
Что такое метаболический синдром
Что такое метаболический синдром?
Для метаболического синдрома характерно висцеральное (внутреннее) ожирение в сочетании с различными нарушениями. Считается, что люди, у которых по всем критериям диагностируют метаболический синдром, в будущем более всего подвержены таким заболеваниям, как инсульт головного мозга и инфаркт миокарда.
Чем опасен тип внутреннего (висцерального) ожирения?
Висцеральный жир секретирует различные биологически активные вещества (например, фактор некроза опухоли-альфа и др.). При висцеральном типе ожирения эта секреция становится аномальной. В результате повышается уровень сахара в крови, артериальное давление, уровень «плохого холестерина» (липопротеиды низкой плотности) и т.д. Чем больше нарушений обмена веществ, тем быстрее прогрессирует атеросклероз и повышается риск развития инфаркта миокарда или инсульта.
Ожирение делится на 2 типа: висцеральное и подкожное.
Тип висцерального ожирения
В этом случае жир накапливается в брюшной полости (вокруг внутренних органов).
При метаболическом синдроме именно данный тип ожирения наносит вред здоровью!
Тип подкожного ожирения
В этом случае жир накапливается в подкожной клетчатке.
Избавиться от подкожного жира сложнее, чем от висцерального.
Как измерить объем висцерального жира?
1. Измерение обхвата живота. Самый простой способ – измерить обхват своего живота на уровне пупка. В этом нам поможет обычная сантиметровая лента.
2. Метод биоэлектрического импеданса. В последнее время продаются как дорогостоящие аппараты для медицинского назначения, так и аппараты для использования в домашних условиях.
3. Метод с использованием компьютерной томографии. На компьютерном томографе сканируется область живота и с помощью специальной программы измеряется объем жира. Данный метод в отличие от других позволяет увидеть внутренний (висцеральный) жир.
Данный метод в отличие от других позволяет увидеть внутренний (висцеральный) жир.
Часто задаваемые вопросы.
— Связан ли метаболический синдром с образом жизни?
Заболевания, связанные с образом жизни, — это заболевания, которые развиваются вследствие неправильного питания, малоподвижного образа жизни, курения, употребления алкоголя. В эту группу заболеваний входят онкологические заболевания, инсульты, сердечно-сосудистые заболевания, сахарный диабет, ожирение и т.д. Метаболический синдром – это висцеральный тип ожирения в сочетании с 2-мя или более следующими состояниями: повышенный уровень глюкозы в крови, повышенный уровень холестерина в крови, гипертония. При метаболическом синдроме увеличивается риск атеросклероза, и как следствие повышается риск возникновения инсульта, инфаркта миокарда.
— У каких людей есть склонность к развитию метаболического синдрома?
Метаболический синдром чаще всего встречается у мужчин среднего и пожилого возраста, которые неправильно питаются, ежедневно употребляют алкоголь, ведут малоподвижный образ жизни. У женщин нарушение метаболических процессов происходит в основном в период менопаузы.
У женщин нарушение метаболических процессов происходит в основном в период менопаузы.
— Кому рекомендуется пройти обследование по поводу метаболического синдрома?
Обследование по поводу метаболического синдрома рекомендуется пройти, если у мужчин обхват талии более 85 см, у женщин – более 90 см.
— Кто может пройти обследование на определение объема висцерального жира на компьютерном томографе?
Любой человек может пройти такое обследование, однако данное обследование нежелательно для беременных женщин, так как проходит с использованием облучения. Тем людям, кто желает пройти такое обследование, следует обратиться в диагностическое учреждение, в котором есть аппарат КТ.
— Сколько времени занимает определение объема висцерального жира с помощью КТ?
Само сканирование занимает около 1 секунды. С учетом времени, затрачиваемого на подготовку, исследование занимает около 5 минут.
— Нужна ли специальная подготовка/диета перед исследованием?
Нет, диета и подготовка не требуется.
Нарушения метаболизма при сосудистых заболеваниях головного мозга
Сосудистые заболевания головного мозга – одна из ведущих причин смертности и инвалидизации населения всего мира. Окклюзия сосудов, питающих головной мозг, является начальным звеном в цепочке неблагоприятных изменений, которые приводят к грубым нарушениям метаболизма нейронов, структурно-функциональным изменениям, нередко заканчивающимся гибелью нервных клеток. Острая или хроническая ишемия мозговой ткани обусловливает целый каскад патологических реакций, которые в конечном итоге приводят к развитию очагового неврологического дефицита, дисциркуляторной энцефалопатии или к гибели больного. Тесная взаимосвязь нарушений энергетического и пластического обмена, их влияние на течение и прогноз заболевания нередко не учитываются при разработке схем лечения, а основой патогенетической терапии считается восстановление гемодинамики.
В последнее время нарушениям метаболизма и возможностям их коррекции уделяется большое внимание.
О возрастных гемодинамических и метаболических особенностях у больных ишемическим инсультом в реабилитационном периоде и способах их коррекции рассказала
директор Украинского реабилитационного центра Института геронтологии АМН Украины, доктор медицинских наук, профессор Светлана Михайловна Кузнецова.
– Неврологи очень много внимания уделяют лечению острого периода инсульта, а также профилактике цереброваскулярных нарушений. Не менее важной остается проблема реабилитации больных, перенесших инсульт.
Стратегия реабилитации пациентов, перенесших инсульт, предусматривает не только ликвидацию или уменьшение устойчивых патологических состояний, но и активацию компенсаторных механизмов в системе церебральной гемодинамики и метаболизма. Чтобы эта стратегия успешно реализовывалась, необходимы фундаментальные знания о состоянии мозга и мозгового кровообращения после инсульта. Изучение метаболических изменений при цереброваскулярных нарушениях позволило разработать патогенетическую терапию острого инсульта, которая в настоящее время с успехом применяется у пациентов, значительно уменьшая летальность и инвалидизацию больных. Такой же принцип (патогенетического лечения) должен быть внедрен и в систему реабилитации после перенесенного инсульта.

При этом очень важно учитывать возрастные особенности метаболизма и гемодинамики. Мы наблюдали пациентов среднего и пожилого возраста в период от 1 месяца до 1 года после острого инсульта. Пациенты от 40 до 59 лет вошли в группу среднего возраста, от 60 до 74 лет – в группу пожилых больных. Нами было выявлено, что течение как острого периода инсульта, так и периода реабилитации в этих двух группах имеет свои особенности. При изучении церебральной гемодинамики было обнаружено, что у больных среднего возраста изменения мозгового кровообращения более выражены по сравнению с контрольной группой такого же возраста и пожилыми людьми. Так, снижение объемной скорости кровотока в пораженных сосудах в средней возрастной группе с перенесенным инсультом составляло 55% по сравнению с той же возрастной группой без инсультов, в то время как в группе пожилых людей с инсультами этот показатель в сравнении с группой без инсультов оказался всего 19%.
О церебральной гемодинамике можно также судить и по структуре атеросклеротических бляшек: гипоэхогенные бляшки – самые агрессивные, наиболее часто приводящие к тромбозам, тромбоэмболиям, сосудистым катастрофам.
 В нашем исследовании показано, что у людей среднего возраста с инсультами такие бляшки встречаются намного чаще, чем у больных старшей возрастной группы, также перенесших инсульт.
В нашем исследовании показано, что у людей среднего возраста с инсультами такие бляшки встречаются намного чаще, чем у больных старшей возрастной группы, также перенесших инсульт.Таким образом, можно утверждать, что у больных, перенесших инсульт в среднем возрасте, атеросклероз имеет более агрессивное и быстропрогрессирующее течение. Еще в 70-е годы в Институте геронтологии (совместно с харьковским Институтом неврологии, психиатрии и наркологии) профессор Никита Борисович Маньковский исследовал проблему раннего церебрального атеросклероза; при этом было выявлено, что у людей 40-50 лет к инсультам и инфарктам приводит высокоагрессивное течение атеросклероза. В настоящее время эта теория продолжает находить свое подтверждение и учитывается при решении вопросов лечения и профилактики мозговых инсультов.
 Мы определяли у наших больных содержание метаболитов (N-ацетилацетата, холина, лактата и креатинина) в лобной и затылочной долях, а также в очаге инсульта. N-ацетилацетат является маркером здоровых нейронов; его определение используется не только для научных целей, но и в практической медицине во многих зарубежных странах для оценки течения и прогноза в острый период инсульта. Содержание этого метаболита в очаге на уровне 0,7 ммоль или ниже говорит о крайне неблагоприятном течении инсульта и глубокой инвалидизации. Холин входит в состав клеточных мембран и является показателем позднего глиоза, который у многих больных в период реабилитации проявляется так называемыми глиозными образованиями. Холин в некоторой степени свидетельствует о функциональной активности холинергической системы. Креатинин – буфер для энергетического обмена, источник АТФ и АМФ, поэтому он характеризует энергетический обмен в нейронах, при старении его содержание снижается. Лактат, как известно, представляет собой маркер ишемии, в здоровом мозге лактата не должно быть.
Мы определяли у наших больных содержание метаболитов (N-ацетилацетата, холина, лактата и креатинина) в лобной и затылочной долях, а также в очаге инсульта. N-ацетилацетат является маркером здоровых нейронов; его определение используется не только для научных целей, но и в практической медицине во многих зарубежных странах для оценки течения и прогноза в острый период инсульта. Содержание этого метаболита в очаге на уровне 0,7 ммоль или ниже говорит о крайне неблагоприятном течении инсульта и глубокой инвалидизации. Холин входит в состав клеточных мембран и является показателем позднего глиоза, который у многих больных в период реабилитации проявляется так называемыми глиозными образованиями. Холин в некоторой степени свидетельствует о функциональной активности холинергической системы. Креатинин – буфер для энергетического обмена, источник АТФ и АМФ, поэтому он характеризует энергетический обмен в нейронах, при старении его содержание снижается. Лактат, как известно, представляет собой маркер ишемии, в здоровом мозге лактата не должно быть. Ранее считалось, что лактат можно выявить только в остром периоде инсульта, а к концу первого месяца после сосудистой катастрофы он практически полностью исчезает. Но наши данные показывают, что лактат нередко остается и в более поздние сроки, причем не только в зоне ишемии, но и в других зонах. Таким образом, вышеперечисленные метаболиты можно использовать для оценки степени ишемического поражения мозга.
Ранее считалось, что лактат можно выявить только в остром периоде инсульта, а к концу первого месяца после сосудистой катастрофы он практически полностью исчезает. Но наши данные показывают, что лактат нередко остается и в более поздние сроки, причем не только в зоне ишемии, но и в других зонах. Таким образом, вышеперечисленные метаболиты можно использовать для оценки степени ишемического поражения мозга. При корреляционном анализе содержания метаболитов и объемной скорости кровотока в различных сосудах каротидного и вертебробазилярного бассейнов нами было обнаружено, что у людей среднего возраста метаболизм зависит от объемной скорости кровотока непораженных сосудов, кровоснабжающих полушарие, противоположное очагу инсульта, а у пожилых людей средняя мозговая артерия, которая является чаще всего источником ишемии и сосудистой катастрофы, влияет на метаболизм как здорового полушария, так и пораженного.
 Таким образом, у пациентов разного возраста совершенно по-разному осуществляются взаимоотношения между гемодинамикой и метаболизмом мозга.
Таким образом, у пациентов разного возраста совершенно по-разному осуществляются взаимоотношения между гемодинамикой и метаболизмом мозга. В патогенезе ишемического инсульта большое значение имеет и кардиальная патология. Не менее четверти случаев инсульта возникают на фоне заболеваний сердца. Назначая терапию при инсульте, следует помнить не только о возрастных особенностях церебральной гемодинамики и метаболизма, но и о воздействии тех или иных препаратов на центральную гемодинамику. Для людей среднего возраста обнаруживаются жесткие корреляционные связи между системной и церебральной гемодинамикой в области пораженных сосудов, у пожилых людей такие корреляции прослеживаются и по отношению к мозговым сосудам интактного полушария. Это говорит о том, что у пожилых пациентов более выражены нарушения авторегуляции мозгового кровообращения, что обусловливает необходимость более осторожно подходить к разработке стратегии лечения таких больных. Небольшое повышение или снижение систолического или диастолического системного артериального давления у таких пациентов значительно изменяет церебральную гемодинамику, в том числе и в регионах кровоснабжения интактных сосудов.
Таким образом, при реабилитации пациентов с инсультом одним из наиболее важных аспектов считается фармакологическая (медикаментозная) реабилитация. Основными методами воздействия на патогенез заболевания можно считать препараты вазоактивного и ноотропного действия, они улучшают гемодинамику и метаболизм мозга. Одним из наиболее перспективных нейропротекторных препаратов на сегодняшний день является препарат комплексного действия Инстенон. Он состоит из нескольких компонентов, которые имеют свою направленность действия: этамиван активно влияет на лимбико-ретикулярный комплекс, гексобендин обладает мощным метаболическим эффектом на нейроны головного мозга, этофиллин активирует метаболизм миокарда, тем самым оказывая положительное влияние на гемодинамику.
 Мы использовали этот препарат сначала внутримышечно в течение 10 дней, затем применяли 20 дней в таблетированной форме. Инстенон выраженно улучшает гемодинамику в интракраниальных сосудах – средней мозговой артерии и сосудах вертебробазилярного бассейна. Нами отмечен интересный эффект препарата: после применения Инстенона устраняется патологическое влияние гемодинамики в пораженных сосудах на электрогенез мозга. Следует отметить, что у больных пожилого возраста с инсультом очень жесткие связи не только между гемодинамикой и метаболизмом, но и между гемодинамикой и электрофизиологией мозга (ЭЭГ-показателями). Даже небольшие нарушения гемодинамики вызывают изменения ЭЭГ-ритмов. При применении Инстенона не только изменяется структура электрогенеза (снижается медленная активность и увеличивается интенсивность альфа-ритма, отмечается некоторое увеличение частоты альфа- и дельта-ритмов), но и устраняется патологическая зависимость электрогенеза от гемодинамики. Это свидетельствует о положительных вазотропных влияниях и антиишемическом эффекте препарата, которые в свою очередь улучшают психоэмоциональное состояние больного (улучшение памяти, речевой функции и других проявлений неврологического дефицита).
Мы использовали этот препарат сначала внутримышечно в течение 10 дней, затем применяли 20 дней в таблетированной форме. Инстенон выраженно улучшает гемодинамику в интракраниальных сосудах – средней мозговой артерии и сосудах вертебробазилярного бассейна. Нами отмечен интересный эффект препарата: после применения Инстенона устраняется патологическое влияние гемодинамики в пораженных сосудах на электрогенез мозга. Следует отметить, что у больных пожилого возраста с инсультом очень жесткие связи не только между гемодинамикой и метаболизмом, но и между гемодинамикой и электрофизиологией мозга (ЭЭГ-показателями). Даже небольшие нарушения гемодинамики вызывают изменения ЭЭГ-ритмов. При применении Инстенона не только изменяется структура электрогенеза (снижается медленная активность и увеличивается интенсивность альфа-ритма, отмечается некоторое увеличение частоты альфа- и дельта-ритмов), но и устраняется патологическая зависимость электрогенеза от гемодинамики. Это свидетельствует о положительных вазотропных влияниях и антиишемическом эффекте препарата, которые в свою очередь улучшают психоэмоциональное состояние больного (улучшение памяти, речевой функции и других проявлений неврологического дефицита).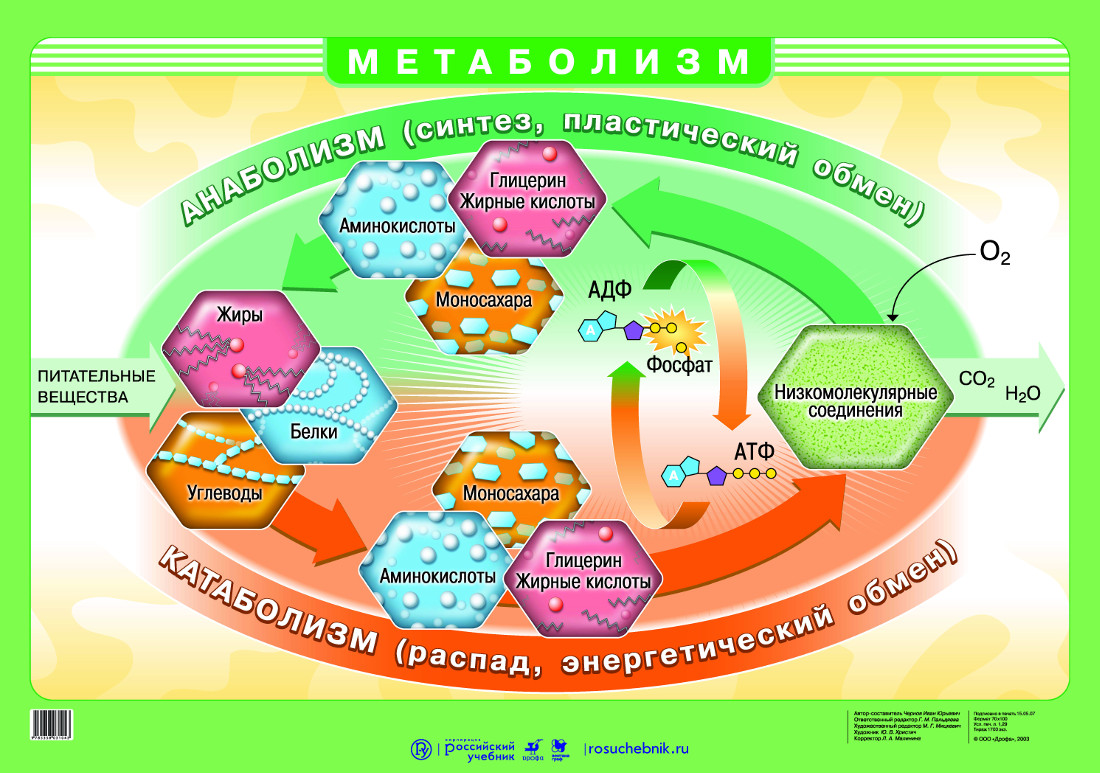
После перенесенного инсульта у многих больных через год, а иногда и раньше, появляется экстрапирамидная недостаточность, которая усугубляет неврологические нарушения и социальную дезадаптацию человека. Еще одним достаточно частым поздним последствием инсульта является депрессия. В связи с этими данными нами проведены длительные наблюдения за больными с целью изучения изменений метаболизма мозга в разные периоды времени после инсульта. Мы обнаружили, что в первое полугодие после перенесенного инсульта у больных наблюдается низкое содержание N-ацетилацетата, что свидетельствует о сниженной функциональной активности нейронов. К 12 месяцам содержание этого метаболита в сером веществе увеличивается, а на второй год после инсульта – резко снижается. Именно это и является основой формирования постинсультных дополнительных синдромов, а также предпосылкой для повторных нарушений мозгового кровообращения. Эти данные свидетельствуют о том, что больным после инсульта даже в отдаленные сроки необходима активная метаболическая и вазоактивная коррекция.
 Мы рекомендуем добавлять к терапии ингибиторы МАО, которые не только эффективно предупреждают развитие экстрапирамидных нарушений, но и улучшают когнитивные процессы, обладают антидепрессивным эффектом. У больных, которым 2-3 раза в год назначают курс лечения ингибиторами МАО, реже развивается экстрапирамидная недостаточность, двигательные нарушения не прогрессируют, а чаще регрессируют. В эксперименте при применении ингибиторов МАО улучшаются не только качество жизни пациентов, но и ее продолжительность.
Мы рекомендуем добавлять к терапии ингибиторы МАО, которые не только эффективно предупреждают развитие экстрапирамидных нарушений, но и улучшают когнитивные процессы, обладают антидепрессивным эффектом. У больных, которым 2-3 раза в год назначают курс лечения ингибиторами МАО, реже развивается экстрапирамидная недостаточность, двигательные нарушения не прогрессируют, а чаще регрессируют. В эксперименте при применении ингибиторов МАО улучшаются не только качество жизни пациентов, но и ее продолжительность.Говоря об антитромбоцитарной терапии как о важнейшей составляющей профилактики повторных инсультов, следует отметить, что в фармакодинамике аспирина важен не только антиагрегантный, но и противовоспалительный эффект препарата. Именно это действие аспирина – основа профилактики развития кист после инсультов.
Подводя итог нашему анализу возрастных гемодинамических и метаболических особенностей при острых инсультах, можно сделать следующие выводы:
– у людей среднего возраста, перенесших инсульт, атеросклеротический процесс идет более агрессивно, в связи с чем ведущим направлением лечения и реабилитации пациентов должна стать противосклеротическая терапия;
– у людей пожилого возраста при инсульте выражены не только нарушения гемодинамики, и особенно метаболизма мозга, но и отмечается жесткая взаимосвязь между центральным и церебральным кровотоком, поэтому наиболее актуальны в лечении препараты вазоактивного и нейропротекторного действия;
– лечение и реабилитация пациентов после инсульта должны быть комплексными, необходимо учитывать особенности гемодинамики и метаболизма мозга.

Результаты сравнительного анализа изменений метаболизма у лиц, перенесших ишемический инсульт однократно и повторно, а также у пациентов, страдающих транзиторными ишемическими атаками, привела в своем докладе заведующая кафедрой нервных болезней Харьковского государственного медицинского университета им. В.Н. Каразина, доктор медицинских наук, профессор Ирина Анатольевна Григорова.
– Одним из наиболее важных принципов реабилитации пациентов после инсульта является оценка тяжести не только неврологического дефицита, но и структурно-функциональных изменений мозга, установленных с помощью КТ, УЗИ, электрофизиологических параметров (ЭЭГ), шкал изменений движения и чувствительности. Нами были дополнительно разработаны шкалы метаболических изменений. Собрав воедино критерии тяжести, полученные с помощью инструментальных методов исследования, мы создали шкалу оценки тяжести больных, перенесших острое нарушение мозгового кровообращения, в которой от качественного представления перешли к количественному, когда измененному параметру присваивается соответствующий балл. Таким образом, все показатели оценивали в баллах, их количество свидетельствовало о тяжести повреждения головного мозга. Это имеет не только научное значение, но и важное практическое применение – медицинское, социальное, экспертное (например, для определения ориентировочных сроков пребывания больного в стационаре, тактики лечения, реабилитационных мероприятий).
Таким образом, все показатели оценивали в баллах, их количество свидетельствовало о тяжести повреждения головного мозга. Это имеет не только научное значение, но и важное практическое применение – медицинское, социальное, экспертное (например, для определения ориентировочных сроков пребывания больного в стационаре, тактики лечения, реабилитационных мероприятий).
Особенностью разработанной шкалы стало то, что оценка степени тяжести состояния больных зависела не столько от клинических проявлений, сколько от глубины метаболических изменений. Мы разделили всех пациентов с высоким риском развития инсульта на три группы: к первой группе пациентов отнесли лиц, которые перенесли один ишемический инсульт; ко второй – перенесших несколько инсультов; к третьей – лиц без инсульта, но страдавших от повторных ТИА. Оказалось, что в третьей группе пациентов, которая клинически была отнесена к сравнительно легкой, развивались грубейшие нарушения метаболического характера, причем степень этих изменений обычно опережала таковую у пациентов первой группы (с одним инсультом в анамнезе). Лица, которые перенесли один инсульт, оказались наиболее подвержены терапии, у них были наилучшие результаты лечения. У лиц, перенесших более одного инсульта, после курса терапии также отмечались регресс симптоматики и улучшение общего состояния. В то же время пациенты, которые не имели ни одного инсульта, но подвергались ТИА, не всегда поддавались лечению; причиной тому было выраженное атеросклеротическое повреждение экстракраниальных сосудов.
Лица, которые перенесли один инсульт, оказались наиболее подвержены терапии, у них были наилучшие результаты лечения. У лиц, перенесших более одного инсульта, после курса терапии также отмечались регресс симптоматики и улучшение общего состояния. В то же время пациенты, которые не имели ни одного инсульта, но подвергались ТИА, не всегда поддавались лечению; причиной тому было выраженное атеросклеротическое повреждение экстракраниальных сосудов.
Состояние пациентов мы оценивали до курса лечения и после него. В схеме лечения применяли вазоактивные препараты, ноотропы и статины. Пациентов с выраженным стенозом общей сонной артерии направляли на консультацию к сосудистому хирургу.
Препарат Инстенон в клинической практике мы применяли внутримышечно по 2 мл 1 раз в сутки на протяжении 10 дней, затем перорально по 1-2 таблетки 3 раза в сутки в течение 35-40 дней. Уникальность действия Инстенона привлекает внимание специалистов давно: три составляющие препарата оказывают комплексное действие на головной мозг. Гексабендин за счет ноотропного эффекта способствует редукции кислородного голодания. Этамиван оказывает великолепное интегративное действие и восстанавливает функцию сознания. Этофиллин известен и кардиологам, и неврологам, так как оптимизирует работу миокарда, увеличивает сердечный выброс и таким образом улучшает центральную и периферическую гемодинамику. Эти эффекты очень важны, так как большинство больных с инсультами имеют кардиальную патологию, которая ухудшает течение заболевания. Инстенон позволяет комплексно воздействовать на всю сердечно-сосудистую систему.
Гексабендин за счет ноотропного эффекта способствует редукции кислородного голодания. Этамиван оказывает великолепное интегративное действие и восстанавливает функцию сознания. Этофиллин известен и кардиологам, и неврологам, так как оптимизирует работу миокарда, увеличивает сердечный выброс и таким образом улучшает центральную и периферическую гемодинамику. Эти эффекты очень важны, так как большинство больных с инсультами имеют кардиальную патологию, которая ухудшает течение заболевания. Инстенон позволяет комплексно воздействовать на всю сердечно-сосудистую систему.
В обязательный курс лечения больных после перенесенного инсульта в период реабилитации входил Актовегин. При комбинировании Актовегина и Инстенона при глубоких метаболических нарушениях (инсультах) метаболическое действие этих препаратов направлено главным образом на функции митохондрий, позволяя осуществлять спасительное в условиях ишемии анаэробное окисление глюкозы, способствуя восстановлению аэробного окисления в нейронах.
В комплексном лечении очень важно применение статинов. В настоящее время статины широко внедряются в неврологию благодаря уникальности своего действия: они обладают не только липидокорригирующим действием, но и положительным влиянием на эндотелиальную дисфункцию. Статины нормализуют липидный спектр крови, создают условия для адекватной агрегации, оптимальной гемодинамики.
При оценке нарушений липидного обмена нами использованы как плазменные, так и клеточные показатели. Липидно-плазменный обмен был нарушен у групп больных с одним инсультом, несколькими инсультами и с перенесенными ТИА приблизительно в одинаковой мере. Это говорит о высочайшем риске развития инсульта и системного атеросклероза у лиц, которые не перенесли инсульт, но имеют множественные ТИА. Кроме изучения плазменных показателей, которые давно используются в медицинской практике, мы впервые предложили метод гистохимического исследования липидосодержащих лейкоцитов крови, который применяли не только для оценки тяжести состояния лиц, перенесших острое нарушение мозгового кровообращения, но и с целью качественно-количественного определения липидного обмена. Внутриклеточные липиды в нейтрофилах являются важным фактором, регулирующим соотношение специфических и неспецифических иммунных сил организма. При гистохимическом исследовании липидосодержащих лейкоцитов мы обнаружили, что до лечения церебральной ишемии способность нейтрофилов к фагоцитозу была значительно ниже. Относительно неплохой компенсаторный коэффициент при гистохимическом исследовании липидосодержащих лейкоцитов определяли у пациентов, перенесших один инсульт (большинство больных активно лечились не только в остром периоде инсульта, но и после выписки из стационара). Наиболее низким коэффициент был у лиц, перенесших повторный инсульт. Больные с повторными ТИА, которые обычно не являются объектами длительного лечения, зачастую поздно обращаются к врачам, имели наиболее неблагоприятные изменения коэффициента фагоцитоза.
Внутриклеточные липиды в нейтрофилах являются важным фактором, регулирующим соотношение специфических и неспецифических иммунных сил организма. При гистохимическом исследовании липидосодержащих лейкоцитов мы обнаружили, что до лечения церебральной ишемии способность нейтрофилов к фагоцитозу была значительно ниже. Относительно неплохой компенсаторный коэффициент при гистохимическом исследовании липидосодержащих лейкоцитов определяли у пациентов, перенесших один инсульт (большинство больных активно лечились не только в остром периоде инсульта, но и после выписки из стационара). Наиболее низким коэффициент был у лиц, перенесших повторный инсульт. Больные с повторными ТИА, которые обычно не являются объектами длительного лечения, зачастую поздно обращаются к врачам, имели наиболее неблагоприятные изменения коэффициента фагоцитоза.
