
У нас есть шесть частей тела, которые мы больше не используем. Зачем они были нужны?
Автор фото, Getty Images
Подпись к фото,Рудименты — отголоски эволюции
Эволюция прошла долгий путь, и процесс этот очень медленный.
Некоторые отличительные признаки организмов сохраняются на протяжении многих поколений даже после того, как соответствующий орган перестал выполнять отведенную ему функцию. Эти эволюционные остатки, или рудиментарные особенности, есть и у людей.
«Ваше тело — это, по сути, музей естествознания!» — написала в «Твиттере» эволюционный антрополог Дорса Амир.
Так почему же эти свойства или органы не пропадают, несмотря на то, что они, судя по всему, утратили свою функцию? Потому что эволюция — это постепенный процесс.
Автор фото, Getty Images
Иногда на них не оказывает достаточного давления естественный отбор, поэтому они переходят из поколения в поколение. В некоторых случаях рудиментарные органы развивают новые функции. Этот процесс называется экзаптацией.
В некоторых случаях рудиментарные органы развивают новые функции. Этот процесс называется экзаптацией.
Откуда мы вообще знаем, для чего эти органы или части тела изначально предназначались?
«Мы можем только предполагать, какова основная функция этих органов, — сказала Дорса Амир в интервью Би-би-си. — Мы можем выяснить, например, важны ли они для выживания, или посмотреть, есть ли они у ближайших к нам приматов и млекопитающих, и если да, то как они функционируют».
Вот шесть этих рудиментов.
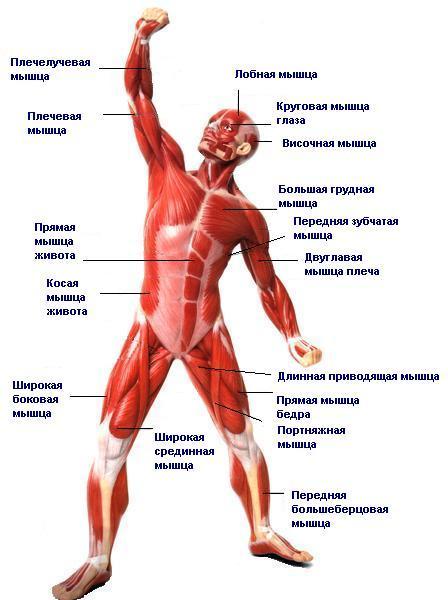
1. Palmaris longus — мускул на запястье
Автор фото, Getty Images
Подпись к фото,С помощью этих мышц люди передвигались по деревьям
Проведем небольшой эксперимент: положите руку ладонью вверх на плоскую поверхность и соедините большой палец с мизинцем.
Видите бугорок, который появился у вас на запястье? Это Palmaris longus — длинная ладонная мышца.
Не волнуйтесь, если не увидите ее. Примерно у 18% людей ее вовсе нет, и это абсолютно ни на что не влияет. Прекрасный пример эволюционного рудимента.
Примерно у 18% людей ее вовсе нет, и это абсолютно ни на что не влияет. Прекрасный пример эволюционного рудимента.
Эта мышца присутствует у живущих в лесу или на деревьях приматов, таких как орангутанги, но есть не у всех приматов, обитающих на других территориях.
«Это свидетельствует о том, что эта мышца нужна, чтобы лазить по деревьям», — говорит Дорса. В наши дни практическое применение этой мышце нашли хирурги.
«Они используют ее в качестве материала при пластических операциях, поскольку сама по себе она не выполняет никакой функции, необходимой для движения рук», — говорит Дорса.
2. Бугорок Дарвина можно найти на верхней части уха
Автор фото, Getty Images
Подпись к фото,Некоторые млекопитающие используют эти мышцы, чтобы определять местонахождение добычи и хищников
«Если вы можете шевелить ушами, вы демонстрируете эволюцию», — пишет Джерри Койн в своей книге «Почему эволюция — это правда».
Речь здесь идет о трех мышцах под кожей головы, которые прикреплены к ушам. Маленькая шишечка на верхней части уха — одна из этих мышц.
У большинства людей они уже не работают, но некоторые до сих пор могут использовать их, чтобы шевелить ушами.
Этот элемент был впервые в общих чертах описан Чарльзом Дарвином и поэтому называется бугорком Дарвина.
«Хотя по-прежнему идут споры о том, является ли сам бугорок рудиментарным, утверждается, что мышцы вокруг уха могут демонстрировать рудиментарность», — говорит Дорса.
Эти мышцы по-прежнему используются многими животными, например, кошками и лошадьми, чтобы двигать ушами, как отмечает Койн.
Это помогает им обнаруживать хищников, определять местонахождение своих детенышей и устанавливать, откуда идут различные звуки.
3. Копчик
Автор фото, Getty Images
Подпись к фото,Копчик был нужен нашим предкам для мобильности и баланса
Как отмечает Дорса Амир, копчик — наиболее очевидный эволюционный пережиток.
«Это напоминание об утерянных нами хвостах, которые были нужны для баланса и передвижения по деревьям», — говорит Дорса.
Он является хорошим примером процесса экзаптации, упомянутого ранее, поскольку теперь служит местом крепежа для мышц.
Другие подобные причуды не совсем выжили в эволюционном процессе.
Дорса говорит: «Определенные черты, такие как перепончатая ткань между пальцами, обнаруживаются на ранних этапах утробного развития, но затем исчезают. Эта ткань обычно уничтожается лейкоцитами».
4. Plica semilunaris — третье веко
Автор фото, Getty Images
Подпись к фото,Третье веко — это свернутая ткань во внутреннем углу глаза
Видите маленькую розовую подушечку во внутреннем углу глаза?
Это отголосок нашего эволюционного прошлого — наша перепончатая мембрана, или третье веко.
«Третье веко моргало бы горизонтально, — говорит Дорса. — У нас оно не функционирует». Но его все еще можно увидеть в действии в животном мире, например, у птиц и кошек.
— У нас оно не функционирует». Но его все еще можно увидеть в действии в животном мире, например, у птиц и кошек.
5. The piloerection — «мурашки»
Автор фото, Getty Images
Подпись к фото,Животные, такие как кошки, используют этот рефлекс, чтобы казаться крупнее
Вы видели, как у кошек шерсть встает дыбом, когда они напуганы?
Это очень похоже на то, как у нас появляются мурашки на коже, когда нам холодно или страшно.
Ученые называют это рефлексом пилоэрекции.
«Учитывая, что мы провели большую часть нашего времени на этой планете в виде покрытых шерстью млекопитающих, рефлекс пилоэрекции — это древний способ либо выглядеть крупнее, чем вы есть на самом деле, либо предотвращать потерю тепла, когда вам холодно», — говорит Дорса.
«Поскольку мы постепенно начали терять волосы на теле, этот рефлекс становился все менее и менее полезным, и теперь он уже не выполняет свою первоначальную функцию».
6. Palmar grasp reflex — хватательный рефлекс
Автор фото, Getty Images
Подпись к фото,Хватательный рефлекс нужнен детенышам приматов для траспортировки
Хватательный рефлекс наблюдается, когда дети крепко сжимают палец взрослого. Этот рефлекс по-прежнему нужен детенышам приматов.
Они рождаются готовыми схватиться за мех родителя для транспортировки.
«Предполагается, что наш собственный хватательный рефлекс ладоней изначально был предназначен именно для этой цели», — добавляет Дорса.
«Но наши дети рождаются преждевременно по сравнению с другими приматами и не могут сами держать голову или двигаться».
Интересно, что у разных людей наблюдаются разные рудиментарные особенности.
«Эволюционные пережитки» варьируются и в разных регионах мира, причем вразброс. И измениться это может только со временем.
Лечение жевательной мышцы — лечение височно челюстного сустава
Обычно мы не задумываемся о том, как двигается наша челюсть. Большинство людей, говоря о прикусе, подразумевает зубы. Но, как упоминалось выше, прикус – это целая система, контролирующая положение челюсти. Вот ее составляющие и их роль:
Большинство людей, говоря о прикусе, подразумевает зубы. Но, как упоминалось выше, прикус – это целая система, контролирующая положение челюсти. Вот ее составляющие и их роль:
Роль мышц
Рисунок №4 Открывание, закрывание рта, жевание и глотание происходит благодаря работе большого количества мышц.
Открывание рта обеспечивают мышцы, располагающиеся в области шеи. Мышцы, удерживающие нижнюю челюсть, находятся позади челюсти и проходят под щеками к лобной и заушной областям.
Самые мощные мышцы, отвечающие за жевание, расположены в щечной области.
Рисунок №5 Наш организм стремится осуществлять все функции с наибольшей рациональностью и наименьшей затратой энергии. Поэтому в норме жевательные мышцы удерживают челюсть в положении, при котором зубы на 1-2 мм разобщены.
Рисунок №6 Из этого рационального положения нижняя челюсть сотни раз в день перемещается до смыкания зубов для осуществления глотания.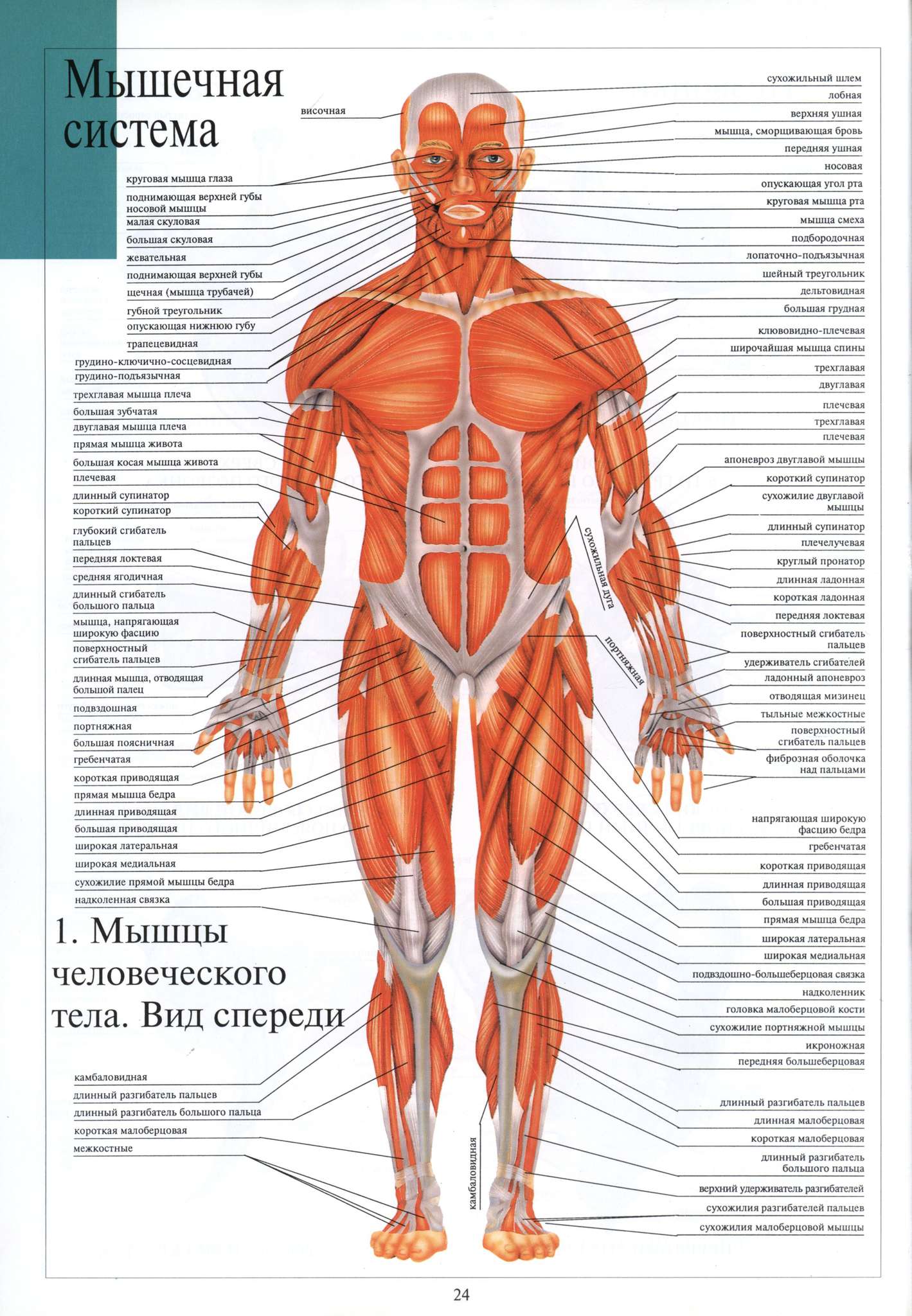
Все это происходит подсознательно. Наш мозг «запрограммирован» на процесс, который называется проприоцепция.
При правильном прикусе и положении челюсти в состоянии покоя большинство мышц находится в расслабленном состоянии.
Если Ваши зубы смыкаются не совсем правильно, мышцы могут приспосабливаться к этому, заставляя челюсть закрываться по траектории, вызывающей чрезмерное напряжение, спазм и со временем усталость мышц.
При этом положение физиологического покоя нижней челюсти (когда зубы не сомкнуты) не сопровождается расслаблением мышц, как это должно быть при нормальной окклюзии, а наоборот, приводит к их постоянному напряжению.
Роль суставов
Рисунок №7 Нижняя челюсть двигается благодаря суставам. называемым височнонижнечелюстными суставами, или ВНЧС.
Представьте сустав как мяч в корзине – суставная головка (мяч) двигается в углублении черепа (корзине). Между ними находится мягкотканная прослойка (суставной диск).
Положение суставной головки (мыщелка) в соответствующем углублении черепа (ямке) во многом определяется прикусом (окклюзией) – положением, где мышцы удерживают челюсть.
Рисунок №8 Если система сбалансирована, мыщелки расположены в центре суставной ямки и диск, находясь между суставной головкой и суставной ямкой, свободно двигается в суставе при открывании и закрывании рта.
Рисунок №9 Если жевательные мышцы приспосабливаются к неправильному положению челюсти, они могут удерживать головку сустава в смещенном положении, негативно влияя на ее функцию. Часто это приводит к смещению суставного диска, что препятствует нормальному его движению в суставе до того момента, пока он полностью не переместится в нормальное положение при открывании рта. Когда это происходит, возникает щелчок.
Интенсивность щелчка может варьироваться от едва осязаемой вибрации до очень громкого звука.
Этот звук часто является признаком ненормальной функции сустава, который в свою очередь, может быть связан с неправильным прикусом.
Роль осанки
Осанка также играет значительную роль в окклюзии. Нижнюю челюсть и нижние конечности можно считать взаимозависимыми частями скелетной системы человека. Если какая-либо из этих частей подвергается негативному влиянию, это может отразиться и на других частях скелета.
Проверим это, слегка сомкнув зубы. Обратите внимание на то, какие зубы сомкнулись первыми. Затем максимально откиньте голову назад и сомкните зубы еще раз. Не удивляйтесь, если на этот раз первый контакт придется на другие зубы. Изменяя положение головы, Вы тем самым влияете на баланс всей скелетной системы (и на Ваш прикус в частности).
То же самое происходит, когда Вам ставят пломбу. Лежа в стоматологическом кресле под анестезией кажется, что пломба не мешает. Но после того, как действие анестезии заканчивается, и Вы встаете с кресла, Вы замечаете, что пломба выше, чем остальные зубы. Вот почему в нейромышечной стоматологии прикус оценивают в положении сидя – положении, в котором обычно находится ваша челюсть. Если оценивать прикус в положении сидя, откинувшись назад, челюсть также смещается кзади. Такое положение отличается от обычного.
Если оценивать прикус в положении сидя, откинувшись назад, челюсть также смещается кзади. Такое положение отличается от обычного.
Фото №10,11 В эксперименте на крысах была изготовлена завышающая пломба на один боковой зуб с правой стороны. Через неделю peнтгенологическое обследование выявило деформацию всего позвоночника. После выравнивания окклюзии изготовлением пломбы на боковой зуб слева, форма позвоночника восстановилась.
Рисунок №12 Поскольку существует непосредственная связь между всеми частями скелетно-мышечной системы, несомненно, положение головы оказывает влияние на окклюзию, и может вызывать напряжение мышц шеи и спины.
Рисунок №13 При смещении нижней челюсти назад в результате аномалии прикуса в детстве или стираемости зубов со временем, шея перемещается вперед, и голова запрокидывается назад. Это, в свою очередь, значительно увеличивает нагрузку на мышцы шеи плечевого пояса. Поэтому у большинства людей с передним положением головы возникают боли в шейном отделе или напряжение мышц шеи.
Рисунок №14, 15 Часто смещение нижней челюсти при неправильном прикусе сопровождается ротацией первого и второго шейного позвонков, что ведет к выраженной деформации всего шейного отдела позвоночника.
Также, как изменения в положении челюсти могут вызывать изменения осанки и приводить к патологии в шейном отделе позвоночника, так и изменения в позвоночнике могут влиять на положение челюсти и окклюзию.
Роль дыхания и ночное апноэ
Рисунок №16 Смещение нижней челюсти назад часто приводит к сужению просвета дыхательных путей вследствие деформации шейного отдела позвоночника и смещения языка кзади, который следует за нижней челюстью. Для того, чтобы увеличить просвет дыхательных путей, мышцы еще больше перемещают шею вперед и запрокидывают голову назад.
При этом артерия, несущая кровь в головной мозг вдоль позвоночника, может резко перегибаться в области первого и второго шейных позвонков, вызывая нарушения кровоснабжения головного мозга.
Сужение дыхательных путей и нарушения кровоснабжения может вызывать состояние ночного апноэ – кратковременную остановку дыхания во время сна.
Повторяющиеся остановки дыхания приводят к значительному изменению метаболизма всего организма и, как следствие, могут стать одним из пусковых механизмов таких заболеваний как гипертония, сахарный диабет, инфаркт миокарда, инсульт и др. Поэтому нелеченное состояние ночного апноэ повинно в снижении продолжительности жизни на 20%.
Теперь Вы понимаете, что прикус гораздо сложнее, чем просто способ смыкания зубов! А неправильный прикус может быть причиной не только ранней потери зубов, но и вызывать такие серьезные патологии, как диабет и инфаркт миокарда и даже снижать продолжительность жизни.
3D-анатомия. На чем учатся студенты-медики в Самаре | ЗДОРОВЬЕ: Медицина | ЗДОРОВЬЕ
Существует ли такая проблема в Самарском государственном медицинском университете и смогут ли виртуальные тренажеры подготовить профессиональных хирургов к операциям, узнал журналист «АиФ-Самара» Ксения Железнова.
Полимерное бальзамирование, или вечный биоматериал
Евгений Петров, научный сотрудник центра «Полимерное бальзамирование в медицине» при СамГМУ предлагает посмотреть лабораторию, в котором проводятся научные эксперименты. По пути ученый рассказывает, что в России полимерным бальзамированием живых тканей занимаются только в Санкт-Петербурге и Самаре.
«Научные разработки полимерных материалов ведутся в Германии доктором и профессором Гюнтер фон Хагенсом. В России этим занимается анатом Иван Гайворонский. В стенах Самарского медицинского университета на кафедре оперативной хирургии и клинической анатомии с курсом инновационных технологий уже два года ведется поиск и улучшение способов полимерного бальзамирования. Такая методика позволяет сделать биологические препараты практически вечными для использования студентами и преподавателями».
Доктор показывает биологические ткани, которые законсервированы резиноподобными препаратами – силоксановыми полимерами. Он мнет их в руках, сгибает и разгибает обработанный биоматериал тонкой и слепой кишки. Ученый говорит, что органы желудочно-кишечного тракта хорошо подаются полимерному бальзамированию, а вот мышечная ткань гораздо сложнее.
Он мнет их в руках, сгибает и разгибает обработанный биоматериал тонкой и слепой кишки. Ученый говорит, что органы желудочно-кишечного тракта хорошо подаются полимерному бальзамированию, а вот мышечная ткань гораздо сложнее.
Затем Евгений Сергеевич показывает препарат коленного сустава – анатомический распил Пирогова, который тоже законсервирован на долгие годы полимерами.
«Посмотрите, это натуральные препараты, но они абсолютно без запаха. Ткани имеют свой естественный цвет, с ними приятно и просто работать преподавателям и студентам. Одно дело извлекать из емкости с формалином влажный, дурнопахнущий препарат, а другое дело брать в руки вот такие полимерные препараты».
Евгений Петров предлагает для сравнения посмотреть, как хранятся препараты «по старинке» в формалине и спирте. Такие методы хранения биоматериала, по мнению ученого, требуют больших усилий в уходе. Евгений Сергеевич открывает емкость, в которой находится препарат толстой кишки. Лабораторию мгновенно заполняет резкий запах формалина, глаза слезятся, начинается насморк.
Такие методы хранения биоматериала, по мнению ученого, требуют больших усилий в уходе. Евгений Сергеевич открывает емкость, в которой находится препарат толстой кишки. Лабораторию мгновенно заполняет резкий запах формалина, глаза слезятся, начинается насморк.
«Самарский медицинский университет получил два патента по полимерному бальзамированию. В планах у заведующего кафедрой оперативной хирургии и клинической анатомии доктора медицинских наук, профессора Александра Колсанова и его коллег законсервировать как можно больше биоматериалов полимерным бальзамированием».
IT-медицина
Евгений Петров закрывает лабораторию, и мы направляемся смотреть уже не биологические, а виртуальные препараты.
Тем временем, в компьютерном кабинете специалисты СамГМУ на больших сенсорных мониторах создают человека, прорисовывают органы, кости и нервную систему. После того, как специалисты завершат работу над 3D-моделированием анатомических объектов, созданные ими сцены окружения (то, над чем проводится операция) войдут в программу симуляторов хирургических операций – «Виртуальный хирург».
Инженер по компьютерной графике Андрей Скрябин работает над ногой человека. Он прорисовывает вены, сухожилия, мышцы, кости, лимфы и нервы. Специалист показывает, как работает «3-D атлас человека».
«Многим студентам, которые изучают анатомию человеческого тела иногда сложно представить в объеме то, что нарисовано в учебниках. Студенты могут посмотреть, как выглядит человеческое тело с разных сторон в 3D атласе и увидеть, как взаимодействуют между собой органы и мышцы».
3D атлас человека Фото: АиФ-Самара / Ксения Железнова«Это первый этап создания виртуального хирурга, давайте посмотрим, что из этого получается», – Евгений Петров открывает дверь следующего кабинета.
В аудитории, куда мы пришли, располагаются тренажеры по виртуальным операциям. Александр Воронин, кандидат медицинских наук, ассистент кафедры оперативной хирургии и клинической анатомии с курсом инновационных технологий СамГМУ проводит для меня обзорную экскурсию.
«Мы разработали линейку из трех симуляционных аппаратов, которые позволяют отрабатывать студенту практические навыки. Посмотрите, это первый кейс, на котором можно проводить операции по удалению желчного пузыря и удалению грыжевого выпячивания. Студент видит на экране компьютера абсолютно ту же картинку, которую видит врач в операционной».
Александр Воронин проводит виртуальную операцию Фото: АиФ-Самара / Ксения ЖелезноваУченый включает тренажер и приступает к операции. На мониторе видны человеческие органы, из колонок слышатся щелчки хирургических ножниц. На 3D изображении отображаются все сосуды, картинка двигается, воспроизводится человеческое дыхание. В это время ассистент кафедры оперативной хирургии и клинической анатомии движениями рук оперирует виртуального человека.
Александр Воронин заканчивает виртуальную операцию и показывает второй кейс – это тренажер для проведения операций на сосудах, кардиохирургическая практика, протезирование.
«Мы можем взять от реального человека данные и внедрить в систему «Виртуального хирурга». Студент, к примеру, берет анализы реального человека, загружает эти данные в тренажер и сначала тренируется на виртуальной модели, а потом может провести операцию на реальном человеке».
Третий кейс – симулятор по открытой хирургии, и здесь Самарский государственный медицинский университет – лидер. Александр говорит, что во всем мире такого симулятора нет.
«У нас есть подкрепленное патентом право, что мы создали первый в мире симулятор по открытой хирургии».
Урок анатомии
У студентов Самарского медицинского университета идет лекция по анатомии «Пищевод. Желудок. Кишечник».
Преподаватель показывает студентам натуральный экспонат, дает пояснения. «Посмотрите на мягкое и твердое небо. Вот это нижняя стенка полости рта и язык».
На партах у студентов лежат биологические материалы Фото: АиФ-Самара / Ксения ЖелезноваДля отработки практической части обучения, студенты отправляются в музей, где хранятся биологические препараты.
Экспонаты, представленные в музее, были собраны до принятия закона «О погребении и похоронном деле». Их хранит кандидат медицинских наук, старший преподаватель СамГМУ Евгений Бадалянц. Сначала идем в зал остеологии, где собраны все кости человека и животных для курса сравнительной анатомии.
Евгений Бадалянц кандидат медицинских наук, старший преподаватель СамГМУ в зале музея Фото: АиФ-Самара / Ксения ЖелезноваСтуденты ежедневно приходят в музей, закрепляют теоретические основы, наглядно изучая строение человека. В помещении находятся многочисленные скелеты взрослых людей и детей.
«Все экспонаты, представленные в музее, настоящие. Здесь только я один нарочный», – шутит ученый.
На одной из полок в емкостях находятся младенцы, которые появились на свет мертвыми в результате патологий. Ученый показывает, какими рождаются дети, у которых родители были наркоманами. Трудно смотреть на эти экспонаты, но будущие врачи должны быть готовы ко всему, считает Евгений.
«На протяжении четырнадцати лет я собирал экспонаты для музея анатомии. Студентам-медикам просто необходимо видеть биологический материал, изучать строение костей и мышц».
