что TS является однородно свернутым, конформации TS демонстрируют различные области порядка и беспорядка в их вторичной и третичной структуре. Снижен экспериментальный м Значения и вокруг равновесной переходной области указывают на возможность частичного разворачивания нативного состояния (11). Последовательно наши симуляции предполагают, что такие флуктуации происходят вне кооперативной единицы, в цепях 1 и 4 и в петле цепей 2-3.В этом смысле TSE можно охарактеризовать как «диффузный» в том смысле, что он демонстрирует в значительной степени разрушенный остов. Однако это не просто расширенная версия исходной топологии. Специфическое зародышеобразование определяет топологию TSE и устанавливает основу для формирования критического контакта. После формирования этой кооперативной единицы структура стремится к складыванию.
Последовательно наши симуляции предполагают, что такие флуктуации происходят вне кооперативной единицы, в цепях 1 и 4 и в петле цепей 2-3.В этом смысле TSE можно охарактеризовать как «диффузный» в том смысле, что он демонстрирует в значительной степени разрушенный остов. Однако это не просто расширенная версия исходной топологии. Специфическое зародышеобразование определяет топологию TSE и устанавливает основу для формирования критического контакта. После формирования этой кооперативной единицы структура стремится к складыванию.
Специфическое ядро представляет собой фиксированный набор контактов, общих для всех конформаций TSE. Другие поддерживающие контакты внутри критического ядра в известной степени неоднородны.Принимая во внимание, что некоторые из этих поддерживающих контактов могут быть сформированы в одном подмножестве TSE ( p раз = 0,5) конформации, другие подмножества конформаций TSE могут иметь другие поддерживающие контакты. В этом смысле гетерогенное облако гидрофобных контактов, поддерживающих ядро, изменчиво или пластично. Существует также пластичность контактов вне кооперативной единицы (например, область петли 2–3 нити), которая, в принципе, может быть достигнута за счет флуктуаций основного (нативного) состояния или локального развертывания.Эти области представляют собой отклонения от глобальной кооперативности, но не от термодинамического поведения двух состояний. Эта наблюдаемая пластичность вне кооперативной единицы может быть предположительно приписана функции связывания РНК и белка в S6. Однако подобные области в других белках могут быть вовлечены в ошибочные побочные реакции, такие как агрегация.
В этом смысле гетерогенное облако гидрофобных контактов, поддерживающих ядро, изменчиво или пластично. Существует также пластичность контактов вне кооперативной единицы (например, область петли 2–3 нити), которая, в принципе, может быть достигнута за счет флуктуаций основного (нативного) состояния или локального развертывания.Эти области представляют собой отклонения от глобальной кооперативности, но не от термодинамического поведения двух состояний. Эта наблюдаемая пластичность вне кооперативной единицы может быть предположительно приписана функции связывания РНК и белка в S6. Однако подобные области в других белках могут быть вовлечены в ошибочные побочные реакции, такие как агрегация.
Выводы. Мы построили модель TSE и эволюционного анализа CoC на атомном уровне, чтобы получить более глубокое понимание укладки, исследуя роль остатков, недоступных для эксперимента.Порядок событий свертывания, который мы выводим из моделирования, коррелирует и обеспечивает поддержку интерпретации экспериментов, которые изменяют значения φ путем модуляции [GdmCl]. Эти данные также представляют убедительные доказательства того, что механистические детали в нашей модели MC-Go реалистичны и, таким образом, позволяют провести детальную структурную интерпретацию φ exp . Простого изучения φ exp недостаточно для полного понимания нуклеации и фолдинга S6 или других белков. Только благодаря синергии эксперимента и моделирования можно построить подробную и полную модель.Самое главное, мы наблюдаем, что, пока кооперативная единица, складчатое ядро, формируется в TSE, другие части структуры могут флуктуировать. В связи с этим конформации вдоль пути укладки проявляют структурную пластичность. Для S6, который связывается с РНК и белком после фолдинга, эта пластичность может быть прямым следствием конформационной гибкости, необходимой для достижения биологической функции.
Эти данные также представляют убедительные доказательства того, что механистические детали в нашей модели MC-Go реалистичны и, таким образом, позволяют провести детальную структурную интерпретацию φ exp . Простого изучения φ exp недостаточно для полного понимания нуклеации и фолдинга S6 или других белков. Только благодаря синергии эксперимента и моделирования можно построить подробную и полную модель.Самое главное, мы наблюдаем, что, пока кооперативная единица, складчатое ядро, формируется в TSE, другие части структуры могут флуктуировать. В связи с этим конформации вдоль пути укладки проявляют структурную пластичность. Для S6, который связывается с РНК и белком после фолдинга, эта пластичность может быть прямым следствием конформационной гибкости, необходимой для достижения биологической функции.
%PDF-1.4 % 52 0 объект > эндообъект 106 0 объект >поток заявка/pdf
2004-05-21T09:45:53ZXPP2022-01-10T00:55:34-08:002022-01-10T00:55:34-08:00StampPDF Пакет 2. 7 для Solaris — SPDF 1045uuid:537f3f1d-1dd2-11b2-0a00-d20827fd5800uuid:537f3f24-1dd2-11b2-0a00-6a0000000000
конечный поток
эндообъект
50 0 объект
>
эндообъект
48 0 объект
>
эндообъект
53 0 объект
>/Font>/ProcSet[/PDF/Text/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
1 0 объект
>/Font>/ProcSet[/PDF/Text/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
14 0 объект
>/Font>/ProcSet[/PDF/Text/ImageB/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
21 0 объект
>/Font>/ProcSet[/PDF/Text/ImageB/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
27 0 объект
>/Font>/ProcSet[/PDF/Text/ImageB/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
33 0 объект
>/Font>/ProcSet[/PDF/Text/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
118 0 объект
>поток
HWYo~ׯH6o>N2f50lI%){_j,bW?VU˽[aU`?>*»?`wQyq३7aP ~C;8/]
/Qx(zF.(
7 для Solaris — SPDF 1045uuid:537f3f1d-1dd2-11b2-0a00-d20827fd5800uuid:537f3f24-1dd2-11b2-0a00-6a0000000000
конечный поток
эндообъект
50 0 объект
>
эндообъект
48 0 объект
>
эндообъект
53 0 объект
>/Font>/ProcSet[/PDF/Text/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
1 0 объект
>/Font>/ProcSet[/PDF/Text/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
14 0 объект
>/Font>/ProcSet[/PDF/Text/ImageB/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
21 0 объект
>/Font>/ProcSet[/PDF/Text/ImageB/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
27 0 объект
>/Font>/ProcSet[/PDF/Text/ImageB/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
33 0 объект
>/Font>/ProcSet[/PDF/Text/ImageC]/XObject>>>/Rotate 0/TrimBox[9 9 603 792]/Type/Page>>
эндообъект
118 0 объект
>поток
HWYo~ׯH6o>N2f50lI%){_j,bW?VU˽[aU`?>*»?`wQyq३7aP ~C;8/]
/Qx(zF.(Обучение с подкреплением — TD(λ) Введение(1) | Джереми Чжан
Применение offline-λ при случайном блуждании
В этой статье мы будем говорить о TD(λ), который является общим методом обучения с подкреплением который объединяет как моделирование Монте-Карло, так и 1-шаговый метод TD. Мы говорили о методе TD исчерпывающе, и, если вы помните, в методе TD (n), я сказал, что это также объединение моделирования MC и 1-шагового TD , но в TD(n) нужно отслеживать все трассы по пути и обновлять текущую оценку на основе значения на n шагов вперед, в TD(λ) мы увидим более элегантную унификацию.
Мы говорили о методе TD исчерпывающе, и, если вы помните, в методе TD (n), я сказал, что это также объединение моделирования MC и 1-шагового TD , но в TD(n) нужно отслеживать все трассы по пути и обновлять текущую оценку на основе значения на n шагов вперед, в TD(λ) мы увидим более элегантную унификацию.
В этой статье мы будем:
- Изучение идеи TD(λ)
- Знакомство с методом прямого просмотра — возврат offline-λ
- Применение метода на примере случайного блуждания
TD(λ) фактически является расширением метода TD(n), помните, что в TD(n) у нас есть накопленное вознаграждение в следующей форме:
Эта оценка значения до шага t+n используется для обновления значение на шаге t , а TD(λ) усредняет значение, например, используя
0.5*Gt:t+2 + 0,5*Gt:t+4
в качестве целевого значения. Но вместо того, чтобы использовать прямые веса, он использует λ в качестве параметра для управления весом и доводит его до 1:
TD(λ) Бьюсь об заклад, изображение выглядит знакомым, агент начинает с состояния, предпринимая действие, он достигает следующего состояния, после чего выбирает другое действие, и процесс SARSA продолжается и продолжается. (Tt-1) (T — длина эпизода).Обратите внимание, что вес уменьшается по мере увеличения n, и общая сумма равна 1. Более общая форма TD(λ):
(Tt-1) (T — длина эпизода).Обратите внимание, что вес уменьшается по мере увеличения n, и общая сумма равна 1. Более общая форма TD(λ):
Из формулы видно, что когда λ = 1 , сохраняется только последний член и по сути, это метод Монте-Карло, так как состояние, процесс действия идет до конца, когда λ = 0 , срок сокращается до G_t:t+1 , что является 1-шаговым методом TD, а для 0 < λ < 1 , метод становится смешанным методом взвешенного TD(n).
Определив цель G_t , мы переходим к определению нашего первого алгоритма.Обобщенная формула обновления может быть определена как:
Правило обновления такое же, как и общий полуградиентный метод, единственное отличие заключается в целевом значении, указанном выше.
Ссылаясь на изображение из книги Саттона, этот метод также называется алгоритмом обучения прямого просмотра , так как в каждом состоянии процесс обновления ожидает значения G_t:t+1 , G_t:t+2 , …, и на основе взвешенного значения, которое обновляет текущее состояние.
Теперь приступим к реализации алгоритма на примере случайного блуждания. Мы изучили пример случайного блуждания здесь, но, к вашему сведению, при случайном блуждании агент начинает со средней позиции, и на каждом шаге он имеет равную вероятность переместиться на один шаг либо влево, либо вправо (политика действий фиксирована), и, заканчиваясь только слева или на большинстве, он может остановить эпизод.
Мы собираемся реализовать случайное блуждание с 19 состояниями, и хотя пространство состояний на самом деле дискретно, мы все же можем применить к нему алгоритм обобщения.
Функция значения
Функция значения проста и явна. У нас есть 19 состояний и 2 конечных состояния, так что всего у нас есть 21 состояние, и каждое состояние имеет вес, который по сути является оценкой его значения. Функция value возвращает значение определенного состояния, а функция Learn обновляет текущую оценку на основе разницы delta , которая в данном случае равна Gt — v(St, wt) ( альфа — скорость обучения ).
Некоторые общие функции
Так как мы не впервые реализуем случайное блуждание, я перечислю некоторые часто используемые функции вместе:
В каждом состоянии агент дойти до конца игры.
Воспроизведение и обновление
Опять же, основная разница заключается в процессе воспроизведения и вычислении дельты для каждого состояния.
В каждом эпизоде нам нужно будет сначала отслеживать все состояния до конца игры, чтобы получить представление вперед при обновлении значения каждого состояния. self.states записывает все состояния на пути, а self.reward хранит только последнее вознаграждение, так как все вознаграждения равны 0, кроме конечного состояния.
Вторая часть — обновить оценку значения состояния после окончания игры.Вспомните формулу:
Для каждого состояния в момент времени t и шага n нам нужно вычислить значение G_t:t+n и объединить их с уменьшающимися весами, чтобы получить целевое значение Ст . Таким образом, функция
Таким образом, функция gt2tn вычисляет значение, заданное t и n , которое определено как:
и gtlambda в предыдущем фрагменте кода является целевым значением. Кроме того, мы также устанавливаем значение atruncate, когда lambda_power слишком мало, мы просто игнорируем это значение.С целью gtlambda и текущим значением из valueFunc мы можем вычислить разницу дельта и обновить оценку, используя функцию узнать , которую мы определили выше.
Помните, что в сеансе TD(n) мы применили n-шаговый метод TD для случайного блуждания с точно такими же настройками. Теперь, когда введен автономный λ-Return , давайте сравним результаты обучения обоих:
Мы видим, что обе кривые среднеквадратичной ошибки похожи, и результат сопоставим.Как правило, наилучший результат обучения обычно достигается при промежуточной скорости обучения и значении λ (я скопировал изображение из книги Саттона, так как оно более четкое, чем то, которое я нарисовал).
Вот и все для обновления просмотра вперед, пожалуйста, ознакомьтесь с полной реализацией здесь. В следующем посте мы изучим обратное обновление, которое является более элегантным методом TD(λ) с использованием трассировки приемлемости .
Справка :
Светлана Колтовская, аспирант кафедры TESL и прикладной лингвистики
Имя: Светлана Колтовская
Офис: 414 Morrill Hall
Электронная почта: skoltov@okstate.образование
Научные интересы
- Письмо L2
- Автоматизированная оценка письма (AWE)
- Изучение языков с помощью компьютера (CALL)
- Письменный корректирующий отзыв
- Межкультурная прагматика
Преподаваемые курсы
Ассистент преподавателя
- ENGL 4013: Грамматика английского языка (содержательный курс) – осень 2020 г./осень 2021 г.
- ENGL 3323: техническое письмо (синхронный и асинхронный онлайн) – осень 2020 г. , весна 2021 г., лето 2021 г., осень 2021 г.
- ENGL 1223: Международный состав первокурсников II - осень 2017 г., весна 2018 г., осень 2018 г.
- ENGL 5693 (ранее 4893): Написание научных исследований для иностранных аспирантов — весна 2019 г. / осень 2019 г.
 , весна 2021 г., лето 2021 г., осень 2021 г.
, весна 2021 г., лето 2021 г., осень 2021 г.Административные роли
- Помощник директора Международной композиторской программы – осень 2020 г.
- Многоязычный специалист Центра письма ОГУ - осень 2018/весна 2019/осень 2019
- Online Supervisor, письменный центр OSU — осень 2018/весна 2019/осень 2019
Аспирант-исследователь
- Научный сотрудник Кэрол Модер и Стефани М.Ссылка (Набор и удержание иностранных студентов в ОГУ; Проект реконструкции Института английского языка
Избранные публикации
Статьи в реферируемых журналах
- Колтовская, С . (2020). Вовлечение учащихся с помощью автоматизированной письменной корректирующей обратной связи (AWCF), предоставленной Grammarly: несколько тематических исследований. Оценка письма, 44 , 1-12. https://doi.org/10.1016/j.asw.2020.100450
 Оценка письма, 44 , 1-12. https://doi.org/10.1016/j.asw.2020.100450
Оценка письма, 44 , 1-12. https://doi.org/10.1016/j.asw.2020.100450 Обзоры книг
- Колтовская С. (ожидается). Интернационализация центра письма: руководство по развитию многоязычного центра письма, автор Норин Грувер Лэйп. Журнал центра письма, 38 (2)
Мультимедийные обзоры
Избранные презентации конференции
- Колтовская, д. (2021, 27 августа). Использование Grammarly учителями 2-го уровня в качестве дополнения к формирующей обратной связи . Документ представлен на EUROCALL, организованном CNAM, Париж.
- Колтовская, д. (2021, июнь). множественное тематическое исследование взаимодействия студентов с отзывами электронных оценщиков . Доклад, представленный на виртуальной конференции CALICO 2021.
- Гесс, Э., Колтовская, С., Такебе, А. (2021, 18 июня). Разговоры о расовой несправедливости через симуляции в международных классах композиции. Документ, представленный на виртуальной конференции Центра языковой и межкультурной коммуникации (CLIC) 2021 года в Университете Райса, Хьюстон, Техас.
- Колтовская, д. (2021, 22 мая). Использование Grammarly учителями 2-го уровня в качестве дополнения к формирующей обратной связи . Документ представлен на исследовательском семинаре L2WR, проведенном Университетом Мурсии, Испания.
- Колтовская, д. (2021, 17 апреля). Использование Grammarly учителями 2-го уровня в качестве дополнения к формирующей обратной связи . Документ, представленный на 6-м ежегодном весеннем форуме TESL и прикладной лингвистике в Университете штата Оклахома, Стилуотер, Оклахома.
- Колтовская, д. (2021, 3 марта). Эффективность исправления ошибок Grammarly. Доклад, представленный на семинаре «Укусы данных / Байты данных», организованном Лабораторией языковых исследований Университета штата Оклахома, Стиллуотер, Оклахома.
- Колтовская, д. (2020, 18 июля). Вовлечение учащихся с помощью автоматизированной письменной обратной связи: несколько тематических исследований . Документ, представленный на виртуальной конференции TESOL & English Language EXPO.
- Колтовская С. (2020, 18 апреля). Участие учащихся в автоматизированной обратной связи Grammarly . Документ представлен на 5-м ежегодном весеннем форуме TESL и прикладной лингвистике в Университете штата Оклахома, Стилуотер, Оклахома (отменен из-за COVID).
- Колтовская, С. (2020, 31 марта - 3 апреля). Вовлечение учащихся с помощью автоматизированной письменной обратной связи: несколько тематических исследований. Доклад принят для устного выступления на конференции TESOL в Денвере, штат Колорадо (отменен из-за COVID).
- Колтовская, д. (2019, 15 апреля). Грамматика. Демонстрация технологий на 4-м ежегодном Весеннем форуме TESL и прикладной лингвистики в Государственном университете Оклахомы, Стилуотер, Оклахома.
- Муранова О., Колтовская С., и Миллер М. (2019, 12 марта). Принятие предложений на онлайн-сессиях письменного центра. Статья представлена на Форуме докторантуры TESOL, Атланта, Джорджия.
- Муранова О., Колтовская С., Кэннон, Дж., и Миллер, М. (2019, 16 февраля). О способах улучшения понимания предложений и повышения эластичности сеансов онлайн-центра письма. Доклад, представленный на конференции SCWCA в Университете Мэри Хардинг-Бейлор, Белтон, Техас.
- Муранова О. и Колтовская С. (2017, ноябрь). Прагматические аспекты общения между агентами и многоязычными клиентами. Доклад, представленный на ежегодной конференции IWCA в Чикаго, штат Иллинойс.
- Муранова О. и Колтовская С. (2017, 1 апреля). От причудливого к гиковскому: роль национальных праздников в пробуждении мотивации учащихся ESL/EFL. Доклад, представленный на 2-м ежегодном весеннем форуме TESL и прикладной лингвистике в Университете штата Оклахома, Стилуотер, Оклахома.
- Колтовская С., и Похарел К. (2015, 12 апреля). Английский встречается с китайским и арабским: изучение акцентов в письме и структурной идентичности между языками .Документ представлен на конференции ECWCA в Университете Нотр-Дам, Нотр-Дам, Индиана.
- Колтовская С., и Похарел К. (2014). Эти неловкие моменты . Документ представлен на конференции MIWCA в Общественном колледже Лансинга, Лансинг, Мичиган.
- Дельгадо Б., Шакья Д. и Колтовская С. (2014, 25 октября). Консультанты ESL в центре письма? Доклад, представленный на конференции MIWCA в Государственном университете Гранд-Вэлли, Аллендейл, Мичиган.
 Документ, представленный на виртуальной конференции Центра языковой и межкультурной коммуникации (CLIC) 2021 года в Университете Райса, Хьюстон, Техас.
Документ, представленный на виртуальной конференции Центра языковой и межкультурной коммуникации (CLIC) 2021 года в Университете Райса, Хьюстон, Техас.


