
антагонист — это… Что такое мышца-антагонист?
- мышца-антагонист
- мышца-антагонист
сущ., кол-во синонимов: 1
Словарь синонимов ASIS. В.Н. Тришин. 2013.
.
- мышкодром
- мышца-вращатель
Смотреть что такое «мышца-антагонист» в других словарях:
мышца-антагонист — мышца антагонист, мышцы антагониста … Орфографический словарь-справочник
мышца — икра, бицепс, сфинктер, флексор, трицепс, антагонист, мясо, эректор, миокард, экстензор, синергист, абдуктор, мускул, аддуктор, констриктор, синергет Словарь русских синонимов.
 мышца мускул Словарь синонимов русского языка. Практический… … Словарь синонимов
мышца мускул Словарь синонимов русского языка. Практический… … Словарь синонимовАНТАГОНИСТ — (греч. antagonistes, от anti против, и agonizomai борюсь). Противник, враг, гонитель чьего либо мнения. Словарь иностранных слов, вошедших в состав русского языка. Чудинов А.Н., 1910. АНТАГОНИСТ греч. antagonistes, от anti, против, и agonizomai,… … Словарь иностранных слов русского языка
антагонист — См … Словарь синонимов
АНТАГОНИСТ — АНТАГОНИСТ, в биологии мышца, которая по своему действию противоположна другой мышце. Например, пару антагонистов составляют бицепс и трицепс. В медицине антагонист это лекарственное вещество или гормон, который противоположен по действию другому … Научно-технический энциклопедический словарь
Мышца поднимающая лопатку — Мышца поднимающая лопатку Латинское название musculus lev … Википедия
Антагонист (Antagonist)
— 1. Мышца, действующая противоположно (противодействующая) другой мышце (называемой агонистом (agonist) или (prime mover)). Работа мышц агонистов и антагонистов позволяет человеку нормально двигаться. 2. Лекарственное или какое либо другое… … Медицинские терминымышца — ▲ орган (животных) ↑ способный, к (чему), сжатие мышцы, мускулы органы, состоящие из мышечной ткани, способной сокращаться под влиянием нервных импульсов; приводят в движение отдельные органы и части тела; мышечная система отсутствует у… … Идеографический словарь русского языка
АНТАГОНИСТ — (antagonist) 1. Мышца, действующая противоположно (противодействующая) другой мышце (называемой агонистом (agonist) или (prime mover)). Работа мышц агонистов и антагонистов позволяет человеку нормально двигаться. 2. Лекарственное или какое либо… … Толковый словарь по медицине
Антагонист — 1) непримиримый противник, соперник; 2) мышца, действующая в паре с основной, но создающая усилие противоположного направления, что позволяет останавливать движение и регулировать его скорость и силу; 3) противостоящие друг другу зубы верхней и… … Теоретические аспекты и основы экологической проблемы: толкователь слов и идеоматических выражений
 мышца мускул Словарь синонимов русского языка. Практический… … Словарь синонимов
мышца мускул Словарь синонимов русского языка. Практический… … Словарь синонимов Мышца, действующая противоположно (противодействующая) другой мышце (называемой агонистом (agonist) или (prime mover)). Работа мышц агонистов и антагонистов позволяет человеку нормально двигаться. 2. Лекарственное или какое либо другое… … Медицинские термины
Мышца, действующая противоположно (противодействующая) другой мышце (называемой агонистом (agonist) или (prime mover)). Работа мышц агонистов и антагонистов позволяет человеку нормально двигаться. 2. Лекарственное или какое либо другое… … Медицинские терминыДефицит калия и магния
У микроэлементов существуют свои «друзья» и свои «противники».
Главный антагонист калия – натрий. Калий для выполнения своих полезных функций накапливается в клетках человеческого тела. Как только он «сдает свои позиции», т.е. когда его запасы истощаются, — место калия тут же занимает натрий.
У этой пары прямо противоположные задачи: если калий выводит воду из организма, то натрий ее задерживает. Поэтому, устремляясь на место калия, натрий заполняет клетки водой, т.е. развивается отечность. В таком состоянии клетки и ткани организма начинают хуже выполнять свои функции. В первую очередь, страдает сердце, т.к. сердечная мышца – миокард – состоит из клеток-кардиомиоцитов, очень чувствительных к потере калия.
Самые распространенные кардиологические заболевания, которые возникают или значительно усугубляются при недостатке калия: ишемическая болезнь сердца и разные виды нарушений ритма.
Работа миокарда восстанавливается, как только в его клетки возвращается необходимое количество калия. Обычно натрий не выдерживает натиска своего соперника и уступает занятое место без лишнего сопротивления.
Обычно натрий не выдерживает натиска своего соперника и уступает занятое место без лишнего сопротивления.
А как насчет «друзей»? У калия складываются отличные отношения с магнием. Калий плохо усваивается из пищи при дефиците магния. И наоборот, в полной мере проявляет свои полезные свойства, если магний содержится в достаточном количестве. Поэтому и потреблять эти два микроэлемента нужно вместе. К счастью, они часто содержатся в одних и тех же продуктах (овощи, зелень, фрукты), да и в составе минеральных препаратов комбинацию калия и магния легко отыскать.
60% магния находится в костях человека, остальная часть — в клетках сердца и головного мозга, а также в мягких мышечных тканях. При нехватке магния на первый план выходит его физиологический антагонист – кальций. Подобно натрию, он оккупирует опустевшую территорию. Работая в избытке, кальций вызывает сильные либо неправильные мышечные сокращения. Судороги и боль в мышцах могут ощущаться во всем теле: чаще всего в икрах ног, стопах. Для таких судорог на фоне дефицита микроэлементов характерно возникновение в ночное время. При избытке калия/недостатке магния также страдает миокард: возникают нарушения ритма сердца, развивается повышенный тонус сосудов, что негативно сказывается на артериальном давлении (тенденция к его повышению), нарушаются обменные процессы в миокарде.
Для таких судорог на фоне дефицита микроэлементов характерно возникновение в ночное время. При избытке калия/недостатке магния также страдает миокард: возникают нарушения ритма сердца, развивается повышенный тонус сосудов, что негативно сказывается на артериальном давлении (тенденция к его повышению), нарушаются обменные процессы в миокарде.
Итак, при дефиците калия и/или магния сердечная мышца (миокард) питается и сокращается неправильно и, можно сказать, не «отдыхает» нормально.
- 1
повышенное потоотделение (спорт, жаркий климат, работа в горячих цехах, частое посещение бани или сауны)
- 2
диарея или применение слабительных препаратов
- 3
применение мочегонных средств (кроме калийсберегающих)
- 4
гормональное лечение, в том числе контрацепция
- 5
сахарный диабет
- 6
заболевания желудочно-кишечного тракта (гастрит, язва, заболевания желчного пузыря и поджелудочной железы)
- 7
хронические стрессы и депрессивные состояния
- 8
регулярное потребление кофе и/или алкоголя
- 9
сердечно-сосудистые заболевания
Главные причины дефицита калия и магния:
- 1
сонливость, апатия
- 2
слабость и боли в мышцах
- 3
отеки
- 4
постоянные запоры
- 5
колебания артериального давления
- 6
сбои в сердечном ритме
- 7
затруднения дыхания
- 8
сухость, выпадение и/или потускнение волос
Какие признаки указывают на пониженный уровень калия?
- 1
повышенная нервная возбудимость, раздражительность
- 2
снижение работоспособности, сильная усталость
- 3
головокружение
- 4
снижение слуха
- 5
судороги в мышцах, подергивания век
- 6
онемение в пальцах рук и/или ног
- 7
перепады артериального давления
- 8
боли в сердце
- 9
тенденция к запорам или диарее
- 10
тонкие, ломкие ногти и /или выпадение волос
Какие признаки указывают на пониженный уровень магния?
К счастью, восстановить недостаток калия и магния несложно: нужно обогатить рацион продуктами с высоким содержанием калия и магния или же можно нормализовать/повысить уровень полезных микроэлементов, принимая специальные комплексы с калием и магнием в органической форме (такие препараты будут лучше всего усваиваться организмом человека).
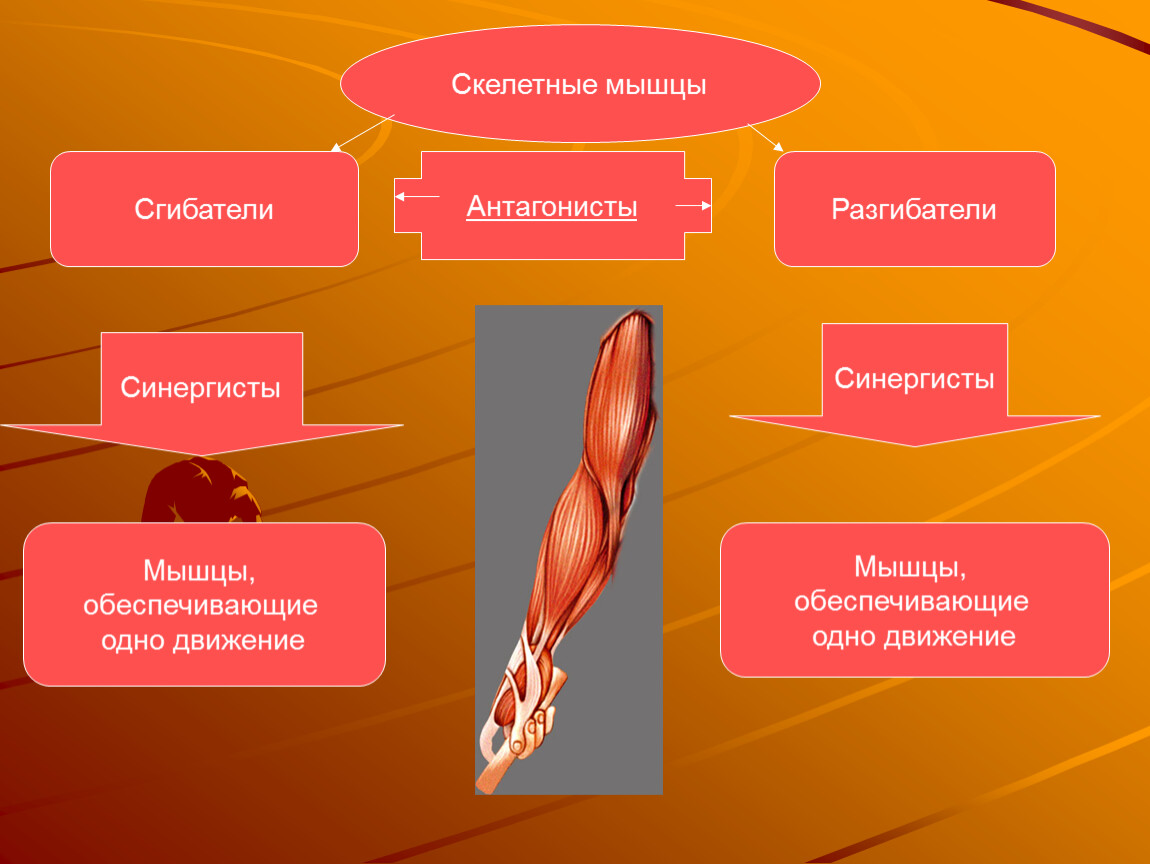
Что такое мышца-антагонист?
Мышца-антагонист — это мышца, которая работает против движения, инициированного мышцей-агонистом. Мышца-антагонист в наборе мышц возвращает конечность или другую анатомическую часть в исходное положение покоя. Эти наборы мышц называются антагонистическими парами, которые необходимы организму, потому что мышцы могут по своей природе только оказывать силу, которая воздействует на связки и кости; ему нужна противоположная мышца в паре, чтобы вернуть его в исходное положение. Мышца, ответственная за перемещение части тела из своего положения, укорачивается или сокращается, а мышца-антагонист отвечает растяжением, которое затем делает возможным второе движение.
Большинство из 600 скелетных мышц, составляющих анатомию человека, существуют в парах агонист / антагонист. Одним из примеров такого сочетания являются бицепс плечевого и трехглавого плеча. Когда бицепс плечевого пояса сокращается, трицепс плечевого пояса расслабляется, что позволяет ему растягиваться обратно в исходное положение; обратное происходит, когда трицепс плечевого пояса сокращается.
Движение групп мышц-агонистов / антагонистов координируется центральной и периферической нервной системой. Моторная кора головного мозга посылает сообщение через спинной мозг и периферическую нервную систему мышце-агонисту. Моторная кора в правом полушарии контролирует движения на левой стороне тела, а левая — на правой. Этот сигнал начинает сложное движение, которое в конечном итоге приводит к тому, что мышцы-антагонисты возвращают часть тела к месту отдыха. Мышца-антагонист помогает организму сохранять состояние расслабления.
Все мышцы содержат рецепторы или мышечные веретена, которые получают специфические сообщения от моторной коры.
ДРУГИЕ ЯЗЫКИ
Клиника Нечаева Владимира Ильича » ХОТИТЕ ТАЛИЮ «ПЕСОЧНЫЕ ЧАСЫ»?
ТАК НЕ ТЕРЯЙТЕ ВРЕМЕНИ!
Когда скульптор приступает к работе и перед ним кусок глины, которому предстоит стать Венерой или Геркулесом, прежде всего, мастер определяет поверхности – переднюю, заднюю, боковые. Затем, вращая материал, он моделирует рельеф каждой поверхности тела, добавляя и убирая объем. А поскольку бодибилдинг — это скульптура тела, давайте тоже начнем с того, что определим поверхности тела и отметим точки коррекции.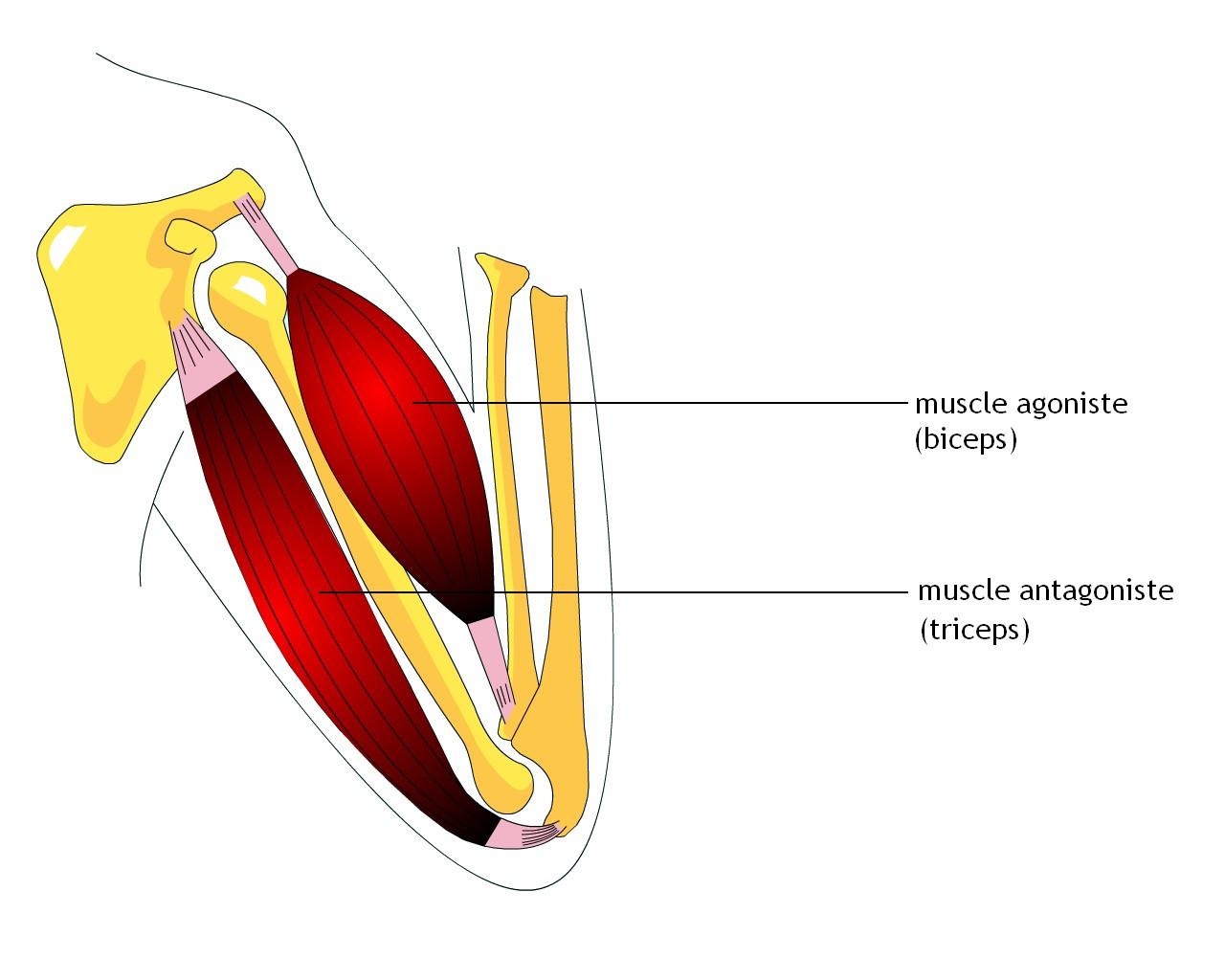
В талии мы выделяем передне-боковую и задне-боковую поверхности. Угол талии отображает выраженность талии, т.е. чем талия тоньше, тем угол острее. Поверхности условно поделим на четыре квадранта.