Звенит звонок, будущие врачи не спешат уходить из музея, рассматривают экспонаты и о чем-то спорят.
Мнение экспертов
Галина Суворова, профессор, заведующая кафедрой анатомии СамГМУ
Галина Суворова, заведующая кафедрой анатомии СамГМУ Фото: АиФ-Самара / Ксения Железнова«Долгое время существовал запрет на взятие трупного материала для обучения студентов-медиков. Мы использовали старые запасы препаратов, на которых и проводилось обучение. Сейчас у высших учебных заведений появилась законодательная возможность получать биологические ткани. Как только мы соберем необходимый пакет документов, трупный материал вновь поступит в Самарский медицинский университет.
В нашем университете есть высококачественные симуляционные препараты, которые потеют, дышат, у экспонатов даже может случиться инфаркт.
К примеру, на кафедре оперативной хирургии студенты сначала учатся шить на искусственных образцах, потом на трупах, а потом уже идут практиковаться в клиники.
Оптимально сейчас выстраивать обучение молодых врачей с использованием и биологического материала, и симуляторов».
Евгений Петров, доцент кафедры оперативной хирургии и клинической анатомии с курсом инновационных технологий СамГМУ, кандидат медицинских наук
Евгений Петров, доцент кафедры оперативной хирургии и клинической анатомии с курсом инновационных технологий Фото: АиФ-Самара / Ксения Железнова«Закон о погребении и похоронном деле ограничивает нас в возможности получения биоматериала. Мы испытываем недостаток в трупах, из которых можно готовить учебные полимерные препараты.
Сейчас появилась возможность консервировать биологические ткани резиноподобными препаратами – силоксановыми полимерами. Такая методика позволяет сделать биологические препараты практически вечными. На протяжении двадцати лет в Самарском медицинском университете работает секция практических медицинских навыков.
На протяжении двадцати лет в Самарском медицинском университете работает секция практических медицинских навыков.
Мы со студентами старших курсов ходим в областное бюро судебно-медицинской экспертизы, где будущим врачам предоставляется возможность работы над трупным материалом. Студенты отрабатывают технику наложения кишечного и сосудистого шва».
Александр Воронин, кандидат медицинских наук, ассистент кафедры оперативной хирургии и клинической анатомии с курсом инновационных технологий СамГМУ
Александр Воронин, ассистент кафедры оперативной хирургии и клинической анатомии с курсом инновационных технологий СамГМУ Фото: АиФ-Самара / Ксения Железнова«Сейчас основная проблема в медицинском образовании заключается в том, что объем теоретических знаний студентов не подкрепляется практической базой, хирургической практикой. Студент не может отрабатывать практические навыки на биологическом материале.
Чтобы получить биологический материал для обучения, нам необходимо собрать около десяти подписей, начиная от главы района, заканчивая прокурором. Перед тем, как биологические ткани попадут в учебное заведение, необходимо, чтобы они хранились определенный срок. В тканях начинаются такие изменения, которые крайне не желательны.
Перед тем, как биологические ткани попадут в учебное заведение, необходимо, чтобы они хранились определенный срок. В тканях начинаются такие изменения, которые крайне не желательны.
Это побудило нас к тому, что мы начали разрабатывать блок симуляционных методик, чтобы студенты отрабатывали практические навыки. Это виртуальные 3D тренажеры с ощущением обратной связи – «Виртуальный хирург», «3D атлас человека». Самарский государственный медицинский университет стал единственным из 130 российских вузов, который получил поддержку на создание центра прорывных технологий в сфере IT-медицины».
Стременная мышца — самая короткая в теле человека: описание и фото
Каждый человек когда-нибудь слышал что-то настолько громкое, что от этого звука даже начинала болеть голова. Может быть, это была пожарная сигнализация, сирена скорой помощи или даже плач ребенка. Совершенно неважно, откуда мог доноситься этот оглушающий звук: самым главным желанием любого слышащего его человека было прекратить это. В человеческом ухе есть маленькая мышца, которая называется стременной: именно она помогает ограничить вибрации в ухе, когда мы слышим очень громкие звуки. Кроме того, она имеет статус самой маленькой и короткой мышцы в теле человека.
В человеческом ухе есть маленькая мышца, которая называется стременной: именно она помогает ограничить вибрации в ухе, когда мы слышим очень громкие звуки. Кроме того, она имеет статус самой маленькой и короткой мышцы в теле человека.
Самая короткая и маленькая мышца человека: описание
У каждого человека более 600 мускулов, которые составляют порядка 40% от общей массы тела. Для людей, ведущих малоподвижный образ жизни и женщин, этот показатель немного ниже и составляет порядка 35% процентов. Для сравнения, кости в организме человека составляют лишь 14%.
Таким образом, все мускулы вместе весят больше, чем скелет. Каждая из них выполняет свою особенную работу и имеет неповторимые особенности:
- наиболее активными мускулами в теле человека являются глазные: они постоянно в движении, даже без тренировки;
- самая крупная мышца — большая ягодичная: а вот ее тренировками озабочено множество людей по всему миру;
- самая выносливая — сердечная;
- самая длинная — портняжная, находящаяся на передней части бедра;
- наиболее склонны к быстрому восстановлению трицепсы, а вот мышцы спины по этому показателю на последнем месте;
- жевательные — одни из самых сильных, чья сила давления может доходить до рекордных 100 кг даже без тренировок.

Стременная же является самой маленькой мышцей и относится к группе поперечно-полосатых. Поперечно-полосатая мускулатура состоит из множества полос мышечных волокон, которые также включают крупные мускулы ног и рук. Длина стременной составляет всего несколько миллиметров, в среднем — 6,3 мм. Площадь поперечного сечения тоже невелика: лишь 4,9 мм2. Главная задача самой короткой и маленькой мышцы — стабилизировать самую маленькую в теле кость: стремечко.
Самая маленькая и короткая из всех скелетных мышц человеческого тела расположена в среднем ухе, которое является открытой областью внутри каждого уха, прямо позади барабанной перепонки. Стременная заключена в конусообразную структуру в барабанной полости, к задней стенке которой самая маленькая мышца крепится стенками собственного канала. Барабанная полость — это открытое пространство, окружающее кости в среднем ухе.
Из этой конусообразной структуры в барабанной полости стременная переходит на костную ткань. Она представляет собой кость в среднем ухе — стремечко, которая помогает передавать звуковые колебания. Интересно, что стремечко — самая маленькая кость в организме человека.
Интересно, что стремечко — самая маленькая кость в организме человека.
Эволюция самой короткой и маленькой мышцы
Подобно кости, к которой она крепится, маленькая стременная мышца связана долгой историей эволюции с другими позвоночными:
- У млекопитающих она развилась из мускула, имевшегося до этого у амфибий, называемого депрессором челюсти, чья функция заключалась в том, чтобы открывать челюсть.
- Депрессор, в свою очередь, появился в результате подъема жаберной кости у костистых рыб и эквивалентен эпихиоиду акул.
- Если смотреть глобально, все они происходят от подъязычной дуги и иннервируются лицевым нервом.
Название самой маленькой и короткой мышцы также напрямую связано с самой маленькой костью, к которой она крепится. По форме стремечко напоминает типичную форму стремян, которые позволяют всадникам закреплять ноги вдоль корпуса лошади во время езды. Также из-за их формы две кости среднего уха, связанные со стременной, называются «молот» и «наковальня».
Как работает самая маленькая мышца
Когда самая короткая мышца в теле сокращается, она тянет за собой стремечко, тем самым регулируя звуковые колебания. Она получает электрические импульсы от головного мозга через черепной нерв VII, который также называют лицевым нервом VII. Когда звуковые волны попадают в ухо, самая маленькая кость начинает вибрировать и помогает проводить и передавать эти звуковые волны в мозг. Затем мозг преобразует эти вибрации в звуки, которые слышит человек.
Если проводить аналогию, стоит представить себе механизм с закрепленной цепью и двумя вертушками. По такой своеобразной цепи звуковая энергия от барабанной перепонки перенаправляется во внутреннее ухо. Там стимулируются сенсорные клетки, которые посылают то, что они слышат, в мозг по электрическим путям в качестве сигнала. Но если шум становится слишком громким в зоне перед барабанной перепонкой, примерно от 75 децибел (что соответствует громкому уличному шуму), самая маленькая в организме мышца напрягается и сжимается примерно на 7 миллиметров, натягивая стремя, и оно становится неподвижным. В подобной ситуации обычно подвижный орган становится жестким и хуже передает вибрации по цепочке: приглушается громкость.
В подобной ситуации обычно подвижный орган становится жестким и хуже передает вибрации по цепочке: приглушается громкость.
Функции самой маленькой мышцы в теле человека
Глубоко в ухе, скрытая костями, самая короткая мышца в теле выполняет свою невероятно важную работу в сочетании со стремечком, к которому прикрепляется. Вместе они обеспечивают человеку мир и покой: по крайней мере, слуховой. Барабанные и маленькие стременные мускулы являются защитными рефлексами. Они уменьшают объем шума, попадающего во внутреннее ухо. Работа одного из самых маленьких органов в теле чем-то похожа на рефлекс моргания и ситуацию, которая возникает, когда один глаз раздражен, а часто моргать начинают оба. Во-первых, даже если воздействие идет на одно ухо, втрое тоже задействовано. Во-вторых, как и моргание, эти защитные рефлексы могут стать более заметными, когда человек находится в состоянии стресса.
Интересно, что одной из ее главных задач является защита человека от шума его собственного голоса, когда он разговаривает. Для этого самая короткая мышца в организме просто приглушает тон, поэтому при разговоре мы слышим сами себя несколько иначе, чем все окружающие. Именно поэтому собственный голос в записи кажется нам чужим и странным: в этот момент маленькая стременная уже не работает, и мы слышим его без приглушения и дополнительных эффектов. С этим же связано, что слишком громко говорящие в повседневной жизни люди не замечают за собой этого недостатка: стоящий на страже маленький орган приглушает издаваемый самим человеком шум, и ему кажется, что все нормально.
Для этого самая короткая мышца в организме просто приглушает тон, поэтому при разговоре мы слышим сами себя несколько иначе, чем все окружающие. Именно поэтому собственный голос в записи кажется нам чужим и странным: в этот момент маленькая стременная уже не работает, и мы слышим его без приглушения и дополнительных эффектов. С этим же связано, что слишком громко говорящие в повседневной жизни люди не замечают за собой этого недостатка: стоящий на страже маленький орган приглушает издаваемый самим человеком шум, и ему кажется, что все нормально.
Также самая короткая мышца в теле может сокращать внутреннюю вокализацию для уменьшения самостимуляции, а также уменьшать силу громкости посредством модуляции сопротивления среднего уха: попросту говоря, защищать человека от внешних шумов.
Акустический рефлекс — важная для комфорта и нормальной жизнедеятельности функция
Тело имеет собственную встроенную систему защиты от действительно громких шумов. Эта защитная система включает в себя самую маленькую мышцу в теле и называется акустический рефлекс. Когда в ухо поступают очень громкие звуки, они вызывают очень сильные вибрации в ухе. Чем сильнее давление из вне, тем интенсивнее вибрация. Таким образом, очень громкие шумы будут вызывать значительную вибрацию кости.
Когда в ухо поступают очень громкие звуки, они вызывают очень сильные вибрации в ухе. Чем сильнее давление из вне, тем интенсивнее вибрация. Таким образом, очень громкие шумы будут вызывать значительную вибрацию кости.
Однако, когда громкие звуки проникают в ухо, акустический рефлекс заставляет маленькую стременную сокращаться и стабилизировать костную ткань, сводя к минимуму уровень вибрации и, таким образом, уменьшая предельную громкость. Без этого рефлекса шумы стали бы почти оглушительными для человека и потенциально вредными для слуха.
Нервом, который соединен с самой короткой мышцей, является лицевой нерв, который является седьмым черепным нервом (CN VII). Акустический рефлекс зависит от этого лицевого нерва. Когда в ухо проникают громкие звуки, мозг через лицевой нерв посылает сообщение стременной, в котором говорится, что маленькая мышца должна сократиться и стабилизировать костную ткань.
Болезни самой короткой мышцы в теле человека
Сокращения маленькой стременной обычно называют трепетанием. Если трепетание связано с лицевыми движениями, это говорит о неправильной работе органа и возникает чаще всего при наличии неврологических заболеваний. Например, такой эффект наблюдается после выздоровления от паралича Белла, или одностороннего лицевого паралича. Когда пораженная сторона лица сокращается, стременная также сокращается (синкинез) из-за аберрантной регенерации лицевого нерва. Говоря простым языком, это абсолютно произвольное и случайное присоединение восстанавливающихся ветвей нервов к не своим мимическим мускулам.
Если трепетание связано с лицевыми движениями, это говорит о неправильной работе органа и возникает чаще всего при наличии неврологических заболеваний. Например, такой эффект наблюдается после выздоровления от паралича Белла, или одностороннего лицевого паралича. Когда пораженная сторона лица сокращается, стременная также сокращается (синкинез) из-за аберрантной регенерации лицевого нерва. Говоря простым языком, это абсолютно произвольное и случайное присоединение восстанавливающихся ветвей нервов к не своим мимическим мускулам.
Могут наблюдаться отклонения в характере измерений акустического рефлекса, соответствующие характеристикам шума в ушах пациента. Нормальный акустический рефлекс у человека возникает только для звуков высокой интенсивности (> 75-85 дБ). Если при шумах высокой интенсивности такой эффект не наблюдается, это обычно говорит о дисфункции самой короткой мышцы в теле и может быть вылечено путем ее высвобождения.
Если самая короткая мышца в теле парализована, человек слышит все, даже тихие или обычные повседневные звуки с большей интенсивностью, ощущая их, как неприятное звяканье. Причиной такого паралича часто является повреждение лицевого нерва, от которого орган обычно получает свои команды. Например, нерв может быть разорван переломом черепа или заражен вирусом герпеса.
Причиной такого паралича часто является повреждение лицевого нерва, от которого орган обычно получает свои команды. Например, нерв может быть разорван переломом черепа или заражен вирусом герпеса.
Еще одним достаточно распространенным заболеванием, связанным с дисфункцией самой короткой мышцы в теле, выступает гиперакузия. Это крайне ослабляющий ее подвижность слуховой синдром, который превращает повседневные звуки окружающей среды в болезненные.
Человек, страдающий гиперакузией, замечает, что привычные звуки становятся болезненными и даже невыносимыми. Это приводит к сокращению личной функциональности или неспособности выполнять повседневные задачи, в зависимости от тяжести течения заболевания у человека: от посещения концертов в более легких случаях, до шелеста листьев в самых тяжелых.
Лечение заключается в использовании звуковых генераторов или, например, прослушивании звукозаписей. В частности, в числе последних могут выступать комбинации звуков в широких полосах, например, белый шум. Первоначально, почти неслышимые уровни используются ежедневно и в течение длительного периода времени. Затем они постепенно увеличиваются, пока слух не потеряет чувствительность и не приобретет допустимую толерантность к звуку. Лечение стресса и расстройств сна также помогает улучшить способность справляться с гиперакузией.
Первоначально, почти неслышимые уровни используются ежедневно и в течение длительного периода времени. Затем они постепенно увеличиваются, пока слух не потеряет чувствительность и не приобретет допустимую толерантность к звуку. Лечение стресса и расстройств сна также помогает улучшить способность справляться с гиперакузией.
Не так сильно распространена, однако не менее неприятна миокимия этого маленького мускула. Она проявляется как звон в ушах, вызванный синхронным повторяющимся сокращением мускулов среднего уха. Звон в ушах обычно характеризуется как щелчок, предположительно возникающий из-за движения барабанной перепонки, или жужжание, предположительно из-за движения мускулатуры. Однако он также был описан как пульсация, постукивание, треск, лопанье пузырьков, тиканье, трепетание бабочки, свист или удары хлыста.
Видео
5 наиболее распространенных проблем осанки (и как их решить)
Эту статью написал Марк Перри — создатель программы BuiltLean и главный редактор и продюсер блога BuiltLean. В ней достаточно емко и коротко отражены современные тенденции заболеваний от сидячего образа жизни. Статья для Вас переведена в Эрготронике.
В ней достаточно емко и коротко отражены современные тенденции заболеваний от сидячего образа жизни. Статья для Вас переведена в Эрготронике.
На прошлой неделе я побывал на семинаре «Анализ осанки и движения» в Нью-Йорке, который длился один день. На нем присутствовали около 25 профессионалов в фитнесе, которые занимаются им постоянно. После того как мы изучили осанку друг друга, мы были в шоке, узнав, что КАЖДЫЙ имел проблему с ней — от умеренной до тяжелой.
Вполне возможно, что у вас проблема осанки от умеренной до тяжелой, что, в свою очередь может повлиять на ваше здоровье и работоспособность. Должен признать, что у меня самого степень составляет от низкой до умеренной!
В этой статье не дается описание того как провести полную оценку осанки; здесь я описываю 5 самых распространенных проблем осанки, которые, как я считаю, вызваны малоподвижным образом жизни. Для каждой проблемы осанки, я называю (1) способ проверки, (2) причину, (3) последствие, и (4) решение.
Имейте в виду, что те варианты, которые тут приведены, являются только «возможными» решениями, так как некоторые из причин и проблем не могут быть исправлены (т.е. врожденные проблемы, или травмы). Наконец, помните, что любая проблема осанки может привести к другим проблемам, так как тело человека — это единая «кинетическая цепь».
Проблема нарушения осанки #1: Гиперпронация
Как проверить – Как указано на фото выше, обе руки положите на расстоянии 1 дюйм от каждой стороны вашей ноги. Выпрямите щиколотку так, чтобы пространство между каждой рукой и щиколоткой было одинаковым. Теперь ослабьте щиколотку и стопу. Если они подались вовнутрь, следовательно у вас гиперпронация стопы.
Причина – ожирение, беременность, неудобная обувь, или постоянное переступание по твердой поверхности — всё это может ослабить свод стопы, что в свою очередь ведет к косолапию и зачастую к плоскостопию.
Последствие – Косолапие создает дополнительную нагрузку на стопы, зажимает мышцы голени и повреждает коленные чашечки. Косолапие часто приводит к подошвенному фасциту, пяточной шпоре, метатарзалгии, тендиниту и / или шишке на ноге. Целых 20-30% американцев имеют плоскостопие, или косолапие.
Косолапие часто приводит к подошвенному фасциту, пяточной шпоре, метатарзалгии, тендиниту и / или шишке на ноге. Целых 20-30% американцев имеют плоскостопие, или косолапие.
Решение – Если свод уже нарушен, лучшее решение – обратиться к ортопеду и подобрать спецобувь и лечение. Если свод только начинает искривляться, укрепить его поможет бег/ходьба босиком. Только предварительно проконсультируйтесь с врачом (один из лучших выходов — обратиться к ортопеду). Про бег босиком см. Barefoot Running Research: Benefits of Barefoot Running
Проблема нарушения осанки #2: Наклон бедра вперед
Как проверить – Один из самых полезных методов — прогиб таза вперед насколько это возможно, а затем так же назад. Прогнуть таз вперед вы можете по максимуму.
Причина – Сидячий образ жизни, отсутствие упражнений для растягивания туловища, что сокращает сгибающие мышцы бедер.
Последствие – Наклоненный таз (оно же «оттопыренная попа») ассоциируется с лишенными гибкости сгибателями бедра, т.е. группой мышц на передней части бёдер, которые тянут колено вверх. Когда вы ходите, жесткие сгибатели бедра не дают включаться ягодичным мышцам, что в свою очередь делает мышцы задней поверхности бедра перегруженными и чрезмерно жесткими. Если у вас перенапряжены мышцы задней поверхности бедра, причиной этого могут быть негибкие сгибатели бедра и наклоненный таз.
Решение – Растягивайте сгибатели бедра с помощью статических выпадов, например, поза лунного серпа, активируйте ягодицы с помощью специальных упражнений и растягивайте мышцы задней поверхности бедра.
Проблема нарушения осанки #3: Искривление верхней части спины/Кифоз
Как проверить – Попросите кого-нибудь сфотографировать вас в профиль. Если верхняя часть спины округлена больше чем на 40-45 градусов, как показано на рисунке справа — это кифоз.
Причина – Сидячий образ жизни, например работа в офисе за компьютером.
Последствие – Сидя сгорбившись за компьютером, мы зажимаем мышцы груди, что может привести к искривлению (кифозу) верхней части спины. Мышцы, отвечающие за осанку в верхнем отделе спины, становятся слабыми.
Решение – Снять зажим можно, массируя область груди теннисным мячиком и выполняя другие упражнения. Мое любимое упражнение — перекатывание по верхней части спины.
Проблема нарушения осанки #4: Сутулость
Как проверить – Можно провести “Тест с карандашом”. Возьмите в каждую руку карандаш или ручку. Как показано на рисунке, если они указывают прямо, с осанкой все в порядке. Если они смотрят друг на друга или повернуты под углом к телу, это говорит о сутулости.
Причина – Сидячий образ жизни, например работа в офисе за компьютером, и неправильное выполнение силовых упражнений с чрезмерной нагрузкой на грудные мышцы.
Последствие – Сидя сгорбившись за компьютером, мы зажимаем мышцы груди, плечи искривляются, что приводит к сутулости. Мышцы, отвечающие за осанку в верхнем отделе спины, ослабевают.
Решение – Очень похоже на исправление искривления – ослабьте стеснение в груди путём миофасциального расслабления (можно использовать мяч для массажа) и растяжения, а также укрепления мышц верхней части спины.
Проблема нарушения осанки #5: Наклон головы вперед
Как проверить – Попросите кого-нибудь сфотографировать вас в профиль. Как показано на рисунке справа, найдите у себя на плече выступающую косточку — акромиально-ключичное сочленение, и проверьте находится ли она под мочкой уха. Если мочка уха выступает вперед, следовательно у вас проблема с осанкой.
Причина — сидение сгорбившись перед экраном компьютера.
Последствие — мышцы задней части шеи и верхнего отдела спины сильно зажимаются.
Решение — во-первых, старайтесь сидеть правильно: спина прямая, голова чуть отклонена назад, взгляд прямо перед собой. Постарайтесь не поднимать голову вверх откидывая ее назад. Во-вторых, регулярно массируйте верхнюю часть спины (можно использовать массажер), чтобы расслабить мышцы шеи.
Эти пять описанных проблем лежат лишь на поверхности столь важной темы, и надеюсь благодаря этому вы задумаетесь о своей осанке.
Анатомия лёгких, строение, функции на ONKO.LV
Лёгкие – это мягкий, губчатый, конусообразный парный орган. Лёгкие обеспечивают дыхание — обмен углекислого газа и кислорода. Так как лёгкие являются внутренней средой организма, которая постоянно соприкасается с внешней средой, они имеют хорошо приспособленное и специализированное строение не только для газообмена, но и для защиты – в дыхательных путях задерживаются и выводятся наружу различные вдыхаемые инфекционные возбудители, пыль и дым. Правое лёгкое образуют три доли, а левое — две. Воздух в лёгкие попадает через носовую полость, горло, гортань и трахею. Трахея разделяется на два главных бронха – правый и левый. Главные бронхи разделяются на более мелкие и образуют бронхиальное дерево. Каждая веточка этого дерева отвечает за небольшую ограниченную часть лёгкого – сегмент. Более мелкие веточки бронхов, которые называются бронхиолами, переходят в альвеолы, в которых происходит обмен кислорода и углекислого газа. В лёгких нет мышц, поэтому они не могут расправляться и сокращаться самостоятельно, но их структура позволяет следовать дыхательным движениям, которые совершают межрёберные мышцы и диафрагма.
Чтобы облегчить движения лёгких, их окружает плевра – оболочка, которая состоит из двух листков – висцеральной и париетальной плевры.
Париетальная плевра присоединяется к стенке грудной клетки. Висцеральная плевра присоединяется к наружней поверхности каждого лёгкого. Между двумя плевральными листками образуется небольшое пространство, которое называется плевральной полостью. В плевральной полости находится небольшое количество водянистой жидкости, которая называется плевральной жидкостью. Она предотвращает трение и держит вместе плевральные поверхности во время вдоха и выдоха.
Строение клеток глубоких дыхательных путей достаточно специализировано и хорошо приспособлено для дыхания. Все дыхательные пути выстланы эпителием, который является специально приспособленными клетками, чтобы выполнять много важных функций:
- защитную;
- секрецию слизи;
- выведение раздражающих веществ;
- начало иммунных реакций.
Вид эпителия отличается в разных частях дыхательных путей. Большую часть слизистой дыхательных путей образует реснитчатый эпителий. Эти клетки – расположены вертикально в один слой с ресничками, направленными в сторону дыхательных путей. Реснички всегда движутся в направлении наружу. Слизистую более мелких дыхательных путей образует эпителий без ресничек.
В эпителии дыхательных путей находятся железы – бокаловидные клетки. Это специализированные клетки, которые производят и выделяют слизь. Слизь, продуцируемая этими клетками необходима, чтобы увлажнять поверхность эпителия и механически защищать слизистую.
Слизь является липкой, поэтому к ней прилипают вдыхаемые микроскопические инородные тела, и потом они выводятся наружу при помощи реснитчатого эпителия.
Артроскопия коленного сустава. Разрыв ПКС
Анатомия
Коленный сустав – не только самый крупный в организме человека, но и самый сложный. Он образован тремя костями: бедренной костью сверху, большеберцовой костью снизу, а спереди от этих костей располагается надколенник (коленная чашечка). Как бедренная, так и большеберцовая кость имеют по два расширяющихся костных выступа – мыщелка: наружный и внутренний. Наружный мыщелок еще называют латеральным (от латинского слова lateralis – наружный), а внутренний – медиальным (от латинского слова medialis — внутренний). Основное движение коленного сустава – это сгибание, при этом надколенник ложится в специальную борозду между наружным и внутренним мыщелками бедренной кости.
