
Как и почему мышцы сокращаются? Новая модель мышечного сокращения «Поворот и замок» ≪ Scisne?
 Рис. 1. Классическая картинка, показывающая структуру мышцы. Изображение с сайта ru.wikipedia.org Рис. 1. Классическая картинка, показывающая структуру мышцы. Изображение с сайта ru.wikipedia.org |
Почти всякая незыблемая общепринятая теория, которую с проклятьями зубрят школьники и которую устало и одинаково рассказывают учителя и даже профессора ВУЗов, при внимательном рассмотрении оказывается отнюдь не однозначной, захватывающей и полной загадок. К теории мышечного сокращения вышесказанное относится в полной мере. В общих чертах она была разработана еще в 50-х годах прошлого века, и классический рисунок (рис. 1) с актиновыми и миозиновыми нитями до сих пор кочует из учебника в учебник. Однако реальная картина сокращения мышцы куда запутаннее, интереснее и непонятнее, со множеством подробностей и неожиданных действующих лиц и со сложными ролями, которые исполняют эти лица. О новой и удивительной отрасли науки, находящейся на стыке физики, математики и биологии и изучающей механизмы мышечного сокращения, рассказали в своих лекциях на проходящей при поддержке РВК, Фонда «Династия» и РФФИ Зимней школе Future Biotech доктор физико-математических наук Андрей Кимович Цатурян и доктор биологических наук, заведующий Лабораторией биологической подвижности Института иммунологии и физиологии УрО РАН Сергей Юрьевич Бершицкий.
Азбучные истины
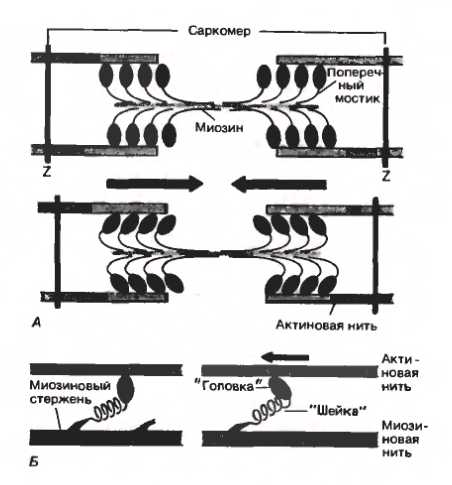
Начнем с азов — собственно, с классической теории мышечного сокращения. Базовая сократительная единица мышечной ткани называется саркомером. Края саркомера — Z-диски — состоят из переплетающихся нитей различных белков. К одному из этих белков цепляются актиновые микрофиламенты, вдоль которых тянутся регуляторные белки тропонин и тропомиозин (рис. 2). Другой белок — титин, самый большой из известных в настоящее время белков, — крепится к соседнему участку Z-диска и служит длинной-длинной основой, с которой связываются молекулы белка миозина. Таким образом, саркомер состоит из чередующихся тонких (образованных многочисленными молекулами актина и регуляторными белками) и толстых (состоящих из тоже многочисленных молекул миозина и вспомогательных белков) нитей.
 Рис. 2. Актиновые микрофиламенты обмотаны регуляторными белками — тропомиозином и прикрепленным к нему тропонином, состоящим из трех различных субъединиц. Когда в мышечной клетке нет ионов кальция, тропомиозин загораживает миозину сайт связывания с актином. Однако когда ионы кальция есть, они присоединяются к тропонину, который меняет свою конформацию и перестает удерживать тропомиозин на актиновых нитях. В результате актиновые сайты оказываются открыты для связывания, миозиновые головки могут прицепиться к ним, и мышечное сокращение становится возможным. Изображение с сайта www.med4you.at |
И вот начинается кое-что интересное. Импульс, подошедший к нервно-мышечному соединению, вызывает повышение внутриклеточного уровня кальция. Кальций присоединяется к регуляторным белкам, которые обматывали актин и загораживали его от миозина, в результате чего эти белки смещаются, и головка миозина, содержащая продукты гидролиза АТФ, приникает к молекуле актина. В результате различных пертурбаций (которые подробно описаны ниже), миозин крепко сцепляется с актином и меняет свою конформацию, поворачивая хвост относительно головки и выплевывая продукты гидролиза. Это происходит на множестве миозиновых головок и приводит к тому, что актиновая нить чуть-чуть сдвигается относительно миозиновой. Затем крепко сцепленный с актиновой нитью миозин связывается с АТФ, отцепляется от актина и претерпевает обратные конформационные изменения — то есть отворачивает хвост обратно (рис. 3).
 Рис. 3. Схема мышечного сокращения (классический цикл Лимна–Тейлора) |
Так, перебирая головками, миозиновые молекулы и обеспечивают работу мышцы. Расслабление же мышц происходит тогда, когда к мышечной клетке перестал подходить импульс и в нее перестал поступать кальций. Тогда отцепившиеся друг от друга актиновые и миозиновые нити постепенно возвращаются в свое первоначальное положение (отчасти благодаря эластичным свойствам молекул титина), и мышца расслабляется.
 Рис. 4. Устрашающая схема, показывающая, какое астрономическое существует количество миозиновых моторов. Изображение с сайта www.motorprotein.de |
Вообще, способность миозина двигаться вдоль актиновой нити — это слишком удобное свойство, чтобы использовать его только для мышечного сокращения. Поэтому множество различных видов миозина (их еще называют «миозиновыми моторами», их филогенетическое дерево показано на рис. 4) применяется разными видами клеток для множества разнообразных функций — помимо собственно сокращения мышц они могут обеспечивать внутриклеточный транспорт, двигать трансмембранные белки и так далее (рис. 5).
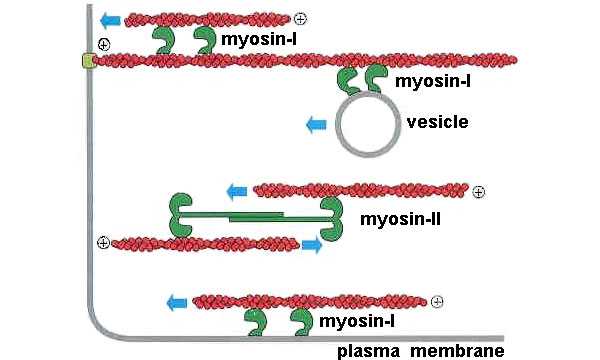 Рис. 5. Вот примеры функций, которые могут выполнять в клетке миозины различных видов. Они могут тянуть вдоль актиновой нити какой-либо груз (в данном случае везикулу), могут сдвигать актиновую нить относительно другой актиновой нити либо относительно мембраны и могут, наконец, обеспечивать мышечное сокращение. Изображение из слайдов к лекции А. К. Цатуряна на Зимней школе |
Различные миозины сильно отличаются друг от друга по строению (рис. 6): они могут быть одноголовыми или двухголовыми, с длинными или короткими хвостами; однако главная функциональная часть — головка — имеет практически одинаковое строение у всех видов миозина. То есть принцип работы миозина одинаков во всех случаях, а детали (например, размер хвоста) обеспечивают ту или иную специализацию.
 Рис. 6. Вот какие разные бывают миозины. Изображение из слайдов к лекции А. К. Цатуряна на Зимней школе |
Миозины — это не единственные моторные белки. Помимо них существует еще два класса моторов — динеины и кинезины. В отличие от миозинов, которые двигаются по актиновой нити, динеины и кинезины бегают по микротрубочкам, причем динеины — только в одну сторону, а кинезины — только в противоположную.
Гипотеза рычага
Теперь пришло время подробней разобраться, что же происходит с миозином при мышечном сокращении. Начнем с общепринятой в настоящее время теории, известной под названием «Гипотеза рычага».
Посмотрим внимательнее на молекулу миозина (а конкретнее — самого удобного для исследований миозина II, рис. 7). Понятно, что в головке миозина должно быть как минимум два важных места — одно, хватающееся за актин, и второе, в которое залезает АТФ. Исследователи, работающие с миозином, остроумно назвали «актиновый» участок головки «пастью», а «АТФный» — «карманом». И пертурбации, происходящие с миозином, можно описать довольно грубым выражением: «закрой пасть и держи карман шире».
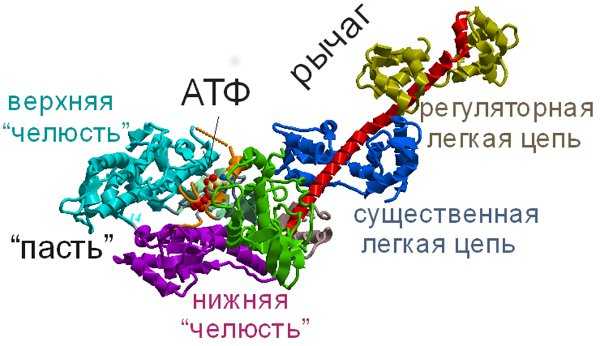 Рис. 7. Структура миозина. Легкие цепи вокруг хвоста выполняют регуляторную роль в гладких мышцах и в немышечных миозинах. Изображение из слайдов к лекции А. К. Цатуряна на Зимней школе |
Дело в том, что, чтобы АТФный карман открылся и в него мог попасть АТФ, актиновая пасть должна быть закрыта (то есть миозин должен сидеть на актине): закрытая верхняя челюсть пасти оттягивает створку кармана, и тот открывается. АТФ влезает в широко раскрытый карман.