Профессиональная служба
В университет
- Координатор по вопросам академической честности — Управление по академическим вопросам, Государственный университет Оклахомы
- Координатор семинара по написанию диссертации – Высший колледж|Библиотека|Письменный центр, Университет штата Оклахома
- Руководитель кабинета экзамена International Teaching Assistant (ITA) — Graduate College, Университет штата Оклахома (2 раза)
- Специалист по языку (оценщик) экзамена International Teaching Assistant (ITA) - Graduate College, Университет штата Оклахома
- Фасилитатор семинара по написанию диссертации – Высшее учебное заведение|Библиотека|Письменный центр, Университет штата Оклахома
В программу
- Модератор и хронометрист 6-го ежегодного весеннего форума th по TESOL, лингвистике и прикладной лингвистике — кафедра английского языка, Университет штата Оклахома
- Соорганизатор семинара по симуляции для учителей письма L2
- Координатор семинара по рабочим материалам — кафедра английского языка, Университет штата Оклахома
- Ведущий инструктор курса последипломного образования (ENGL 5695 ) - Международная программа композиции, факультет английского языка, Государственный университет Оклахомы
- Соруководитель исследовательской группы многоязычных студентов - Центр письма, Государственный университет Оклахомы
- Group Mentor - Центр письма, Университет штата Оклахома
- Главный инструктор по международной композиции II (ENGL 1223 ) - Международная программа по композиции, Университет штата Оклахома
- Вице-президент - Студенческая организация TESLing, Университет штата Оклахома
- Сопредседатель программы — Весенний форум TESL и прикладной лингвистики, Университет штата Оклахома
- Главный редактор информационного бюллетеня - Студенческая организация TESLing, Университет штата Оклахома
Награды и признание
- Стипендия доктора философии Салли Хинрих, кафедра английского языка, Государственный университет Оклахомы, Стилуотер, Оклахома – весна 2021 г.
- Стипендия TESL/лингвистики, факультет английского языка, Государственный университет Оклахомы, Стилуотер, Оклахома – весна 2021 г.
- 2021 г. Стипендия Рут Краймс для обучения в аспирантуре, Международная ассоциация преподавания английского языка носителям других языков (TESOL) – осень 2020 г.
- 3-е место в 7-м заезде в предварительном раунде трехминутной диссертации, Высший колледж Университета штата Оклахома – осень 2020 г.
- Премия женского факультета за научные исследования, Государственный университет Оклахомы – весна 2020 г.
- Стипендия Houston-Truax-Wentz Travel Fellowship, Государственный университет Оклахомы (отменено из-за COVID) – весна 2020 г.
- Премия Рави Шери TESL за исследования, Государственный университет Оклахомы – весна 2017 г./весна 2019 г.
- Премия EGSA Research Travel Award, Государственный университет Оклахомы, Стилуотер, Оклахома – весна 2019 г. /весна 2020 г.
- Ричард Г.Ingham Scholarship (стипендия для выдающихся аспирантов), Государственный университет Оклахомы – осень 2016 г.