Контактирующие поверхности бедренной, большеберцовой костей и надколенника покрыты гладким хрящом, который облегчает скольжение.
Между бедренной и большеберцовой костью есть мениски – хрящевые прослойки полулунной формы, которые увеличивают стабильность сустава, повышая площадь контакта костей, работают как амортизаторы и выполняют некоторые другие важные функции.
Стабильность костей друг относительно друга обеспечивается за счет связок коленного сустава. К наиболее важным связкам коленного сустава относятся:
- Крестообразные связки, которые удерживают голень от смещения кпереди (передняя крестообразная связка) и кзади (задняя крестообразная связка).
- Большеберцовая коллатеральная связка (внутренняя боковая связка), которая удерживает голень от отклонения кнаружи.
- Малоберцовая коллатеральная связка (наружная боковая связка), которая удерживает голень от отклонения кнутри.
Передняя крестообразная связка находится в самом центре коленного сустава, сверху прикрепляется к наружному мыщелку бедренной кости, после чего идет вниз и немного кнаружи и прикрепляется к углублению на большеберцовой кости (переднее межмыщелковое поле), где часть ее волокон соединяются с менисками. Перпендикулярно передней крестообразной связке кзади от нее располагается задняя крестообразная связка, и если посмотреть на эти связки спереди, то можно увидеть, как они образуют крест, что и дало этим связкам такое название — крестообразные связки. Средняя длина передней крестообразной связки — 3 сантиметра, а ширина — 7-12 мм.
Передняя крестообразная связка, как и другие связки, в основном состоит из прочных коллагеновых волокон, которые практически не растягиваются. Эти волокна внутри связки закручены по спирали под углом в 110 градусов. Исходя из мест прикрепления связки и ее хода (сверху вниз, спереди назад и снаружи кнутри) становиться понятна ее роль: передняя крестообразная связка удерживает голень от смещения кпереди и кнутри.
Анатомия связок коленного сустава: ПКС — передняя крестообразная связка, ЗКС -задняя крестообразная связка
В составе передней крестообразной связки выделяют два пучка: передне-внутренний и задне-наружный. Такая двухпучковая структура передней крестообразной связки обеспечивает лучшую стабильность в коленном суставе при разных углах сгибания. Некоторые ученые даже выделяют третий — промежуточный пучок.
Передне-внутренний пучок в полтора раза длиннее задне-наружного (в среднем 37,7 мм против 20,7 мм) и шире (8,5 мм и 7,7 мм). При работе сустава эти два пучка находятся в сложном взаимодействии. При разогнутом колене они почти параллельны друг другу, если смотреть на них сбоку и перекрещиваются, если смотреть на них спереди. Если колено согнуть, то передне-внутренний пучок удлинняется, а задне-наружный — укорачивается.
Передняя крестообразная связка коленного сустава: двухпучковая структура. ПВ — передне-внутренний пучок (синяя линия), ЗН — задне-наружный (зеленая линия). Слева — вид сбоку, колено разогнуто; в центре — вид сбоку, колено согнуто; справа — вид спереди, колено разогнуто
Помимо того, что передняя крестообразная связка коленного сустава выполняет стабилизационную функцию (удерживает голень от смещения вперед и кнутри), в ней есть еще и нервные окончания, которые сигнализируют о том, в камо положении, согнутом или разогнутом находится коленный сустав.
Передняя крестообразная связка практически не имеет кровеносных сосудов.
Причины разрыва передней крестообразной связки и причины
Растяжения и повреждения передней крестообразной связки являются очень распространенной травмой. Например в США ежегодно диагностируется около 200 тысяч разрывов передней крестообразной связки, из них в 100 тысячах случаев выполняется операция по реконструкции (пластике) передней крестообразной связки. Среди всех других связок колена передняя крестообразная травмируется чаще всего, например, разрывы передней крестообразной связки происходят в 15-30 раз чаще, чем разрывы задней крестообразной связки. В спорте разрыв передней крестообразной связки у женщин происходит в 4-10 раз чаще, чем мужчины.
Исходя из функции, которую выполняет передняя крестообразная связка (удерживание голени от смещения вперед и кнутри), становится понятным и механиз травмы, при котором происходит растяжение или разрыв передней крестообразной связки. Как правило кручение на порной ноге, когда корпус с бедром вращается наружу, а голень со стопой остаются на месте.
Однако на самом деле механизм и причины разрыва передней крестообразной связки сложнее. Принципиально в качестве причин разрыва передней крестообразной связки можно выделить прямую травму (контактный механизм: удар по голени, бедру) и непрямую травму (неконтактный механизм: кручение на ноге при резком торможении, приземление после прыжка и т.д.).
Отклонение голени кнаружи и кручение бедра кнутри. Этот механизм разрыва передней крестообразной связки самый распространенный. Часто такой разрыв передней крестообразной связки происходит в гандболе, баскетболе, футболе и волейболе, когда при беге нужно резко развернуться или при приземлении после прыжка с разворотом корпуса кнутри от опорной ноги. При таком механизме травмы может произойти и разрыв внутреннего мениска. Если такое движение будет очень сильным, то возможен разрыв трех структур: передней крестообразной связки, внутреннего мениска и внутренней боковой связки. Такое повреждение коленного сустава еще называют «несчастливой триадой» или «взрывом коленного сустава». Это название прижилось по имени хирурга O’Donoghue, которые его впервые описал в 1950 году.
«Несчастливая триада»: при сильном отклонении голени кнаружи и вращении бедра кнутри возможен разрыв трех структур: передней крестообразной связки (1), внутреннего мениска (2) и внутренней боковой связки (3)
Отклонение голени кнутри и кручение бедра кнаружи. Этот механизм разрыва передней крестообразной связки коленного сустава противоположен предыдущему, при этом тоже возможен разрыв мениска, но наружного.
Фантом-стопа. Этот механизм разрыва возможен при падении с горных лыж. Например, при падении назад правая лыжа задирается вверх и только ее задний конец соприкасается со снегом. Как только лыжа входит в снег (край лыжи и олицетворяет «призрачную стопу»), она делает поворот и обусловливает внешнее вращение голени (пронация). Коленный сустав при этом согнут под прямым углом. Этот механизм может привести и к ихолировнному повреждению (полному или частичному разрыву) передней крестообразной связки без повреждения менисков и других структур (задне-латерального угла).
Механизм, обусловленный лыжным ботинком. Чаще всего разрывы передней крестообразной связки по этому механизму происходят у горнолыжников, которые носят более высокие и жесткие ботинки. Например, при падении назад верхняя часть ботинка передает нагрузку на верхнюю часть голени большеберцовой кости по типу «выдвижного ящика». Бедро кость смещается назад, а голень удерживается задним краем высокого ботинка. Передняя крестообразная связка натягивается и рвется. Фиксированный наклон вперед во всех современных лыжных ботинках способствует такому типу разрыва.
Контактные механизмы. Разрыв передней крестообразной связки может произойти в результате прямого удара по колену, бедру или голени. При этом может произойти отклонение голени кнаружи, кнутри или кпереди. Разрыв передней крестообразной связки может произойти и в результате переразгибания колена (гиперэкстензии). Гиперэкстензия может быть вызвана как прямым ударом по колену спереди, так и ударом по нижней части голени. Кроме того разрыв передней крестообразной связки при сильном ударе ко верхней части голени сзади. Контактные механизмы травмы встречаются редко.
Факторы, которые способствуют разрыву передней крестообразной связки
Угол между бедром и голенью. Если смотреть на скелет спереди, то можно увидеть как бедро соединяется с голенью под углом, который называют углом квадрицепса или Q-углом. Размер угла Q определяется шириной таза. У женщин таз более широкий, чем у мужчин, поэтому у женщин и Q-угол больше, чем у мужчин. Большой угол Q приводит к тому, что при отклонении голени кнаружи нагрузка на переднюю крестообразную связку больше и поэтому она легче рвется.
Q-угол у мужчин и женщин
Ширина межмыщелковой вырезки (Notch-вырезка). Передняя крестообразная связка находится в коленном суставе в межмыщелковой вырезке. У женщин она уже, чем у мужчин и при движениях в коленном суставе пространство вокруг передней крестообразной связки меньше, и она может даже тереться о край наружного мыщелка бедренной костичто способствует разрыву. При вращении голени с распрямлением в колене узкая межмыщелковая вырезка может зажать переднюю крестообразную связку и разорвать ее.
Межмыщелковая вырезка (обведена красным цветом) у женщин уже, чем у мужчин, что способствует разрыву передней крестообразной связки
Сила мышц бедра. Помимо связок важную роль в стабильности коленного сустава играют мышцы бедра, которые своими сухожилиями крепятся к большеберцовой кости голени. У женщин мышцы бедра слабее чем у мужчин, и поэтому в обеспечении стабильности коленного сустава большая нагрузка приходится на переднюю крестообразную связку.
Согласованность мышц бедра. Передние мышцы бедра (четырехглавая мышца бедра или квадрицепс) и задние мышцы бедра у женщин работают по другому, чем у мужчин. При согнутом колене женский квадрицепс сокращается сильнее, что выдвигает голень вперед, натягивая переднюю крестообразную связку. В тоже время задние мышцы бедра, которые препятствуют сдвиганию голени вперед, реагируют медленнее, чем у мужчин. Тем самым создаются предпосылки для разрыва передней крестообразной связки.
Гормональный профиль. Возможно, что разрыву передней крестообразной связки способствуют некоторые гормоны (эстроген и прогестерон). Некоторые ученые считают, что они уменьшают и прочность вех связок, не только передней крестообразной, и их эластичность. Эта эластичность (связки способны растягиваться на 4-5% от своей длины, не разрываясь при этом) вообще защищает от многих травм, позволяя поглотить больше энергии, прежде чем порвутся связки.
Все эти факторы в комплексе объясняют большую вероятность разрыва передней крестообразной связки у женщин, однако каждый из этих фактором может работать и у мужчин? например, у мужчин тоже бывает увеличенный Q-угол или узкая межмыщелковая вырезка.
Профилактика
В спорте разрывы передней крестообразной связки чаще встречаются у женщин, но в обычной жизни они встречаются одинаково часто как у мужчин, так и у женщин. Профилактика разрывов передней крестообразной связки касается, в основном, только спорта, и включает в себя обязательные разминки, укрепляющие и проприоцептивные тренировки, а также изменение техники бега, прыжков и рывковых движений. К сожалению, в настоящее время общепринятой программы профилактики травм передней крестообразной связки не существует.
Какие бывают повреждения передней крестообразной связки?
Как мы уже упоминали, передняя крестообразная связка до определенного предела может растягиваться (на 4-5% от своей длины). Если сила будет большей, то может произойти частичный разрыв связки (микроразрыв отдельных волокон), а если сила будет еще большей, то произойдет частичный разрыв, и, наконец, если сила будет еще увеличиваться, то произойдет полный разрыв передней крестообразной связки.
|
Классификация повреждений передней крестообразной свзки американской медицинской ассоциации спотивных травм (American Medical Association for Athletic Injuries) |
|
|
I степень: |
малые растяжения передней крестообразной связки (микроразрывы). Характеризуются болью, умеренным ограничением движений в суставе, умеренным отеком коленного сустава, сохранением стабильности сустава |
|
II степень: |
умеренные растяжения связки (частичные разрывы). Имеют такие же признаки, что и малые растяжения, но эта травма склонна многократно повторяться, так как после частичного разрыва связка становится менее прочной и ее повторная травма может произойти гораздо легче |
|
III степень: |
полные разрывы. Выраженная боль, отек, ограничение движений, часто нарушение опороспособности, нестабильность коленного сустава. |
Однако на самом деле вариантов повреждений передней крестообразной связки больше. Например, как мы уже упоминали, у передней крестообразной связки есть два пучка. Бывают разрывы одного из пучков (передне-внутреннего или задне-наружного). При этом оторвавшийся пучок передней крестообразной связки может болтаться в коленном суставе и блокировать движения, давая картину, похожую на блок коленного сустава при разрыве мениска.
Кроме того, возможен и так называемый перелом Сегонда. Иногда его называют отрывным переломом межмыщелкового возвышения.
Перелом Сегонда (отрыв передней крестообразной связки от места прикрепления к большеберцовой кости с костным блоком). На рентгенограмме справа показан не только перелом Сегонда в месте прикрепления передней крестообразной связки (синяя стрелка), но и в месте прикрепления наружной боковой связки (рыжая стрелка)
Кроме того, как мы мы уже отмечали, разрыв передней крестообразной связки может сочетаться с разрывами менисков и других связок коленного сустава (задней крестообразной, наружной боковой, внутренней боковой).
Симптомы разрыва передней крестообразной связки
Повреждению передней крестообразной связки обычно предшествует травма коленного сустава, сразу после которой возникает боль и сильный отек колена. При разрыве передней крестообразной связки часто слышно треск, однако это неспецифический признак, который бывает и при разрывах других связок коленного сустава. Кроме того, в момент травмы возможно ощущение «вывихивания» голени кпереди или вбок. В таких случаях нужно обратиться к врачу.
Прежде всего врач-травматолог выяснит механизм травмы, который поможет заподозрить не только повреждение передней крестообразной связки, но и других структур (например, задней крестообразной связки, наружной и внутренней боковых коллатеральных связок).
При разрыве передней крестообразной связки в полость сустава попадает кровь — такое состояние называют гемартрозом. В первые несколько дней после травмы этот гемартроз и боль могут быть настолько выражены, что полноценное обследование сустава руками врача невозможно, а именно благодаря обследованию руками врача и может быть поставлен диагноз разрыва передней крестообразной связки. Такое обследование может быть выполнено позже, когда острая боль и гемартроз пройдут. Как правило к этому моменту и обнаруживается нестабильность коленного сустава или ощущение «провала» в нем. Это происходит ввиду того, что разованная передняя крестообразная связка не удерживает голень от смещения кпереди и кнутри (антеромедиальная или передне-внутренняя нестабильность голени в коленном суставе).
Диагноз разрыва передней крестообразной связки
После того, как врач тщательно ознакомиться с механизмом травмы, он начнет тестировать Ваши коленные суставы. В первую очередь обследуют здоровое колено, чтобы ознакомить пациента с техникой осмотра, а так же для того, чтобы имелась возможность сравнить результаты тестов на здоровой и больной ноге. Для выявления передней нестабильности голени в коленном суставе существует ряд специальных тестов. Принцип этих тестов состоит в том, что врач-травматолог провоцирует голень смещаться кпереди, и, если передняя крестообразная связка разорвана и не выполняет своей функции, то голень будет поддаваться и смещаться. Существует три основных теста, которые определяют передне-внутреннюю нестабильность голени в коленном суставе: тест переднего выдвижного ящика, тест «pivot shift» и тест Лахмана (Lachman).
Тест переднего выдвижного ящика. Нога согнута в колене под прямым углом, врач тащит голень на себя, оценивая ее смещение в сравнении со здоровым коленом. Дополнительно тест выполняется с поворотом стопы внутрь и наружу
Если тест переднего выдвижного ящика дал сомнительные результаты, то для более точного измерения нестабильности голени в коленном суставе используется специальный прибор — артрометр, который позволяет оценить подвижность голени с точностью до миллиметра
Pivot shift тест или тест Jerk
Тест Лахмана (Lachman). Пробу проводят при согнутом под углом 20—30 градусов колене. Оценивают степень смещения голени вперед, а также ощущения в момент остановки. Недостаточность функции переднецй крестообразной связк4и или переднюю нестабильность голени разделяют на три степени, основываясь на сравнении со здоровой ногой. Первой степени соответствует увеличение смещения на 1—5 мм (т.е. подвижность голени больной ноги на 1-5 мм больше, чем на здоровой), второй — на 6—10 мм, третьей — более чем на 10 мм.
Как мы уже отмечали, в первые несколько дней после травмы боль в колене и гемартроз делают такое обследование невозможным, но, если сразу после травмы не упустить время, то есть провести осмотр до того, как разовьется отек и больной начнет инстинктивно сопротивляться манипуляциям врачато, то эти тесты воможны. Уменьшить боль и улучшить качество обследования можно, аспирировав (удалив шприцом) из сустава кровь.
При тщательном сборе анамнеза и внимательном осмотре, тестировании сустава разрыв передней крестообразной связки можно диагностировать без дополнительных методов исследования. Но, так как необходимо исключить и другие травмы (переломы мыщелков большеберцовой и мыщелков бедренной костей, переломы надколенника, перелом Сегонда, разрывы менисков, боковых связок и др.), врач использует и другие, инструментальные методы обследования (рентгенографию, магнитно-резонансную томографию, УЗИ).
Лучевая диагностика. Для исключения переломов выполняют рентгенографию коленного сустава в стандартных проекциях. Перелом Сегонда, упомянутый выше, представляет собой отрывной перелом участка мыщелка большеберцовой кости. Чаще этот перелом встречается у детей и подростков. Следующий по ценности метод для обнаружения сопутствующих травм — магнитно-резонансная томография. Точность ее для диагностики разрыва передней крестообразной связки составляет 95% или более, но обычно использование магнитно-резонансной томографии (МРТ) является подстраховкой, поскольку диагноз часто очевиден при обследовании руками. Более того, МРТ может быть даже вредным, поскольку исследование может показать «страшный» разрыв связки, а на самом деле он не будет давать нестабильности — и в такой ситуации врач и пациент могут склониться к ненужной операции. Среди ортопедов, травматологов, спортивных врачей есть даже специальное обозначение таких случаев — VOMIT, что является англоязычной аббревиатурой victim of modern imaging techniques (жертва современных изуализирующих методов обследования, к которым относится и МРТ).
В 80% случаев на магнитно-резонансных томограммах обнаруживают также поднадкостничные гематомы в области наружного мыщелка бедренной кости и наружной части большеберцовой кости.
Магнитно-резонансная томография коленного сустава. Слева — нормальная передняя крестообразная связка. На снимке она выглядит как равномерный темный тяж. Справа — разрыв передней крестообразной связки в ее верхней части. Связка на снимке не целая и светлая.
Лечение
Разрыв передней крестообразной связки совершенно не означает, что единственным возможным методом лечения такой травмы будет операция. Показанием к операции является не сам факт разрыва передней крестообразной связки, а развившаяся вследствие разрыва передне-внутренняя нестабильность голени в коленном суставе. Частичные разрывы передней крестообразной связки в большинстве случаев не приводят к нестабильности голени, поскольку оставшаяся часть связки может вполне успешно стабилизировать голень в коленном суставе. Полные разрывы также не всегда приводят к нестабильности. Например, оторвавшаяся передняя крестообразная связка может подпаяться к задней крестообразной связке и при неспротивных нагрузках в таком случае нестабильности не будет, и, соотеветственно, такое состояние не будет требовать операции. Стоит еще раз отметить, что передняя крестообразная связка хоь и главный стабилизатор, но не единственный (существуют и другие связки, мышцы, капсула сустава), и даже при полном разрыве оставшиеся неповрежденными структуры могут обеспечивать стабильность при определенном уровне физических нагрузок.
Прежде чем мы поговорим о лечении, стоит разделить повреждения и разрывы передней крестообразной связки по давности. Можно выделить свежие повреждения, когда после травмы колена прошло несколько дней, в колене имеется кровь (гемартроз) и еще ярко выражена боль. Несвежие случаи (до 3-5 недель), когда отек спадает, но связка еще не срослась (имеется ввиду возможность «подпаивания» передней крестообразной связки к задней или срастание частичного микроразрыва). И, наконец, спустя 3-5 недель после травмы можно выделить период, когда все нарушения функции сустава обусловлены только недостаточностью поврежденной передней крестообразной связки, а не болью или отеком острой травмы.
В остром периоде, когда повреждение (растяжение, частичный или полный разрыв) передней крестообразной связки свежее, лечение направлено на снятие боли и отека (гемартроза) коленного сустава. Сразу после травмы не пытайтесь передвигаться без посторонней помощи, желательно вообще не наступать на травмированную ногу — ведь нагрузка может усугубить внутрисуставные повреждения. Сразу после травмы (и в первые 2-3 суток) нужно прикладывать холод, применять противовоспалительные препараты (обезбаливающие таблетки, капсулы). Важен покой для коленного сустава — который подразумевает ограничение и осевой нагрузки на ногу (нельзя наступать на ногу или можно наступать с частичной опорой), и амплитуды движений. Для ограничения амплиитуды движений может использоваться гипсовая лонгета, которая полностью исключит движения к коленном суставе, или ортез, который позволяет как полностью запрещать движения, так и ограничивать их в заданной амплитуде за счет специальных регулируемых шарниров. При наличии выраженного гемартроза необходимо отсасывать шприцом скапливающуюся в коленном суставе кровь — это позволит значительно снизить боль.
Ортез на коленный сустав
Во втором периоде (несвежий разрыв), который начинается после купирования острой боли в коленном суставе и гемартроза, начинают постепенно восстанавливать движения и приступают к тренировке мышц. Физические упражнения, которые укрепляют подколенные мышцы и четырехглавую мышцу бедра, помогут быстрее возвратить нормальную подвижность в суставе. Тренированные мышцы сами по себе могут стабилизировать коленный сустав, что очень важно при повреждении передней крестообразной связки. Если нет каких-либо других повреждений (связок, менисков), то упражнения выполняют с нарастающей амплитудой и интеснивностью, исключая только лишь те движения, которые провоцируют нестабильность. Для ограничения этих движений целесообразно все время использовать ортез для коленного сустава. Спустя 3-5 недель после травмы, после восстановления амплитуды движений и силы мышц, ортез снимают и еще больше увеличивают активность.
Широко распространенная практика полного обездвиживания коленного сустава гипсовой лонгетой на 5-6 недель неправильна, так как она может привести к стойкому ограничению движений в суставе (контрактуре), атрофии мышц. Конечно же, если повреждению передней крестообразной связки сопутствуют другие травмы структур колена (мениски, другие связки), то тактика лечения во втором периоде может быть другой, например, врач может обоснованно полностью запретить движения в коленном суставе или, наоборот, рекомендовать безотлагательную операцию.
В третьем периоде сустав оценивают с «чистого листа». Грубо говоря, если на нужном уровне физической активности нет признаков нестабильности, то консервативное лечение можно считать успешным. Если при нужном уровне физической активности имеются признаки нестабильности (боль, непослушность колена, выскальзывание, подкашивание и т.д.), то переднюю крестообразную связку можно признать несостоятельной и выходом в такой ситуации может быть операция. Однако важно понимать, что уровень физической активности после прекращения второго периода, когда проводилась иммобилизация, т.е. полное или частичное обездвиживание ортезом, восстанавливается не сразу, а за несколько недель. При этом важно продолжать тренировать передние и задние мышцы бедра, которые могут компенсировать разорванную переднюю крестообразную связку. Если после всего этого сохраняется нестабильность коленного сустава при нужных физических нагрузках, то ставится диагноз хронической передне-медиальной нестабильности, которая лечится только операцией.
Консервативное, т.е. безоперационное лечение изолированных повреждений передней крестообразной связки рекомендуется или может быть эффективным в случаях, когда:
- при частичных разрыв без признаков нестабильности в остром периоде
- при полных разрывах без признаков нестабильности в повседневной жизни у спортсменов, которые более не планируют возвращаться в спорт
- при низких физических запросах (малоподвижный образ жизни).
- у детей и подростков с незакрытыми зонами роста кости
- у пожилых людей (разрывы связки у детей и у пожилых встречаются редко — около 5% случаев от числа всех разрывов)
Консервативное лечение травм передней крестообразной связки обычно не дает хороших результатов у людей, возвращающихся к интенсивным спортивным занятиям. Боль, отек и нестабильность периодически возникают у 56—89% спортсменов с разрывами передней крестообразной связки после консервативного лечения. Важно отметить, что нестабильность увеличивает риск последующих травм внутрисуставных структур (менисков, других связок). Кроме того, в нестабильном коленном суставе сильнее изнашивается хрящ, что приводит к развитию артроза коленного сустава. Без операции вероятность возвращения в профессиональный спорт после полных разрывов передней крестообразной связки составляет не более 20%.
У профессиональных спортсменов операция по восстановлению передней крестообразной связки может быть выполнена и сразу после травмы, т.е. без этапа консервативного лечения. Кроме того, безотлагательная операция может быть целесообразной и в том случае, если разрыву передней крестообразной связки сопутствует и повреждение других внутрисуставных структур. Например, при сопутствующем разрыве мениска его оторванный и болтающийся лоскут может блокировать движения в коленном суставе и, соответственно, полноценное консервативное движение в таком случае не возможно в принципе.
Хирургическое лечение. Операцию делают в том случае, если после консервативного лечения стабильность сустава не соответствует требованиям физической активности. Стоит отметить, что операция дает наилучшие результаты на фоне хороших движений в коленном суставе и сильных мышц, что еще раз подчеркивает важность консервативного этапа лечения. В среднем операции по восстановлению передней крестообразной связки у неспортсменов делают через 6 месяцев после разрыва, но это не значит, что позже операцию делать не нужно. Бывает, что ее делают и через 5-7 лет после травмы. В принципе операция может быть выполнена на любом сроке после травмы, за исключением тех случаев, когда в коленном суставе на фоне разрыва передней крестообразной связки и последовавшей за ним нестабильности коленного сустава развился выраженный артроз.
Идеальный кандидат на операцию — это молодой подвижный человек с объективными (т.е. обнаруживаемыми врачом) и субъективными (ощущаемыми пациентом) признаками нестабильности, желающий заниматься спортом, где необходимы частые рывковые движения и прыжки. Напротив, для больных постарше, с имеющимся дегенеративными изменениями в суставе, не участвующих в соревнованиях и без жалоб на нестабильность сустава больше подходят консервативное лечение, лечебная физкультура.