И вот тут начинается самое интересное. Гидролиз АТФ может происходить только в закрытом кармане, а для того, чтобы карман закрылся, должна открыться пасть — то есть миозин должен отвалиться от актина. Но это еще не всё. Чтобы обеспечить гидролиз, окрестности кармана должны немного перестроиться, сдвинуться. Сдвигаясь, околокарманные участки вызывают небольшие изменения соседних областей, которые, в свою очередь, приводят к тому, что жесткий домен миозина под названием «конвертер» перебрасывается из одного устойчивого положения в другое и тянет за собой миозиновый хвост, отклоняя его на целых 60°. Курок взводится.
Теперь начинается следующий акт. Миозин с карманом, набитым АДФ и фосфатом, должен обязательно прильнуть к актину и закрыть пасть, потому что иначе выплюнуть фосфат он не в состоянии (то есть чисто теоретически он его когда-нибудь выплюнет, но очень нескоро; поэтому миозин — это, по сути, актин-зависимая АТФаза). Миозин сначала слабо связывается с актином при помощи электростатических взаимодействий, а затем запускается процесс закрытия пасти. Происходит это так. В результате конформационных изменений миозиновая головка разворачивается к актиновой нити таким образом, что, во-первых, образует контакт, очень большой по площади (больше 18 нм
Итак, миозин выплевывает фосфат и, крепко-накрепко вцепившись в актин, претерпевает обратные конформационные изменения — хвост его «выстреливает», сдвигается относительно головки. Это происходит сразу на множестве молекул миозина и потому приводит к движению актиновой нити относительно миозиновой, а следовательно — и к сокращению мышцы. После этого АДФ выбрасывается из кармана. Миозин остается сцеплен с актином; пасть его закрыта — даже заперта! — крепко-накрепко, и если клетка мертва и все АТФ в ней уже закончились, то на этом грустном моменте история заканчивается, миозин и актин так и остаются в навсегда сцепившемся, застывшем состоянии, а у организма начинается трупное окоченение. Более оптимистичный сценарий, характерный для живой клетки с солидным запасом АТФ, предполагает, что в карман (который, как мы помним, головка с закрытой пастью держит шире) влезает новая молекула АТФ, пасть открывается, миозин отлипает от актина и цикл повторяется заново.
Roll and lock: поворачиваем и запираем
По гипотезе рычага, в мышечном сокращении существует только один момент генерации силы — когда поворачивается миозиновый хвост (рычаг). Однако некоторые данные рентгенографии и томографии мышц не то чтобы не согласуются с этой теорией, а свидетельствуют о том, что существует еще какой-то непонятный момент в сокращении мышцы, который гипотеза рычага не объясняет. Поэтому группа исследователей под руководством А. К. Цатуряна предложила теорию мышечного сокращения под названием «Roll and lock» — «Поворачиваем и запираем» (см.: Michael A. Ferenczi et al., 2005. The «Roll and Lock» Mechanism of Force Generation in Muscle). По этой теории, миозиновые головки садятся на актин еще до гидролиза АТФ, причем садятся не стройно и организованно, а как попало. На головке миозина есть длинный выступающий домен — «щуп», — который «нащупывает» подходящую себе (кислую и отрицательно заряженную) часть актиновой нити и прилипает к ней — как придется, под первым попавшимся углом. Однако стоит произойти гидролизу АТФ, как миозин меняет свою конформацию, головки поворачиваются под нужным углом и крепко и четко, как ключ с замком, сцепляются с актиновой нитью, а из миозинового кармана выбрасывается фосфат. И вот только после этого происходит поворот рычага. Иными словами, модель получается двухстадийной — на первом этапе головка миозина крепко и четко вцепляется в актин и при этом немного поворачивается, а на втором — поворачивается рычаг, причем сила, которая потом приведет к движению мышцы, генерируется на обоих этих этапах.
Помимо рентгеноструктурных и томографических данных, которые очень хорошо согласуются с теорией «Roll and lock», существует и несколько косвенных, но очень красивых доказательств ее правоты. Например, известно, что во время мышечного сокращения, в том случае, если мышца не меняет свою длину, всего чуть более 40% миозиновых головок сидит на актине, а остальные болтаются ни к чему не присоединенными. Однако когда сжатую мышцу насильно растягивают (например, такое бывает при беге, когда человек приземляется на напряженную мышцу), то жесткость мышцы резко увеличивается из-за того, что почти все свободные миозиновые головки резко сцепляются с актиновой нитью. Однако, судя по рентгеноструктурным данным, сцепляются они отнюдь не «намертво», как ключ с замком, а просто как попало. Объяснить это можно как раз с помощью теории «Roll and lock». Гидролиз АТФ при растяжении мышцы прекращается (оно и понятно: какой смысл тратить АТФ, если работа совершается не мышцей, а над мышцей), и все миозиновые головки переходят в состояние «активного актинового поиска» — их торчащий щуп ищет актиновую нить, нащупывает на ней подходящее место и сцепляется с ним — не крепко-накрепко, не как ключ с замком, а как попало. Однако для того, чтобы увеличить жесткость мышцы (и этим защитить кости от перелома) этого оказывается достаточно.
Итог
Гипотеза «Roll and lock» уточняет гипотезу рычага. Она лучше согласуется с экспериментальными данными и описывает мышечное сокращение более подробно. Но совершенно точно можно сказать, что и эта теория может быть уточнена и расширена — однако каким именно образом, мы пока еще не знаем. Мышечное сокращение, которое интенсивно и кропотливо исследуется уже многие годы, во многом по-прежнему остается неразгаданной загадкой.
Видеоиллюстрации:
Видеоиллюстрация гипотезы рычагаМиозин цепляется к актиновой нити, вцепляется в нее, поворачивает хвост, затем отцепляется и возвращает хвост в прежнее положение. Видео Кеннета Холмса. |
Видеоиллюстрация гипотезы «Roll and lock»Миозин вначале ищет подходящее положение на актиновой нити, затем слабо соединяется с ней, потом сцепляется крепко — при этом вся головка поворачивается и развивает некоторое усилие, и только затем поворачивается хвост. Видео Мэри Риди. |
Источники:
1) Лекции А. К. Цатуряна и С. Ю. Бершицкого на Зимней школе FutureBiotech.
2) Н. А. Кубасова, А. К. Цатурян. Молекулярный механизм работы актин-миозинового мотора в мышце // Успехи биологической химии. 2011. Т. 51. С. 233–282 — относительно свежий, очень подробный и хорошо написанный русскоязычный обзор по теме, находится в свободном доступе.
3) К. Бэгшоу. Мышечное сокращение // М.: Мир. 1985.
4) Н. Б. Гусев. Молекулярные механизмы мышечного сокращения // Соросовский образовательный журнал, т. 6, №8, 2000. С. 24–32.
5) А. Н. Тихонов. Молекулярные моторы. Часть 2. Молекулярные основы биологической подвижности // Соросовский образовательный журнал, с. 18–24, 1999.
Вера Башмакова
«Элементы»
scisne.net
Сокращение мышц. Принцип работы мышцы человека.
О том, как устроена мышечная клетка и что представляет из себя мышца, Вы уже имеете понятие. Но, как же осуществляется сокращение мышцы? Что заставляет наши мышцы работать?
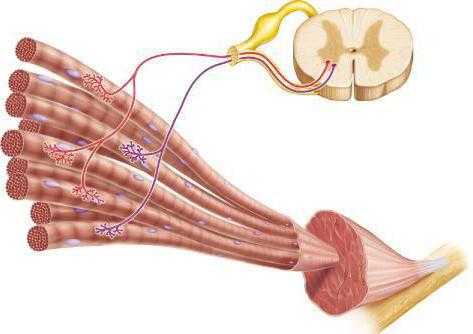
Говоря доступным языком, сокращение мышц происходит под воздействием нервных импульсов, которые активируют нервные клетки спинного мозга – мотонейроны, ответвления которых — аксоны подведены к мышце. Если разобраться подробнее, то внутри мышцы аксон разделяется и образует сеть ответвлений, которые, подобно электрическим контактам, «подсоединены» к мышечной клетке. Посредством таких контактов и осуществляется сокращение мышц.

Получается, что каждый мотонейрон управляет группой мышечных клеток. Такие группы получили название – нейромоторные единицы, благодаря которым человек может задействовать в работе часть мышцы. Поэтому, мы можем сознательно контролировать скорость и силу сокращения мышц.