 /весна 2020 г.
/весна 2020 г.Текущие исследовательские проекты
Колтовская, С. (в разработке). Эффективность обнаружения/исправления ошибок Grammarly и использование Учителями письма L2 для дополнения их формирующей обратной связи.
Муранова О., Колтовская С., и Миллер М. (в разработке). Тематическое исследование по восприятию предложений на онлайн-сессиях центра синхронного письма.
Анализ последовательности информации о сегментах инициации укладки в ферредоксиноподобных белках укладки | BMC Structural Biology
Дилл К.А., МакКаллум Дж.Л.: Проблема сворачивания белков, 50 лет спустя. Наука 2012, 338: 1042–1046. 10.1126/наука.1219021
CAS Статья пабмед Google Scholar

Араи М., Ито К., Инобе Т., Накао М., Маки К., Камагата К., Кихара Х., Амемия Й., Куваджима К.: Быстрое уплотнение α-лактальбумина во время складывания изучено с помощью рентгенографии с остановленным потоком рассеяние. J Mol Biol 2002, 321: 121–132. 10.1016/S0022-2836(02)00566-1
КАС Статья пабмед Google Scholar
Nickson AA, Clarke J: Какие уроки можно извлечь из изучения фолдинга гомологичных белков? Методы 2010, 52: 38–50. 10.1016/j.ymeth.2010.06.003
PubMed Central КАС Статья пабмед Google Scholar
Nishimura C, Prytulla S, Dyson HJ, Wright PE: Сохранение путей укладки в эволюционно отдаленных последовательностях глобина. Nat Struct Biol 2000, 7: 679–686. 10.1038/77985
КАС Статья пабмед Google Scholar

Cavagnero S, Dyson HJ, Wright PE: Влияние дестабилизирующих мутаций H-спирали на кинетическую и равновесную укладку апомиоглобина. J Mol Biol 1999, 285: 269–282.10.1006/джмби.1998.2273
КАС Статья пабмед Google Scholar
Jennings PA, Wright PE: Формирование промежуточного расплавленного глобулы на ранней стадии кинетического пути сворачивания апомиоглобина. Наука 1993, 262: 892–896. 10.1126/наука.8235610
CAS Статья пабмед Google Scholar
Бернс Л.Л., Далессио П.М., Ропсон И.Дж.: Механизм складывания трех структурно сходных белков бета-листа. Белки 1998, 33: 107–118. 10.1002/(SICI)1097-0134(19981001)33:1<107::AID-PROT10>3.0.CO;2-P
CAS Статья пабмед Google Scholar
Kim DE, Fisher C, Baker D: Нарушение симметрии в переходном состоянии фолдинга белка L. J Mol Biol 2000, 298: 971–984. 10.1006/джмби.2000.3701
J Mol Biol 2000, 298: 971–984. 10.1006/джмби.2000.3701
КАС Статья пабмед Google Scholar
Park SH, Shastry MCR, Roder H: Динамика укладки домена B1 белка G, исследованная с помощью сверхбыстрого перемешивания. Nat Struct Biol 1999, 6: 943–947. 10.1038/13311
КАС Статья пабмед Google Scholar
Radford SE, Dobson CM, Evans PA: Сворачивание куриного лизоцима включает частично структурированные промежуточные соединения и несколько путей. Природа 1992, 358: 302–307.10.1038/358302а0
КАС Статья пабмед Google Scholar
Накамура Т., Макабе К., Томойори К., Маки К., Мукаяма А., Кувадзима К.: Различные пути укладки высоко гомологичных белков, козьего α-лактальбумина и лизоцима собачьего молока. Дж Мол Биол 2010, 396: 1361–1378. 10.1016/j.jmb.2010.01.021
Дж Мол Биол 2010, 396: 1361–1378. 10.1016/j.jmb.2010.01.021
КАС Статья пабмед Google Scholar
Hills RD Jr, Brooks CL III: Конкуренция поддоменов, кооперативность и топологическая фрустрация при складывании CheY. Дж Мол Биол 2008, 382: 485–495. 10.1016/j.jmb.2008.07.007
PubMed Central КАС Статья пабмед Google Scholar
Gangadhara BN, Laine JM, Kathuria SV, Massi F, Matthews CR: Кластеры разветвленных алифатических боковых цепей служат ядрами стабильности в нативном состоянии бочкообразного белка TIM HisF. Дж Мол Биол 2013, 425: 1065–1081. 10.1016/j.jmb.2013.01.002
PubMed Central КАС Статья пабмед Google Scholar
Hills RD Jr, Kathuria SV, Wallace LA, Day IJ, Brooks CL III, Matthews CR: Топологическое нарушение в белках с βα-повторами: разнообразие последовательностей модулирует консервативные механизмы фолдинга α/β/α сэндвич-белков . Дж Мол Биол 2010, 398: 332–350.10.1016/j.jmb.2010.03.001
Дж Мол Биол 2010, 398: 332–350.10.1016/j.jmb.2010.03.001
PubMed Central КАС Статья пабмед Google Scholar
Хаглунд Э., Линдберг М.О., Оливеберг М.: Изменения путей сворачивания белков путем круговой перестановки. Перекрывающиеся ядра способствуют глобальному сотрудничеству. J Biol Chem 2008, 283: 27904–27915. 10.1074/джбк.М801776200
КАС Статья пабмед Google Scholar
Тернстрем Т., Майор У., Акке М., Оливеберг М.: От снимка к фильму: фи-анализ переходных состояний фолдинга белка — еще один шаг вперед. Proc Natl Acad Sci USA 1999, 96: 14854–14859. 10.1073/pnas.96.26.14854
PubMed Central Статья пабмед Google Scholar
Villegas VV, Martínez JCJ, Avilés FXF, Serrano LL: Структура переходного состояния в процессе сворачивания домена активации прокарбоксипептидазы A2 человека. Дж Мол Биол 1998, 283: 1027–1036. 10.1006/джмби.1998.2158
Дж Мол Биол 1998, 283: 1027–1036. 10.1006/джмби.1998.2158
КАС Статья пабмед Google Scholar
Bernstein FC, Koetzle TF, Williams GJ, Meyer EF, Brice MD, Rodgers JR, Kennard O, Shimanouchi T, Tasumi M: The Protein Data Bank: компьютерный архивный файл макромолекулярных структур. J Mol Biol 1977, 112: 535–542. 10.1016/S0022-2836(77)80200-3
КАС Статья пабмед Google Scholar
Хамфри В., Далке А., Шультен К.: VMD: Визуальная молекулярная динамика. J Mol Graph 1996, 14: 33–38. 10.1016/0263-7855(96)00018-5
КАС Статья пабмед Google Scholar
Xi L, Li S, Liu H, Li J, Lei B, Yao X: Глобальное и локальное предсказание скорости фолдинга белка на основе информации об автокорреляции последовательностей. J Theor Biol 2010, 264: 1159–1168.10.1016/j.jtbi.2010.03.042
J Theor Biol 2010, 264: 1159–1168.10.1016/j.jtbi.2010.03.042
КАС Статья пабмед Google Scholar
Lin G, Wang Z, Xu D, Cheng J: SeqRate: классификация типов фолдинга белков на основе последовательностей и предсказание скоростей. БМС Биоинф 2010, 11: С1.
Артикул Google Scholar
Guo J-X, Rao N: Предсказание скорости фолдинга белка по аминокислотной последовательности. J Bioinform Comput Biol 2011, 09: 1–13.
КАС Статья Google Scholar
Gao J, Zhang T, Zhang H, Shen S, Ruan J, Kurgan L: Точное предсказание скорости фолдинга белка на основе последовательности и гибкости остатков, полученных из последовательности, и доступности растворителя. Белки 2010, 78: 2114–2130.
КАС пабмед Google Scholar
Ma B-G, Guo J-X, Zhang H-Y: Прямая корреляция между скоростью фолдинга белков и их аминокислотным составом: предсказание скорости фолдинга ab initio. Белки 2006, 65: 362–372. 10.1002/прот.21140
CAS Статья пабмед Google Scholar
Chang L, Wang J, Wang W: Эффективная длина цепи на основе состава для прогнозирования скорости фолдинга белка. Phys Rev E 2010, 82: 051930.
Артикул Google Scholar
Huang J-T, Tian J: Аминокислотная последовательность предсказывает скорость фолдинга белков среднего размера с двумя состояниями. Белки 2006, 63: 551–554. 10.1002/прот.20911
Артикул пабмед Google Scholar

Huang JT, Xing DJ, Huang W: Взаимосвязь между кинетикой сворачивания белков и свойствами аминокислот. Аминокислоты 2012, 43: 567–572. 10.1007/s00726-011-1189-3
КАС Статья пабмед Google Scholar
Calosci N, Chi CN, Richter B, Camilloni C, Engström A, Eklund L, Travaglini-Allocatelli C, Gianni S, Vendruscolo M, Jemth P: Сравнение последовательных переходных состояний для складывания показывает альтернативное раннее складывание пути двух гомологичных белков. Proc Natl Acad Sci USA 2008, 105: 19241–19246.10.1073/pnas.0804774105
PubMed Central КАС Статья пабмед Google Scholar
Fowler SB, Clarke J: Картирование пути сворачивания домена иммуноглобулина: структурные детали из анализа значений Phi и движения переходного состояния. Структура 2001, 9: 355–366. 10.1016/S0969-2126(01)00596-2
10.1016/S0969-2126(01)00596-2
КАС Статья пабмед Google Scholar
Мурзин А.Г., Бреннер С.Е., Хаббард Т., Чотия С.: SCOP: база данных структурной классификации белков для исследования последовательностей и структур. J Mol Biol 1995, 247: 536–540.
КАС пабмед Google Scholar
Kikuchi T, Némethy G, Scheraga HA: Предсказание расположения структурных доменов в глобулярных белках. J Protein Chem 1988, 7: 427–471.10.1007/БФ01024890
КАС Статья пабмед Google Scholar
Kikuchi T: Анализ трехмерных структурных различий в IgG-связывающих доменах на основе статистики среднего расстояния между остатками. Аминокислоты 2008, 35: 541–549. 10.1007/s00726-008-0082-1
КАС Статья пабмед Google Scholar