Сшить разорвавшуюся переднюю крестообразную связку невозможно — для ее восстановления используются трансплантаты, т.е. другие сухожилия (аутотрансплантат из связки надколенника, аутотрансплантат из подколенных сухожилий, аллотрансплантаты) или синтетические протеы. Исключение, пожалуй, только одно — перелом Сегонда (отрыв межмыщелкового возвышения). В таком случае оторвавшийся вместе со связкой костный блок можно фиксировать на свое место.
Схема операции при переломе Сегонда
Еще при первых попытках хирургического восстановления передней крестообразной связки было замечено, что простое сшивание не дает хороших результатов, да это часто и невозможно технически. Начались поиски оптимального способа реконструкции с помощью различных материалов: от искусственных до ауто- и аллотрансплантатов. Аутотрансплантаты — это связки или сухожилия, которые берутся у самого пациента из другого места (связка надколенника, сухожилия хамстринг-мышц и др.). Аллотрасплантаты — специально обработанные связки или сухожилия других людей.
Операцию по восстановлению передней крестообразной связки называют «пластикой передней крестообразной связки» или «стабилизацией коленного сустава». Суть операции заключается в том, что разорвавшуюся связку убирают, а вместо нее ставят заменитель. Сейчас такие операции выполняются малотравматично благодаря артроскопии. Суть артроскопических операций заключается в том, что их выполняют без разрезов, а через маленькие проколы длинной по 1-2 сантиметра. Через один из проколов в сустав вводят артроскоп (оптическую часть видеокамеры), что позволяет осматривать коленный сустав изнутри. Во время операции хирург смотрит на монитор и видит все, что происходит в данный момент в суставе, с большим увеличением – от 40 до 60 раз. Через другой прокол вводят минниатюрные инструменты, которыми и выполняют операцию. Артроскопия позволяет выполнять тончайшие манипуляции на коленном суставе с минимальным повреждением окружающих структур и самого сустава (например, сшивание или удаление части менисков, пересадка хряща, реконструкция связок). Во время операции хирург смотрит на монитор и видит все, что происходит в данный момент в суставе, с большим увеличением – от 40 до 60 раз. Использование современных инструментов и высокочувствительной оптики позволяет выполнять тончайшие манипуляции на коленном суставе с минимальным повреждением окружающих структур и самого сустава (например, сшивание или удаление части менисков, пересадка хряща, реконструкция связок) – и все это через 2–3 небольших разреза.
Артроскопия коленного сустава
Восстановленная передняя крестообразная связка в идеале должна по прочности, расположению и функции соответствовать неповрежденной. Проблема прочности решается за счет адекватного выбора материала для пластики, и первостепенным становится расположение трансплантата. Обычно его проводят через каналы в большеберцовой и бедренной костях таким образом, чтобы позиция трансплантата максимально точно соотвтетствовала нормальной связке.
Для успешной реконструкции необходимо подобрать правильную степень натяжения трансплантата и прочно фиксировать его. Натяжение трансплантата определяет его функциональность: слабо натянутый трансплантат не обеспечивает стабильности коленного сустава, туго натянутый — может порваться или ограничить амплитуду движений в коленном суставе.
Реконструкция связкой надколенника. Эта связка соединяет надколенник с большеберцовой костью. Аутотрансплантат отсекают от большеберцовой кости и надколенника с костными фрагментами, таким образом получается связка с костными блоками на концах. В большеберцовой и бедренной костях просверливаются каналы, выходящие в полость коленного сустава. Внутренние отверстия этих каналов в суставе находятся в тех же самых местах, где находились места прикрепления передней крестообразной связки. Трансплантат связки проводится в полость сустава через канал большеберцовой кости. Концы трансплантата протеза фиксируются в костных каналах при помощи специальных металлических или биополимерных рассасывающихся винтов. Титановый винт очень прочный, но он не рассасывается, что при возможных последующих операциях будет создавать технические трудности. В целом у молодых пациентов мы считаем более предпочтительными рассасывающиеся винты. Иногда такой трансплантат называют BTB-трансплантатом от английской аббревиатуры BTB: bone-tendon-bone (кость-сухожилие-кость). Именно эти костные блоки трансплантата фиксируются в каналах бедренной и большеберцовой кости винтами. Такая фиксация имеет важное преимущество: костный блок трансплантата быстрее срастается со стенками канала — за 2—3 недели, что значительно меньше срока прочного прирастания сухожилия к кости, что требуется, например, у трансплантата из подколенных сухожилий. Края разрезанной связки надколенника, откуда был взят трансплантат, зашиваются.
Схема операции пластики передней крестообразной связки трансплантаом из связки надколенника (BTB-трансплантат)
Принцип фиксации костного блока трансплантата титановым или рассасывающимся винтом
Рентгенограмма после операции стабилизации коленного сустава BTB-трансплантатом. Костные блоки фикированы титановыми винтами, которые хорошо видны на рентгенограмме. Рассасывающиеся винты рентгенпрозрачны и поэтому не видны.
Артроскопическая стабилизация коленного сустава: пластика передней крестообразной связки BTB — трансплантатом (из связки надколенника с костными блоками надколенника и бугристости большеберцовой кости)
Аутотрансплантат из подколенных сухожилий. Иногда этот трансплантат называют еще хамстринг-трансплантатом. В качестве материала для пересадки используется сухожилие полусухожильной мышцы бедра, которое забтирают специальным инструментом (стриппером) через разрез длинной 3-4 сантиметра.
После забора сухожилия полусухожильной мышцы его складывают пополам, прошивают и, точно также как и при описанном выше BTB-трансплантате, просверливают каналы в бедренной и большеберцовой костях, через которые проводят новую связку, натягивают и фиксируют ее. Иногда для пластики забирают не одно сухожилие, а два (из полусухожильной, нежной или полуперепончатой мышц). Вариантов фиксации такого трансплантата больше, чем BTB — скобы, пуговицы, пины, винты и т.д.
Варианты фиксации бедренной части трансплантата: A — EndoButton, Б — винт Mulch В — TransFix, Г — RigidFix, Д — рассасывающийся винт, Е — EZLoc.
Варианты фиксации большеберцовой части трансплантата: A — накладка AO с винтами, Б — WasherLoc, В — накладка с шипами и винт, Г — скобы, Д — нитями к винту (Suture-post fixation), Е — рассасывающийся винт, Ж — IntraFix, З — система GTS (гильза и распирающий винт).
Послеоперационные рентгенограммы (слева — проекция спереди, справа — боковая проекция): аутотрансплантат не виден, так как он состоит из мягкой сухожильной ткани. Фиксация бедренной части трансплантата системой эндобаттон (Еndobutton фирмы
Smith & Nephew, США), а большеберцовая часть фиксирована скобой.
Среди травматологов до сих пор не существует единого мнения о том, какой аутотрансплантат лучше. Реконструкция аутотрансплантатом из связки надколенника травматичнее и восстановление после такой операции протекает сложнее из-за травмирования данной связки. Но зато считается, что такая операция надежнее, костные блоки трансплантата быстрее срастаются со стенками каналов, сколено более стабильно, лучше выдерживает нагрузки. Хотя если хирург хорошо освоил технику выполнения операции по реконструкции из подколенных сухожилий, получаются сравнимые результаты. При второй методике операции (из сухожилия полусухожильной мышцы) получается меньше разрезов и в будущем будет практически незаметно, что была операция на колене. При первой методике (из связки надколенника) об операции будет напоминать 5-сантиметровый рубец на месте разреза, через который забирали часть связки надколенника. Но и он часто малозаметен.
Аллотрансплантаты — это ткани, полученные от донора. После смерти человека передняя крестообразная связка или другая связка забирается и отсылается в банк тканей. Там она проверяется на все инфекции, стерилизуется и замораживается. Когда необходима операция врач отправляет запрос в банк тканей и получает нужный аллотрансплантат. Источником аллотрансплантата могут быть связка надколенника, подколенные сухожилия или ахиллово сухожилие. Преимущество этого метода заключается в том, что хирургу не приходится вырезать трансплантат из организма пациента, нарушая его нормальные связки или сухожилия. Такая операция длится меньше, т.к. не тратится время на выделение трансплантата. У аллотрансплантатов существует риск неприживления. В нашей стране такие операции практически не выполняются.
Для лучшего восстановления двухпучковой структуры передней крестообразной связки существуют и методики двухпучковой реконструкции, когда устанавливаются два трансплантата или один, состоящий из двух ручков.
Осложнения
Вероятность успеха при реконструкции передней крестообразной связки очень высока, тем не менее осложнения все же возможны. Одно из наиболее частых — ограничение подвижности коленного сустава (контрактура). Для профилактики сустав полностью разгибают сразу после операции и поддерживают в разогнутом состоянии. Как можно раньше начинают упражнения, увеличивающие амплитуду движений, стараясь достигнуть 90° сгибания за неделю. Кроме того, важно сохранить подвижность надколенника, чтобы по возможности уменьшить рубцевание связок, соединяющих его с бедренной костью. Другое возможное осложнение — боль в переднем отделе коленного сустава (пателло-феморальный артроз), которая возможна после забора BTB-трансплантата. Также после забора BTB-трансплантата бывают редкие переломы надколенника или разрывы его связки, откуда забирался трансплантат.
Кроме того операция может быть неуспешной -трансплантат может порваться или от может вырваться из костных каналов. В таком случае приходится делать еще одну операцию, ревизионную. Для предотвращения этого осложнения важно тщательно выбирать места проведения костных каналов и жестко фиксировать трансплантат, а сам пациент должен четко соблюдать все рекомендации в послеоперационном периоде, во время реабилитации. В литературе описаны единичные случаи компартмент синдрома после пластики передней крестообразной связки.
Прогноз
Цель операции по пластике восстановления передней крестообразной связки — как можно быстрее вернуть больного к желаемому уровню физической активности и избежать осложнений, к которым в первую очередь относится артроз. Совершенствование хирургической техники и методов реабилитации привело к тому, что более 90% больных продолжают заниматься спортом и полностью удовлетворены результатами лечения. Средний срок реабилитации составляет 4—6 месяцев, но некоторые профессиональные спортсмены с успехом приступают к соревнованиям и через 3 мес. Критерии допуска к спортивным занятиям могут отличаться, но всегда ориентируются в той или иной мере на результаты функциональных проб, ощущения больного и данные осмотра. Наиболее общепринятые критерии следующие: восстановление амплитуды движений, увеличение смещаемости голени по данным гониометрии не более чем на 2—3 мм по сравнению со здоровой ногой, сила четырехглавой мышцы не менее 85% от нормы, восстановление силы задней группы мышц бедра, все функциональные показатели составляют не менее 85% нормы.
Тест по анатомии мышц
Тест по анатомии основных мышц для класса анатомии и физиологии!
Когда вы изучаете анатомию и физиологию, вам потребуется определить основные мышцы человеческого тела. Этот тест требует и , поэтому он проверит ваши знания о том, как идентифицировать эти мышцы (широчайшая мышца спины, трапеция, дельтовидная мышца, двуглавая мышца плеча, трицепс плеча, брахиорадиалис, большая грудная мышца, передняя зубчатая мышца, прямая мышца живота и т. Д.).
Перед тем, как пройти этот тест, вы можете посмотреть песню о мышцах, которая дает вам несколько стишков, которые помогут вам запомнить расположение и действие мышц.Кроме того, вы можете просмотреть текст песни анатомии мышц и примечания.
(ПРИМЕЧАНИЕ. Когда вы нажмете «Отправить», эта же страница обновится. Прокрутите вниз, чтобы увидеть свои результаты.)
Тест по анатомии мышц
1. Найдите мышцу, обозначенную цифрой «1» на схеме выше:
A. Latissimus Dorsi
Б. Трапеция
C. Serratus Anterior
D. Vastus Medialis
Ответ: A: Latssimus Dorsi
2.Найдите мышцу, обозначенную цифрой «2» на схеме выше:
A. Rectus Abdominis
Б. Широчайшая мышца спины
К. Трапеция
D. Semitendinosus
Ответ: C: Trapezius
3. Найдите мышцу, обозначенную цифрой «3» на схеме выше:
A. Двуглавая мышца плеча
Б. Дельтовидная
C. Трицепс плеча
D. Brachioradialis
Ответ: B: Дельтоид
4. Найдите мышцу, обозначенную цифрой «4» на схеме выше:
А.Трицепс плеча
B. Brachioradialis
C. Двуглавая мышца бедра
D. Двуглавая мышца плеча
Ответ: D: Biceps Brachii
.5. Найдите мышцу, обозначенную цифрой 5 на схеме выше:
А. Трицепс плеча
B. Gastrocnemius
C. Semimembranosus
D. Двуглавая мышца бедра
Ответ: A: Трицепс плеча
.6. Найдите мышцу, обозначенную цифрой «6» на схеме выше:
A. Vastus Lateralis
Б.Двуглавая мышца бедра
C. Brachioradialis
D. Pectoralis Major
Ответ: C: Brachioradialis
.7. Найдите мышцу, обозначенную цифрой «7» на схеме выше:
A. Gluteus Maximus
Б. Серратус передний
C. Pectoralis Major
D. Широчайшая мышца спины
Ответ: C: Большая грудная мышца
8. Найдите мышцу, обозначенную цифрой «8» на схеме выше:
A. Rectus Abdominis
Б.Серратус передний
C. Косые
D. Semitendinosus
Ответ: B: Serratus Anterior
.9. Найдите мышцу, обозначенную цифрой «9» на схеме выше:
A. Косые
Б. Серратус передний
C. Rectus Femoris
D. Прямые мышцы живота
Ответ: D: Rectus Abdominis
.10. Найдите мышцу, обозначенную цифрой «10» на схеме выше:
A. Косые
Б. Трапеция
С.Полумембранозный
D. Rectus Femoris
Ответ: A: косые
11. Найдите мышцу, обозначенную цифрой «11» на схеме выше:
A. Pectoralis Major
Большая ягодичная мышца
С. Широчайшая мышца спины
D. Gastrocnemius
Ответ: B: Gluteus Maximus
12. Найдите мышцу, обозначенную цифрой «12» на схеме выше:
A. Vastus Medialis
B. Rectus Femoris
С.Большой латеральный пояс
D. Прямые мышцы живота
Ответ: B: Rectus Femoris
.13. Найдите мышцу, обозначенную цифрой 13 на схеме выше:
A. Rectus Femoris
B. Vastus Intermedius
C. Vastus Lateralis
D. Vastus Medialis
Ответ: C: Vastus Lateralis
.14. Найдите мышцу, обозначенную цифрой «14» на схеме выше:
A. Rectus Femoris
B. Vastus Intermedius
С.Большой латеральный пояс
D. Vastus Medialis
Ответ: D: Vastus Medialis
15. Найдите мышцу, обозначенную цифрой «15» на схеме выше:
A. Двуглавая мышца бедра
Б. Двуглавая мышца плеча
C. Semimembranosus
D. Semitendinosus
Ответ: A: Бицепс бедра
16. Найдите мышцу, обозначенную цифрой «16» на схеме выше:
A. Semimembranosus
B. Semitendinosus
С.Двуглавая мышца бедра
D. Vastus Lateralis
Ответ: B: Semitendinosus
.17. Найдите мышцу, обозначенную цифрой «17» на схеме выше:
A. Rectus Femoris
B. Semitendinosus
C. Semimembranosus
D. Vastus Lateralis
Ответ: C: Semimembranosus
.18. Найдите мышцу, обозначенную цифрой 18 на схеме выше:
Прямая мышца живота
A. Latissimus Dorsi
Б.Трапеция
C. Semitendinosus
D. Gastrocnemius
Ответ: D: Gastrocnemius
.Не забудьте рассказать об этой викторине своим друзьям, поделившись ею в Facebook, Twitter и других социальных сетях. Вы также можете пройти более увлекательные медсестринские викторины.
* Заявление об ограничении ответственности. Хотя мы делаем все возможное, чтобы предоставить учащимся точные и углубленные учебные викторины, эта викторина / тест предназначена только для образовательных и развлекательных целей. Пожалуйста, обратитесь к последним обзорным книгам NCLEX для получения последних обновлений по сестринскому делу.Авторские права на эту викторину принадлежат RegisteredNurseRn.com. Пожалуйста, не копируйте этот тест напрямую; однако, пожалуйста, поделитесь ссылкой на эту страницу со студентами, друзьями и другими людьми.
Генно-инженерный трансплантат мышц человека усиливает неоваскуляризацию и миогенез мыши-хозяина
Культура клеток
Первичные венозные эндотелиальные клетки взрослых и первичные венозные гладкомышечные клетки взрослых (взрослые SMC) были выделены из сегмента вены нижних конечностей пожилых пациентов, перенесших операции шунтирования. как описано ранее 19,25,28 .Взрослые венозные эндотелиальные клетки, экспрессирующие флуоресцентный белок ANGPT1 и ZsGreen (эндотелиальная клетка ANGPT1 ), культивировали в эндотелиальной базальной ростовой среде-2 с добавлением фактора роста фибробластов-2, инсулиноподобного фактора роста-1 и эпидермального фактора роста из EGM. -2 BulletKit (Lonza, США) и 3% фетальная бычья сыворотка (FBS; HyClone, Thermo Fisher Scientific, США). Как экспрессирующие VEGF (SMC VEGF ), так и наивные взрослые SMC культивировали в среде Игла, модифицированной по Дульбекко, с добавлением 20% FBS и раствора пенициллина / стрептомицина.Использование сегментов вены, не используемых в хирургии, было одобрено Наблюдательным советом Медицинского центра леди Дэвис Кармель. Информированное согласие было получено от пациентов, которым было назначено шунтирование. Первичные клетки скелетных мышц человека (hMyo) были приобретены у PromoCell и культивированы в рекомендованной среде для роста клеток скелетных мышц PromoCell. Клетки инкубировали в увлажненной атмосфере с 5% CO 2 при 37 ° C. Клетки разделяли (1: 3) для субкультивирования и использовали на 8 или 9 пассаже для всех экспериментов.
Лентивирус и ретровирусная трансдукция
Все эндотелиальные клетки, использованные в этом исследовании, были предварительно помечены нецелевым флуоресцентным белком с помощью частиц лентивируса ZsGreen, как описано ранее 12 . Для эндотелиальной клетки ANGPT1 взрослые эндотелиальные клетки сначала трансдуцировали ретровирусным вектором для экспрессии ANGPT1, а затем трансдуцировали частицами лентивируса ZsGreen. Взрослые SMC трансдуцировали ретровирусным вектором для экспрессии VEGF 165 (SMC VEGF ), как описано ранее 19,43 .Экспрессию трансгена определяли с помощью иммуногистохимии (IHC) и ELISA. Минимальные уровни экспрессии ИГХ были установлены на уровне 55% для эндотелиальных клеток, экспрессирующих ANGPT1, и 60% для SMC, экспрессирующих VEGF 165. Минимальный уровень экспрессии трансгена с помощью ELISA составлял 0,05 пг для ANGPT1 и VEGF 165 белок / одиночная клетка / 24 ч 20 . Эти минимальные уровни были установлены на основании результатов, полученных в исследованиях эффективности на животных и исследованиях прорастания in vitro 25 . Комбинация эндотелиальных клеток ANGPT1 –SMC VEGF была установлена на основе результатов, полученных в анализе сфероидов с залитым коллагеном (дополнительный рис.4).
Изготовление каркасов из PLLA / PLGA
Метод солевого выщелачивания был использован для изготовления трехмерных пористых поли-1-молочной кислоты (PLLA) (Polysciences, Warrington) и полимолочной гликолевой кислоты (PLGA; Boehringer Ingelheim) (1: 1) каркасов , с размером пор 212-600 мкм и пористостью 93%, как описано ранее 12,15,21,44 . Вкратце, был приготовлен раствор полимера PLLA и PLGA, растворенного в хлороформе, 1: 1. Затем 0,24 мл этого раствора добавляли к 0,4 г частиц хлорида натрия в тефлоновых формах диаметром 18 мм.Хлороформу давали испариться в течение ночи, а каркасы промывали дистиллированной водой в течение 8 часов. Перед каждым экспериментом каркасы толщиной 1 мм разрезали на круги диаметром 6 мм, вымачивали в 70% (об. / Об.) Этиловом спирте в течение 2 часов, а затем промывали 3 раза физиологическим раствором с фосфатным буфером (PBS).
Создание васкуляризованных трехмерных конструкций
В этом исследовании были исследованы шесть многоклеточных культур: (i) эндотелиальные клетки (0,5 × 10 6 клеток) и SMC (0,1 × 10 6 клеток) культивировались для создания трехмерных сосудистых сетей; (ii) эндотелиальные клетки (0.5 × 10 6 клеток) и SMC VEGF (0,1 × 10 6 клеток) культивировали для создания трехмерных сосудистых сетей; (iii) эндотелиальные клетки ANGPT1 (0,5 × 10 6 клеток) и SMC (0,1 × 10 6 клеток) культивировали для создания трехмерных сосудистых сетей; (iv) эндотелиальные клетки ANGPT1 (0,5 × 10 6 клеток) и SMC VEGF (0,1 × 10 6 клеток) культивировали для создания трехмерных сосудистых сетей; (v) эндотелиальные клетки (0.5 × 10 6 клеток), SMC (0,1 × 10 6 клеток) и hMyo (0,2 × 10 6 клеток) культивировали для создания трехмерных сосудистых сетей в конструкциях скелетных мышц; и (vi) эндотелиальные клетки ANGPT1 (0,5 × 10 6 клеток), SMC VEGF (0,1 × 10 6 клеток) и hMyo (0,2 × 10 6 клеток) культивировали для создания трехмерных сосудистых сетей. внутри конструкций скелетных мышц.
Соотношения и концентрации клеток в совместной и трех культурах были определены на основании предыдущих исследований 15,21,24 .Клетки трипсинизировали на 8 или 9 пассаже и повторно суспендировали в 4 мкл раствора тромбина 5 Ед / мл (Johnson & Johnson Medical, Израиль). Затем к смеси клетки с тромбином добавляли 4 мкл 15 мг / мл фибриногена (Johnson & Johnson Medical, Израиль) и каждую клеточную суспензию высевали на каркасы PLLA / PLGA, а затем давали возможность затвердеть в течение 30 минут (37 ° C). , 5% CO 2 ) внутри 6-луночного планшета для нетканевых культур. После затвердевания в каждую лунку добавляли 4 мл культуральной среды. Среду меняли через день.
Оценка удлинения сосудистой сети
Для оценки удлинения сосудоподобных структур, образованных различными комбинациями клеток in vitro, каркасы были визуализированы с помощью конфокального микроскопа через 4 и 7 дней после посева и эксцентриситета ZsGreen-положительных сосудовидных Структуры были рассчитаны с использованием самописного алгоритма MATLAB (MATLAB © , MathWorks). Вкратце, каждое изображение стопки Z было преобразовано в двоичное изображение, а затем разделено на отдельные элементы до определения эксцентриситета с использованием функции «regionprops».Элемент в форме круга получил значение ноль, а более удлиненные элементы получили значение эксцентриситета, близкое к единице (дополнительный рисунок 1 и дополнительное программное обеспечение с 1 по 4).
Иммунофлуоресцентное окрашивание целиком
Целые залитые клетками каркасы окрашивали, как описано ранее 12 . Сначала скаффолды фиксировали в 4% параформальдегиде (Electron Microscopy Sciences, США) в течение 10 мин, а затем промывали PBS. Затем клетки были проницаемы путем добавления 0.3% Triton X-100 (Bio Lab Ltd.) в течение 10 мин при комнатной температуре (КТ). Затем каркасы промывали PBS и вымачивали на 1 час в блокирующей сыворотке (10% FBS, 0,1% Triton X-100, 1% глицин) при комнатной температуре. Затем каркасы инкубировали в течение ночи при 4 ° C со следующими первичными антителами (разведенными в блокирующем растворе): мышиные антитела против αSMA человека (1:50, Dako), мышиные антитела против человеческого коллагена IV (1: 500, Sigma-Aldrich). и мышиный анти-человеческий VE-кадгерин (1: 150, Santa Cruz). После четырех промывок по 10 минут каждая в PBS наносили Cy3-конъюгированный ослиный антимышиный IgG (1: 100, иммуно-исследовательская лаборатория Джексона, Пенсильвания) на 3 часа.Ядра контрастировали 4 ’, 6-диамидино-2-фенилиндолом (DAPI) (1: 1000, Sigma-Aldrich). Затем каркасы промывали в PBS и хранили в 24-луночных планшетах в PBS при 4 ° C до наблюдения под инвертированным конфокальным микроскопом Zeiss LSM700 (Carl Zeiss) с линзами объектива 5x и 10x с использованием программного обеспечения ZEN (Carl Zeiss). Дальнейший анализ изображений проводился с использованием программного обеспечения FIJI (Fiji Is Just ImageJ).
Определение стабильности и зрелости сосудов
Для оценки стабильности и зрелости сосудов скаффолды окрашивали на αSMA, коллаген IV и VE-кадгерин.Каркасы получали с помощью конфокального микроскопа через 4 и 7 дней после посева и анализировали конфокальные проекционные изображения z -стека каркасов.
Общая площадь экспрессии αSMA была определена количественно с использованием самописного алгоритма MATLAB.
Удлинение VE-кадгерин-положительных сосудов количественно определяли с использованием написанного самим собой алгоритма MATLAB для определения эксцентриситета (дополнительный рисунок 1 и дополнительное программное обеспечение с 1 по 4). Процент коллагеновых IV- и αSMA-положительных сосудов определялся независимым экспертом, не знающим компонентов культуры.