Рассматриваем тело с точки зрения художественной анатомии!
- На передней поверхности живота центральная линия представлена прямой мышцей живота (красный вектор), той самой, образующей желанные кубики пресса. Она является эстетической осью талии и определяет форму передней поверхности живота, создавая мышечный базис для подкожной жировой клетчатки. Проработка прямой мышцы живота также необходима для прорисовки средней линии между правой и левой половинами.
- Наружная косая мышца живота (синий вектор), определяет объем и рельеф верхней части живота в квадрантах №1 и №2.
- Внутренняя косая мышца живота (желтый вектор), формирует нижнюю часть живота в квадрантах №3 и №4.
При наличии мощных, объемных косых мышц живота угол талии сглаживается и приближается к 180, что характерно для мужского пресса. Не скажу, что это некрасиво. Эстетика тела, его построение – это процесс сугубо индивидуальный, так что не слушайте никого! Но, если вы формируете осиную талию – не перегружайте косые мышцы живота!
Не скажу, что это некрасиво. Эстетика тела, его построение – это процесс сугубо индивидуальный, так что не слушайте никого! Но, если вы формируете осиную талию – не перегружайте косые мышцы живота!
Итак, мы рассмотрели девушку спереди, она прошла мимо, самое время повернуться и посмотреть вслед!
Заднебоковая поверхность талии также представлена четырьмя квадрантами.
Эстетической осью талии при взгляде сзади является позвоночник и группа мышц разгибателей позвоночника (красный вектор).
Это мышцы антагонисты прямой мышцы живота, но с эстетической точки зрения их функции схожи. Линия позвоночника делит поясницу на две половины. Мышцы разгибатели нам интересны при взгляде сбоку, при их должном развитии они обеспечивают умеренный прогиб поясницы. Крайне важно поддержать баланс между ними и прямой мышцей живота!
Эстетическая верхних квадрантов (1 и 2) составляющая зависит от состояния широчайшей мышцы (синий вектор), именно она формирует наружный контур и верхнюю сторону угла талии. Так же она является опорой для жировой клетчатки области «крыльев». При недостаточном тонусе и объеме данной мышцы, жировая клетчатка под действием силы тяжести опускается вниз, образуя складку и создавая определенный эстетический дефект. Границу между верхними и нижними квадрантами, а именно самую узкую часть талии определяет тонус поперечной мышцы живота (зеленый вектор). И, совместно с ягодичной группой мышц (черный вектор) поперечная мышца формирует нижнюю сторону угла талии. При недостаточном тонусе мышц данной группы контур сглаживается, и талия становится невыраженной.
Так же она является опорой для жировой клетчатки области «крыльев». При недостаточном тонусе и объеме данной мышцы, жировая клетчатка под действием силы тяжести опускается вниз, образуя складку и создавая определенный эстетический дефект. Границу между верхними и нижними квадрантами, а именно самую узкую часть талии определяет тонус поперечной мышцы живота (зеленый вектор). И, совместно с ягодичной группой мышц (черный вектор) поперечная мышца формирует нижнюю сторону угла талии. При недостаточном тонусе мышц данной группы контур сглаживается, и талия становится невыраженной.
НЕСКОЛЬКО МИФОВ О ФОРМИРОВАНИИ ТАЛИИ:
Миф №1: «Хочешь узкую талию – делай скручивания!»
Да! Скручивания делать нужно во всех вариантах исполнения. Двойные концентрические, скручивания на полу, на наклонной скамье, скручивания в висе, скручивания в тренажере, обратные скручивания и многие другие варианты являются эффективными упражнениями для развития прямой мышцы живота в первую очередь.
Но узкую талию и фигуру «песочные часы» прямая мышца живота не формирует.
Прямая мышца живота является мышцей фазической, т.е. мышцей быстрого сокращения. Она не любит статические нагрузки, и уж, тем более не будет заниматься удержанием плоского живота. И выполняет прямая мышца одно простое движение – сгибание поясницы при наклоне вперед, когда вы уронили ваш iphone 8 на асфальт.
Миф №2: «Хочешь узкую талию – развивай косые мышцы живота!»
И опять – да! Боковые наклоны корпуса, скручивания с поворотом корпуса, боковые подъемы — это необходимые упражнения для укрепления наружной и внутренней косых мышц живота. Но данная мускулатура также относится к фазической и совершает быструю силовую работу. Косые мышцы производят сближение передней половины грудной клетки к соответствующей половине таза. Соответственно они необходимы для бега, ударов, бросков, кувырков, но они не выносят статической работы, а соответственно не делают талию узкой.
Миф №3: «Хочешь узкую талию – делай подъемы ног, укрепляй нижний пресс!»
Даже в 21 веке люди верят в существование «нижнего пресса», который включается только при подъемах ног. Нижним прессом принято называть нижнюю часть прямой мышцы живота, которая располагается ниже уровня пупка. Есть поверие, что данную область можно укрепить, делая такие упражнения как уголок, подъем коленей или ног в висе, подъем ног лежа и т.д. В общем, без подъема ног нижнего пресса вам не видать!
Нижним прессом принято называть нижнюю часть прямой мышцы живота, которая располагается ниже уровня пупка. Есть поверие, что данную область можно укрепить, делая такие упражнения как уголок, подъем коленей или ног в висе, подъем ног лежа и т.д. В общем, без подъема ног нижнего пресса вам не видать!
А теперь внимание вопрос! А точнее два.
1. Как прямая мышца живота поднимает ноги, если она к ним не крепится?
2. Как может преимущественно сокращаться нижняя часть мышцы, если к ней подходит один нерв и запускается она либо вся целостно, либо никак?
Путаница с нижним прессом вызвана наличием подвздошно-поясничной мышцы, соединяющей переднюю поверхность позвоночника с бедренной костью. Именно она и осуществляет подъем ног, а прямая мышца живота в данном упражнении является стабилизатором и удерживает таз от переднего наклона. Аналогично прямая мышца живота работает в подтягиваниях и других упражнениях в висе. Поэтому подъемы ног мы относим к упражнениям для общей физической подготовки, они не являются специфическими для развития брюшного пресса, а тем более узкой талии.
Конечно, это не повод списывать эти упражнения со счетов, но есть некоторые, связанные с работой позвоночника, при которых они не только не окажут положительного эффекта, но и приведут вашего клиента к боли в спине.
Визуальная оценка статики и динамики
Нет плохих упражнений – есть неправильная методология. Всю информацию, необходимую для начала тренировочного процесса может вам дать визуальная оценка. Для нашей задачи она должна состоять из двух частей биомеханической и эстетической. Правильная оценка биомеханики позволит избежать осложнений связанных с болевыми синдромами поясницы, а грамотная эстетическая оценка позволит правильно разработать программу и избежать недовольства взыскательного клиента. Подробно методология визуальной оценки будет показана на предстоящем вебинаре.
Армирование кора
Прямая и косые (наружная и внутренняя) мышцы живота – это мышцы движения, фазическая мускулатура, которая не может долго находится в сокращенном состоянии, а, следовательно, не может выполнять функцию корсета. Не будем забывать, что большую часть женщин, приходящих в фитнес-клуб занимает вопрос узкой талии, а не рельефного пресса. Следовательно, им нужен корсет, опора для прямой и косых мышц живота, тоническая мышца, способная поддерживать сокращенное состояние долгое время. И у нас есть такая, это поперечная мышца живота (зеленый вектор).
Не будем забывать, что большую часть женщин, приходящих в фитнес-клуб занимает вопрос узкой талии, а не рельефного пресса. Следовательно, им нужен корсет, опора для прямой и косых мышц живота, тоническая мышца, способная поддерживать сокращенное состояние долгое время. И у нас есть такая, это поперечная мышца живота (зеленый вектор).
Именно она работает при выполнении таких упражнений как вакуум.
Именно «вакуум» сделает талию узкой и создаст базу для формирования мышц кора. Кроме того, поперечная мышца — это корсет, который сохранит поясничный отдел позвоночника от травм, а коже передней поверхности живота – от растяжек во время беременности. Ключ к формированию осиной талии – укрепление именно поперечной мышцы живота. Вакуум — это практически изолирующее упражнение на поперечную мышцу.
Так же следует отметить «планку».
Планка это статическое упражнение, но при его выполнении необходимо подключение динамической мускулатуры, этим и объясняется небольшое время удержания планки. Можно смело ставить данную практику на второе место в рейтинге упражнений для развития идеальной талии. Не будем забывать о самом главном действии «планки» — создание баланса между группой разгибателей позвоночника и прямой мышцей живота.
Можно смело ставить данную практику на второе место в рейтинге упражнений для развития идеальной талии. Не будем забывать о самом главном действии «планки» — создание баланса между группой разгибателей позвоночника и прямой мышцей живота.
Формирование верхнего контура талии
Обратим свое внимание на задневнутреннюю поверхность. Верхнюю сторону угла талии образует широчайшая мышца спины, поэтому для красивого наружного контура необходима работа с данной мышечной группой. Спектр упражнений достаточно широк. В основе лежат тяги: подтягивание в гравитроне, тяга вертикального блока, тяга штанги в наклоне и т.д.
Отвечаю на немой вопрос. Нет! Массивной спина не будет, а сформируется четкий верхний контур и угол талии.
Нижний контур талии и стабилизация таза
Ягодичная группа мышц – уникальная конструкция, динамическая подвеска таза, формирует нижний контур талии. Но, для нас более важна другая составляющая – это стабилизация таза, так как кости таза — это места крепления мышц кора, соответственно некорректная работа ягодичных мышц приведет к потере тонуса мышц брюшного пресса. По тренировке ягодичной группы сказано много, но есть неосвещенные вопросы проблем ягодичной группы, которые мы подробно осветим на предстоящем вебинаре.
По тренировке ягодичной группы сказано много, но есть неосвещенные вопросы проблем ягодичной группы, которые мы подробно осветим на предстоящем вебинаре.
Автор: Александр Рязанцев
| Другие результаты | |
| Это небольшая плотная мышца у основания матки. | |
| Мы знаем, что сердце — всего лишь мышца, но сотни или десятки тысяч лет мы говорили. | |
| Единственная мышца в организме, которую можно разбить резким словом и вылечить нежным прикосновением. | |
| Каждая моя мышца, казалось, устала и болела. | |
| У меня болело плечо и мышца бедра, которую я слегка потянул. | |
| Тонкая мышца отделяется от реберного хряща. | |
| Поперечная фасция, поперечная мышца живота, внутренняя косая мышца и до конца паховой связки. | |
Напрягатель широкой фасции и косая внутренняя брюшная мышца удаляются по естественным линиям сращения.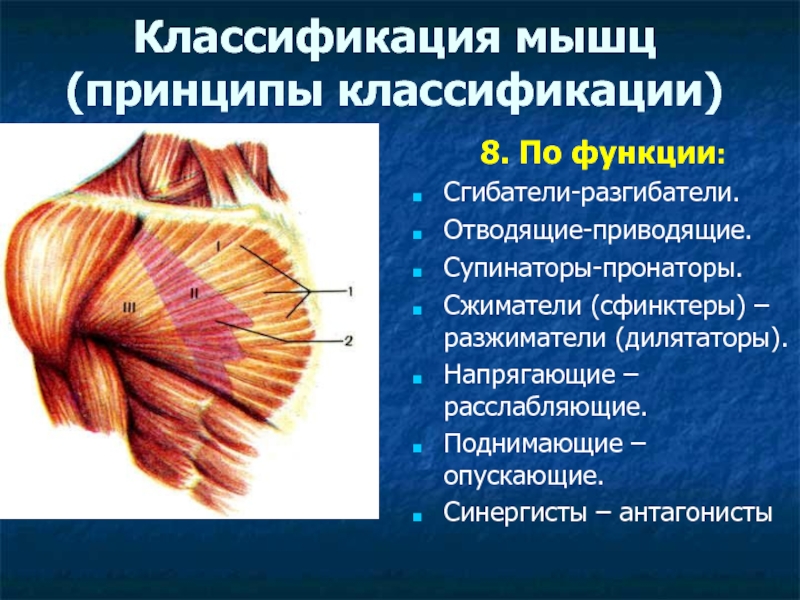 | |
| Полусухожильная мышца порционного отруба из говяжьего бедра — б/к. | |
| На заднем краю не должна быть видна трехглавая мышца плеча. | |
| Отбивные из верхней части отруба корейки — аналогично отбивным из корейки, при этом удаляются вырезка и часть хребтового края, причем основной является центральная мышца. | |
| Сердце — это мышца, а ваши мышцы ослаблены. | |
| Он наблюдал изнутри, как отказывал его организм, конечность за конечностью, мышца за мышцей. | |
| Также используется копытная мышца, которая срезается с нижней части ссека, и конусообразная мышца. | |
| Сердечная мышца полностью отделена от артерий. | |
| Наконец, если вы везунчик, вам предоставят лечение в течение 3-4 часов после приступа, но сердечная мышца уже погибла. | |
| Мышца в моем сердце перекачивает недостаточно крови в остальное тело. | |
| О ты, тремя обручами скрепленная, прочно сбитая мышца мощи! | |
Самая важная мышца -это Orbicularis oris.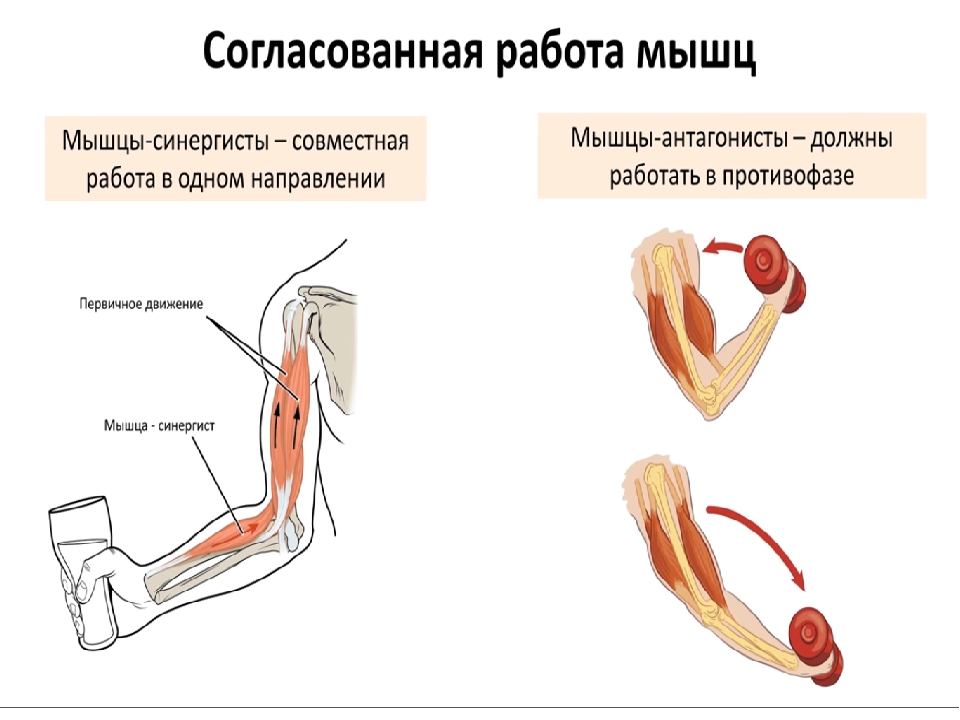 Потому что она управляет губами. Потому что она управляет губами. | |
| Я найду свою поднимающую лопатку (мышца) и разорву её попозже вечером | |
| Его левая рука, дельтовидная мышца. | |
| Зато как сильно будет болеть дельтовидная мышца! | |
| Да, когда-то здоровая мышца будет медленно увядать в одиночестве, окруженная толстым слоем жира. | |
| Да, но это намного лучше, чем оранжевая роба и сокамерник по кличке Даун или Мышца. | |
| Нет, сердечная мышца слишком слабая. | |
| Скажи мне, моя большая ягодичная мышца, каково это — так себя ощущать, а? | |
| Сердечная мышца без рубцов и некроза. | |
| Странность в том, что я не нашел следов некроза в миокарде(сердечная мышца) | |
| Должно быть это мышца или диск. | |
| Врач команды говорит, что порвана грудная мышца. | |
Кадры для фитнес-центра. Урок теории физической культуры в 8-м классе на тему «Атлетизм и его основы»
Цели и задачи: 1) углублять и расширять знания учащихся о мышцах человека;2) развивать стремление к здоровому образу жизни, вырабатывать умение сотрудничать в группе;3) воспитывать культуру здоровья и стремление к эстетической красоте тела человека.