Nakajima S, Kikuchi T: Анализ различий в механизмах сворачивания лизоцимов с-типа на основе карт контактов, построенных с учетом средних расстояний между остатками. Дж Мол Модель 2007, 13: 587–594. 10.1007/s00894-007-0185-8
КАС Статья пабмед Google Scholar
Ichimaru T, Kikuchi T: Анализ различий в кинетике фолдинга структурно-гомологических белков на основе прогнозов грубых признаков контактов остатков. Белки 2003, 51: 515–530. 10.1002/прот.10378
CAS Статья пабмед Google Scholar
Kawai Y, Matsuoka M, Kikuchi T: Анализ белковых последовательностей с использованием статистики среднего расстояния между остатками для изучения процессов сворачивания и значимости их частичных последовательностей. Protein Pept Lett 2011, 18: 979–990. 10.2174/0
6511107010979
КАС Статья пабмед Google Scholar

Matsuoka M, Kabata M, Kawai Y, Kikuchi T: Анализ последовательностей (β/α) бочкообразных белков на основе статистики среднего расстояния между остатками для выяснения процессов сворачивания. В Химическая биология . Риека, Хорватия: InTech; 2012: 83–98.
Google Scholar
Oubridge C, Ito N, Evans PR, Teo CH, Nagai K: Кристаллическая структура при разрешении 1,92 Å РНК-связывающего домена сплайсосомного белка U1A в комплексе со шпилькой РНК. Природа 1994, 372: 432–438. 10.1038/372432а0
КАС Статья пабмед Google Scholar
Хименес М.А., Вильегас В., Санторо Дж., Серрано Л., Вендрелл Дж., Авилес Ф.Х., Рико М.: ЯМР-структура раствора домена активации прокарбоксипептидазы человека А2. Protein Sci 2003, 12: 296–305. 10.1110/ps.0227303
PubMed Central Статья пабмед Google Scholar
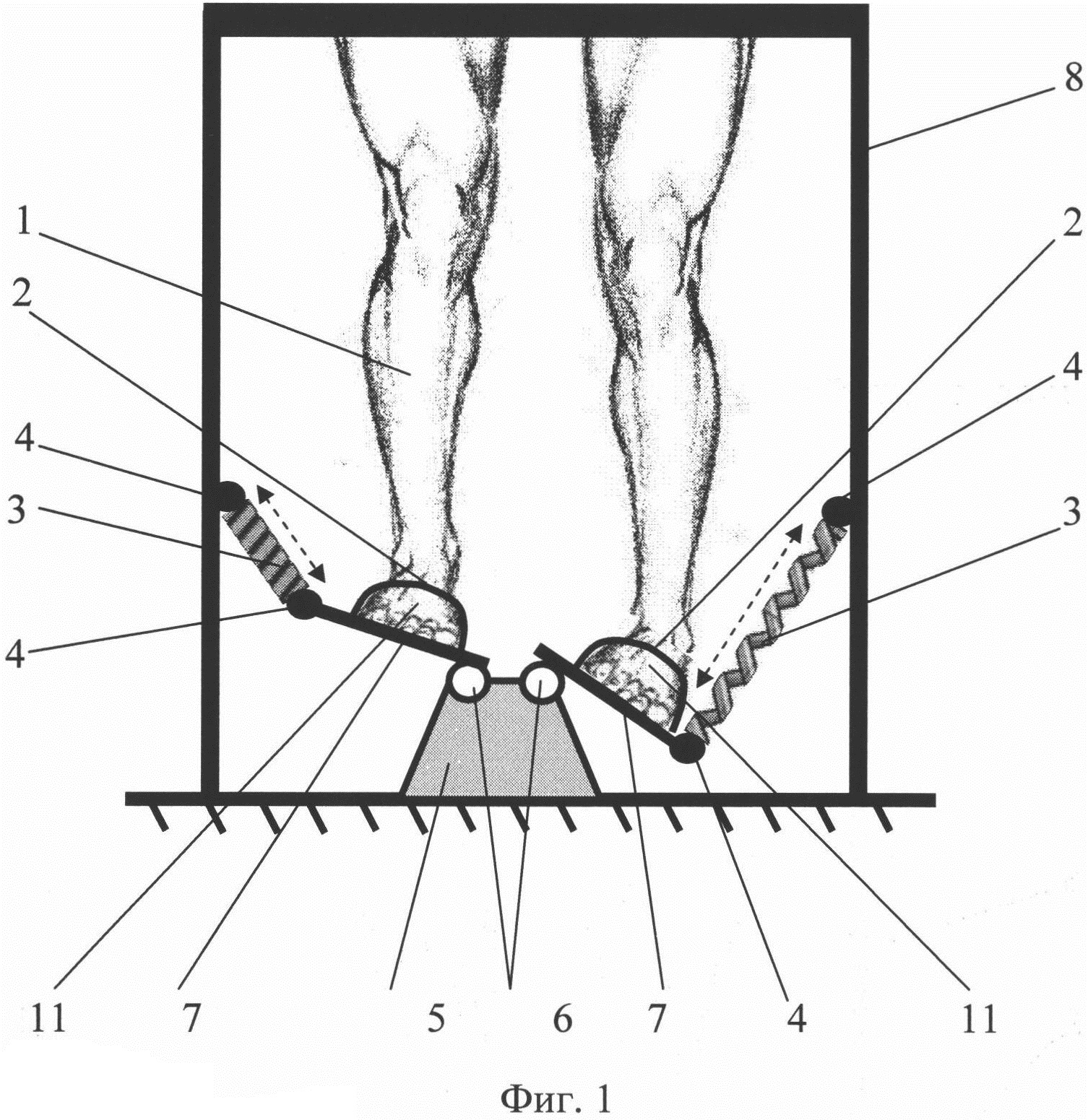
Линдал М., Свенссон Л.А., Лильяс А., Седельникова С.Е., Елисейкина И.А., Фоменкова Н.П., Невская Н., Никонов С.В., Гарбер М.Б., Муранова Т.А.: Кристаллическая структура рибосомного белка S6 из Thermus thermophilus. EMBO J 1994, 13: 1249–1254.
Центральный пабмед КАС пабмед Google Scholar
Пасторе А., Саудек В., Рампони Г., Уильямс Р.Дж.: Трехмерная структура ацилфосфатазы.Уточнение и структурный анализ. J Mol Biol 1992, 224: 427–440. 10.1016/0022-2836(92)
-АКАС Статья пабмед Google Scholar
Fulton KF, Bate MA, Faux NG, Mahmood K, Betts C, Buckle AM: База данных по фолдингу белков (PFD 2.0): онлайн-среда для Международного консорциума по фолдемике. Nucleic Acids Res 2007, 35: D304-D307.10.1093/nar/gkl1007
Центр PubMed КАС Статья пабмед Google Scholar
Chiti F, Taddei N, White PM, Bucciantini M, Magherini F, Stefani M, Dobson CM: Мутационный анализ ацилфосфатазы предполагает важность топологии и порядка контактов при сворачивании белка. Nat Struct Biol 1999, 6: 1005–1009. 10.1038/14890
КАС Статья пабмед Google Scholar
Kikuchi T: Расшифровка аминокислотных последовательностей белков с использованием статистики средних расстояний между остатками для извлечения информации о механизмах сворачивания белков. В Сворачивание белков . Под редакцией: Уолтерс ЕС. Нью-Йорк, США: Nova Science Publishers Inc.; 2011: 465–487.
Google Scholar
Hills RD, Brooks CL: Коэволюция функции и складчатого ландшафта: корреляция с плотностью местных контактов. Biophys J 2008, 95: L57-L59. 10.1529/biophysj.108.143388
PubMed Central КАС Статья пабмед Google Scholar

Miyazawa S, Jernigan RL: Потенциалы остатка-остатка с благоприятным термином контактной пары и неблагоприятным термином высокой плотности упаковки, для моделирования и нарезания резьбы. J Mol Biol 1996, 256: 623–644. 10.1006/джмби.1996.0114
КАС Статья пабмед Google Scholar
Skolnick J: В поисках эмпирического потенциала для предсказания структуры белка. Curr Opin Struct Biol 2006, 16: 166–171. 10.1016/j.sbi.2006.02.004
CAS Статья пабмед Google Scholar
Ishizuka Y, Kikuchi T: Анализ локальных последовательностей сайтов укладки в сэндвич-белках со статистикой среднего расстояния между остатками. Open Bioinform J 2011, 5: 59–68.10.2174/1875036201105010059
КАС Статья Google Scholar
Dasmeh P, Serohijos AWR, Kepp KP, Shahnovich EI: Положительно выбранные сайты в миоглобинах китообразных способствуют стабильности белка. PLoS Comput Biol 2013, 9: e1002929. 10.1371/journal.pcbi.1002929
PubMed Central КАС Статья пабмед Google Scholar
Мирный Л., Шахнович Э.: Эволюционная консервация ядра складчатости. J Mol Biol 2001, 308: 123–129. 10.1006/джмби.2001.4602
КАС Статья пабмед Google Scholar
Rorick MM, Wagner GP: Структурная модульность и надежность белков связаны с эволюционируемостью. Геном Биол Эвол 2011, 3: 456–475. 10.1093/gbe/evr046
Центр PubMed КАС Статья пабмед Google Scholar
Liao H, Yeh W, Chiang D, Jernigan RL, Lustig B: Энтропия белковой последовательности тесно связана с плотностью упаковки и гидрофобностью. Protein Eng Des Sel 2005, 18: 59–64. 10.1093/белок/gzi009
Protein Eng Des Sel 2005, 18: 59–64. 10.1093/белок/gzi009
PubMed Central КАС Статья пабмед Google Scholar
Ting K-LH, Jernigan RL: Определение складчатого ядра для семейства лизоцим/альфа-лактальбумин из кластеров сохранения последовательности. J Mol Evol 2002, 54: 425–436. 10.1007/s00239-001-0033-x
CAS Статья пабмед Google Scholar
Птицын О.Б., Тинг К.Л.: Нефункциональные консервативные остатки в глобинах и их возможная роль в качестве укладочного ядра. J Mol Biol 1999, 291: 671–682. 10.1006/джмби.1999.2920
КАС Статья пабмед Google Scholar
Альтшул С.Ф., Гиш В., Миллер В., Майерс Э.В., Липман Д.Дж.: Базовый инструмент локального поиска выравнивания. Дж Мол Биол 1990, 215: 403–410. 10.1016/S0022-2836(05)80360-2
10.1016/S0022-2836(05)80360-2
КАС Статья пабмед Google Scholar
Edgar RC: MUSCLE: множественное выравнивание последовательностей с высокой точностью и высокой производительностью. Nucleic Acids Res 2004, 32: 1792–1797. 10.1093/nar/gkh440
PubMed Central КАС Статья пабмед Google Scholar
Saitou N, Nei M: Метод объединения соседей: новый метод реконструкции филогенетических деревьев. Мол Биол Эвол 1987, 4: 406–425.
КАС пабмед Google Scholar
Ян Z: PAML 4: филогенетический анализ методом максимального правдоподобия. Мол Биол Эвол 2007, 24: 1586–1591. 10.1093/молбев/msm088
CAS Статья пабмед Google Scholar
Джонс Д.Т., Тейлор В.Р., Торнтон Дж.М.: Быстрое создание матриц данных о мутациях из белковых последовательностей. Comput Appl Biosci 1992, 8: 275–282.
КАС пабмед Google Scholar
Shrake A, Rupley JA: Окружающая среда и воздействие растворителя белковых атомов. Лизоцим и инсулин. J Mol Biol 1973, 79: 351–371. 10.1016/0022-2836(73)
-9КАС Статья пабмед Google Scholar
Матушек А., Келлис Дж. Т. мл., Серрано Л., Фершт А. Р.: Картирование переходного состояния и путей сворачивания белка с помощью белковой инженерии. Природа 1989, 340: 122–126. 10.1038/340122а0
КАС Статья пабмед Google Scholar
Hamill SJ, Steward A, Clarke J: Сворачивание иммуноглобулиноподобного греческого ключевого белка определяется ядром с общим ядром и областями, ограниченными топологией. Дж Мол Биол 2000, 297: 165–178. 10.1006/джмби.2000.3517
Дж Мол Биол 2000, 297: 165–178. 10.1006/джмби.2000.3517
КАС Статья пабмед Google Scholar
Bemporad F: Исследования фолдинга и агрегации в ацилфосфатазоподобном семействе . Флоренция, Италия: Издательство Флорентийского университета; 2009.
Google Scholar
Абкевич В.И., Гутин А.М., Шахнович Е.И.: Специфическое ядро как переходное состояние для сворачивания белков: данные решеточной модели. Биохимия 1994, 33: 10026–10036. 10.1021/би00199а029
КАС Статья пабмед Google Scholar
Faísca PFN, Travasso RDM, Ball RC, Shahnovich EI: Идентификация критических остатков в сворачивании белка: выводы из φ-значения и анализа P[subfold]. J Chem Phys 2008, 129: 095108. 10.1063/1.2973624
PubMed Central Статья пабмед Google Scholar