Определение секреции ANGPT1, ANGPT2 и VEGF
Супернатанты из всех групп каркасов собирали через 4 дня после посева, и концентрации ANGPT1, ANGPT2 и VEGF определяли с использованием наборов для обнаружения ANGPT1, ANGPT2 и VEGF ELISA (R&D SYSTEMS) человека соответственно. Оптическая плотность (OD) ростовой среды без культивированных клеток служила контролем и вычиталась из значений OD тестируемых групп.
Окно визуализации брюшной полости
AIW был разработан нами путем модификации протокола, описанного Ritsma et al. 45,46 и изготовлены для визуализации живых мышц. AIW состоял из кольца из нержавеющей стали многоразового использования (внешний диаметр 17 мм, внутренний диаметр 13 мм и толщина 2,3 мм) с канавкой 1,3 мм на боковой стороне (рис. 6). После имплантации AIW закрывали круглым покровным стеклом диаметром 13 мм, покрытым поли-L-лизин-g-поли (этиленгликолем). Покровное стекло фиксировали стопорным кольцом для последующей прижизненной микроскопии (рис. 5).
Стабилизирующее устройство визуализации
Стабилизирующее устройство визуализации из полиметилметакрилата (SID) было разработано нами (Рис.5i), чтобы стабилизировать мышь во время прижизненной визуализации и минимизировать дыхательные движения, обеспечивая сфокусированные изображения.
Трансплантация модифицированной ткани
Все хирургические процедуры проводились в соответствии с протоколами, утвержденными Комитетом по уходу и использованию животных Института Техниона. Размер выборки был определен с помощью анализа мощности (β = 0,1 и α = 0,05). Размер стандартизованного эффекта был рассчитан на основе предыдущих исследований и пилотного эксперимента; В результате размер выборки составил 6.Четыре животных были исключены из исследования, поскольку их AIW упал до достижения конечной точки. Было проведено два независимых эксперимента, и в каждом эксперименте было проанализировано не менее трех животных на группу лечения. Отбор животных для распределения в опытную и контрольную группы производился случайным образом. Три-культуры взрослых эндотелиальных клеток, SMC и hMyo ( n = 6) и взрослых эндотелиальных клеток ANGPT1 , SMC VEGF и hMyo ( n = 6) культивировали на каркасах PLLA / PLGA в течение 4 дней. перед имплантацией.Самок голых 9-недельных мышей (Harlan Laboratories) анестезировали путем внутрибрюшинной инъекции смеси кетамин-ксилазин (100 мг / кг и 10 мг / кг соответственно) с помощью иглы 30-го калибра. Бупренорфин (0,05 мг / кг) вводили подкожно за 20 мин до процедуры и каждые 12 ч после этого в течение 3 дней. Место запланированного разреза было очищено спиртом и йодом для создания асептического рабочего поля. Затем была обнажена брюшная стенка посредством вентрального разреза кожи и удален сегмент прямой мышцы живота на полную толщину диаметром 6 мм.Инженерный трансплантат скелетных мышц был зашит полипропиленовой нитью 8-0.
Имплантация окна визуализации брюшной полости
Перед использованием AIW вымачивали на ночь в 70% (об. / Об.) Этиловом спирте и трижды промывали PBS. Имплантация AIW проводилась по протоколу, представленному Ritsma et al. 45,46 с некоторыми модификациями. Вкратце, после ушивания искусственной ткани брюшной стенки вокруг разреза через кожу наложили кисетный шов с использованием шелковых швов 4-0.Затем цианоакрилатный клей наносили на внутреннюю кольцевую поверхность AIW, и AIW приклеивали вокруг трансплантированной ткани, прилагая легкое давление. Кожу осторожно поместили в бороздку AIW. Затем петли кисетного шва вытягивали одну за другой, затягивая нить в канавке AIW. Затем сверху помещали круглое покровное стекло и фиксировали стопорным кольцом. За всеми мышами внимательно наблюдали в течение 1-2 часов, чтобы гарантировать полное выздоровление после анестезии. За мышами ежедневно наблюдали для оценки общего состояния здоровья и впоследствии включали в исследования прижизненной микроскопии.
Прижизненная визуализация
Прижизненная микроскопия выполнялась через 4, 7, 10 и 14 дней после трансплантации с использованием конфокального микроскопа LSM700. Для визуализации сосудистой сети хозяина у мышей конъюгированные с Alexa Fluor® 647 антитела против CD31 мыши (mCD31-X647; Biolegend) (0,5 мг / мл) вводили внутривенно в хвостовую вену и давали возможность циркулировать в течение ~ 15 мин. Затем мышей анестезировали (используя кетамин-ксилазин, как описано выше) и внутривенно вводили TRITC-декстран (10 мг / мл) (средняя молекулярная масса 155 000, Sigma-Aldrich) в хвостовую вену для визуализации функциональных сосудов.Мышей помещали в SID и проводили прижизненную микроскопию. В течение всего сеанса визуализации температура поддерживалась на уровне 28 ° C с помощью камеры для нагрева. В конечной точке мышей умерщвляли, а трансплантаты извлекали и фиксировали в 10% формалине (Sigma-Aldrich).
Количественная оценка длины сосудов
Общая длина (мм) сосудов в конструкциях через 14 дней после трансплантации была рассчитана путем анализа конфокальных проекционных изображений стека z с использованием программного обеспечения AngioTool (AngioTool ® ) 47 .Было проведено два независимых эксперимента, и в каждом эксперименте было проанализировано не менее трех животных на группу лечения.
Иммуногистохимическое и иммунофлуоресцентное окрашивание
Трансплантаты, извлеченные через 14 дней после трансплантации, инкубировали в течение ночи в 30% -ном (мас. / Об.) Растворе сахарозы, залитом в состав оптимальной температуры резки (OCT) (Tissue-Tec, США) и заморозили для последующего использования. криосрезов на срезы толщиной 5 и 10 мкм, как описано ранее 12 . Вкратце, для иммунофлуоресцентного окрашивания срезы инкубировали при 0.5% раствор твина в течение 20 минут, промывают PBS и затем блокируют 5% (масс. / Об.) Раствором бычьего сывороточного альбумина (Sigma-Aldrich) в течение дополнительных 30 минут. Затем срезы инкубировали в течение ночи при 4 ° C с козьим антидесмином (1:50, Santa Cruz), разведенным в блокирующем растворе. Затем срезы промывали PBS и метили IgG, конъюгированным с Alexa 405 (1: 400; Molecular Probes), перед помещением во Fluromount-G (Southern Biotechnology) и исследовали под конфокальным микроскопом. Для окрашивания H&E предметные стекла сушили на воздухе в течение нескольких минут и окрашивали фильтрованным 0.1% гематоксилин Майерса (Sigma-Aldrich) в течение 10 мин. Затем предметные стекла промывали водопроводной водой и окрашивали 0,5% эозином (Sigma-Aldrich) в течение 1 мин. Затем предметные стекла были погружены в бидистиллированную воду (DDW) и обезвожены последовательными погружениями в этанол с возрастающими концентрациями. Наконец, слайды были погружены в ксилол и покрыты Vectamount (Vector Labs). Для окрашивания трихромом по Массону слайды сушили на воздухе, а затем окрашивали фильтрованным 0,1% гематоксилином Майерса в течение 5 мин. Затем предметные стекла промывали дистиллированной водой и окрашивали трихромным красителем (Sigma-Aldrich) в течение 2 минут.Затем предметные стекла дважды промывали 0,2% ледяной уксусной кислотой, а затем DDW. После этого предметные стекла были обезвожены путем последовательного погружения в этанол с возрастающей концентрацией и, наконец, окунули в ксилол и покрыли Vectamount. Слайды получали с помощью автоматического цифрового сканера слайдов Pannoramic MIDI (3DHISTECH, Венгрия). Изображения анализировали с помощью программы Pannoramic Viewer (3DHISTECH, Венгрия).
Количественная оценка плотности сосудов Ex vivo
Плотность сосудов-хозяев в трансплантатах, извлеченных через 14 дней после трансплантации, и в окружающей нативной мышце, рассчитывали путем анализа конфокальных проекционных изображений стека z с использованием программного обеспечения FIJI.Было проведено два независимых эксперимента, и в каждом эксперименте было проанализировано не менее трех животных на группу лечения.
Статистический анализ
Статистический анализ выполняли с использованием компьютеризированной статистической программы (GraphPad Software, Inc.). Данные представлены в виде графиков в виде прямоугольников, где центральные линии обозначают медианы, края — верхний и нижний квартили, а усы — минимальные и максимальные значения. Групповые различия определяли с помощью непарного двустороннего критерия Стьюдента t .При необходимости данные анализировали с помощью одностороннего дисперсионного анализа (ANOVA) с последующим тестом множественного сравнения Холма – Сидака. Значения p ниже 0,05 были приняты, чтобы указать на статистически значимое различие между группами.
Доступность кода
Исходный код MATLAB для оценки удлинения сосудистой сети предоставляется в качестве дополнительного программного обеспечения 3.
Набор данных архитектуры мышц нижней конечности человека in vivo, полученный с помощью визуализации тензора диффузии
Abstract
«Золотой стандарт» эталонных наборов архитектуры мускулов человека основан на образцах трупов пожилых людей, которые вряд ли будут репрезентативными для значительной части человеческой популяции.Это важно для моделирования опорно-двигательного аппарата, где свойства общих моделей генерировать мышечную силу определяются этими данными, но могут быть неприменимы при применении к моделям молодых, здоровых людей. Получение данных об индивидуальной архитектуре мышц in vivo затруднительно, однако в последнее время в качестве надежного метода достижения этой цели стала использоваться диффузионно-тензорная магнитно-резонансная томография (DTI). DTI был использован здесь для предоставления набора данных об архитектуре 20 мышц нижних конечностей от 10 здоровых взрослых, включая длину мышечных волокон, которые являются важными входными данными для моделей мышц типа Хилла, обычно используемых при моделировании опорно-двигательного аппарата.Было обнаружено, что максимальная изометрическая сила и длина мышечных волокон не соотносятся с антропометрией объекта, что позволяет предположить, что эти факторы может быть трудно предсказать с использованием алгоритмов масштабирования или оптимизации. Эти данные также подчеркивают высокий уровень анатомических различий, которые существуют между людьми с точки зрения архитектуры мышц нижних конечностей, что подтверждает необходимость включения свойств создания силы в модели опорно-двигательного аппарата для оптимизации их точности для клинической оценки.
Образец цитирования: Charles JP, Suntaxi F, Anderst WJ (2019) Набор данных по архитектуре мышц нижней конечности человека in vivo , полученный с помощью визуализации тензора диффузии. PLoS ONE 14 (10): e0223531. https://doi.org/10.1371/journal.pone.0223531
Редактор: Джулия Мольнар, Нью-Йоркский технологический институт, США
Поступила: 17 марта 2019 г .; Принята к печати: 23 сентября 2019 г .; Опубликовано: 15 октября 2019 г.
Авторские права: © 2019 Charles et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией. Скрипт MATLAB, использованный в этом исследовании, находится в свободном доступе по адресу DOI: 10.6084 / m9.figshare.96.
Финансирование: Работа выполнена при финансовой поддержке кафедры ортопедической хирургии Питтсбургского университета.Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Костно-мышечная архитектура ( и . и . Макроскопическое расположение мышечных волокон [1]) нижней конечности человека хорошо определена, и опубликовано несколько обширных наборов данных [2, 3]. Однако эти наборы справочных данных «золотого стандарта» основаны на образцах трупов пожилых людей, которые по разным причинам, таким как возможные изменения в архитектуре мышц из-за старения [4], вряд ли будут репрезентативными для молодых, активных и здоровых взрослых [5 ].Эти различия были подчеркнуты в отношении объемов мышц [5, 6], хотя степень вариации свойств архитектуры мышц, таких как длина мышечных волокон, угол перистости и максимальная изометрическая сила, в значительной степени неизвестна. Это особенно важно в контексте моделирования опорно-двигательного аппарата с использованием безразмерных моделей мышц типа Хилла [7], которые определяются этими свойствами. Важно отметить, что различные анализы чувствительности показали, что эти модели особенно чувствительны даже к небольшим изменениям мышечных волокон и, в частности, длины провисания сухожилий [8–14].Более того, насколько хорошо эти параметры масштабируются по отношению к антропометрическим факторам тела, таким как масса тела или конечностей, также подробно не сообщалось, хотя было высказано предположение, что длина волокон может не особенно сильно зависеть от длины кости [15]. Хотя параметры архитектуры мышц можно оценить с помощью оптимизации [16–19], прямое измерение этих in vivo может повысить точность вычислительных моделей.
Используя ранее установленную основу диффузионно-тензорной визуализации (DTI) и волоконной трактографии, в сочетании с другими последовательностями магнитно-резонансной томографии [20], это исследование направлено на развитие предыдущей литературы и предоставление подробного описания нижней конечности человека in vivo набор данных.Это также подчеркнет уровень межсубъектной изменчивости параметров мышечной архитектуры, которая существует у молодых здоровых взрослых, а также масштабные отношения между мышечной архитектурой и пропорциями тела.
Методы
Для настоящего исследования данные были собраны по 20 мышцам правых нижних конечностей 10 здоровых, непрофессионально атлетически подготовленных взрослых (5 мужчин, 5 женщин; возраст 27 ± 4 года. Масса тела 76 ± 12) кг; Таблица 1), которые подписали документы об информированном согласии до участия в этом исследовании, одобренном IRB.Анализируемые мышцы были разделены на 5 отдельных функциональных групп на основе основных функций (таблица 2), которые были основаны на предыдущих исследованиях архитектуры мышц человека [3]. Длина мышечных волокон, угол перистости волокон и объемы мышц оценивались для каждой из этих мышц с использованием проверенной структуры магнитно-резонансной томографии (МРТ) и DTI [20]; см. ниже).
Таблица 1. Информация об участниках исследования.
Длина бедра — расстояние между самой проксимальной стороной большого вертела бедренной кости и самой дистальной стороной латерального мыщелка бедренной кости.Длина ноги — расстояние между плато большеберцовой кости и центром голеностопного (тибиоталарного) сустава. L L — Полная длина нижней конечности. V LM — Общий объем мышц нижней конечности (сумма объемов исследуемых мышц). V L — Общий объем нижней конечности (сумма объемов мышц плюс жир, фасция и кожа).
https://doi.org/10.1371/journal.pone.0223531.t001
Получение МРТ и DTI
Все МРТ-изображения были получены от гребня подвздошной кости до голеностопного сустава с использованием сканера 3 Т (Biograph mMR, Siemens, Мюнхен, Германия), при этом каждый субъект находился в положении лежа на спине, а нижние конечности — в анатомическом положении.Визуализация состояла из двух последовательностей (рис. 1A и 1B): T1-взвешенное анатомическое турбо спин-эхо (TSE) (размер вокселя 0,47 × 0,47 × 6,5 мм 3 , время повторения [TR] — 650 мс, время эхо [TE] — 23 мс, количество срезов — 35 на сегмент, количество средних сигналов (NSA) — 1, коэффициент ускорения — 2), и планарное спин-эхо с однократным повторным фокусом и двойным перефокусированием (размер вокселя 2,96 × 2,96 × 6,5 мм). 3 , TR / TE 7900/65 мс, градиенты диффузии в 12 направлениях, b Значение — 0 и 400 с / мм 2 , сильное подавление жира — восстановление спектральной ослабленной инверсии [SPAIR], количество срезов — 35 на сегмент, NSA — 2, коэффициент ускорения — 2, полоса пропускания — 2440 Гц / пиксель).Усовершенствованная регулировка шиммирования B 0 была выполнена для каждого сегмента, чтобы уменьшить пространственное искажение и минимизировать химический сдвиг остаточного жира в изображениях, взвешенных по диффузии, в направлении фазового кодирования (от переднего к заднему). Для каждого объекта изображения были получены в аксиальной ориентации срезов и повторены в общей сложности от пяти до шести сегментов, которые были объединены во время постобработки с использованием плагина Stitching для Fiji / ImageJ [21, 22]. Общее время получения изображения составляло ~ 37 минут на одного человека.
Рис 1.
Типичное Т1-взвешенное МРТ анатомическое изображение (A) и изображение тензора диффузии (B) сегмента бедра одного пациента. Мышцы и кости были сегментированы в цифровом виде из изображений T1 для создания трехмерных представлений нижних конечностей (C) (определения сокращений мышц см. В таблице 2). Мышечные пучки (волокна) отслеживались по диффузионно-взвешенным МР-изображениям (D). С помощью этих трехмерных моделей на основе облака точек можно было измерить длину волокна (L f ) и угол перистости поверхностных волокон (θ, угол волокон относительно линии действия мышцы (синяя линия)).
https://doi.org/10.1371/journal.pone.0223531.g001
МРТ-изображения, взвешенные по T1, были сегментированы в цифровой форме в программе Mimics (Materialise, Левен, Бельгия) для создания трехмерных сеток каждой мышцы (рис. 1C). , что позволило определить объем отдельных мышц (живота) ( 3 мм).
Предварительная обработка DTI и волоконная трактография
Изображения тензора диффузии были предварительно обработаны для уменьшения артефактов изображения и улучшения отношения сигнал / шум.Чтобы уменьшить артефакты изображения, вызванные возможным движением объектов или пространственным искажением (вихревые токи и / или неоднородность магнитного поля), каждое взвешенное по диффузии изображение было зарегистрировано в недиффузионно-взвешенное изображение (со значением b 0) с использованием аффинного преобразования. в DTI-studio [23].
Чтобы уменьшить отношение сигнал / шум изображений, алгоритм подавления шума Rician был применен к изображениям DTI [24] в MedINRIA (www.med.inria.fr), где тензоры диффузии для каждого объекта были оценены и сглажены.Ручная установка пороговых значений удалила фоновые пиксели из оценки тензора. Мышечные пучки для каждой мышцы оценивались на основе этих тензоров с помощью трактографии в программном обеспечении Camino [25], создавая тракты волокон из областей интереса (ROI), построенных на основе анатомических изображений T1 MR (рис. 1D). Эти тракты отслеживались в двух направлениях (размер шага 1 мм) от интересующих областей засева (ROI) и продолжались до тех пор, пока не прекращались на основании определенных критериев остановки кривизны волокна (изменение угла> 10 градусов на 5 мм).Эти параметры трактографии были одинаковыми для мышц и испытуемых.
Хотя мышечные волокна не обязательно могут проходить по всей длине мышечного пучка (пучки из ~ 5–10 волокон) и вместо этого могут быть соединены последовательно, было показано, что волокна в таком расположении могут активироваться одновременно, чтобы действовать как единое целое. волокно [26]. Поэтому здесь предполагалось, что длина волокна функционально эквивалентна длине пучка, и эти термины используются как синонимы. Исходя из этого предположения, пользовательский код MATLAB (доступен на www.figshare.com — DOI: 10.6084 / m9.figshare.96) был использован для измерения L f из этих волоконных трактов (эквивалентных мышечным пучкам), и приведенные здесь значения являются средними для всего диапазона длин волоконных трактов в каждой мышце. [20]. Это стандартная практика при измерении архитектуры мышц для входных данных в модели мышц типа Хилла [3, 27], и в этом контексте было показано, что средняя точность L f составляет <1-7 мм [20].
Угол перистости этих волокон был измерен здесь как угол волокон относительно линии действия мышцы.Линия действия каждой мышцы оценивалась с использованием функции «подгонки центральной линии» на каждой трехмерной мышечной сетке в Mimics (из T1-взвешенных МРТ-изображений), которая оценивает линию, проходящую через осевые центроиды каждой сетки, и, следовательно, учитывает их часто -изогнутые формы. Предположение, что эта линия эквивалентна анатомической линии действия, было высказано ранее [6]. Пять поверхностных (2D) измерений угла перистости были записаны вручную в проксимальной, средней и дистальной областях каждой мышцы с помощью ImageJ [28] для получения репрезентативного среднего значения.Это также стандартная практика для оценки этого параметра для моделей опорно-двигательного аппарата, и было показано, что углы перистости поверхности оцениваются со средней точностью 4 ± 1 ° [20].
Все эти методы были выполнены одним и тем же исследователем для каждого субъекта, что обеспечило согласованность представленных данных по архитектуре мышц.
Прогнозирование оптимальной длины волокна
Ранее признанное ограничение измерения длины волокон по изображениям тензора диффузии заключается в том, что оценки оптимальной длины волокон (важный вход для моделей опорно-двигательного аппарата) не могут быть получены с использованием только этого метода.Это связано с тем, что длины саркомеров, которые нормализованы к стандартизированной оптимальной длине саркомера в состоянии покоя для оценки оптимальной длины волокон, не могут быть напрямую измерены по отслеживаемым волокнам. Поэтому оптимальные длины волокон были оценены с использованием длин саркомеров, указанных в [3], с использованием следующего уравнения [29]: где L f — оптимальная длина волокна, L f ’- длина сырого волокна (измеренная по DTI), L s — длина саркомера, а 2,7 мкм — общее значение для оптимальной длины саркомера [29].Значения L s были получены от Ward et al. [3], которые измерили L s в фиксированных мышцах, отсеченных от конечностей с большинством суставов (кроме голеностопного) в анатомическом положении, как в настоящем исследовании.
Эти параметры затем использовались для расчета физиологической площади поперечного сечения (PCSA, мм 2 ), основного определяющего фактора выхода мышечной силы, с использованием уравнения (из [30]): где V m — объем мышцы (живота) (мм 3 ), L f — оптимальная длина мышечного волокна (мм), θ — угол перистости мышечного волокна.Чтобы оценить максимальную изометрическую силу (важный входной параметр для скелетно-мышечных моделей, F max ), индивидуальные значения PCSA умножались на изометрическое напряжение скелетных мышц (или удельное напряжение; 0,3 Н · мм -2 ; [7]). Использование этого общего значения для изометрического стресса хорошо установлено в исследованиях по моделированию опорно-двигательного аппарата [13, 31], и было показано, что оно не зависит от размера тела и сохраняется в филогении позвоночных [32]. Учитывая, что оценка этого значения для каждой отдельной мышцы нижней конечности выходила за рамки данного исследования, здесь предполагалось, что оно будет постоянным для всех мышц.Однако признается, что в действительности это может быть не так, поскольку в литературе сообщается о широком диапазоне значений (0,04–0,6 Нмм 2 ) для мышц нижних конечностей человека, в зависимости от функции или типа волокна [33, 34 ].
Специализация в параметрах мышечной архитектуры внутри функциональных групп (т.е. определенные функциональные группы мышц, такие как разгибатели коленного сустава, демонстрируют в целом сходную ориентацию мышечных волокон и функциональные возможности расширения) ранее была продемонстрирована в опорно-двигательной системе позвоночных [27, 30, 35–41 ].Таким образом, данные об архитектуре мышц, полученные здесь для каждой мышцы, были усреднены по каждой функциональной группе в пределах каждого индивидуума, а также в пределах сгруппированных средних значений (таблицы 1, 2 и таблицы S1 – S10). Это дает общее представление о степени специализации этих функциональных групп мышц в нижних конечностях людей, участвовавших в этом исследовании, а также позволяет проводить сравнения со средними значениями аналогичных функциональных групп в предыдущих наборах данных по архитектуре (3).
Как эти переменные архитектуры мышц масштабируются в зависимости от массы тела, роста, общего объема конечности (V L ) и длины конечности (L L ) у разных людей в нашей исследуемой популяции, было протестировано с использованием линейной регрессии в GraphPad Prism (La Jolla). , Калифорния, США; www.graphpad.com). Длина конечности определялась как длина от самой проксимальной части большого вертела бедренной кости до самой дистальной части боковой лодыжки малоберцовой кости.
Результаты
Было определено среднее значение (± стандартное отклонение) in vivo архитектурных свойств для 20 мышц нижних конечностей, измеренных с помощью последовательностей DTI и T1 МРТ у 10 человек (таблица 3). Данные по архитектуре мышц для отдельных субъектов перечислены в таблицах S1 – S10.
Таблица 3.Средние (± стандартное отклонение) архитектурные характеристики 20 мышц нижних конечностей от 10 человек (5 мужчин, 5 женщин; возраст 27,3 ± 3,95 года.
Масса тела 76 ± 12,5 кг) плюс средние значения функциональной группы. L F : L м — отношение длины мышечного волокна к длине мышцы. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https: // doi.org / 10.1371 / journal.pone.0223531.t003
В среднем мышцами нижней конечности с наибольшим PCSA были латеральная широкая мышца бедра (3206 ± 1559 мм 2 ), средняя широкая мышца бедра (2938 ± 926 мм 2 ) , и камбаловидной мышцы (3226 ± 1042 мм 2 ). Мышцы с наименьшим PCSA у всех испытуемых были длинного разгибателя большого пальца стопы (196 ± 78 мм 2 ), подколенной мышцы (202 ± 76 мм 2 ) и портняжной мышцы (333 ± 84 мм 2 ).
Мышцы с самой длинной L f были в среднем; портняжная мышца (408 ± 30 мм), большая приводящая мышца (231 ± 61 мм) и двуглавая мышца бедра (длинная головка) (204 ± 38 мм).Самыми короткими мышцами L f были подколенная мышца (74 ± 14 мм), короткая приводящая мышца (76 ± 22 мм) и медиальная икроножная мышца (97 ± 22 мм). Когда длина волокна была нормализована к длине мышцы (L f : L м ), мышцы с самыми большими отношениями L f : L м были; sartorius (0,85 ± <0,01), длинная головка двуглавой мышцы бедра (0,74 ± 0,15), подколенная мышца (0,78 ± 0,17), большая приводящая мышца (0,74 ± 0,17) и латеральная широкая мышца бедра (0,59 ± 0,14). Мышцы с наименьшими L f : L m были камбаловидной (0.32 ± 0,07), медиальная икроножная мышца (0,39 ± 0,08) и прямая мышца бедра (0,39 ± 0,12).