Оборудование: проектор, экран, компьютер.Ход урока1. Организационный момент. Подготовка учащихся к уроку. Сообщение темы. Учитель: Ваша цель на данном уроке – расширять и углублять знания об особенностях человеческого организма, в данном случае о мышцах и о способах их развития с помощью самостоятельно составленного комплекса упражнений для занятий в тренажерном зале.2. Изучение нового материала.Сообщение с элементами беседы.Задачи. Повторить изученное ранее по данной теме, пополнить свои знания, усвоить особенности развития мышц. Учитель: Атлетизм – это система упражнений с отягощениями: штангой, гантелями, амортизаторами, блочными устройствами. Атлетическая гимнастика – одно из наиболее эффективных средств, всесторонне воздействующих на человеческий организм. Одна из главных задач в атлетической гимнастике – совершенствование своей фигуры, развитие мускулатуры. Какой процент массы тела составляют мышцы? Примерно 35-40 % веса человеческого тела. Атлетическая тренировка изменяет соотношение в сторону увеличения веса мускулатуры до 50% и более.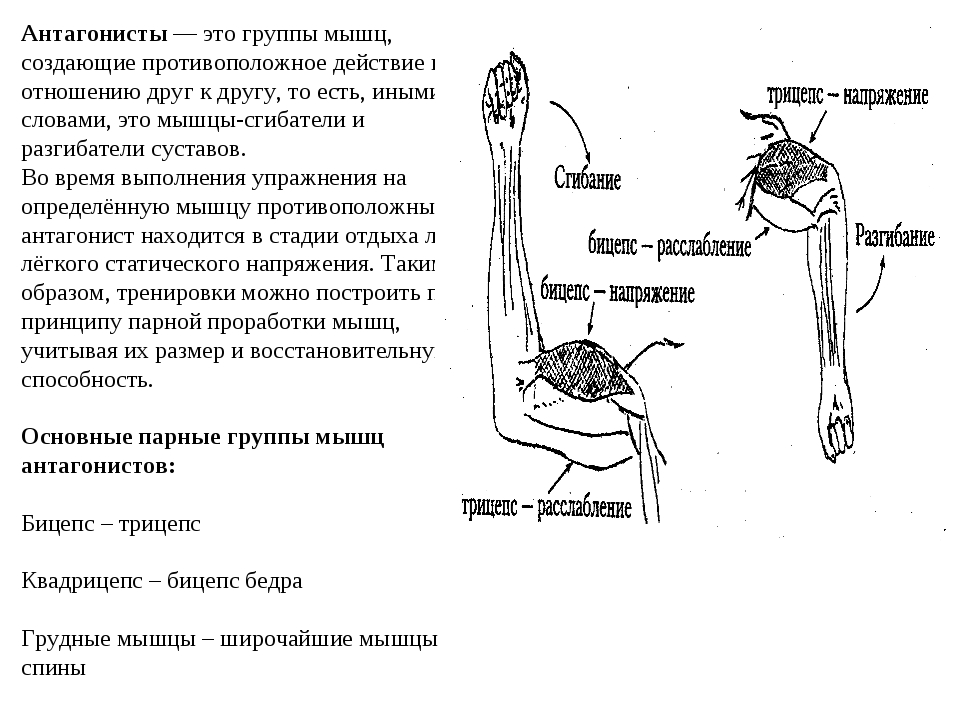 Знание функций и местоположения отдельных мышечных групп дает атлету большую свободу в подборе упражнений и тренировочных схем исходя из своих индивидуальных особенностей. Дельтовидная мышца. Она покрывает плечевой сустав. Состоит из трех пучков: переднего, среднего и заднего. Каждый пучок двигает руку в сторону, одноименную своему названию. В атлетизме увеличение ширины плеч происходит в основном за счет дельтовидных мышц, которые в хорошо развитом виде имеют шарообразные очертания. Бицепс, или двуглавая мышца плеча. Как вы думаете, где находится бицепс? Правильно! Он расположен на передней поверхности руки, выше локтя. Сгибает руку в локтевом суставе. Выйдите два парня, продемонстрируйте свои бицепсы. Как вы думаете, у кого бицепс развит сильнее?Трехглавая мышца плеча, или трицепс. Эта мышца – антагонист бицепса. Покажите, где она находится. Молодцы! Она расположена на задней поверхности плеча. Разгибает руку в локтевом суставе. В хорошо развитом виде имеет подковообразную форму. Сгибатели и разгибатели пальцев.
Знание функций и местоположения отдельных мышечных групп дает атлету большую свободу в подборе упражнений и тренировочных схем исходя из своих индивидуальных особенностей. Дельтовидная мышца. Она покрывает плечевой сустав. Состоит из трех пучков: переднего, среднего и заднего. Каждый пучок двигает руку в сторону, одноименную своему названию. В атлетизме увеличение ширины плеч происходит в основном за счет дельтовидных мышц, которые в хорошо развитом виде имеют шарообразные очертания. Бицепс, или двуглавая мышца плеча. Как вы думаете, где находится бицепс? Правильно! Он расположен на передней поверхности руки, выше локтя. Сгибает руку в локтевом суставе. Выйдите два парня, продемонстрируйте свои бицепсы. Как вы думаете, у кого бицепс развит сильнее?Трехглавая мышца плеча, или трицепс. Эта мышца – антагонист бицепса. Покажите, где она находится. Молодцы! Она расположена на задней поверхности плеча. Разгибает руку в локтевом суставе. В хорошо развитом виде имеет подковообразную форму. Сгибатели и разгибатели пальцев.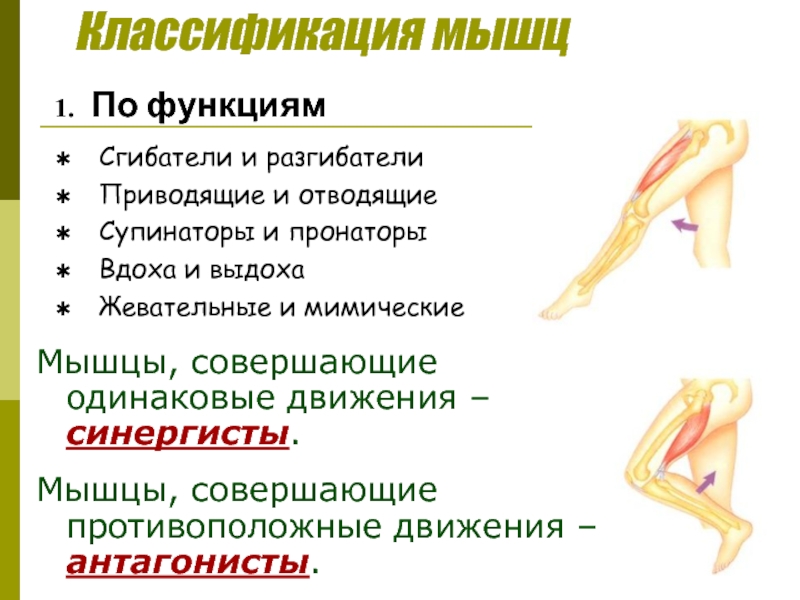 Из них состоит мышечная масса предплечья. Мышцы на внутренней стороне предплечья сгибают пальцы и кисть, а на наружной – разгибают. Большая грудная мышца. Один из важнейших компонентов атлетического развития. Как вы думаете, где она находится? Молодцы! Расположена на передней поверхности грудной клетки. Приводит руку к туловищу и вращает ее внутрь. Передняя зубчатая. Находится на боковой поверхности грудной клетки. Вращает лопатку и отводит ее от позвоночного столба. Межреберные мышцы. Находятся на ребрах и между ними. Участвуют в акте дыхания.Трапециевидная мышца. Находится на задней поверхности шеи и грудной клетки. Поднимает и опускает лопатки, приводит их к позвоночному столбу, тянет голову назад и наклоняет голову в стороны. Широчайшая мышца спины. Находится на задней поверхности грудной клетки. Приводит плечо к туловищу, вращает руку внутрь, тянет ее назад. Эта мышца придает торсу конусообразную форму. Выйдите два человека, повернитесь спиной. Ребята, у кого из них лучше развита широчайшая мышца спины?Длинные мышцы спины.
Из них состоит мышечная масса предплечья. Мышцы на внутренней стороне предплечья сгибают пальцы и кисть, а на наружной – разгибают. Большая грудная мышца. Один из важнейших компонентов атлетического развития. Как вы думаете, где она находится? Молодцы! Расположена на передней поверхности грудной клетки. Приводит руку к туловищу и вращает ее внутрь. Передняя зубчатая. Находится на боковой поверхности грудной клетки. Вращает лопатку и отводит ее от позвоночного столба. Межреберные мышцы. Находятся на ребрах и между ними. Участвуют в акте дыхания.Трапециевидная мышца. Находится на задней поверхности шеи и грудной клетки. Поднимает и опускает лопатки, приводит их к позвоночному столбу, тянет голову назад и наклоняет голову в стороны. Широчайшая мышца спины. Находится на задней поверхности грудной клетки. Приводит плечо к туловищу, вращает руку внутрь, тянет ее назад. Эта мышца придает торсу конусообразную форму. Выйдите два человека, повернитесь спиной. Ребята, у кого из них лучше развита широчайшая мышца спины?Длинные мышцы спины. Расположены вдоль позвоночного столба. Разгибают, вращают и наклоняют туловище в стороны. Даже достигнув весьма значительного развития, эти мышцы не кажутся чересчур громоздкими и массивными.Физкультурная пауза. Зарядка для глаз (см. приложение).3. Закрепление новой темы. Работа в группах.Цель: применить полученные на уроке знания на практике. Учитель: Ребята, теперь представьте себе, что вы работаете в фитнес-центре инструкторами в тренажерном зале. К вам приходят посетители с различными проблемами и просят им помочь.Задание 1-й группе: «У меня слабо развита мускулатура рук. Подберите мне упражнения для развития двуглавой и трехглавой мышц плеча».Задание 2-й группе: «У меня слабо развита дельтовидная мышца и большая грудная. Подберите мне упражнения для их развития».Задание 3-й группе: «У меня слабо развита мускулатура широчайшей мышцы спины и длинной мышцы спины. Подберите мне комплекс упражнений».Цель каждой группы – защитить свои наработки; цель экспертов – определить степень полноты и научности разработок.
Расположены вдоль позвоночного столба. Разгибают, вращают и наклоняют туловище в стороны. Даже достигнув весьма значительного развития, эти мышцы не кажутся чересчур громоздкими и массивными.Физкультурная пауза. Зарядка для глаз (см. приложение).3. Закрепление новой темы. Работа в группах.Цель: применить полученные на уроке знания на практике. Учитель: Ребята, теперь представьте себе, что вы работаете в фитнес-центре инструкторами в тренажерном зале. К вам приходят посетители с различными проблемами и просят им помочь.Задание 1-й группе: «У меня слабо развита мускулатура рук. Подберите мне упражнения для развития двуглавой и трехглавой мышц плеча».Задание 2-й группе: «У меня слабо развита дельтовидная мышца и большая грудная. Подберите мне упражнения для их развития».Задание 3-й группе: «У меня слабо развита мускулатура широчайшей мышцы спины и длинной мышцы спины. Подберите мне комплекс упражнений».Цель каждой группы – защитить свои наработки; цель экспертов – определить степень полноты и научности разработок.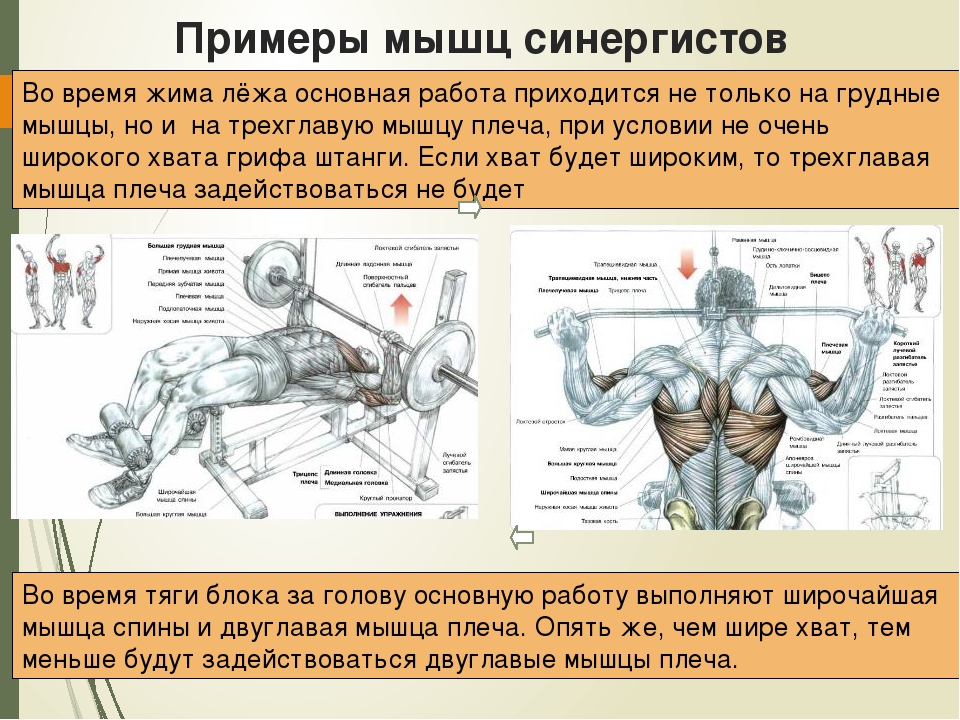 4. Экспертная оценка групповых работ.5. Подведение итогов. Домашнее задание.Учитель: Что вы узнали нового? Составьте комплекс из 20 упражнений, по два упражнения на развитие каждой мышцы.Алексей ОСИПОВ, учитель физической культуры средней школы №16 с углубленным изучением отдельных предметов имени Н.Ф.Семизорова города Тольятти Самарской области, победитель Всероссийского конкурса «Учитель года России-2011»ПриложениеЗарядка для глаз1. Круговые движения. Сделайте круговые движения открытыми глазами: сначала по часовой стрелке, затем – против.2. Движения по прямым линиям. Интенсивно подвигайте глазами по горизонтали: направо-налево, и по вертикали: вверх-вниз.3. Моргание. Интенсивно сжимайте и разжимайте глаза.4. Диагонали. Направьте взгляд в левый нижний угол, то есть посмотрите как бы на левое плечо, и сосредоточьте взгляд на этой точке. После трех морганий повторите в правую сторону.5. «Темное расслабление». Положите на закрытые глаза теплые ладони, пальцы скрещены на лбу. Расслабьтесь и постарайтесь добиться глубокого черного цвета.
4. Экспертная оценка групповых работ.5. Подведение итогов. Домашнее задание.Учитель: Что вы узнали нового? Составьте комплекс из 20 упражнений, по два упражнения на развитие каждой мышцы.Алексей ОСИПОВ, учитель физической культуры средней школы №16 с углубленным изучением отдельных предметов имени Н.Ф.Семизорова города Тольятти Самарской области, победитель Всероссийского конкурса «Учитель года России-2011»ПриложениеЗарядка для глаз1. Круговые движения. Сделайте круговые движения открытыми глазами: сначала по часовой стрелке, затем – против.2. Движения по прямым линиям. Интенсивно подвигайте глазами по горизонтали: направо-налево, и по вертикали: вверх-вниз.3. Моргание. Интенсивно сжимайте и разжимайте глаза.4. Диагонали. Направьте взгляд в левый нижний угол, то есть посмотрите как бы на левое плечо, и сосредоточьте взгляд на этой точке. После трех морганий повторите в правую сторону.5. «Темное расслабление». Положите на закрытые глаза теплые ладони, пальцы скрещены на лбу. Расслабьтесь и постарайтесь добиться глубокого черного цвета. 8. Моргания. Не менее ста раз легко и быстро поморгайте глазами.9. «Косые глаза». Сведите глаза к носу. Для выполнения этого упражнения к переносице поставьте кончик указательного пальца и посмотрите на него – тогда глаза легко «соединятся».10. «Близко-далеко, или работа глаз на расстоянии». Подойдите к окну, внимательно посмотрите на близкую деталь. Ею может быть листик дерева, растущего за окном, или небольшая бумажная точка, наклеенная на стекло на уровне глаз. Затем над выбранной точкой проведите воображаемую прямую линию, уходящую вдаль, и направьте свой взгляд далеко вперед, стараясь увидеть максимально удаленные предметы.