Итак, мы рассмотрели процесс «запуска» сокращения мышц. Теперь давайте детально разберемся, что же происходит непосредственно внутри мышцы во время сокращения. Этот материал несколько сложен для восприятия, но весьма важен. Вам необходимо разобраться в нем, иначе Вы не сможете до конца уяснить, каким образом растут наши мышцы.
Сокращение мышц в грубом приближении
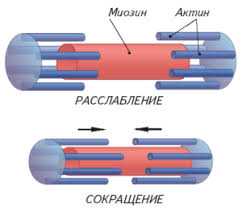
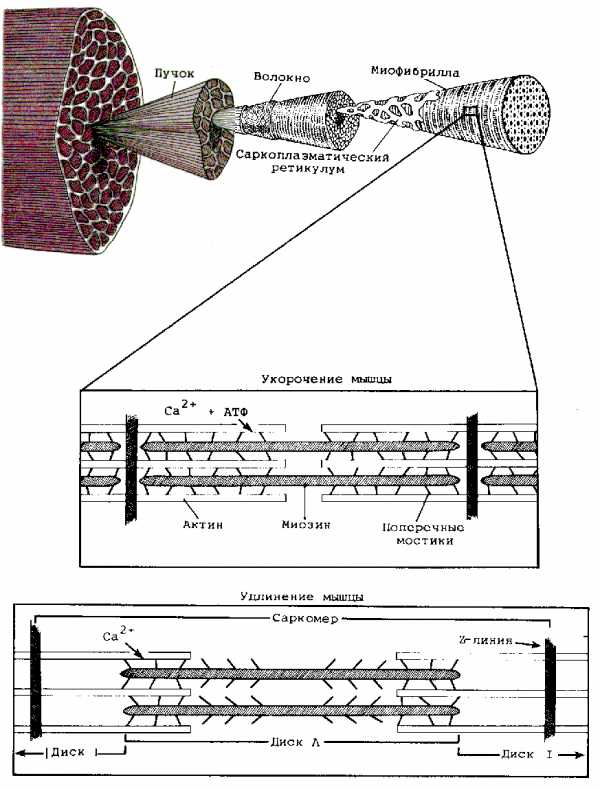
В первую очередь необходимо уяснить, что миофибрилла состоит из многочисленных нитей двух белков: миозина и актина, которые располагаются вдоль миофибриллы. Причем, миозин – толстые нити, а актин – тонкие нити. Этим и объясняется светло-темное полосатое строение миофибриллы (темные полосы – миозин, светлые полосы – актин).
В литературе темные участки миофибриллы получили название А-диск, а светлые участки именуются I-диск. Актиновые нити крепятся к так называемой Z-линии, которая расположена в центре I-диска. Сегмент миофибриллы между Z-линииями, включающий миозиновый А-диск называется саркомером, который можно считать некой сократительной единицей миофибриллы.
Саркомер сокращается следующим образом: при помощи боковых ответвлений (мостиков) толстые нити миозина втягивают вдоль себя тонкие нити актина.
То есть головки мостиков входят в зацепление с актиновой нитью и втягивают ее между нитями миозина. По окончанию движения головки отсоединяются и входят в новое зацепление, продолжая втягивание. Получается, что сокращение мышц – совокупность сокращений множества саркомеров.
Если рассмотреть отдельно тонкую нить актина, то она представляет собой двойную спираль актиновых нитей, между которыми расположена двойная цепь тропомиозина.
Тропомиозин – это также белок, который блокирует зацепления миозиновых мостиков с актином в расслабленном состоянии мышцы. Как только нервный импульс через мотонейрон подается в мышцу, происходит смена полярности заряда мембраны мышечной клетки, в результате чего саркоплазма клетки насыщается ионами кальция (Ca++), которые высвобождаются из специальных хранилищ, находящихся вдоль каждой миофибриллы. Тропомиозиновая нить, в присутствии ионов кальция, мгновенно углубляется между актиновыми нитями, и мостики миозина получают возможность зацепления с актином – сокращение мышц становится возможным.
Однако после поступления Са++ в клетку, он тут же возвращается в свои хранилища и происходит расслабление мышцы. Только при постоянных импульсах, исходящих от нервной системы, мы можем поддерживать длительное сокращение – это состояние получило определение тетаническое сокращение мышц.
Разумеется, сокращение мышц требует энергии. А откуда же она берется, как формируется энергия, поддерживающая движение миозинового мостика? Об этом Вы узнаете в следующей статье Энергетические процессы в мышечной клетке. Энергия сокращения мышц.
© Твой Тренинг
Материалы данной статьи охраняются законом о защите авторских прав. Копирование без указания ссылки на первоисточник и уведомления автора ЗАПРЕЩЕНО!
tvoytrening.ru
Механизм мышечного сокращения — SportWiki энциклопедия
Нервно-мышечная реакция на силовую тренировку[править | править код]
Структура мышц[править | править код]
Мышца — это комплексная структура, отвечающая за движение. Мышцы состоят из саркомеров, которые содержат определенное сочетание фибриллярных белков — миозина (толстые нити) и актина (тонкие нити), которые играют важную роль в мышечных сокращениях. Таким образом, саркомер — это сократительный элемент мышечного волокна, состоящий из миозиновых и актиновых белковых нитей.
Помимо этого, способность мышцы сокращаться и прилагать силу зависит конкретно от ее вида, площади поперечного сечения, а также длины и количества волокон внутри мышцы. Число волокон определяется генетикой, и на него невозможно повлиять с помощью тренировок; однако тренировки в состоянии изменить другие переменные. Например, число и толщина миозиновых нитей увеличивается посредством упорных тренировок с максимальной силовой нагрузкой. Увеличение толщины мышечных нитей увеличивает размер мышцы и силу сокращений.
Человеческое тело состоит из различных типов мышечных волокон, подразделяющихся на группы, и каждая группа относится к одной двигательной единице. В общем и целом в нашем организме имеются тысячи двигательных единиц, в которых находятся десятки тысяч мышечных волокон. Каждая двигательная единица содержит сотни или тысячи мышечных волокон, пребывающих в покое до тех пор, пока им не нужно действовать. Двигательная единица управляет совокупностью волокон и направляет их действия по закону «все или ничего». Этот закон означает, что при раздражении двигательной единицы импульс, направляемый в ее мышечные волокна, либо распространяется полностью — таким образом раздражая всю совокупность волокон, — либо не распространяется вообще.
Разные двигательные единицы реагируют на разные нагрузки при тренировках. Например, выполнение жима лежа с 60% повторного максимума задействует определенную совокупность двигательных единиц, тогда как более крупные двигательные единицы ожидают более высокой нагрузки. Поскольку последовательное задействование двигательных единиц зависит от нагрузки, необходимо разрабатывать специальные программы, чтобы активизировать и адаптировать основные группы двигательных единиц и мышечных волокон, играющих доминирующую роль в избранном виде спорта. К примеру, в тренировках для спринта на короткую дистанцию и легкоатлетических дисциплин (таких как толкание ядра) следует использовать тяжелые нагрузки, чтобы способствовать развитию силы, необходимой для оптимизации скорости и взрывных действий.
Мышечные волокна выполняют разные биохимические (метаболические) функции; выражаясь конкретнее, одни лучше приспособлены с физиологической точки зрения к работе в анаэробных условиях, а другие лучше работают в аэробных условиях. Волокна, которые используют кислород для выработки энергии, называются аэробными, тип I, красными или медленными. Волокна, которым кислород не требуется, называются анаэробными, тип II, белыми или быстрыми. Быстрые мышечные волокна, в свою очередь, делятся на подтипы IIА и IIХ (иногда называемые IIВ, хотя у людей тип IIВ практически не встречается[1]).
Медленные и быстрые волокна существуют примерно в равной пропорции. Однако в зависимости от их функций, в некоторых группах мышц (например, подколенные сухожилия, бицепсы) содержится больше быстрых волокон, тогда как в других (например, в камбаловидной мышце) содержится больше медленных волокон. В таблице 2.1 мы сравниваем характеристики быстрых и медленных волокон.
Сравнение быстрых и медленных волокон
МЕДЛЕННЫЕ ВОЛОКНА | БЫСТРЫЕ ВОЛОКНА |
Красные, тип I, аэробные | Белые, тип II, анаэробные |
• Медленно устают • Нервная клетка меньше — иннервирует от 10 до 180 мышечных волокон • Развивают долгие, продолжительные сокращения • Применяются для развития выносливости • Активизируются во время низко- и высокоинтенсивной деятельности | • Быстро устают • Большая нервная клетка — иннервирует от 300 до 500 (или более) мышечных волокон • Развивают короткие, сильные сокращения • Применяются для развития скорости и силы • Активизируются только во время высокоинтенсивной деятельности |
Тренировки могут влиять на эти характеристики. Датские ученые Андерсен и Аагаард[2][3][4][5][6] в своих исследованиях показывают, что при объемных нагрузках или лактатных по природе тренировках волокна IIХ приобретают характеристики волокон IIА. То есть богатая миозином цепочка этих волокон становится более медленной и более эффективно справляется с лактатной деятельностью. Эти изменения можно повернуть вспять, снижая тренировочную нагрузку (тейперинг), в результате чего волокна IIХ возвращаются к изначальным характеристикам наиболее быстрых волокон[3]. Силовые тренировки также увеличивают размер волокон, благодаря чему вырабатывается больше силы.