После проведенного курса полифакторного лечения Актовегином, Инстеноном и статинами в высоких дозах отмечали снижение атерогенности крови и повышение способности клеток к фагоцитозу.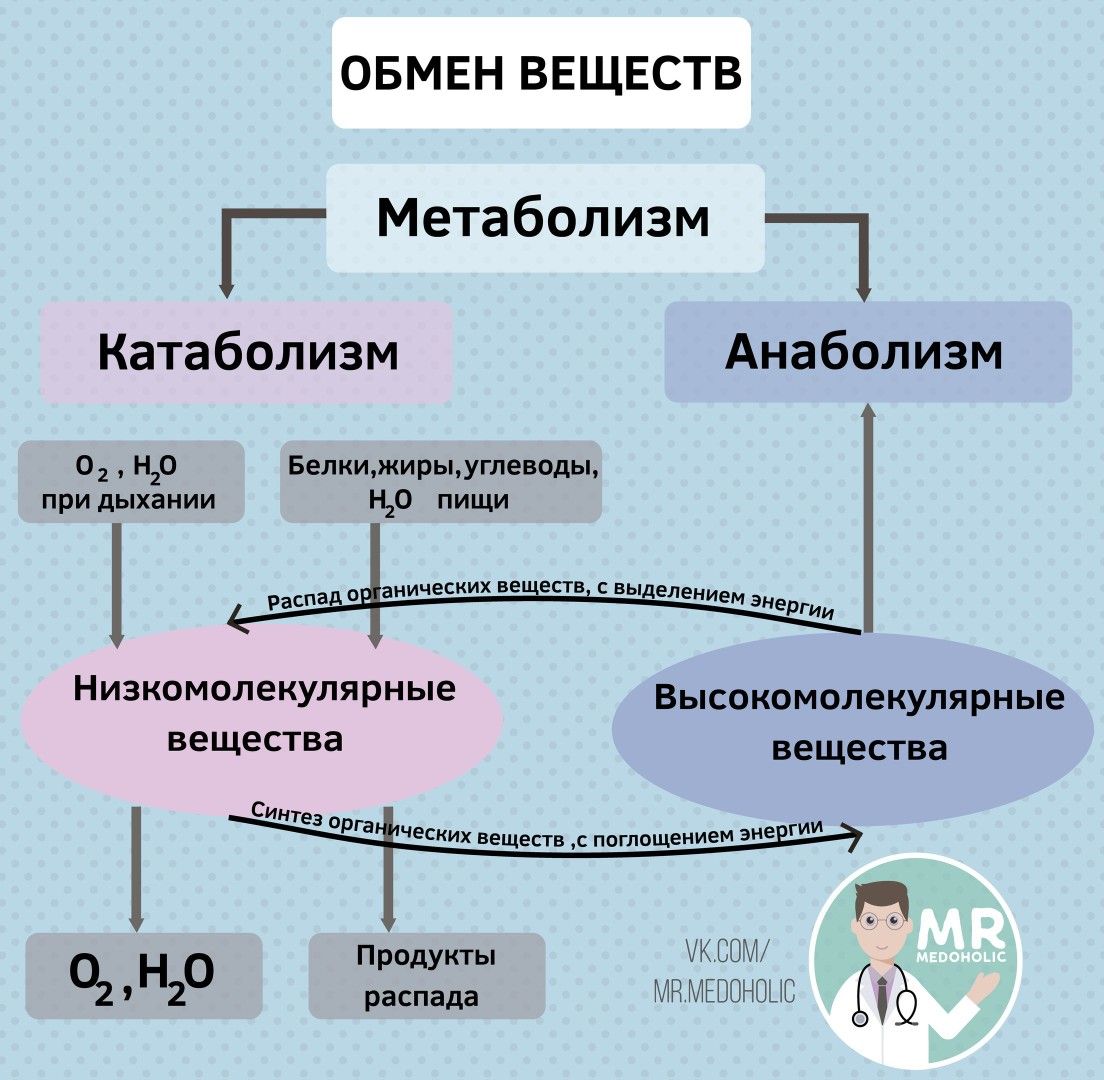
Помимо этих показателей, мы изучали состояние оксидантно-антиоксидантной системы, поскольку в настоящее время существуют неоспоримые доказательства, что одним из основных повреждающих факторов при ишемии является оксидантный стресс, который запускает серию патологических процессов с вовлечением всех видов обмена веществ – белкового, липидного и углеводного. В настоящее время принято считать, что самые агрессивные воздействия в структурно-молекулярной биологии оказывают свободные радикалы, атакующие клеточные мембраны. Мы использовали интегративный метод биохемолюминесценции, который дает возможность выявлять наиболее агрессивные перекиси и свободные радикалы. У всех больных с сосудистыми заболеваниями головного мозга преобладали агрессивные факторы над антиоксидантными. До лечения наибольшее содержание свободных радикалов определялось в группе больных, перенесших повторные инсульты. После лечения у больных наблюдалось относительное улучшение данных показателей, хотя далеко не полноценное.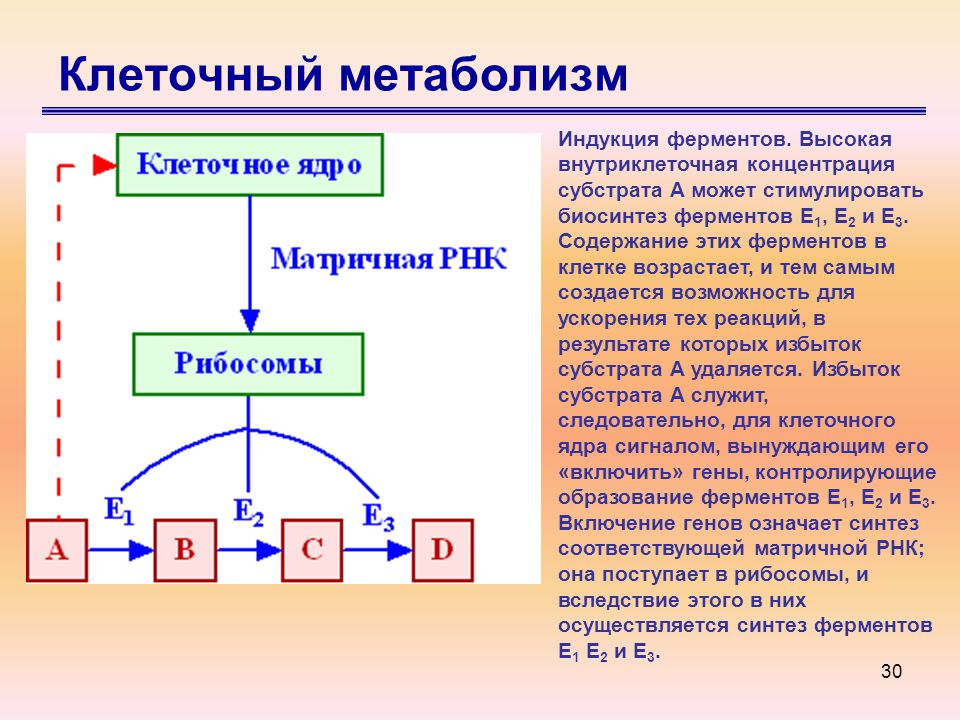 Это еще раз доказывает, что лица, перенесшие инсульт, требуют тщательного наблюдения и адекватного беспрерывного лечения даже в отдаленные сроки.
Это еще раз доказывает, что лица, перенесшие инсульт, требуют тщательного наблюдения и адекватного беспрерывного лечения даже в отдаленные сроки.
В качестве метода определения состояния физического здоровья и основы для рекомендаций по целесообразности того или иного комплекса терапевтических воздействий мы внедрили так называемый биофизический метод, с помощью которого определяли степень подвижности нативных клеток буккального эпителия при электрофорезе. С помощью этого метода можно судить об эффективности проведенного лечения, о степени тяжести больного в данный момент и необходимости включения в схему лечения тех или иных препаратов. Абсолютно у всех пациентов электроотрицательность клеток была ниже нормы. После лечения Актовегином, Инстеноном и статинами она повышалась, что сопровождалось улучшением состояния физического здоровья.
Таким образом, у больных, которые перенесли ТИА, выявлены более глубокие клинико-биохимические и энергетические нарушения, чем у лиц, перенесших один ишемический инсульт. Это связано со многими причинами: прежде всего, играет роль чрезмерное экстракраниальное сужение сосудов, питающих головной мозг; кроме того, у пациентов с неоднократными ТИА развивается дисциркуляторная энцефалопатия II и III стадии с микроструктурными нарушениями, которые приводят к выраженным метаболическим изменениям. Пациентов с ТИА необходимо выявлять скрининговыми методами исследования, включать в группу диспансерного наблюдения, они подлежат активному лечению. Однако в настоящее время такие пациенты зачастую попадают к врачу лишь после второй или третьей ишемической атаки, а порой и вообще не обращаются за медицинской помощью. Да и неврологи часто считают, что ТИА – намного менее значимая патология, чем инсульты, поэтому уделяют ее лечению и профилактике гораздо меньше внимания.
Это связано со многими причинами: прежде всего, играет роль чрезмерное экстракраниальное сужение сосудов, питающих головной мозг; кроме того, у пациентов с неоднократными ТИА развивается дисциркуляторная энцефалопатия II и III стадии с микроструктурными нарушениями, которые приводят к выраженным метаболическим изменениям. Пациентов с ТИА необходимо выявлять скрининговыми методами исследования, включать в группу диспансерного наблюдения, они подлежат активному лечению. Однако в настоящее время такие пациенты зачастую попадают к врачу лишь после второй или третьей ишемической атаки, а порой и вообще не обращаются за медицинской помощью. Да и неврологи часто считают, что ТИА – намного менее значимая патология, чем инсульты, поэтому уделяют ее лечению и профилактике гораздо меньше внимания.
Реабилитацию цереброваскулярных больных и подбор медикаментозных препаратов должны осуществлять только специалисты, в том числе кардиологи. Полученные нами данные позволяют патогенетически обосновать значение вазоактивного и ноотропного лечения в сочетании с гиполипидемической терапией. Для коррекции нарушений метаболического гомеостаза мы рекомендуем схему, включающую Актовегин, Инстенон и препарат из группы статинов. Многим пациентам также показана экстракраниальная эндартерэктомия, которая позволяет предотвратить грубые метаболические изменения головного мозга.
Для коррекции нарушений метаболического гомеостаза мы рекомендуем схему, включающую Актовегин, Инстенон и препарат из группы статинов. Многим пациентам также показана экстракраниальная эндартерэктомия, которая позволяет предотвратить грубые метаболические изменения головного мозга.
СТАТТІ ЗА ТЕМОЮ
27.01.2022 Психіатрія Терапія та сімейна медицина Погляд психіатра на постінсультні порушення у пацієнтівПоширеними наслідками інсульту мозку є виникнення післяінсультних розладів. Серед них найчастіше зустрічаються депресія, тривога, судинні когнітивні розлади та втома. На жаль, не завжди вдається своєчасно виявити ці порушення, що суттєво ускладнює реабілітацію та знижує якість життя хворих. У межах фахової школи «UkraineNeuroGlobal‑2021», що відбулася 23 вересня 2021 р. в онлайн-форматі, д. мед. н., професорка кафедри медичної психології, психосоматичної медицини та психотерапії Національного медичного університету імені О.О. Богомольця (м. Київ) Олена Олександрівна Хаустова розглянула постінсультні розлади у пацієнтів із погляду психіатра.
…
Серед них найчастіше зустрічаються депресія, тривога, судинні когнітивні розлади та втома. На жаль, не завжди вдається своєчасно виявити ці порушення, що суттєво ускладнює реабілітацію та знижує якість життя хворих. У межах фахової школи «UkraineNeuroGlobal‑2021», що відбулася 23 вересня 2021 р. в онлайн-форматі, д. мед. н., професорка кафедри медичної психології, психосоматичної медицини та психотерапії Національного медичного університету імені О.О. Богомольця (м. Київ) Олена Олександрівна Хаустова розглянула постінсультні розлади у пацієнтів із погляду психіатра.
…
Дефіцит когнітивної сфери – актуальна проблема сьогодення через зниження якості життя пацієнтів, активної комунікації в соціумі та потребу в сторонній допомозі при догляді. На особливу увагу в умовах нинішньої пандемії COVID‑19 заслуговує нова стратегія щодо когнітивних порушень.
…
На особливу увагу в умовах нинішньої пандемії COVID‑19 заслуговує нова стратегія щодо когнітивних порушень.
…
ПЭТ-исследование – для чего?
ПЭТ-исследования широко применяются при онкологических заболеваниях и позволяют оценить распространенность опухолевого процесса.
ПЭТ/КТ-исследование до (сверху) и после лучевой и химио- терапии (снизу).
Полученная при ПЭТ исследовании информация о характере метаболических процессов в опухолях дает возможность целенаправленно воздействовать на их жизнедеятельность, повышает избирательность и точность предстоящего лечения.
Уточнение объема активной опухолевой ткани позволяет максимально сократить отрицательное влияние на здоровые клетки организма при выборе лучевых методов лечения.
ПЭТ позволяет на ранних сроках выявлять рецидивы злокачественных опухолей и дифференцировать их с посттерапевтическими изменениями (в частности, постлучевыми некрозами и фиброзами).
Метаболическая активность опухоли отображается в виде количественной оценки уровня накопления различных РФП и измеряется в единицах SUV (Standardized Uptake Values – стандартизированный уровень накопления, чаще используются максимальные значения). Динамика изменения уровня накопления РФП в опухолях и их метастазах до лечения и в процессе терапии позволяет с высокой достоверностью оценивать эффективность лечения.
Вследствие разнородности метаболических изменений внутри самой опухоли различные участки злокачественных новообразований могут отличаться по уровню включения РФП. Исходя из этого, ПЭТ позволяет выявлять наиболее агрессивные участки опухоли для выбора оптимального места для биопсии.
Полученная информация даст возможность Вашему лечащему врачу в кратчайшие сроки установить диагноз и правильно спланировать лечение.
В нашем отделении исследования проводятся на современной аппаратуре в режиме ПЭТ/КТ. Полученные в ходе исследования данные значительно сокращают общее время диагностики и позволяют адекватно корректировать проводимую терапию.
Особенно важное значение метод имеет при раке легких, опухолях желудочно-кишечного тракта, раке простаты, нейроэндокринных опухолях, опухолях головного мозга, а также при лимфомах. При солидных опухолях (опухоли в паренхиматозных органах, таких как легкие, печень, простата, почки, головной мозг, поджелудочная железа, яичники и др.) результаты ПЭТ могут использоваться для планирования лечения на роботизированной стереотаксической радиохирургической установке, представленной на территории РОНЦ имени Н.Н.Блохина кабинетом «КиберНож».
Болезни XXI века. Что мы приобрели вместе со сменой образа жизни
Век технологий упростил жизнь человека. Но принес с собой и новые заболевания, о которых раньше даже не слышали, а также омолодил проблемы со здоровьем, которые называли болезнями стариков.
Стремительное развитие медицинской техники позволило в начавшемся столетии еще лучше изучить тело человека. Так, например, офтальмологи благодаря современным линзам смогли обнаружить ранние признаки кератоконуса (заболевания, связанного с изменением формы роговицы глаза), о котором раньше практически ничего не знали, отмечает хирург-офтальмолог Татьяна Шилова.
Но есть и заболевания, которые мы, наоборот, приобрели в век новых технологий. Столичные медики выделили три основных повлиявших на это фактора: пассивный образ жизни, обновившийся рацион питания и хронический стресс.
Метаболический синдром
По словам врача-терапевта Надежды Чернышовой, бич нового времени – метаболический синдром, с которым в XX в. не сталкивались. Гиподинамия, а также преобладание в рационе жиров и сахаров, особенно простых сахаров в виде сладкой газировки, печенья, конфет, булок и белого хлеба, привели к нарушению обмена сахаров в организме.
«Эти нарушения откладывают жир в области талии, во внутренние органы, и в конечном итоге развиваются сердечно-сосудистые заболевания и сахарный диабет. Это встречается очень часто, и особенно часто в последнее время – у молодежи», – уверяет Чернышова.
Диабет второго типа
Как отмечает Алексей Федоров, врач-кардиохирург, заведующий операционным отделением Центра сердечно-сосудистой хирургии ГВКГ им. Бурденко, еще 30 лет назад диабет второго типа развивался у людей в 50–60 лет. Сейчас выявление заболевания у сорокалетних – не редкость. Причина – также в ожирении и пассивном образе жизни.
Бурденко, еще 30 лет назад диабет второго типа развивался у людей в 50–60 лет. Сейчас выявление заболевания у сорокалетних – не редкость. Причина – также в ожирении и пассивном образе жизни.
«Это вообще основные факторы риска всех сердечно-сосудистых заболеваний, которые у нас в стране прочно удерживают первое место среди причин смерти», – рассказал Алексей Федоров. Среди прочих факторов кардиохирург выделил перегрузку организма алкоголем.
Болезнь стариков
Еще одна болезнь современности – нарушение пуринового обмена и, как следствие, – подагра. Раньше считавшаяся болезнью стариков, сегодня она стала чаще встречаться как раз у молодых и людей средних лет, говорит врач-диетолог, нутрициолог Евгений Арзамасцев.
«Даже если болезнь еще никак себя визуально не проявляет, мы стали часто замечать в своей практике у пациентов в анализах крови повышенный уровень мочевой кислоты. А это говорит о том, что нарушение пуринового обмена уже есть», – указывает Арзамасцев.
Более молодой возраст болезни специалист объяснил тем, что ожирение и нарушение обмена веществ вследствие именно неправильного питания, а не возрастных факторов стали встречаться у молодых людей гораздо чаще, чем в прошлом столетии. Еще 5–7 лет назад ситуация была лучше, считает диетолог.
Синдром карпального канала
Это заболевание связано с регулярным использованием компьютерной мыши. Синдром карпального канала – это заболевание рук, характеризующееся защемлением срединного нерва на уровне запястья.
По словам врача-невролога, остеопата Александра Евдокимова, болезнь появляется вследствие постоянного давления на кисть.
«Когда мы кладем на стол руку и очень долго пользуемся мышкой, за счет контакта руки и стола происходит ограничение кровотока и начинает страдать нерв. Развитие болезни – процесс не быстрый, но за счет длительности и регулярности нагрузки нервная ткань не успевает к ней адаптироваться, что и приводит к защемлению», – рассказал специалист.
При синдроме карпального канала может происходить онемение кистей рук, слабость, невозможность удерживать в руке даже легкие предметы, а в более запущенных случаях – появление болевых спазмов по ночам.
Это болезнь молодых. «Для людей 60 плюс обычно она не свойственна, так как они зачастую не успели еще перестроиться в этот ритм беспрерывного пользования компьютером», – заключил Евдокимов.
Пандемия близорукости
«Мы научились пользоваться техникой, однако пока не научились делать это в безопасном режиме», – считает хирург-офтальмолог Татьяна Шилова. Это, по ее мнению, стало одной из причин стремительного роста количества людей с близорукостью в текущем столетии.
Среди главных факторов не генетического развития болезни – работа с гаджетами в неправильном положении и на близком расстоянии. «У нас глазная мышца не расслабляется. А глаз устроен таким образом, что должен постоянно менять фокусное расстояние», – пояснила Шилова.
Также повлияла кропотливость работы, например постоянное чтение документов с экрана. Проблема заключается в том, что человек при этом очень редко моргает, говорит офтальмолог. Это приводит к компьютерному зрительному синдрому, который характеризуется дополнительной нагрузкой на глазную мышцу.
Проблема заключается в том, что человек при этом очень редко моргает, говорит офтальмолог. Это приводит к компьютерному зрительному синдрому, который характеризуется дополнительной нагрузкой на глазную мышцу.
Негативно влияет мерцание экрана. «Наш глаз привык к другой картинке, более приглушенным тонам, поэтому, конечно, работая за компьютером, нужно ставить правильные режимы защиты экрана, настраивать контрасты в зависимости от времени суток, однако так мало кто заморачивается», – рассказывает Шилова.
Эти факторы плюс редкое пребывание при естественном освещении, которое полезно для глаза, приводят к развитию близорукости даже у людей, не имеющих генетической предрасположенности к заболеванию, заключила терапевт. И сегодня это болезнь большинства.
Молодые неврозы
Увеличение скорости жизни и постоянная информационная перегрузка привели к развитию повышенной тревожности у значительной части населения, считает невролог Евдокимов. С его слов, это одна из главных проблем XXI в. , которая страшна тем, что все чаще проявляется у людей молодого и подросткового возраста.
, которая страшна тем, что все чаще проявляется у людей молодого и подросткового возраста.
«Следствием повышенной тревожности являются неврозы. И в таком состоянии сегодня находятся практически все люди. Нервная система пребывает в постоянном перенапряжении. И подавляющее большинство просто не может с ним справиться, адаптироваться. Как следствие, мы видим широкий спектр психосоматических расстройств и, что самое главное, молодых онкологических заболеваний, причину которых можно объяснить именно длительным негативным психоэмоциональным состоянием», – указывает Евдокимов.
По словам врача, сегодня неврозы требуют особого внимания. Пандемия коронавируса, ее непредсказуемость, изоляция и недостаток живого общения усугубили ситуацию, особенно среди подростков.
Что такое метаболический синдром?
В настоящее время сердечно-сосудистые заболевания (инфаркт миокарда, инсульт и т. д.) и сахарный диабет 2 типа лидируют среди причин смертности, поэтому профилактика этих заболеваний — важная проблема современности. В основе профилактики любого заболевания лежит борьба с факторами риска. Термин метаболический синдром применяется в медицине именно с целью раннего выявления и устранения факторов риска сердечно-сосудистых заболеваний и диабета.
В основе профилактики любого заболевания лежит борьба с факторами риска. Термин метаболический синдром применяется в медицине именно с целью раннего выявления и устранения факторов риска сердечно-сосудистых заболеваний и диабета.
Нарушения, входящие в рамки метаболического синдрома, длительно остаются незамеченными, часто начинают формироваться в детском и юношеском возрасте, неизбежно приводя к атеросклеротическим заболеваниям, диабету, артериальной гипертонии. Зачастую пациентам с ожирением, «немного» повышенным уровнем глюкозы, артериальным давлением на верхней границе нормы не уделяется должного внимания. Только когда эти факторы риска перейдут в серьезное заболевание, пациент удостаивается внимания здравоохранения.
Важно, чтобы факторы риска выявлялись и корригировались как можно раньше, до того, как они приведут к сердечно-сосудистым катастрофам. В этом огромную роль играет введение и применение такого понятия как метаболический синдром.
Метаболический синдром — это комплекс изменений, связанный с нарушением обмена веществ. Гормон инсулин перестает восприниматься клетками и не выполняет свои функции. В таком случае развивается инсулинорезистентность или нечувствительность к инсулину, что приводит к нарушению усвоения клетками глюкозы, а также патологическим изменениям всех систем и тканей. На сегодняшний день, согласно 10-му международному классификатору болезней, метаболический синдром не считается отдельным заболеванием. Это состояние, когда организм одновременно страдает от четырех болезней (нарушений):
Гормон инсулин перестает восприниматься клетками и не выполняет свои функции. В таком случае развивается инсулинорезистентность или нечувствительность к инсулину, что приводит к нарушению усвоения клетками глюкозы, а также патологическим изменениям всех систем и тканей. На сегодняшний день, согласно 10-му международному классификатору болезней, метаболический синдром не считается отдельным заболеванием. Это состояние, когда организм одновременно страдает от четырех болезней (нарушений):
- гипертонии
- ожирения
- ишемической болезни сердца
- сахарного диабета 2 типа
В развитых странах, где большинство населения ведет малоподвижный образ жизни, 10–25% людей старше 30 лет страдают от данных нарушений. В старшей возрастной группе показатели возрастают до 40%. Так в Европе количество больных превысило 50 млн человек. За ближайшие четверть века заболеваемость повысится на 50%. За последние два десятилетия количество больных среди детей и подростков увеличилось до 6,5%. Эту тревожную статистику связывают с пристрастием к углеводной диете.
Эту тревожную статистику связывают с пристрастием к углеводной диете.
Метаболический синдром поражает преимущественно мужчин. Женщинам эта болезнь грозит во время и после менопаузы. У представительниц слабого пола после 50-ти лет риск развития метаболического синдрома возрастает в 5 раз. К сожалению, современная медицина не в состоянии вылечить метаболический синдром. Однако есть и хорошие новости. Большинство изменений, развившихся в результате метаболического синдрома, обратимы. Грамотное лечение, правильное питание и здоровый образ жизни помогают надолго стабилизировать состояние.
Причины метаболического синдрома
Инсулин в организме выполняет множество функций. Но его основная задача — связаться в инсулиночувствительными рецепторами, которые есть в оболочке каждой клетки. После этого запускается механизм транспортировки глюкозы из межклеточного пространства внутрь клетки. Таким образом, инсулин «открывает двери» в клетку для глюкозы. Если рецепторы не реагируют на инсулин, то и сам гормон и глюкоза накапливаются в крови. В основе развития метаболического синдрома лежит нечувствительность к инсулину — инсулинорезистентность. Это явление может вызываться рядом причин.
В основе развития метаболического синдрома лежит нечувствительность к инсулину — инсулинорезистентность. Это явление может вызываться рядом причин.
1. Генетическая предрасположенность. У некоторых людей нечувствительность к инсулину заложена на генетическом уровне. Ген, который несет ответственность за развитие метаболического синдрома, находится в 19 хромосоме. Его мутации могут привести к тому, что
- у клеток недостаточно рецепторов, отвечающих за связывание инсулина;
- рецепторы не чувствительны к инсулину;
- иммунная система вырабатывает антитела, которые блокируют инсулиночувствительные рецепторы;
- поджелудочная железа вырабатывает аномальный инсулин.
2. Диета с высоким содержанием жиров и углеводов — самый важный фактор развития метаболического синдрома. Насыщенные жирные кислоты, поступающие с животными жирами в большом количестве, способствуют развитию ожирения. Кроме того жирные кислоты вызывают изменения в оболочках клеток, делая их нечувствительными к действию инсулина. Чрезмерно калорийное питание приводит к тому, что в кровь поступает много глюкозы и жирных кислот. Их избыток откладывается в жировых клетках в подкожной жировой клетчатке, а также в других тканях. Это приводит к снижению их чувствительности к инсулину.
Чрезмерно калорийное питание приводит к тому, что в кровь поступает много глюкозы и жирных кислот. Их избыток откладывается в жировых клетках в подкожной жировой клетчатке, а также в других тканях. Это приводит к снижению их чувствительности к инсулину.
3. Малоподвижный образ жизни. Уменьшение физической активности влечет за собой снижение скорости всех обменных процессов, в том, числе и расщепления и усвоения жиров. Жирные кислоты блокируют транспорт глюкозы в клетку и снижают чувствительность ее мембраны к инсулину.
4. Длительная не леченная артериальная гипертония. Вызывает нарушение периферического кровообращения, которое сопровождается снижением чувствительности тканей к инсулину.
5. Пристрастие к низкокалорийным диетам. Если калорийность суточного рациона составляет менее 300 ккал, это приводит к необратимым нарушениям обмена веществ. Организм «экономит» и наращивает запасы, что приводит к усиленному жироотложению.
6. Длительные психические нагрузки и стресс нарушают нервную регуляцию органов и тканей. В результате нарушается производство гормонов, в том числе инсулина, и реакция клеток на них.
7. Приём препаратов-антагонистов инсулина:
- глюкагон;
- кортикостероиды;
- пероральные контрацептивы;
- тиреоидные гормоны.
Эти лекарственные препараты уменьшают поглощение тканями глюкозы, что сопровождается снижением чувствительности к инсулину.
8. Передозировка инсулина при лечении сахарного диабета. Неправильно подобранное лечение приводит к тому, что в крови находится большое количество инсулина. Это вызывает привыкание рецепторов. Инсулинорезистентность в этом случае является своеобразной защитной реакцией организма от высокой концентрации инсулина.
9. Гормональные нарушения. Жировая ткань является эндокринным органом и выделяет гормоны, которые снижают чувствительность к инсулину. Причем, чем более выражено ожирение, тем ниже чувствительность. У женщин при повышенной выработке тестостерона и сниженной эстрогена жиры накапливаются по «мужскому» типу, нарушается работа сосудов и развивается артериальная гипертензия. Снижение уровня гормонов щитовидной железы также способно вызвать повышение уровня липидов (жиров) в крови и развитие инсулинорезистентности.
Причем, чем более выражено ожирение, тем ниже чувствительность. У женщин при повышенной выработке тестостерона и сниженной эстрогена жиры накапливаются по «мужскому» типу, нарушается работа сосудов и развивается артериальная гипертензия. Снижение уровня гормонов щитовидной железы также способно вызвать повышение уровня липидов (жиров) в крови и развитие инсулинорезистентности.
10. Возрастные изменения у мужчин. С возрастом снижается выработка тестостерона, что приводит к инсулинорезистентности, ожирению и гипертонии.
11. Апное во сне. Задержка дыхания во сне вызывает кислородное голодание мозга и усиленную выработку соматотропного гормона. Это вещество способствуют развитию нечувствительности к инсулину.
Симптомы метаболического синдрома
Рассмотрим все критерии метаболического синдрома.
1. Главный и обязательный критерий — абдоминальное ожирение, т. е. ожирение, при котором жировая ткань откладывается преимущественно в области живота. Иногда такое ожирение называют ожирением «по типу яблока» или «андроидным». Жировая клетчатка скапливается не только под кожей, но и вокруг внутренних органов. Она не только сдавливает их, затрудняя их работу, но и играет роль эндокринного органа. Жир выделяет вещества, способствующие появлению воспаления, повышению уровня фибрина в крови, что повышает риск развития тромбов. Отложение жира преимущественно в области бедер и ягодиц («по типу груши», «гиноидное») не имеет столь неблагоприятных последствий и в качестве критерия метаболического синдрома не рассматривается. Определить абдоминальное ожирение очень просто, достаточно измерить объем талии на уровне середины расстояния между краями реберных дуг и подвздошных костей. Для европеоидной нации показателями абдоминального ожирения являются объем талии у мужчин более 94–102 см, у женщин — более 80–88 см. Для азиатской популяции показатель ожирения у мужчин более строгий — объем талии более 90 см, а для женщин также — более 80 см.
Иногда такое ожирение называют ожирением «по типу яблока» или «андроидным». Жировая клетчатка скапливается не только под кожей, но и вокруг внутренних органов. Она не только сдавливает их, затрудняя их работу, но и играет роль эндокринного органа. Жир выделяет вещества, способствующие появлению воспаления, повышению уровня фибрина в крови, что повышает риск развития тромбов. Отложение жира преимущественно в области бедер и ягодиц («по типу груши», «гиноидное») не имеет столь неблагоприятных последствий и в качестве критерия метаболического синдрома не рассматривается. Определить абдоминальное ожирение очень просто, достаточно измерить объем талии на уровне середины расстояния между краями реберных дуг и подвздошных костей. Для европеоидной нации показателями абдоминального ожирения являются объем талии у мужчин более 94–102 см, у женщин — более 80–88 см. Для азиатской популяции показатель ожирения у мужчин более строгий — объем талии более 90 см, а для женщин также — более 80 см.
2. Артериальная гипертензия диагностируется, если систолическое артериальное давление более или равно 130 мм. рт. ст., диастолическое более или равно 85 мм. рт. ст. Красные пятна на груди и шее. Это признаки повышения давления связанного со спазмом сосудов, который вызван избытком инсулина.
Артериальная гипертензия диагностируется, если систолическое артериальное давление более или равно 130 мм. рт. ст., диастолическое более или равно 85 мм. рт. ст. Красные пятна на груди и шее. Это признаки повышения давления связанного со спазмом сосудов, который вызван избытком инсулина.
3. Нарушения липидного спектра. Для диагностики потребуется бихимический анализ крови: определение уровня триацилглицеридов и холестерина липопротеинов высокой плотности.
4. Нарушением углеводного обмена считается уровень глюкозы крови натощак более 5,6 ммоль/л, или лечение сахароснижающими препаратами.
Диагностика метаболического синдрома
При необходимости лечащий врач назначит дополнительное обследование:
— суточный мониторинг артериального давления, ЭКГ-исследование, ультразвуковое исследование сердца и сосудов, определение биохимических показателей уровня липидов крови, исследования функции печени и почек, определение глюкозы крови через 2 часа после еды или после проведения орального глюкозтолерантного теста.
Лечение метаболического синдрома
Лечение метаболического синдрома заключается в ведении здорового образа жизни и медикаментозной терапии. Изменение образа жизни означает изменение питания, режима физических нагрузок и отказ от вредных привычек. Фармакотерапия (назначение лекарственных препаратов) не будет иметь эффекта, если пациент не соблюдает правила питания и режима физической активности.
Рекомендации по питанию при метаболическом синдроме
— Крайне не рекомендуются чрезмерно строгие диеты и голодание. Снижение массы тела должно быть постепенным (на 5–10% за первый год). При быстрых темпах снижения массы тела пациенту трудно удержать полученный результат, практически всегда потерянные килограммы также быстро возвращаются.
— Более полезным и эффективным будет изменение состава питания: снижение употребления животных жиров, замена животных жиров растительными, увеличение потребления растительной клетчатки, волокон и снижение потребления поваренной соли.
— Следует практически полностью исключить сладкие газированные напитки, кондитерские изделия, фаст-фуд.
— Употребление хлеба лучше ограничить до 150–200 граммов в день.
— Супы должны быть преимущественно овощные.