Faísca PFN: Механизм зародышеобразования при сворачивании белков: обзор исследований компьютерного моделирования. J Phys Condens Matter 2009, 21: 373102. 10.1088/0953-8984/21/37/373102
Статья пабмед Google Scholar
Parzen E: Об оценке функции и моды плотности вероятности. Ann Math Stat 1962, 33: 1065–1076. 10.1214/aoms/1177704472
Артикул Google Scholar
Линдберг М.О., Оливеберг М.: Гибкость путей сворачивания белка: простая причина сложного поведения. Curr Opin Struct Biol 2007, 17: 21–29. 10.1016/j.sbi.2007.01.008
CAS Статья пабмед Google Scholar
Truong HH, Kim BL, Schafer NP, Wolynes PG: Направление и разочарование в энергетических ландшафтах некоторых разработанных и упрощенных белков. J Chem Phys 2013, 139: 121908.10.1063/1.4813504
J Chem Phys 2013, 139: 121908.10.1063/1.4813504
Центральный пабмед Статья пабмед Google Scholar
Selvaraj S, Gromiha MM: Важность образования гидрофобных кластеров посредством контактов дальнего действия в переходном состоянии фолдинга белков с двумя состояниями. Белки 2004, 55: 1023–1035. 10.1002/прот.20109
CAS Статья пабмед Google Scholar
Parrini C, Bemporad F, Baroncelli A, Gianni S, Travaglini-Allocatelli C, Kohn JE, Ramazzotti M, Chiti F, Taddei N: Процесс сворачивания ацилфосфатазы из Escherichia coli заметно ускоряется благодаря наличию дисульфидной связи. J Mol Biol 2008, 379: 1107–1118. 10.1016/j.jmb.2008.04.051
КАС Статья пабмед Google Scholar
Taddei N, Capanni C, Chiti F, Stefani M, Dobson CM, Ramponi G: На сворачивание и агрегацию выборочно влияют конформационные предпочтения альфа-спиралей мышечной ацилфосфатазы. J Biol Chem 2001, 276: 37149–37154. 10.1074/джбк.М105720200
J Biol Chem 2001, 276: 37149–37154. 10.1074/джбк.М105720200
КАС Статья пабмед Google Scholar
Taddei N, Chiti F, Fiaschi T, Bucciantini M, Capanni C, Stefani M, Serrano L, Dobson CM, Ramponi G: Стабилизация альфа-спиралей с помощью сайт-направленного мутагенеза показывает важность вторичной структуры в переходном состоянии для сворачивания ацилфосфатазы. Дж Мол Биол 2000, 300: 633–647.10.1006/джмби.2000.3870
КАС Статья пабмед Google Scholar
Paci E, Vendruscolo M, Dobson CM, Karplus M: Определение переходного состояния с атомным разрешением на основе данных белковой инженерии. J Mol Biol 2002, 324: 151–163. 10.1016/S0022-2836(02)00944-0
КАС Статья пабмед Google Scholar
Арад-Хаасе Г. , Чуартцман С.Г., Даган С., Нево Р., Коуза М., Май Б.К., Нгуен Х.Т., Ли М.С., Райх З.: Механическое разворачивание ацилфосфатазы изучено с помощью силовой спектроскопии одиночных молекул и моделирования МД. Biophys J 2010, 99: 238–247. 10.1016/j.bpj.2010.04.004
, Чуартцман С.Г., Даган С., Нево Р., Коуза М., Май Б.К., Нгуен Х.Т., Ли М.С., Райх З.: Механическое разворачивание ацилфосфатазы изучено с помощью силовой спектроскопии одиночных молекул и моделирования МД. Biophys J 2010, 99: 238–247. 10.1016/j.bpj.2010.04.004
PubMed Central КАС Статья пабмед Google Scholar
Sugita M, Kikuchi T: Анализ складчатых свойств ферредоксиноподобных складчатых белков с помощью крупнозернистой модели Gō: взаимосвязь между профилями свободной энергии и ядрами складок. Белки Структура Функция Биоинформ 2014, 82: 954–965. 10.1002/прот.24469
CAS Статья Google Scholar
Накадзима С., Альварес-Сальгадо Э., Кикучи Т., Арредондо-Питер Р.: Прогнозирование пути и кинетики складывания гемоглобинов растений с использованием метода карты среднего расстояния. Белки 2005, 61: 500–506. 10.1002/прот.20658
Белки 2005, 61: 500–506. 10.1002/прот.20658
CAS Статья пабмед Google Scholar
Замена рыбной муки растительным белком в рационе атлантического лосося (Salmo salar) путем добавления гидролизата рыбного белка
Hertrampf, J. W. & Piedad-Pascual, F. Справочник по ингредиентам для кормов для аквакультуры. (Springer Science & Business Media, 2012 г.).
Турчини, Г. М., Трушенски, Дж. Т. и Гленкросс, Б. Д. Мысли о будущем питания аквакультуры: переориентация перспектив с учетом современных проблем, связанных с разумным использованием морских ресурсов в кормах для аквакультуры. Североамериканский журнал аквакультуры 81 , 13–39, https://doi.org/10.1002/naaq.10067 (2019).
Артикул Google Scholar
Hardy, R. W. Использование растительных белков в рационах рыб: влияние глобального спроса и предложения рыбной муки.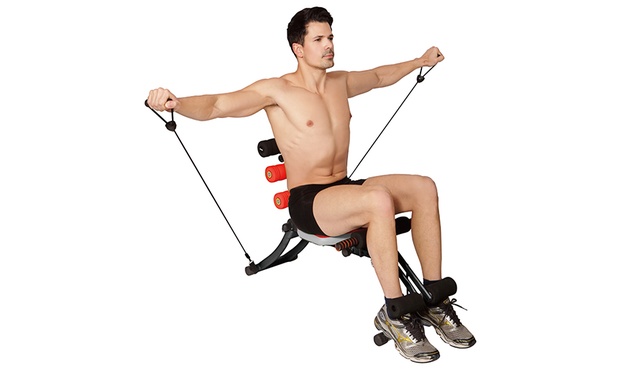 Исследования аквакультуры 41 , 770–776, https://doi.org/10.1111/j.1365-2109.2009.02349.x (2010).
Исследования аквакультуры 41 , 770–776, https://doi.org/10.1111/j.1365-2109.2009.02349.x (2010).
КАС Статья Google Scholar
Гахардо, К. и др. . Альтернативные источники белка в рационе модулируют микробиоту и функциональность дистального отдела кишечника атлантического лосося ( Salmo salar ). Прикладная и экологическая микробиология 83 , https://doi.org/10.1128/AEM.02615-16 (2017).
Коллинз, С.А., Оверланд, М., Скреде, А. и Дрю, М.Д. Влияние источников растительного белка на скорость роста лососевых: мета-анализ включения в рацион сои, гороха и шротов канолы/рапса и белковые концентраты. Аквакультура 400-401 , 85–100, https://doi.org/10.1016/j.aquaculture.2013.03.006 (2013).
КАС Статья Google Scholar
Шеперд, С. Дж., Монройг, О. и Точер, Д. Р. Будущая доступность сырья для кормов для лосося и последствия цепочки поставок: пример лосося, выращиваемого в Шотландии. Аквакультура 467 , 49–62, https://doi.org/10.1016/j.aquaculture.2016.08.021 (2017).
Аквакультура 467 , 49–62, https://doi.org/10.1016/j.aquaculture.2016.08.021 (2017).
Артикул Google Scholar
Ягуби, М., Мозанзаде, М.Т., Мараммази, Дж.Г., Сафари, О. и Гисберт, Э. Диетическая замена рыбной муки соевыми продуктами (соевая мука и изолированный соевый белок) у молоди серебристо-черных порги ( Sparidentex hasta ). Аквакультура 464 , 50–59, https://doi.org/10.1016/j.aquaculture.2016.06.002 (2016).
КАС Статья Google Scholar
Lall, S.P. & Anderson, S. Аминокислотное питание лососевых рыб: диетические потребности и биодоступность. Cahiers Options Méditerranéennes 63 , 73–90 (2005).
Google Scholar
Michl, S.C. и др. . Податливый кишечный микробиом молоди радужной форели ( Oncorhynchus mykiss ): зависящие от диеты сдвиги структур бактериального сообщества. PloS one 12 , e0177735, https://doi.org/10.1371/journal.pone.0177735 (2017).
PloS one 12 , e0177735, https://doi.org/10.1371/journal.pone.0177735 (2017).
КАС Статья пабмед ПабМед Центральный Google Scholar
Хартвиксен, М. и др. . Альтернативные источники диетического белка для атлантического лосося ( Salmo salar L.) влияние на кишечную микробиоту, гистологию и рост кишечника и печени. Питание для аквакультуры 20 , 381–398, https://doi.org/10.1111/anu.12087 (2014).
КАС Статья Google Scholar
Кононова С. В., Зинченко Д. В., Муранова Т. А., Белова Н. А., Мирошников А. И. Кишечная микробиота лососевых рыб и ее изменения при введении в корм рыб соевых белков. Aquaculture International 27 , 475–496, https://doi.org/10.1007/s10499-019-00341-1 (2019).
КАС Статья Google Scholar
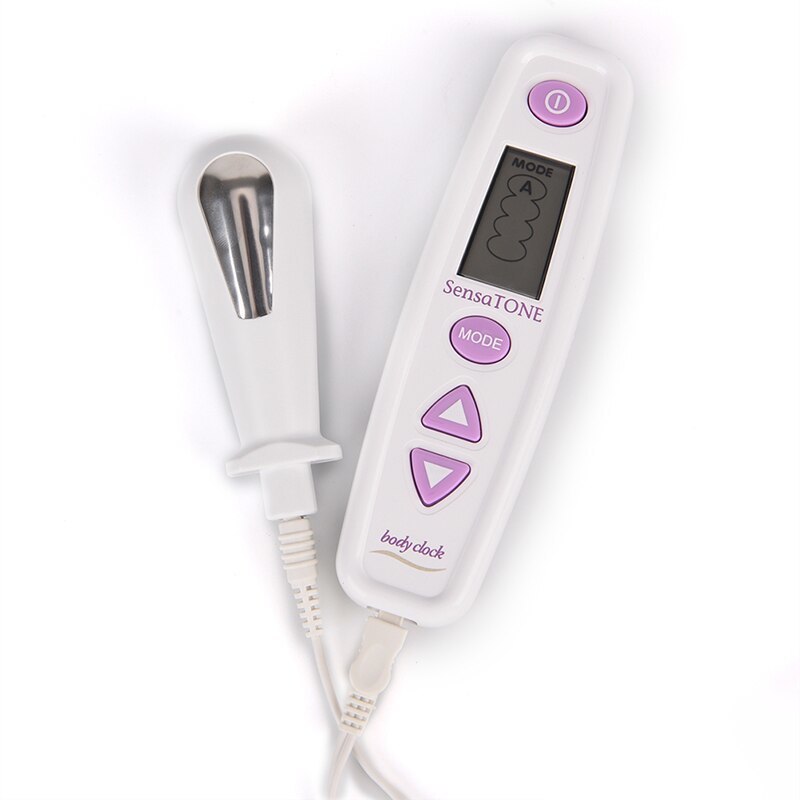
Zhou, Z., Ringø, E., Olsen, R. E. & Song, S. K. Диетические эффекты соевых продуктов на микробиоту кишечника и иммунитет водных животных: обзор. Питание для аквакультуры 24 , 644–665, https://doi.org/10.1111/anu.12532 (2018).
КАС Статья Google Scholar
Чжан, К. и др. . Замена рыбной муки соевой мукой в рационе японского морского окуня ( Lateolabrax japonicus ): влияние на рост, активность пищеварительных ферментов, гистологию кишечника и экспрессию генов воспаления кишечника и генов-транспортеров. Аквакультура 483 , 173–182 (2018).
КАС Статья Google Scholar
Крогдаль, А., Бакке-Маккеллеп, А. М. и Баеверфьорд, Г.Влияние дозированного количества стандартной соевой муки на структуру кишечника, активность ферментов слизистой оболочки и реакцию поджелудочной железы у атлантического лосося ( Salmo salar L. ). Питание для аквакультуры 9 , 361–371, https://doi.org/10.1046/j.1365-2095.2003.00264.x (2003).
). Питание для аквакультуры 9 , 361–371, https://doi.org/10.1046/j.1365-2095.2003.00264.x (2003).
Артикул Google Scholar
Dalsgaard, J. и др. . Влияние экзогенных ферментов на кажущуюся усвояемость питательных веществ у радужной форели ( Oncorhynchus mykiss ), получавших рационы с высоким содержанием растительного белка. Наука и технология кормов для животных 171 , 181–191 (2012).
КАС Статья Google Scholar
Ringø, E. & Song, S.K. Применение пищевых добавок (синбиотиков и пробиотиков в сочетании с растительными продуктами и β-глюканами) в аквакультуре. Питание для аквакультуры 22 , 4–24 (2016).
Артикул КАС Google Scholar
Лю, Ю. и др. . Добавка гидроксипролина на эффективность рационов с высоким содержанием растительного белка для тюрбо ( Scophthalmus maximus L. ). Аквакультура 433 , 476–480 (2014).
). Аквакультура 433 , 476–480 (2014).
КАС Статья Google Scholar
Sarker, MSA, Satoh, S. & Kiron, V. Включение лимонной кислоты и/или аминокислотных хелатных микроэлементов в рационы с альтернативными источниками растительного белка влияет на рост и выделение азота и фосфора у красного морского леща. Пагрус большой . Аквакультура 262 , 436–443 (2007).
КАС Статья Google Scholar
Эгертон С., Каллоти С., Хули Дж., Стэнтон С. и Росс Р. П. Микробиота кишечника морских рыб. Границы микробиологии 9 , 873, https://doi.org/10.3389/fmicb.2018.00873 (2018).
Артикул пабмед ПабМед Центральный Google Scholar
Меррифилд, Д.Л. и др. . Текущее состояние и будущее применение пробиотиков и пребиотиков для лососевых. Аквакультура 302 , 1–18 (2010).
Аквакультура 302 , 1–18 (2010).
Артикул Google Scholar
Эспе, М., Лемме, А., Петри, А. и Эль-Мовафи, А. Может ли атлантический лосось ( Salmo salar ) расти на диетах, лишенных рыбной муки? Аквакультура 255 , 255–262, https://doi.org/10.1016/j.aquaculture.2005.12.030 (2006 г.).
КАС Статья Google Scholar
Кусулаки, К. и др. . Высокие темпы роста у атлантического лосося ( Salmo salar L.), при кормлении 7,5% рыбной муки. Микро-, ультра- и нанофильтрация клеевой воды и влияние различных фракций и соединений на качество гранул и продуктивность рыбы. Аквакультура 338-341 , 134–146, https://doi.org/10.1016/j.aquaculture.2012.01.017 (2012 г.).
КАС Статья Google Scholar
Эспе, М., Лемме, А., Петри, А. и Эль-Мовафи, А. Оценка потребности в лизине для максимального прироста белка у атлантического лосося при использовании диеты с растительным белком. Аквакультура 263 , 168–178, https://doi.org/10.1016/j.aquaculture.2006.10.018 (2007).
и Эль-Мовафи, А. Оценка потребности в лизине для максимального прироста белка у атлантического лосося при использовании диеты с растительным белком. Аквакультура 263 , 168–178, https://doi.org/10.1016/j.aquaculture.2006.10.018 (2007).
КАС Статья Google Scholar
Green, T.J., Smullen, R. & Barnes, A.C. Расстройство кишечника, вызванное диетическим концентратом соевого белка, у атлантического лосося, выращиваемого на морских фермах, Salmo salar связано с изменениями в микробиоте кишечника. Ветеринарная микробиология 166 , 286–292, https://doi.org/10.1016/j.vetmic.2013.05.009 (2013).
КАС Статья пабмед ПабМед Центральный Google Scholar
Шмидт В., Амарал-Зеттлер, Л., Дэвидсон, Дж., Саммерфельт, С. и Гуд, К. Влияние рационов без рыбной муки на микробные сообщества в системах рециркуляционной аквакультуры атлантического лосося ( Salmo salar ). Прикладная экологическая микробиология 82 , 4470–4481 (2016).
Прикладная экологическая микробиология 82 , 4470–4481 (2016).
КАС пабмед Статья ПабМед Центральный Google Scholar
Bakke-McKellep, A.M. et al . Влияние диетической соевой муки, инулина и окситетрациклина на кишечную микробиоту и стресс эпителиальных клеток, апоптоз и пролиферацию костистых атлантических лососей. (Салмо салар Л.). 97 , 699–713 (2007).
КАС пабмед ПабМед Центральный Google Scholar
Dehler, C.E., Secombes, C.J. & Martin, S.A. Экологические и физиологические факторы формируют микробиоту кишечника молоди атлантического лосося ( Salmo salar L.). Аквакультура 467 , 149–157, https://doi.org/10.1016/j.aquaculture.2016.07.017 (2017).
КАС Статья пабмед ПабМед Центральный Google Scholar