Сокращение быстрой двигательной единицы более быстрое и мощное, чем сокращение медленной двигательной единицы. В результате пропорция быстрых волокон, как правило, выше в организме успешных спортсменов, занимающихся скоростно-силовыми видами спорта, но они также быстрее утомляются. Спортсмены с более высоким скоплением медленных волокон, напротив, обычно преуспевают в видах спорта на выносливость, поскольку они могут выполнять нагрузки низкой интенсивности в течение более продолжительного времени.
Активизация мышечных волокон происходит по принципу величины, известному также как принцип Хеннемана[7], согласно которому двигательные единицы и мышечные волокна активизируются начиная с меньшей в сторону большей. Активация всегда начинается с медленных волокон. При низкой или умеренно интенсивной нагрузке активируются медленные волокна и выполняют большую часть работы. При сильной нагрузке сначала сокращаются медленные волокна, затем в процесс вовлекаются быстрые волокна. При повторениях до отказа с умеренной нагрузкой двигательные единицы, состоящие из быстрых волокон, постепенно активизируются, чтобы поддерживать выработку силы, тогда как ранее задействованные двигательные единицы утомляются (см. рис. 1).
 рис. 1. Последовательная активизация двигательных единиц в подходе упражнений до концентрического отказа
рис. 1. Последовательная активизация двигательных единиц в подходе упражнений до концентрического отказаВ распределении типов мышечных волокон у спортсменов, занимающихся разными видами спорта, могут наблюдаться различия. Это иллюстрируют рис. 2 и 2.3, представляющие общий процент содержания быстрых и медленных мышечных волокон у спортсменов в избранных видах спорта. Например, существенная разница между спринтерами и марафонцами четко дает понять, что успех в некоторых видах спорта хотя бы частично определяется генетическим составом мышечных волокон спортсмена.
 рис. 2. Распределение типов волокон у мужчин в разных видах спорта. Обратите внимание на преобладание медленных волокон у спортсменов, занимающихся аэробными видами спорта, и на преобладание быстрых волокон у спортсменов, занимающихся скоростно-силовыми видами спорта
рис. 2. Распределение типов волокон у мужчин в разных видах спорта. Обратите внимание на преобладание медленных волокон у спортсменов, занимающихся аэробными видами спорта, и на преобладание быстрых волокон у спортсменов, занимающихся скоростно-силовыми видами спортаСледовательно, пиковая мощность, вырабатываемая спортсменами, также имеет отношение к распределению типов волокон — чем выше процент быстрых волокон, тем большую мощность развивает спортсмен. Процент быстрых волокон также имеет отношение к скорости: чем выше скорость спортсмена, тем выше процент имеющихся у него быстрых волокон. Из таких людей получаются превосходные спринтеры и прыгуны, а подобный природный талант следует направлять в русло скоростно-силовых видов спорта. Попытка тренировать их, скажем, для бега на дистанцию означает трату таланта; в таких дисциплинах их ждет лишь средний успех, тогда как из них могут выйти отличные спринтеры, бейсболисты или футболисты (на этом список скоростносиловых видов спорта не кончается).
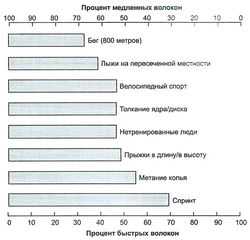 рис. 3. Распределение типов волокон у женщин в разных видах спорта
рис. 3. Распределение типов волокон у женщин в разных видах спортаКак мы описывали раньше, мышечные сокращения происходят в результате цепочки событий с участием белковых нитей — миозина и актина. В миозиновых нитях содержатся поперечные мостики — крошечные перемычки, выступающие вбок по направлению к актиновым нитям. Возбуждение, приводящее к сокращениям, стимулирует все волокно, создавая химические изменения, позволяющие актиновым нитям соединяться с миозиновыми поперечными мостиками. Связывание миозина с актином посредством поперечных мостиков высвобождает энергию, из-за чего поперечные мостики поворачиваются, таким образом подтягивая или совершая скользящее движение, связывающее миозиновые нити с актиновыми. Это скользящее движение вызывает мышечное сокращение, которое вырабатывает силу.
Чтобы визуализировать это иначе, вообразите гребную лодку. Весла представляют собой миозиновые нити, а воды — актиновые. Когда весла ударяются о воду, лодка с силой тянется вперед — и чем больше в воде весел, чем выше физическая сила гребца, тем больше вырабатываемая сила. Увеличение количества и толщины миозиновых нитей таким же образом повышает выработку силы.
Описанная ранее теория скользящих нитей дает понять, как работают мышцы, чтобы выработать силу. Эта теория включает в себя механизмы, способствующие эффективным мышечным сокращениям. Например, освобождение запаса эластичной энергии и рефлекторная адаптация играют ключевую роль в оптимизации спортивной работоспособности, но подобная адаптация происходит только тогда, когда в процессе тренировки происходит правильная стимуляция. Например, способность спортсмена использовать запас энергии для того, чтобы прыгать выше или толкать ядро дальше, оптимизируется посредством взрывных движений, как те, которые используются в плиометрическом тренинге. Однако мышечные компоненты — как, например, эластичные компоненты (сюда входят сухожилия, мышечные волокна и поперечные мостики) — не могут осуществлять эффективную транспортировку энергии, если спортсмен не укрепляет параллельные эластичные компоненты (напр., связки) и коллагеновые структуры (обеспечивающие стабильность и предохраняющие от травм). Если телу нужно выдерживать силы и воздействия, которым спортсмен подвергается, чтобы оптимизировать эластичные качества мышц, анатомическая адаптация должна предшествовать силовому тренингу.
Рефлекс — это непроизвольное мышечное сокращение, вызванное внешним стимулом[8]. Два основных компонента контроля рефлексов — это мышечные веретена и нервносухожильное веретено. Мышечные веретена реагируют на величину и скорость мышечного растяжения[9], тогда как нервно-сухожильное веретено (которое находится в местах соединения мышечных волокон с сухожильными пучками [8]) реагирует на мышечное напряжение. Когда в мышцах развивается высокая степень напряжения или растяжения, мышечные веретена и нервно-сухожильное веретено непроизвольно расслабляют мышцу, чтобы защитить ее от повреждения и травмы.
При пресечении этих ингибиторных реакций повышается спортивная работоспособность. Единственный способ добиться этого — адаптировать организм к более высокой степени напряжения, что повышает порог активизации рефлексов. Этой адаптации можно добиться посредством силового тренинга с использованием постепенно утяжеляющейся нагрузки (до 90 процентов повторного максимума или даже выше), таким образом вынуждая нервно-мышечную систему выдерживать более высокое напряжение, постоянно задействуя большее число быстрых волокон. В быстрых волокнах вырабатывается больше белка, что способствует увеличению силы.
Все спортивные движения выполняются по двигательной модели, которая называется циклом растяжение — сокращение и характеризуется тремя основными типами сокращения: эксцентрическим (удлинение), изометрическим (статичное положение) и концентрическим (сокращение). Например, волейболист, который быстро приседает и сразу подпрыгивает, чтобы блокировать атакующий удар, выполнил весь цикл растяжение — сокращение. То же касается и спортсмена, который опускает штангу на грудь и быстро выполняет взрывное движение, вытягивая руки. Чтобы полноценно пользоваться физиологическими качествами цикла растяжение — сокращение, мышца должна быстро переходить от удлинения к сокращению[10] (Schmidtble-icher, 1992).
Мышечный потенциал оптимизируется, когда активизируются все сложные факторы, влияющие на цикл растяжение — сокращение. Их влияние можно использовать для улучшения спортивных показателей только тогда, когда нервно-мышечная система стратегически стимулируется в правильной последовательности. Именно для достижения этой цели периодизация тренировки силы основывает планирование этапов на физиологической базе выбранного вида спорта. После составления эргогенного профиля (оценки вклада энергетических систем) выбранного вида спорта нужно пошагово распланировать этапы тренировки, чтобы перенести положительную нервно-мышечную адаптацию на практические показатели деятельности человека. Таким образом, понимание прикладной человеческой физиологии и установление цели в конце каждого этапа помогают тренерам и спортсменам интегрировать физиологические принципы в конкретную спортивную тренировку.
Повторим: скелетно-мышечная система тела — это сочетание костей, прикрепляемых друг к другу с помощью связок в области суставов. Пересекающие эти суставы мышцы дают силу для движения тела. Однако скелетные мышцы не сокращаются независимо друг от друга. Движения, выполняемые вокруг сустава, производятся несколькими мышцами, каждая из которых выполняет определенную роль, как уже было упомянуто выше.