— Из мясных продуктов лучше выбрать нежирные сорта говядины, птицы или рыбу в отварном или заливном виде.
— Из круп лучше использовать гречневую и овсяную, возможны также рисовая, пшённая, перловая, крупы, манную лучше максимально ограничить.
— Картофель, морковь, свёклу, рекомендуется употреблять не более 200 граммов в день. Овощи, богатые клетчаткой (помидоры, огурцы, болгарский перец, капусту, салат, редис, кабачки) и зелень возможно употреблять практически без ограничений в сыром и варёном или печёном виде.
— Яйца рекомендуется использовать не более 1–2 штуки в день.
— Фрукты и ягоды можно употреблять до 200–300 граммов в день.
— Молоко минимальной жирности, нежирные кисломолочные продукты и творог — 1–2 стакана в день. Сливки, жирные сорта сыров, сметану рекомендуется употреблять изредка.
Сливки, жирные сорта сыров, сметану рекомендуется употреблять изредка.
— Из напитков разрешены чай, некрепкий кофе в умеренных количествах, томатный сок, компоты и соки из ягод и фруктов кислых сортов, лучше домашнего приготовления без сахара.
Борьба с вредными привычками: ограничение алкоголя, отказ от курения.
Список рекомендованных продуктов:
- нежирные сорта мяса животных (телятина, кролик, курица без шкурки) и птицы 150–200 г в сутки;
- рыба и морепродукты 150 г;
- яйца — 1–2 в день в виде омлета или сваренные вкрутую;
- нежирные молочные продукты;
- творог 100–200 г;
- нежирные и неострые сорта твердого сыра — ограниченно до 30 г;
- нежирные говяжьи сосиски или вареная колбаса 2 раза в неделю;
- овощи 25% в сыром виде, остальные в тушеном, вареном, запеченном, приготовленном на пару (не менее 400 г).
Рекомендованы зеленые овощи;
- фрукты и ягоды несладкие сорта до 400 г.
 Свежие, замороженные или консервированные без сахара.
Свежие, замороженные или консервированные без сахара. - квашеная капуста, промытая водой;
- каши из перловой, гречневой, ячной крупы, бурый рис. 150–200 г на порцию с условием ограничения хлеба;
- первые блюда (250–300 г) на слабом обезжиренном мясном, рыбном или грибном бульоне, вегетарианские супы;
- хлеб из муки грубого помола, изделия с отрубями до 200 г;
- чаи, фруктовые и овощные соки без сахара;
- полоска черного шоколада, желе и муссы заменителях сахара;
- потребление жидкости ограничивают до 1,5 л. Это вызывает усиленный распад жиров в организме.
 Свежие, замороженные или консервированные без сахара.
Свежие, замороженные или консервированные без сахара.Необходимо отказаться от таких продуктов:
- кондитерские изделия: конфеты, печенье, изделия с кремом;
- выпечка, особенно вредна сдобная и слоеная;
- жирные сорта мяса: свинина, баранина, утка;
- консервы, копченое мясо и рыба, колбаса, ветчина;
- рис, манная и овсяная крупы, макароны;
- сливки, сладкий йогурт, жирный творог и изделия из него;
- маргарин, кулинарный жир;
- изюм, бананы, виноград, финики и другие сладкие фрукты;
- майонез, жирные острые соусы, пряности;
- сладкие газированные напитки, соки и нектары с сахаром.
1 раз в 1–2 недели можно делать выходной и умеренно употреблять «нежелательные» продукты.
Рекомендации по режиму физических нагрузок при метаболическом синдроме
Рекомендуется постепенное увеличение физической активности. Следует предпочесть такие виды спорта как ходьба, бег, гимнастика, плавание. Главное, чтобы физические нагрузки были регулярными и соответствовали вашим возможностям.
Медикаментозное лечение метаболического синдрома
Фармакотерапия метаболического синдрома направлена на лечение ожирения, нарушений углеводного обмена, артериальной гипертонии и дислипидемии.
Лечением метаболического синдрома занимаются врачи-эндокринологи. Но учитывая, что в организме больного происходят разнообразные патологические изменения, может потребоваться консультация: терапевта, кардиолога, диетолога.
Профилактика метаболического синдрома
Что нужно делать?
- Питайтесь правильно. Ешьте 5–6 раз в день небольшими порциями. У вас не должно возникать чувство голода. В противном случае, организм, получая питательные вещества, откладывает их про запас, что способствует развитию ожирения.
 Ешьте 5–6 раз в день небольшими порциями. У вас не должно возникать чувство голода. В противном случае, организм, получая питательные вещества, откладывает их про запас, что способствует развитию ожирения.
Ешьте 5–6 раз в день небольшими порциями. У вас не должно возникать чувство голода. В противном случае, организм, получая питательные вещества, откладывает их про запас, что способствует развитию ожирения.- Больше двигайтесь. Это помогает сжигать калории и нормализует обмен веществ. Используйте каждую возможность проявить физическую активность: ходите на работу пешком, поднимайтесь по ступенькам, мойте пол руками, а не шваброй.
- Купите абонемент в тренажерный зал или бассейн. Потраченные деньги будут вас стимулировать посещать занятия.
- Массаж и самомассаж. Эта процедура нормализует кровообращение в тканях, особенно в нижних конечностях, что делает клетки более чувствительными к воздействию инсулина.
- Пройдите курс физиотерапии: барокамера, миостимуляция, криотерапия, грязелечение прекрасно ускоряют обмен веществ. Направление в физкабинет можно получить у вашего лечащего врача.

- Гирудотерапия. Лечение пиявками улучшает все характеристики крови и нормализует кровоток, что важно для поддержания метаболизма в организме.
- Следите за уровнем холестерина. После 40 лет не реже чем раз в год проверяйте уровень «хорошего» и «плохого» холестеринов, чтобы при необходимости своевременно начать лечение.
Почему возникают нарушения обмена веществ и что с ними делать
Статью можно послушать. Если вам так удобнее, включайте подкаст:
О метаболизме мы задумываемся в основном тогда, когда не получается похудеть. И по большей части волнуемся лишь о его скорости. Медленный — плохо, от него заводятся лишние кило. Быстрый — прекрасно: ешь и худеешь!
Между тем обмен веществ — процесс куда более сложный, чем кажется. Он описывается не только скоростью, но и качеством.
Как работает обмен веществ
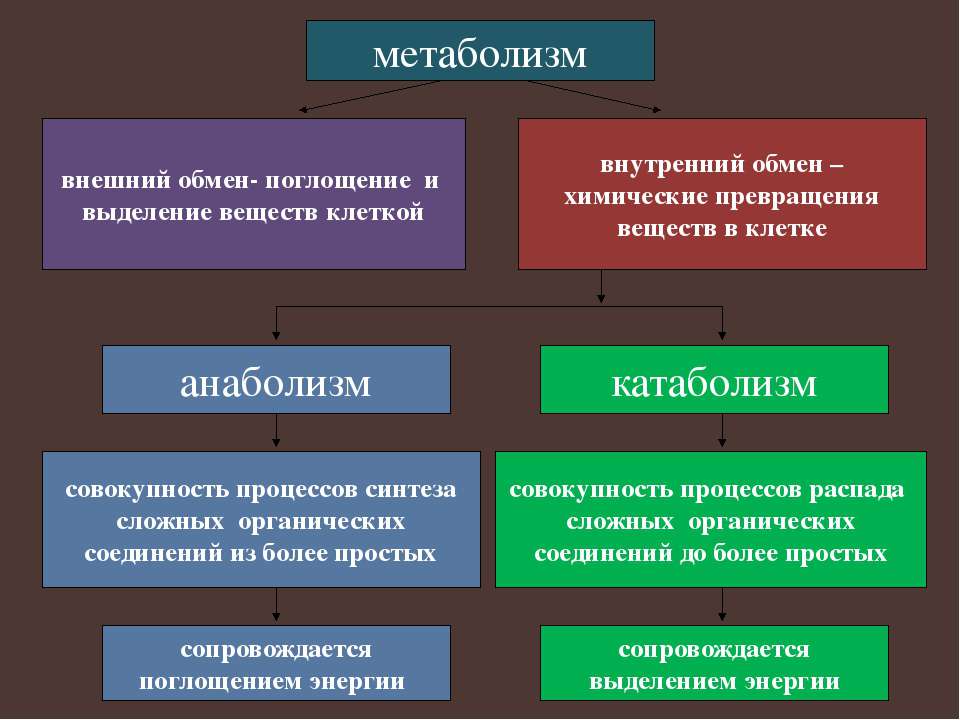
В общих чертах метаболизм — это совокупность физико-химических процессов , с помощью которых организм превращает пищу в необходимую для жизни энергию.
Если говорить более подробно, процесс выглядит примерно так. Пища состоит из белков, жиров, углеводов, витаминов, микроэлементов и минералов. Эти вещества расщепляются ферментами в пищеварительной системе, поступают в кровь и с её помощью переносятся к каждой клеточке тела.
Клетки реагируют на сигнал «кушать подано», открываются, пропускают необходимое питание, с помощью кислорода сжигают (окисляют) его, а в процессе получают энергию. Именно она позволяет клеткам функционировать, а организму в целом — дышать, двигаться, думать, жить.
Это ещё не всё. В результате окисления в клетке образуются продукты распада: углекислый газ, вода, молочная кислота, разнообразные соли. Все они возвращаются в кровь, а та транспортирует их к органам выделительной системы: печени, почкам, лёгким.
Так работает обмен веществ в норме.
Что такое нарушение обмена веществ
Это сбой на любом этапе метаболизма. Например, в пищеварительной системе может не хватать ферментов, чтобы расщепить пищу и обеспечить доставку питания в кровь. Или в крови может оказаться слишком мало кислорода, из-за чего клетки не смогут окислить питательные вещества и получить нужное количество энергии. А может, клетки, к которым кровь принесёт питание, не примут его. Или, положим, не полностью выведут продукты распада и начнут отравлять сами себя.
Или в крови может оказаться слишком мало кислорода, из-за чего клетки не смогут окислить питательные вещества и получить нужное количество энергии. А может, клетки, к которым кровь принесёт питание, не примут его. Или, положим, не полностью выведут продукты распада и начнут отравлять сами себя.
Вариантов нарушений — сотни. При любом из них клетки получают слишком мало или слишком много тех или иных веществ. И это серьёзно сказывается на их работоспособности, а в итоге — на нашем самочувствии и здоровье.
Как распознать нарушения обмена веществ
Человеческое тело очень чувствительно к любым нарушениям метаболизма. Недостаток питания или избыток продуктов распада затрагивает клетки всех органов и тканей, включая мозг, нервные окончания, мышцы, сердце и сосуды, кожу, печень, эндокринные железы и так далее. Кроме того, он влияет на выработку гормонов, управляющих процессами во всём организме. Поэтому симптомы могут быть самыми разными: от постоянной тяги к сладкому до заболеваний внутренних органов.
Тем не менее предположить нарушение обмена веществ даже на начальной стадии всё же можно. Вот самые распространённые симптомы:
- быстрая утомляемость;
- упадок сил, хроническая усталость;
- нарушения менструального цикла у женщин;
- беспричинный (на первый взгляд, естественно) набор или потеря веса;
- ухудшения памяти и когнитивных способностей;
- ухудшение состояния кожи (появление акне и раздражений) и ногтей, ломкость и выпадение волос;
- сердечная аритмия, скачки давления;
- перепады настроения;
- постоянные запоры или диарея;
- учащённое мочеиспускание;
- пониженная или повышенная температура, которая долго держится.
По отдельности каждый из этих симптомов может иметь собственное, неметаболическое обоснование. Но если вы наблюдаете у себя сразу несколько признаков, необходимо как можно быстрее обратиться к терапевту.
Обменные нарушения не обязательно что-то страшное: часто они легко корректируются. Но бывают случаи, когда сбои говорят о развитии серьёзного заболевания, например диабета. Важно этот сигнал не пропустить.
Но бывают случаи, когда сбои говорят о развитии серьёзного заболевания, например диабета. Важно этот сигнал не пропустить.
Почему происходит нарушение обмена веществ
Вот список наиболее распространённых причин метаболических сбоев:
- Генетическая предрасположенность. Возможно, ваше тело изначально не вырабатывает те или иные ферменты, необходимые для полноценного пищеварения. Или неправильно выводит продукты распада из клеток. А может, определённые продукты повреждают ворсинки в кишечнике, не позволяя тому пропускать переваренную пищу в кровь (на этом механизме основана, например, непереносимость глютена).
- Возраст. Годы меняют гормональный баланс. Например, во время климакса у женщин снижается выработка эстрогена. Это вызывает проблемы с обменом веществ.
- Паразиты. Глисты и патогенные микроорганизмы нарушают процесс преобразования пищи в энергию.
- Нервное перенапряжение. Стрессы — частые провокаторы нарушений обмена веществ.
- Вредные привычки. В частности, злоупотребление алкоголем и курением или страсть к сидяче-лежачему образу жизни.
- Диеты или нерегулярное питание. Они могут спровоцировать гипогликемию — состояние, характеризующееся постоянно низким уровнем сахара в крови, что тоже влияет на метаболизм.
- Некоторые перенесённые заболевания, в том числе вирусные — грипп, корь, ветрянка.
- Приём лекарственных препаратов, например оральных контрацептивов.
Что делать, если у вас нарушение обмена веществ
Главное — не разбрасываться диагнозами самостоятельно. Установить, что ваш метаболизм действительно дал сбой, может только терапевт. Для этого он заглянет в вашу медицинскую карту, расспросит об образе жизни, изменениях в самочувствии и внешности, предложит сдать анализы. В зависимости от результатов медик может отправить вас к эндокринологу или иному узкому специалисту.
Если причина нарушения обмена веществ кроется в болезни какого-то органа или генетической предрасположенности, схему лечение должен составить профессионал, учитывая ваши индивидуальные особенности.
Если же нарушения обмена веществ не связаны с генетикой и заболеваниями, их можно скорректировать самостоятельно. Для этих целей терапевт порекомендует вам внести некоторые изменения в образ жизни:
- Следите за тем, чтобы ваш рацион был полноценным. Овощи, фрукты, мясо и кисломолочные продукты помогут вам поддерживать необходимый баланс питательных веществ.
- Питайтесь регулярно. Завтрак, обед, ужин, пара перекусов между ними.
- Откажитесь от фастфуда и полуфабрикатов. Они заставляют нас накапливать жировую ткань, которая, в свою очередь, влияет на гормональный фон и в итоге на метаболизм в целом.
- Больше двигайтесь. Если нет времени на спортзал, хотя бы начните подниматься в офис и домой по лестнице, а не на лифте и по возможности больше гуляйте в выходные.
Кстати, этот же список мероприятий — эффективная профилактика нарушений обмена веществ.
Читайте также 🤕🧐😉
Лечение нарушений обмена веществ и профилактика сопутствующих заболеваний
С нарушением обмена веществ связаны многие проблемы в функционировании всех систем организма. Замедленный метаболизм ведет к набору лишнего веса и ожирению, к возникновению и развитию сахарного диабета и других эндокринных заболеваний, сердечно-сосудистым патологиям и многим другим нарушениям.
Замедленный метаболизм ведет к набору лишнего веса и ожирению, к возникновению и развитию сахарного диабета и других эндокринных заболеваний, сердечно-сосудистым патологиям и многим другим нарушениям.
К нарушению обмена веществ приводит совокупность нескольких факторов:
- наследственная предрасположенность;
- малоподвижный образ жизни, работа, связанная с постоянным сидением за компьютером;
- неправильное нерациональное питание: преобладание в рационе жирной пищи, быстрых углеводов, сладких газированных напитков, сдобы и сладостей;
- недостаточный непродуктивный сон, неправильный режим дня;
- вредные привычки – курение, излишнее употребление алкогольных напитков;
- отсутствие привычки к прогулкам на свежем воздухе, занятиям спортом;
- частые стрессы, постоянное нервное напряжение;
- некоторые заболевания эндокринной системы.
Санаторий «Красноусольск» имеет все необходимые ресурсы для коррекции метаболических нарушения и профилактики их возникновения. Нормализация режима дня и формирование привычки к здоровому образу жизни и правильному питанию, использование природных климатических факторов позволяют ускорить обменные процессы в организме и значительно улучшить самочувствие.
Нормализация режима дня и формирование привычки к здоровому образу жизни и правильному питанию, использование природных климатических факторов позволяют ускорить обменные процессы в организме и значительно улучшить самочувствие.
Метаболические нарушения успешно лечатся при помощи:
- минеральной воды, выводящей из организма соли и токсины;
- физиотерапии, направленной на ускорение кровообращения и лимфодренажа;
- грязелечения, улучшающего состояние сосудов и оказывающего общеукрепляющее действие;
- климатотерапии, активизирующей все процессы в организме.
Метаболический процесс – обзор
8.1 Введение
Цитата из известного учебника по биохимии [1] гласит: «Метаболизм – это общий процесс, посредством которого живые системы приобретают и используют свободную энергию для выполнения своих различных функций». Метаболизм осуществляется через метаболические пути: цепочки последовательных ферментативных реакций, которые производят определенные продукты для использования организмом. Как выяснили биологи и биохимики, существуют сотни таких «цепочек реакций», соединяющихся воедино многими сложными (и иногда недостаточно понятными) способами.Например, известно, что одна бактерия Escherichia coli имеет 600–700 метаболических реакций. Стандартные (био)химические диаграммы таких систем реакций для множественных клеточных реакций могут легко занимать диаграммы размером с стену через несколько стен. Чтобы получить образец этого (с относительно несложными диаграммами) в случае E. coli , перейдите на сайт эталонного пути Kegg http://www.genome.jp/kegg/pathway/map/map01100.html. Подобные диаграммы существуют для метаболизма клеток человека и животных, но они еще более сложны; часть показана ниже на рисунке 8.1. метаболитов в метаболическом пути обычно рассматриваются как субстраты, промежуточные продукты и реагенты в цепи реакций.
Метаболизм осуществляется через метаболические пути: цепочки последовательных ферментативных реакций, которые производят определенные продукты для использования организмом. Как выяснили биологи и биохимики, существуют сотни таких «цепочек реакций», соединяющихся воедино многими сложными (и иногда недостаточно понятными) способами.Например, известно, что одна бактерия Escherichia coli имеет 600–700 метаболических реакций. Стандартные (био)химические диаграммы таких систем реакций для множественных клеточных реакций могут легко занимать диаграммы размером с стену через несколько стен. Чтобы получить образец этого (с относительно несложными диаграммами) в случае E. coli , перейдите на сайт эталонного пути Kegg http://www.genome.jp/kegg/pathway/map/map01100.html. Подобные диаграммы существуют для метаболизма клеток человека и животных, но они еще более сложны; часть показана ниже на рисунке 8.1. метаболитов в метаболическом пути обычно рассматриваются как субстраты, промежуточные продукты и реагенты в цепи реакций.
Рисунок 8.1. Представление части метаболических путей метаболизма клеток человека и животных из http://www.genome.jp/kegg-bin/show_pathway?org_name=map&mapno=01100&mapscale=1.0&show_description=show.
Так почему интерес? Клеточный метаболизм представляет собой сложный набор химических реакций, которые позволяют клетке извлекать энергию и другие необходимые для жизни вещества из питательных веществ, а также строить новые структуры, необходимые для жизни и размножения.Хотя это может и не дать метафизического ответа на вопрос «Почему мы живы?» Метаболизм, безусловно, дает физический ответ на вопрос о том, как наши клетки, а следовательно, и мы сами, способны существовать, расти и, в конечном счете, что дает сбой и приводит к смерти. Изучение клеточного метаболизма лежит в основе многочисленных вопросов и фундаментальных исследований в области здоровья, таких как старение, а также в появляющихся второстепенных вопросах, таких как аутизм. Степень взаимосвязанности наших тел и окружающей среды посредством метаболических взаимодействий в клетках нашего кишечника и многих других клетках 1 , которые мы там размещаем, стала горячей темой исследований благодаря изучению микробиома (соотв. , metabalome), как и биосфера кишечника (соответственно полный профиль на метаболическом уровне). Такие исследования показывают, что широкий спектр современных заболеваний, таких как диабет, может быть результатом того, что метаболические процессы не функционируют должным образом, возможно, из-за отсутствия или дисбаланса того, что было эволюционно точно настроенным вкладом этих нечеловеческих клеток. в нашем убивающем микробы, существующем антибиотике, антибактериальном мыльном мире. См., например, [2] для недавнего доступного введения.
, metabalome), как и биосфера кишечника (соответственно полный профиль на метаболическом уровне). Такие исследования показывают, что широкий спектр современных заболеваний, таких как диабет, может быть результатом того, что метаболические процессы не функционируют должным образом, возможно, из-за отсутствия или дисбаланса того, что было эволюционно точно настроенным вкладом этих нечеловеческих клеток. в нашем убивающем микробы, существующем антибиотике, антибактериальном мыльном мире. См., например, [2] для недавнего доступного введения.
Даже если оставить в стороне такие новые спекуляции, метаболические процессы в течение некоторого времени корректировались и корректировались с пользой, и не только для лечения болезней. Метаболическая инженерия определяется как направленная модификация клеточного метаболизма и свойств путем введения, удаления и модификации метаболических путей с использованием рекомбинантной ДНК и других молекулярно-биологических инструментов. В настоящее время ведутся поиски «зеленых» альтернатив для многих соединений, производимых химическим путем с использованием нефти, а также более «зеленых» методов. Методы биохимического производства, заставляющие биологические организмы или компоненты производить избыточное производство определенных желаемых соединений, являются одной из таких альтернатив, достижимых с помощью метаболической инженерии.Некоторые из организмов, используемых в качестве производственных хозяев, включают E. coli , Mycobacterium tuberculosis , 2 и Saccharomyces cerevisiae (дрожжи). Большинство из нас хорошо знакомы с преимуществами метаболизма дрожжей, но, как еще один пример, процессы биохимической инженерии «скармливают» глюкозу и кукурузный экстракт E. coli или другим бактериям и производят в результате метаболических процессов янтарную кислоту, предшественник производства фармацевтических препаратов, тонких химикатов, биоразлагаемых полимеров и многого другого.Цель состоит в том, чтобы лучше понять множество метаболических путей в организмах и использовать эти знания для увеличения потока посредством полезных реакций (таким образом, производя больше продукции) или даже для обнаружения ранее не подозревавшихся цепочек реакций, которые могут производить желаемые метаболиты каким-либо другим способом.
Методы биохимического производства, заставляющие биологические организмы или компоненты производить избыточное производство определенных желаемых соединений, являются одной из таких альтернатив, достижимых с помощью метаболической инженерии.Некоторые из организмов, используемых в качестве производственных хозяев, включают E. coli , Mycobacterium tuberculosis , 2 и Saccharomyces cerevisiae (дрожжи). Большинство из нас хорошо знакомы с преимуществами метаболизма дрожжей, но, как еще один пример, процессы биохимической инженерии «скармливают» глюкозу и кукурузный экстракт E. coli или другим бактериям и производят в результате метаболических процессов янтарную кислоту, предшественник производства фармацевтических препаратов, тонких химикатов, биоразлагаемых полимеров и многого другого.Цель состоит в том, чтобы лучше понять множество метаболических путей в организмах и использовать эти знания для увеличения потока посредством полезных реакций (таким образом, производя больше продукции) или даже для обнаружения ранее не подозревавшихся цепочек реакций, которые могут производить желаемые метаболиты каким-либо другим способом. Полезные модели также позволили бы нам проверять альтернативные гипотезы, скажем, с помощью компьютера, с меньшими затратами, чем проведение нескольких экспериментов, и могли бы дать представление о том, какие типы экспериментов будут наиболее полезными.Можно также надеяться получить более глобальную перспективу, системную перспективу , которая, например, позволяет увидеть и предсказать последствия множества взаимосвязанных реакций на менее редукционистском уровне, чем реакция за реакцией.
Полезные модели также позволили бы нам проверять альтернативные гипотезы, скажем, с помощью компьютера, с меньшими затратами, чем проведение нескольких экспериментов, и могли бы дать представление о том, какие типы экспериментов будут наиболее полезными.Можно также надеяться получить более глобальную перспективу, системную перспективу , которая, например, позволяет увидеть и предсказать последствия множества взаимосвязанных реакций на менее редукционистском уровне, чем реакция за реакцией.
Как можно исследовать и понять эти взаимосвязанные реакции, эту сеть биохимических реакций систематическим и вычислительным способом? Что значит иметь такую систему (кроме рисования мультфильмов)? Для того чтобы клетка, а значит, и организм в целом находились в живом, процветающем состоянии, он вообще должен иметь возможность поддерживать некоторое равновесие (гомеостаз).Каждая реакция преобразует фиксированный набор входов в фиксированный набор выходов, но «поток» или «поток» через реакцию описывает, как происходит это преобразование или поток через реакцию. Если сосредоточить внимание на части этих биохимических реакций, формирующих интересующую (под)сеть, то в сбалансированном состоянии общая концентрация всех химических соединений в этой (под)системе не изменится. Каковы в таком состоянии цепочки реакций, для которых «общий поток», совокупная мера потока во всех реакциях, не изменяется, т.е.е., равно 0? Именно исследованию этого запроса и математической настройке путем применения стандартных инструментов линейной алгебры к так называемой стехиометрической матрице реакционной системы будет посвящена эта глава. Таким образом, у нас есть возможность увидеть, как можно построить математическую модель метаболических сетей и определенных путей внутри них. Математические модели, подобные этой, могут помочь нам понять, прояснить и сделать прогнозы относительно очень сложной внутренней работы клеточных процессов, общих для всех существ, которые изучаются клиницистами и врачами, биологами, химиками, инженерами и многими другими.
Если сосредоточить внимание на части этих биохимических реакций, формирующих интересующую (под)сеть, то в сбалансированном состоянии общая концентрация всех химических соединений в этой (под)системе не изменится. Каковы в таком состоянии цепочки реакций, для которых «общий поток», совокупная мера потока во всех реакциях, не изменяется, т.е.е., равно 0? Именно исследованию этого запроса и математической настройке путем применения стандартных инструментов линейной алгебры к так называемой стехиометрической матрице реакционной системы будет посвящена эта глава. Таким образом, у нас есть возможность увидеть, как можно построить математическую модель метаболических сетей и определенных путей внутри них. Математические модели, подобные этой, могут помочь нам понять, прояснить и сделать прогнозы относительно очень сложной внутренней работы клеточных процессов, общих для всех существ, которые изучаются клиницистами и врачами, биологами, химиками, инженерами и многими другими.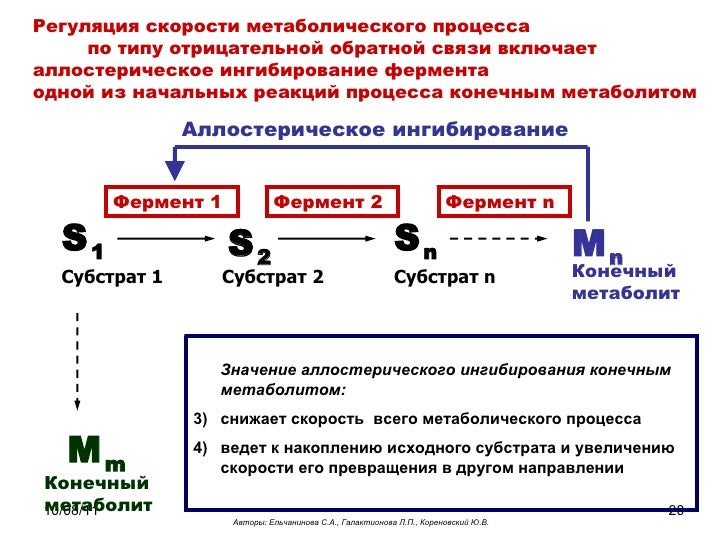
6.1C: Метаболические пути — Биология LibreTexts
- Последнее обновление
- Сохранить как PDF
- Ключевые моменты
- Ключевые термины
- Метаболические пути
- Анаболические пути
- Катаболические пути
- Важность ферментов
Катаболические пути требуют энергии и расщепления молекул.
Цели обучения
- Описать два основных типа метаболических путей
Ключевые моменты
- Метаболический путь представляет собой серию химических реакций в клетке, в ходе которых строятся и разрушаются молекулы для клеточных процессов.
- Анаболические пути синтезируют молекулы и требуют энергии.
- Катаболические пути расщепляют молекулы и производят энергию.
- Поскольку почти все метаболические реакции протекают неспонтанно, белки, называемые ферментами, помогают облегчить эти химические реакции.