Наваррете П., Эспехо Р. Т. и Ромеро Дж. Молекулярный анализ микробиоты пищеварительного тракта молоди атлантического лосося ( Salmo salar L.). Микробная экология 57 , 550–561, https://doi.org/10.1007/s00248-008-9448-x (2009).
КАС Статья пабмед ПабМед Центральный Google Scholar
Dehler, C.E., Secombes, C.J. & Martin, S.A.M. Перенос морской воды изменяет профиль микробиоты кишечника атлантического лосося ( Salmo salar L.). Научные отчеты 7 , 13877, https://doi.org/10.1038/s41598-017-13249-8 (2017).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google Scholar
Llewellyn, M. S. и др. . Биогеография кишечного микробиома атлантического лосося ( Salmo salar ). Журнал ISME 10 , 1280–1284, https://doi.org/10.1038/ismej.2015.189 (2016).
Артикул пабмед ПабМед Центральный Google Scholar
Кристинссон, Х.Г. и Раско, Б.А. Гидролизаты белков рыб: получение, биохимические и функциональные свойства. Критические обзоры в области пищевой науки и питания 40 , 43–81, https://doi.org/10.1080/104086189266 (2000).
КАС Статья пабмед ПабМед Центральный Google Scholar
Egerton, S., Culloty, S., Whooley, J., Stanton, C. & Ross, R.P. Характеристика белковых гидролизатов путассу ( Micromesistius poutassou ) и их применение для обогащения напитков. Пищевая химия 245 , 698–706 (2018).
КАС пабмед Статья ПабМед Центральный Google Scholar
Кристинссон, Х., Теодор, А. и Ингадоттир, Б. в Максимизация ценности морских побочных продуктов стр. 144-168 (Woodhead Publishing, 2007).
144-168 (Woodhead Publishing, 2007).
Benjakul, S., Yarnpakdee, S., Senphan, T., Halldorsdottir, S.M. & Kristinsson, H.G. Гидролизаты белков рыб: производство, биологическая активность и применение. Антиоксиданты и функциональные компоненты в пищевых продуктах на водной основе , 1-е изд. Рейкьявик, Исландия: Matil Ltd , 237Á283 (2014).
Хеврой, Э. М. и др. . Использование питательных веществ у атлантического лосося ( Salmo salar L.) при кормлении повышенным уровнем гидролизата рыбьего белка в период быстрого роста. Питание для аквакультуры 11 , 301–313 (2005).
Артикул Google Scholar
Bøgwald, J., Dalmo, R., McQueen Leifson, R., Stenberg, E. & Gildberg, A. Стимулирующее действие гидролизата мышечного белка из атлантической трески, Gadus morhua L ., на атлантического лосося, Salmo salar L ., лейкоциты головы почки. Иммунология рыбы и моллюсков 6 , 3–16, https://doi.org/10.1006/fsim.1996.0002 (1996).
Иммунология рыбы и моллюсков 6 , 3–16, https://doi.org/10.1006/fsim.1996.0002 (1996).
Артикул Google Scholar
Каху, К.Л., Инфанте, Дж.Л., Квазугель, П. и Ле Галл, М. М. Гидролизат белка по сравнению с рыбной мукой в сложных рационах для 10-дневных личинок морского окуня Dicentrarchus labrax . Аквакультура 171 , 109–119 (1999).
КАС Статья Google Scholar
Espe, M., Sveier, H., Høgøy, I. & Lied, E. Поглощение питательных веществ и рост атлантического лосося ( Salmo salar L .), питающегося концентратом рыбьего белка. Аквакультура 174 , 119–137 (1999).
КАС Статья Google Scholar
Мюррей, А. Л. и др. . Влияние различных кормовых добавок, содержащих гидролизат рыбного белка или побочные продукты переработки рыбы, на функции врожденного иммунитета молоди кижуча ( Oncorhynchus kisutch ). Аквакультура 220 , 643–653, https://doi.org/10.1016/s0044-8486(02)00426-x (2003).
Аквакультура 220 , 643–653, https://doi.org/10.1016/s0044-8486(02)00426-x (2003).
КАС Статья Google Scholar
Акснес А., Хоуп Б., Йонссон Э., Бьернссон Б. Т. и Альбректсен С. Фракционированный по размеру гидролизат рыбы в качестве кормового ингредиента для радужной форели ( Oncorhynchus mykiss ), получающей рационы с высоким содержанием растительного белка. I: Рост, регуляция роста и использование корма. Аквакультура 261 , 305–317, https://doi.org/10.1016/j.aquaculture.2006.07.025 (2006).
КАС Статья Google Scholar
Лян М., Wang, J., Chang, Q. & Mai, K. Влияние различных уровней гидролизата рыбного белка в рационе на неспецифический иммунитет японского морского окуня, Lateolabrax japonicus (Cuvieret Valenciennes, 1828). Исследования аквакультуры 37 , 102–106, https://doi. org/10.1111/j.1365-2109.2005.01392.x (2006).
org/10.1111/j.1365-2109.2005.01392.x (2006).
КАС Статья Google Scholar
Berge, G. M. & Storebakken, T. Гидролизат рыбного белка в стартовом рационе для мальков атлантического лосося ( Salmo salar ). Аквакультура 145 , 205–212 (1996).
КАС Статья Google Scholar
Gildberg, A., Johansen, A. & Bøgwald, J. Рост и выживание мальков атлантического лосося ( Salmo salar ) на диетах, дополненных гидролизатом рыбьего белка и молочнокислыми бактериями, во время контрольного испытания с Aeromonas salmonicida . Аквакультура 138 , 23–34, https://doi.org/10.1016/0044-8486(95)01144-7 (1995).
Артикул Google Scholar
Rollin, X., Mambrini, M., Abboudi, T., Larondelle, Y. & Kaushik, S. J. Оптимальный рацион незаменимых аминокислот для выращивания мальков атлантического лосося ( Salmo salar L. ). British Journal of Nutrition 90 , 865–876 (2003).
). British Journal of Nutrition 90 , 865–876 (2003).
КАС пабмед Статья ПабМед Центральный Google Scholar
Sissener, N.H., Sanden, M., Bakke, A.M., Krogdahl, Å. и Hemre, G. I. Длительное испытание с атлантическим лососем ( Salmo salar L.), которого кормили генетически модифицированной соей; уделяя особое внимание общему здоровью и производительности до, во время и после преобразования пеструшки в смолта. Аквакультура 294 , 108–117, https://doi.org/10.1016/j.aquaculture.2009.05.002 (2009).
КАС Статья Google Scholar
Бекман, Б. Р. и др. . Влияние различных условий выращивания в заводских условиях на выживание весенней чавычи от смолта до взрослой особи. Transactions of the American Fisheries Society 146 , 539–555, https://doi.org/10.1080/00028487.2017.1281168 (2017).
Артикул Google Scholar
Е, Л., Амберг, Дж., Чепмен, Д., Гайковски, М. и Лю, В.-Т. Анализ микробиоты кишечника рыб различает физиологию и поведение инвазивных азиатских карпов и местных американских рыб. Журнал ISME 8 , 541 (2014).
КАС пабмед Статья ПабМед Центральный Google Scholar
Bransden, M., Carter, C. & Nowak, B. Влияние источника пищевого белка на рост, иммунную функцию, химический состав крови и устойчивость к болезням атлантического лосося ( Salmo salar L.) parr. Зоотехника 73 , 105–113 (2001).
КАС Статья Google Scholar
Оверланд, М. и др. . Концентрат горохового белка, заменяющий рыбную муку или соевую муку в рационе атлантического лосося ( Salmo salar ) — влияние на показатели роста, усвояемость питательных веществ, состав туши, здоровье кишечника и физическое качество корма. Аквакультура 288 , 305–311, https://doi.org/10.1016/j.aquaculture.2008.12.012 (2009).
Аквакультура 288 , 305–311, https://doi.org/10.1016/j.aquaculture.2008.12.012 (2009).
КАС Статья Google Scholar
Берр, Г.S., Wolters, W. R., Barrows, F. T. & Hardy, R. W. Замена рыбной муки смесями альтернативных белков на показатели роста радужной форели ( Oncorhynchus mykiss ) и ранней или поздней молоди атлантического лосося ( Salmo salar ). Аквакультура 334-337 , 110–116, https://doi.org/10.1016/j.aquaculture.2011.12.044 (2012).
КАС Статья Google Scholar
Пратумиот, Дж., Бендиксен Э.А., Белл Дж.Г. и Точер Д.Р. Влияние увеличения замены рыбной муки в рационе источниками растительного белка на показатели роста и липидный состав тела атлантического лосося ( Salmo salar L.). Аквакультура 305 , 124–132, https://doi.org/10.1016/j.aquaculture.2010.04.019 (2010).
КАС Статья Google Scholar
Рефсти С., Олли Дж. Дж. и Стандал Х.Потребление корма, рост и использование белка атлантическим лососем после смолты ( Salmo salar ) в зависимости от уровня гидролизата рыбного белка в рационе. Аквакультура 239 , 331–349, https://doi.org/10.1016/j.aquaculture.2004.06.015 (2004).
КАС Статья Google Scholar
Espe, M., Ruohonen, K. & El-Mowafi, A. Гидролизованный концентрат рыбного белка (FPC) снижает массу внутренних органов у атлантического лосося ( Salmo salar ), питающегося рационами на основе растительного белка. Питание для аквакультуры 18 , 599–609, https://doi.org/10.1111/j.1365-2095.2012.00944.x (2012).
КАС Статья Google Scholar
Рюйтер, Б., Мойя-Фалькон, К., Розенлунд, Г. и Вегусдал, А. Содержание жира и морфология печени и кишечника атлантического лосося ( Salmo salar ): Влияние температуры и диеты соевое масло. Аквакультура 252 , 441–452, https://doi.org/10.1016/j.aquaculture.2005.07.014 (2006 г.).
Содержание жира и морфология печени и кишечника атлантического лосося ( Salmo salar ): Влияние температуры и диеты соевое масло. Аквакультура 252 , 441–452, https://doi.org/10.1016/j.aquaculture.2005.07.014 (2006 г.).
КАС Статья Google Scholar
Nordgarden, U., Hemre, G.-I. & Hansen, T. Рост и состав тела атлантического лосося ( Salmo salar L.), кормящиеся молодняком и смолтом с разным содержанием белка и липидов. Аквакультура 207 , 65–78 (2002).
КАС Статья Google Scholar
Ву, Н., Берн, Х. и Нишиока, Р. Изменения в составе тела, связанные со смолтификацией и преждевременным переходом в морскую воду у кижуча ( Oncorhynchus kisutch ) и королевского лосося ( O. tschawytscha ). Journal of Fish Biology 13 , 421–428 (1978).
КАС Статья Google Scholar
Виртанен Э., Седерхольм-Тана Л., Сойвио А., Форман Л. и Муона М.Влияние физиологического состояния и статуса смолтификации при выпуске смолта на последующие уловы взрослых лососей. Аквакультура 97 , 231–257 (1991).
Артикул Google Scholar
Гао, В. и др. . Белковосберегающая способность пищевых липидов у травоядных и всеядных пресноводных рыб: сравнительное исследование белого амура ( Ctenopharyngodon idella ) и тиляпии ( Oreochromis niloticus × O.золотистый ). Питание для аквакультуры 17 , 2–12 (2011).
КАС Статья Google Scholar
Сантигоса, Э. и др. . Модификации пищеварительных ферментов у форели ( Oncorhynchus mykiss ) и морского леща ( Sparus aurata ) в ответ на замену рыбной муки в рационе растительными источниками белка. Аквакультура 282 , 68–74, https://doi. org/10.1016/j.aquaculture.2008.06.007 (2008).
org/10.1016/j.aquaculture.2008.06.007 (2008).
КАС Статья Google Scholar
Ларсен Б.К., Далсгаард Дж. и Педерсен П.Б. Влияние растительных белков на постпрандиальные концентрации свободных аминокислот в плазме у радужной форели ( Oncorhynchus mykiss ). Аквакультура 326–329 , 90–98, https://doi.org/10.1016/j.aquaculture.2011.11.028 (2012).
КАС Статья Google Scholar
Storebakken, T., Shearer, KD & Roem, AJ. Доступность белка, фосфора и других элементов в рыбной муке, концентрате соевого белка и корме на основе концентрата соевого белка, обработанного фитазой, для атлантического лосося, Salmo salar . Аквакультура 161 , 365–379, https://doi.org/10.1016/S0044-8486(97)00284-6 (1998).
КАС Статья Google Scholar
Усмани Н. и Джафри А.K. Влияние фитиновой кислоты в рационе на рост, эффективность преобразования и состав туши мальков mrigal Cirrhinus mrigala (Hamilton). Журнал Всемирного общества аквакультуры 33 , 199–204 (2002).
и Джафри А.K. Влияние фитиновой кислоты в рационе на рост, эффективность преобразования и состав туши мальков mrigal Cirrhinus mrigala (Hamilton). Журнал Всемирного общества аквакультуры 33 , 199–204 (2002).
Артикул Google Scholar
Юсиф, О. М., Альхадрами, Г. А. и Пессаракли, М. Оценка обезвоженной люцерны и листьев солончака ( Atriplex ) в рационах для тиляпии ( Oreochromis aureus L.). Аквакультура 126 , 341–347 (1994).
Артикул Google Scholar
Гатлин, Д. М. III и Филлипс, Х. Ф. Взаимодействие кальция, фитата и цинка с пищей у канального сома. Аквакультура 79 , 259–266 (1989).
КАС Статья Google Scholar
Паризи, Г. и др. . Влияние полной замены пищевой рыбной муки источниками растительного белка на ранние послеубойные изменения биохимических и физических показателей радужной форели. Veterinary Research Communications 28 , 237–240 (2004).
Veterinary Research Communications 28 , 237–240 (2004).
ПабМед Статья ПабМед Центральный Google Scholar
Mambrini, M., Roem, AJ, Carvedi, J., Lalles, J. & Kaushik, S. Влияние замены рыбной муки концентратом соевого белка и добавки DL-метионина в высококалорийные экструдированные продукты. диеты на рост и использование питательных веществ радужной форелью Oncorhynchus mykiss . Зоотехника 77 , 2990–2999 (1999).
КАС Статья Google Scholar
Каушик, С., Ковес, Д., Дутто, Г. и Блан, Д. Почти полная замена рыбной муки источниками растительного белка в рационе морского костистого скелета, европейского морского окуня, Dicentrarchus labrax . Аквакультура 230 , 391–404 (2004).
КАС Статья Google Scholar