8. Моргания. Не менее ста раз легко и быстро поморгайте глазами.9. «Косые глаза». Сведите глаза к носу. Для выполнения этого упражнения к переносице поставьте кончик указательного пальца и посмотрите на него – тогда глаза легко «соединятся».10. «Близко-далеко, или работа глаз на расстоянии». Подойдите к окну, внимательно посмотрите на близкую деталь. Ею может быть листик дерева, растущего за окном, или небольшая бумажная точка, наклеенная на стекло на уровне глаз. Затем над выбранной точкой проведите воображаемую прямую линию, уходящую вдаль, и направьте свой взгляд далеко вперед, стараясь увидеть максимально удаленные предметы.
»Антагонист, мышечный агонист и мышечный синергизм!
Одно из самых частых сомнений людей, которые мало знают о бодибилдинге и особенно биомеханика бодибилдинга и, в основном, одна из основных причин ошибок, допущенных во время настройки тренировки, — это соотношение антагоснимо, агонизм и, следовательно, мышечная синергия.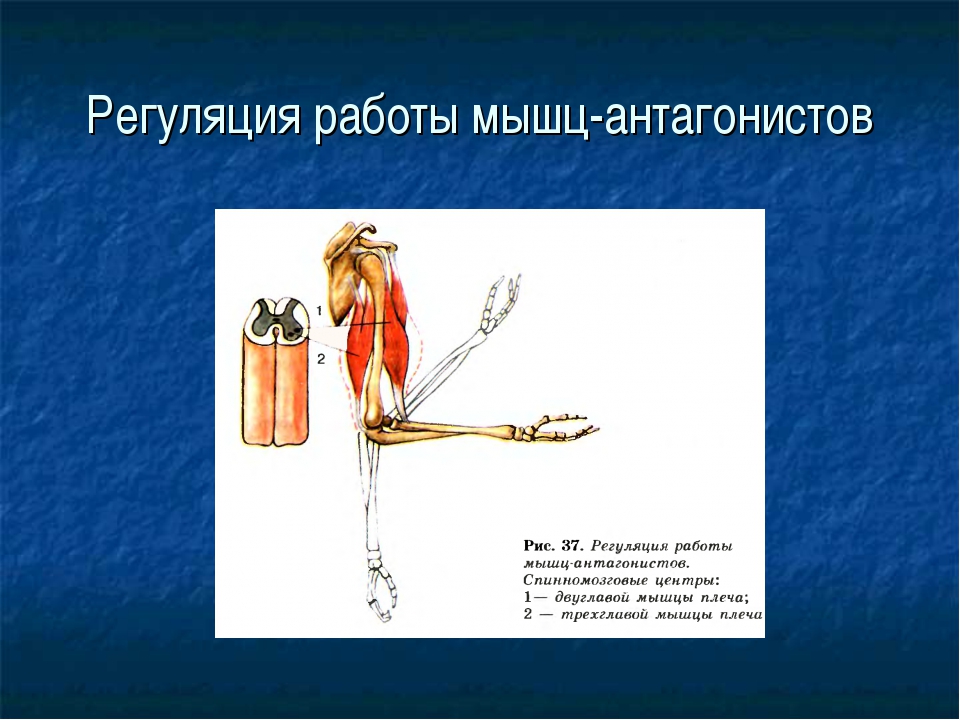
Давайте проведем небольшой тест, чтобы начать иллюстрировать: возьмите любой предмет весом не менее 1 кг и держите его в ладони. Поверните руку и оставьте ее в анатомическом положении (тыльная сторона кисти назад, ладони вперед). Затем согните руки в локтях, как будто хотите сделать гантели чередующимися нитками.
Что ты заметил?
Среди нескольких реакций мы можем наблюдать бицепс работа, мышцы пронатора, предплечья, запястья и даже часть дельтовидных мышц, обеспечивающие вращение, а почему бы не сказать сокращение? Все эти мышцы работают синергетический, один помогает и / или дает возможность двигаться другому. Итак, у нас есть то, что мы называем агонисты, то есть мышцы, которые одновременно работают на определенное движение (в данном случае на сгибание в переднебоковой оси).
Однако это движение возможно только из-за расслабления других мышц, например, трицепсов. В это время α-мотонейроны трицепса подавлены, что делает невозможным сокращение волокон и, как следствие, сокращение трицепса.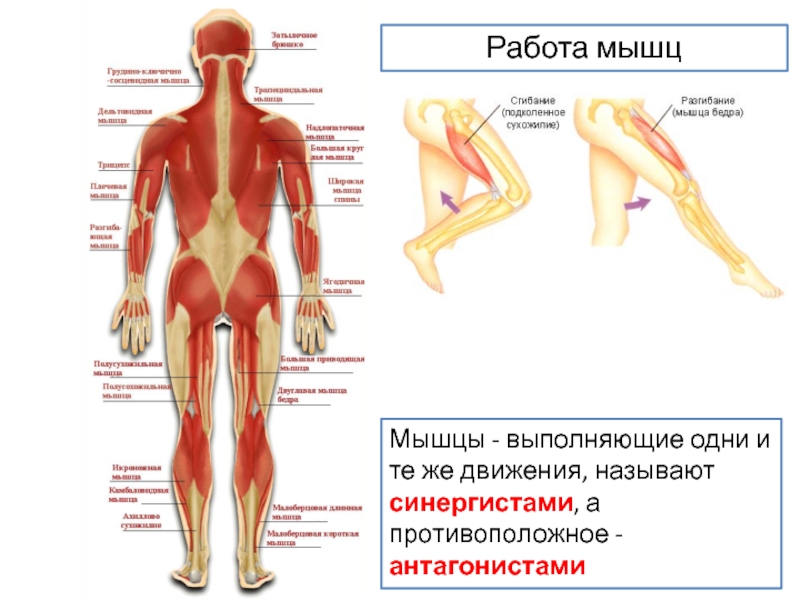 Если бы этого не произошло, мы не смогли бы согнуть руку, иначе она застряла бы, жесткая. В этом случае мы называем трицепс мышца-антагонист.
Если бы этого не произошло, мы не смогли бы согнуть руку, иначе она застряла бы, жесткая. В этом случае мы называем трицепс мышца-антагонист.
Как видим, эти и многие другие функции бывают одновременно во всех мускулах тела практически при каждом движении. Работа одной мышцы практически невозможна без подавления других мышц.
Однако чем больше мышц задействовано в движении, тем сложнее анализ. Например, анализировать жим лежа: Какие мышцы-агонисты? А антагонисты? Соответственно, можно упомянуть трицепсы, дельтовидные мышцы, большую / малую грудную мышцу, трапеции и бицепсы, некоторые мышцы спины и т. Д. Конечно все еще говорим о агонизмНельзя не упомянуть живот, который присутствует практически во всех (если не сказать всех) движениях бодибилдинга, обеспечивая стабильность и фиксацию туловища.
Понимание, что это такое агонизм и антагонизм так что намного проще разработать планы тренировок последовательно организованы (а также осознают многие ошибки, которые там делают).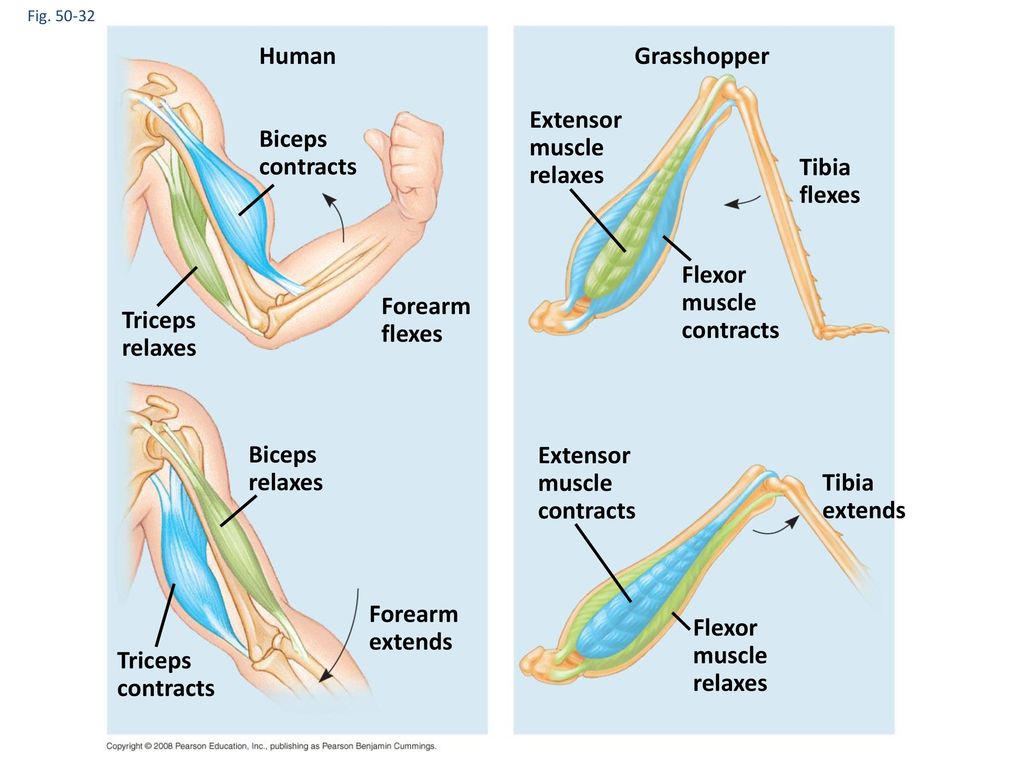 Тренировка, например, ABC 2X в неделю (кроме того, я считаю это большим преувеличением), если она плохо разделена, может вызвать такой большой износ и такое отсутствие прогресса.
Тренировка, например, ABC 2X в неделю (кроме того, я считаю это большим преувеличением), если она плохо разделена, может вызвать такой большой износ и такое отсутствие прогресса.
Нередко можно увидеть людей, которые тренируют грудь и бицепс в понедельник, а затем во вторник, тренируют спину и трицепс.
Какой результат? Конечно, в понедельник мы косвенно проработаем трицепсы на тренировке груди и будем тренировать их специально днем позже, не давая достаточного времени для отдыха. То же самое произойдет с бицепсами, которые будут проработаны и, в свою очередь, будут переучены на следующий день. Кроме того, на четвертом тренируются плечи, а на пятом тренируются плечи. Помимо отсутствия прогресса, это вызовет износ сухожилий, суставов и даже может стать причиной красивой травмы.
Важность знания биомеханика бодибилдинга это не только способствует правильной тренировке, но и дает наилучшие результаты.
Другой хороший пример — люди, которые забывают, например, что молотковая резьба также работает предплечья с лучевыми мышцами.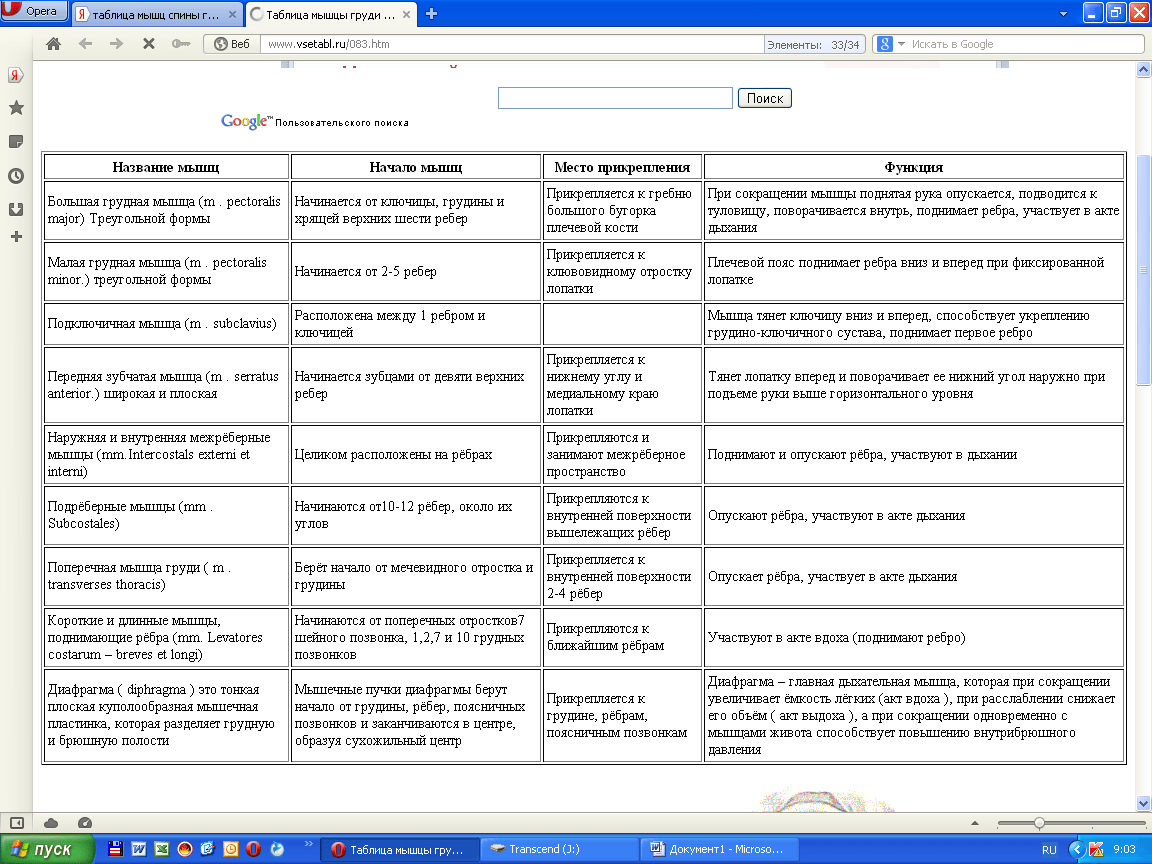 Затем человек вместо того, чтобы сосредоточиться на работе других мышц предплечья, настаивает, например, на обратной нити в конце тренировки рук. Результат: перетренированность., Чисто!
Затем человек вместо того, чтобы сосредоточиться на работе других мышц предплечья, настаивает, например, на обратной нити в конце тренировки рук. Результат: перетренированность., Чисто!
Наконец, разумно также предупредить две группы мышц, которые многие игнорируют синергию: Живот, как уже сказано, и поясничный. Это стабилизирующие мышцы, которые присутствуют практически во всех других упражнениях. Более того, они изо дня в день участвуют в наших самых простых и самых сложных действиях, таких как подъем по лестнице, спуск или даже ходьба. А в случае с поясницей восстановление все еще идет намного медленнее (около 7 дней). Значит, на тренировках они заслуживают особого внимания, чтобы не совершались эксцессы (да, в этом случае гораздо проще грешить избытком, чем недостатком).
Таким образом, всегда помните обо всех задействованных мышцах и знайте, как наилучшим образом спланировать тренировку, добиться хороших результатов и избежать травм или ненужного износа.
Хорошая тренировка!
Понравился контент? Нажмите на звездочки ниже и сообщите нам от 1 до 5, какой у вас балл за этот контент!
Средний рейтинг:
Всего голосов:
Антагонистический мышечный, агонистический и мышечный синергизм!
Мышца-антагонист — обзор
4.3.1 Введение
Комплаенс играет важную роль в передвижении людей и животных. Есть много источников податливости в биологических системах, включая мышцы, сухожилия, мягкие ткани и т. Д. Податливость суставов может даже модулироваться различными уровнями совместного сокращения мышц-агонистов и антагонистов. У комплаенс есть много преимуществ: это позволяет лучше адаптироваться к данной задаче; он позволяет накапливать энергию и высвобождать ее в другой точке цикла движения и, следовательно, может помочь в повышении эффективности передвижения (Alexander, 1988).Кроме того, податливость может улучшить стабильность системы, поскольку она создает естественные движущие силы на механическом уровне, если податливость должным образом контролируется для движения.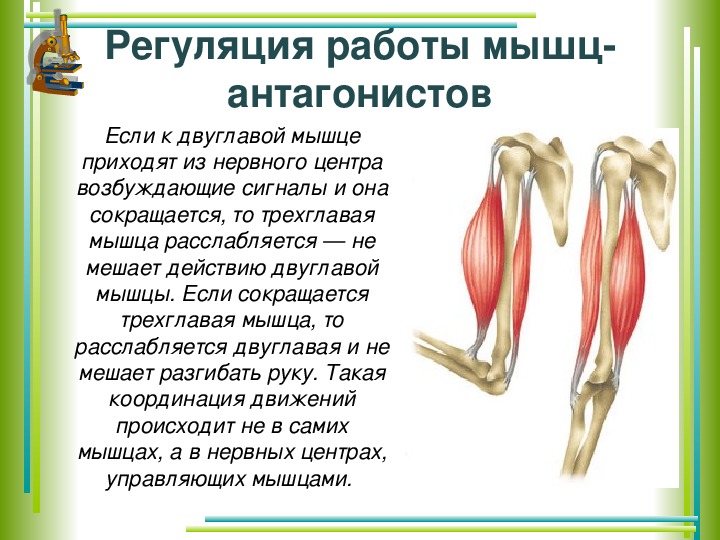 Податливые внешние поверхности приводят к более мягким контактам и большей безопасности при взаимодействии.