Степень масштабирования объема мышц, длины волокон и максимальной изометрической силы в зависимости от общего объема конечности (V LM ) и длины конечности (L L ) значительно различалась между функциональными группами мышц (рисунки 2 и 3; таблица S11. ). Средний объем мышечных групп сильно масштабировался с помощью V LM (рис. 2A и 2B), хотя только сгибатели колена и разгибатели колена показали статистически значимое масштабное соотношение между F max и M L (R 2 > 0.5, р <0,05; Рис. 3C, таблица S11). Длина живота мышцы измерена с помощью L L (рис. 3A и 3B), однако L f не особенно сильно масштабировалась с L L в любой функциональной группе, при этом приводящие мышцы бедра показали самую сильную и только статистически значимую корреляцию (R 2 = 0,49, p = 0,02; рис. 3C, таблица S11).
Рис. 2. Масштабные соотношения между: индивидуальным общим объемом мышц конечности и общим объемом мышц живота в (A) приводящих, сгибателях и разгибателях колена бедра; (B) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава; и максимальная изометрическая сила в (C) приводящих мышцах бедра, сгибателях колена и разгибателях колена; (D) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава.
Пунктирные линии представляют собой среднее значение стандартной ошибки ± 1.
https://doi.org/10.1371/journal.pone.0223531.g002
Рис. 3. Масштабные соотношения между: индивидуальной длиной конечности и длиной живота мышцы в (A) приводящих, сгибателях и разгибателях колена бедра; (B) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава; и длину мышечных волокон в приводящих, сгибателях и разгибателях колена (C) бедра; (D) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава.
Пунктирные линии представляют собой среднее значение стандартной ошибки ± 1.
https://doi.org/10.1371/journal.pone.0223531.g003
Обсуждение
В этом исследовании использовалась проверенная методика для получения обширного набора данных из in vivo данных архитектуры мышц нижней конечности человека исключительно из МРТ-изображений 10 молодых здоровых людей. Эти данные определяют объем мышц, длину, оптимальную длину волокна, угол перистости волокна, PCSA и максимальную изометрическую силу. Техника использования DTI и трактографии мышечных волокон для сбора подробных данных об архитектуре мышц была ранее описана и показала свою достоверность и повторяемость [20, 42–56].В исследовании валидности метода сбора архитектуры мышц специально для моделей опорно-двигательного аппарата Чарльз и др. [20] обнаружили, что DTI может воспроизводить мышечную массу, длину волокон и PCSA в пределах 4%, 1% и 6% от соответствующих значений. переменные, измеренные при ручном вскрытии, соответственно. Точность этого метода повысила уверенность в нашей способности генерировать точный и надежный набор данных из in vivo мышечной архитектуры нижних конечностей из популяции молодых здоровых взрослых людей (таблица 2; таблицы S1 – S10).Этот набор данных основан на предыдущих попытках количественно оценить анатомию мышц нижних конечностей с помощью МРТ [6], сосредоточив внимание на сборе данных об архитектуре, необходимых для построения скелетно-мышечных моделей и моделирования.
Степень, в которой эти данные об архитектуре мышц масштабируются с антропометрическими параметрами, такими как длина конечности, объем конечности и масса тела, а также возраст, в функциональных группах мышц, может указывать на необходимость сбора такого обширного набора данных in vivo архитектуры мышц. на молодых, здоровых лицах.Объем мышц живота и длина живота достаточно хорошо соотносились с массой конечностей и длиной конечностей, соответственно, в большинстве функциональных групп. Эти соотношения масштабирования особенно очевидны в мышцах подошвенного сгиба лодыжки, где F max и объем мышц показывают сильную корреляцию с ростом пациента, массой тела, объемом конечности и длиной конечности. Эти результаты согласуются с результатами Хэндсфилда и др. [6], которые сообщили о столь же сильных масштабных взаимосвязях между объемом / длиной живота и длиной конечностей и массой тела.Однако эти данные показывают, что длина мышечных волокон плохо согласуется с длиной конечностей в любой функциональной группе мышц, что согласуется с предыдущими исследованиями [15].
В контексте моделирования опорно-двигательного аппарата это предполагает, что взаимосвязь между длиной мышечного волокна и длиной конечности не обязательно может быть точно предсказана с использованием алгоритмов масштабирования или оптимизации и может быть более сложной, чем другие переменные архитектуры мышц. Таким образом, в то время как антропометрическое масштабирование может использоваться для оценки общих анатомических свойств, таких как объем и длина мышц, визуализация анатомии нижних конечностей для конкретного субъекта, вероятно, необходима для точной оценки более сложных параметров архитектуры мышц, таких как длина мышечных волокон, особенно для использования в опорно-двигательном аппарате. моделирование.Отсутствие прямой корреляции между длиной волокон и длиной конечностей можно объяснить различиями между участниками в длине внешней сухожильной части мышечно-сухожильного блока, что, как было показано, связано с диапазоном движений сустава, особенно в дистальной части мышцы группы нижней конечности [57].
Сбор тематических данных дополнительно оправдан различиями между этими данными и ранее опубликованными данными о трупной архитектуре, например, описанными Ward et al., [3]. В то время как общие тенденции в средних характеристиках архитектуры в наших данных во многом совпадают с ранее описанными (многие из тех же мышц имеют большие значения PCSA и большие оптимальные длины волокон), можно увидеть много различий в абсолютных значениях (см. Дополнительную информацию для более подробной информации). . Учитывая анатомические вариации, наблюдаемые в нашем наборе данных, эти различия, скорее всего, связаны с разной степенью масштабирования мышечной архитектуры между функциональными группами мышц, а также с потенциальными эффектами старения.В исследовании кинематических и кинетических эффектов старения ДеВита и Хортобадьи [58] предположили, что старение приводит к перераспределению крутящего момента в суставах и мышцах по всей нижней конечности, при относительно более низком голеностопном суставе, но большем крутящем моменте в тазобедренном суставе и силе мышц у пожилых людей по сравнению с молодым людям. Хотя это, вероятно, связано с изменениями кинематики походки с возрастом, это изменение крутящего момента в суставах может также возникать в результате изменений в сложной архитектуре мышц (т. Е. Уменьшения L f : L m ), которые были показаны здесь посредством сравнений. к архитектуре трупных мышц и были особенно очевидны в дистальных мышцах.
Это подтверждает точность наших данных, однако, поскольку объемы мышц и длина волокон, согласно прогнозам, уменьшатся только на 25% и 10% соответственно из-за эффектов старения [4], анатомических изменений, в дополнение к эффектам фиксации формалина или возможным патологиям в образцы трупов в предыдущих исследованиях вскрытия [3], вероятно, является еще одной важной причиной различий между этими данными и данными о трупах. Этот высокий уровень анатомических вариаций также подтверждает потенциальную потребность в индивидуальном моделировании опорно-двигательного аппарата для клинической оценки.Индивидуализированные модели стали более распространенными [11, 12, 59–62], и с новыми методами сбора архитектуры мышц in vivo эти модели потенциально могут обеспечить более точные и надежные оценки мышечной функции по сравнению с универсальными или масштабированными универсальными моделями.
Ограничения
В то время как этот метод сбора архитектуры мышц in vivo становится все более распространенным [20, 53–56], все еще существуют некоторые ограничения, которые необходимо преодолеть перед широким использованием в сообществе моделирования и моделирования опорно-двигательного аппарата.Многие из этих ограничений, такие как предположения о том, как оценивались углы перистости и длина волокон, аналогичны тем, которые обсуждались ранее [20]. Одним из важных недостатков этого метода, который особенно важен для его приложений для моделирования мышц, является то, что невозможно оценить оптимальную длину волокон. Это часто делается в исследованиях рассечения с использованием лазерной дифракции для измерения длины саркомера [29], однако этот параметр не может быть измерен напрямую по последовательностям DTI. Хотя оптимальные длины волокон были рассчитаны здесь на основе ранее опубликованных длин саркомеров [3], сочетание DTI с дальнейшей медицинской визуализацией, такой как микроэндоскопия [63, 64], для получения in vivo длин саркомеров из поверхностных мышц может обеспечить более точные оценки длины саркомера. оптимальная длина волокна в будущих исследованиях.Без индивидуальной поправки на длину саркомера представленные здесь данные о длине волокон требуют дальнейшего тестирования и оптимизации в рамках скелетно-мышечных моделей, чтобы гарантировать, что мышцы работают на правильной части кривой сила-длина [7].
Следует отметить, что ошибка измерения / наблюдателя могла способствовать отсутствию корреляции, наблюдаемой здесь между антропометрией объекта и архитектурой мышц. Скорее всего, это повлияло на ручную сегментацию мышц для определения объемов мышц (и, соответственно, расчета F max ) и измерения углов перистости (которые мало влияют на результаты модели опорно-двигательного аппарата — см. Обсуждение ниже).Однако, поскольку определение in vivo длин мышечных волокон было в основном автоматизировано и для каждой мышцы каждого испытуемого использовались одни и те же критерии остановки волоконной трактографии, любые ошибки в этом параметре, вероятно, связаны с различиями в качестве изображения тензора диффузии изображения, а не человеческая ошибка. Поскольку ручная сегментация мышц позвоночных является часто используемым методом для измерения объема мышц [6], и все изображения тензора диффузии были предварительно обработаны с использованием одного и того же метода (см. Методы) перед анализом, влияние этих потенциальных ошибок на общую представленные здесь результаты, вероятно, были небольшими.
Точность и ограничения используемой здесь структуры волоконной трактографии обсуждались ранее [20]. Однако это оказало прямое влияние на представленные здесь данные. В то время как средняя точность оценки длины мышечных волокон составляла <1-7 мм, она варьировалась между испытуемыми и между группами мышц (2 мм в приводящих мышцах бедра, но 17 мм в разгибателях колена). Эта вариабельность может быть связана с качеством изображения тензора диффузии или артефактами частичного объема от костей или подкожно-жировой клетчатки, которые будут иметь различные эффекты в зависимости от местоположения или размера анализируемой мышцы (см. [45] для подробного обсуждения возможных источников. вариации измерения в волоконной трактографии DTI).Несмотря на это изменение, даже большие средние расхождения в точности измерения длины волокон и углов перистости в основном оказываются ниже коэффициентов повторяемости, указанных Heemskerk et al. [45] (<50 мм для L f , <10,2 ° для θ), предполагая, что этот каркас является относительно точным и воспроизводимым. Однако следует отметить, что валидация этого каркаса проводилась на трупах [20], которые не подвергались воздействию тех же физиологических факторов, которые могли повлиять на повторяемость, описанную Heemskerk et al.[45] (например, движение, артефакты дыхания или температура тела), что может объяснить относительно высокую точность, о которой сообщалось ранее. Тем не менее, хотя этот метод демонстрирует несомненный потенциал для области биомеханического моделирования, для его широкого применения необходимо улучшение согласованности трактографии волокон между мышцами и людьми.
Что касается углов перистости, это исследование сообщает о более низких углах, чем ограниченные трехмерные углы, полученные из трактографии DTI, которые в настоящее время доступны в литературе.Ранее сообщалось о значениях ~ 30 ° для камбаловидной и медиальной икроножных мышц [53, 56], по сравнению со средними значениями 12 ° и 9 °, соответственно, указанными здесь, которые больше похожи на углы, измеренные с помощью ультразвуковых или трупных данных [3, 65–67]. Хотя эти отличия от других исследований DTI кажутся существенными, известно, что угол перистости в значительной степени зависит от положения сустава, и, по оценкам, с помощью DTI он изменялся от 9 ° [56] до 46 ° [46] с поворотами на 30 ° при тыльном / подошвенном сгибании голеностопного сустава. .Хотя субъектов в этом исследовании просили оставаться в анатомическом положении во время получения изображения (с тазобедренным, коленным и голеностопным суставами при 0 ° сгибания / разгибания), возможно, что голеностопный сустав не находился точно в этом положении для продолжительность каждого сканирования. Даже небольшие отклонения от нейтрального положения в голеностопном суставе могли вызвать большие изменения в углах перистости, особенно в тыльных или подошвенных мышцах голеностопного сустава, и, следовательно, могли объяснить эти различия.Однако, учитывая низкую чувствительность прогнозов мышечной функции в скелетно-мышечных моделях к вариациям входного параметра угла перистости [10, 13], двухмерные углы перистости поверхности, представленные здесь, могут быть достаточными для прогнозирования функций мышц, если эти данные будут использоваться в качестве входы в такие модели. Тем не менее, дальнейшее уточнение этой схемы для более точной оценки оптимальной длины волокна и углов перистости может оказаться полезным в будущих исследованиях.
Воздействие и будущие исследования
Это исследование представляет собой первый экземпляр обширного набора данных из in vivo мышечной архитектуры нижних конечностей человека, созданных исключительно на основе медицинских изображений (DTI и MRI), с особым акцентом на последствиях для биомеханики и моделирования опорно-двигательного аппарата.Путем исследования масштабных соотношений между антропометрическими параметрами и важными свойствами генерирования мышечной силы, такими как длина мышечного волокна и максимальная изометрическая сила, эти данные показывают отсутствие корреляции между длиной мышечного волокна и антропометрией среди большинства функциональных групп мышц нижней конечности. Это означает, что алгоритмы оптимизации или масштабирования, часто используемые для оценки архитектуры мышц при моделировании опорно-двигательного аппарата, могут быть ненадежными, и что то, как длина мышечных волокон соотносится с пропорциями тела, может быть более сложным по сравнению с аналогичными отношениями с другими переменными архитектуры мышц.Тем не менее, учитывая различия между этими данными и ранее опубликованными данными об архитектуре трупа, возможно, что применение представленных здесь мышечных параметров к скелетно-мышечным моделям людей аналогичного возраста или антропометрии может дать более точные оценки мышечной функции, чем аналогичные данные из предыдущих исследований. . Кроме того, точные траектории мышечных волокон, восстановленные с помощью этого метода, также могут улучшить прогнозирование мышечной функции за счет более точного представления рычагов мышечного момента и могут быть включены в биомеханические модели с использованием таких методов, как описанные Chen et al., [68].
В то время как это исследование было сосредоточено на оценке архитектуры мышц у молодых, здоровых людей с использованием DTI, эта схема могла бы принести дополнительную пользу в области моделирования опорно-двигательного аппарата, измеряя аналогичные параметры в патологических популяциях (например, у людей с церебральным параличом [54], мышечной атрофией [69], мышечной дистрофия [70] и пожилые люди [71]), походка и функции мышц которых часто исследуются с помощью биомеханических моделей и симуляций [72, 73].
Кроме того, различия с предыдущими данными, а также различия в нашем наборе данных также подтверждают появление предметно-ориентированного моделирования опорно-двигательного аппарата.Хотя общие и масштабно-обобщенные модели обычно эффективны при проверке общих прогнозов опорно-двигательного аппарата, более подробный анализ моделирования, такой как прогнозирование реабилитационных или послеоперационных исходов, может потребовать включения данных по архитектуре мышц для максимальной эффективности. Дальнейшая работа будет сосредоточена на проверке этих предположений и дальнейшей валидации этой структуры. Несмотря на то, что методы, используемые здесь для измерения архитектуры мышц in vivo , были ранее проверены специально для использования в моделировании опорно-двигательного аппарата [20], до сих пор неясно, насколько точно данные будут моделировать функции мышц в рамках этих моделей.Достоверность этих методов может быть дополнительно оценена путем сравнения мышечных сил, прогнозируемых с помощью моделей опорно-двигательного аппарата для конкретных субъектов, с теми, которые измеряются экспериментально, например, с помощью изокинетического динамометра. Точные прогнозы мышечных сил на основе моделей для конкретных субъектов еще больше повысили бы уверенность в достоверности этой схемы при измерении архитектуры мышц in vivo и формировании основы для индивидуальных скелетно-мышечных моделей.
Вспомогательная информация
S1 Таблица.Архитектурные характеристики 20 мышц нижних конечностей субъекта 01 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы. L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s001
(DOCX)
S2 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 02 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы. L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения.F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s002
(DOCX)
S3 Таблица. Архитектурные характеристики 20 мышц нижних конечностей Субъекта 03 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s003
(DOCX)
S4 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 04 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s004
(DOCX)
S5 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 05 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s005
(DOCX)
S6 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 06 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s006
(DOCX)
S7 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 07 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s007
(DOCX)
S8 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 08 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s008
(DOCX)
S9 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 09 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s009
(DOCX)
S10 Таблица. Архитектурные характеристики 20 мышц нижних конечностей испытуемого 10 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышцы к длине волокна. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s010
(DOCX)
S11 Таблица. Результаты линейной регрессии (значения R
2 ) для проверки масштабных соотношений между параметрами архитектуры мышц (L f- длина мышечного волокна; L f : L м — отношение длины волокна к длине мышцы; F max- расчетная максимальная изометрическая сила; V м — объем мышц ; L м — длина мышцы) и возраст субъекта, рост, масса тела, общая масса конечности и длина нижней конечности.Значения P указаны в скобках. Значения, выделенные курсивом и жирным шрифтом, указывают на статистическую значимость (p ≤ 0,05).
https://doi.org/10.1371/journal.pone.0223531.s011
(DOCX)
S12 Таблица. Средние различия (% различий) между данными по архитектуре мышц нижних конечностей, полученными здесь из МРТ, и данными из предыдущего исследования трупов [3].
Мышечная масса по нашим данным была оценена по объему мышц. L f : L m — отношение длины мышечного волокна к длине мышцы.PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s012
(DOCX)
S2 Рис. Средние расчетные значения максимальной изометрической силы (F
max ; выражены в процентах массы тела) для каждой функциональной группы мышц для субъектов 1–10.Классификацию мышечных функциональных групп см. В таблице 1.Горизонтальная пунктирная линия представляет собой среднее значение из Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s014
(TIF)
Благодарности
Авторы хотели бы поблагодарить Chan Hong Moon за помощь в создании протоколов МРТ, используемых здесь, и Tom Gale за написание сценария MATLAB, используемого для измерения длины волокон мышечных волокон, отслеживаемых DTI. Мы также хотели бы поблагодарить трех рецензентов первоначальных версий этой рукописи, чьи конструктивные комментарии помогли значительно улучшить окончательную версию.Работа выполнена при финансовой поддержке отделения ортопедической хирургии Питтсбургского университета.
Список литературы
- 1. Либер Р.Л., Фриден Дж. Функциональное и клиническое значение архитектуры скелетных мышц. Мышечный нерв. 2000. 23 (11): 1647–66. pmid: 11054744.
- 2. Вицкевич Т.Л., Рой Р.Р., Пауэлл П.Л., Эджертон В.Р. Мышечная архитектура нижней конечности человека. Clin Orthop Relat Res. 1983; (179): 275–83. pmid: 6617027.
- 3.Ward SR, Eng CM, Smallwood LH, Lieber RL. Точны ли текущие измерения архитектуры мышц нижних конечностей? Clin Orthop Relat Res. 2009. 467 (4): 1074–82. pmid: 18972175.
- 4. Наричи М.В., Маганарис С.Н., Ривз Н.Д., Каподаглио П. Влияние старения на архитектуру мышц человека. J. Appl Physiol (1985). 2003. 95 (6): 2229–34. Epub 2003/07/08. pmid: 12844499.
- 5. Тейт CM, Уильямс GN, Барранс П.Дж., Бьюкенен Т.С. Морфология мышц нижних конечностей у юных спортсменов: анализ на основе МРТ.Медико-спортивные упражнения. 2006. 38 (1): 122–8. Epub 2006/01/06. pmid: 16394964.
- 6. Хэндсфилд Г.Г., Мейер СН, Харт Дж.М., Абель М.Ф., Блемкер СС. Связь 35 мышц нижних конечностей с ростом и массой тела количественно определена с помощью МРТ. J Biomech. 2014; 47 (3): 631–8. Epub 2013/12/26. pmid: 24368144.
- 7. Zajac FE. Мышцы и сухожилия: свойства, модели, масштабирование и применение в биомеханике и моторном управлении. Crit Rev Biomed Eng. 1989. 17 (4): 359–411. pmid: 2676342.
- 8.Сковил CY, Ронски JL. Чувствительность модели мышц Хилла к отклонениям в параметрах модели. J Biomech. 2006. 39 (11): 2055–63. Epub 2005/08/09. pmid: 16084520.
- 9. Акленд, округ Колумбия, Лин Ю.К., Панди М.Г. Чувствительность модельных прогнозов мышечной функции к изменениям в моментальных руках и свойствах мышц-сухожилий: анализ Монте-Карло. J Biomech. 2012. 45 (8): 1463–71. Epub 2012/04/14. pmid: 22507351.
- 10. О’Нил М.С., Ли Л.Ф., Ларсон С.Г., Демес Б., Стерн Дж. Т., Умбергер Б.Р.Трехмерная модель опорно-двигательного аппарата таза и задней конечности шимпанзе ( Pan troglodytes ). J Exp Biol. 2013; 216 (Pt 19): 3709–23. pmid: 24006347.
- 11.
Валенте Дж., Питто Л., Тести Д., Сет А., Делп С. Л., Стагни Р. и др. Устойчивы ли индивидуальные модели опорно-двигательного аппарата к неопределенностям в идентификации параметров? PLoS One. 2014; 9 (11): e112625. Epub 2014/11/12. pmid: 253
.
- 12. Navacchia A, Myers CA, Rullkoetter PJ, Shelburne KB.Прогнозирование нагрузок на коленный сустав in vivo с помощью глобального вероятностного анализа. J Biomech Eng. 2016; 138 (3): 4032379. pmid: 26720096.
- 13. Чарльз Дж. П., Каппеллари О., Спенс А. Дж., Уэллс Д. Д., Хатчинсон-младший. Мышечный момент рук и анализ чувствительности модели опорно-двигательного аппарата задних конечностей мыши. J Anat. 2016; 229 (4): 514–35. pmid: 27173448.
- 14. Буяльски П., Мартинс Дж., Стирлинг Л. Анализ методом Монте-Карло оценки чувствительности оценки мышечной силы к свойствам мышцы-сухожилия с использованием модели мышц на основе Хилла.J Biomech. 2018. Epub 2018/08/28. pmid: 30146173.
- 15. Уорд С.Р., Смоллвуд Л.Х., Либер Р.Л. Масштабирование архитектуры мышц нижних конечностей человека до размеров скелета. В: XX Конгресс ISB, Кливленд, Огайо. 2005.
- 16. Modenese L, Ceseracciu E, Reggiani M, Lloyd DG. Оценка параметров опорно-двигательного аппарата для масштабированных и конкретных моделей опорно-двигательного аппарата с использованием метода оптимизации. J Biomech. 2016; 49 (2): 141–8. Epub 2015/11/18. pmid: 26776930.
- 17. Манал К., Бьюкенен Т. Оценка длины провисания сухожилия по конкретным предметам: численный метод. Журнал прикладной биомеханики. 2004. 20 (2): 195–203.
- 18. Винби ЧР, Ллойд Д.Г., Кирк ТБ. Оценка различных аналитических методов для индивидуального масштабирования параметров опорно-двигательного аппарата. J Biomech. 2008. 41 (8): 1682–8. Epub 2008/05/06. pmid: 18456272.
- 19. Ву В., Ли П.В., Брайант А.Л., Галеа М., Окленд, округ Колумбия. Индивидуальное моделирование опорно-двигательного аппарата при оценке функции плечевых мышц и суставов.J Biomech. 2016; 49 (15): 3626–34. Epub 2016/09/23. pmid: 28327299.
- 20. Чарльз Дж. П., Мун СН, Андерст В. Определение данных об архитектуре мышц нижних конечностей для моделей опорно-двигательного аппарата с использованием МРТ с тензором диффузии. J Biomech Eng. 2019; 141 (6): 060905–9. pmid: 30098157.
- 21. Прейбиш С., Заальфельд С., Томанчак П. Оптимальное сшивание мозаичных трехмерных изображений при получении микроскопических изображений. Биоинформатика. 2009. 25 (11): 1463–5. Epub 2009/04/07. pmid: 19346324.
- 22. Schindelin J, Arganda-Carreras I, Frize E, Kaynig V, Longair M, Pietzsch T. и др. Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Нат методы. 2012. 9 (7): 676–82. Epub 2012/06/30. pmid: 22743772.
- 23. Jiang H, van Zijl PC, Kim J, Pearlson GD, Mori S. DtiStudio: ресурсная программа для вычисления тензора диффузии и отслеживания пучков волокон. Вычислительные методы Программы Biomed. 2006. 81 (2): 106–16. Epub 2006/01/18. pmid: 16413083.
- 24.Аха-Фернандес С., Нитхаммер М., Кубики М., Шентон М.Э., Вестин С.Ф. Восстановление данных DWI с использованием оценщика Rician LMMSE. IEEE Trans Med Imaging. 2008. 27 (10): 1389–403. Epub 2008/09/26. pmid: 18815091.
- 25. Cook P, Bai Y, Nedjati-Gilani S, Seunarine K, Hall M, Parker G, et al. Camino: Реконструкция и обработка диффузионной МРТ с открытым исходным кодом. 14-е научное собрание Международного общества магнитного резонанса в медицине, Сиэтл, Вашингтон, США, 2006 г. п. 2759.
- 26.Бодин С.К., Рой Р.Р., Медоуз Д.А., Зернике Р.Ф., Сакс Р.Д., Фурнье М. и др. Архитектурные, гистохимические и сократительные характеристики уникальной двусуставной мышцы: полусухожильной мышцы кошки. J Neurophysiol. 1982; 48 (1): 192–201. pmid: 7119845.