Хосрави, С. и др. . Пищевая добавка гидролизатов морского белка в рационы на основе рыбной муки для красного морского леща ( Pagrus major ) и оливковой камбалы ( Paralichthys olivaceus ). Аквакультура 435 , 371–376, https://doi.org/10.1016/j.aquaculture.2014.10.019 (2015).
КАС Статья Google Scholar
Conceicao, L. E. и др. . Пищевой азот и благополучие рыб. Физиология рыб и . Биохимия 38 , 119–141, https://doi.org/10.1007/s10695-011-9592-y (2012).
КАС Статья Google Scholar
НРК. Потребность рыбы и креветок в питательных веществах. Серия кормов для животных (2011 г.).
Хосрави С. и др. . Влияние добавления белковых гидролизатов в рационы с низким содержанием рыбной муки на показатели роста, врожденный иммунитет и устойчивость к болезням красного морского леща Pagrus major . Иммунология рыбы и моллюсков 45 , 858–868, https://doi.org/10.1016/j.fsi.2015.05.039 (2015).
Иммунология рыбы и моллюсков 45 , 858–868, https://doi.org/10.1016/j.fsi.2015.05.039 (2015).
КАС Статья Google Scholar
Fournier, V., Huelvan, C. & Desbruyeres, E. Включение смеси растительных кормов в качестве заменителя рыбной муки в рацион молоди тюрбо ( Psetta maxima ). Аквакультура 236 , 451–465 (2004).
Артикул Google Scholar
Leal, A.L.G., de Castro, P.F., de Lima, J.P.V., de Souza Correia, E. & de Souza Bezerra, R. Использование гидролизата белка креветок в кормах для нильской тиляпии ( Oreochromis niloticus , L.). Aquaculture International 18 , 635–646, https://doi.org/10.1007/s10499-009-9284-0 (2010).
Артикул Google Scholar
Сюй, Х. и др. . Регулируемые уровни гидролизата рыбного белка в рационах с высоким содержанием растений для палтуса ( Scophthalmus maximus ): влияние на показатели роста и накопление липидов. Аквакультура 454 , 140–147, https://doi.org/10.1016/j.aquaculture.2015.12.006 (2016).
Аквакультура 454 , 140–147, https://doi.org/10.1016/j.aquaculture.2015.12.006 (2016).
КАС Статья Google Scholar
де Вердаль, Х. и др. . Измерения пищеварительного тракта и гистологическая адаптация у бройлерных линий, отобранных по дивергентной эффективности пищеварения. Poultry Science 89 , 1955–1961, https://doi.org/10.3382/ps.2010-813 (2010).
КАС Статья пабмед ПабМед Центральный Google Scholar
Zarkasi, KZ и др. . Атлантический лосось ( Salmo salar L. ) динамика микробного сообщества желудочно-кишечного тракта в зависимости от свойств пищеварения и рациона. Микробная экология 71 , 589–603, https://doi.org/10.1007/s00248-015-0728-y (2016).
КАС Статья пабмед ПабМед Центральный Google Scholar