Податливые внешние поверхности приводят к более мягким контактам и большей безопасности при взаимодействии.
В робототехнике в последние десятилетия также произошел значительный сдвиг от жестких шарниров с регулируемым положением к управляемым по крутящему моменту и совместимым приводам и использованию пассивных податливых элементов. Цель состоит в том, чтобы также воспользоваться преимуществами, перечисленными выше, путем введения соответствия в конструкцию робота.Однако правильный выбор конструктивных параметров совместимых элементов и управляющих входов для соответствующих исполнительных механизмов — непростая задача из-за большого количества степеней свободы (DOF) систем передвижения и большого количества параметров, которые необходимо настраивать. Для решения этой проблемы в приложениях робототехники оптимальное управление представляет собой эффективный инструмент.
Цель этой главы — дать обзор преимуществ оптимального управления для правильной настройки соответствия движений робота, а также подхода к пониманию соответствия у людей. Мы резюмируем несколько исследований комплаентности в передвижении и связанных с ними динамических движений, которые были выполнены независимо разными авторами, направленных на оптимизацию переменного и постоянного соответствия. Таким образом, эта глава является дополнением к подразделам 4.4 и 4.2 по управлению импедансом и оптимальному управлению, соответственно. Основное внимание здесь уделяется податливости стыков, а не внешних контактных поверхностей. Поэтому представленные здесь исследования требуют моделей передвижения со всеми соответствующими суставами или реальной системой, но не упрощенных шаблонных моделей.
Мы резюмируем несколько исследований комплаентности в передвижении и связанных с ними динамических движений, которые были выполнены независимо разными авторами, направленных на оптимизацию переменного и постоянного соответствия. Таким образом, эта глава является дополнением к подразделам 4.4 и 4.2 по управлению импедансом и оптимальному управлению, соответственно. Основное внимание здесь уделяется податливости стыков, а не внешних контактных поверхностей. Поэтому представленные здесь исследования требуют моделей передвижения со всеми соответствующими суставами или реальной системой, но не упрощенных шаблонных моделей.
В последние годы наблюдается растущий интерес к аспектам соблюдения при передвижении человека и животных. Первоначально считалось, что она актуальна в основном для бега, но была проанализирована и при ходьбе и была продемонстрирована с помощью простых моделей пружины и массы, в которых целые ноги заменяются линейными пружинами на ровной поверхности (Geyer et al., 2006). ) и пересеченной местности (Liu et al., 2015) или рессорах переменной жесткости (Visser et al., 2013). Другие исследования были сосредоточены на роли двусуставных мышц (Iida et al., 2008; Mombaur, 2014) и сухожилий (Endo et al., 2006) с использованием моделей пружина-демпфер.
) и пересеченной местности (Liu et al., 2015) или рессорах переменной жесткости (Visser et al., 2013). Другие исследования были сосредоточены на роли двусуставных мышц (Iida et al., 2008; Mombaur, 2014) и сухожилий (Endo et al., 2006) с использованием моделей пружина-демпфер.
Комплаенс на уровне суставов также играет центральную роль в локомоции (Латаш и Зациорский, 1993), где многие исследователи обращаются к анализу жесткости суставов, изучая соотношение крутящего момента и угла в суставах ног (Weiss et al., 1986a, 1986b ), а именно бедра, колена и лодыжки. Целью многих из этих исследований является поиск возможного способа воссоздать то же движение при ходьбе с помощью простых механизмов, таких как линейные пружины, что было продемонстрировано на определенных этапах ходьбы (Shamaei et al., 2013a, 2013b, 2013c) и бегом (Günther, Blickhan, 2002). Вращающие моменты суставов вычисляются с обратной динамикой, и был проведен статистический анализ большого числа субъектов обоих полов, но без выявления различий между ними, что затем было выполнено Gabriel et al. (2008). Однако исследования показали, что комплаентность у людей варьируется и модулируется из-за совместного сокращения мышц-агонистов и мышц-антагонистов, действующих на суставы во время выполнения движений (Ferris et al., 1998; Хоган, 1984).
(2008). Однако исследования показали, что комплаентность у людей варьируется и модулируется из-за совместного сокращения мышц-агонистов и мышц-антагонистов, действующих на суставы во время выполнения движений (Ferris et al., 1998; Хоган, 1984).
Люди могут регулировать импеданс (в дополнение к своим кинематическим планам) как по направлению, так и по величине в соответствии с требованиями задачи. Это можно продемонстрировать, подвергнув человека случайным силовым полям при движении и наблюдая, как изменяется измеренное сопротивление в руке (Burdet et al., 2001; Selen et al., 2009; Franklin et al., 2007). Эти эксперименты демонстрируют, как и ожидалось из теоретических принципов управления, что адаптация импеданса может служить по крайней мере двум целям: (1) Она может использоваться для стабилизации кинематической задачи от случайных возмущений за счет увеличения жесткости .(2) С помощью понижая жесткость тела или частей, он может быть отделен от внешних возмущений. Хотя такие эксперименты из практических соображений чаще всего проводились с верхними конечностями на основе общих теоретических принципов управления, мы можем предположить, что такие принципы являются общими и применимы также к нижним конечностям. Требования задачи могут заключаться в достижении определенной цели (т. Е. Кинематических / геометрических ограничений), отсутствии падения (т. Е. Стабильности, смешанных динамических / геометрических ограничениях), скорости (переводя во время или скорости в качестве ограничений).Чаще всего в любой момент времени могут быть важны несколько таких целей, которые затрудняют поиск правильного решения задачи. Однако недавние результаты все более и более убедительно доказывают, что оптимальное управление может находить движения и связанные с ними контроллеры для решения таких сложных задач планирования и управления. Вышеупомянутые ограничения задачи обычно обрабатываются либо как мягкие ограничения через функцию стоимости, либо как «реальные» ограничения через транскрипцию.
Хотя такие эксперименты из практических соображений чаще всего проводились с верхними конечностями на основе общих теоретических принципов управления, мы можем предположить, что такие принципы являются общими и применимы также к нижним конечностям. Требования задачи могут заключаться в достижении определенной цели (т. Е. Кинематических / геометрических ограничений), отсутствии падения (т. Е. Стабильности, смешанных динамических / геометрических ограничениях), скорости (переводя во время или скорости в качестве ограничений).Чаще всего в любой момент времени могут быть важны несколько таких целей, которые затрудняют поиск правильного решения задачи. Однако недавние результаты все более и более убедительно доказывают, что оптимальное управление может находить движения и связанные с ними контроллеры для решения таких сложных задач планирования и управления. Вышеупомянутые ограничения задачи обычно обрабатываются либо как мягкие ограничения через функцию стоимости, либо как «реальные» ограничения через транскрипцию.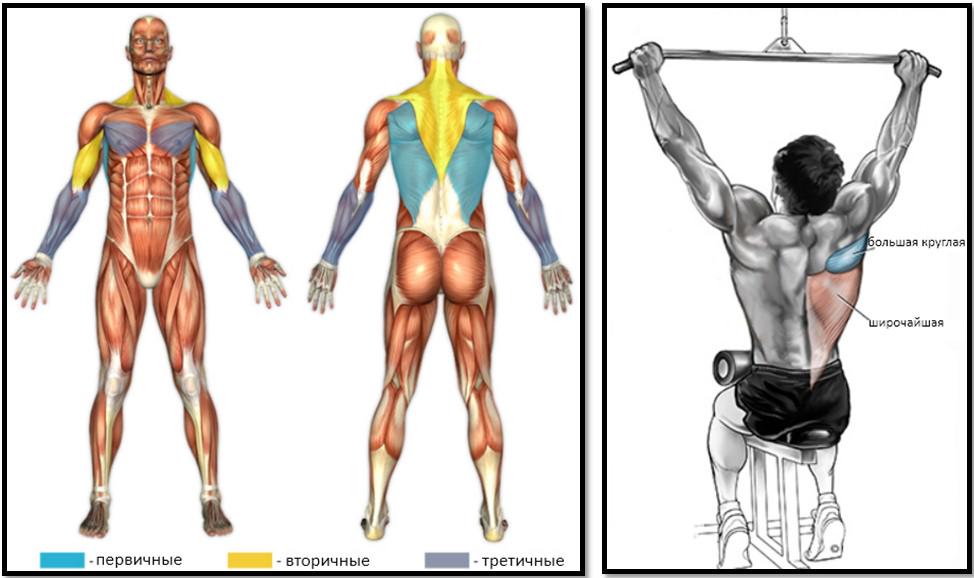
В повседневной жизни люди ходят в самых разных средах, наиболее распространенными из которых являются ровная поверхность, подъем и спуск с разными наклонами, лестницы разных размеров и разные типы пересеченной местности.Итак, чтобы лучше понять передвижение, необходимо проанализировать ходьбу во всех этих различных сценариях. Но, несмотря на большое количество литературы по жесткости на уровне суставов, большинство из них сосредоточено на ходьбе по ровной поверхности, и лишь несколько работ посвящены другим сценариям ходьбы. В биомеханике есть работы по анализу кинематики и кинетики ходьбы по склону (Franz et al., 2012; Silder et al., 2012) и лазания по лестнице (Andriacchi et al., 1980; Amirudin et al., 2014), но в этом контексте недостаточно исследований, посвященных жесткости суставов.
В робототехнике основная цель, связанная с соблюдением требований, состоит в сборе фундаментальной информации для разработки соответствующих принципов управления и приведения в действие, некоторые из которых имеют упругие элементы с фиксированной жесткостью, а другие — с переменной жесткостью.
In Mombaur et al. (2009) три разные гипотезы об использовании податливости были сформулированы для двуногого передвижения и продемонстрированы на примерах моделей многотельных систем различной сложности с использованием методов оптимизации.Было высказано предположение, что оптимально настроенная податливость может значительно снизить стоимость транспортировки, может производить естественно выглядящие движения, а также может улучшить стабильность передвижения.
Контроль соответствия и, в частности, соответствие переменных, может быть реализован в robots несколькими способами. Одним из наиболее гибких подходов является имитация соответствия внешнему контуру положения вокруг контура управления внутренним усилием / крутящим моментом (Semini et al., 2015) (см. Также подраздел 4.4. Здесь коэффициенты усиления регулятора положения соответствуют импедансу с положением усиление — это жесткость, а производное усиление — это демпфирование.Теперь задача состоит в том, чтобы найти подходящие импедансы для любого заданного времени, что сводится к нахождению графика усиления. Переменный импеданс также может быть реализован с помощью полупассивных систем. Здесь существуют две основные категории: (1) активное изменение жесткости пружины (Wolf et al., 2016) и (2) динамическая регулировка импеданса через последовательный двигатель в последовательном упругом приводе (Pratt and Williamson, 1995a). Первый подход имеет типичное ограничение, заключающееся в том, что жесткость пружины не может быть изменена мгновенно и что устройства обычно все еще слишком громоздки, чтобы их можно было хорошо интегрировать в сложные роботизированные системы, например.g., например, маленькие роботы, протезы или экзоскелеты. Второй подход имеет ограничение, заключающееся в том, что теоретические границы устойчивости строгого контроля значительно ограничивают достижимые уровни соответствия.
Переменный импеданс также может быть реализован с помощью полупассивных систем. Здесь существуют две основные категории: (1) активное изменение жесткости пружины (Wolf et al., 2016) и (2) динамическая регулировка импеданса через последовательный двигатель в последовательном упругом приводе (Pratt and Williamson, 1995a). Первый подход имеет типичное ограничение, заключающееся в том, что жесткость пружины не может быть изменена мгновенно и что устройства обычно все еще слишком громоздки, чтобы их можно было хорошо интегрировать в сложные роботизированные системы, например.g., например, маленькие роботы, протезы или экзоскелеты. Второй подход имеет ограничение, заключающееся в том, что теоретические границы устойчивости строгого контроля значительно ограничивают достижимые уровни соответствия.
Когда роботы начали выходить с заводов и входить в среду обитания людей, начали появляться гуманоидные роботы, использующие соответствующие приводы (Pratt and Williamson, 1995a), такие как робот Люси, использующий пневматические искусственные мышцы (Verrelst et al. , 2005). , Roboray с использованием приводов с сухожильным приводом (Kim et al., 2012) и M2V2 (Pratt, Krupp, 2008), COMAN (Colasanto et al., 2012), WALK-MAN (Tsagarakis et al., 2016) и iCub (Metta et al., 2010; Parmiggiani et al. , 2012) с использованием последовательных упругих приводов (SEA) (Pratt and Williamson, 1995b). В частности, робот-гуманоид COMAN показал способность стабильно ходить с SEA (Li et al., 2012; Moro et al., 2014; Dallali et al., 2012). Внедрение совместимых приводов направлено на поглощение ударов и облегчение создания движений, подобных человеку, и повышения энергоэффективности.Часто цитируемая мотивация повышения безопасности за счет добавления соответствующих элементов не является общим принципом (Semini et al., 2015).
, 2005). , Roboray с использованием приводов с сухожильным приводом (Kim et al., 2012) и M2V2 (Pratt, Krupp, 2008), COMAN (Colasanto et al., 2012), WALK-MAN (Tsagarakis et al., 2016) и iCub (Metta et al., 2010; Parmiggiani et al. , 2012) с использованием последовательных упругих приводов (SEA) (Pratt and Williamson, 1995b). В частности, робот-гуманоид COMAN показал способность стабильно ходить с SEA (Li et al., 2012; Moro et al., 2014; Dallali et al., 2012). Внедрение совместимых приводов направлено на поглощение ударов и облегчение создания движений, подобных человеку, и повышения энергоэффективности.Часто цитируемая мотивация повышения безопасности за счет добавления соответствующих элементов не является общим принципом (Semini et al., 2015).
Остальная часть этой главы организована следующим образом. В Разделе 4.3.2 мы представляем различные модели соответствия в системе передвижения, включая различные типы постоянного и переменного соответствия. В разделе 4.3.3 описаны возможности и преимущества использования оптимального контроля для исследований соответствия. В Разделе 4.3.4 мы представляем несколько примеров исследований комплаентности в передвижении человека, основанных на оптимальном управлении.Раздел 4.3.5 посвящен примерам оптимизации соответствия в движениях робота. В разделе 4.3.6, наконец, сформулированы выводы и некоторые исследовательские перспективы.
В Разделе 4.3.4 мы представляем несколько примеров исследований комплаентности в передвижении человека, основанных на оптимальном управлении.Раздел 4.3.5 посвящен примерам оптимизации соответствия в движениях робота. В разделе 4.3.6, наконец, сформулированы выводы и некоторые исследовательские перспективы.