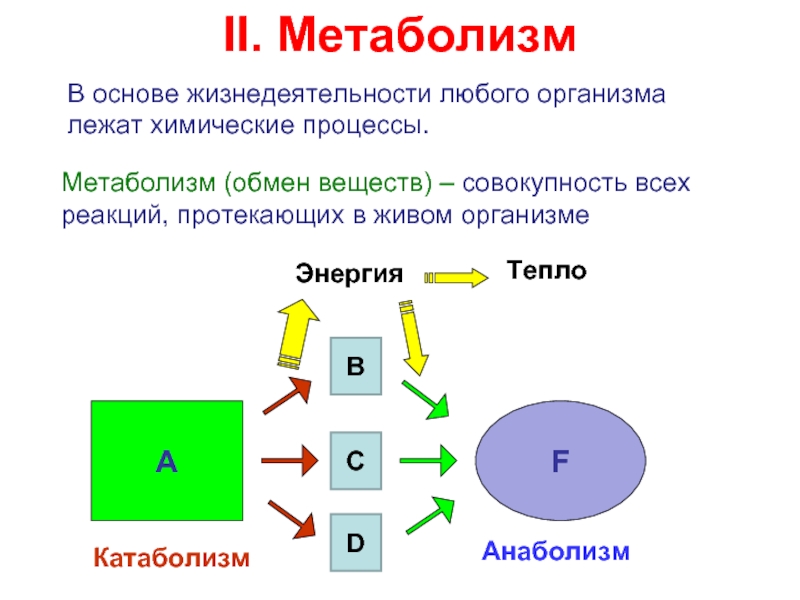
Основные термины
- катаболизм : деструктивный метаболизм, обычно включающий высвобождение энергии и расщепление материалов
- фермент : глобулярный белок, катализирующий биологическую химическую реакцию
- анаболизм : конструктивный метаболизм организма в отличие от катаболизма
Метаболические пути
Процессы создания и расщепления молекул углеводов иллюстрируют два типа метаболических путей.Метаболический путь представляет собой пошаговую серию взаимосвязанных биохимических реакций, которые превращают молекулу или молекулы субстрата через ряд метаболических промежуточных продуктов, в конечном итоге давая конечный продукт или продукты.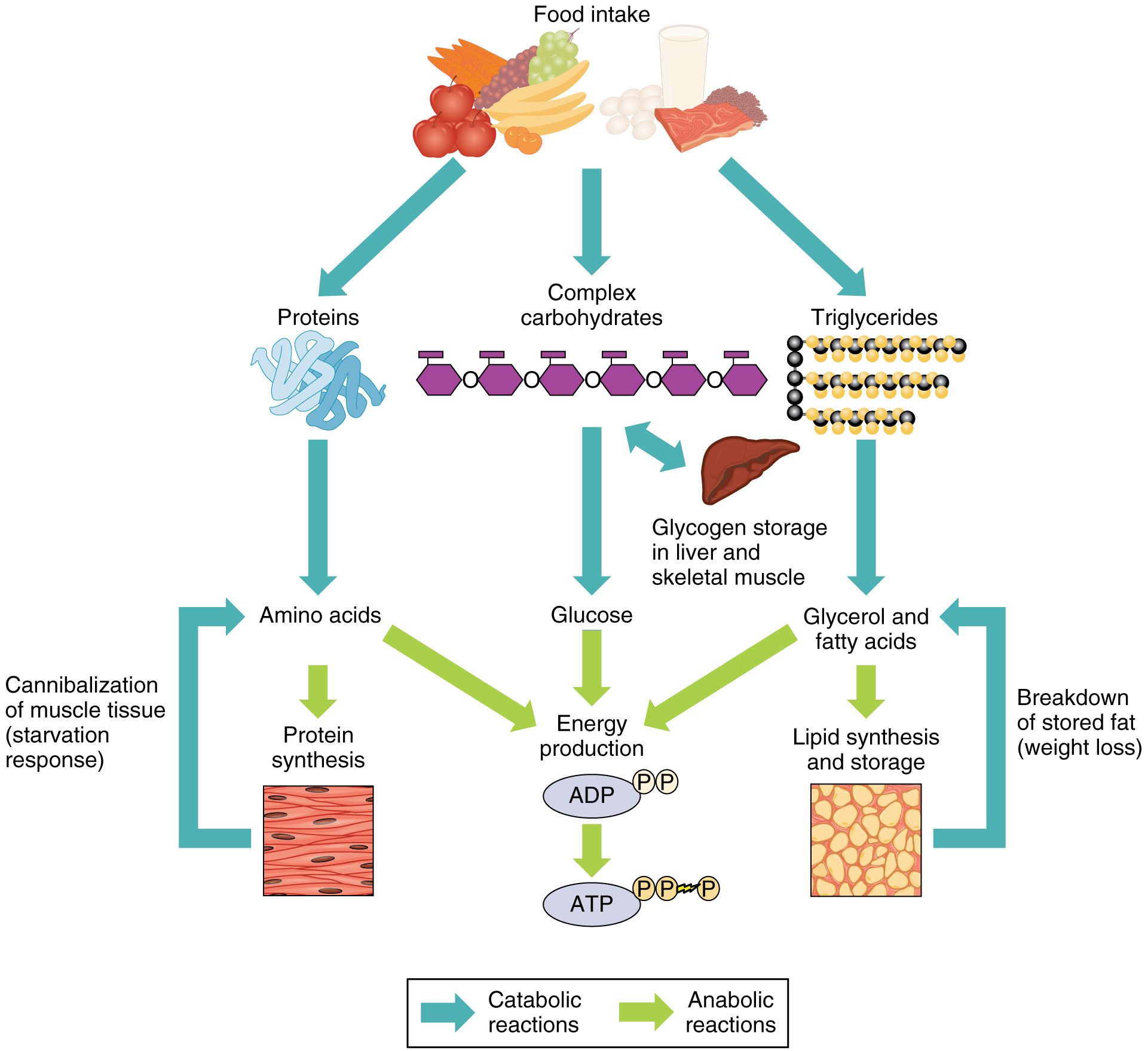 Например, один метаболический путь углеводов расщепляет большие молекулы до глюкозы. Другой метаболический путь может превращать глюкозу в большие молекулы углеводов для хранения. Первый из этих процессов требует энергии и называется анаболическим. Второй процесс производит энергию и называется катаболическим.Следовательно, метаболизм состоит из этих двух противоположных путей:
Например, один метаболический путь углеводов расщепляет большие молекулы до глюкозы. Другой метаболический путь может превращать глюкозу в большие молекулы углеводов для хранения. Первый из этих процессов требует энергии и называется анаболическим. Второй процесс производит энергию и называется катаболическим.Следовательно, метаболизм состоит из этих двух противоположных путей:
- Анаболизм (строительные молекулы)
- Катаболизм (расщепление молекул)
Анаболические пути
Анаболические пути требуют затрат энергии для синтеза сложных молекул из более простых. Одним из примеров анаболического пути является синтез сахара из CO 2 . Другие примеры включают синтез крупных белков из строительных блоков аминокислот и синтез новых цепей ДНК из строительных блоков нуклеиновых кислот. Эти процессы имеют решающее значение для жизни клетки, происходят постоянно и требуют энергии, обеспечиваемой АТФ и другими высокоэнергетическими молекулами, такими как НАДН (никотинамидадениндинуклеотид) и НАДФН.
Другие примеры включают синтез крупных белков из строительных блоков аминокислот и синтез новых цепей ДНК из строительных блоков нуклеиновых кислот. Эти процессы имеют решающее значение для жизни клетки, происходят постоянно и требуют энергии, обеспечиваемой АТФ и другими высокоэнергетическими молекулами, такими как НАДН (никотинамидадениндинуклеотид) и НАДФН.
Катаболические пути
Катаболические пути включают расщепление сложных молекул на более простые, высвобождая химическую энергию, хранящуюся в связях этих молекул. Некоторые катаболические пути могут улавливать эту энергию для производства АТФ, молекулы, используемой для питания всех клеточных процессов. Другие запасающие энергию молекулы, такие как липиды, также расщепляются в ходе аналогичных катаболических реакций с высвобождением энергии и образованием АТФ.
Важность ферментов
Химические реакции в метаболических путях редко протекают спонтанно.Каждый этап реакции облегчается или катализируется белком, называемым ферментом. Ферменты важны для катализа всех типов биологических реакций: как требующих энергии, так и высвобождающих энергию.
Ферменты важны для катализа всех типов биологических реакций: как требующих энергии, так и высвобождающих энергию.
Metabolic Pathways
В интерактивном режиме, показанном в верхней части страницы, мышечные клетки рассматриваются как один тип клеток. Но на самом деле у нас есть несколько типов мышц. Все мышцы похожи в том, что они сокращаются и могут осуществлять большинство одних и тех же метаболических реакций.Тем не менее, каждый тип отличается.
Сердце состоит из клеток сердечной мышцы. Эти клетки сжигают очень мало глюкозы. В качестве топлива они используют в основном жирные кислоты и не запасают гликоген.
Пласты гладкомышечных клеток окружают кровеносные сосуды, пищеварительный тракт и многие наши органы. Они работают вне нашего контроля и осознания, например, продвигают пищу по мере ее переваривания и регулируют кровяное давление. Клетки гладкой мускулатуры могут запасать гликоген и сжигать как глюкозу, так и жирные кислоты, как описано выше.
Скелетные мышцы являются наиболее распространенным типом. Эти мышцы прикрепляются к нашим костям и координируют все наши произвольные движения. Они хранят большую часть гликогена в организме и могут сжигать как глюкозу, так и жирные кислоты. Работающая мышечная клетка может пройти через АТФ примерно в 100 раз быстрее, чем мышца в состоянии покоя. По мере увеличения рабочей нагрузки скелетные мышцы начинают сжигать больше глюкозы.
Скелетные мышцы делятся на быстро сокращающиеся и медленно сокращающиеся клетки. У всех нас есть оба типа, но у людей они в разных пропорциях.Медленно сокращающиеся клетки более многочисленны у марафонцев, и они постоянно сжигают топливо в течение длительных периодов времени. Эти клетки заполнены митохондриями, и во время упражнений они склонны полностью расщеплять сахар на углекислый газ и воду.
Быстросокращающиеся клетки более многочисленны у спринтеров. В этих клетках меньше митохондрий, но они сокращаются сильнее, чем медленно сокращающиеся клетки. Быстросокращающиеся клетки работают короткими всплесками и производят большую часть своего АТФ за счет гликолиза.Вместо полного окисления глюкозы они расщепляют ее до трехуглеродного пирувата, а затем до лактата (также называемого молочной кислотой). Лактат может быть поглощен и сожжен сердечной мышцей и медленно сокращающимися мышечными клетками, или же он может быть поглощен печенью и снова соединен с образованием глюкозы.
Быстросокращающиеся клетки работают короткими всплесками и производят большую часть своего АТФ за счет гликолиза.Вместо полного окисления глюкозы они расщепляют ее до трехуглеродного пирувата, а затем до лактата (также называемого молочной кислотой). Лактат может быть поглощен и сожжен сердечной мышцей и медленно сокращающимися мышечными клетками, или же он может быть поглощен печенью и снова соединен с образованием глюкозы.
Типы мышечных клеток
Происхождение и эволюция метаболических путей: почему и как первичные клетки построили метаболические пути? | Эволюция: образование и просветительская деятельность
Увлекательно.Захватывающе. Реконструкция истории жизни на Земле представляет собой одну из самых интригующих проблем науки. И еще более интригующим является попытка понять (самые) первые молекулярные шаги, ведущие к первичным клеткам и их ранней эволюции. Существующие клетки представляют собой довольно сложные образования, состоящие из множества различных молекул, которые, однако, должны действовать и взаимодействовать согласованным образом, чтобы обеспечить выживание и размножение клеток (и многоклеточных организмов). В каждый момент жизни клетки миллиарды молекул превращаются в разные посредством реакций, которые ускоряются (катализируются) так называемыми ферментами, большая часть которых представлена белками. Несмотря на то, что эти белки могут взаимодействовать с множеством различных молекул во время своего хаотического путешествия внутри клетки, они связываются только со специфическими молекулами, представляющими их субстрат , и превращают его в другие и другие молекулы, называемые продуктом (реакции).В целом, это верно не для всех ферментов; каждый фермент взаимодействует с одним субстратом, образуя определенный продукт. Следовательно, в каждый момент жизни клетки млрд субстратов превращаются в млрд продуктов млрд молекул ферментов. Эти реакции чрезвычайно быстры, и мы можем представить себе клетку как вязкую среду, в которой эти реакции протекают упорядоченно (и только внешне хаотично). Совокупность этих реакций называется метаболизмом , круговой «сущностью» в том смысле, что молекулы могут быть разрушены (катаболизм) для получения энергии и «кирпичиков», необходимых для построения других различных молекул (анаболизм) (рис.
В каждый момент жизни клетки миллиарды молекул превращаются в разные посредством реакций, которые ускоряются (катализируются) так называемыми ферментами, большая часть которых представлена белками. Несмотря на то, что эти белки могут взаимодействовать с множеством различных молекул во время своего хаотического путешествия внутри клетки, они связываются только со специфическими молекулами, представляющими их субстрат , и превращают его в другие и другие молекулы, называемые продуктом (реакции).В целом, это верно не для всех ферментов; каждый фермент взаимодействует с одним субстратом, образуя определенный продукт. Следовательно, в каждый момент жизни клетки млрд субстратов превращаются в млрд продуктов млрд молекул ферментов. Эти реакции чрезвычайно быстры, и мы можем представить себе клетку как вязкую среду, в которой эти реакции протекают упорядоченно (и только внешне хаотично). Совокупность этих реакций называется метаболизмом , круговой «сущностью» в том смысле, что молекулы могут быть разрушены (катаболизм) для получения энергии и «кирпичиков», необходимых для построения других различных молекул (анаболизм) (рис. 1). Отсюда ясно, что внутри клетки существует «равновесие» между катаболическими и анаболическими реакциями. Таким образом, метаболизм современных клеток достаточно сложен, но его также можно считать предельно упорядоченным. На рисунке 2 показан пример катаболической (расщепление глюкозы во время гликолиза) и анаболической (биосинтез аминокислоты гистидина) систем. Как видно из рис. 2, и гликолиз, и биосинтез гистидина протекают как своего рода «каскад» реакций, где для разрушения глюкозы и образования гистидина требуется последовательное действие разных ферментов, каждый из которых способен катализировать один шаг этого каскада.Совокупность реакций, начинающихся с субстрата и ведущих к конечному продукту реакции, называется метаболическим путем . В большинстве случаев каждый этап метаболического пути катализируется одним ферментом, который (в трети случаев) представляет собой один белок, кодируемый одним геном (Holliday et al. 2011).
1). Отсюда ясно, что внутри клетки существует «равновесие» между катаболическими и анаболическими реакциями. Таким образом, метаболизм современных клеток достаточно сложен, но его также можно считать предельно упорядоченным. На рисунке 2 показан пример катаболической (расщепление глюкозы во время гликолиза) и анаболической (биосинтез аминокислоты гистидина) систем. Как видно из рис. 2, и гликолиз, и биосинтез гистидина протекают как своего рода «каскад» реакций, где для разрушения глюкозы и образования гистидина требуется последовательное действие разных ферментов, каждый из которых способен катализировать один шаг этого каскада.Совокупность реакций, начинающихся с субстрата и ведущих к конечному продукту реакции, называется метаболическим путем . В большинстве случаев каждый этап метаболического пути катализируется одним ферментом, который (в трети случаев) представляет собой один белок, кодируемый одним геном (Holliday et al. 2011).
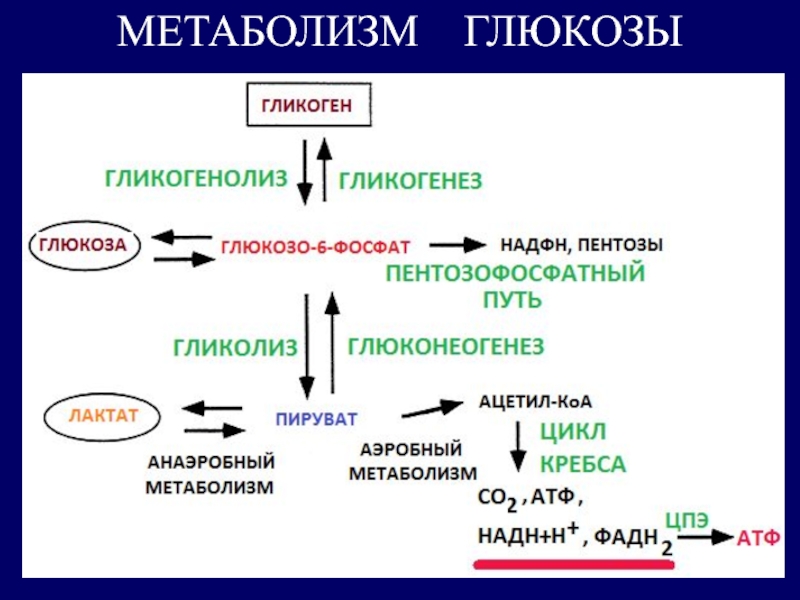 1
1 Схематическое изображение метаболических сетей, существующих в существующих клетках (из http://manet.illinois.edu/pathways.php)
Рис.2Схематическое изображение катаболического (гликолиз) (из http://www.genome.jp/dbget-bin/www_bget?pathway+hsa00010) ( a ) и анаболического пути (биосинтез гистидина) (из http: //www.genome.jp/kegg/pathway/map/map00340.html) ( b ) у гамма-протеобактерий Escherichia coli K12
Если предположить, что современные и очень сложные клетки произошли от гораздо более простых предковых клеток, также правдоподобно предположить, что последний имел более простой метаболизм по сравнению с существующим.Это, в свою очередь, означает, что они должны обладать гораздо более простыми геномами, состоящими, скорее всего, из нескольких сотен генов. Если это так, возникает вопрос: почему и как формировались первичные клетки и развивались их метаболические пути? Вопрос можно перефразировать следующим образом: почему и как ранние клетки увеличили количество своих генов и усложнили свои геномы? Ответ(ы), которые мы можем попытаться дать на эти вопросы, явно зависят от условий первобытной Земли и от того, как выглядели первобытные живые существа. Однако это один из самых туманных вопросов; на самом деле, хотя были предприняты значительные усилия, чтобы понять появление первых живых существ, мы до сих пор не знаем, когда и как возникла жизнь (Перето и др., 1998). Тем не менее принято считать, что ранние организмы возникли и заселили водные среды (океаны, реки, пруды и т. д.), богатые органическими соединениями, спонтанно образовавшиеся в добиотическом мире. Это гетеротрофное происхождение жизни обычно предполагается и часто упоминается как теория Опарина-Халдейна (Опарин, 1924; Ласкано и Миллер, 1996).Если эта идея верна, жизнь развилась из первобытного бульона, содержащего различные органические молекулы (многие из которых используются существующими формами жизни). Этот бульон из питательных соединений был доступен ранним гетеротрофным организмам, поэтому биосинтез у них был минимальный. Экспериментальное подтверждение этого предположения было получено в 1953 г., когда Миллер (Miller, 1953) и Юри показали, что аминокислоты и другие органические молекулы образуются в атмосферных условиях, которые, как считается, характерны для условий ранней Земли.
Однако это один из самых туманных вопросов; на самом деле, хотя были предприняты значительные усилия, чтобы понять появление первых живых существ, мы до сих пор не знаем, когда и как возникла жизнь (Перето и др., 1998). Тем не менее принято считать, что ранние организмы возникли и заселили водные среды (океаны, реки, пруды и т. д.), богатые органическими соединениями, спонтанно образовавшиеся в добиотическом мире. Это гетеротрофное происхождение жизни обычно предполагается и часто упоминается как теория Опарина-Халдейна (Опарин, 1924; Ласкано и Миллер, 1996).Если эта идея верна, жизнь развилась из первобытного бульона, содержащего различные органические молекулы (многие из которых используются существующими формами жизни). Этот бульон из питательных соединений был доступен ранним гетеротрофным организмам, поэтому биосинтез у них был минимальный. Экспериментальное подтверждение этого предположения было получено в 1953 г., когда Миллер (Miller, 1953) и Юри показали, что аминокислоты и другие органические молекулы образуются в атмосферных условиях, которые, как считается, характерны для условий ранней Земли.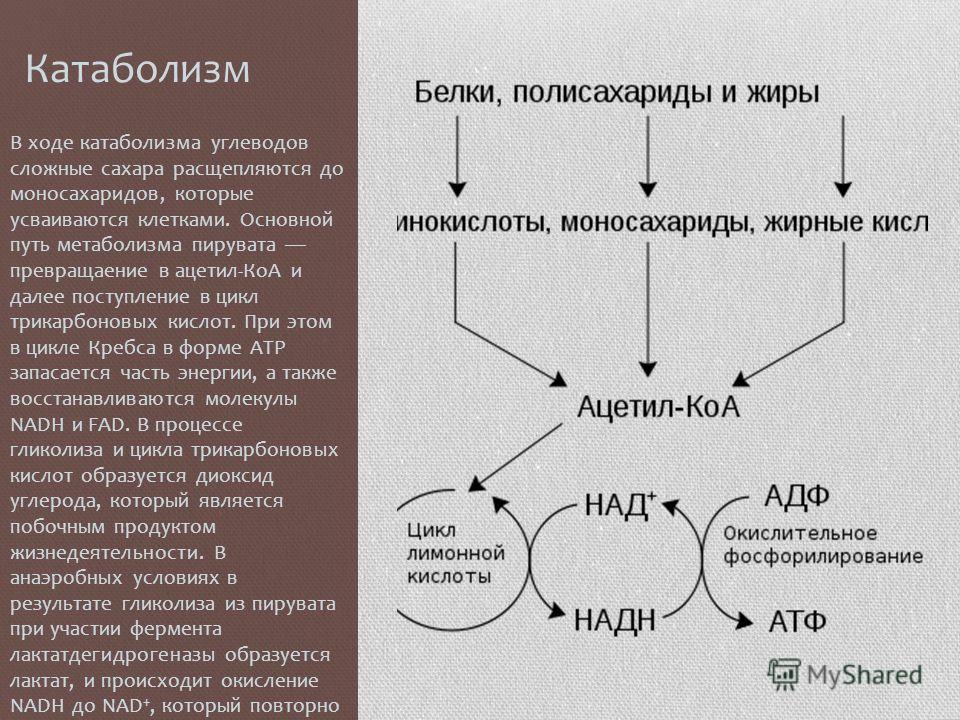 Первые живые системы, вероятно, произошли непосредственно от первобытного бульона и относительно быстро эволюционировали до общего предка, обычно называемого последним универсальным общим предком (LUCA), сущности, представляющей отправную точку расхождения всех существующих форм жизни на Земле. (Рис. 3). Если мы предположим, что жизнь возникла в пребиотическом бульоне, содержащем большинство, если не все, необходимых малых молекул, то можно предположить, что на первобытной Земле существовала большая потенциальная доступность питательных веществ, обеспечивающих как рост, так и снабжение энергией большого числа живых организмов. предковые организмы.Мы можем представить себе существование «раннего плавающего живого мира», состоящего из первичных клеток, которые могли бы выглядеть как «мыльные пузыри», включающие в себя одну или несколько информационных молекул и выполняющие ограниченное количество метаболических реакций. Эти пузыри были способны делиться, взаимодействовать друг с другом, сливаться и делиться своими геномами и метаболическими способностями, создавая все более сложные живые существа.
Первые живые системы, вероятно, произошли непосредственно от первобытного бульона и относительно быстро эволюционировали до общего предка, обычно называемого последним универсальным общим предком (LUCA), сущности, представляющей отправную точку расхождения всех существующих форм жизни на Земле. (Рис. 3). Если мы предположим, что жизнь возникла в пребиотическом бульоне, содержащем большинство, если не все, необходимых малых молекул, то можно предположить, что на первобытной Земле существовала большая потенциальная доступность питательных веществ, обеспечивающих как рост, так и снабжение энергией большого числа живых организмов. предковые организмы.Мы можем представить себе существование «раннего плавающего живого мира», состоящего из первичных клеток, которые могли бы выглядеть как «мыльные пузыри», включающие в себя одну или несколько информационных молекул и выполняющие ограниченное количество метаболических реакций. Эти пузыри были способны делиться, взаимодействовать друг с другом, сливаться и делиться своими геномами и метаболическими способностями, создавая все более сложные живые существа. Если этот сценарий верен, то есть первичные организмы были гетеротрофными и не нуждались в развитии новых и улучшенных метаболических способностей, поскольку большинство необходимых питательных веществ было доступно, мы можем вернуться к двум вопросам, которые можно решить, то есть . почему и как первичные клетки расширили свои метаболические способности и геномы?
Если этот сценарий верен, то есть первичные организмы были гетеротрофными и не нуждались в развитии новых и улучшенных метаболических способностей, поскольку большинство необходимых питательных веществ было доступно, мы можем вернуться к двум вопросам, которые можно решить, то есть . почему и как первичные клетки расширили свои метаболические способности и геномы?
Ориентировочная временная шкала эволюции от происхождения Земли до диверсификации жизни
Ответ на первый вопрос довольно интуитивен. В самом деле, увеличение количества ранних клеток, процветающих на первичном бульоне, должно было привести к истощению основных питательных веществ, создавая все более сильное селективное давление, которое, в свою очередь, благоприятствовало (в дарвиновском смысле) тем микроорганизмам, которые стали способны синтезировать эти молекулы. концентрация которых уменьшалась в первичном бульоне.Следовательно, происхождение и эволюция основных метаболических путей представляли собой решающий шаг в молекулярной и клеточной эволюции, поскольку они сделали первичные клетки менее зависимыми от экзогенных источников питательных веществ (рис. 4).
4).
Схематическое изображение наследственного клеточного сообщества с селективным давлением, позволяющим приобретать и распространять новый метаболический признак (с изменениями из Fondi et al. 2009a)
Но как происходило расширение геномов? В следующем разделе основное внимание будет уделено молекулярным механизмам, которые руководили этим переходом, т.е.э., расширение и уточнение наследственных метаболических путей, ведущих к структуре существующих метаболических путей.
Самый быстрый словарь в мире | Vocabulary.com
метаболический процесс органические процессы, необходимые для жизни
адиабатический процесс любой процесс, протекающий без поступления или потери тепла
 62″>
62″>математический процесс (математика) расчет математическими методами
метаболический алкалоз
патологический процесс органический процесс, возникающий как следствие болезни
нарушение обмена веществ нарушение или дефект обмена веществ
скорость метаболизма скорость метаболизма
геологический процесс естественный процесс, посредством которого изменяются геологические особенности
 6″>
6″>психический процесс выполнение некоторой сложной когнитивной деятельности
патологический процесс органический процесс, возникающий вследствие болезни
биологический процесс процесс, происходящий в живых организмах
шиловидный отросток отходит от основания височной кости
телесный процесс органический процесс, происходящий в организме
крыловидный отросток два костных отростка, отходящие от тела клиновидной кости
естественный процесс Процесс, существующий в природе или произведенный ею
 4″>
4″>геологический процесс естественный процесс, посредством которого изменяются геологические особенности
метаболизм Органические процессы, необходимые для жизни
фотожелатиновый процесс Процесс фотомеханической печати, в котором используется стеклянная пластина с желатиновой поверхностью, на которую нанесено воспроизводимое изображение; можно использовать с одним или несколькими цветами
Процесс Маркова простой случайный процесс, в котором распределение будущих состояний зависит только от текущего состояния, а не от того, как оно пришло в настоящее состояние
Синхронизация метаболических процессов у растений с толстянковокислым метаболизмом | Журнал экспериментальной ботаники
Аннотация
У растений с метаболизмом крассуловой кислоты прямое разделение процессов карбоксилирования, опосредованных фосфо енол пируваткарбоксилазой (PEPC) и Rubisco, оптимизирует фотосинтетические характеристики и накопление углерода в потенциально ограничивающих условиях. В этом обзоре рассматриваются механизмы, которые синхронизируют спрос и предложение на углерод при сохранении пластичности фотосинтеза в течение 24-часового цикла САМ. Циркадные часы играют центральную роль в контроле многих метаболических, транспортных и физиологических компонентов САМ. Уровень контроля, осуществляемого часами, может варьироваться от транскрипционной до посттрансляционной регуляции, в зависимости от рассматриваемых генов, белков и даже видов растений. Еще один уровень контроля обеспечивают метаболиты, в том числе органические кислоты и углеводы, которые обнаруживают существенные взаимные колебания содержания в течение дневного цикла.Обсуждаются механизмы, ответственные за определение содержания метаболитов, а также требования к сигнализации для координации потоков углерода. Эволюционные последствия рассматриваются с точки зрения того, как циркадный и метаболический контроль цикла САМ мог быть получен от растений C 3 .
В этом обзоре рассматриваются механизмы, которые синхронизируют спрос и предложение на углерод при сохранении пластичности фотосинтеза в течение 24-часового цикла САМ. Циркадные часы играют центральную роль в контроле многих метаболических, транспортных и физиологических компонентов САМ. Уровень контроля, осуществляемого часами, может варьироваться от транскрипционной до посттрансляционной регуляции, в зависимости от рассматриваемых генов, белков и даже видов растений. Еще один уровень контроля обеспечивают метаболиты, в том числе органические кислоты и углеводы, которые обнаруживают существенные взаимные колебания содержания в течение дневного цикла.Обсуждаются механизмы, ответственные за определение содержания метаболитов, а также требования к сигнализации для координации потоков углерода. Эволюционные последствия рассматриваются с точки зрения того, как циркадный и метаболический контроль цикла САМ мог быть получен от растений C 3 .
Поступила 28 октября 2003 г.; Принято 19 января 2004 г.
Введение
Метаболизм крассуловой кислоты представляет собой особый способ фотосинтетической ассимиляции углерода, развившийся в ответ на исключительные условия окружающей среды.На сегодняшний день известно, что примерно 7% видов растений, охватывающих 33 семейства и 328 родов, обладают способностью к САМ (Winter and Smith, 1996, и ). Такое таксономическое разнообразие отражается в различных средах обитания, предпочитаемых САМ-растениями: от засушливых пустынь до тропических лесов и водных экосистем. CAM представляет собой механизм концентрации CO 2 , который использует фермент фосфо енол пируваткарбоксилазу (PEPC) для улавливания CO 2 из дыхательных путей и атмосферы в ночное время.Физиологическое значение CAM заключается в сохранении углерода и воды в растениях, растущих в условиях, которые ограничивают доступность одного или обоих ресурсов на периодической или долгосрочной основе. В то время как ферментативный аппарат, необходимый для САМ, присутствует у всех высших растений, эволюция этого пути потребовала изменения регуляторной способности ключевых ферментов и транспортеров, чтобы поддерживать временное разделение процессов карбоксилирования, которые являются центральными для САМ. Целью этого обзора является рассмотрение механизмов, которые могут синхронизировать спрос и предложение на углерод в течение 24-часового цикла CAM.Молекулярные подходы и появляющиеся геномные ресурсы обеспечивают беспрецедентный потенциал для использования цикла САМ-диэля для выяснения компонентов циркадного контроля и контроля метаболитов, которые оптимизируют фотосинтез в потенциально ограничивающих и экстремальных условиях.
Целью этого обзора является рассмотрение механизмов, которые могут синхронизировать спрос и предложение на углерод в течение 24-часового цикла CAM.Молекулярные подходы и появляющиеся геномные ресурсы обеспечивают беспрецедентный потенциал для использования цикла САМ-диэля для выяснения компонентов циркадного контроля и контроля метаболитов, которые оптимизируют фотосинтез в потенциально ограничивающих и экстремальных условиях.
В сущности, CAM может экспрессироваться на фоне фотосинтеза C 3 посредством развертывания процессов ночного карбоксилирования и последующего дневного декарбоксилирования (Fig. 1a). Ночью, когда скорость эвапотранспирации мала, атмосферный СО 2 и/или дыхательный СО 2 фиксируются в цитозоле ферментом фосфо енол пируваткарбоксилазой (ФЭПК).Субстрат 3-C, фосфо--енол--пируват (ФЕП), образуется в результате гликолитического расщепления углеводов, образовавшихся в течение предыдущего дня. Конечный продукт 4-C, яблочная кислота, хранится в большой центральной вакуоли. В течение дня малат выходит из вакуоли, и декарбоксилирование может происходить за счет одиночного или комбинированного действия трех карбоксилаз (в зависимости от вида растений): НАДФ-яблочного фермента (НАДФ-МЭ), НАД-МЭ и фосфо--енол--пируваткарбоксикиназы ( ПЕПК). В дополнение к 3-C продуктам PEP или пирувату, CO 2 высвобождается при высоком внутреннем парциальном давлении ( p CO 2 ), которого часто бывает достаточно для закрытия устьиц и, таким образом, сохранения воды.Высокое содержание p CO 2 , образующееся при декарбоксилировании, также подавляет фотодыхание. Восстановление углеводов посредством глюконеогенеза требует больших энергетических затрат на этот путь, но обеспечивает производство субстрата для последующего ночного карбоксилирования и разделения для роста. Углеводы, которые обеспечивают субстраты для ночных реакций, транспортируются либо в хлоропласт и хранятся в виде крахмала, либо в вакуоли и сохраняются в виде сахарозы и/или гексозы, в зависимости от вида (Christopher and Holtum, 1996).