Агонисты — или синергисты — это мышцы, которые взаимодействуют друг с другом при выполнении движения. В большинстве случаев, особенно если речь идет об умелом и опытном спортсмене, мышцы-антагонисты расслабляются, облегчая движение. Поскольку взаимодействие мышц группы агонистов и антагонистов напрямую влияет на спортивные движения, неправильное взаимодействие между этими группами может привести к порывистому или скованному движению. Следовательно, гладкость мышечного сокращения можно улучшить, если сосредоточиться на расслаблении антагонистов.
По этой причине одновременное сокращение (одновременная активизация мышц-агонистов и антагонистов, чтобы стабилизировать сустав) рекомендуется только на ранних стадиях реабилитации после травмы. Здоровому же спортсмену, особенно если он занимается силовыми видами спорта, не нужно выполнять упражнения (например, на нестабильной поверхности), вызывающие одновременные сокращения. К примеру, одной из основных характеристик элитных спринтеров является очень низкая миоэлектрическая активность мышц-антагонистов в каждой фазе цикла шага[11].
Первичные мышцы в первую очередь отвечают за суставное действие, которое является частью объемного силового движения или технической способности. Например, во время флексии локтя (сгибание бицепса) первичной мышцей является двуглавая мышца, тогда как трехглавая мышца (трицепс) выступает в роли антагониста и должна быть расслаблена, чтобы обеспечить беспрепятственное действие. В дополнение к этому стабилизаторы, или фиксаторы (обычно это меньшие мышцы), сокращаются изометрически, чтобы закрепить кость так, чтобы у первичных мышц была прочная база, откуда начинать натяжение. Мышцы других конечностей также могут принимать в этом участие, выступая в роли стабилизаторов, позволяющих первичным мышцам выполнять необходимые движения. Например, когда дзюдоист тянет соперника на себя, удерживая его за дзюдоги, мышцы его спины, ног и живота сокращаются изометрически, чтобы обеспечить стабильное основание для действия локтевых сгибателей (бицепсов), плечевых разгибателей (задние дельты) и лопаточных аддукторов и депрессоров (трапециевидная мышца и широчайшая мышца спины).
Если мышцу стимулировать коротким электрическим импульсом, спустя небольшой латентный период происходит ее сокращение. Такое сокращение называется «одиночное сокращение мышцы». Одиночное мышечное сокращение длится около 10-50 мс, причем оно достигает максимальной силы через 5-30 мс.
Каждое отдельное мышечное волокно подчиняется закону «все или ничего», т. е. при силе раздражения выше порогового уровня происходит полное сокращение с максимальной для данного волокна силой, а ступенчатое повышение силы сокращения по мере увеличения силы раздражения невозможно. Поскольку смешанная мышца состоит из множества волокон с различным уровнем чувствительности к возбуждению, сокращение всей мышцы может быть ступенчатым в зависимости от силы раздражения, при этом при сильных раздражениях происходит активация глубжележащих мышечных волокон.
Механизм скольжения филаментов[править | править код]
рис. 1. Схема образования поперечных связей — молекулярной основы сокращения саркомераУкорочение мышцы происходит за счет укорочения образующих ее саркомеров, которые, в свою очередь, укорачиваются за счет скольжения относительно друг друга актиновых и миозиновых филаментов (а не укорочения самих белков). Теория скольжения филаментов была предложена учеными Huxley и Hanson (Huxley, 1974; рис. 1). (В 1954 г. две группы исследователей — X. Хаксли с Дж. Хэнсон и А. Хаксли с Р. Нидергерке — сформулировали теорию, объясняющую мышечное сокращение скольжением нитей. Независимо друг от друга они обнаружили, что длина диска А оставалась постоянной в расслабленном и укороченном саркомере. Это позволило предположить, что есть два набора нитей — актиновые и миозиновые, причем одни входят в промежутки между другими, и при изменении длины саркомера эти нити каким-то образом скользят друг по другу. Сейчас эта гипотеза принята почти всеми.)
Актин и миозин — два сократительных белка, которые способны вступать в химическое взаимодействие, приводящее к изменению их взаимного расположения в мышечной клетке. При этом цепочка миозина прикрепляется к актиновой нити с помощью целого ряда особых «головок», каждая из которых сидит на длинной пружинистой «шее». Когда происходит сцепление между миозиновой головкой и актиновой нитью, конформация комплекса этих двух белков изменяется, миозиновые цепочки продвигаются между актиновыми нитями и мышца в целом укорачивается (сокращается). Однако, чтобы химическая связь между головкой миозина и активной нитью образовалась, необходимо подготовить этот процесс, поскольку в спокойном (расслабленном) состоянии мышцы активные зоны белка актина заняты другим белком — тропохмиозином, который не позволяет актину вступить во взаимодействие с миозином. Именно для того, чтобы убрать тропомиозиновый «чехол» с актиновой нити, требуется быстрое выливание ионов кальция из цистерн саркоплазматического ретикулума, что происходит в результате прохождения через мембрану мышечной клетки потенциала действия. Кальций изменяет конформацию молекулы тропомиозина, в результате чего активные зоны молекулы актина открываются для присоединения головок миозина. Само это присоединение осуществляется с помощью так называемых водородных мостиков, которые очень прочно связывают две белковые молекулы — актин и миозин — и способны в таком связанном виде находиться очень долго.
Для отсоединения миозиновой головки от актина необходимо затратить энергию аденозинтрифосфа-та (АТФ), при этом миозин выступает в роли АТФазы (фермента, расщепляющего АТФ). Расщепление АТФ на аденозиндифосфат (АДФ) и неорганический фосфат (Ф) высвобождает энергию, разрушает связь между актином и миозином и возвращает головку миозина в исходное положение. В дальнейшем между актином и миозином могут снова образовываться поперечные связи.
При отсутствии АТФ актин-миозиновые связи не разрушаются. Это и является причиной трупного окоченения (rigor mortis) после смерти, т. к. останавливается выработка АТФ в организме — АТФ предотвращает мышечную ригидность.
Даже при мышечных сокращениях без видимого укорочения (изометрические сокращения, см. выше) активируется цикл формирования поперечных связей, мышца потребляет АТФ и выделяет тепло. Головка миозина многократно присоединяется на одно и то же место связывания актина, и вся система миофиламентов остается неподвижной.
Внимание: Сократительные элементы мышц актин и миозин сами по себе не способны к укорочению. Мышечное укорочение является следствием взаимного скольжения миофиламентов относительно друг друга (механизм скольжения филаментов).
Как же образование поперечных связей (водородных мостиков) переходит в движение? Одиночный саркомер за один цикл укорачивается приблизительно на 5-10 нм, т.е. примерно на 1 % своей общей длины. За счет быстрого повторения цикла поперечных связей возможно укорочение на 0,4 мкм, или 20% своей длины. Поскольку каждая миофибрилла состоит из множества саркомеров и во всех них одновременно (но не синхронно) образуются поперечные связи, суммарно их работа приводит к видимому укорочению всей мышцы. Передача силы этого укорочения происходит через Z-линии миофибрилл, а также концы сухожилий, прикрепленных к костям, в результате чего и возникает движение в суставах, через которые мышцы реализуют перемещение в пространстве частей тела или продвижение всего тела.
Связь между длиной саркомера и силой мышечных сокращений[править | править код]
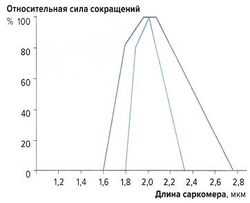 рис. 2. Зависимость силы сокращений от длины саркомера
рис. 2. Зависимость силы сокращений от длины саркомераНаибольшую силу сокращений мышечные волокна развивают при длине 2-2,2 мкм. При сильном растяжении или укорочении саркомеров сила сокращений снижается (рис. 2). Эту зависимость можно объяснить механизмом скольжения филаментов: при указанной длине саркомеров наложение миозиновых и актиновых волокон оптимально; при большем укорочении миофиламенты перекрываются слишком сильно, а при растяжении наложение миофиламентов недостаточно для развития достаточной силы сокращений.
Скорость укорочения мышечных волокон[править | править код]
рис.3. Зависимость скорости укорочения от нагрузкиСкорость укорочения мышцы зависит от нагрузки на эту мышцу (закон Хилла, рис. 3). Она максимальна без нагрузки, а при максимальной нагрузке практически равна нулю, что соответствует изометрическому сокращению, при котором мышца развивает силу, не изменяя своей длины.
Влияние растяжения на силу сокращений: кривая растяжения в покое[править | править код]
рис. 4. Влияние предварительного растяжения на силу сокращения мышцы. Предварительное растяжение повышает напряжение мышцы. Результирующая кривая, описывающая взаимоотношения длины мышцы и силы ее сокращения при воздействии активного и пассивного растяжения, демонстрирует более высокое изометрическое напряжение, чем в покоеВажным фактором, влияющим на силу сокращений, является величина растяжения мышцы. Тяга за конец мышцы и натяжение мышечных волокон называются пассивным растяжением. Мышца обладает эластическими свойствами, однако в отличие от стальной пружины зависимость напряжения от растяжения не линейна, а образует дугообразную кривую. С увеличением растяжения повышается и напряжение мышцы, но до определенного максимума. Кривая, описывающая эти взаимоотношения, называется кривой растяжения в покое.