Влияние сенсорного воздействия мышц-антагонистов на регуляцию силы
Abstract
Целью этого исследования было понять, как связанная с растяжением сенсорная обратная связь от мышцы-антагониста влияет на выработку мышц-агонистов при различных уровнях сокращения у здоровых взрослых. Десять молодых (25,3 ± 2,4 года) здоровых испытуемых выполняли постоянные изометрические сокращения сгибания колена (агонист) на 6 уровнях крутящего момента: 5%, 10%, 15%, 20%, 30% и 40% от их максимального произвольного сокращения.В половине испытаний испытуемые получали постукивание по сухожилию надколенника (сенсорная обратная связь антагониста) во время сокращения.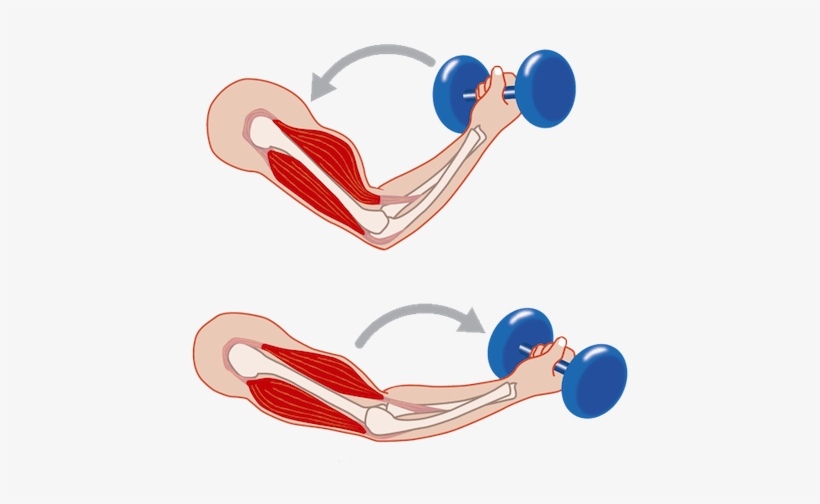 Мы сравнили погрешность в целевом крутящем моменте сгибания колена и активности мышц подколенного сухожилия с постукиванием по сухожилию надколенника и без него на 6 уровнях крутящего момента. При более низких уровнях крутящего момента (5%, 10% и 15%) испытуемые производили большую ошибку крутящего момента колена после постукивания по сухожилию по сравнению с такими же уровнями крутящего момента без постукивания по сухожилию. Напротив, мы не обнаружили никакой разницы в выходном крутящем моменте на более высоких целевых уровнях (20%, 30% и 40%) между испытаниями с нарезанием резьбы по сухожилию и без него.Мы также наблюдали зависящее от нагрузки увеличение величины активности мышц-агонистов после постукивания сухожилий, без связанного с ним увеличения ко-активации агонистов и антагонистов, зависящего от нагрузки, или подавления рефлекса от постукивания антагониста. Полученные данные свидетельствуют о том, что при относительно низкой мышечной активности наблюдается дефицит способности корректировать двигательный результат после сенсорных нарушений, и, вероятно, задействованы корковые центры (по сравнению с подкорковыми).
Мы сравнили погрешность в целевом крутящем моменте сгибания колена и активности мышц подколенного сухожилия с постукиванием по сухожилию надколенника и без него на 6 уровнях крутящего момента. При более низких уровнях крутящего момента (5%, 10% и 15%) испытуемые производили большую ошибку крутящего момента колена после постукивания по сухожилию по сравнению с такими же уровнями крутящего момента без постукивания по сухожилию. Напротив, мы не обнаружили никакой разницы в выходном крутящем моменте на более высоких целевых уровнях (20%, 30% и 40%) между испытаниями с нарезанием резьбы по сухожилию и без него.Мы также наблюдали зависящее от нагрузки увеличение величины активности мышц-агонистов после постукивания сухожилий, без связанного с ним увеличения ко-активации агонистов и антагонистов, зависящего от нагрузки, или подавления рефлекса от постукивания антагониста. Полученные данные свидетельствуют о том, что при относительно низкой мышечной активности наблюдается дефицит способности корректировать двигательный результат после сенсорных нарушений, и, вероятно, задействованы корковые центры (по сравнению с подкорковыми).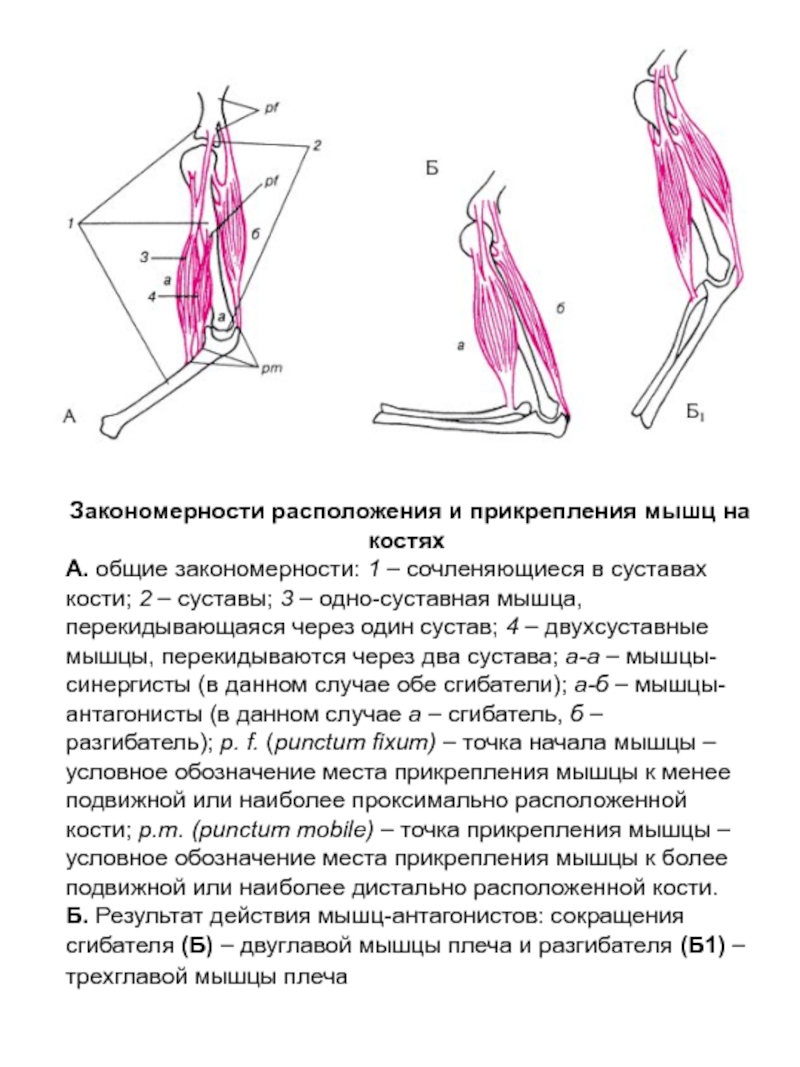
Образец цитирования: Онушко Т., Шмит Б.Д., Хингстрем А. (2015) Влияние сенсорного воздействия мышц-антагонистов на регуляцию силы.PLoS ONE 10 (7): e0133561. https://doi.org/10.1371/journal.pone.0133561
Редактор: Франсуа Хуг, Университет Квинсленда, АВСТРАЛИЯ
Поступила: 11 марта 2015 г .; Принята к печати: 27 июня 2015 г .; Опубликован: 17 июля 2015 г.
Авторские права: © 2015 Onushko et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника
Доступность данных: Все соответствующие данные находятся в пределах документ и вспомогательные информационные файлы к нему.
Финансирование: Эта публикация [или проект] была поддержана Национальным центром развития трансляционных наук Национальных институтов здравоохранения в рамках гранта номер 8KL2TR000056. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Интеграция сенсорной обратной связи с нисходящими моторными командами важна для управления выходной силой во время произвольных движений.Мышечные афферентные пути и нисходящие пути от супраспинальных центров регулируют величину и время активности мотонейронов [1,2]. В то время как общеизвестно, как афферентная обратная связь мышц влияет на регуляцию силы внутри мышцы-агониста, меньше известно о сенсорной интеграции между группами мышц-агонистов и антагонистов у людей. Понимание эффектов сенсорной интеграции от афферентных путей мышц между мышцами-агонистами и антагонистами во время субмаксимальных сокращений важно не только с точки зрения базового моторного контроля, но также потому, что это имеет клиническое значение. Изменения возбудимости этих путей были отмечены у популяций пациентов, таких как инсульт, и могут быть вредными для регуляции силы и движения [3,4].
Изменения возбудимости этих путей были отмечены у популяций пациентов, таких как инсульт, и могут быть вредными для регуляции силы и движения [3,4].
В последние годы несколько исследовательских групп количественно оценили регуляцию субмаксимальных сил ног и продемонстрировали различия между популяциями пациентов и здоровыми людьми в контрольной группе [5–8]. Регуляция субмаксимальной силы использовалась как проба для понимания стратегий волевого контроля и выявления субклинических нарушений в регуляции силы.Типичные измерения включают способность достичь заданной силы (погрешность) и способность поддерживать заданную силу (устойчивость). Хотя регулировка силы может быть специфической для конкретной задачи и мышц, более крупные группы мышц ноги, как правило, хуже регулируют силу при более низких уровнях относительной силы (<~ 20% от максимальных произвольных сокращений) по сравнению с более высокими уровнями силы [8,9]. Это проявляется в снижении устойчивости или увеличении ошибок при более низких уровнях силы и коррелирует с клиническими показателями функции [7].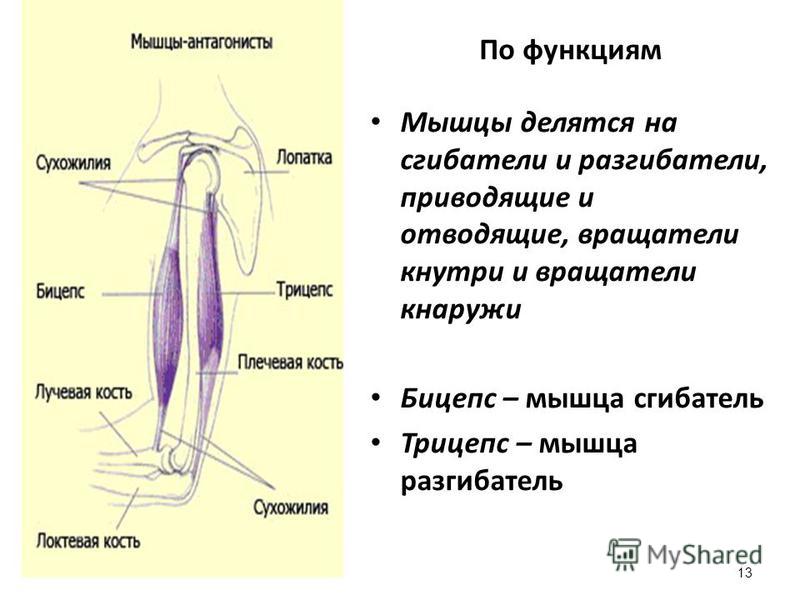 Зависимый от нагрузки эффект на регуляцию субмаксимальной силы может быть приписан соотношению мотонейрон: иннервация мышечных волокон (более низкое разрешение контроля) [8] и, в некоторых случаях, плохой временной регуляции пар агонист: антагонист [9–11].
Зависимый от нагрузки эффект на регуляцию субмаксимальной силы может быть приписан соотношению мотонейрон: иннервация мышечных волокон (более низкое разрешение контроля) [8] и, в некоторых случаях, плохой временной регуляции пар агонист: антагонист [9–11].
У людей взаимодействие сенсорной обратной связи и волевого регулирования силы в значительной степени исследовалось путем манипулирования сенсорной обратной связью внутри мышцы-агониста. Во время изометрических сокращений вибрация мышцы-агониста возбуждает проводящие пути группы Ia и, следовательно, создает силовые ошибки в двигательном выходе [12].В частности, Джонс и Хантер [12] продемонстрировали зависимый от нагрузки эффект сенсорной обратной связи агонистов во время постоянных изометрических сокращений сгибателей локтя. Субъекты производили более высокие ошибки при уровнях силы <60% от их максимального произвольного сокращения (MVC), в то время как при уровнях силы> 60% субъекты MVC генерировали меньшие ошибки в силе [12]. Точно так же было показано, что максимальное сокращение мышцы в анамнезе снижает реактивность в органах сухожилий Гольджи агониста и приводит к большим ошибкам в последующей выходной силе [13].Меньше известно о сенсорной интеграции и силовой регуляции между агонистами: антагонистами.
Точно так же было показано, что максимальное сокращение мышцы в анамнезе снижает реактивность в органах сухожилий Гольджи агониста и приводит к большим ошибкам в последующей выходной силе [13].Меньше известно о сенсорной интеграции и силовой регуляции между агонистами: антагонистами.
Есть некоторые свидетельства того, что интеграция сенсорной информации от мышечных афферентов, особенно путей группы Ia, и нисходящих входов существует в парах реципрокных мышц. Aimonetti et al. [14] продемонстрировали, что стимуляция смешанных нервов в мышце-антагонисте во время изометрического сокращения влияет на вероятность срабатывания различных типов двигательных единиц-агонистов в руке человека. Стимуляция увеличивала вероятность активации более быстрых двигательных единиц (Тип II), которые обычно задействуются при более высоких силах, одновременно снижая вероятность активации более медленных двигательных единиц (Тип I) [14].Эти результаты предлагают стратегию контроля, которая зависит от величины сокращения. В другом исследовании Кудина [15] изучала постукивание по сухожилию надколенника на активность двигательных единиц при слабом сокращении двуглавой мышцы бедра. Хотя методология не включала меры регулирования силы, авторы показали, что новые двигательные единицы задействованы в двуглавой мышце бедра. Авторы приходят к выводу, что в анамнезе рефлекторная реакция на постукивание надколенника увеличивала возбудимость мышечных двигательных единиц-агонистов при последующем сокращении.Остается важный вопрос: может ли сенсорная интеграция входных сигналов Группы Ia и волевое регулирование силы также существовать между парами мышц агонист-антагонист, и каков может быть эффект нагрузки (и, следовательно, степень активации мышц-агонистов).
В другом исследовании Кудина [15] изучала постукивание по сухожилию надколенника на активность двигательных единиц при слабом сокращении двуглавой мышцы бедра. Хотя методология не включала меры регулирования силы, авторы показали, что новые двигательные единицы задействованы в двуглавой мышце бедра. Авторы приходят к выводу, что в анамнезе рефлекторная реакция на постукивание надколенника увеличивала возбудимость мышечных двигательных единиц-агонистов при последующем сокращении.Остается важный вопрос: может ли сенсорная интеграция входных сигналов Группы Ia и волевое регулирование силы также существовать между парами мышц агонист-антагонист, и каков может быть эффект нагрузки (и, следовательно, степень активации мышц-агонистов).
Для этого исследования мы изучили, как сенсорная обратная связь от путей Ia в разгибателях коленного сустава влияет на активность сгибателей колена во время различных уровней изометрического сокращения. Наша цель состояла в том, чтобы понять, как связанная с растяжением сенсорная обратная связь от мышцы-антагониста влияет на моторную мощность агониста при различных уровнях сокращения у здоровых взрослых. Чтобы проверить это, субъекты выполняли изометрические сгибательные сокращения коленного сустава (агонист) с разными уровнями крутящего момента с постукиванием по сухожилию надколенника и без него (сенсорная обратная связь антагониста). Мы предположили, что отводы сухожилия надколенника усугубят ошибку и изменчивость при более низких уровнях нагрузки (<20% MVC), потому что погрешность и изменчивость выходной мощности двигателя больше при более низких уровнях сокращения, а сенсорная обратная связь влияет на регулирование силы.
Чтобы проверить это, субъекты выполняли изометрические сгибательные сокращения коленного сустава (агонист) с разными уровнями крутящего момента с постукиванием по сухожилию надколенника и без него (сенсорная обратная связь антагониста). Мы предположили, что отводы сухожилия надколенника усугубят ошибку и изменчивость при более низких уровнях нагрузки (<20% MVC), потому что погрешность и изменчивость выходной мощности двигателя больше при более низких уровнях сокращения, а сенсорная обратная связь влияет на регулирование силы.
Методы
Участники
Десять здоровых мужчин (средний возраст ± стандартное отклонение: 25.3 ± 2,4 года) были набраны для этого исследования. Ни один из субъектов не сообщил о какой-либо текущей или прошлой неврологической патологии. Все исследования были одобрены институциональным наблюдательным советом Университета Маркетт (IRB, HR-1196). Все клинические исследования проводились в соответствии с принципами, изложенными в Хельсинкской декларации.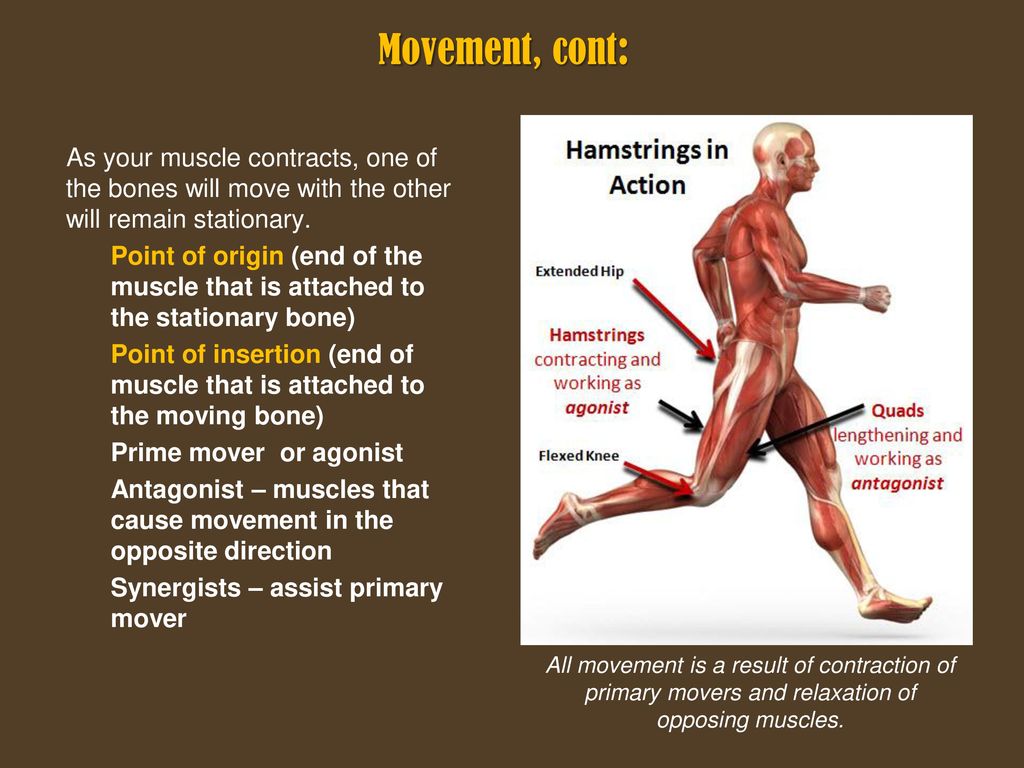 Письменное информированное согласие было получено от участников.