В течение дня малат выходит из вакуоли, и декарбоксилирование может происходить за счет одиночного или комбинированного действия трех карбоксилаз (в зависимости от вида растений): НАДФ-яблочного фермента (НАДФ-МЭ), НАД-МЭ и фосфо--енол--пируваткарбоксикиназы ( ПЕПК). В дополнение к 3-C продуктам PEP или пирувату, CO 2 высвобождается при высоком внутреннем парциальном давлении ( p CO 2 ), которого часто бывает достаточно для закрытия устьиц и, таким образом, сохранения воды.Высокое содержание p CO 2 , образующееся при декарбоксилировании, также подавляет фотодыхание. Восстановление углеводов посредством глюконеогенеза требует больших энергетических затрат на этот путь, но обеспечивает производство субстрата для последующего ночного карбоксилирования и разделения для роста. Углеводы, которые обеспечивают субстраты для ночных реакций, транспортируются либо в хлоропласт и хранятся в виде крахмала, либо в вакуоли и сохраняются в виде сахарозы и/или гексозы, в зависимости от вида (Christopher and Holtum, 1996). Что касается чистого потока углерода, результатом реакций, показанных на рис. 1а, являются существенные суточные и возвратно-поступательные колебания содержания малата и углеводов (рис. 1б), которые могут составлять до 20% сухой массы листа.
Что касается чистого потока углерода, результатом реакций, показанных на рис. 1а, являются существенные суточные и возвратно-поступательные колебания содержания малата и углеводов (рис. 1б), которые могут составлять до 20% сухой массы листа.
Несмотря на энергетические затраты, связанные с CAM, во многих случаях потенциал высокой производительности не подвергается риску. Агрономически важные виды САМ, включая ананас ( Ananas comosus ) и несколько агав, могут демонстрировать продуктивность, сравнимую с продуктивностью сахарного тростника (Bartholomew and Kadzimin, 1977; Nobel, 1996).Такие желательные атрибуты являются следствием пластичности, благодаря которой САМ может включаться или отключаться в ответ на прерывистые или долгосрочные (сезонные) возмущения окружающей среды. Факторы развития и окружающей среды (например, наличие воды, интенсивность света) сильно влияют на долю CO 2 , поглощаемую ночью через PEPC и непосредственно днем через Rubisco в растениях САМ (Cushman and Borland, 2002; Dodd et al . , 2002). Рост и продуктивность большинства САМ-растений максимальны, когда преобладает прямая дневная фиксация CO 2 через Rubisco (фаза IV газообмена).Как определяющая черта CAM, фотосинтетическая пластичность должна быть достигнута при сохранении синхронизации между процессами карбоксилирования, декарбоксилирования и транспорта, показанными на рис. 1a, чтобы минимизировать бесполезные циклы оборота углерода в течение диэльного цикла.
, 2002). Рост и продуктивность большинства САМ-растений максимальны, когда преобладает прямая дневная фиксация CO 2 через Rubisco (фаза IV газообмена).Как определяющая черта CAM, фотосинтетическая пластичность должна быть достигнута при сохранении синхронизации между процессами карбоксилирования, декарбоксилирования и транспорта, показанными на рис. 1a, чтобы минимизировать бесполезные циклы оборота углерода в течение диэльного цикла.
Циркадные часы задают фазы дня CAM
Хорошо задокументированные ритмы газообмена у видов рода Kalanchoë , выращенных в условиях постоянной темноты и CO 2 на воздухе, показали, что эндогенные циркадные часы играют кардинальную роль в установлении и синхронизации разделенных во времени метаболических процессов. компоненты CAM (Wilkins, 1992; Lüttge, 2000).Циркадный контроль потока углерода через PEPC обычно рассматривается как ключевой компонент, лежащий в основе разделения дневных и ночных процессов карбоксилирования, которые определяют CAM (Nimmo, 2000). PEPC активируется ночью посредством фосфорилирования остатка серина вблизи N-конца белка, что делает фермент более чувствительным к PEP и положительным эффекторам, глюкозе-6-P и триозе-P, и менее чувствительным к аллостерическому ингибитору, малату. (Рис. 2; Nimmo и др. ., 1986, 1987; Chollet и др. ., 1996). Таким образом, временные рамки, в течение которых PEPC остается активным, на что указывают модели ночного накопления малата, отражают изменения в явном состоянии фосфорилирования фермента, на что указывает [малат], необходимый для 50% ингибирования фермента (т.е. K и малат). Кроме того, степень фосфорилирования ФЭПК является основным фактором, определяющим количество CO 2 , поглощенного и хранящегося в виде малата в течение ночи у разных видов САМ, у которых экстрагируемая активность ФЭПК варьировала не более чем на 10% (рис.3).
PEPC активируется ночью посредством фосфорилирования остатка серина вблизи N-конца белка, что делает фермент более чувствительным к PEP и положительным эффекторам, глюкозе-6-P и триозе-P, и менее чувствительным к аллостерическому ингибитору, малату. (Рис. 2; Nimmo и др. ., 1986, 1987; Chollet и др. ., 1996). Таким образом, временные рамки, в течение которых PEPC остается активным, на что указывают модели ночного накопления малата, отражают изменения в явном состоянии фосфорилирования фермента, на что указывает [малат], необходимый для 50% ингибирования фермента (т.е. K и малат). Кроме того, степень фосфорилирования ФЭПК является основным фактором, определяющим количество CO 2 , поглощенного и хранящегося в виде малата в течение ночи у разных видов САМ, у которых экстрагируемая активность ФЭПК варьировала не более чем на 10% (рис.3).
Состояние фосфорилирования PEPC определяется наличием или отсутствием специализированной Ca 2+ -независимой Ser/Thr протеинкиназы, которая, в свою очередь, регулируется на уровне экспрессии генов циркадианным осциллятором (рис. 2; Carter и др. ., 1991; Hartwell и др. ., 1996, 1999; Taybi и др. ., 2000). Теоретически контроль активности киназы PEPC (PPCK) с помощью эндогенных часов должен обеспечивать предвидение фотопериода и быструю инактивацию PEPC в начале дня, тем самым избегая бесполезных циклов синтеза малата и декарбоксилирования.Однако экологическое преимущество такого циркадного контроля поглощения CO 2 менее очевидно, учитывая присущую САМ-растениям пластичность для модуляции потока углерода в ответ на изменение условий окружающей среды. Полевые измерения мгновенной дискриминации изотопов углерода у гемиэпифитных душителей рода Clusia показали, что PEPc может оставаться активным в течение 4–5 часов после рассвета (Borland et al , 1993; Roberts et al ). , 1997). На рисунке 3 показано, что время инактивации PEPC может различаться у разных видов САМ, даже если они выращиваются в одинаковых условиях окружающей среды.
2; Carter и др. ., 1991; Hartwell и др. ., 1996, 1999; Taybi и др. ., 2000). Теоретически контроль активности киназы PEPC (PPCK) с помощью эндогенных часов должен обеспечивать предвидение фотопериода и быструю инактивацию PEPC в начале дня, тем самым избегая бесполезных циклов синтеза малата и декарбоксилирования.Однако экологическое преимущество такого циркадного контроля поглощения CO 2 менее очевидно, учитывая присущую САМ-растениям пластичность для модуляции потока углерода в ответ на изменение условий окружающей среды. Полевые измерения мгновенной дискриминации изотопов углерода у гемиэпифитных душителей рода Clusia показали, что PEPc может оставаться активным в течение 4–5 часов после рассвета (Borland et al , 1993; Roberts et al ). , 1997). На рисунке 3 показано, что время инактивации PEPC может различаться у разных видов САМ, даже если они выращиваются в одинаковых условиях окружающей среды.
Метаболиты обеспечивают дополнительный уровень контроля над циклом САМ
Экспериментальные наблюдения показывают, что экспрессия и активность PPCK могут изменяться в зависимости от метаболического статуса листа. В листьях Kalanchoë daigremontiana , которые были предотвращены от накопления малата путем помещения в атмосферу N 2 в течение части или всей ночи, после переноса наблюдались резкие сдвиги в амплитуде и продолжительности поглощения CO 2 PEPC. окружающему воздуху (рис.4а; Borland и др. , 1999). Стимуляцию поглощения CO 2 через PEPC ночью и в начале дня объясняли сдвигами в величине активности PPCK (рис. 4b), которые контролируются на уровне экспрессии генов (Hartwell et al ., 1999; Borland и др. ., 1999). Наблюдения согласуются с мнением о том, что цитоплазматический малат (или родственный метаболит) вызывает ингибирование экспрессии гена PPCK по принципу обратной связи. Таким образом, у растений с ограниченным накоплением малата активность и экспрессия PPCK выше, чем в контроле, и наблюдается задержка в подавлении PPCK у растений с пониженным содержанием малата (рис.4; Borland и др. , 1999). Следовательно, предполагается, что циркадный контроль экспрессии гена PPCK в САМ является вторичным ответом на циркадные изменения транспорта малата через тонопластную мембрану вакуоли (Nimmo, 2000).
В листьях Kalanchoë daigremontiana , которые были предотвращены от накопления малата путем помещения в атмосферу N 2 в течение части или всей ночи, после переноса наблюдались резкие сдвиги в амплитуде и продолжительности поглощения CO 2 PEPC. окружающему воздуху (рис.4а; Borland и др. , 1999). Стимуляцию поглощения CO 2 через PEPC ночью и в начале дня объясняли сдвигами в величине активности PPCK (рис. 4b), которые контролируются на уровне экспрессии генов (Hartwell et al ., 1999; Borland и др. ., 1999). Наблюдения согласуются с мнением о том, что цитоплазматический малат (или родственный метаболит) вызывает ингибирование экспрессии гена PPCK по принципу обратной связи. Таким образом, у растений с ограниченным накоплением малата активность и экспрессия PPCK выше, чем в контроле, и наблюдается задержка в подавлении PPCK у растений с пониженным содержанием малата (рис.4; Borland и др. , 1999). Следовательно, предполагается, что циркадный контроль экспрессии гена PPCK в САМ является вторичным ответом на циркадные изменения транспорта малата через тонопластную мембрану вакуоли (Nimmo, 2000). Независимые доказательства в поддержку того, что мембрана тонопласта является «главным переключателем» циркадной регуляции САМ, были получены в результате компьютерного моделирования ритмов САМ, основанного на осмотических соображениях обмена малата и механизма натяжения/расслабления тонопласта (Lüttge, 2000, 2002 a ).Поскольку сдвиги в дневном/ночном содержании малата являются отличительной чертой пластичности САМ, метаболитный контроль фосфорилирования PEPC обеспечит эффективное средство точной настройки поглощения CO 2 в течение дневного/ночного цикла в зависимости от изменений условий окружающей среды. . Такая интеграция циркадных сигналов и сигналов окружающей среды может обеспечить основу для синхронизации и пластичности метаболизма, присущей САМ. Однако такая гипотеза поднимает дополнительные вопросы в отношении того, сколько метаболических компонентов цикла САМ напрямую связаны с циркадианным осциллятором, а сколько зависит от дневных и ночных потоков метаболитов через тонопласт?
Независимые доказательства в поддержку того, что мембрана тонопласта является «главным переключателем» циркадной регуляции САМ, были получены в результате компьютерного моделирования ритмов САМ, основанного на осмотических соображениях обмена малата и механизма натяжения/расслабления тонопласта (Lüttge, 2000, 2002 a ).Поскольку сдвиги в дневном/ночном содержании малата являются отличительной чертой пластичности САМ, метаболитный контроль фосфорилирования PEPC обеспечит эффективное средство точной настройки поглощения CO 2 в течение дневного/ночного цикла в зависимости от изменений условий окружающей среды. . Такая интеграция циркадных сигналов и сигналов окружающей среды может обеспечить основу для синхронизации и пластичности метаболизма, присущей САМ. Однако такая гипотеза поднимает дополнительные вопросы в отношении того, сколько метаболических компонентов цикла САМ напрямую связаны с циркадианным осциллятором, а сколько зависит от дневных и ночных потоков метаболитов через тонопласт?
Какие компоненты CAM регулируются часами?
Гены, контролируемые часами
Анализ примерно 40 генов в индуцируемом CAM-растении, Mesembryanthemum crystallinum , показал ритмические изменения в распространенности транскриптов более чем 30 выбранных генов с пиками экспрессии на различных фазах в течение всего цикла диэля (Boxall et al . , 2001, 2002). Более того, в растениях, экспрессирующих САМ, больше генов обнаруживают ритмичные изменения количества транскриптов по сравнению с растениями в состоянии С 3 (Boxall et al. ., 2001, 2002). Одним из способов синхронизации метаболических компонентов САМ с часами может быть фазирование транскрипции определенных генов в определенное время в дневном/ночном цикле, таким образом гарантируя, что соответствующие ферменты и переносчики будут наиболее распространены, когда это необходимо. Было обнаружено, что гены контролируются часами в CAM-исполнении M.crystalinum кодируют ферменты, участвующие в фотосинтезе, гликолизе, поглощении CO 2 в ночное время, декарбоксилировании, метаболизме сахарозы и крахмала (Dodd et al ., 2003), транспорте метаболитов хлоропластов и вакуолярной АТФазе (Boxall et al ., 2001, 2002). Таким образом, основные метаболические компоненты САМ, как показано на рис. 1а, по-видимому, подлежат элементу контроля, осуществляемому циркадными часами.
, 2001, 2002). Более того, в растениях, экспрессирующих САМ, больше генов обнаруживают ритмичные изменения количества транскриптов по сравнению с растениями в состоянии С 3 (Boxall et al. ., 2001, 2002). Одним из способов синхронизации метаболических компонентов САМ с часами может быть фазирование транскрипции определенных генов в определенное время в дневном/ночном цикле, таким образом гарантируя, что соответствующие ферменты и переносчики будут наиболее распространены, когда это необходимо. Было обнаружено, что гены контролируются часами в CAM-исполнении M.crystalinum кодируют ферменты, участвующие в фотосинтезе, гликолизе, поглощении CO 2 в ночное время, декарбоксилировании, метаболизме сахарозы и крахмала (Dodd et al ., 2003), транспорте метаболитов хлоропластов и вакуолярной АТФазе (Boxall et al ., 2001, 2002). Таким образом, основные метаболические компоненты САМ, как показано на рис. 1а, по-видимому, подлежат элементу контроля, осуществляемому циркадными часами. Текущий анализ генных чипов M. crystallinum , содержащих 8400 генов, несомненно, даст захватывающее представление о том, какие гены попадают под контроль часов, когда индуцируется САМ (J Cushman, личное сообщение).
Текущий анализ генных чипов M. crystallinum , содержащих 8400 генов, несомненно, даст захватывающее представление о том, какие гены попадают под контроль часов, когда индуцируется САМ (J Cushman, личное сообщение).
Хотя заманчиво предположить, что ключевым событием в эволюции САМ было соединение большего количества/различных генов с центральным осциллятором, требуется осторожность при интерпретации функционального значения ритмических изменений в обилии транскриптов. В растении C 3 , Arabidopsis thaliana эксперименты с микроматрицами показали, что по крайней мере 6% генов ритмично экспрессируются (Harmer et al. ., 2000; Schaffer et al. ., 2001). Однако более поздние подходы с использованием захвата энхансера in vivo показали, что 36% генома Arabidopsis потенциально находится под контролем транскрипции с помощью циркадных часов (Michael and McClung, 2003).Расхождение между этими сообщениями может быть связано с различиями в стабильности транскриптов между генами. Т.о., для того чтобы колебания транскрипции приводили к колебаниям количества мРНК, требуется, чтобы транскрипт был достаточно нестабилен, чтобы переворачиваться в течение циркадного цикла. Индукция САМ в M. crystallinum сопровождается повышением стабильности транскриптов САМ-специфичной изоформы PEPC, но снижением стабильности транскриптов, кодирующих малую субъединицу Rubisco (Cushman et al ., 1990; ДеРоше и Бонерт, 1993). Таким образом, регуляция сложного метаболического пути, такого как CAM, с помощью циркадных часов, вероятно, включает несколько уровней контроля, от транскрипции до посттрансляционной модификации белка, воздействуя на любое количество ферментов или транспортеров.
Т.о., для того чтобы колебания транскрипции приводили к колебаниям количества мРНК, требуется, чтобы транскрипт был достаточно нестабилен, чтобы переворачиваться в течение циркадного цикла. Индукция САМ в M. crystallinum сопровождается повышением стабильности транскриптов САМ-специфичной изоформы PEPC, но снижением стабильности транскриптов, кодирующих малую субъединицу Rubisco (Cushman et al ., 1990; ДеРоше и Бонерт, 1993). Таким образом, регуляция сложного метаболического пути, такого как CAM, с помощью циркадных часов, вероятно, включает несколько уровней контроля, от транскрипции до посттрансляционной модификации белка, воздействуя на любое количество ферментов или транспортеров.
Посттранскрипционный контроль с помощью часов
Чтобы определить физиологическую значимость циркадных колебаний количества транскриптов, необходимо также оценить уровни количества/активности соответствующих ферментов.Например, в то время как сообщается, что активность PPCK регулируется на уровне экспрессии генов (Hartwell et al.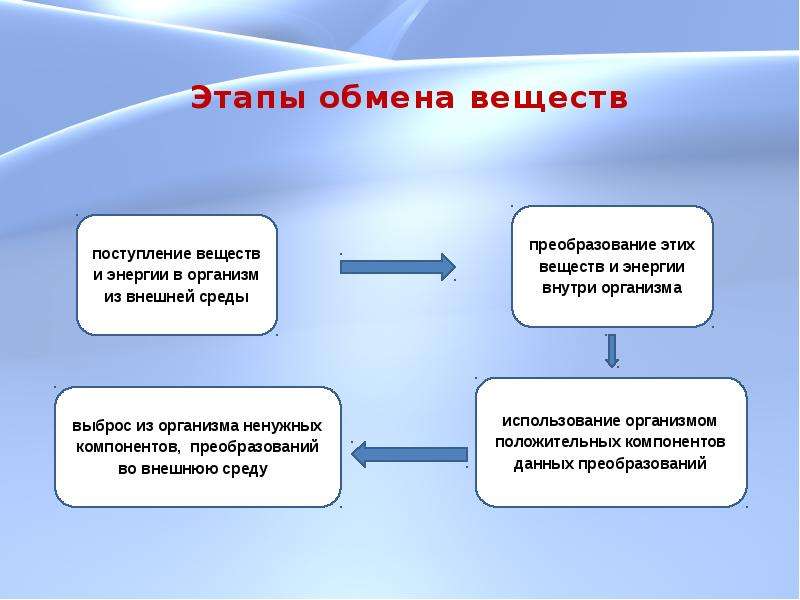 ., 1999; Taybi et al. ., 2000), обилие транскриптов САМ-специфичной изоформы PEPC также демонстрирует суточную активность. и циркадные колебания у M. crystallinum (с пиком к концу фотопериода), хотя белок PEPC и экстрагируемая активность остаются относительно постоянными (J Hartwell, личное сообщение; Boxall et al. ., 2001). Повышенное количество транскриптов PEPC в дневное время также было отмечено у видов Clusia , выполняющих САМ, но не у видов C 3 C.multiflora (T Taybi, неопубликованные наблюдения). У всех этих видов САМ одной из важных функций часов может быть контроль оборота белка PEPC и обновление уровня фермента каждую ночь. Дальнейший уровень циркадного контроля затем обеспечивается ритмическими сдвигами в экспрессии и активности PPCK, которые, в свою очередь, ночью активируют белок PEPC. По мере того, как появляется больше данных об экспрессии ритмичных генов в результате сравнения микрочипов C 3 и CAM-performing M.
., 1999; Taybi et al. ., 2000), обилие транскриптов САМ-специфичной изоформы PEPC также демонстрирует суточную активность. и циркадные колебания у M. crystallinum (с пиком к концу фотопериода), хотя белок PEPC и экстрагируемая активность остаются относительно постоянными (J Hartwell, личное сообщение; Boxall et al. ., 2001). Повышенное количество транскриптов PEPC в дневное время также было отмечено у видов Clusia , выполняющих САМ, но не у видов C 3 C.multiflora (T Taybi, неопубликованные наблюдения). У всех этих видов САМ одной из важных функций часов может быть контроль оборота белка PEPC и обновление уровня фермента каждую ночь. Дальнейший уровень циркадного контроля затем обеспечивается ритмическими сдвигами в экспрессии и активности PPCK, которые, в свою очередь, ночью активируют белок PEPC. По мере того, как появляется больше данных об экспрессии ритмичных генов в результате сравнения микрочипов C 3 и CAM-performing M. crystallinum важно связать такие исследования с обычными биохимическими измерениями количества и активности соответствующих ферментов и переносчиков. Такая информация поможет определить, требует ли экспрессия САМ не только изменения в наборе генов, связанных с часами, но также необходимо изменение уровня контроля (транскрипционного или посттранскрипционного), осуществляемого ферментом. Часы.
crystallinum важно связать такие исследования с обычными биохимическими измерениями количества и активности соответствующих ферментов и переносчиков. Такая информация поможет определить, требует ли экспрессия САМ не только изменения в наборе генов, связанных с часами, но также необходимо изменение уровня контроля (транскрипционного или посттранскрипционного), осуществляемого ферментом. Часы.
Физиологические процессы, контролируемые часами
Дополнительный подход к решению вопроса о том, какие компоненты пути САМ регулируются часами, заключался в использовании явных циркадных ритмов газообмена листа, обнаруженных у некоторых видов, для изучения более высоких уровней метаболической организации, которые подпадают под действие часов. контроль циркадного осциллятора.Недавний анализ стехиометрии поглощения CO 2 и накопления малата в M. crystallinum и K. daigremontiana во время свободных ритмов газообмена показывает, что карбоксилирование C 3 вносит основной вклад в генерацию ритмических Поглощение CO 2 при постоянном освещении (Dodd et al. ., 2002, 2003; Wyka and Lüttge, 2003). При продолжительных периодах непрерывного освещения затухание активности PEPC происходит быстрее, чем затухание явного ритма в чистом поглощении CO 2 .Это указывает на то, что в то время как эндогенный контроль фосфорилирования PEPC не является устойчивым при постоянном освещении, другой и, возможно, отдельный C 3 -родственный осциллятор поддерживает ритмы в чистом поглощении CO 2 (Wyka and Lüttge, 2003). В настоящее время неясно, может ли эта ритмическая активность при карбоксилировании C 3 быть непосредственно связана с циркадным контролем активации/активности Rubisco или с устьичной проводимостью. Оба эти аспекта метаболизма, как известно, подвергаются по крайней мере некоторому элементу циркадного контроля у C 3 spp (Liu et al ., 1996; Уэбб, 2003). Представление о том, что общий контроль явных ритмов при поглощении CO 2 растениями САМ опосредован колебаниями устьичного отверстия, обсуждалось в другом месте (Lüttge, 2002 a ; Wyka and Lüttge, 2003).
., 2002, 2003; Wyka and Lüttge, 2003). При продолжительных периодах непрерывного освещения затухание активности PEPC происходит быстрее, чем затухание явного ритма в чистом поглощении CO 2 .Это указывает на то, что в то время как эндогенный контроль фосфорилирования PEPC не является устойчивым при постоянном освещении, другой и, возможно, отдельный C 3 -родственный осциллятор поддерживает ритмы в чистом поглощении CO 2 (Wyka and Lüttge, 2003). В настоящее время неясно, может ли эта ритмическая активность при карбоксилировании C 3 быть непосредственно связана с циркадным контролем активации/активности Rubisco или с устьичной проводимостью. Оба эти аспекта метаболизма, как известно, подвергаются по крайней мере некоторому элементу циркадного контроля у C 3 spp (Liu et al ., 1996; Уэбб, 2003). Представление о том, что общий контроль явных ритмов при поглощении CO 2 растениями САМ опосредован колебаниями устьичного отверстия, обсуждалось в другом месте (Lüttge, 2002 a ; Wyka and Lüttge, 2003).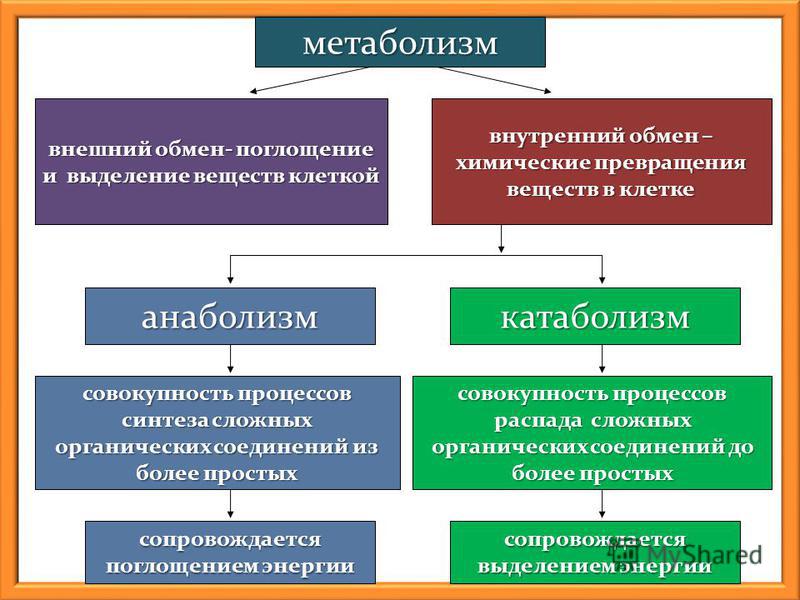 В рамках этой структуры циркадные осцилляторы, которые непосредственно контролируют PEPC и Rubisco, могут представлять собой другие компоненты сети, которые стабилизируют и координируют поглощение CO 2 в ответ на петли обратной связи метаболитов и сигналы окружающей среды (Dodd et al ., 2003). Такие соображения согласуются с идеей, что устойчивые ритмы газообмена листа не обязательно контролируются одним центральным часовым механизмом, который мог бы действовать на уровне транскрипции генов. Скорее, свободные колебания в чистом поглощении CO 2 , по-видимому, управляются сложными регуляторными сетями, интегрированными во времени и пространстве (Lüttge, 2000, 2002 a ; Rascher et al , 2001).
В рамках этой структуры циркадные осцилляторы, которые непосредственно контролируют PEPC и Rubisco, могут представлять собой другие компоненты сети, которые стабилизируют и координируют поглощение CO 2 в ответ на петли обратной связи метаболитов и сигналы окружающей среды (Dodd et al ., 2003). Такие соображения согласуются с идеей, что устойчивые ритмы газообмена листа не обязательно контролируются одним центральным часовым механизмом, который мог бы действовать на уровне транскрипции генов. Скорее, свободные колебания в чистом поглощении CO 2 , по-видимому, управляются сложными регуляторными сетями, интегрированными во времени и пространстве (Lüttge, 2000, 2002 a ; Rascher et al , 2001).
Метаболитный контроль САМ
Для ряда генов характерны ритмические изменения количества транскриптов только после индукции САМ в М.crystallinum (Boxall et al. ., 2001, 2002), и возможно, что многие из этих ритмов могут возникать как последующие последствия цикла метаболитов, связанного с CAM. Вопрос о контроле метаболитов также актуален для понимания того, как отдельные осцилляторы, как было предложено выше, могут быть интегрированы в пространстве и времени, чтобы координировать обмен diel CO 2 в различных условиях окружающей среды. Существенный взаимный цикл органических кислот и углеводов происходит в течение 24-часового цикла САМ (рис.1b), и известно, что обе категории метаболитов влияют на экспрессию разнообразных генов (Smeekens, 2000; Stitt et al. ., 2002).
Вопрос о контроле метаболитов также актуален для понимания того, как отдельные осцилляторы, как было предложено выше, могут быть интегрированы в пространстве и времени, чтобы координировать обмен diel CO 2 в различных условиях окружающей среды. Существенный взаимный цикл органических кислот и углеводов происходит в течение 24-часового цикла САМ (рис.1b), и известно, что обе категории метаболитов влияют на экспрессию разнообразных генов (Smeekens, 2000; Stitt et al. ., 2002).
Датчики органических кислот и сигнализация
Недавние доказательства в поддержку индуцированного метаболитами цикла существующих генов C 3 получены путем сравнения экспрессии киназы PEPC (PPCK) в диапазоне видов Clusia , которые демонстрируют заметные различия в способности к САМ. Способность к САМ в Clusia , по-видимому, определяется количеством белка PEPC (Borland et al ., 1998), который, в свою очередь, регулируется на уровне обилия транскриптов PEPC (Taybi et al ., 2004). Однако ген, кодирующий PPCK, белок, ответственный за активацию PEPC, экспрессируется на сравнимых уровнях в листьях как C 3 , так и САМ-производящих видов Clusia , и дневные/ночные изменения в экспрессии PPCK, по-видимому, последствие круговорота органических кислот и растворимых сахаров (рис. 5; Taybi et al. ., 2004). Более того, diel-изменения содержания транскриптов PPCK, по-видимому, контролируются за счет подавления активности гена в течение дня, а не повышения активности в ночное время (рис.5). Эти результаты контрастируют с результатами, полученными с M. crystallinum , в которых транскрипты PPCK присутствуют в небольшом количестве в листьях C 3 и где индукция САМ сопровождается повышающей регуляцией количества транскриптов PPCK в ночное время (Taybi et al. al ., 2000). Дневной выброс малата (или какого-либо другого метаболита) из вакуоли может быть первичным сигналом для подавления экспрессии PPCK во время фотопериода у видов Clusia после индукции САМ.Последующее декарбоксилирование органических кислот будет ключевым фактором, влияющим на увеличение экспрессии PPCK на следующую ночь. Интересно отметить, что видов Clusia могут накапливать высокие концентрации как яблочной, так и лимонной кислот, а фоновые концентрации обеих органических кислот могут быть относительно высокими у растений, осуществляющих фотосинтез C 3 (Borland et al ., 1996). Опять же, это контрастирует с ситуацией в M. crystallinum , где содержание как малата, так и цитрата низкое в режиме C 3 .Таким образом, у Clusia оказывается, что мобилизация органических кислот в дневное время является ключевым фактором, регулирующим как экспрессию САМ, так и PEPC-киназы (Borland et al. ., 1996; Roberts et al. ., 1997). ). Таким образом, заметная фенотипическая пластичность растений, таких как C. minor , для быстрого и обратимого переключения между фотосинтезом C 3 и САМ может быть связана с транспортными и/или ферментативными процессами, которые регулируют распределение метаболитов, особенно органических кислот, между вакуолями. и цитозоль.
Транспортные этапы, которые опосредуют приток и отток яблочной и лимонной кислот через мембрану тонопласта, будут центральным компонентом в восприятии органических кислот и передаче сигналов у САМ-растений. Ночное накопление малата включает стехиометрический транспорт 2H + на малат с транспортом H + , управляемым тонопластной H + АТФазой и, дополнительно, H + PP i азой (Smith et ). ., 1996). Циркадные колебания количества транскриптов субъединицы V-АТФазы C были описаны у M.crystalinum (Rockel et al. ., 1997), но неизвестно, подвергается ли активность V-АТФазы циркадной регуляции. Хотя фактические переносчики органических кислот еще не идентифицированы на молекулярном уровне, кажется вероятным, что в тонопласте САМ-растений существуют различные системы притока и оттока (Smith et al. ., 1996). Приток малата через тонопласт, по-видимому, опосредуется потенциал-зависимым анионселективным каналом (Hafke et al ., 2003). Этот канал реагирует на рН цитоплазмы таким образом, который согласуется с ночным поглощением аниона малата и помогает свести к минимуму бесполезный возврат малата обратно в вакуоль во время дневной фазы раскисления. Транспортер карбоксилата тонопласта, который, по-видимому, способен транспортировать как малат, так и цитрат, недавно был клонирован из A. thaliana (Emmerlich et al. ., 2003). Более того, количество транскриптов этого транспортера может модулироваться в ответ на концентрацию малата (Emmerlich et al ., 2003). САМ-ортолог карбоксилатного переносчика Arabidopsis потенциально может действовать как ключевой сенсор для передачи сигналов, опосредованной органическими кислотами, в течение цикла diel CAM, регулируя поток через тонопласт в ответ на концентрацию малата.