Данный физиологический механизм объясняется эластическими элементами мышцы — эластичностью сарколеммы и соединительной ткани, располагающимися параллельно сократительным мышечным волокнам.
Также при растяжении изменяется и наложение друг на друга миофиламентов, однако это не оказывает влияния на кривую растяжения, т. к. в покое не образуются поперечные связи между актином и миозином. Предварительное растяжение (пассивное растяжение) суммируется с силой изометрических сокращений (активная сила сокращений).
- ↑ Harrison BC. et al. 2011. lib or not lib? Regulation of myosin heavy chain gene expression in mice and men. Skeletal Muscle. 1 (1): 5. doi: 10.1186/2044-5040-1-5.
- ↑ Andersen, J.L., et al. 1994. Myosin heavy chain isoforms in single fibres from m. vastus lateralis of sprinters: Influence of training. Acta Physiologica Scandinavica 151 (2): 135-42.
- ↑ 3,03,1 Andersen T.L, Aagaard P. 2000. Myosin heavy chain IIX overshoot in human skeletal muscle. Muscle Nerve. 23 (7): 1095-104.
- ↑ Andersen, L.L., et al. 2010. Early and late rate of force development: Differential adaptive responses to resistance training? Scandinavian Journal of Medicine and Science in Sports 20 (1): el62-69. doi:10.1111/j.l600-0838.2009.00933.x.
- ↑ Anderson, K., and Behm, D.G. 2004. Maintenance of EMG activity and loss of force output with instability. Journal of Strength and Conditioning Research 18:637-40.
- ↑ Aagaard, R, et al. 2011. Effects of resistance training on endurance capacity and muscle fiber composition in young top-level cyclists. Scandinavian Journal of Medicine and Science in Sports 21 (6): e298-307. doi:10.1111/j. 1600-0838.2010.01283.x.
- ↑ Henneman, E., Somjen, G., and Carpenter, D.O. 1965. Functional significance of cell size in spinal motoneurons./. Neurophysiol. 28:560-580.
- ↑ 8,08,1 Latash, M.L. 1998. Neurophysiological basis of movement. Champaign, IL: Human Kinetics.
- ↑ Brooks, G.A., Fahey, T.D., and White, T.P. 1996. Exercise physiology: Human bioenergetics and its applications. 2nd ed. Mountainview, CA: Mayfield.
- ↑ Schmidtbleicher, D. 1992. Training for power events. In Strength and power in sport, ed. P.V. Komi, 381-95. Oxford, UK: Blackwell Scientific.
- ↑ Wiemann, K., and Tidow, G. 1995. Relative activity of hip and knee extensors in sprinting—Implications for training. New Studies in Athletics 10 (1): 29-49.
sportwiki.to
Сокращение мышц
Специфическая деятельность мышечной ткани — ее сокращение при возбуждении. При сокращении мышца укорачивается и развивает силу, которая вызывает растяжение упругого компонента. Последний играет роль буфера при передаче усилий, возникающих в сократительном компоненте, к подвижным звеньям, вследствие этого движения звеньев тела становятся плавными.
Механизм мышечного сокращения обусловлен взаимодействием актина и миозина. Взаимодействие актина и миозина тормозится системой мышечных белков. На поверхности актиновых нитей имеется два белка — тропонин и тропомиозин. Поступление импульса к мышце сопровождается выходом из саркоплазматического ретикулума мышечного волокна ионов Са2+, которые, взаимодействуя с белком тропонином, образуют комплекс, и он толкает тропомиозин в желобки между двумя цепями актина. За счет гребковых движений головок (специального белка) миозиновых нитей актиновые нити втягиваются на миозиновые, и мышца укорачивается. Кальциевый насос транспортирует ионы Са2+ в систему саркоплазматического ретикулума, происходит отсоединение поперечных мостиков миозина от актина, и мышца расслабляется. Непосредственным источником энергии для сокращения является АТФ. Энергия АТФ обеспечивает перемещение поперечных мостиков. Молекула АТФ связывается с поперечным мостиком после завершения его гребкового движения. Расщепление АТФ до АДФ и фосфата — обязательное условие для поперечного мостика к актину.
Рис. 6. Поперечные мостики
Различают одиночное и тетаническое сокращение мышцы.
Одиночное сокращение мышцы. На однократное кратковременное раздражение, например электрическим током, мышца отвечает одиночным сокращением. При записи этого сокращения на кимографе отмечают три периода; латентный—от раздражения до начала сокращения, период сокращения и период расслабления.
Общая продолжительность одиночного сокращения икроножной мышцы лягушки составляет 0,1 с. Из этого времени 0,01 с приходится на латентный период, 0,04 — на сокращение и 0,05 с — на расслабление. У млекопитающих одиночное сокращение скелетных мыши длится 0,04-0,1 с.
Во время латентного периода в мышце происходят процессы, при которых освобождается энергия для мышечного сокращения. Пик потенциала действия по времени совпадает с латентным периодом и началом сокращения.
Время одиночного сокращения неодинаково в различных мышцах у одного и того же животного. Так, в красных волокнах мышц оно значительно больше, чем в белых. Сила мышечного сокращения в определенной степени зависит от силы раздражения. При раздражении током пороговой силы сокращение мышцы едва заметно. Дальнейшее увеличение силы тока вызывает усиление сокращения мышцы до некоторой максимальной величины. Сила мышечного сокращения зависит от количества возбужденных мышечных волокон, которые обладают различной возбудимостью. Слабое раздражение действует на наиболее возбудимые волокна, по мере усиления раздражения начинают реагировать и другие волокна, наконец наступает момент, когда возбуждаются все мышечные волокна — наступает максимальное сокращение мышцы.
Тетаническое сокращение мышцы. Если к мышцам поступают несколько возбуждающих импульсов, ее одиночные сокращения суммируются, в результате этого происходит сильное и длительное сокращение мышцы. Длительное сокращение мышцы при ее ритмическом раздражении называется тетаническим сокращением или тетанусом. Meханизм тетанического сокращения мышцы был выяснен Гельмгольцем в опытах на нервно-мышечном препарате. При нанесении на мышцу одиночных раздражений, следующих друг за другом с различными интервалами времени, наблюдают сокращения разной формы. В том случае, когда раздражения отделены друг от друга интервалом времени, превышающим продолжительность одиночного сокращения, возникают одиночные сокращения. При более частых раздражениях, если каждый импульс возбуждения действует на мышцу в тот момент, когда она уже начинает расслабляться, отмечают неполный, или зубчатый, тетанус.
Гладкий тетанус. Если же раздражения настолько частые, что они воздействуют на мышцу до начала ее расслабления, то получится длительное непрерывное сокращение мышцы — гладкий тетанус. При очень большой частоте раздражений каждое очередное раздражение будет попадать на фазу абсолютной рефракторности и мышца вообще не сократится.
Высота сокращения мышцы при тетанусе больше, чем при одиночном сокращении. Объясняя этот эффект, Гельмгольц рассматривал тетанус как простое геометрическое накладывание (суперпозицию) одиночных сокращений. При тетанусе каждое последующее сокращение мышцы в ряду складывающихся одиночных сокращений начинается от той точки, где мышцу застает новое раздражение, и ее сокращение идет от этой точки, как от точки покоя. Дальнейшие исследования показали, что явление суперпозиции при тетанусе нельзя сводить к простому складыванию, то есть суммированию механических эффектов. Эффект от двух следующих друг за другом раздражений не совпадает с арифметической суммой одиночных сокращений, он может быть больше или меньше данной суммы. Следовательно, способность к новому сокращению после каждого предшествующего импульса возбуждения неодинакова.
Рис. 7. Сокращение скелетной мышцы при различной частоте раздражения:
I – одиночное сокращение; II – зубчатый (неполный) тетанус; III – гладкий (полный) тетанус; М – механограмма; ПД – потенциал действия; 1 – фаза сокращения; 2 – фаза расслабления
Рис. 8. Структура сократительного механизма нормальной скелетной мышцы.
Высота мышечного сокращения при тетанусе зависит от ритма раздражения, а также от возбудимости и лабильности, которые изменяются в процессе деятельности мышцы. Тетанус наиболее высокий при оптимальном ритме, когда каждый последующий импульс действует на мышцу в фазу экзальтации, вызванной предыдущим импульсом. В этом случае создаются наилучшие (оптимальные) условия для работы мышцы.
Существуют два вида сокращения мышц: изотоническое и изометрическое.
Когда мышца при раздражении сокращается, не поднимая никакого груза, напряжение ее мышечных волокон не изменяется и равно нулю; такое сокращение называют изотоническим (isos — равный, tonus — напряжение). Если концы мышцы закреплены, то при раздражении она ни укорачивается, а лишь сильно напрягается.