Письменное информированное согласие было получено от участников.
Экспериментальная установка и подход
Испытуемые лежали на спине на столе с регулируемой высотой, их ноги были закреплены в роботизированном тестовом устройстве для измерения крутящего момента в коленях (рис. 1A) [16].Роботизированный аппарат состоял из специально изготовленных опор для ног, прикрепленных к двум системам серводвигателей (Коллморген, Нортгемптон, Массачусетс). Ноги участников были закреплены в скобах для ног с помощью ремня вокруг бедра, ремня вокруг лодыжки и зажима на тыльной стороне стопы, который прикреплял ступню к подножке на конце бандажа. Правую ногу использовали в качестве тестовой для всех испытуемых, и все испытуемые сами сообщили о правостороннем доминировании.
Рис. 1. Экспериментальная установка.
(A) Роботизированное устройство, используемое для измерения крутящего момента колена во время изометрических сокращений.Субъекты получали визуальную обратную связь на мониторе компьютера в течение первых 5 секунд испытания, чтобы убедиться, что они достигли правильного уровня сокращения.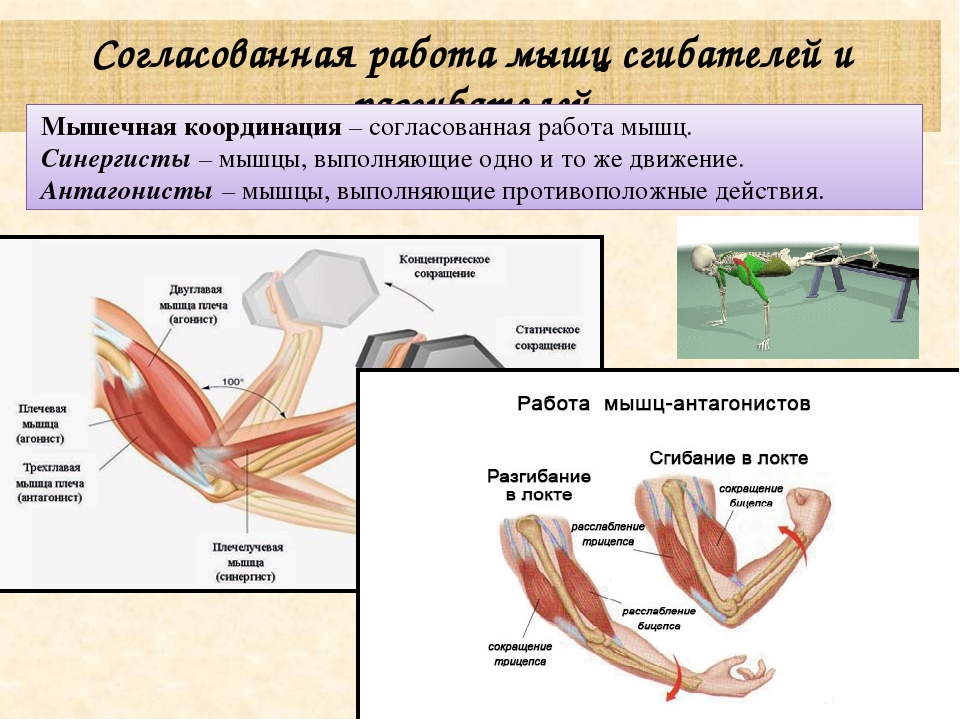 (B) Линейный двигатель обеспечивал контролируемые отводы сухожилия к сухожилию правой надколенника. Десять ударов по сухожилиям (2 Гц и 25 мс продолжительностью) были применены через 2 секунды после того, как визуальная обратная связь была удалена. (C) Экспериментальная временная шкала для изометрического сокращения сгибания колена с (i) и без (ii) постукиванием по сухожилию. Через 5 секунд визуальная обратная связь момента сгибания колена была удалена на оставшуюся часть испытания.
(B) Линейный двигатель обеспечивал контролируемые отводы сухожилия к сухожилию правой надколенника. Десять ударов по сухожилиям (2 Гц и 25 мс продолжительностью) были применены через 2 секунды после того, как визуальная обратная связь была удалена. (C) Экспериментальная временная шкала для изометрического сокращения сгибания колена с (i) и без (ii) постукиванием по сухожилию. Через 5 секунд визуальная обратная связь момента сгибания колена была удалена на оставшуюся часть испытания.
https://doi.org/10.1371/journal.pone.0133561.g001
Специально разработанный моторизованный рефлекторный молоток был прикреплен к бандажу правой ноги и использовался для устранения контролируемых нарушений сухожилия надколенника (рис. 1B). Постукивание по сухожилию активирует афферент Ia [17], а постукивание по сухожилию или рефлекс в ответ на постукивание по сухожилию является золотым стандартом в клинике для измерения реакции рефлекса растяжения при неврологическом физикальном обследовании [18,19]. Моторизованный молот состоял из линейного двигателя LinMot серии P (P01-23×160 / 70×70), питаемого от усилителя LinMot E1010 (LinMot Inc., Делаван, Висконсин). В конец вала двигателя ввертывался небольшой резиновый наконечник (диаметром 12 мм). После пальпации колена для определения местоположения сухожилия надколенника резиновый наконечник помещали на расстоянии 5 см от сухожилия и выровняли перпендикулярно (~ 90 °) по отношению к большеберцовой кости. Резиновая прокладка (2,8 х 3,1 см) была прикреплена к сухожилию надколенника пациента, чтобы распределить силу возмущения сухожилия. Линейный регулируемый дифференциальный трансформатор (LVDT; Accusens Series 2000 DC-EC, Measurement Specialists Inc., Хэмптон, Вирджиния), прикрепленный к валу линейного двигателя, использовался для измерения смещения двигателя во время испытаний с нарезанием резьбы по сухожилию.Написанные на заказ программы LabVIEW управляли скоростью линейного двигателя.
Моторизованный молот состоял из линейного двигателя LinMot серии P (P01-23×160 / 70×70), питаемого от усилителя LinMot E1010 (LinMot Inc., Делаван, Висконсин). В конец вала двигателя ввертывался небольшой резиновый наконечник (диаметром 12 мм). После пальпации колена для определения местоположения сухожилия надколенника резиновый наконечник помещали на расстоянии 5 см от сухожилия и выровняли перпендикулярно (~ 90 °) по отношению к большеберцовой кости. Резиновая прокладка (2,8 х 3,1 см) была прикреплена к сухожилию надколенника пациента, чтобы распределить силу возмущения сухожилия. Линейный регулируемый дифференциальный трансформатор (LVDT; Accusens Series 2000 DC-EC, Measurement Specialists Inc., Хэмптон, Вирджиния), прикрепленный к валу линейного двигателя, использовался для измерения смещения двигателя во время испытаний с нарезанием резьбы по сухожилию.Написанные на заказ программы LabVIEW управляли скоростью линейного двигателя.
Измерение крутящего момента.
Крутящий момент в колене измерялся датчиком крутящего момента (S. Himmelstein and Company, Hoffman Estates, Иллинойс), встроенным в опору для ног. Ось датчика крутящего момента была совмещена с приблизительным анатомическим центром вращения колена. Сигналы крутящего момента были отобраны с частотой 2 кГц с использованием карты сбора данных (National Instruments Corp., Остин, Техас) и отфильтрованы нижними частотами (аналоговый фильтр 500 Гц) до сбора данных.
Himmelstein and Company, Hoffman Estates, Иллинойс), встроенным в опору для ног. Ось датчика крутящего момента была совмещена с приблизительным анатомическим центром вращения колена. Сигналы крутящего момента были отобраны с частотой 2 кГц с использованием карты сбора данных (National Instruments Corp., Остин, Техас) и отфильтрованы нижними частотами (аналоговый фильтр 500 Гц) до сбора данных.
измерений ЭМГ.
Поверхностные электромиограммы (ЭМГ) были записаны для пяти мышц правой ноги: медиальной широкой мышцы бедра (VM), латеральной широкой мышцы бедра (VL), прямой мышцы бедра (RF), медиального подколенного сухожилия (MH) и бокового подколенного сухожилия (LH). Одноразовые предварительно гелеобразные электроды (Vermed Medical Inc., Bellows Falls, VT) помещали на мышечный живот биполярно. Перед наложением электродов кожа очищалась спиртом. Перед оцифровкой сигналы ЭМГ были предварительно усилены (x 1000) и отфильтрованы (полоса пропускания 10–1000 Гц) (Bortec Medical AMT-16; Калгари, Альберта, Канада).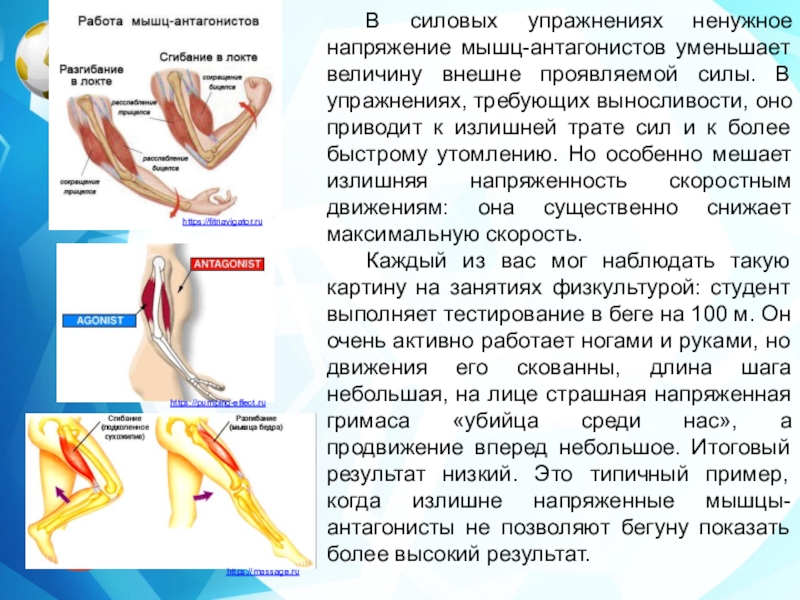 Сигналы ЭМГ регистрировались с частотой 2 кГц с использованием карты сбора данных (National Instruments Corp., Остин, Техас) и ПК.
Сигналы ЭМГ регистрировались с частотой 2 кГц с использованием карты сбора данных (National Instruments Corp., Остин, Техас) и ПК.
Максимальные произвольные сокращения.
Перед началом экспериментального тестирования испытуемые выполняли изометрические максимальные произвольные сокращения (MVC) сгибателей правого колена. Визуальная обратная связь, отображаемая на мониторе компьютера, подсказывала испытуемым, когда нужно сокращаться и расслабляться для каждого испытания MVC. Субъекты удерживали сокращение в течение 5 секунд и их устно поощряли на протяжении каждого испытания MVC.Испытания повторялись (до 5 испытаний) до тех пор, пока максимальный момент сгибания колена из 2 из 3 последних испытаний не стал меньше 5% друг от друга. MVC был определен как средний крутящий момент, поддерживаемый в течение 2 секунд испытания с максимальным крутящим моментом. Во время испытаний MVC правая нога (тестовая нога) располагалась с углом сгибания бедра 20 °, а левая нога — сгибанием бедра 0 °.
Постоянная изометрическая задача.
Испытуемые выполняли субмаксимальные изометрические сокращения сгибания колена правой ногой.Их попросили согласовать 6 постоянных целевых уровней крутящего момента (5, 10, 15, 20, 30 и 40% MVC) и поддерживать сокращение в течение 20 с. Цель отображалась на мониторе компьютера в виде горизонтальной линии, и испытуемым давали визуальную обратную связь об их крутящем моменте, чтобы убедиться, что они достигли правильного уровня. После того, как субъекты достигли и поддержали уровень сокращения в течение 2 секунд, визуальная обратная связь была предоставлена на 5 дополнительных секунд, а затем удалена на время испытания (15 с; рис. 1C). Для каждого целевого уровня участники выполнили 3 испытания с постукиванием по сухожилию надколенника (TAP) и 3 испытания без постукивания по сухожилию надколенника (NTAP) при сохранении сокращения.Во время испытаний TAP, через 2 секунды после удаления визуальной обратной связи были применены 10 ударов по сухожилиям (2 Гц и 25 мс) (рис.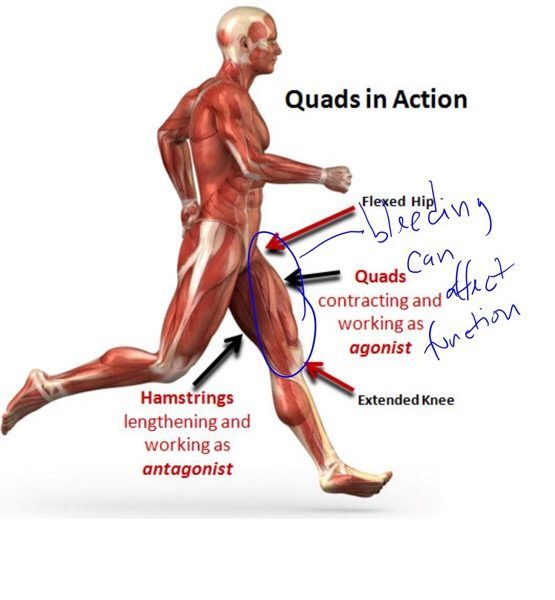 1C). Дополнительное испытание с постукиванием сухожилий было выполнено, когда субъекты находились в состоянии покоя (0% MVC). Каждый целевой уровень был представлен в случайном порядке блоков (13 испытаний на блок x 3 блока), всего 39 испытаний. Между испытаниями испытуемым давали 30–60 секунд отдыха, чтобы ограничить эффект утомления.
1C). Дополнительное испытание с постукиванием сухожилий было выполнено, когда субъекты находились в состоянии покоя (0% MVC). Каждый целевой уровень был представлен в случайном порядке блоков (13 испытаний на блок x 3 блока), всего 39 испытаний. Между испытаниями испытуемым давали 30–60 секунд отдыха, чтобы ограничить эффект утомления.
Анализ данных
Данные были проанализированы с использованием специально написанных программ Matlab (Math Works Inc., Натик, Массачусетс, США). Перед дальнейшим анализом записи крутящего момента были отфильтрованы нижними частотами (20 Гц), а записи ЭМГ — полосовыми фильтрами (60 Гц) и полосовыми фильтрами (10–300 Гц) с использованием 2 фильтров Баттерворта -го порядка ( двунаправленный). Затем сигналы крутящего момента и ЭМГ были разделены на два 5-секундных сегмента — первые 5 секунд (PRE) и последние 5 секунд (POST) постоянной части сокращения.
Измерение крутящего момента.