Определение содержания углеводов и сигнализация
Доступность углеводов является основным фактором, ограничивающим поглощение темного CO 2 растениями САМ (Borland and Dodd, 2002). В листьях M. crystallinum , которые были истощены по углеводам на 50% при воздействии CO 2 на открытом воздухе в течение 24 часов, последующее суммарное поглощение CO 2 в темноте в окружающем воздухе было снижено на 50% по сравнению с контролем, несмотря на заметное увеличение экспрессии PEPC-киназы (Dodd et al ., 2003). Доказательства обнаружения дефицита углеводов (и, возможно, органических кислот) у растений, сниженного при ночном поглощении CO 2 , также подтверждаются сдвигами в последующем дневном чистом поглощении CO 2 и распределении углеводов, которые могут компенсировать предыдущий ночной дефицит в приросте углерода (Roberts et al. ., 1997; Borland and Griffiths, 1997; Dodd et al. ., 2002). Таким образом, углеводный статус, по-видимому, является ключевым элементом в синхронизации метаболических потоков в течение цикла САМ в соответствии с изменениями условий окружающей среды.
Углеводный статус определяется взаимодействием циркадного контроля и контроля метаболитов, которые регулируют оборот и субклеточное распределение в течение дневного цикла. Наиболее изученные модели САМ (например, M. crystallinum , K. daigremontiana ) продуцируют преимущественно крахмал в хлоропластах в качестве временного запаса углерода для поддержки ночного карбоксилирования и синтеза малата. У растений C 3 ранее предполагалось, что дневное накопление крахмала находится под циркадным контролем (Li et al ., 1992; Гейгер и др. ., 1995). Для CAM-выполняющих M. crystallinum был предложен циркадный контроль основного этапа биосинтеза крахмала, катализируемого АДФ-глюкоза-пирофосфорилазой (Boxall et al. ., 2001, 2002; J Hartwell, личное сообщение). На ключевую роль переносчиков хлоропластов в регуляции углеводного обмена в диэльном цикле указывает тот факт, что хлоропласты САМ-производящих листьев M. crystallinum уникальны тем, что содержат три класса транслокаторов фосфата (т.е. PEP, транслокаторы триозы-P и глюкозы-6-P), в дополнение к хлоропластному переносчику глюкозы (Häusler et al. ., 2000). Более того, динамическая суточная регуляция количества транскриптов и активности этих хлоропластных транслокаторов очевидна, когда CAM индуцируется (Häusler et al. ., 2000; Boxall et al. ., 2001, 2002). ФЕП, триозо-P, глюкоза-6-P и хлоропластные переносчики глюкозы могут играть ключевую роль в восприятии уровней углеводов и связанных с ними метаболитов и, следовательно, модулировать транспортную активность таким образом, чтобы поддерживать возвратно-поступательные метаболические потоки во время цикла CAM (Häusler). и др. ., 2000).
Ритмические изменения количества транскриптов нескольких генов, кодирующих ферменты, участвующие в расщеплении крахмала, также были зарегистрированы у M. crystallinum , с пиком количества транскриптов до конца (субъективного) фотопериода (Boxall et al ., 2001, 2002; Dodd и др. ., 2003). Циркадный контроль деградации крахмала у M. crystalinum может служить средством прогнозирования потребности в субстрате для ночного карбоксилирования, обеспечивая сохранение адекватных запасов углеводов в течение фотопериода.Однако вполне вероятно также, что существуют механизмы для «ощущения» доступности углеводов в течение ночи, поскольку переход от C 3 к САМ приводит к устойчивому «измерению» запасов углеводов, которые поддерживают доступность субстрата в течение всего темного периода. Додд и др. ., 2003). В настоящее время неясно, определяется ли это кажущееся ощущение уровня углеводов притяжением 3-C субстрата от PEPC или прямой модуляцией скорости деградации углеводов.Однако недавние результаты, полученные с CAM-дефицитным мутантом M. crystallinum , показывают, что активность ферментов, разлагающих крахмал, может модулироваться в соответствии с доступностью субстрата. В мутантной линии, выделенной группой Джона Кушмана, неспособность накапливать малат в течение ночи, по-видимому, коррелирует с низким содержанием крахмала (Branco et al ., 2003). В этих САМ и растениях с дефицитом крахмала активность и колебание суточной активности хлоропластной фосфорилазы крахмала также снижены по сравнению с растениями дикого типа (A Borland и J Cushman, неопубликованные наблюдения).Сходным образом мутант проявляет более низкую активность и уменьшенные колебания диэля ряда глюкангидролаз, указывая на то, что активность разложения крахмала можно регулировать в соответствии с доступностью субстрата (A Borland and J Cushman, неопубликованные наблюдения). Таким образом, деградация крахмала в M. crystallinum , по-видимому, подлежит циркадианному контролю на уровне экспрессии генов и контролю метаболитов на уровне ферментативной активности.
Во многих САМ-растениях, включая сельскохозяйственные виды Ananas comosus (ананас), Aloe vera и виды Agave , вакуолярные растворимые сахара являются преобладающей формой углеводов, накапливаемых в течение дня для поддержания цикла САМ (Winter). и Smith, 1996 b ).Дневные/ночные потоки сахаров через мембрану тонопласта у ананаса могут быть значительными (до 20% сухой биомассы листа) и являются основным фактором, определяющим экспрессию САМ. Предполагаемый вакуолярный переносчик сахарозы был описан в везикулах тонопласта A. comosus , который проявляет кинетические характеристики унипортера сахарозы и, по-видимому, способен облегчать значительные потоки сахаров в вакуоль (McRae et al. ., 2002). Исследования плазматической мембраны растений C 3 показали, что экспрессия транскриптов транспорта сахара может сильно реагировать на метаболические сигналы и сигналы окружающей среды, а продукты генов могут участвовать в таких функциях, как распознавание сахара и передача сигналов в дополнение к транспорту. Уильямс и др. ., 2000). Таким образом, поток сахаров через тонопласт может представлять собой стратегическую контрольную точку для интеграции циркадного и метаболитного контроля диэлитного цикла у видов САМ, которые накапливают растворимые сахара.
CO
2 датчики и сигнализацияКрупные сильно вакуолизированные и плотно упакованные клетки листьев многих САМ-растений существенно ограничивают диффузию CO 2 как в лист, так и из него. Это потенциально может привести к 450-кратному снижению p CO 2 в течение нескольких часов с момента максимального декарбоксилирования в фазе III до момента, когда во время фазы IV происходит прямое поглощение атмосферного CO 2 (Максвелл и др. ., 1997). Таким образом, в ходе диэл-САМ-цикла сдвиги режимов карбоксилирования С 3 и С 4 приводят к колебаниям внутренней концентрации СО 2 , которые могут составлять от 0,011% до 5%. Считается, что увеличение p CO 2 является основным внутренним фактором контроля закрытия устьиц в фазе III (Bohn et al. ., 2001). Ощущение p CO 2 во время перехода от фазы III к фазе IV также проявляется на метаболическом уровне увеличением состояния активации Rubisco, которое, как предполагалось, поддерживает выработку CO 2 во время фазы IV (Максвелл и др. ., 1999: Максвелл, 2002; Griffiths и др. ., 2002). Lüttge (2002 b ) также рассмотрел роль p CO 2 в качестве сигнала для координации скорости потребления CO 2 Rubisco и ремобилизации яблочной кислоты из вакуоли во время фазы III CAM. Таким образом, p CO 2 восприятие и передача сигналов могут представлять собой еще один неотъемлемый компонент механизмов, которые синхронизируют потоки углерода в течение суточных циклов у САМ-растений.
Интеграция экологических и метаболических сигналов с циркадными часами
Механизм, лежащий в основе циркадных часов во всех организмах, основан на замедленных отрицательных петлях, регулирующих гены, вовлеченные в ядро осциллятора (Dunlap, 1999; Harmer et al ., 2001). В этих петлях задействованы негативные регуляторы, которые в ответ подавляют собственное выражение, блокируя положительно действующие элементы (Harmer et al. , 2001; Young and Kay, 2001). Различные уровни контроля, включая посттранскрипционную регуляцию и связанные с ней взаимосвязанные петли обратной связи, обеспечивают механизмы, обеспечивающие стабильность и надежность часов в изменяющихся условиях, а также для сброса фазы часов сигналами окружающей среды, особенно светом (Shearman et al ., 2000). У Arabidopsis, циркадные часы включают набор генов, некоторые из которых являются хорошо охарактеризованными факторами транскрипции (например, регулятором псевдоответа TOC1: Timing of CAB Expression ) и факторами семейства Myb ( CCA1 ). : Циркадные часы, связанные и LHY1 : Поздний удлиненный гипокотиль ). Эти факторы регулируют экспрессию друг друга в петле отрицательной обратной связи, что приводит к примерно 24-часовому периоду.Отсутствие экспрессии TOC1 вызывает аритмию, в то время как генов CCA1 и LHY1 , которые имеют перекрывающиеся функции, участвуют в контроле периода часов. Мутанты, лишенные экспрессии CCA1 и LHY1 , неспособны поддерживать устойчивые колебания ни при постоянном свете, ни в темноте (Alabadi et al. ., 2002). Циркадная экспрессия этих часовых генов контролирует нижестоящую экспрессию множества генов, участвующих во многих жизненно важных процессах, включая фотосинтетический метаболизм, движения листьев и цветение.В настоящее время ведутся работы по идентификации гомологичных генов TOC1 , CCA1 и LHY1 в индуцибельном САМ-растении M. crystallinum (J Hartwell, личное сообщение). Полученные результаты помогут выяснить, как включается часовой контроль метаболических процессов при индукции САМ. Более того, исследования таких часовых генов могут выявить дополнительную роль часов в регуляции долговременной индукции САМ. У таких видов, как K. blossfeldiana и M.crystallinum, , долгосрочный онтогенетический контроль экспрессии САМ может быть связан с изменением длины фотопериода (Brulfert et al , 1975; Edwards et al , 1996). В то время как этот эволюционный переход к САМ может быть ускорен различными факторами окружающей среды (например, засухой, соленостью, ярким светом), ощущая изменение длины фотопериода, циркадные часы могут устанавливать метаболические и физиологические приспособления в ожидании изменения. в условиях окружающей среды (Taybi и др. ., 1995, 2002; Тайби и Кушман, 1999). Как у K. blossfeldiana , так и у M. crystallinum фотопериодическая индукция САМ всегда сопровождается цветением и образованием семян. В будущем будет интересно установить, контролируются ли долгосрочные (сезонные) реакции, такие как индукция САМ и цветение, и краткосрочные (диэль) реакции с помощью одних и тех же часов или наборов часовых генов.
Понятно, что внешние (например, свет, температура) и внутренние факторы (метаболиты, гормоны) могут воздействовать как на амплитуду, так и на период основных ритмов, генерируемых часами у САМ-растений.Такая модуляция необходима для оптимизации использования ресурсов в ежедневной шкале времени и для обеспечения координации развития САМ с сезонными изменениями. В настоящее время неясно, действуют ли эти внешние и внутренние факторы непосредственно на гены часов или действуют ниже по течению, захватывая метаболический выход часов. Тем не менее, тонопластовая мембрана остается сильным кандидатом на интеграцию метаболического и циркадианного контроля пути diel CAM. Lüttge (2000, 2002 a ) предположил, что биофизическое напряжение/расслабление тонопласта представляет собой первичный осциллятор или кардиостимулятор для САМ.Другая возможная модель гипотезы осциллятора тонопласта включает синтез пептида, который включается в мембрану, где он имеет отрицательную обратную связь с собственным синтезом (Lüttge et al. ., 2002 a ). Прямой циркадный контроль пептида, связанного с транспортом малата, пептида, который, в свою очередь, регулирует собственную экспрессию, может быть одним из возможных механизмов, с помощью которого тонопласт может интегрировать циркадные и метаболические сигналы для контроля цикла САМ.
Эволюционные последствия и выводы
Два основных уровня контроля контролируют метаболические компоненты САМ. Контроль с помощью циркадных часов устанавливает суточные фазы САМ и обеспечивает соответствующую синхронизацию метаболических и транспортных процессов путем фазирования транскрипции определенных генов в определенное время в дневном/ночном цикле. На этот циркадный контроль накладывается контроль метаболитов, действующий как на транскрипционном, так и на посттранскрипционном уровнях, что способствует фотосинтетической пластичности, привязывая выходные данные часов к колебаниям окружающей среды.Возвратно-поступательные пулы органических кислот и углеводов играют центральную роль в метаболическом контроле САМ. Локализованные в мембране переносчики и сенсоры органических кислот, сахаров и промежуточных продуктов метаболизма, таким образом, занимают стратегические контрольные точки для интеграции циркадных и метаболических сигналов, которые синхронизируют и модулируют фазы диэля в ответ на окружающую среду. Тонопласт, в частности, играет ключевую роль в управлении метаболизмом в течение цикла день/ночь, контролируя поглощение и высвобождение органических кислот в цитозоль.В самом деле, у некоторых видов САМ (например, Clusia ) суточные колебания количества транскриптов генов, кодирующих ключевые ферменты САМ (например, киназу PEPC), по-видимому, являются последующим следствием круговорота метаболитов в тонопласте (Fig. 5).
Интегрированный механистический обзор цикла САМ, описанный выше, поднимает вопрос о том, как особенности метаболического контроля САМ развились из фотосинтеза C 3 . Если САМ развился в результате серии инкрементных модификаций существующих признаков, важно учитывать атрибуты САМ-предшественников, которые представляли предрасположенность к развитию как циркадных, так и компонентов метаболического контроля пути.Считается, что начальным этапом эволюции САМ является ночное удаление CO 2 из дыхательных путей с помощью PEPC, чему, вероятно, способствовала сочная и крупная вакуолизированная анатомия листьев, типичная для видов САМ (Griffiths, 1989; Sage, 2002). ). Суккулентность могла быть ранее существовавшей чертой, которая улучшала гидравлический транспорт и емкость в местах обитания с высокой потребностью в транспирации (Sage, 2002). Плотно упакованные клетки суккулентных листьев минимизировали бы утечку CO 2 из листьев, в то время как увеличенная вакуоль была необходима для хранения фиксированного CO 2 в виде органических кислот в течение ночи.Поскольку вакуоль обычно занимает c . 97% объема клетки и цитоплазмы c . 1% в листьях САМ-растений цитоплазма легко могла быть залита вакуолярным содержимым, нарушая тем самым метаболический гомеостаз. Т.о., транспорт через тонопласт д. строго регулироваться, что указывает на то, что ранним событием с точки зрения метаболического контроля САМ может быть увеличение регуляторной способности транспортеров тонопласта. Это могло быть достигнуто за счет диверсификации состава транспортеров тонопластов (например,г. отдельные транспортеры для притока и оттока органических кислот) вместе с изменением регуляторных свойств транспортеров. На молекулярном уровне такие качественные и количественные изменения в переносчиках тонопластов могли развиться посредством дупликации генов с последующей специфической регуляцией одного или нескольких изогенов, что обычно предполагается для нескольких основных метаболических компонентов САМ (Cushman and Bohnert, 1999). Важность вакуоли в поддержании клеточного гомеостаза, возможно, также сделала тонопласт ключевым (если не основным) компонентом циркадианного осциллятора, который контролирует CAM.
При рассмотрении эволюционного происхождения циркадного контроля САМ также уместно рассмотреть, обладали ли уже предшественники САМ часами с всеобъемлющим контролем над метаболизмом. В то время как циркадианная экспрессия гена в прокариотическом организме Synechococcus , по-видимому, является универсальным феноменом (Liu et al ., 1995), у высших растений A. thaliana ~36% генома потенциально находится под контролем транскрипции с помощью циркадные часы (Michael and McClung, 2003).Тем не менее, из генов Arabidopsis , которые, как известно, демонстрируют циркадные колебания численности транскриптов, большая часть (почти 70%) также напрямую реагирует на стресс окружающей среды (т.е. низкую температуру, соль и засуху; Kreps et al ., 2002). Это наблюдение подтолкнуло к предположению, что ритмичная экспрессия таких генов, связанных со стрессом, в ожидании предсказуемых изменений окружающей среды может подготовить растение к противостоянию стрессу или наилучшим образом использовать потенциально ограничивающий ресурс (Eriksson and Millar, 2003).Поскольку обычно считается, что САМ возникла в экстремальных условиях (Raven and Spicer, 1996), можно предположить, что предшественники САМ уже обладали часами с всеобъемлющим контролем над метаболизмом как средством поддержания гомеостаза в потенциально ограничивающих условиях. Последующая эволюция пути могла происходить либо за счет соединения большего количества/других компонентов метаболического выброса с центральным генератором и/или за счет изменений в путях (и, возможно, механизмах), которые связывают центральный(е) осциллятор(ы) с метаболическим выбросом.Будущие подходы к пониманию фотосинтеза у САМ-растений потребуют применения постгеномных технологий, включая транскриптомику, протеомику и метаболомику, для идентификации генов и белков, которые контролируются часами, и для определения роли метаболитов (например, органические кислоты и сахара) в синхронизации потоков углерода в дизеле. Технология генных чипов в настоящее время применяется для анализа относительного вклада часов и контроля метаболитов у мутантов дикого типа и CAM-дефицитных мутантов M.crystallinum (Дж. Кушман, личное сообщение). Дальнейший прогресс требует разработки подходящей и поддающейся трансформации системы для проверки гипотез о циркадных и метаболических сигналах, которые контролируют как экспрессию, так и синхронизацию метаболических компонентов, составляющих САМ.
Благодарности
Мы благодарны многим коллегам, которые внесли свой вклад в идеи, высказанные выше, особенно Джону Кушману, Джеймсу Хартвеллу, Хью Ниммо и Эндрю Смиту.Наше исследование поддерживается Советом по исследованиям окружающей среды, Великобритания.
Рис. 1. Основные метаболические компоненты цикла САМ. Схематический план (а), иллюстрирующий дневное (светлый фон) и ночное (темный фон) разделение основных метаболических и транспортных процессов для углерода. Распределение углеводов между ростом и хранением (в виде растворимых сахаров в вакуолях или в виде крахмала в хлоропластах) является важным пунктом контроля диэлицикла.Конечными результатами такого разделения метаболизма (b) являются возвратно-поступательные уровни органических кислот (в основном малата) и углеводов, которые могут составлять до 20% сухой массы листа. Сплошная полоса на оси x указывает на период темноты, а данные о метаболитах были получены из Clusia fluminensis (A Borland, неопубликованные результаты).
Рис. 1. Основные метаболические компоненты цикла САМ. Схематический план (а), иллюстрирующий дневное (светлый фон) и ночное (темный фон) разделение основных метаболических и транспортных процессов для углерода.Распределение углеводов между ростом и хранением (в виде растворимых сахаров в вакуолях или в виде крахмала в хлоропластах) является важным пунктом контроля диэлицикла. Конечными результатами такого разделения метаболизма (b) являются возвратно-поступательные уровни органических кислот (в основном малата) и углеводов, которые могут составлять до 20% сухой массы листа. Сплошная полоса на оси x указывает на период темноты, а данные о метаболитах были получены из Clusia fluminensis (A Borland, неопубликованные результаты).
Рис. 2. Регуляция фосфо енол пируваткарбоксилазы в растениях САМ. Ген Ppc может быстро индуцироваться факторами окружающей среды (водным стрессом, солевым стрессом и т. д.) или в качестве долговременной реакции на изменение фотопериода посредством контроля с помощью циркадианного осциллятора. Транскрипты Ppc также колеблются на циркадной основе, но это изменение не приводит к изменению количества белка PEPC в течение 24-часового цикла.Diel-регуляция PEPC контролируется ночным фосфорилированием, катализируемым специальной киназой (PEPC-киназа). Ген Ppck может индуцироваться солевым стрессом у некоторых видов ( Mesembryanthemum crystallinum, sorghum) и контролируется циркадианным осциллятором и/или метаболитами (органическими кислотами и сахарами). Транскрипты Ppck накапливаются в основном ночью в растениях САМ, что приводит к ночному накоплению PEPC-киназы и фосфорилированию PEPC. Фосфорилированный PEPC более активен, более чувствителен к активации глюкозо-6-фосфатом и триозофосфатом и менее чувствителен к ингибированию малатом.
Рис. 2. Регуляция фосфо енол пируваткарбоксилазы в растениях САМ. Ген Ppc может быстро индуцироваться факторами окружающей среды (водным стрессом, солевым стрессом и т. д.) или в качестве долговременной реакции на изменение фотопериода посредством контроля с помощью циркадианного осциллятора. Транскрипты Ppc также колеблются на циркадной основе, но это изменение не приводит к изменению количества белка PEPC в течение 24-часового цикла. Diel-регуляция PEPC контролируется ночным фосфорилированием, катализируемым специальной киназой (PEPC-киназа).Ген Ppck может индуцироваться солевым стрессом у некоторых видов ( Mesembryanthemum crystallinum, sorghum) и контролируется циркадианным осциллятором и/или метаболитами (органическими кислотами и сахарами). Транскрипты Ppck накапливаются в основном ночью в растениях САМ, что приводит к ночному накоплению PEPC-киназы и фосфорилированию PEPC. Фосфорилированный PEPC более активен, более чувствителен к активации глюкозо-6-фосфатом и триозофосфатом и менее чувствителен к ингибированию малатом.
Рис. 3. Регуляция Diel и кажущийся статус фосфорилирования PEPC у контрастирующих видов САМ. Чувствительность PEPC к малату (а) и содержание малата в листьях (b) в листьях Kalanchoë daigremontiana (закрашенные кружки), Clusia minor (незаштрихованные кружки) и Tillandsia usneoides (незаштрихованные ромбы), выращенных в идентичных контролируемых условиях с сплошная полоса на оси x указывает период темноты. Графики были составлены на основе данных, представленных Borland and Griffiths (1997) и Haslam et al .(2002).
Рис. 3. Регуляция Diel и кажущийся статус фосфорилирования PEPC у контрастирующих видов САМ. Чувствительность PEPC к малату (а) и содержание малата в листьях (b) в листьях Kalanchoë daigremontiana (закрашенные кружки), Clusia minor (незаштрихованные кружки) и Tillandsia usneoides (незаштрихованные ромбы), выращенных в идентичных контролируемых условиях с сплошная полоса на оси x указывает период темноты. Графики были составлены на основе данных, представленных Borland and Griffiths (1997) и Haslam et al .(2002).
Рис. 4. Метаболитный контроль фосфорилирования PEPC и чистого поглощения CO 2 . Схемы чистого поглощения CO 2 (a) листьями Kalanchoë daigremontiana при атмосферном воздухе (контроль) или в атмосфере N 2 в течение первых 6 ч (половина N 2 ) или всего 12 ч (полный N 2 ) темного периода (обозначен сплошной полосой на оси x ). Закрытие в N 2 предотвращает накопление малата, а последующее высвобождение из N 2 приводит к удвоению чистого поглощения CO 2 через PEPC по сравнению с контролем.Увеличение чистого поглощения CO 2 в листьях, обработанных N 2 , сопровождается увеличением активности (b) и экспрессии (данные не показаны) киназы PEPC, белка, ответственного за фосфорилирование PEPC. Данные были повторно взяты из Borland и др. . (1999).
Рис. 4. Метаболитный контроль фосфорилирования PEPC и чистого поглощения CO 2 . Схемы чистого поглощения CO 2 (a) листьями Kalanchoë daigremontiana при атмосферном воздухе (контроль) или в атмосфере N 2 в течение первых 6 ч (половина N 2 ) или всего 12 ч (полный N 2 ) темного периода (обозначен сплошной полосой на оси x ).Закрытие в N 2 предотвращает накопление малата, а последующее высвобождение из N 2 приводит к удвоению чистого поглощения CO 2 через PEPC по сравнению с контролем. Увеличение чистого поглощения CO 2 в листьях, обработанных N 2 , сопровождается увеличением активности (b) и экспрессии (данные не показаны) киназы PEPC, белка, ответственного за фосфорилирование PEPC. Данные были повторно взяты из Borland и др. . (1999).
Рис.5. Diel экспрессия киназы PEPC у видов Clusia . Дневные (3 ч и 6 ч) и ночные (15 ч и 18 ч) равновесные уровни транскриптов Ppck1 и Ubq1 (контроль) в листьях Clusia multiflora (C 3 видов), C .aripoensis (C 3 ), C. minor (САМ) и C. rosea (САМ), выращенных в идентичных контролируемых условиях.
Рис. 5. Diel экспрессия киназы PEPC у видов Clusia .Дневные (3 ч и 6 ч) и ночные (15 ч и 18 ч) равновесные уровни транскриптов Ppck1 и Ubq1 (контроль) в листьях Clusia multiflora (C 3 видов), C .aripoensis (C 3 ), C. minor (САМ) и C. rosea (САМ), выращенных в идентичных контролируемых условиях.
Каталожные номера
Алабади D , Яновский М.Дж., Мас П., Хармер С.Л., Кей С.А.
2002
. Критическая роль CCA1 и LHY1 в поддержании циркадной ритмичности у Arabidopsis .Современная биология
12
,757
–761.Варфоломей ДП , Кадзимин С.Б.
1977
. Ананас. В: Альвим П. де Т., Козловский Т.Т., ред. Экофизиология тропических культур . Нью-Йорк: Academic Press,113
–156.Bohn A , Geist A, Rascher U, Lüttge U.
2001
. Реакция циркадного ритма метаболизма крассуловой кислоты на различные внешние световые ритмы у Kalanchoë daigremontiana .Растения, клетки и окружающая среда
24
,811
–820.Borland AM , Додд А.Н.
2002
. Разделение углеводов в САМ-установках: урегулирование потенциальных конфликтов интересов.Функциональная биология растений
29
,707
–716.Borland AM , Гриффитс Х.
1997
. Сравнительное исследование регуляции процессов карбоксилирования C 3 и C 4 в конститутивном CAM растении Kalanchoë daigremontiana и C 3 — CAM интермедиате Clusia minor .Планта
201
,368
–378.Borland AM , Griffiths H, Broadmeadow MSJ, Fordham MC, Maxwell C.
1993
. Краткосрочные изменения в различении изотопов углерода у C 3 ‐CAM интермедиата Clusia minor L., произрастающего на Тринидаде.Экология
95
,444
–453.Borland AM , Griffiths H, Maxwell C, Broadmeadow MSJ, Fordham MC.
1996
. Индукция САМ у Clusia minor L. при переходе от влажного к засушливому сезону на Тринидаде: роль образования органических кислот и декарбоксилирования.Растения, клетки и окружающая среда
19
,655
–664.Borland AM , Хартвелл Дж., Дженкинс Г.И., Уилкинс М.Б., Ниммо Х.Г.
1999
. Метаболитный контроль преобладает над циркадной регуляцией киназы PEPc и фиксацией CO 2 в метаболизме крассуловой кислоты (CAM).Физиология растений
121
,889
–896.Borland AM , Tecsi L, Leegood RC, Walker RP.
1998
. Индуцируемость метаболизма крассуловой кислоты (CAM) у видов Clusia ; физиологическая/биохимическая характеристика и межклеточная локализация процессов карбоксилирования у трех видов, которые демонстрируют разную степень САМ.Планта
205
,342
–351.Boxall SF , Bohnert HJ, Cushman JC, Nimmo HG, Hartwell J.
2001
. Циркадные часы и метаболизм крассуловой кислоты у Mesembryanthemum crystallinum .Плакат S18-D12. 12-й Международный конгресс по фотосинтезу , Брисбен.Boxall SF , Bohnert HJ, Cushman JC, Nimmo HG, Hartwell J.
2002
. На пути к пониманию молекулярных событий, лежащих в основе циркадного контроля фиксации CO 2 в метаболизме крассуловой кислоты с использованием модельного САМ-растения Mesembryanthemum crystallinum . Плакат 520. Биология растений ASPB 2002 , Денвер.Бранко JB , Агари С., Эллиотт С., Борланд А.М., Кушман Дж.К.
2003
. Выделение и анализ CAM-дефектных мутантов в растении обыкновенном льдине, Mesembryanthemum crystallinum . Американское общество биологов растений. Плакат 403. ASPB Биология растений 2003 , Гонолулу, Гавайи.Brulfert J , Guerrier D, Queiroz O.
1975
. Фотопериодизм и ферментативные ритмы. Кинетические характеристики фотопериодической индукции метаболизма крассуловой кислоты.Планта
125
,987
–995.Картер PJ , Ниммо Х.Г., Фьюсон К.А., Уилкинс М.Б.
1991
. Циркадные ритмы в активности растительной протеинкиназы.Журнал EMBO
10
,2063
–2068.Чолле R , Видал Дж., О’Лири М.Х.
1996
. Фосфо енол пируваткарбоксилаза: вездесущий, строго регулируемый фермент в растениях.Ежегодный обзор физиологии растений и молекулярной биологии растений
47
,273
–298.Кристофер JT , Holtum JAM.
1996
. Закономерности распределения углеводов в листьях крассуловых видов с кислотным обменом при раскислении.Физиология растений
112
,393
–399.Cushman JC , Bohnert HJ.
1999
. Метаболизм крассуловой кислоты: молекулярная генетика.Ежегодный обзор физиологии растений и молекулярной биологии растений
50
,305
–332.Cushman JC , Borland AM.
2002
. Индукция метаболизма крассуловой кислоты ограничением воды.Растения, клетки и окружающая среда
25
,297
–312.Cushman JC , Михаловски CB, Bohnert HJ.
1990
. Онтогенетический контроль индуцируемости метаболизма крассуловой кислоты солевым стрессом у обыкновенного ледяного растения.Физиология растений
94
,1137
–1142.DeRocher EJ , Bohnert HJ.
1993
. Стресс, связанный с развитием и окружающей средой, использует разные механизмы экспрессии семейства генов растений.Растительная клетка
5
,1611
–1625.Додд AN , Borland AM, Haslam RP, Griffiths H, Maxwell K.
2002
. Метаболизм крассуловой кислоты: пластичный, фантастический.Журнал экспериментальной ботаники
53
,569
–580.Додд AN , Гриффитс Х., Тайби Т., Кушман Дж. К., Borland AM.
2003
. Интеграция метаболизма диального крахмала с циркадной регуляцией САМ в среде Mesembryanthemum crystallinum .Планта
216
,789
–797.Данлэп JC .
1999
. Молекулярные основы циркадных часов.Сотовый
96
,271
–290.Эдвардс GE , Dai Z, Cheng SH, Ku MSB.