- 27. Чарльз Дж. П., Каппеллари О., Спенс А. Дж., Хатчинсон-младший, Уэллс ди-джей. Скелетно-мышечная геометрия, архитектура мышц и функциональные особенности задней конечности мыши. PLoS One. 2016; 11 (4): e0147669. pmid: 27115354.
- 28.Шнайдер CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 лет анализа изображений. Нат методы. 2012; 9 (7): 671–5. pmid: 22930834.
- 29. Фельдер А., Уорд С.Р., Либер Р.Л. Измерение длины саркомера позволяет с высоким разрешением нормализовать длину мышечных волокон в архитектурных исследованиях. J Exp Biol. 2005; 208 (Pt 17): 3275–9. Epub 2005/08/20. pmid: 16109889.
- 30. Мешки Р. Д., Рой Р. Р.. Архитектура мышц задних конечностей кошек: функциональное значение.J Morphol. 1982. 173 (2): 185–95. pmid: 7120421.
- 31. Хатчинсон-младший. Биомеханическое моделирование и анализ чувствительности двуногого бега. I. Сохранившиеся таксоны. J Morphol. 2004. 262 (1): 421–40. pmid: 15352201.
- 32. Медлер С. Сравнительные тенденции в сокращении скорости и производства силы в скелетных мышцах. Am J Physiol Regul Integr Comp Physiol. 2002; 283 (2): R368–78. pmid: 12121850.
- 33. Фукунага Т., Рой Р. Р., Шеллок Ф. Г., Ходжсон Дж. А., Эджертон В. Р..Удельное напряжение подошвенных сгибателей и дорсифлексоров человека. J. Appl Physiol (1985). 1996. 80 (1): 158–65. Epub 1996/01/01. pmid: 8847297.
- 34. Maganaris CN, Baltzopoulos V, Ball D, Sargeant AJ. Специфическое напряжение скелетных мышц человека in vivo. J. Appl Physiol (1985). 2001; 90 (3): 865–72. Epub 2001/02/22. pmid: 11181594.
- 35. Пауэлл П.Л., Рой Р.Р., Каним П., Белло М.А., Эджертон В.Р. Предсказуемость напряжения скелетных мышц на основе архитектурных определений задних конечностей морских свинок.J Appl Physiol Respir Environ Exerc Physiol. 1984. 57 (6): 1715–21. pmid: 6511546.
- 36. Либер Р.Л., Блевинс Ф.Т. Архитектура скелетных мышц задней конечности кролика: функциональные последствия дизайна мышц. J Morphol. 1989; 199 (1): 93–101. pmid: 2921772.
- 37. Пейн Р.К., Хатчинсон-младший, Робиллиард Дж. Дж., Смит, Северная Каролина, Уилсон А.М. Функциональная специализация анатомии тазовых конечностей у лошадей ( Equus caballus ). J Anat. 2005. 206 (6): 557–74. pmid: 15960766.
- 38.Уильямс С.Б., Уилсон А.М., Родс Л., Эндрюс Дж., Пейн Р.С. Функциональная анатомия и мышечный момент рук тазовой конечности элитного спортсмена-спринтера: гоночной борзой ( Canis ownis ). J Anat. 2008. 213 (4): 361–72. pmid: 18657259.
- 39. Уильямс С.Б., Пейн Р.С., Уилсон А.М. Функциональная специализация тазовой конечности зайца ( Lepus europeus ). J Anat. 2007. 210 (4): 472–90. pmid: 17362487.
- 40. Аллен В., Элси Р.М., Джонс Н., Райт Дж., Хатчинсон-младший.Функциональная специализация и онтогенетическое масштабирование анатомии конечностей у Alligator mississippiensis . J Anat. 2010. 216 (4): 423–45. pmid: 20148991.
- 41. Пакстон Х., Энтони Н.Б., Корр С.А., Хатчинсон-младший. Влияние селективного разведения на архитектурные свойства тазовой конечности у цыплят-бройлеров: сравнительное исследование современных и исконных популяций. J Anat. 2010. 217 (2): 153–66. pmid: 20557402.
- 42. Хсу Э. У., Мори С. Аналитические выражения для коэффициентов кажущейся диффузии ЯМР в анизотропной системе и упрощенный метод определения ориентации волокна.Magn Reson Med. 1995. 34 (2): 194–200. pmid: 7476078.
- 43. Дэймон Б.М., Динг З., Андерсон А.В., Фрейер А.С., Гор Дж. К.. Подтверждение отслеживания мышечных волокон на основе тензора диффузии с помощью МРТ. Magn Reson Med. 2002. 48 (1): 97–104. pmid: 12111936.
- 44. Deux JF, Malzy P, Paragios N, Bassez G, Luciani A, Zerbib P и др. Оценка сокращения икроножной мышцы с помощью тензорной диффузной визуализации. Eur Radiol. 2008. 18 (10): 2303–10. Epub 2008/05/09. pmid: 18463875.
- 45. Heemskerk AM, Sinha TK, Wilson KJ, Ding Z, Damon BM.Повторяемость отслеживания волокон скелетных мышц на основе DTI. ЯМР Биомед. 2010. 23 (3): 294–303. pmid: 20099372.
- 46. Sinha U, Sinha S, Hodgson JA, Edgerton RV. Архитектура камбаловидной мышцы человека под разными углами голеностопного сустава по данным магнитно-резонансной диффузной тензорной визуализации. J. Appl Physiol (1985). 2011; 110 (3): 807–19. pmid: 21164150.
- 47. Froeling M, Nederveen AJ, Heijtel DF, Lataster A, Bos C, Nicolay K и др. Диффузионно-тензорная МРТ выявляет сложную мышечную архитектуру предплечья человека.J Магнитно-резонансная томография. 2012. 36 (1): 237–48. pmid: 22334539.
- 48. Соарес Дж. М., Маркес П., Алвес В., Соуза Н. Автостопом по визуализации диффузионного тензора. Front Neurosci. 2013; 7: 31. pmid: 23486659.
- 49. Bolsterlee B, Veeger HE, van der Helm FC, Gandevia SC, Herbert RD. Сравнение измерений архитектурных параметров медиальной икроножной мышцы по ультразвуковым и диффузным тензорным изображениям. J Biomech. 2015; 48 (6): 1133–40. Epub 2015/02/16. pmid: 25682540.
- 50. Froeling M, Oudeman J, Strijkers GJ, Maas M, Drost MR, Nicolay K и др. Изменения мышц, обнаруженные с помощью диффузионно-тензорной визуализации после бега на длинные дистанции. Радиология. 2015. 274 (2): 548–62. Epub 2014/10/03. pmid: 25279435.
- 51. Дэймон Б.М., Фроилинг М., Бак А.К., Аудеман Дж., Динг З., Недервен А.Дж. и др. Тензор диффузии скелетных мышц — отслеживание волокон МРТ: обоснование, методы сбора и анализа данных, приложения и будущие направления. ЯМР Биомед.2016. Epub 2016/06/03. pmid: 27257975.
- 52. Sieben JM, van Otten I., Lataster A, Froeling M, Nederveen AJ, Strijkers GJ, et al. Реконструкция in vivo архитектуры поясничного эректора позвоночника с использованием диффузно-тензорной МРТ. Clin Spine Surg. 2016; 29 (3): E139–45. pmid: 27007789.
- 53. Болстерли Б., Финни Т., Д’Суза А., Эгучи Дж., Кларк Е.К., Герберт Р.Д. Трехмерная архитектура всей камбаловидной мышцы человека in vivo. PeerJ. 2018; 6: e4610. Epub 2018/04/24.pmid: 29682414.
- 54. Сахрманн А.С., Стотт Н.С., Безье Т.Ф., Фернандес Дж.В., Хэндсфилд Г.Г. Слабость камбаловидной мышцы при церебральном параличе: архитектура мышц выявлена с помощью диффузно-тензорной визуализации. PLoS One. 2019; 14 (2): e0205944. Epub 2019/02/26. pmid: 30802250.
- 55. Bolsterlee B, D’Souza A, Herbert RD. Надежность и надежность измерений архитектуры мышц, полученных с помощью тензорной диффузной визуализации с анатомически ограниченной трактографией. J Biomech. 2019; 86: 71–8.Epub 2019/02/12. pmid: 30739766.
- 56. Bolsterlee B, D’Souza A, Gandevia SC, Herbert RD. Как пассивное удлинение меняет архитектуру медиальной икроножной мышцы человека? J. Appl Physiol (1985). 2017; 122 (4): 727–38. Epub 2017/01/21. pmid: 28104754.
- 57. Мацукиё А., Гох А.С., Асагай Ю. Взаимосвязь между длиной мышечного сухожилия, диапазоном движений и сопротивлением пассивному движению у детей с нормальным и повышенным тонусом. J Phys Ther Sci.2017; 29 (2): 349–55. Epub 2017/03/08. pmid: 28265172.
- 58. ДеВита П., Хортобадьи Т. Возраст вызывает перераспределение суставных моментов и сил во время ходьбы. J. Appl Physiol (1985). 2000. 88 (5): 1804–11. Epub 2000/05/08. pmid: 10797145.
- 59. Scheys L, Spaepen A, Suetens P, Jonkers I. Расчетные длины момент-рука и мышца-сухожилие во время походки существенно различаются с использованием МРТ по сравнению с масштабированными стандартными скелетно-мышечными моделями нижних конечностей. Поза походки. 2008. 28 (4): 640–8.Epub 2008/06/04. pmid: 18534855.
- 60. Scheys L, Loeckx D, Spaepen A, Suetens P, Jonkers I. Нежесткая регистрация изображений на основе Атласа для автоматического определения моделей мышц линии действия: исследование для проверки. J Biomech. 2009. 42 (5): 565–72. Epub 2009/02/24. pmid: 19232618.
- 61. Scheys L, Desloovere K, Suetens P, Jonkers I. Уровень детализации предмета в скелетно-мышечных моделях влияет на расчет длины плеча тазобедренного сустава во время походки у детей с повышенной антеверсией бедренной кости.J Biomech. 2011; 44 (7): 1346–53. Epub 2011/02/03. pmid: 21295307.
- 62. Prinold JA, Mazzà C, Di Marco R, Hannah I, Malattia C, Magni-Manzoni S, et al. Модель стопы для конкретного пациента для оценки силы голеностопного сустава у пациентов с ювенильным идиопатическим артритом. Энн Биомед Eng. 2016; 44 (1): 247–57. Epub 2015/09/15. pmid: 26374518.
- 63. Чен Икс, Санчес Г.Н., Шнитцер М.Дж., Delp SL. Изменение длины саркомера латеральной широкой мышцы бедра человека при сгибании колена, измеренное с помощью микроэндоскопии in vivo.J Biomech. 2016; 49 (13): 2989–94. Epub 2016/08/03. pmid: 27481293.
- 64. Чен X, Delp SL. Длина саркомера камбаловидной мышцы человека измерена с помощью микроэндоскопии in vivo при двух углах сгибания голеностопного сустава. J Biomech. 2016; 49 (16): 4164–7. Epub 2016/11/22. pmid: 27866676.
- 65. Herbert RD, Heroux ME, Diong J, Bilston LE, Gandevia SC, Lichtwark GA. Изменения длины и трехмерной ориентации мышечных пучков и апоневрозов при пассивных изменениях длины икроножных мышц человека.J Physiol. 2015; 593 (2): 441–55. Epub 2015/01/30. pmid: 25630264.
- 66. Maganaris CN, Baltzopoulos V, Sargeant AJ. Измерения in vivo сложной архитектуры трицепса surae у человека: последствия для мышечной функции. J Physiol. 1998; 512 (Pt 2): 603–14. Epub 1998/10/09. pmid: 9763648.
- 67. Наричи М.В., Бинзони Т., Хильтбранд Э., Фазель Дж., Терьер Ф., Черретелли П. Архитектура икроножной мышцы человека in vivo с изменяющимся углом сустава в состоянии покоя и во время ступенчатого изометрического сокращения.J Physiol. 1996; 496 (Pt 1): 287–97. Epub 1996/10/01. pmid: 8
- 6.
- 68. Чен Дж. С., Басава Р. Р., Чжан Ю., Чапо Р., Малис В., Синха Ю. и др. Пиксельное моделирование скелетных мышц без сетки. Comput Methods Biomech Biomed Eng Imaging Vis. 2016; 4 (2): 73–85. Epub 2016/01/01. pmid: 28748126.
- 69. Малис В., Синха У., Чапо Р., Наричи М., Смитаман Э, Синха С. Визуализация тензора диффузии и моделирование диффузии: применение для мониторинга изменений в медиальной икроножной мышце при атрофии неиспользования, вызванной односторонним подвешиванием конечностей.J Магнитно-резонансная томография. 2019; 49 (6): 1655–64. Epub 2018/12/21. pmid: 30569482.
- 70. Ponrartana S, Ramos-Platt L, Wren TA, Hu HH, Perkins TG, Chia JM и др. Эффективность визуализации тензора диффузии в оценке тяжести заболевания при мышечной дистрофии Дюшенна: предварительное исследование. Pediatr Radiol. 2015; 45 (4): 582–9. Epub 24.09.2014. pmid: 25246097.
- 71. Sinha U, Csapo R, Malis V, Xue Y, Sinha S. Возрастные различия в индексах тензора диффузии и архитектуре волокон в медиальной и латеральной икроножной мышце.J Магнитно-резонансная томография. 2015; 41 (4): 941–53. Epub 2014/04/29. pmid: 24771672.
- 72. Розенберг М, Стил К. Моделирование воздействия ортезов голеностопного сустава на потребность в мышцах и их набор у типично развивающихся детей и детей с церебральным параличом и походкой приседания. PLoS One. 2017; 12 (7): e0180219. Epub 2017/07/14. pmid: 28704464.
- 73. Стил К.М., Сет А., Хикс Дж. Л., Шварц М. С., Дельп С. Л.. Вклад мышц в поддержку и прогресс во время ходьбы на одной конечности.J Biomech. 2010. 43 (11): 2099–105. Epub 2010/05/25. pmid: 20493489.
Фотографии анатомии шеи Кости, мышцы, нервы
Назначение позвоночника — поддерживать тело, чтобы мы могли стоять прямо. Во-вторых, он защищает спинной мозг (который является продолжением головного мозга) и все нервы, ответвляющиеся от спинного мозга.
Шейный отдел позвоночника (шея), в частности, поддерживает вес вашей головы, позволяет вам смотреть прямо и защищает спинной мозг и нервы.
Читайте дальше, чтобы узнать больше об анатомии шейного отдела позвоночника.
Мышцы шеи:
Анатомия шеи Фотографии мышц
Вокруг шеи множество мышц, которые помогают поддерживать шейный отдел позвоночника и позволяют двигать головой в разных направлениях.
Вот список многих мышц шеи.
Longus Colli & Capitis — Отвечает за сгибание головы и шеи.
Передняя прямая мышца головы — отвечает за сгибание шеи
Rectus Capitis Lateralis — Помогает шее сгибаться в стороны.
Чешуйчатые мышцы — Отвечают за подъем первого и второго ребер, то есть помогают при дыхании
Levator Scapulae — отвечает за движение лопатки (лопатки) вверх и вниз.
Rectus Capitis Lateralis — Позволяет шее сгибаться из стороны в сторону.
Obliques Capitis Superior — Позволяет шее разгибаться и сгибаться в стороны.
Obliques Capitis Inferior — Помощь при повороте головы / шеи
Шейный отдел позвоночника (шейные кости):
Кости шеи Изображение
Анатомия шейного отдела позвоночника довольно сложна.Мы попытаемся дать упрощенный обзор этой сложной анатомии.
Есть семь шейных позвонков, которые обеспечивают большую подвижность шеи.
Если смотреть сзади, у большинства людей позвоночник выглядит прямым. Однако, если смотреть на позвоночник сбоку, можно заметить отчетливые изгибы каждой части позвоночника. Назначение этих изгибов — придать позвоночнику дополнительную гибкость и способность амортизировать удары.
В шейном отделе позвоночник обычно выгибается назад.Это искривление известно как лордоз . Степень лордоза у каждого человека варьируется, но обычно находится в диапазоне 20-40 градусов. Есть много состояний позвоночника, которые могут повлиять на нормальную кривизну шейного отдела позвоночника, что приводит к боли и инвалидности. Некоторые из этих условий перечислены в конце этой статьи.
Подобно остальному позвоночнику (грудному и поясничному), каждый позвонок шейного отдела позвоночника состоит из тела, двух ножек, пластинки и множества костных выступов (называемых отростками ).Позвонки шейного отдела позвоночника самые маленькие из позвоночника, так как они выдерживают только вес головы. Отростки служат точками крепления различных связок и мышц, которые важны для устойчивости позвоночника.
Шейный отдел позвоночника состоит из 7 позвонков, пронумерованных от 1 до 7 сверху вниз, то есть C1, C2, C3, C4 и т. Д. В то время как в грудном и поясничном отделах позвоночника анатомия позвонков относительно одинакова для каждого позвонка, шейного отдела позвоночника. анатомия позвоночника весьма разнообразна.Ниже приводится описание каждого из позвонков:
C1 — называется tlas. Череп лежит прямо на нем. Это самый верхний из шейных позвонков. Он имеет кольцевую структуру. Около 50% сгибания / разгибания шеи происходит в суставе C1 / затылочного бугра (черепа).
C2 — ось . Он имеет специальный процесс, называемый dens , который действует как точка поворота, на которой включается атлас (C1). Около 50% вращения шеи происходит между C1-C2 в атланто-осевом суставе .
C3- C7- Позвонки в этой области очень похожи по своему строению на остальной позвоночник. Остальные движения шеи происходят одинаково между каждым из этих позвоночных уровней.
Нервы шейного отдела позвоночника:
Анатомия шеи Изображение нервов
От шейного отдела позвоночника отходят 8 спинномозговых нервов.
Большинство этих нервов контролируют функции верхних конечностей и позволяют чувствовать руки, плечо и затылок.Каждый нерв обеспечивает ощущение определенной области тела, называемой дерматомом.
Возможно, наиболее важно то, что несколько нервов, которые берут начало в шейном отделе позвоночника, контролируют диафрагму , мышцу, которая позволяет нам дышать. Таким образом, серьезная травма шеи может привести к катастрофическим последствиям.
Заболевания шейного отдела позвоночника
- Остеопороз
- Остеоартроз
- Стеноз шейки матки
- Радикулопатия шейки матки
- Грыжа шейного отдела позвоночника
- Цервикальная миелопатия
- Задняя продольная окостеневшая связка
- Болезнь межпозвонковых дисков
Не волнуйтесь из-за этого вирусного изображения «молочного протока», это не совсем правильно
Яркое анатомическое изображение «женской мышечной системы» и «молочных протоков» стало популярным благодаря вирусному твиту, появляющемуся повсюду от Buzzfeed до BBC.Но если вас пугает это внезапное открытие внутреннего мира человеческой груди, не волнуйтесь — изображение и его контекст не совсем правильные.
Драматическая картина впервые появилась в Интернете в этом посте Imgur 2016 года, но, как и в случае с большинством вирусного контента, постер не был оригинальным автором изображения. На самом деле это был скриншот из этого малоизвестного приложения для iPad; и нет, это , а не женская мышечная система:
(Снимок экрана / Анатомия и физиология — анатомия частей человеческого тела)
Во-первых, грудь фактически не содержит скелетных мышц.(Там есть некоторые клетки и ткани, которые вроде как гладкие мышцы, но это не то, что, как утверждается, изображено на этой диаграмме.)
Итак, что это за формы, похожие на лепестки? В описании приложения красные капли неопределенно называются «молочной железой», что бесполезно, поскольку вся грудь на самом деле является молочной железой.
(Снимок экрана / Анатомия и физиология — анатомия частей человеческого тела)
Вот как это на самом деле устроено. Расположенная в изрядной дозе жира, ткань груди содержит ряд клеток, ответственных за выработку молока, когда гормоны человека сигнализируют о том, что он готов к кормлению новорожденного.
Эти клетки, называемые альвеолами, группируются в комки, известные как дольки, которые, в свою очередь, соединяются с соском через пучок млечных протоков (или молочных протоков), узких каналов, которые в основном направляют молоко к мундштуку ребенка — соску.
Итак, эти штуки в форме цветов определенно не воздуховоды. Вероятно, они представляют собой дольки, за исключением того, что в настоящей человеческой груди они вообще не расположены таким аккуратным узором.
Многие интернет-пользователи отреагировали на картинку как на какое-то безумное упущение на уроках анатомии в школе, хотя, вероятно, они столкнулись с чем-то более похожим на один из этих:
(MedicineNet)
(ER Services)
(Macea & Fregnani, Int.J. Morphol., 2006)
Как видите, распределение долек гораздо более разнообразно и беспорядочно, чем показано на 3D-графике. Поскольку приложение по анатомии не является работой авторитетного медицинского или образовательного издателя (на самом деле, мы вообще не смогли найти в нем никаких научных ссылок!), Разумно отнестись к его изображениям с недоверием.
Кроме того, форма и размер долек различаются в зависимости от того, кормит женщина или нет — важная деталь, которая также не указана в исходном материале или любой из его вирусных итераций.
Кроме того, есть по крайней мере одно «развенчание», которое тоже ошибается. В сообщении на научно-популярной странице в Facebook утверждается, что «фотография [sic] основана на молочных протоках, извлеченных из трупов в 1800-х годах, введенных воском и расположенных в аккуратном круге».
За исключением того, что исходное изображение вскрытия с введением воска в 1840 г. мало похоже на вирусную 3D-визуализацию:
(Рамзи и др., Journal of Anatomy, 2005) грудь.Перед диссекцией в протоки вводили цветной воск (от Cooper, 1840).
(Если вы хотите увидеть больше 1840 изображений женской и мужской груди сэром Астли Пастоном Купером, они находятся в свободном доступе здесь.) женская грудь упоминалась в современных исследованиях по этой теме — предмет, который сложно исследовать, поскольку большинство методов медицинской визуализации не рекомендуется использовать у кормящих матерей.
Страница поддержки грудного вскармливания в Facebook, критикующая 3D-рендеринг, ссылается на этот факт и подчеркивает ультразвуковое исследование 2005 года ткани груди кормящей женщины как лучшее представление о том, что на самом деле происходит внутри наших молочных желез.
(Ramsay et al., Journal of Anatomy, 2005)
Вверху: изображение общей анатомии кормящей груди на основе ультразвуковых наблюдений системы молочных протоков и распределения различных тканей в груди.
Это научное изображение явно менее жуткое и, следовательно, имеет гораздо меньший вирусный потенциал — но оно намного точнее, чем странные цветы из плоти, пугающие Интернет. Пожалуйста.
Мишель Старр внесла свой вклад в этот отчет.
Примечание редактора: В более ранней версии этой статьи утверждалось, что грудь не содержит ни одной мышцы . Это было уточнено, чтобы указать, что грудь не содержит, в частности, скелетных мышц.
Обзор мышечной системы
Взаимодействие скелетных мышц
Скелетные мышцы взаимодействуют, чтобы производить движения посредством анатомического позиционирования и скоординированного суммирования сигналов иннервации.
Цели обучения
Объясните суммарное взаимодействие скелетных мышц и то, как они влияют на движение
Основные выводы
Ключевые моменты
- Сокращения мышц можно назвать подергиванием, суммированием или столбняком.
- Подергивание — это период
сокращения и расслабления мышцы после однократной стимуляции. - Суммирование — это
случаев возникновения дополнительных сокращений перед тем, как предыдущие сокращения полностью расслабились. - Суммирование может быть достигнуто путем увеличения частоты стимуляции или привлечения дополнительных мышечных волокон в мышцу.
- Столбняк
возникает, когда частота мышечных сокращений такова, что максимальная сила возникает при напряжении без какого-либо расслабления мышцы.
Ключевые термины
- столбняк : Когда частота мышечных сокращений такова, что максимальная сила создается напряжением без какого-либо расслабления мышцы.
- суммирование : Возникновение дополнительных сокращений перед тем, как предыдущие сокращения полностью расслабились.
- подергивание : период сокращения и расслабления мышцы после однократной стимуляции.
Сокращения скелетных мышц можно сгруппировать по длине и частоте сокращений.
Twitch
При стимуляции одним потенциалом действия мышца сокращается, а затем расслабляется. Время между стимулом и началом сокращения называется латентным периодом, за которым следует период сокращения. При пиковом сокращении мышца расслабляется и возвращается в исходное положение. Взятые вместе, эти три периода называются подергиванием.
Сокращение мышц : Время между стимуляцией и сокращением называется латентным периодом.После сокращения мышца снова расслабляется до уровня напряжения покоя. Вместе эти три периода образуют одно мышечное сокращение,
.Суммирование
Если бы дополнительный потенциал действия должен был стимулировать сокращение мышцы до того, как предыдущее мышечное сокращение полностью расслабилось, то он суммировался бы с этим предыдущим сокращением, увеличивая общее количество напряжения, производимого в мышце. Это сложение называется суммированием. Суммирование мышц может происходить между моторными единицами, чтобы задействовать больше мышечных волокон, а также внутри моторных единиц за счет увеличения частоты сокращения.
Суммирование нескольких волокон
Когда центральная нервная система посылает слабый сигнал о сокращении мышцы, сначала стимулируются меньшие двигательные единицы, будучи более возбудимыми, чем более крупные. По мере увеличения силы сигнала возбуждаются все больше (и более крупные) двигательные единицы. У самых крупных моторных единиц сократительная сила в 50 раз выше, чем у меньших; таким образом, по мере того, как активируется все больше и больше двигательных единиц, сила сокращения мышц становится все сильнее.Концепция, известная как принцип размера, позволяет градации силы мышц во время слабого сокращения происходить небольшими шагами, которые постепенно увеличиваются по мере того, как требуется большее количество силы.