Ley, R. E. et al . Ожирение изменяет микробную экологию кишечника. Proceedings of the National Academy of Sciences of the United States of America 102 , 11070–11075, https://doi.org/10.1073/pnas.0504978102 (2005).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google Scholar
Menni, C. и др. . Разнообразие микробиома кишечника и потребление большого количества клетчатки связаны с более низким долгосрочным набором веса. International Journal Of Obesity 41 , 1099, https://doi.org/10.1038/ijo.2017.66 (2017).
КАС Статья пабмед ПабМед Центральный Google Scholar
Turnbaugh, P. J. & Gordon, J. I. Основной микробиом кишечника, энергетический баланс и ожирение. Журнал физиологии 587 , 4153–4158, https://doi.org/10.1113/jphysiol.2009.174136 (2009).
КАС Статья пабмед ПабМед Центральный Google Scholar

Семова И. и др. . Микробиота регулирует кишечную абсорбцию и метаболизм жирных кислот у рыбок данио. Клеточный хозяин и микроб 12 , 277–288 (2012).
КАС Статья Google Scholar
Ni, J., Yan, Q., Yu, Y. & Zhang, T. Факторы, влияющие на микробиом кишечника белого амура и его влияние на метаболизм. FEMS микробиология экология 87 , 704–714 (2014).
КАС пабмед Статья ПабМед Центральный Google Scholar
Ye, JD, Wang, K., Li, FD & Sun, YZ Единичное или комбинированное воздействие добавок фрукто- и маннанолигосахаридов и Bacillus clausii на рост, использование корма, состав тела, активность пищеварительных ферментов, врожденный иммунный ответ и липидный обмен японской камбалы Paralichthys olivaceus . Питание для аквакультуры 17 , e902–e911 (2011).
Артикул Google Scholar

Banerjee, G. & Ray, A.K. Бактериальный симбиоз в кишечнике рыб и его роль в здоровье и обмене веществ. Symbiosis 72 , 1–11, https://doi.org/10.1007/s13199-016-0441-8 (2017).
КАС Статья Google Scholar
Webster, T.M.U., Consuegra, S., Hitchings, M. & de Leaniz, C.G.Межпопуляционная изменчивость микробиома атлантического лосося отражает экологическое и генетическое разнообразие. Прикладная экологическая микробиология 84 , e00691–00618 (2018).
Google Scholar
Дером, Н., Готье, Дж., Бутин, С. и Ллевеллин, М. в Эффект Распутина: когда комменсалы и симбионты становятся паразитами 81-108 (Springer, 2016).
Лавуа, К., Курсель, М., Redivo, B. & Derome, N. Структурное и композиционное несоответствие микробиоты кишечника атлантического лосося, содержащегося в неволе, и дикого атлантического лосося ( Salmo salar ) подчеркивает актуальность интеграции молекулярной экологии в методы управления и сохранения. Эволюционные приложения 11 , 1671–1685 (2018).
Эволюционные приложения 11 , 1671–1685 (2018).
ПабМед ПабМед Центральный Статья Google Scholar
Гупта С., Фернандес Дж. и Кирон В.Антибиотикоиндуцированные возмущения проявляются в доминирующих кишечных бактериальных типах атлантического лосося. Микроорганизмы 7 , 233 (2019).
Центральный пабмед Статья Google Scholar
Руди К. и др. . Стабильная основная кишечная микробиота при переходе от пресной воды к морской для выращиваемого на ферме атлантического лосося. Прикладная экологическая микробиология 84 , https://doi.org/10.1128/AEM.01974-17 (2018).
Локеш, Дж., Кирон, В., Сипкема, Д., Фернандес, Дж. М.О. и Моум, Т. Последовательность эмбриональных и кишечных бактериальных сообществ атлантического лосося ( Salmo salar ) выявляет стадийно-специфические микробные подписи. Microbiology Open , e00672, https://doi.org/10.1002/mbo3.672 (2018).
Microbiology Open , e00672, https://doi.org/10.1002/mbo3.672 (2018).
ПабМед ПабМед Центральный Статья КАС Google Scholar
Лобо, С. и др. . Пищевые пробиотические добавки ( Shewanella putrefaciens Pdp11) модулируют микробиоту кишечника и способствуют росту и состоянию личинок сенегальской камбалы. Физиология и биохимия рыб 40 , 295–309, https://doi.org/10.1007/s10695-013-9844-0 (2014).
КАС Статья пабмед ПабМед Центральный Google Scholar
Брант, Дж. и Остин, Б.Использование пробиотика для борьбы с лактококкозом и стрептококкозом у радужной форели, Oncorhynchus mykiss (Walbaum). Журнал болезней рыб 28 , 693–701 (2005).
КАС пабмед Статья ПабМед Центральный Google Scholar
Гахардо, К. и др. . Карта микробиоты кишечника атлантического лосося с высоким разрешением ( Salmo salar ): основа для сравнительного исследования микробиоты кишечника. Научные отчеты 6 , 30893, https://doi.org/10.1038/srep30893 (2016).
и др. . Карта микробиоты кишечника атлантического лосося с высоким разрешением ( Salmo salar ): основа для сравнительного исследования микробиоты кишечника. Научные отчеты 6 , 30893, https://doi.org/10.1038/srep30893 (2016).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google Scholar
Диас, С. и др. . Отчет о глобальной оценке биоразнообразия и экосистемных услуг. (Межправительственная научно-политическая платформа Организации Объединенных Наций по биоразнообразию и экосистемным услугам, 2019 г.).
Ассоциация официальных химиков-аналитиков, И. Официальные методы анализа . 17-е издание, 2200 г. (AOAC International, 2002 г.).
Смедес, Ф. Определение общего содержания липидов с использованием нехлорированных растворителей. Аналитик 124 , 1711–1718 (1999).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar

McDermott, A. и др. . Прогнозирование содержания отдельных молочных белков, включая свободные аминокислоты, в коровьем молоке с использованием спектроскопии в среднем инфракрасном диапазоне и их корреляции с характеристиками переработки молока. Journal of Dairy Science 99 , 3171–3182, https://doi.org/10.3168/jds.2015-9747 (2016).
КАС Статья пабмед ПабМед Центральный Google Scholar
Шнайдер, К. А., Расбанд, В. С. и Элисейри, К. В. NIH Image to ImageJ: 25 лет анализа изображений. Nature Methods 9 , 671 (2012).
КАС пабмед ПабМед Центральный Статья Google Scholar
Caporaso, JG и др. . QIIME позволяет анализировать данные секвенирования с высокой пропускной способностью. Nature Methods 7 , 335 (2010).
КАС пабмед ПабМед Центральный Статья Google Scholar