1996
. Факторы, влияющие на индукцию метаболизма крассуловой кислоты у Mesembryanthemum crystallinum . В: Винтер К., Смит JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer-Verlag,119
–143.Эммерлих В , Линка Н., Райнхольд Т., Хюрт М.А., Трауб М., Мартиноя Э., Нойхаус Х.Е.
2003
. Растительным гомологом котранспортера натрия/дикарбоновой кислоты человека является вакуолярный переносчик малата.Proceedings of the National Academy of Sciences, USA
(в печати).Эрикссон ME , Миллар А.Дж.
2003
. Циркадные часы. Лучший друг растения в прядильном мире.Физиология растений
132
,732
–738.Гейгер DR , Шейх В-Дж, Ю Х-М.
1995
. Фотосинтетический метаболизм углерода и транслокация у мутанта дикого типа и с дефицитом крахмала Nicotiana sylvestris L.Физиология растений
107
,507
–514.Гриффитс Н .
1989
. Механизмы концентрации углекислого газа и эволюция САМ в сосудистых эпифитах. В: Lüttge U, изд. Сосудистые растения как эпифиты . Берлин: Springer-Verlag,42
–86.Гриффитс H , Хелликер Б., Робертс А., Хаслам Р.П., Гирнус Дж., Роуб В.Е., Борланд А.М., Максвелл К.
2011 900. Регуляция активности Rubisco в растениях САМ: лучше поздно, чем никогда.
Функциональная биология растения
29
,689
-696 -696Hafke JB , Hafke Y, Smith Jac, Lüttge U, Thiel G.
2003
. Вакуолярное поглощение малата опосредовано анионселективным внутренним выпрямителем.Журнал завода
35
,116
–128.Harmer SL , Hogenesch JB, Straume M, Chang H-S, Han B, Zhu T, Wang Z, Kreps JA, Kay SA.
2000
. Организованная транскрипция ключевых путей в Arabidopsis с помощью циркадных часов.Наука
290
,2110
–2113.Harmer SL , Panda S, Kay SA.
2001
.Молекулярные основы циркадных ритмов.Annual Review of Cell and Developmental Biology
17
,215
–253.Хартвелл Дж , Гилл А., Ниммо Г.А., Уилкинс М.Б., Дженкинс Г.И., Ниммо Х.Г.
1999
. Фосфо енол киназа пируваткарбоксилазы представляет собой новую протеинкиназу, регулируемую на уровне экспрессии.Журнал завода
20
,333
–342.Хартвелл Дж , Смит Л.Х., Уилкинс М.Б., Дженкинс Г.И., Ниммо Х.Г.
1996
. Высшая растительная фосфо--енол--пируваткарбоксилазная киназа регулируется на уровне транслируемой мРНК в ответ на свет или циркадный ритм.Журнал завода
10
,1071
–1078.Haslam R , Borland A, Griffiths H.
2002
. Кратковременная пластичность экспрессии метаболизма крассуловой кислоты у эпифитной бромелиевой Tillandsia usneoides L.Функциональный завод Биология
29
,749
-756 949 -756Häusler Re , Baur B, Scharte J, Teichmann T, Eicks M, Fischer KL, Flügge U-I, Schubert S, Weber A , Фишер К.
2000
. Транспортеры пластидных метаболитов и их физиологические функции в индуцибельном метаболизме крассуловых кислот Mesembryanthemum crystallinum .Журнал завода
24
,285
–296.Крепс JA , Ву И, Чан Х-С, Чжу Т, Ван Х, Харпер Дж. Ф.
2002
. Изменения транскриптома Arabidopsis в ответ на солевой, осмотический и холодовой стресс.Физиология растений
130
,2129
–2141.Li B , Geiger DR, Shieh W-J.
1992
. Доказательства циркадной регуляции синтеза крахмала и сахарозы в листьях сахарной свеклы.Физиология растений
99
,1393
–1399.Лю Y , Циноремас Н.Ф., Джонсон Ч., Лебедека Н.В., Голден С.С., Ишиура М., Кондо Т.
1995
. Циркадная оркестровка экспрессии генов у цианобактерий.Гены и развитие
9
,1469
–1478.Лю Z , Taub CC, McClung CR.
1996
. Идентификация минимального промотора рибулозо-1,5-бифосфаткарбоксилазы/оксигеназы (RCA) Arabidopsis thaliana , регулируемого светом и суточными часами.Физиология растений
112
,43
–51.Люттге U .
2000
. Тонопласт функционирует как главный переключатель циркадной регуляции метаболизма крассуловой кислоты.Планта
211
,761
–769.Люттге U .
2002
и . Циркадный ритм: «Биологические часы» аппаратные или программные?Прогресс в ботанике
64
,277
–319.Люттге U .
2002
б . CO 2 – концентрация: последствия метаболизма крассуловой кислоты.Журнал экспериментальной ботаники
53
,2131
–2142.McRae SR , Christopher JT, Smith JAC, Holtum JAM.
2002
. Транспорт сахарозы через вакуолярную мембрану Ananas comosus .Функциональная биология растений
29
,717
–724.Максвелл К .
2002
. Устойчивость полезна: суточные закономерности фотосинтеза у C 3 и толстянковые эпифитные бромелиевые метаболизма кислот.Функциональная биология растений
29
,679
–687.Максвелл К , фон Каммерер С., Эванс Дж.Р.
1997
. Является ли низкая внутренняя проводимость по CO 2 следствием суккулентности растений с метаболизмом крассуловой кислоты?Австралийский журнал физиологии растений
24
,777
–786.Максвелл К , Борланд А.М., Хаслам Р.П., Хелликер Б.Р., Робертс А., Гриффитс Х.
1999
. Модуляция активности Rubisco во время дневных фаз растения толстянковых кислотного метаболизма Kalanchoë daigremontiana .Физиология растений
121
,849
–856.Майкл TP , McClung CR.
2003
. Ловушка энхансера выявляет широко распространенный контроль транскрипции циркадных часов у Arabidopsis .Физиология растений
132
,629
–639.Ниммо Г.А. , Ниммо Х.Г., Гамильтон И.Д., Фьюсон К.А., Уилкинс М.Б.
1986
. Очистка фосфорилированной ночной формы и дефосфорилированной дневной формы фосфо энол пируваткарбоксилазы из Bryophylum fedtschenkoi .Биохимический журнал
239
,213
–220.Ниммо Г.А. , Уилкинс М.Б., Фьюсон К.А., Ниммо Х.Г.
1987
. Стойкий циркадный ритм в состоянии фосфорилирования фосфо енол пируваткарбоксилазы листьев Bryophylum fedtschenkoi и ее чувствительность к ингибированию малатом.Планта
170
,408
–415.Ниммо HG .
2000
. Регуляция фосфо--енол--пируваткарбоксилазы в САМ-растениях.Тенденции в растениеводстве
5
,75
–80.Нобель ПС .
1996
. Высокая продуктивность некоторых агрономических видов САМ. В: Винтер К., Смит JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer-Verlag,255
–265.Rascher U , Hütt MT, Siebke K, Osmond CB, Beck F, Lüttge U.
2001
. Пространственно-временная изменчивость метаболизма в циркадном ритме растений: биологические часы как совокупность связанных отдельных осцилляторов.Proceedings of the National Academy of Sciences, USA
98
,11801
–11805.Ворон JA , Спайсер Р.А.
1996
. Эволюция метаболизма крассуловой кислоты. В: Винтер К., Смит JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer-Verlag,360
–385.Робертс А , Borland AM, Гриффитс Х.
1997
. Дискриминационные процессы и сдвиги карбоксилирования в фазах метаболизма крассуловой кислоты.Физиология растений
113
,1283
–1292.Rockel B , Blasius B, Beck F, Ratajcek R, Lüttge U.
1997
. Эндогенные колебания количества транскриптов субъединицы с V-АТФазы Mesembryanthemum crystallinum с гармоническими частотными резонансами при непрерывном освещении.Письма по клеточной и молекулярной биологии
2
,69
–76.Шалфей РФ .
2002
. Метаболизм крассуловой кислоты и фотосинтез C 4 несовместимы?Функциональная биология растений
29
,775
–785.Шаффер R , Ландграф Дж., Акчерби М., Саймон В., Ларсон М., Висман Э.
2001
. Микроматричный анализ суточных и циркадианно-регулируемых генов у Arabidopsis .Растительная клетка
13
,113
–123.Ширман LP , Шрирам С., Уивер Д.Р., и др. .
2000
. Взаимодействующие молекулярные петли в циркадных часах млекопитающих.Наука
288
,1013
–1019Смикенс S .
2000
. Передача сигнала, индуцированная сахаром, у растений.Ежегодный обзор физиологии растений и молекулярной биологии растений
51
,49
–81.Смит JAC , Ингрэм Дж., Циатис М.С., Баркла Б.Дж., Варфоломей Д.М., Бетти М., Пантоха О., Пеннингтон А.Дж.
1996
. Транспорт через вакуолярную мембрану у САМ-растений. В: Винтер К., Смит JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer-Verlag,53
–71.Ститт М , Мюллер К., Мэтт П., Гибон И., Карилло П., Моркуэнде Р., Шейбл В.-Р., Крапп А.
2002
. Шаги к комплексному взгляду на азотистый обмен.Журнал экспериментальной ботаники
53
,959
–970.Тайби Т , Кушман Дж.К.
1999
. Сигнальные события, приводящие к индукции метаболизма крассуловой кислоты у ледяного растения.Физиология растений
121
,545
–555.Taybi T , Cushman JC, Borland AM.
2002
. Экологическая, гормональная и циркадианная регуляция экспрессии метаболизма крассуловой кислоты.Функциональная биология растений
29
,669
–678.Taybi T , Nimmo HG, Borland AM.
2004
. Экспрессия генов фосфо--енол--пируваткарбоксилазы (PEPC) и PEPC-киназы: последствия для генотипической способности и фенотипической пластичности в экспрессии метаболизма крассуловой кислоты.Физиология растений
(в печати).Тайби T , Патил С., Чолле Р., Кушман Дж.К.
2000
. Минимальная серин/треониновая протеинкиназа циркадианно регулирует активность фосфо--енол--пируваткарбоксилазы в листьях ледяного растения, вызванного метаболизмом крассуловой кислоты.Физиология растений
123
,1471
–1481.Тайби T , Сотта Б., Гериг Х.Х., Гюклю С., Клуге М., Брулферт Дж.
1995
. Дифференциальные эффекты абсцизовой кислоты на фосфо енол пируваткарбоксилазу и действие САМ у Kalanchoë blossfeldiana .Botanica Acta
198
,240
–246.Уэбб ААР .
2003
. Физиология циркадных ритмов у растений.Новый фитолог
160
,281
–304.Уилкинс МБ .
1992
. Циркадные ритмы: их происхождение и контроль.Новый фитолог
121
,347
–375.Уильямс LE , Лемуан Р., Зауэр Н.
2000
. Переносчики сахара у высших растений — разнообразие ролей и сложная регуляция.Тенденции в растениеводстве
5
,283
–290.Зима K , Smith JAC.
1996
и .Таксономическое распределение метаболизма крассуловой кислоты. В: Винтер К., Смит JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer-Verlag,1
–13.Зима K , Smith JAC.
1996
б . Метаболизм крассуловой кислоты: современное состояние и перспективы. В: Винтер К., Смит JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция .Берлин: Springer-Verlag,389
–426.Wyka TP , Lüttge U.
2003
. Вклад карбоксилирования C 3 в циркадный ритм поглощения углекислого газа в растении Crassulacean с метаболизмом кислоты Kalanchoë daigremontiana .Журнал экспериментальной ботаники
54
,1471
–1480.Янг МВт , Кей С.А.
2001
.Часовые пояса: сравнительная генетика циркадных часов.Nature Reviews Genetics
2
,702
–715.Общество экспериментальной биологии
Влияние циркадного ритма на метаболические процессы и регуляцию энергетического баланса — Полный текст — Annals of Nutrition and Metabolism 2019, Vol. 74, № 4
Аннотация
Справочная информация: Циркадная система синхронизации или циркадные часы играют решающую роль во многих биологических процессах, таких как цикл сна-бодрствования, секреция гормонов, здоровье сердечно-сосудистой системы, гомеостаз глюкозы и регуляция температуры тела.Энергетический баланс также является одним из важнейших краеугольных камней метаболических процессов, тогда как энергетический дисбаланс связан со многими заболеваниями (например, ожирением, диабетом, сердечно-сосудистыми заболеваниями). Циркадные часы являются основным регулятором метаболизма, и этот анализ дает представление о двунаправленном влиянии циркадных ритмов на метаболические процессы и энергетический баланс. Резюме: Циркадная система синхронизации или циркадные часы играют решающую роль во многих биологических процессах, но рост деятельности, которая осуществляется круглосуточно и без выходных, и повсеместное использование телевидения, Интернета и мобильных телефонов почти 24 часа в сутки приводит к постепенное уменьшение адекватного времени сна.Согласно последним исследованиям, долгосрочные нарушения циркадных ритмов связаны со многими патологическими состояниями, такими как преждевременная смертность, ожирение, нарушение толерантности к глюкозе, диабет, психические расстройства, тревога, депрессия и прогрессирование рака, тогда как кратковременные нарушения связаны с ухудшением самочувствия. , усталость и потеря концентрации. В этом обзоре были рассмотрены циркадные ритмы метаболических процессов и их влияние на энергетический баланс. Ключевые сообщения: Суточный ритм имеет двунаправленное взаимодействие практически со всеми метаболическими процессами.Таким образом, понимание основной причины, влияющей на циркадные часы, и создание рекомендаций по лечению с использованием циркадных ритмов может повысить успех лечения заболевания. Хронофармакология, хронопитание и хроноупражнения являются новыми подходами к лечению метаболического баланса.
© 2019 S. Karger AG, Базель
Введение
Термин циркадный происходит от двух латинских слов circa (приблизительно) и dies (день), обозначающих примерно «один день» [1].Циркадный ритм также называют биологическими/циркадными часами [2] и относится к поведенческим, физиологическим и молекулярным изменениям с продолжительностью цикла примерно 24 часа [3]. Циркадные часы можно разделить на 2 части: центральные часы, находящиеся в супрахиазматическом ядре (СХЯ) гипоталамуса, которое получает световые сигналы, и периферические часы, находящиеся в различных тканях по всему телу. Периферические часы играют неотъемлемую и уникальную роль в каждой из соответствующих тканей, управляя циркадной экспрессией специфических генов, участвующих в различных физиологических функциях [4].
Основным раздражителем СХЯ является свет [5]. Однако у слепых есть циркадные циклы (например, циклы сна и бодрствования), и этот цикл длится дольше 24 часов. Это открытие привело к мысли, что другие раздражители, помимо света, могут выступать в качестве стимула для биологических часов человека [6]. Обзорное исследование показало, что температура, гормоны, питательные вещества, распределение питательных веществ, некоторые питательные вещества (отдельно; например, глюкоза, аминокислоты, этанол и ретиноевая кислота), состояние питания/натощак, состояние сна-бодрствования, физическая активность являются эффективными стимулами для циркадный цикл в различных периферических путях [5].Интересно, что сообщалось о различиях в эффектах циркадного ритма у мужчин по сравнению с женщинами [7].
В основном цикл сна-бодрствования регулирует циркадный ритм. Однако в современном мире рост деятельности, которая осуществляется круглосуточно и без выходных, и повсеместное использование телевидения, интернета и мобильных телефонов почти 24 часа в сутки приводит к постепенному уменьшению адекватного времени сна [8]. Эпидемиологическое исследование показало, что продолжительность ночного сна уменьшилась на 18 минут за последние 30 лет [9].Ухудшение цикла сон-бодрствование, особенно у здоровых людей, может быть основной причиной многих заболеваний, таких как преждевременная смертность, ожирение, нарушение толерантности к глюкозе, сахарный диабет, психические расстройства, тревожность, депрессия и прогрессирование рака, утомляемость, потери. концентрации [10, 11]. Другое эпидемиологическое исследование показало, что работа не менее 3 ночей в месяц в течение 15 и более лет может увеличить риск колоректального рака у женщин [12]. У крыс нарушение циркадных обмороков ускоряет развитие диабета, вызывая потерю функции и массы бета-клеток [13].В экспериментальном исследовании расстройство циркадного ритма вызывало сердечно-сосудистые и почечные повреждения у хомяков [14]. В исследовании, проведенном с пациентами с шизофренией, у участников были серьезные циркадные расстройства сна-бодрствования, хотя их настроение, психический статус и психотические эпизоды были стабильными [15]. В этом обзоре были рассмотрены роль циркадианного ритма в метаболических процессах и двунаправленное влияние циркадного ритма на энергетический баланс. В соответствии с этой целью были проанализированы исследования, опубликованные с 2000 по 2018 год.Кроме того, были добавлены 3 важные статьи, опубликованные в 1984, 1993 и 1996 годах. В основном для сканирования научных статей использовались базы данных Google Academic (библиографическая база данных), PubMed, Scopus, Web of Science и Science Direct. Кроме того, были просканированы ресурсы Центральной библиотеки Университета Гази. Такие термины, как «циркадные часы или циркадный ритм» или «биологические часы» и «гормоны или обмен веществ, или энергетический баланс, или расход энергии, или термогенез, метаболический гомеостаз или метаболическая регуляция, или часовые гены, или диетический состав, режим приема пищи, время приема пищи или физическая активность». активность, упражнения или спорт» были использованы в качестве ключевых слов.В конце этого сканирования было найдено 4456 статей. После проверки заголовков и аннотаций дублирующиеся статьи и статьи, опубликованные не на английском языке, были исключены. Окончательное количество статей составило 82, включая статьи, которые были бесплатными и доступными в виде полного текста. Кроме того, главы книги также использовались для изучения предмета.
Циркадная синхронизация в метаболическом гомеостазе
Многие исследования объясняют взаимосвязь между физиологией человека, некоторыми заболеваниями и циркадными ритмами [12-15].Метаболический гомеостаз является важнейшим компонентом, регулирующим энергетический обмен, особенно в жировой ткани. Жировая ткань является центральным метаболическим органом, который регулирует энергетический гомеостаз всего организма. Белая жировая ткань функционирует как ключевой резервуар энергии для других органов, тогда как бурая жировая ткань накапливает липиды для адаптивного термогенеза, индуцированного холодом. Жировая ткань секретирует различные гормоны, цитокины и метаболиты (называемые адипокинами), которые контролируют системный энергетический баланс, регулируя сигналы аппетита от центральной нервной системы, а также метаболическую активность в периферических тканях [16].Например, лептин имеет специфические рецепторы в гипоталамусе и высвобождается из основных адипоцитов. Этот гормон играет регулирующую роль в энергетическом обмене, усиливая активацию симпатической нервной системы и повышая термогенез за счет увеличения гормонов щитовидной железы. В термогенезе белок UCP (разобщающий) ингибирует синтез АТФ в митохондриях, позволяя энергии потребляться в виде тепла. Лептин повышает уровень гормонов щитовидной железы и активирует симпатическую центральную нервную систему, что приводит к большему образованию UCP и, следовательно, к большему потреблению энергии [17].Высвобождение гормона лептина происходит в циркадном цикле, а уровень лептина в сыворотке достигает пика ночью [18]. Таким образом, нарушение циркадного баланса может косвенно влиять на секрецию лептина, термогенез и энергетический гомеостаз.
Напротив, некоторые гормоны, выделяемые гипоталамусом, проявляют большую активность ночью. Наиболее ярким примером этого является «гормон роста». Уровни гормона роста достигают пика между 2:00 и 4:00 утра. Если перекрестная связь между циркадным ритмом и гормоном роста нарушается из-за нарушений сна, гормон роста не может высвобождаться на нормальном уровне.Поэтому необходимо уделять особое внимание режиму сна детей [19].
Кортизол — это стероидный гормон, секретируемый надпочечниками. Он регулирует многие метаболические процессы, такие как гликогенолиз, липолиз и протеолиз [20]. Количество и частота секреции кортизола регулируется циркадным ритмом. Концентрация кортизола в кровотоке достигает пика непосредственно перед пробуждением утром. Уровень кортизола постепенно снижается в течение дня.Самый низкий уровень он достигает во время сна после полуночи [21]. Кортизол является основным гормоном, который регулирует метаболические процессы в организме. Он увеличивает использование кортизола, глюкозы, свободных жирных кислот и аминокислот из запасов эндогенного топлива. Таким образом, высокий уровень кортизола действует как катаболический гормон, который снижает мышечную и мышечную массу, а также увеличивает потребление энергии [22]. Кроме того, толерантность к глюкозе и секреция инсулина меняются в течение дня. При естественном течении метаболизма как чувствительность к инсулину, так и секреция инсулина снижаются ночью (особенно между 3 и 5 часами утра).м.) по сравнению с утренними часами. Этот метаболический процесс, феномен рассвета, подчеркивает влияние циркадного контроля на метаболизм глюкозы [23]. В естественных физиологических процессах организма гормоны, работающие как антагонисты инсулина (особенно гормон роста), проявляют гиперинсулинемическую активность из-за снижения секреции инсулина между 3:00 и 5:00 утра, так что уровень сахара в крови возвращается к норме. Этому противодействует дополнительная физиологическая секреция инсулина у людей, не страдающих диабетом или инсулинозависимых.И наоборот, когда высвобождение инсулина нарушено, действие гормона роста, высвобождаемого в ночное время, особенно у больных сахарным диабетом, не может быть смягчено. Это приводит к патологическому циркадному ритму, который может привести к утренней гипергликемии независимо от режима питания [24].
Мелатонин является важным гормоном в циркадной синхронизации. Этот гормон участвует во многих биологических и физиологических процессах в организме. Это эффективный гормон для человеческого биоритма (циркадного ритма).Основная роль этого гормона заключается в поддержании биологических часов и регулировании ритма тела [25]. Синтез и высвобождение мелатонина стимулируется в темноте, ночью, а днем подавляется на свету [1]. Особенно между 23:00. и 5:00 утра пик секреции мелатонина и его концентрация в крови увеличивается в 3-10 раз [25]. Однако воздействие света в ночное время вызывает падение уровня мелатонина в плазме [1].
На метаболизм липидов также влияют циркадные колебания.Исследования показывают, что многие белки, связанные с метаболизмом липидов (например, ApoB, ApoA1 и ApoA4), кишечный микросомальный транспортный белок триглицеридов и кишечный белок, связывающий жирные кислоты, изменяются в течение дня [26-28]. Кроме того, исследования на мышах показывают, что поглощение холестерина и липидов в темновой фазе выше, чем в светлой фазе [28]. Некоторые продукты метаболизма липидов также имеют циркадный ритм. Например, циркулирующие неэтерифицированные жирные кислоты у людей выше ночью из-за повышенной липолитической активности [29].Кроме того, эпизоды инфаркта миокарда и астмы связаны с циркадным циклом метаболизма. Эти приступы достигают пика ночью или рано утром [30, 31]. В ранние утренние часы улучшается работа сердечно-сосудистой системы в ответ на активацию деятельности симпатического нерва, а в вечерние часы наблюдается пик артериального давления и частоты сердечных сокращений. Сердечные приступы, особенно рано утром или вечером, могут быть вызваны этим суточным ритмом сердечно-сосудистой системы.Суточный ритм гормонов и некоторых обменных процессов представлен на рис. 1.
Рис. 1.
Суточный ритм гормонов и некоторых обменных процессов. Высвобождение гормона лептина происходит в соответствии с циркадным циклом, и пик уровня лептина в сыворотке крови приходится на ночное время. Уровни гормона роста достигают пика между 02:00 и 04:00 утра. Концентрация кортизола в кровообращении достигает пикового уровня непосредственно перед пробуждением утром. Секреция инсулина снижается ночью (особенно между 03:00 и 05:00 утра).м.). Синтез и высвобождение мелатонина стимулируется ночью в темноте, а днем подавляется светом. Поглощение липидов в темновой фазе выше, чем в светлой. Пик инфаркта миокарда приходится на ночное или раннее утро.
Метаболическая регуляция циркадных ритмов
У млекопитающих циркадные ритмы контролируются в основном СХЯ, которые называются главными часами. Супрахиазматические ядра состоят из множества одноклеточных циркадианных осцилляторов, расположенных в передней области гипоталамуса головного мозга, и при синхронизации производят скоординированные циркадные сигналы [32].SCN активируется через нервный пучок, называемый «ретиногипоталамическим трактом». Таким образом, СХЯ регулирует биологические часы живых существ и способствует физиологическим процессам, стимулируя другие области мозга [2].
Супрахиазматические ядра синапсируют непосредственно с вентральной и дорсальной субвентрикулярными областями, телами клеток, расположенными в вентральной и дорсальной субвентрикулярных областях, и дорсомедиальным гипоталамусом. Несмотря на то, что эти области взаимодействуют друг с другом, нейроны, расположенные в дорсальной наджелудочковой области, более эффективно регулируют термогенез, тогда как вентральная наджелудочковая область в основном играет роль в регуляции циклов сон-бодрствование и активности.Кроме того, паравентрикулярный гипоталамус отвечает за высвобождение кортикостероидов, тогда как латеральный гипоталамус отвечает за питание и бдительность [33].
Хотя СХЯ функционирует как основные биологические часы метаболизма, исследования, проведенные в 2000-х годах, показали, что вегетативные циркадианные осцилляторы, присутствующие в периферических органах и тканях, таких как печень, кишечник, сердце и сетчатка, участвуют в метаболических процессах через клеточные часовых генов в этих органах/тканях [32, 34, 35].Супрахиазматические ядра играют важную роль в регуляции метаболизма глюкозы. Чувствительность к инсулину и усвоение глюкозы нарушены у крыс с поражением СХЯ [36]. Ухудшение циркадных часов может привести к нарушению секреции инсулина и гипоинсулинемии [29]. Белки CLOCK и BMAL1, связанные с циркадным ритмом, участвуют в производстве и высвобождении инсулина путем связывания с дистальными областями β-клеток поджелудочной железы, регулирующими циркадный ритм [37]. Как при диабете 1, так и при диабете 2 типа наблюдается недостаточность или отсутствие инсулина из-за повреждения β-клеток.Поэтому роль циркадных ритмов в этиологии диабета не следует упускать из виду с точки зрения повреждения β-клеток [38]. Часовые гены и задачи, регулирующие циркадные ритмы у млекопитающих, приведены в таблице 1 [2].
Таблица 1.
Гены циркадных ритмов и их роль [2]
У млекопитающих гомеостаз циркадных часов обеспечивается механизмами обратной связи (отрицательной) и прямой связи (положительной), влияющими на транскрипцию, трансляцию и посттрансляционную события [5, 29].Паттерн транскрипционной обратной связи опосредован белками Cry1, Cry2, Per1 и Per2. Белки CLOCK и BMAL1 связываются с Е-промоторной областью генов Per и Cry и индуцируют их экспрессию. На более поздней стадии Per и Cry гетеродимеризуются и перемещаются из цитоплазмы в ядро, чтобы ингибировать CLOCK/BMAL1-индуцированную экспрессию генов [37, 39]. Этот механизм отрицательной обратной связи важен для эффективной работы циркадной системы.
Суточный ритм и энергетический гомеостаз
Энергетический обмен регулируется многими гормонами, ферментами и транспортными системами, а циркадный ритм эффективно модулирует их экспрессию, секрецию и/или активацию [40].Метаболические пути, обеспечивающие энергетический гомеостаз, координируются метаболитами, которые предлагают изменения и активно подготавливают молекулярную среду, а также сигнальными системами, которые мгновенно реагируют на изменения в циркадных часах [41].
Гормональные рецепторы, взаимодействия между генами и внутриклеточные реакции окисления/восстановления управляют регуляцией энергетического метаболизма на клеточном уровне. Эти пути имеют важные взаимодействия с часами биологического метаболизма.Циркадные часы могут влиять на функцию рецепторов гормонов (рецептор, активирующий пролиферацию пероксисом [PPAR]α, PPARγ и REV-ERBα) и некоторых генов на клеточном уровне (сиртуин) [41, 42].
Рецепторы ядерных гормонов (PPARα, PPARγ, REV-ERBα, RORα, HNF4α, TRα и NURR1) и лиганды, присутствующие в метаболических тканях, функционируют как сенсоры, объединяющие циркадные и метаболические пути [41]. Например, PPAR являются связующим звеном между циркадными часами и энергетическим обменом. PPARγ локализуется в жировой ткани и активирует транскрипционные факторы, которые усиливают липогенез и накопление липидов.PPARα от ядерных гормональных рецепторов запускает кетогенез и окисление жирных кислот в печени в ответ на голодание. PPARδ является наиболее распространенным рецептором ядерных клеток в организме и способен коррелировать ежедневные изменения температуры тела с циркадными часами [43].
Ритмическая экспрессия и активация метаболических путей в основном связаны с координацией часовых генов (BMAL1, Per2, Per1, Per3, Cry1 и Cry2) в печени и жировой ткани. Существует связь между белком BMAL1, липогенными путями и механизмами клеточных часов.REV-ERBα (репрессор транскрипции BMAL1) и RORα (позитивный регулятор BMAL1) являются ядерными рецепторами гормонов, регулирующими липогенез. Кроме того, оба модулируются CLOCK: BMAL1. PPARα играет роль в метаболизме липидов и липопротеинов. PPARα напрямую связывается с промоторной областью белка BMAL1, который регулирует экспрессию PPARα через гетеродимер CLOCK:BMAL1 [44].
Другим фактором, влияющим на функцию циркадного ритма, являются гены сиртуина (SIRT). Гены семейства SIRT представляют собой NAD+-зависимые ферменты деацетилазы класса III, которые влияют на многие клеточные функции, включая метаболизм человека, старение, рак и старение клеток.Существует 7 вариантов семейства SIRT (SIRT 1–7). Регуляция экспрессии генов является наиболее важным механизмом, на который влияет семейство SIRT. SIRT1, SIRT6 и SIRT7 преимущественно локализуются в ядре клетки, SIRT2 — в цитоплазме, SIRT3, SIRT4 и SIRT5 — в митохондриях. SIRT1 играет значительную роль в регуляции метаболических процессов, таких как чувствительность к инсулину, метаболизм липидов и глюконеогенез, а также продолжительности жизни человека [45]. SIRT1 модулирует активность CLOCK: BMAL1 и вносит свой вклад в циркадный цикл.Гены SIRT 3–5 регулируют внутриклеточные пути, такие как окисление жирных кислот, кетогенез, цикл мочевины и окислительное фосфорилирование [33].
Статус внутриклеточного окисления-восстановления (редокс) является важным фактором, который регулирует тактовые гены в периферических тканях. Белки CLOCK могут эффективно связываться только в присутствии восстановленных последовательностей NADH и NADPH, BMAL1 и E-box. Напротив, формы никотинамидадениндинуклеотидоксидазы (НАД+ и НАДФ+) ингибируют связывание комплекса ЧАСЫ: BMAL1 с ДНК [32].Таким образом, окислительно-восстановительный статус NAD/NADH клетки может приводить к изменениям циркадных фаз, влияя на транскрипционную активность генов BMAL1:CLOCK [46].