Сокращение мышцы, при котором ее длина остается постоянной, называется изометрическим (isos — равный. metron — мера, размер). В этом случае сократительный компонент укорачивается за счет растяжения пассивного упругого компонента. Если у изометрически сокращающейся мышцы освободить сухожилие, то мышца станет сокращаться изотонически, а предварительно растянутый упругий компонент очень быстро укорачивается. Упругий компонент при изометрическом сокращении может увеличивать свою длину на 2-6% от длины покоя.
studfiles.net
Мышца: почему она сокращается
|
www.nazdor.ru
Непроизвольные сокращения мышц: причины и лечение
На правах рекламы:
Невроз – это настоящая «болезнь XXI века», образ жизни современного человека способствует стремительному распространению этой патологии. Неврозы или невротические состояния – это обширная группа заболеваний нервной системы, которые носят психогенный характер. И, хотя, невроз не является угрозой для жизни, этот недуг в состоянии значительно ухудшить качество жизни человека, создать ему множество проблем в общении с окружающими людьми.
Несмотря на психогенную природу невроза, существуют и многочисленные вегетативные симптомы этого заболевания. Одними из наиболее очевидных и неприятных признаков невроза является непроизвольное изменение тонуса мышц: их постоянное напряжение или периодическое мышечные подергивания.

Содержание статьи
Похожее на сайте:
Общее описание
Тики или мышечные подергивания – это непроизвольные, периодические возникающие подергивания мышц, которые могут иметь разную локализацию. Непроизвольно сокращаться могут мимические мышцы, дыхательные мышцы (при этом человек может издавать разные звуки: покашливать, попискивать, тяжело вздыхать, внезапно прерывать свою речь, что напоминает симптомы заикания). Также бывают судорожные подергивания мышц верхних и нижних конечностей. Подобные симптомы могут встречаться как у детей, так и у взрослых.
Следует отметить, что не только невроз вызывает подергивания мышц. Подобные симптомы могут быть результатом резидуальной церебральной органической недостаточности, вегетососудистой дистонии, алкоголизма, наркомании, эпилепсии, шизофрении или дисфункции щитовидной железы. Точную причину непроизвольного сокращения мышц может установить только опытный врач.
Еще одним симптомом, который часто вызывает невроз, является напряжение мышц. Так же как и мышечные подергивания, напряжение мышц может быть вызвано разными причинами. Защемление нервов (невралгия) тоже может вызывать подобные симптомы. Она может быть шейной, грудной и поясничной. Причина этого заболевания – сдавливание спинномозговых нервов в результате действия разных факторов. Чаще всего это остеохондроз разных отделов позвоночника. Однако защемление могут вызывать и другие факторы.
Вызывают болезненное напряжение мышц повреждения черепно-мозговых нервов, которые иннервируют различные группы мышц. Примером может служить неврит лицевого нерва.
Психоневрозы также вызывают излишнее мышечное напряжение, которое часто приводит к нарушению осанки. Можно часто увидеть людей, движения которых очень зажаты и похожи на движения механизмов. Психологи установили четкую корреляцию между психическим состоянием человека и тонусом его мышц.
Так, например, повышенное напряжение мышц рук часто связано с повышенной агрессивностью, а мышечные зажимы мышц таза и нижних конечностей – с проблемами в сексуальной сфере.

Причины возникновения
Причиной возникновения непроизвольных сокращений мышц при неврозе является состояние беспокойства, тревоги и страха, которые характерны для невротического состояния. Именно эти причины приводят к состоянию психического возбуждения, которое вызывает подергивания и мышечные спазмы.
Это было подтверждено поведением больных под влиянием внушения или гипноза. Мышечные тики, как и напряжение – это своего рода форма психомоторной разрядки накопившегося болезненного внутреннего напряжения.
Обычно люди, у которых наблюдаются подобные симптомы, отличаются высокой эмоциональной активностью и ранимостью. При этом они стараются держать свои эмоции глубоко в себе. Они очень болезненно относятся к критике, эмоционально неустойчивы.
Очень часто причиной возникновения непроизвольного сокращения мышц являются наследственные факторы, подобные симптомы нередко передаются от родителей к детям. Но генетические факторы могут обуславливать только предрасположенность к подобным симптомам. Основной причиной их появления являются внешние факторы и реакция человека на них.
Современный человек живет весьма напряженной жизнью. Постоянная спешка, хроническая нехватка времени, стрессовые ситуации отрицательно сказываются на работе нервной системы. Кроме того, в последние десятилетия постоянно увеличивается количество людей, которые заняты умственным трудом.
Человек, проживающий в большом городе, ежедневно перерабатывает огромное количество информации, что также создает ощутимую нагрузку для нервной системы. Все вышеперечисленные факторы приводят к неврозам.
Следует помнить, что непроизвольные сокращения мышц – это всего лишь один из многочисленных симптомов этого заболевания. Вот основные причины невроза:
- Сильные умственные или физические нагрузки, которые связаны с постоянной работой, отсутствие нормально отдыха (особенно полноценного сна) в течение долгого времени. В итоге у человека возникает состояние постоянного (хронического) стресса, который вызывает невроз. Еще одной причиной возникновения подобных симптомов могут быть переживания личного характера, обе причины вместе дают поистине «адский коктейль» и почти гарантировано приводят к неврозу.
- Невозможность правильно выполнить определенную задачу или добиться поставленной цели. Очень частая причина неврозов, которые могут вызывать непроизвольное сокращение мышц.
- Также подобные симптомы могут быть вызваны нежеланием или неумением человека отдыхать. Такой график приводит к нервному истощению и является одной из причин начала невроза.
- Злоупотребление спиртными напитками или употребление наркотиков. Подобные вещества очень сильно истощают нервную систему и являются одним из факторов развития заболевания. Очень часто люди прибегают к алкоголю для того, чтобы снять нервное напряжение, расслабиться. Это неверная тактика. Алкоголь расшатывает нервную систему и усугубляет невроз.
Лечение
Лечение нервного напряжения мышц или их непроизвольного подергивания заключается в устранении причины, которой они вызваны. Ибо эти признаки – это всего лишь симптомы заболевания. Лечение можно разделить на две больших группы: немедикаментозное и лечение с помощью лекарственных препаратов.
Снять (полностью или частично) решить проблему позволяет применение седативных препаратов, транквилизаторов и антидепрессантов. Назначать лекарственные препараты должен врач, самолечение недопустимо.
К довольно эффективным методам лечения неврозов и устранения неприятных симптомов относится психотерапия. Однако этот метод лечения требует много времени и напряженной работы как врача, так и пациента. Для того чтобы победить болезнь пациенту часто необходимо изменить свой прежний образ жизни: снизить ежедневные нагрузки, отказаться от вредных привычек, научится по-другому относиться к жизненным трудностям и невзгодам. Очень важен настрой больного, его желание победить недуг, освободиться от него. Это весьма сложный процесс, но иначе победить невроз невозможно.
Чаще всего пациенты избавляются от непроизвольного сокращения мышц, если оно вызвано именно невротическими состояниями. Но преодоление этой проблемы может занять достаточно много времени.
Советуем посмотреть:
lechenienevroza.ru
Механизм мышечных сокращений кратко :: SYL.ru
Процессы мышечной работы представляют собой многоуровневый комплекс физиологических и биохимических функций, жизненно важных для полноценной работы человеческого организма. Внешне подобные процессы можно наблюдать на примерах произвольных движений при ходьбе, беге, изменении мимики и т. д. Однако они охватывают гораздо больший спектр функций, в числе которых также значится работа дыхательного аппарата, органов пищеварения и выделительной системы. В каждом случае механизм мышечных сокращений подкрепляется работой миллионов клеток, в которой задействуются химические элементы и физические волокна.
Структурная организация мышцы
Мышцы формируются множеством волокон ткани, которые имеют узлы крепления к костям скелета. Они располагаются параллельно и в процессе мышечной работы взаимодействуют между собой. Именно волокна при поступлении импульсов обеспечивают механизм мышечного сокращения. Кратко структуру мышцы можно представить как систему, состоящую из молекул саркомер и миофибрилла. Важно понимать, что каждое мышечное волокно образуется множеством субъединиц миофибрилл, располагающихся продольно по отношению друг к другу. Теперь стоит отдельно рассмотреть саркомеры и филаменты. Поскольку они играют важную роль в двигательных процессах.
Саркомеры и филаменты
Саркомеры представляют собой сегменты волокон, которые отделяются так называемыми Z-пластинами, содержащими бета-актинин. От каждой пластины отходят актиновые филаменты, а промежутки заполняются толстыми миозиновыми аналогами. Актиновые элементы, в свою очередь, похожи на ниточки бус, закрученных в двойную спираль. В этой структуре каждая бусинка является молекулой актина, а в участках с углублениями в спирали находятся молекулы тропонина. Каждая из этих структурных единиц формирует механизм сокращения и расслабления мышечного волокна, связываясь друг с другом. Ключевую роль в возбуждении волокон играет клеточная мембрана. В ней заключены поперечные трубочки-инвагинации, которые активизируют функцию саркоплазматического ретикулума – это и будет возбуждающий эффект для мышечной ткани.