Суммирование частот
Для скелетных мышц сила, оказываемая мышцами, может контролироваться путем изменения частоты, с которой потенциалы действия посылаются к мышечным волокнам. Потенциалы действия не поступают в мышцы синхронно, и во время сокращения только определенный процент волокон в мышце будет сокращаться в любой момент времени.В типичных обстоятельствах, когда человек прилагает столько мышечной силы, сколько он сознательно способен, примерно одна треть волокон в этой мышце будет сокращаться одновременно. Этот относительно низкий уровень сокращения является защитным механизмом для предотвращения повреждения мышечной ткани и прикрепления сухожилий и структур.
Столбняк
Если частота генерируемых потенциалов действия увеличивается до такой степени, что напряжение мышц достигает своего пика и выходит на плато и не наблюдается расслабления, то сокращение мышц описывается как столбняк.
Суммирование и сокращения столбняка : Повторяющиеся сокращения, когда предыдущие сокращения не расслабились полностью, называются суммированием. Если частота этих сокращений увеличивается до точки, при которой создается максимальное напряжение и не наблюдается расслабления, то сокращение называется столбняком.
Как называются скелетные мышцы
Анатомическое расположение пучков скелетных мышц можно описать как параллельное, сходящееся, перистое или сфинктерное.
Цели обучения
Различать параллельные, перистые, конвергентные и сфинктерные типы мышц
Основные выводы
Ключевые моменты
- Параллельные мышцы являются наиболее многочисленными и типичными, пучки которых расположены параллельно друг другу.
- Конвергентные мышцы похожи на параллельные мышцы в прикреплении, хотя пучки не идут параллельно друг другу, образуя более широкую мышцу.
- В перистой мышце сухожилие проходит по всей длине мышцы, при этом пучки прикрепляются под углом.
- Мышцы сфинктера характеризуются круговым расположением пучков вокруг отверстия. При сокращении отверстие становится меньше.
Ключевые термины
- Параллельно : мышца с общей точкой прикрепления, пучки которой проходят параллельно друг другу.
- Круговой : Кольцевая полоса мышц, которая окружает отверстие тела, сужается и расслабляется для контроля потока.
- Pennate : мышца в форме пера с пучками, которые косо (под углом) прикрепляются к центральному сухожилию.
- Конвергентная : мышца с общей точкой прикрепления, хотя отдельные пучки не обязательно проходят параллельно друг другу.
Скелетные мышцы можно разделить на четыре группы в зависимости от их анатомического расположения.
Параллельный
Параллельные мышцы характеризуются пучками, которые проходят параллельно друг другу, и сокращение этих групп мышц действует как продолжение сокращения одного мышечного волокна. Большинство скелетных мышц тела — это параллельные мышцы; хотя их можно увидеть в различных формах, таких как плоские полосы, в форме веретена, а некоторые могут иметь большие выступы в середине, известные как брюшко мышцы.
Параллельные мышцы можно разделить на веретенообразные и нефузиформные типы в зависимости от их формы. Веретенообразные мышцы имеют более веретенообразную форму (их диаметр в центре больше, чем на обоих концах), тогда как неправильные мышцы имеют более прямоугольную форму с постоянным диаметром.
Двуглавая мышца плеча является примером веретенообразной параллельной мышцы и отвечает за сгибание предплечья.
сходящийся
Конвергентные мышцы имеют общую точку прикрепления, от которой мышечные пучки отходят наружу, не обязательно в определенной пространственной структуре, позволяя мышце покрывать широкую поверхность.Эти мышцы не оказывают такого большого усилия на свои сухожилия. Мышечные волокна во время сокращения часто могут оказывать противоположное воздействие, например, не тянуть в одном направлении, в зависимости от расположения мышечного волокна. Эти волокна, покрывающие широкую поверхность, позволяют совершать более разнообразные движения. Поскольку пучки натягивают сухожилия под углом, они не перемещают сухожилие до их параллельных мышечных аналогов. Несмотря на это, они создают большее напряжение, поскольку обладают большим количеством мышечных волокон, чем параллельные мышцы аналогичного размера.
Большая грудная мышца, расположенная в груди, является примером сходящейся мышцы и отвечает за сгибание плеча.
Пеннат
В Pennate мышцах сухожилие проходит по всей длине мышцы. Фасцикулы натягивают сухожилие под углом, таким образом, не перемещаясь так далеко от параллельных мышц во время сокращения. Однако в этих мышцах обычно относительно больше мышечных волокон, чем в параллельных мышцах аналогичного размера, и поэтому они несут большее напряжение.
Если все пучки перистой мышцы находятся на одной стороне сухожилия, перистая мышца называется монопородной.Если пучки лежат по обе стороны от сухожилия, мышца называется двуплодной. Если центральное сухожилие разветвляется внутри перистой мышцы, эта мышца называется многоплодной.
Прямая мышца бедра, находящаяся в бедре и отвечающая за его сгибание, является примером двуплодной мышцы.
Циркуляр
Волокна круговых мышц или мышц сфинктера расположены концентрически вокруг отверстия или углубления. По мере того как мышца сокращается, отверстие, которое она обходит, становится меньше.По этой причине эти мышцы часто встречаются на входах и выходах внешних и внутренних проходов. Скелетные круговые мышцы отличаются от эквивалентов гладких мышц своей структурой и тем, что они находятся под произвольным контролем
Orbicularis oris, контролирующая открывание рта, является примером круговой мышцы.
Типы мышц тела : четыре типа мышц; параллельные (веретенообразные и не веретенообразные), круглые, сходящиеся и перистые (однотонные, би и мульти).
Как скелетные мышцы производят движения
Мышцы организованы в группы из агонистов, антагонистов и синергистов, которые производят и модулируют движение.
Цели обучения
Различение мышц-агонистов и мышц-антагонистов
Основные выводы
Ключевые моменты
- Мышцы-агонист укорачиваются при сокращении, чтобы произвести движение.
- После сокращения мышца-антагонист, соединенная с мышцей-агонистом, возвращает конечность в предыдущее положение.
- Мышцы-синергисты действуют вокруг подвижного сустава, производя движение, аналогичное или согласованное с мышцами-агонистами, что обеспечивает широкий диапазон возможных движений.
Ключевые термины
- антагонист : Этот тип мышц действует как мышца, противостоящая агонистам, обычно сокращаясь, чтобы вернуть конечность в исходное положение покоя.
- агонист : Эти мышцы обычно связаны с самим движением и иногда называются первичными двигателями.Они сокращаются, в то время как другая мышца расслабляется.
- синергист : Этот тип мышц действует вокруг подвижного сустава, вызывая движение, аналогичное или согласованное с мышцами-агонистами.
Мышцы существуют в группах, которые производят движения за счет сокращения мышц. Мышцы классифицируются в зависимости от их действий во время сокращений как агонисты, антагонисты или синергисты.
Для пар мышц, называемых антагонистическими парами, одна мышца обозначается как мышца-разгибатель, которая сокращается, чтобы открыть сустав, и мышца-сгибатель, которая действует напротив мышцы-разгибателя.Эти пары существуют в местах тела, в которых тело не может вернуть конечность в исходное положение из-за простого отсутствия сокращения. Типичные пары мышц включают двуглавую мышцу плеча и трехглавую мышцу плеча, которые сгибают или разгибают предплечье.
Мышцы-агонисты
Мышцы-агонисты — это мышцы, которые мы обычно ассоциируем с самим движением, и поэтому их иногда называют первичными двигателями. Мышцы-агонисты производят основное движение или серию движений за счет собственных сокращений.Чтобы вызвать движение, мышцы-агонисты должны быть физически расположены так, чтобы они пересекали сустав через сухожилие. Сокращение будет двигать конечностями, связанными с этим суставом. В этом смысле кость действует как рычаг при сокращении прикрепленного мышечного волокна, приводя в движение движение.
Во время сгибания предплечья двуглавая мышца плеча является мышцей-агонистом, подтягивая предплечье к плечу.
Мышцы-антагонисты
Большинство мышц сгруппированы в пары, с антагонистом каждой мышцы-агониста.Исключение составляют такие мышцы, как мышцы сфинктера, которые сокращаются способом, противоположным состоянию покоя. Мышцы-антагонисты действуют как мышцы, противостоящие агонистам, обычно сокращаясь, чтобы вернуть конечность в исходное положение покоя.
Во время сгибания предплечья трехглавая мышца плеча является мышцей-антагонистом, сопротивляющейся движению предплечья вверх по направлению к плечу.
Мышцы-синергисты
Мышцы-синергисты действуют вокруг подвижного сустава, производя движение, аналогичное или согласованное с мышцами-агонистами.Они часто действуют, чтобы уменьшить чрезмерную силу, создаваемую мышцей-агонистом, и называются нейтрализаторами. Синергисты полезны, потому что они фиксируют определенные суставы, чтобы разрешить диапазон сокращений, в отличие от явной силы сокращения агонистов, ограничивающей диапазон возможных движений.
Во время сгибания предплечья плечевые и плечевые мышцы действуют как мышцы-синергисты, помогая двуглавой мышце плеча подтягивать предплечье к плечу. Мышцы вращающей манжеты также являются синергистами в том смысле, что они фиксируют плечевой сустав, позволяя двуглавой мышце плеча проявлять большую силу.
Сгибание предплечья двуглавой мышцей плеча : Двуглавая мышца плеча является агонистом или движителем праймера, ответственным за сгибание предплечья. Трехглавая мышца плеча (не показана) действует как антагонист. Brachioradialis и brachialis являются мышцами-синергистами, а вращающая манжета (не показана) фиксирует плечевой сустав, позволяя двуглавой мышце плеча прикладывать большую силу.
Мест прикрепления мышц
Сухожилия состоят из соединительной ткани, которая прикрепляет мышцы к кости.
Цели обучения
Опишите функцию сухожилий
Основные выводы
Ключевые моменты
- Сухожилия обладают эластичностью, что позволяет им выдерживать натяжение и действовать как пружины.
- Сухожилия в основном состоят из плотно упакованных коллагеновых волокон, идущих параллельно силе, создаваемой мышцами, к которым они прикреплены.
- Сухожилия также содержат волокна эластина для улучшения эластичных свойств и протеогликаны, которые поддерживают организацию сухожилий при растяжении и сжатии.
- Апоневрозы — это большие плоские листы соединительной ткани, похожие на сухожилия. Они отвечают за связывание мускулов с костью и фасциями других мускулов.
Ключевые термины
- сухожилие : плотная полоса фиброзной ткани, которая обычно соединяет мышцу с костью.
- апоневрозы : плотный плоский лист фиброзной ткани, который соединяет мышцу с костями или фасциями других мышц.
Большая часть скелетных мышц прикрепляется к кости для движения.Однако некоторые скелетные мышцы прикрепляются непосредственно к другим мышцам, фасциям или тканям, например к коже.
Сухожилия
Ахиллово сухожилие : Ахиллово сухожилие обеспечивает стабильность и ограничивает диапазон движений в голеностопном суставе. Это самое толстое и сильное сухожилие в теле. Сухожилия — это обычная ткань, соединяющая мышцы с костью.
Сухожилие представляет собой шнуровидную волокнистую соединительную ткань, которая соединяет мышцу с костью и способна выдерживать напряжение.На любом конце сухожилия его волокна переплетаются с фасцией мышцы или надкостницей (плотное волокнистое покрытие кости), позволяя рассеивать силу по кости или мышце.
Сухожилия в основном состоят из плотно упакованных коллагеновых волокон, идущих параллельно силе, создаваемой мышцами, к которым они прикреплены. С коллагеновыми волокнами переплетены молекулы эластина, улучшающие эластичность сухожилий, и различные протеогликаны, белки, к которым присоединены многие молекулы углеводов.Эти белки играют ключевую роль в поддержании организации сухожилия, особенно во время сжатия и разгибания.
Когда-то считалось, что сухожилия играют только пассивную связующую роль. Однако исследования их упругих свойств показали, что они также могут действовать как пружины. Эластичность сухожилий позволяет им пассивно накапливать энергию для последующего высвобождения. Наиболее широко исследуемым примером является ахиллово сухожилие, которое накапливает и высвобождает упругую энергию во время ходьбы, повышая эффективность и снижая мышечную нагрузку.
Апоневрозы
Не все мышцы прикрепляются сухожилиями. Апоневрозы — это большие листообразные слои соединительной ткани, по составу похожие на сухожилия. Апоневрозы также могут прикрепляться к костям, например апоневрозам волосистой части головы, и к фасциям других мышц или тканей, например к передним апоневрозам живота. Их большая форма и форма обеспечивают структуру и распределяют напряжение по более широкой площади или большому количеству групп мышц.
Другое навесное оборудование
Мышцы также могут прикрепляться непосредственно к другим тканям, что особенно заметно на лице.Скелетные мышцы, участвующие в контроле экспрессии, прикрепляются непосредственно к фасции кожи.
Расположение пучков
Скелетные мышцы сгруппированы в пучки, которые представляют собой пучки мышечных волокон, окруженных перимизием.
Цели обучения
Очертание строения мышечного пучка
Основные выводы
Ключевые моменты
- Скелетная мышца окружена толстым соединительным листом, называемым фасцией
- Под ним находится еще один слой соединительной ткани, называемый эпимизием, который простирается внутрь в виде перимизиума в мышцу, разделяя волокна на пучки, называемые пучками.
- Каждый пучок окружен другим слоем соединительной ткани, называемым эндомизием.
- Эта структура разделяет и защищает мышцы, а также распределяет силу по мышцам, предотвращая повреждения.
Ключевые термины
- перимизий : продолжение эпимизия в мышцу, разделяющее волокна на пучки.
- эпимизий : лист соединительной ткани, лежащий ниже фасции, также окружающий мышцу.
- фасция : лист толстой соединительной ткани, окружающий мышцу.
- эндомизий : лист соединительной ткани, который обертывает каждый пучок.
- пучок : Группа мышечных волокон, окруженных перимизием.
Мышечная фасция
Структура мышцы : Скелетная мышца окружена толстым внешним слоем соединительной ткани, называемым фасцией. Внутри него находится слой, называемый эпимизием, который расщепляется внутрь на мышцу как перимизий, разделяющий мышечные волокна на группы, называемые пучками.Каждый пучок окружен другим слоем соединительной ткани, называемым эндомизием.
Ткань скелетных мышц состоит из многочисленных мышечных волокон, которые отделены от прилегающих мышц и других тканей слоем плотной эластичной соединительной ткани, называемой фасцией. Эта фасция может выступать за конец мышцы и прикрепляться к костям, другим мышцам и другим тканям. Ключевые группы мышц и связанные с ними сосудистая и нервная системы также могут быть отделены от других тканей, например, в плече.Эти группы называются фасциальными отделениями.
Эта фасция взаимосвязана с серьезными фасциями, встречающимися по всему телу, включая поверхностную фасцию, которая является самым нижним слоем кожи, и висцеральную фасцию, окружающую внутренние органы. Фасция, окружающая мышцу или группу мышц, не содержит многих кровеносных сосудов, но богата сенсорными рецепторами.
Мышечная фасция преимущественно состоит из поперечно сшитых волокон коллагена и эластина, ориентированных параллельно направлению мышечной силы, что позволяет им противостоять силам высокого напряжения, оставаясь при этом в некоторой степени эластичными.
Волосы
Под фасцией скелетных мышц находится еще один слой соединительной ткани, называемый эпимизием, который тесно связан с фасцией. Он простирается внутрь и становится перимизием, затем в мышцу, разделяя мышечные волокна на небольшие пучки, называемые пучками. Фасцикулы могут быть расположены в различных анатомических положениях внутри мышцы, производя различные движения.
Каждое отдельное волокно в пучке окружено тонким соединительным слоем, называемым эндомизием, который помогает поддерживать тесную связь между мышечным волокном и соответствующими сосудистыми и нервными системами.
Организация соединительной ткани по всей мышце и вокруг нее обеспечивает силу и гибкость при равномерном распределении силы. Он также поддерживает тесную связь сосудистой и нервной системы с мышцами, которая необходима для доставки необходимых метаболитов и нервных импульсов.
Сердечная ткань и гладкая мышечная ткань
Хотя сердечные и гладкие мышцы также обернуты соединительной тканью, они не дифференцируются так же, как скелетные мышцы.
Рычажные системы
Расположение мышц позволяет им двигаться относительно друг друга, в то время как соединительный элемент действует как точка поворота для рычажной системы.
Цели обучения
Различать сгибатели и разгибатели, а также приводящие и отводящие мышцы
Основные выводы
Ключевые моменты
- Для описания действия скелетных мышц можно использовать простую рычажную систему. Рычаги состоят из оси, неподвижного стержня и нагрузки, к которой приложена сила.
- Относительные положения оси поворота, нагрузки и силы определяют тип рычага и последующее действие.
- Сгибатели и разгибатели регулируют угол между двумя частями тела. Сгибание уменьшает угол, а разгибание увеличивает угол.
- Отведение и приведение — это движения относительно средней линии тела. Отведение — это движение от этой средней линии, а приведение — это движение к средней линии.
- Внутреннее вращение перемещает дистальные части конечностей внутрь к средней линии.Наружное вращение — это противоположное движение, указывающее дистальную часть конечностей от средней линии.
- Возвышение конечности или части тела перемещает ее в верхнем направлении, в то время как депрессия перемещает части тела в нижнем направлении.
Ключевые термины
- отведение : Движение от средней линии тела.
- вращение : Акт поворота вокруг центра или оси.
- Первоклассный рычаг : Усилие и нагрузка по обе стороны от оси.
- рычаг второго класса : Нагрузка между усилием и стержнем.
- рычаг третьего класса : Усилие между шарниром и грузом.
- происхождение : Относительно неподвижная точка прикрепления мышцы.
- насадка : подвижная точка прикрепления мышцы.
- flexion : Уменьшает угол между двумя частями тела.
- удлинитель : Увеличивает угол между двумя частями тела.
- приведение : Движение к средней линии тела.
Скелетная мышца обычно прикрепляется к относительно неподвижной части тела на одном конце и к более подвижной области на другом конце сустава. Крепление на неподвижном конце называется исходной точкой, а на подвижном конце — вставкой. При сокращении вставка тянется к исходной точке. Это движение можно описать с помощью простой рычажной системы. Мышцы могут иметь несколько источников и прикреплений, которые модулируют тип движения, которое они производят.
Рычаги
Рычаг состоит из трех частей: неподвижного стержня, прикрепленного к точке опоры (оси), и груза.В зависимости от относительного положения трех компонентов рычаги могут перемещать тяжелые грузы или перемещать грузы дальше или быстрее при приложении силы.
Если нагрузка находится близко к оси поворота, а сила приложена далеко от оси, то считается, что рычаг работает с механическим преимуществом. Большая, но относительно небольшая сила может переместить тяжелый объект. Классический пример такого рычага — автомобильный домкрат. При больших перемещениях рычага тяжелый автомобиль небольшими приращениями поднимается от земли.
Если нагрузка находится далеко от оси поворота и сила приложена рядом с ней, то считается, что рычаг работает с механическими недостатками.Для перемещения относительно небольшого груза требуется большая сила, но скорость и расстояние, на которое этот груз может перемещаться, значительно увеличиваются. Примером этого является сочетание лопаты с сильными движениями мышц руки, что приводит к большим движениям головки лопаты.
В мышцах суставы являются стержнями, а кости — неподвижными стержнями. Нагрузка — это вес кости, связанных тканей и других перемещаемых объектов, а сила прилагается мышцей в точке ее прикрепления.
Классы рычагов
Рычагитакже могут изменяться в зависимости от относительного положения нагрузки, оси поворота и точки приложения силы. Классы рычагов включают:
Первоклассный рычаг
В рычаге первого класса нагрузка и усилие располагаются по обе стороны от оси, как качели. Первоклассные рычаги относительно редко встречаются в организме, но одним из примеров является трехглавая мышца плеча плеча, которая действует для разгибания предплечья. Сила прилагается в точке ее приложения к локтевой кости предплечья, локоть является стержнем, а нагрузка — плечевой костью в плече.Думая об относительном расстоянии между точками прикрепления, можно сказать, что трехглавая мышца плеча имеет механический недостаток.
Рычаг второго класса
В рычаге второго класса сила прилагается к одному концу, шарнир — на другом, а нагрузка находится между ними. Рычаги второго класса тоже относительно редки в кузове. Один из примеров — приподняться на цыпочках. Ось опирается на переднюю часть стопы, нагрузка — это вес тела, а сила прилагается через ахиллово сухожилие в пятке.Все рычаги второго класса в корпусе действуют механически, поскольку сила всегда прикладывается ближе к нагрузке, чем к оси.
Рычаг третьего класса
В рычаге третьего класса сила приложена между грузом и стержнем. Большинство мышц тела — это рычаги третьего класса, и все они действуют с механическими недостатками, поскольку сила прилагается ближе к оси вращения, чем нагрузка. Примером рычага третьего класса в теле является двуглавая мышца плеча, которая сгибает предплечье по направлению к плечу.Начинающаяся от лопатки, точка опоры — это локоть, при этом сила, прикладываемая сразу после локтя в точке прикрепления к радиусу предплечья. Нагрузка — это предплечье и любые предметы, которые несет человек.
Типы движения
Работая вместе, используя принципы рычага, описанные выше, скелетные мышцы могут вызывать широкий диапазон движений.
Сгибатели и разгибатели
Сгибатели и разгибатели регулируют и изменяют угол между двумя частями тела.Сгибание уменьшает угол, а разгибание увеличивает угол. Например, разгибание руки открывает угол в локтевом суставе, а сгибание позволяет сгибать руку. Сгибание также может двигаться внутрь к телу или вперед, например, бедрами или плечами. Разгибание в этом случае перемещает конечности к задней стороне рычага тела.
Похищение и приведение
Отведение и приведение — это движения относительно средней линии тела. Отведение — это движение от средней линии тела, а приведение — это движение к этой линии.Например, отведение рук или ног в сторону от тела — это отведение, а возвращение конечностей к средней линии — приведение.
Внутреннее вращение
Внутренняя или медиальная ротация характерна для плеча или бедра и приводит дистальные части конечностей внутрь к средней линии. Внутреннее вращение также может перемещать плечевую и бедренную кость внутрь. Наружное или латеральное вращение — это противоположное движение, указывающее дистальную часть конечностей, а также плечевую и бедренную кость от средней линии.
Депрессия и возвышение
Конечность или часть тела могут перемещаться вверх (или в более высоком направлении) посредством возвышения. Например, трапеция поднимает верхушку плеча вверх. Депрессия противоположна возвышению или движению частей тела в нижнем направлении.
Типы движений тела : Расположение мышц вокруг сустава определяет тип производимого движения.
Онлайн-визуализатор анатомии тела зиготы 3D
Онлайн-визуализатор анатомии тела зиготы | Анатомия человека 3DZygoteBody ™ и 3D-данные © 2012-2017 Zygote Media Group, Inc.
Краткое руководствоНавигация
- Щелкните и перетащите мышью, чтобы повернуть, прокрутите для увеличения.
- Или используйте кнопки в верхнем левом углу. Кнопка «Домой» сбрасывает вид.
- Перейдите из режима капсулы в режим орбиты в правом верхнем углу, чтобы включить полное 3D Поверните на
- и удерживайте Ctrl, чтобы панорамировать вид.(Только для премиум-пользователей)
- Используйте ползунок непрозрачности слева, чтобы отобразить слои.
- Щелкните переключатель под ползунком, чтобы управлять слоями по отдельности.
- Используйте поле поиска в правом верхнем углу для поиска или щелкните объекты, чтобы выбрать их.
- Щелкните фон или X, чтобы отменить выбор.
- Ctrl + щелчок по объектам, чтобы быстро скрыть объекты.(Cmd + щелкните на Mac.)
- Shift + щелкните объекты или метки (или щелкните значок «булавка» на этикетке), чтобы закрепить объект. Это будет держать его выбранным, пока вы выбираете больше.
- Используйте значок видимости на имени объекта, чтобы скрыть элемент. В отличие от Ctrl + щелчка по объекту, инструмент видимости будет скрывать элементы до тех пор, пока не будет нажата кнопка «Показать все» в правом верхнем углу.
- Используйте инструмент «Увеличить до» на имени элемента, чтобы центрировать изображение на элементе.
- Используйте стрелку вниз на имени элемента, чтобы отобразить информацию об элементе.
- Используйте инструмент «Закрепить» на имени элемента, чтобы закрепить элемент, позволяя выбрать несколько элементов.
- Используйте значок выбора модели над ползунком анатомии слева для загрузки различных моделей.
- Мои сцены позволяет загружать и сохранять созданные вами сцены.Все аннотации, булавки и видимые элементы будут сохранены.
- Zygote Scenes — это коллекция сцен, созданная Zygote Media Group, с аннотациями, определяющими анатомические ориентиры.
- Иерархия — это список всех объектов, которые есть в вашей текущей сцене. Вы можете включать и выключать детали, используя галочки у каждого объекта или группы объектов.
- Аннотации позволяет вам создавать свои собственные заметки и маркеры, которые будут сохранены при сохранении вашей сцены.
- инструментов:
- Pick возвращает вас в режим по умолчанию для выбора деталей и поворота камеры.
- Slice позволяет вам разрезать части сцены в плоскости X, Y или Z.
- Explode перемещает все части сцены в сторону от центральной точки, которую вы можете расположить в интерактивном режиме.
- Викторина при включении, когда вы выбираете деталь, а не отображаете ее название, появляется меню с множественным выбором, позволяющее вам проверить себя.
- Значок снимка вверху по центру позволяет сделать снимок сцены, который затем можно сохранить в формате jpg или нарисовать с помощью прилагаемых инструментов пера.