Внутриклеточные уровни НАД+, а также повышенные уровни АМФ (аденозинмонофосфата) являются индикаторами низкого уровня энергии. Когда уровни внутриклеточного АТФ снижаются, AMPK (AMP-активирующая протеинкиназа) действует как пищевой сенсор и активирует пути внутриклеточного снабжения энергией. Таким образом, соотношение АМФ/АТФ также может быть связующим звеном между циркадным ритмом и энергетическим обменом [47].
Влияние циркадианного ритма на энергетический баланс
Энергия, вырабатываемая и запасаемая в процессе метаболизма, используется для поддержания метаболической активности, такой как основной уровень метаболизма, физическая активность и тепловое воздействие пищевых продуктов [48].
Фрэнсис Г. Бенедикт впервые описал циркадные изменения энергетического метаболизма в 1915 году [49]. Кроме того, Хауген и соавт. [50] обнаружили, что уровень метаболизма в покое был на 6% выше в полдень, чем в утренние часы. Одним из наиболее важных факторов, влияющих на скорость основного обмена, является характер сна.Сон и циркадный ритм являются основными компонентами регуляции энергетического обмена [48, 51]. Различают 2 фазы сна: быстрое движение глаз (БДГ) и не-БДГ [52]. Активность симпатической нервной системы и сновидения увеличиваются в период БДГ. Температура тела, частота сердечных сокращений, частота дыхания и артериальное давление увеличиваются во время БДГ. Нарушения в БДГ-периоде нарастают за счет активности симпатической нервной системы [53]. Из-за повышения температуры тела и расхода энергии в головном мозге (затраты энергии мозга в этот период составляют примерно 25 %) скорость метаболизма во время сна достигает наивысшей точки в фазе БДГ [54].Следовательно, ухудшение цикла сна из-за позднего сна, смены часовых поясов, сменной работы и т. д. может привести к снижению основного обмена за счет изменения времени фазы быстрого сна.
В дополнение к основному уровню метаболизма в состоянии покоя физическая активность является важным компонентом общего расхода энергии [55]. Тем не менее, исследования подчеркивают различные метаболические эффекты типа, продолжительности и продолжительности упражнений. Например, в обзорном исследовании подчеркивается, что максимальная производительность в краткосрочных анаэробных упражнениях достигается ближе к вечеру, обычно в полдень [56], что также соответствует пиковой температуре тела [56, 57].Температура тела считается «базовой переменной» циркадного ритма и используется в качестве маркера циркадного ритма [58]. Разница в температуре тела между утренними и вечерними часами составляет 0,9°С [59]. Температура тела влияет на мышечную активность [60]. Эта разница между утренними и вечерними часами может повлиять на физическую работоспособность и, косвенно, на скорость основного обмена.
Время приема пищи влияет на физическую работоспособность. Упражнения до или после еды по-разному влияют на окисление жирных кислот и метаболизм аппетита [61-63].Тем не менее, до сих пор неясно, когда (упражнения до еды или после еды) самое эффективное время для похудения. Подход к снижению массы тела и облегчению потери жира заключается в выполнении постпрандиальных аэробных упражнений после ночного голодания [64]. Упражнения во время голодания вызывают использование отложений гликогена для увеличения окисления жиров и снижают уровень инсулина в плазме за счет повышения уровня адреналина и норадреналина в плазме, вызывая липолиз. Напротив, некоторые исследования были сосредоточены на положительном влиянии постпрандиальных упражнений на контроль массы тела по сравнению с упражнениями во время голодания из-за их положительного влияния на аппетит и метаболизм в состоянии покоя [65].Исследование показало, что 36-минутные умеренные упражнения на беговой дорожке значительно увеличивают скорость метаболизма в покое через 40 минут после завтрака в средиземноморском стиле в первые 24 часа [66]. Во многих исследованиях общее потребление энергии, режим питания, физические характеристики, а также продолжительность и тяжесть упражнений различаются, что приводит к различным интерпретациям результатов.
Еще одним компонентом общего расхода энергии является тепловой эффект питательных веществ. На тепловой эффект продуктов оказывает влияние макроэлементный состав рациона.Липиды обладают самым низким (0-3%), а белки — самым высоким (20-30%) термогенным эффектом [67]. Кроме того, важным фактором, влияющим на тепловой эффект продуктов, является время приема пищи. Диетический термогенез выше в утренние часы по сравнению с вечерними и ночными часами [68]. Точно так же в исследовании Morris et al. [69], тепловой эффект от пищи утром был на 44% выше, чем вечером. Снижение теплового эффекта пищи с утра к вечеру может быть в первую очередь связано с влиянием эндокринной циркадной системы на физиологию желудочно-кишечного тракта.Моторика кишечника в утренние часы более эффективна, чем в вечерние. Два исследования на здоровых взрослых показали, что скорость опорожнения желудка утром (8:00) была выше, чем вечером (20:00–23:00) [70]. Кроме того, нарушения циркадного ритма, такие как посменная работа и синдром смены часовых поясов, вызывают расстройства желудочно-кишечного тракта, такие как боль в животе, вздутие живота, диарея или запор. Эти наблюдения показывают функциональную корреляцию между суточными ритмами и физиологией желудочно-кишечного тракта.Хугерверф и др. [71] показали, что экспрессия белков PER2 и BMAL1 была связана с циркадным ритмом в области межмышечного сплетения, который играет важную роль в координации эпителиальных клеток толстой кишки и подвижности толстой кишки.
Влияние рациона питания и физической активности на циркадные ритмы
Здесь подчеркивается влияние циркадных ритмов на компоненты расхода энергии и их роль в энергетическом балансе. Однако некоторые исследования показали, что некоторые факторы окружающей среды (т.е., пища, состав пищи, время кормления и упражнения) могут быть эффективны в отношении периферических часов, которые существуют в нескольких частях тела, таких как печень, поджелудочная железа или сердце [72-75]. Периферические часы играют неотъемлемую и уникальную роль в каждой из соответствующих тканей, управляя циркадной экспрессией специфических генов, участвующих в различных физиологических функциях. Существование всех этих часов, работающих вместе и синхронизированных центральными часами, со многими гормонами и физиологическими или экологическими переменными, изменяющимися в течение дня, делает это двунаправленное взаимодействие в циркадной системе довольно сложным [4].
Еда — один из внешних синхронизаторов наших периферийных часов. Основная роль циркадных часов состоит в том, чтобы увлечь организм сигналами окружающей среды; это позволяет организмам предсказывать доступность пищи. Ограничение доступа к пище в определенное время суток оказывает сильное влияние на поведение и физиологию организмов [72]. Дамиола и др. [76] показали, что временное ограничение питания в условиях свет-темнота или темнота-темнота может изменять фазу циркадной экспрессии генов в периферических типах клеток на срок до 12 часов, оставляя фазу циклической экспрессии генов в СХЯ неизменной.
Время кормления оказывает существенное влияние на репертуар, фазу и амплитуду ритмической экспрессии генов. В исследовании было показано, что как временной характер потребления пищи, так и циркадные часы влияют на транскрипцию печеночных генов у мышей дикого типа [73].
Состав рациона — еще один важный фактор, влияющий на циркадные часы. Косака и др. [74] показали, что диета с высоким содержанием жиров у мышей приводила к изменению периода ритма локомоторной активности, изменению экспрессии и цикличности канонических генов циркадных часов, ядерных рецепторов, регулирующих факторы транскрипции часов, и задействованных часов-контролируемых генов. в утилизации топлива в гипоталамусе, печени и жировой ткани.
Одним из примечательных несветовых сигналов для регуляции периферийных часов является упражнение. Предполагается, что физическая активность или упражнения вызывают несколько физиологических изменений, таких как изменение температуры тела и гормонального статуса, которые, как известно, влияют на периферические часы посредством активации симпатической нервной системы и высвобождения глюкокортикоидов [75]. Повышение температуры тела может действовать как входной сигнал для циркадного водителя ритма млекопитающих [77]. Кроме того, физические упражнения способствуют выработке и высвобождению мелатонина.Положительный эффект 4-недельного лечения мелатонином для модуляции циркадных компонентов цикла сна-бодрствования обычно приводит к улучшению качества сна [78].
Напротив, молекулярные циркадные часы в периферических тканях могут реагировать на время упражнений, что позволяет предположить, что физическая активность обеспечивает важную информацию о времени для синхронизации циркадных часов во всем теле. Хотя точная продолжительность и интенсивность упражнений, необходимых для изменения циркадных ритмов, не были определены, одно исследование на мышах показало, что низкоинтенсивные упражнения на выносливость, выполняемые в течение 2 часов в день в течение 4 недель, были достаточными для включения циркадных часов и изменить циркадный ритм [79].
Как правило, когда периферийные часы десинхронизируются с центральными часами, это приводит к хроноразрыву [80]. Это физиологическое изменение связано с различными заболеваниями, такими как рак, сердечно-сосудистые заболевания, депрессия, ожирение и метаболический синдром [72]. Например, при лечении ожирения основным подходом к диетотерапии является ограничение потребления энергии [81]. Как правило, факторы, непосредственно влияющие на биологические ритмы, такие как время приема пищи и время сна, обычно не учитываются при планировании диеты.Суточные часы играют важную роль в энергетическом гомеостазе и метаболических процессах. Таким образом, оценка факторов (посменная работа, нерегулярный сон, бессонница и т. д.), которые могут привести к нарушениям циркадных ритмов у лиц с метаболическими заболеваниями, такими как ожирение, и планирование физических упражнений и времени приема пищи в соответствии с нормальными биологическими ритмами. например, выбор белковосодержащих продуктов по утрам для повышения термогенеза) может повысить эффективность лечения.
В последние годы для поддержания метаболического здоровья были разработаны методы лечения, зависящие от циркадных ритмов, диетические вмешательства и подходы к физическим упражнениям, которые называются «хронофармакологией», «хронопитанием» и «хроноупражнениями» соответственно.Хронофармакология изучает правильное время введения доз препарата для повышения его эффективности, всасывания и/или действенности [65]. Например, фермент HMG-COA, фермент, ограничивающий уровень холестерина, демонстрирует циркадный ритм у людей. Пик активности этого фермента приходится на ночь, поэтому рекомендуется принимать препараты, снижающие уровень холестерина, такие как статины, ночью, чтобы максимизировать их эффективность [19]. Хронопитание — это подход к определению оптимального потребления питательных веществ для поддержания здоровья и регулирования циркадного ритма [82].Например, кофеин, нобилетин (флавоноид, присутствующий в цитрусовых) и ресвератрол в пищевых продуктах могут вызывать изменения циркадного ритма на молекулярном или поведенческом уровне [83]. Хроноупражнения в первую очередь исследуют влияние продолжительности упражнений на поддержание здоровья и спортивные результаты, быстрые изменения в системе внутренних часов или перерегуляцию циркадных часов [84].
В результате циркадный ритм имеет двунаправленное взаимодействие практически со всеми метаболическими процессами и является основным фактором, влияющим на цикл сна-бодрствования.Таким образом, изучение и использование характера сна, информации о качестве и создание рекомендаций по лечению с использованием циркадных ритмов могут повысить успех лечения заболевания. По этой причине могут быть разработаны новые подходы, перспективы и стратегии лечения метаболического баланса.
Благодарности
Нет.
Заявление об этике
Эта статья не содержит никаких исследований с участием людей или животных, проведенных кем-либо из авторов.
Заявление о раскрытии информации
Авторы заявляют об отсутствии конфликта интересов.
Источник финансирования
Нет.
Взносы авторов
Ю.С. провел обзор литературы и Н.А.Т. организована и оформлена рукопись. Все авторы читали и одобрили окончательный вариант рукописи.
Ссылки
- Акынджи Э., Орхан Фё.Sirkadiyen ritim uyku bozuklukları. Псикиятр Гюнцел Якласимлар. 2016;8(2):178–89.
- Озбайер С, Дегирменджи И. Sirkadiyen saat, hücre döngüsü ve kanser. Дикле Тип Дергиси; 2011. с. 38.
- Гумз М.Л., редактор.Циркадные часы: роль в здоровье и болезни. Спрингер; 2016.
- Ричардс Дж., Гамз М.Л. Прогресс в понимании периферических циркадных часов. FASEB J. 2012 Sep; 26 (9): 3602–13.
- Гюльдур Т., Отлу Х.Г.Циркадный ритм у млекопитающих: время есть и время спать. Биол Ритм Рез. 2017;48(2):243–61.
- Аллен РП. Рассмотренная статья: увлечение мелатонином свободных циркадных ритмов у слепых. Сон Мед. 2001 март; 2 (2): 167–8.
- Санти Н., Лазар А.С., Маккейб П.Дж., Ло Дж.С., Гроегер Дж.А., Дейк Д.Дж.Половые различия в циркадной регуляции сна и бодрствования у людей. Труды Национальной академии наук. 2016:201521637.
- Ферри Дж. Э., Кумари М., Сало П., Сингх-Ману А., Кивимяки М. Эпидемиология сна — быстро развивающаяся область. Издательство Оксфордского университета; 2011.
- Кронхольм Э., Партонен Т., Лаатикайнен Т., Пелтонен М., Хярма М., Хаблин С. и др. Тенденции в самооценке продолжительности сна и симптомов, связанных с бессонницей, в Финляндии с 1972 по 2005 год: сравнительный обзор и повторный анализ выборок финского населения. J Сон Res. 2008 март; 17 (1): 54–62.
- Чжу Л., Зи ПК. Нарушения циркадного ритма сна. Нейрол клин. 2012 ноябрь; 30 (4): 1167–91.
- Авраам Ф.Обзор функциональных причин бесплодия у коров. JFIV Reprod Med Genet. 2017;5(2):203.
- Шернхаммер Э.С., Ладен Ф., Спейзер Ф.Е., Уиллетт В.К., Хантер Д.Дж., Кавачи И. и др. Работа в ночную смену и риск колоректального рака в исследовании здоровья медсестер. J Natl Cancer Inst.2003 г., июнь; 95 (11): 825–8.
- Гейл Дж. Э., Кокс Х. И., Цянь Дж., Блок Г. Д., Колвелл К. С., Матвеенко А. В. Нарушение циркадных ритмов ускоряет развитие диабета из-за потери и дисфункции бета-клеток поджелудочной железы. J Биол Ритмы. 2011 окт; 26 (5): 423–33.
- Мартино Т.А., Оудит Г.Ю., Герценберг А.М., Тата Н., Колетар М.М., Кабир Г.М. и др.Дезорганизация циркадных ритмов вызывает серьезные сердечно-сосудистые и почечные заболевания у хомяков. Am J Physiol Regul Integr Comp Physiol. 2008 г., май; 294 (5): R1675–83.
- Бакли П. Сон и нарушение циркадного ритма при шизофрении. Ежегодник психиатрии и прикладного психического здоровья; 2013.стр. 411–2.
- Чхве С.С., Ху Дж.И., Хван И.Дж., Ким Джи, Ким Дж.Б. Ремоделирование жировой ткани: ее роль в энергетическом обмене и метаболических нарушениях. Фронт Эндокринол (Лозанна). 2016 апр;7:30.
- Чон М., Далгин Д., Дженезиз М., Дженезис С.Лептин ве адипонектинин enerji ве egzersiz ilişkisi. 2017.
- Цудзино Н., Сакураи Т. [Суточный ритм лептина, орексина и грелина]. Нихон Ринсё. 2012 июль; 70 (7): 1121–5.
- Сато Т., Ида Т., Кодзима М.Роль биологических ритмов в выполнении двигательной активности. J Phys Fit Sports Med. 2017;6(3):125–34.
- МакГиннис Г.Р., Янг М.Э. Суточная регуляция метаболического гомеостаза: причины и последствия. Естественный научный сон. 2016 г., май; 8: 163–80.
- Ибрагим Эрдемир ET.Kortizol Sirkadiyen Ritmini Etkileyen Bazı Fiziksel ve Fizyolojik Parametrelerin Karşılaştırılması. Balıkesir Üniversitesi Sosyal Bilimler Enstitüsü Dergisi. 2008; 11:1–10.
- Christiansen JJ, Djurhuus CB, Gravholt CH, Iversen P, Christiansen JS, Schmitz O, et al. Влияние кортизола на углеводный, липидный и белковый обмен: исследования острой абстиненции кортизола при адренокортикальной недостаточности.J Clin Endocrinol Metab. 2007 г., сен; 92 (9): 3553–9.
- Болли Г.Б., Де Фео П., Де Космо С., Перриелло Г., Вентура М.М., Кальчинаро Ф. и др. Демонстрация феномена рассвета у нормальных людей-добровольцев. Сахарный диабет. 1984 г., декабрь; 33 (12): 1150–3.
- Рыбицкая М., Крысяк Р., Окопень Б.Феномен утренней гипергликемии и эффект Шомоди — два феномена утренней гипергликемии. Эндокринол пол. 2011;62(3):276–84.
- Озчелик Ф., Эрдем М., Болу А., Гюльсун М. Мелатонин: генель озелликлери ве психиятрик бозуклардаки ролю. Псикиятр Гюнцел Якласимлар. 2013;5(2).
- Пан Х, Чжан И, Ван Л, Хуссейн ММ.Суточная регуляция MTP и триглицеридов плазмы с помощью CLOCK опосредована SHP. Клеточный метаб. 2010 авг; 12 (2): 174–86.
- Пан Х, Хуссейн ММ. Суточная регуляция микросомального белка-переносчика триглицеридов и уровней липидов в плазме. Дж. Биол. Хим. 2007 г., август; 282 (34): 24707–19.
- Пан Х, Хуссейн ММ.Часы важны для пищевой и циркадной регуляции поглощения макронутриентов у мышей. J липидный рез. 2009 сен; 50 (9): 1800–13.
- Бейли С.М., Удох США, Янг МЭ. Циркадная регуляция метаболизма. J Эндокринол. 2014 г., август; 222 (2): R75–96.
- Стефенсон Р.Циркадные ритмы и расстройства дыхания, связанные со сном. Сон Мед. 2007 г., сен; 8 (6): 681–7.
- Muller JE, Stone PH, Turi ZG, Rutherford JD, Czeisler CA, Parker C, et al. Суточные колебания частоты возникновения острого инфаркта миокарда. N Engl J Med. 1985 ноябрь; 313 (21): 1315–22.
- Фрой О. Метаболизм и циркадные ритмы – последствия ожирения. Endocr Rev. 2010 Feb; 31(1):1–24.
- Крамер А., Мерроу М., редакторы.Циркадные часы. Спрингер; 2013.
- Браун С.А., Аззи А. Периферийные циркадные осцилляторы у млекопитающих; Циркадные часы. Спрингер; 2013. С. 45–66.
- Дэвидсон А.Дж., Лондон Б., Блок Г.Д., Менакер М.Сердечно-сосудистые ткани содержат независимые циркадные часы. Клин Эксперт Гипертенс. 2005 г., февраль-апрель; 27 (2–3): 307–11.
- Ла Флер С.Е., Калсбек А., Вортель Дж., Феккес М.Л., Буйс Р.М. Суточный ритм толерантности к глюкозе: роль супрахиазматического ядра. Сахарный диабет. 2001 г., июнь; 50 (6): 1237–43.
- Харада Н., Инагаки Н. Роль часовых генов в секреции инсулина. J Исследование диабета. 2016 ноябрь; 7 (6): 822–3.
- Миджилы С., Озогул С.Диябет Кок Хюкрелер. Dokuz Eylül Üniversitesi Tıp Fakültesi Dergisi. 2007; 21:109–17.
- Созлю С., Шанлиер Н. Сиркадийен Ритим, Саглик и Бесленме Илишкиси. Журнал медицинских наук Турции Klinikleri. 2017;2:100–9.
- Кесслер К, Пивоварова О, Пфайффер АФ.[Циркадные часы и энергетический обмен: последствия для здоровья]. Dtsch Med Wochenschr. 2014 г., апрель; 139 (14): 684–6.
- Санкар Г., Бруннер М. Циркадные часы и энергетический обмен. Cell Mol Life Sci. 2014 г., июль; 71 (14): 2667–80.
- Масри С.Сиртуин-зависимый контроль часов: новые достижения в области метаболизма, старения и рака. Curr Opin Clin Nutr Metab Care. 2015 ноябрь; 18 (6): 521–7.
- Yang X, Downes M, Yu RT, Bookout AL, He W, Straume M, et al. Экспрессия ядерных рецепторов связывает циркадные часы с метаболизмом. Клетка. 2006 г., август; 126 (4): 801–10.
- Фрой О., Мискин Р. Взаимосвязь питания, циркадных ритмов и старения. Прог Нейробиол. 2007 г., июнь; 82 (3): 142–50.
- Байрам А, Мехри И.Sirtuin Genleri ve İşlevleri. Фират Тип Дерг. 2013;18:136–40.
- Раттер Дж., Рейк М., Ву Л.С., Макнайт С.Л. Регуляция связывания ДНК часов и NPAS2 окислительно-восстановительным состоянием кофакторов НАД. Наука. 2001 г., июль; 293 (5529): 510–4.
- Ум Дж. Х., Ян С., Ямадзаки С., Кан Х., Виоллет Б., Форетц М. и др.Активация 5′-АМФ-активируемой киназы лекарством от диабета метформином индуцирует зависимую от казеинкиназы Iepsilon (CKIepsilon) деградацию часового белка mPer2. Дж. Биол. Хим. 2007 г., июль; 282 (29): 20794–8.
- Кумар Джха П., Чалле Э., Калсбек А. Суточные ритмы метаболизма глюкозы и липидов у ночных и дневных млекопитающих.Мол Селл Эндокринол. 2015, декабрь; 418 (часть 1): 74–88.
- Бенедикт ФГ. Факторы, влияющие на основной обмен. Proc Natl Acad Sci USA. 1915 г., февраль; 1 (2): 105–9.
- Хауген Х.А., Мелансон Э.Л., Тран З.В., Кирни Дж.Т., Хилл Дж.О.Вариабельность измеренной скорости метаболизма в состоянии покоя. Am J Clin Nutr. 2003 г., декабрь; 78 (6): 1141–5.
- Лапоски А.Д., Басс Дж., Косака А., Турек Ф.В. Сон и циркадные ритмы: ключевые компоненты регуляции энергетического обмена. ФЭБС лат. 2008 г., январь; 582 (1): 142–51.
- Алгин Ди, Акдаг Г, Эрдинч ОО.Калители уйку ве уйку бозуклюклари / Качественный сон и нарушения сна. Медицинский журнал Османгази. 2016;38.
- Шахин Л., Ащоглу М., Ташкин Э. Уйку ве уйкунун düzenlenmesi. Саглык Билимлери Дергиси. 2013;22:93–98.
- Босколо Р.А., Эстевес А.М., де Сантана М.Г., Виана В.А.Р., Грассманн В., Туфик С., де Мелло М.Т.Существует ли связь между составом тела, уровнем основного обмена и сном у пожилых пациентов с синдромом обструктивного апноэ во сне и без него? Наука сна. 2013;6(4):129–134.
- Кнутсон К.Л., Шпигель К., Пенев П., Ван Каутер Э. Метаболические последствия лишения сна.Sleep Med Rev. 2007 Jun; 11 (3): 163–78.
- Chtorou H, Souissi N. Эффект от тренировок в определенное время дня: обзор. J Прочность Конд Рез. 2012 г., июль; 26 (7): 1984–2005 гг.
- Суисси Н., Готье А., Сесбуэ Б., Ларю Дж., Давенн Д.Циркадные ритмы при двух видах анаэробных циклических упражнений на ноги: скоростно-силовом тесте и 30-секундном тесте Вингейта. Int J Sports Med. 2004 г., янв; 25 (1): 14–9.
- Kinişler A. Anaerobik Performansta sirkadiyen değişimlerin incelenmesi. Спорт Билимлери Дергиси. 2005; 16: 174–84.
- Краучи К.Как регулируется циркадный ритм внутренней температуры тела? Клин Автон Рез. 2002 г., июнь; 12 (3): 147–9.
- Райт К.П. младший, Халл Дж. Т., Чейслер К.А. Взаимосвязь между настороженностью, производительностью и температурой тела у людей. Am J Physiol Regul Integr Comp Physiol. 2002 г., декабрь; 283 (6): R1370–7.
- Беннард П., Дусе Э. Острое влияние времени тренировки и гликемического индекса завтрака на окисление жира, вызванное физическими упражнениями. Appl Physiol Nutr Metab. 2006 г., 31 октября (5): 502–11.
- Катсанос К.С., Моффат Р.Дж.Острые эффекты упражнений до еды по сравнению с упражнениями после еды на постпрандиальную гипертриглицеридемию. Клин Джей Спорт Мед. 2004 г., янв; 14 (1): 33–9.
- Фарах Н.М., Гилл Дж.М. Влияние физических упражнений до или после приема пищи на жировой баланс и постпрандиальный метаболизм у мужчин с избыточным весом. Бр Дж Нутр.2013 г., июнь; 109 (12): 2297–307.
- Канг Дж., Рейнс Э., Розенберг Дж., Ратамесс Н., Наклерио Ф., Файгенбаум А. Метаболические реакции во время постпрандиальных упражнений. Рес Спорт Мед. 2013;21(3):240–52.
- Шибата С., Тахара Ю.Циркадный ритм и физические упражнения. J Phys Fit Sports Med. 2014;3(1):65–72.
- Паоли А., Марколин Г., Зонин Ф., Нери М., Сивьери А., Пачелли К.Ф. Упражнения натощак или кормление для усиления потери жира? Влияние приема пищи на коэффициент дыхания и избыточное потребление кислорода после тренировки на выносливость.Int J Sport Nutr Exerc Metab. 2011 февраль; 21 (1): 48–54.
- Таппи Л. Термический эффект пищи и активность симпатической нервной системы у человека. Репрод Нутр Дев. 1996;36(4):391–7.
- Ромон М., Эдме Дж.Л., Буленгес С., Лескроарт Дж.Л., Фримат П.Циркадные вариации термогенеза, вызванного диетой. Am J Clin Nutr. 1993 г., апрель; 57 (4): 476–80.
- Моррис С.Дж., Гарсия Д.И., Майерс С., Ян Д.Н., Триенекенс Н., Шеер Ф.А. Циркадная система человека играет доминирующую роль в возникновении разницы между утром и вечером в термогенезе, вызванном диетой.Ожирение (Серебряная весна). 2015 окт; 23 (10): 2053–2058.
- Grammaticos PC, Doumas A, Koliakos G. Полупериод опорожнения желудка утром и ночью отличался более чем на 220% у двух молодых здоровых взрослых. Ад Джей Нукл Мед. 2015 янв.-апрель;18(1):60–2.
- Хугерверф В.А.Роль часовых генов в моторике желудочно-кишечного тракта. Am J Physiol Gastrointest Liver Physiol. 2010 г., сен; 299 (3): G549–55.
- Garaulet M, Gómez-Abellán P. Время приема пищи и ожирение: новая ассоциация. Физиол Поведение. 2014 июль; 134: 44–50.
- Воллмерс С., Гилл С., Ди Таккио Л., Пуливарти С.Р., Ле Х.Д., Панда С.Время кормления и внутренние циркадные ритмы привода часов в экспрессии печеночных генов. Proc Natl Acad Sci U S A. 2009 Dec 15;106(50):21453–8.
- Косака А., Лапоски А.Д., Рэмси К.М., Эстрада С., Джошу С., Кобаяши Ю. и др. Диета с высоким содержанием жиров нарушает поведенческие и молекулярные циркадные ритмы у мышей.Клеточный метаб. 2007 ноябрь; 6 (5): 414–21.
- Тахара Ю., Аояма С., Шибата С. Циркадные часы млекопитающих и их увлечение стрессом и физическими упражнениями. J Physiol Sci. 2017 Январь; 67 (1): 1–10.
- Дамиола Ф., Ле Минь Н., Прейтнер Н., Корнманн Б., Флери-Олела Ф., Шиблер Ю.Ограниченное питание разъединяет циркадные осцилляторы в периферических тканях и центральный водитель ритма в супрахиазматическом ядре. Гены Дев. 2000 г., декабрь; 14 (23): 2950–61.
- Браун С.А., Зумбрунн Г., Флери-Олела Ф., Прейтнер Н., Шиблер У. Ритмы температуры тела млекопитающих могут поддерживать периферические циркадные часы.Карр Биол. 2002 г.; 12 (18): 1574–83.
- Леонардо-Мендонса Р.С., Мартинес-Николас А., де Тереза Гальван С., Оканья-Вильгельми Дж., Русанова И., Герра-Эрнандес Э. и др. Влияние четырехнедельного лечения мелатонином на циркадные ритмы у спортсменов, тренирующихся с отягощениями. Хронобиол Инт.2015;32(8):1125–34.
- Вольф Г., Эссер К.А. Запланированная фаза упражнений сдвигает циркадные часы в скелетных мышцах. Медицинские спортивные упражнения. 2012 г., сен; 44 (9): 1663–70.
- Гарулет М., Ордовас Дж. М., Мадрид Дж. А.Хронобиология, этиология и патофизиология ожирения. Инт Дж. Обес. 2010 г., декабрь; 34 (12): 1667–83.
- Фрой О. Циркадные ритмы и ожирение у млекопитающих. ISRN Обес. 2012 18 ноября; 2012:437198.
- Тахара Ю, Шибата С.Хронобиология и питание. Неврология. 2013 декабрь; 253: 78–88.
- Ойк Х. Модуляция циркадных часов питательными веществами и пищевыми факторами. Биоски Биотехнолог Биохим. 2017 г., май; 81 (5): 863–70.
- Накао Р.Циркадные часы скелетных мышц: текущие идеи. Хронофизиология и терапия. 2017;7:47–57.
Автор Контакты
Nilüfer Acar Tek
Факультет медицинских наук, кафедра питания и диетологии
Университет Гази, округ Эмниет, улица Муаммер Яшар Бостанджи
Номер 16, TR-06560 Анкара (Турция)
Электронная почта acarnil@hotmail.ком
Информация о статье / публикации
Предварительный просмотр первой страницы
Получено: 26 июня 2018 г.
Принято: 30 марта 2019 г.
Опубликовано онлайн: 23 апреля 2019 г.
Дата выпуска выпуска: июнь 2019 г.
Количество печатных страниц: 9
Количество фигурок: 1
Количество столов: 1
ISSN: 0250-6807 (печать)
eISSN: 1421-9697 (онлайн)
Для получения дополнительной информации: https://www.karger.com/ANM
Авторское право / Дозировка препарата / Отказ от ответственности
Авторское право: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме и любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование или любую систему хранения и поиска информации, без письменного разрешения издателя. .
Дозировка препарата: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор препарата и дозировка, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Тем не менее, в связи с продолжающимися исследованиями, изменениями в правительственных постановлениях и постоянным потоком информации, касающейся лекарственной терапии и реакций на лекарства, читателю настоятельно рекомендуется проверять вкладыш в упаковке для каждого лекарства на предмет любых изменений в показаниях и дозировке, а также для дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендуемый агент является новым и/или редко используемым лекарственным средством.
Отказ от ответственности: заявления, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и участникам, а не издателям и редакторам.Появление рекламы и/или ссылок на продукты в публикации не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор(ы) отказываются от ответственности за любой ущерб, нанесенный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в содержании или рекламе.