Двигательная единица

Теперь стоит отойти от углубленной структуры мышцы и рассмотреть двигательную единицу в общей конфигурации скелетной мышцы. Это будет совокупность мышечных волокон, иннервируемых отростками мотонейрона. Работа ткани мышцы независимо от характера действия будет обеспечиваться волокнами, включенными в состав одной двигательной единицы. То есть при возбуждении мотонейрона срабатывает механизм мышечных сокращений в рамках одного комплекса с иннервируемыми отростками. Такое разделение на мотонейроны позволяет целенаправленно задействовать конкретные мышцы, не возбуждая без надобности соседние двигательные единицы. По сути, вся мышечная группа одного организма делится на сегменты мотонейронов, которые могут объединяться в работе над сокращением или расслаблением, а могут действовать разнопланово или поочередно. Главное, что они независимы друг от друга и работают только с сигналами своей группы волокон.
Молекулярные механизмы мышечной работы
В соответствии с молекулярной концепцией о скольжении нитей, работа мышечной группы и, в частности, ее сокращение реализуется в ходе скользящего действия миозинов и актинов. Реализуется сложный механизм взаимодействия этих нитей, в котором можно выделить несколько процессов:
- Центральная часть миозиновой нити соединяются со связками актинов.
- Достигнутый контакт актина с миозином способствует конформационному перемещению молекул последнего. Головки вступают в фазу активности и разворачиваются. Таким образом осуществляются молекулярные механизмы мышечного сокращения на фоне перестройки нитей активных элементов по отношению друг к другу.
- Затем происходит взаимное расхождение миозинов и актинов с последующим восстановлением головной части последних.
Весь цикл выполняется несколько раз, в результате чего происходит смещение вышеупомянутых нитей, а Z-сегменты саркомеров сближаются и укорачиваются.
Физиологические свойства работы мышц
Среди основных физиологических свойств мышечной работы выделяют сократимость и возбудимость. Эти качества, в свою очередь, обуславливаются проводимостью волокон, пластичностью и свойством автоматии. Что касается проводимости, то она обеспечивает распространение процесса возбудимости между миоцитами по нексусам – это специальные электропроводящие контуры, отвечающие за проведение импульса сокращения мышцы. Однако после сокращения или расслабления тоже совершается работа волокон.
За их спокойное состояние в определенной форме отвечает пластичность, определяющая сохранение постоянного тонуса, в котором на текущий момент находится механизм мышечного сокращения. Физиология пластичности может проявляться как в виде сохранения укороченного состояния волокон, так и в их растянутом виде. Интересно и свойство автоматии. Она определяет способность мышц входить в рабочую фазу без подключения нервной системы. То есть миоциты самостоятельно вырабатывают ритмически повторяющиеся импульсы для тех или иных действий волокон.
Биохимические механизмы мышечной работы
В работе мышц участвует целая группа химических элементов, среди которых кальций и сократительные белки наподобие тропонина и тропомиозина. На базе этого энергетического обеспечения и выполняются рассмотренные выше физиологические процессы. Источником же этих элементов выступает аденозинтрифосфорная кислота (АТФ), а также ее гидролиз. При этом запас АТФ в мышце способен обеспечивать сокращение мышцы лишь в течение доли секунды. Несмотря на это, волокна могут отвечать на нервные импульсы в постоянном режиме.
Дело в том, что биохимические механизмы мышечного сокращения и расслабления с поддержкой АТФ связаны с процессом выработки резервного запаса макроэрга в виде креатинфосфата. Объем этого резерва в несколько раз превышает запас АТФ и в то же время способствует его генерации. Также помимо АТФ энергетическим источником для мышцы может выступать гликоген. К слову, на мышечные волокна приходится около 75% всего запаса данного вещества в организме.
Сопряжение возбудительных и сократительных процессов

В спокойном состоянии нити волокон не взаимодействуют друг с другом посредством скольжения, так как центры связок закрываются молекулами тропомиозина. Возбуждение может иметь место только после электромеханического сопряжения. Данный процесс также делится на несколько этапов:
- При активации нейромышечного синапса на мембране миофибриллы формируется так называемый постсинаптический потенциал, накапливающий энергию для действия.
- Возбуждающий импульс благодаря системе трубок расходится по мембране и активизирует ретикулум. Этот процесс в итоге способствует снятию барьеров с каналов мембраны, по которым выпускаются ионы, связывающиеся с тропонином.
- Белок тропонин, в свою очередь, открывает центры связок актина, после чего становится возможным механизм мышечных сокращений, но для его начала также потребуется соответствующий импульс.
- Использование открывшихся центров начнется в момент, когда к ним присоединятся головки миозина по описанной выше модели.
Полный цикл этих операций происходит в среднем за 15 мс. Период от начальной точки возбуждения волокон до полного сокращения называется латентным.
Процесс расслабления скелетной мышцы
При расслаблении мышц происходит обратный перенос ионов Са++ с подключением ретикулума и кальциевых каналов. В процессе выхода ионов из цитоплазмы количество центров связки сокращается, в результате чего происходит разъединение актиновых и миозиновых филаментов. Иными словами, механизмы мышечного сокращения и расслабления подключают те же функциональные элементы, но оперируют ими разными способами. После расслабления может наступать процесс контрактуры, при котором отмечается устойчивое сокращение мышечных волокон. Это состояние может сохраняться до момента, пока не наступит очередное действие раздражающего импульса. Бывает и контрактура краткого действия, предпосылками для которой становится тетаническое сокращение в условиях скопления ионов с большими объемами.

Фазы сокращения
Когда мускулатура приводится в действие раздражающим импульсом сверхпороговой силы, происходит одиночное сокращение, в котором можно выделить 3 фазы:
- Уже упомянутый выше период сокращения латентного типа, в процессе которого волокна накапливают энергию для совершения последующих действий. В это время проходят процессы электромеханического сопряжения и открываются центры связок. На данной стадии подготавливается механизм сокращения мышечного волокна, который активизируется после распространения соответствующего импульса.
- Фаза укорочения – длится 50 мс в среднем.
- Фаза расслабления – также длится примерно 50 мс.
Режимы мышечного сокращения
Работа при одиночном сокращении была рассмотрена как пример «чистой» механики мышечных волокон. Однако в естественных условиях такая работа не совершается, поскольку волокна находятся в постоянном отклике на сигналы двигательных нервов. Другое дело, что в зависимости от характера этого отклика может происходить работа в следующих режимах:
- Сокращения возникают при пониженной частоте импульсов. Если электрический импульс распространяется после завершения расслабления, то следует серия одиночных актов сокращения.
- Высокая частота импульсных сигналов может совпадать с расслабляющей фазой предшествующего цикла. В этом случае амплитуда, в которой работал механизм сокращения мышечной ткани, будет суммироваться, что обеспечит длительное сокращение с неполными актами расслабления.
- В условиях повышения частоты импульсов новые сигналы будут действовать в периоды укорочения, что спровоцирует длительное сокращение, которое не будет прерываться расслаблениями.
Оптимум и пессимум частоты

Амплитуды сокращений определяются частотой импульсов, которые раздражают мышечные волокна. В этой системе взаимодействия сигналов и откликов можно выделить оптимум и пессимум частоты. Первым обозначается частота, которая в момент действия будет накладываться на фазу повышенной возбудимости. В таком режиме может активизироваться механизм сокращения мышечного волокна с большой амплитудой. В свою очередь, пессимум определяет более высокую частоту, импульс которой приходится на фазу рефрактерности. Соответственно, в этом случае амплитуда уменьшается.
Виды работы скелетной мышцы
Мышечные волокна могут осуществлять работу динамически, статически и динамически-уступающе. Стандартная динамическая работа является преодолевающей – то есть мышца в момент сокращения перемещает объекты или его составные части в пространстве. Статическое действие мышцы в некотором роде избавлено от нагрузок, поскольку в этом случае не предусматривается изменение его состояния. Динамически-уступающий механизм мышечного сокращения скелетной мышцы срабатывает, когда волокна функционируют в условиях растяжения. Потребность в параллельном растяжении также может быть обусловлена тем, что работа волокон предполагает выполнение операций со сторонними телами.
В заключение
Процессы организации мышечного действия подключают самые разные функциональные элементы и системы. В работе задействуется сложный комплекс участников, каждый из которых выполняет свою задачу. Можно видеть, как в процессе активации механизма мышечных сокращений срабатывают и косвенные функциональные блоки. Например, это касается процессов генерации энергетического потенциала для совершения работы или системы блокировки центров связок, через которые происходит соединение миозинов и актинов.
Основная же нагрузка приходится непосредственно на волокна, которые выполняют те или иные действия по командам двигательных единиц. Причем характер выполнения определенной работы может быть разным. На него будут влиять параметры направляемого импульса, а также текущее состояние мышцы.
www.syl.ru