Для сегмента POST данных крутящего момента мы количественно определили: изменчивость, погрешность и величину.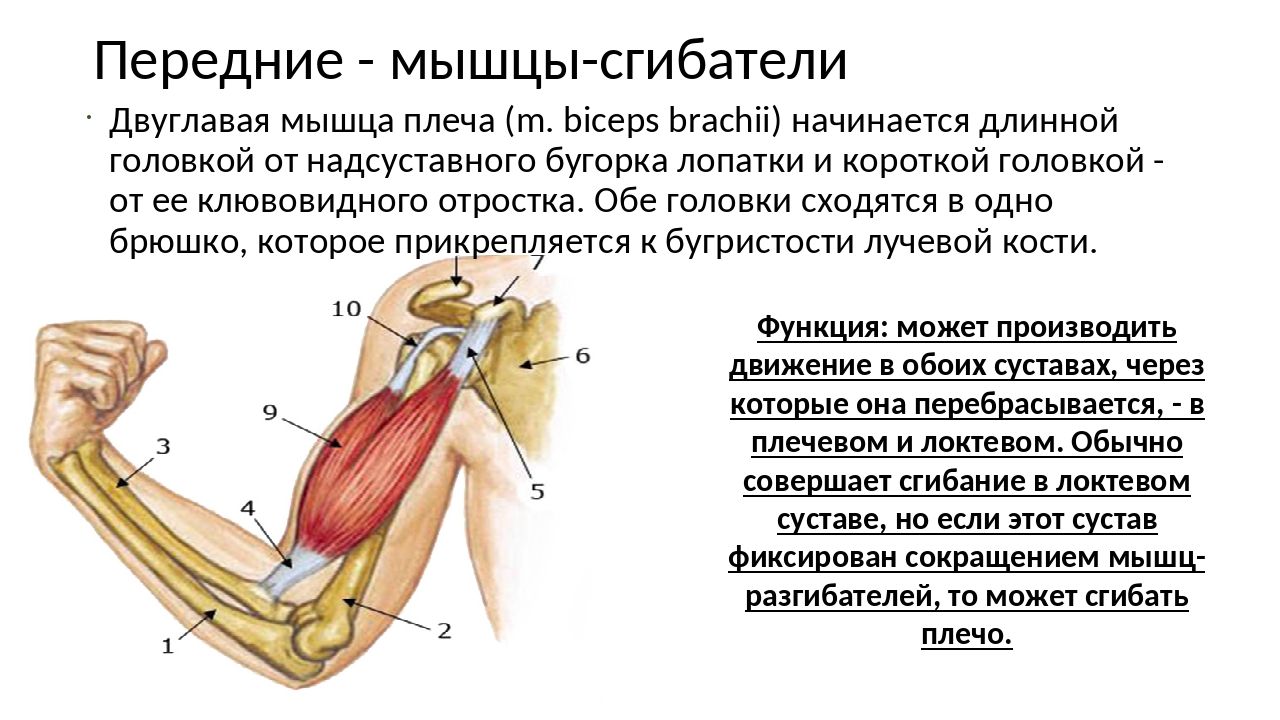 Вариабельность определялась количественно как коэффициент вариации (CV, стандартное отклонение / среднее значение) сигнала крутящего момента. Перед вычислением CV сигнал крутящего момента был сначала линейно изменен, чтобы минимизировать любой дрейф сигнала. Исключение тренда данных является принятым протоколом для определения CV силы, потому что любой дрейф, происходящий в записи крутящего момента, особенно во время отсутствия визуальной обратной связи, может повлиять на относительную величину изменчивости [8, 20–22]. Ошибки в точности отслеживания были количественно определены с использованием среднеквадратичной ошибки (RMSE) по отношению к целевому уровню крутящего момента.Величина крутящего момента была определена количественно, чтобы определить количество крутящего момента, создаваемого объектами по отношению к цели. Мы количественно оценили величину крутящего момента, вычислив площадь сигнала крутящего момента выше и ниже целевого значения, а затем взяв разницу между площадью больше и меньше целевого уровня (рис.
Вариабельность определялась количественно как коэффициент вариации (CV, стандартное отклонение / среднее значение) сигнала крутящего момента. Перед вычислением CV сигнал крутящего момента был сначала линейно изменен, чтобы минимизировать любой дрейф сигнала. Исключение тренда данных является принятым протоколом для определения CV силы, потому что любой дрейф, происходящий в записи крутящего момента, особенно во время отсутствия визуальной обратной связи, может повлиять на относительную величину изменчивости [8, 20–22]. Ошибки в точности отслеживания были количественно определены с использованием среднеквадратичной ошибки (RMSE) по отношению к целевому уровню крутящего момента.Величина крутящего момента была определена количественно, чтобы определить количество крутящего момента, создаваемого объектами по отношению к цели. Мы количественно оценили величину крутящего момента, вычислив площадь сигнала крутящего момента выше и ниже целевого значения, а затем взяв разницу между площадью больше и меньше целевого уровня (рис. 2). Для сегмента PRE данных крутящего момента мы количественно оценили RMSE по отношению к целевому уровню.
2). Для сегмента PRE данных крутящего момента мы количественно оценили RMSE по отношению к целевому уровню.
Нервная активация мышц.
Мы исследовали активацию мышц, используя три показателя ЭМГ: 1) величину ЭМГ подколенных сухожилий во время сокращения, 2) размах амплитуды ЭМГ четырехглавой мышцы в ответ на постукивание по сухожилию надколенника и 3) коэффициент коактивации для МН и ЭМГ четырехглавой мышцы.Величина активации MH EMG была основана на среднеквадратических значениях (RMS), нормированных на соответствующую MVC EMG. Затем мы рассчитали процентное изменение (Δ%) величины MH и LH EMG от POST до PRE (100 * [POST-PRE] / PRE). Затем мы определили амплитуду размаха рефлекса (P-P) от ЭМГ RF, VL и VM во время испытаний с постукиванием сухожилий. Амплитуда от пика до пика (разница в вольтах между положительным и отрицательным пиком сигнала ЭМГ) использовалась для измерения рефлекторной реакции, потому что записанный рефлекторный сигнал ЭМГ является трехфазным, а отклик от пика к пику учитывает максимум обоих. положительная и отрицательная фазы [18, 23–24].ЭМГ-ответ на прикосновение к сухожилию отличался от фонового ЭМГ на основании задержки от прикосновения (~ 20 мс) и его стандартной трехфазной формы. Амплитуда P-P-рефлекса на каждом целевом уровне была нормализована к соответствующему рефлексу P-P-ЭМГ-рефлекса во время испытания 0% MVC. Мы усреднили рефлекторные ответы P-P RF, VL и VM в пределах каждого целевого уровня. Мы сделали это, чтобы минимизировать вариабельность ЭМГ четырехглавой мышцы [25, 26]. Наконец, мы рассчитали коэффициент совместной активации четырехглавой мышцы и подколенного сухожилия.Это было вычислено как сумма средней амплитуды квадрицепса EMG (RF, VM и VL) RMS, деленная на MH EMG [27, 28]. Мы не использовали ЛГ ЭМГ в нашем анализе, потому что ЛГ не постоянно набирался между субъектами и внутри субъектов. Мы использовали MH EMG для анализа, потому что он постоянно активировался. Коэффициент совместной активации рассчитывался только во время сегмента POST для каждого целевого уровня.
положительная и отрицательная фазы [18, 23–24].ЭМГ-ответ на прикосновение к сухожилию отличался от фонового ЭМГ на основании задержки от прикосновения (~ 20 мс) и его стандартной трехфазной формы. Амплитуда P-P-рефлекса на каждом целевом уровне была нормализована к соответствующему рефлексу P-P-ЭМГ-рефлекса во время испытания 0% MVC. Мы усреднили рефлекторные ответы P-P RF, VL и VM в пределах каждого целевого уровня. Мы сделали это, чтобы минимизировать вариабельность ЭМГ четырехглавой мышцы [25, 26]. Наконец, мы рассчитали коэффициент совместной активации четырехглавой мышцы и подколенного сухожилия.Это было вычислено как сумма средней амплитуды квадрицепса EMG (RF, VM и VL) RMS, деленная на MH EMG [27, 28]. Мы не использовали ЛГ ЭМГ в нашем анализе, потому что ЛГ не постоянно набирался между субъектами и внутри субъектов. Мы использовали MH EMG для анализа, потому что он постоянно активировался. Коэффициент совместной активации рассчитывался только во время сегмента POST для каждого целевого уровня.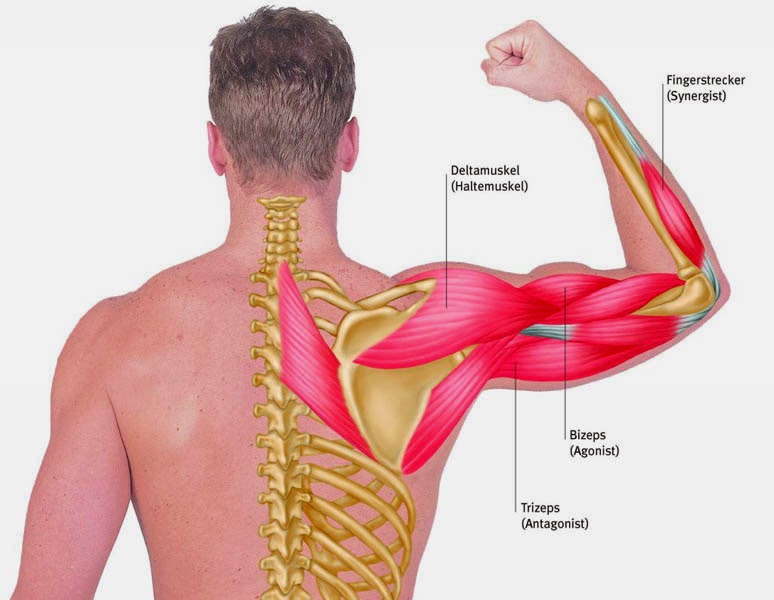
Частотный анализ.
Был проведен спектральный анализ крутящего момента, а также сегментов MH EMG PRE и POST для проверки колебаний выходной мощности двигателя.Были проведены сравнения между условиями TAP и NTAP. Спектральные данные были получены с использованием метода средней периодограммы Велча с неперекрывающимся окном Хеннинга (Matlab). Длина каждого сегмента составляла 5 секунд, а размер окна составлял 10 000 точек, в результате чего разрешение по частоте составляло 0,2 Гц. Перед спектральным анализом сегменты крутящего момента были отклонены линейно. Для статистических сравнений частота сигнала крутящего момента была разделена на полосы частот 0,1–1, 1–3, 3–7 и 7–10 Гц [29].Мы количественно определили процент пиковой мощности в каждом из диапазонов. Процент пиковой мощности рассчитывался как пиковая мощность в каждой полосе частот относительно суммы пиковых мощностей во всех полосах (0,1–10 Гц). Сегменты ЭМГ анализировали аналогично. Перед спектральным анализом сегменты ЭМГ сначала исключали тренд. Затем спектр был разделен на 3 частотных диапазона (13–30, 30–60 и 60–100 Гц), потому что эти частоты мышечных разрядов ранее были связаны со специфическими корковыми движениями [30].Мощность ЭМГ в каждой полосе нормировалась на общую пиковую мощность во всех полосах частот (13–100 Гц). Процентное изменение (Δ%) от POST к PRE (100 * ((POST-PRE) / PRE) было рассчитано для крутящего момента и данных MH.
Затем спектр был разделен на 3 частотных диапазона (13–30, 30–60 и 60–100 Гц), потому что эти частоты мышечных разрядов ранее были связаны со специфическими корковыми движениями [30].Мощность ЭМГ в каждой полосе нормировалась на общую пиковую мощность во всех полосах частот (13–100 Гц). Процентное изменение (Δ%) от POST к PRE (100 * ((POST-PRE) / PRE) было рассчитано для крутящего момента и данных MH.
Статистический анализ
Двусторонний дисперсионный анализ (6 целевых уровней x 2 состояния постукивания по сухожилию) с повторными измерениями обоих факторов был использован для сравнения вариабельности крутящего момента, погрешности и величины ЭМГ подколенных сухожилий и коэффициента совместной активации. Для сравнения относительной пиковой мощности в спектре мощности крутящего момента использовался трехфакторный дисперсионный анализ (4 полосы частот x 2 состояния отвода сухожилия x 6 целевых уровней) с повторными измерениями всех факторов.Аналогичным образом, линейный дисперсионный анализ смешанной модели (3 полосы частот x 2 состояния касания сухожилия x 6 целевых уровней x субъект (случайный фактор)) с повторными измерениями частотных диапазонов, состояния отвода сухожилия и целевых уровней был использован для сравнения нормализованной мощности ЭМГ в сигналы MH EMG. Рефлекторная ЭМГ P-P привела к ненормальному распределению. Таким образом, мы использовали ANOVA Фридмана, чтобы определить, была ли нормализованная амплитуда P-P рефлекса ЭМГ статистически различающейся между 6 целевыми уровнями.
Рефлекторная ЭМГ P-P привела к ненормальному распределению. Таким образом, мы использовали ANOVA Фридмана, чтобы определить, была ли нормализованная амплитуда P-P рефлекса ЭМГ статистически различающейся между 6 целевыми уровнями.
Статистический анализ был выполнен с использованием IBM SPSS Statistics 21.0 (IBM Corp., Армонк, Нью-Йорк, США). Соответствующие апостериорные тесты были выполнены для значимых взаимодействий из моделей ANOVA. Множественные попарные сравнения корректировали с помощью поправок Бонферрони. Альфа-уровень для всех статистических тестов составлял 0,05. Представлены только важные основные эффекты и взаимодействия, если не указано иное.
Результаты
Влияние сенсорной обратной связи антагониста Ia на мощность двигателя
Чтобы определить влияние постукивания по сухожилию надколенника на мощность двигателя сгибателя коленного сустава, мы исследовали измерения величины крутящего момента, погрешности и вариабельности в течение последних 5 секунд сокращения сгибания колена (POST).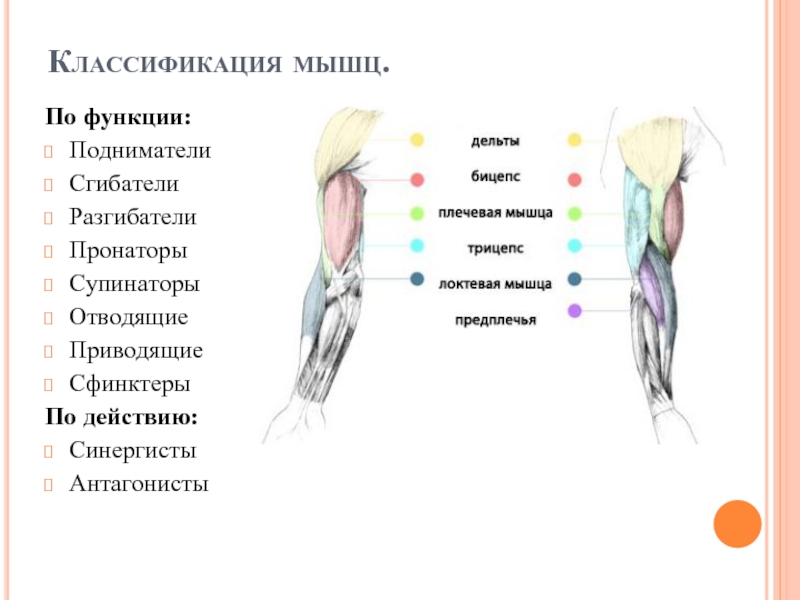 Величина крутящего момента сгибания коленного сустава была больше во время состояния TAP по сравнению с состоянием NTAP (основной эффект условия постукивания сухожилия, p = 0,001; см. Рис. 3). Мы также наблюдали взаимосвязь между постукиванием по сухожилию и целевым уровнем (% MVC) на величину крутящего момента (рис. 4). Величина крутящего момента относительно целевого значения была больше на более низких целевых уровнях (5%, 10%, 15%) после постукивания по сухожилию, в то время как не было разницы в величине крутящего момента на более высоких целевых уровнях между условиями TAP и NTAP (20%, 30%). %, 40%; величина: состояние постукивания по сухожилию × взаимодействие усилий, p = 0.045; Рис 4A). Точно так же ошибка крутящего момента была больше во время условия TAP по сравнению с NTAP (RMSE: основной эффект состояния отвода сухожилия, p = 0,002). Ошибки усугублялись после постукивания по сухожилию при более низких целевых уровнях крутящего момента (RMSE: состояние постукивания по сухожилию × взаимодействие усилия, p <0,001; рис.
Величина крутящего момента сгибания коленного сустава была больше во время состояния TAP по сравнению с состоянием NTAP (основной эффект условия постукивания сухожилия, p = 0,001; см. Рис. 3). Мы также наблюдали взаимосвязь между постукиванием по сухожилию и целевым уровнем (% MVC) на величину крутящего момента (рис. 4). Величина крутящего момента относительно целевого значения была больше на более низких целевых уровнях (5%, 10%, 15%) после постукивания по сухожилию, в то время как не было разницы в величине крутящего момента на более высоких целевых уровнях между условиями TAP и NTAP (20%, 30%). %, 40%; величина: состояние постукивания по сухожилию × взаимодействие усилий, p = 0.045; Рис 4A). Точно так же ошибка крутящего момента была больше во время условия TAP по сравнению с NTAP (RMSE: основной эффект состояния отвода сухожилия, p = 0,002). Ошибки усугублялись после постукивания по сухожилию при более низких целевых уровнях крутящего момента (RMSE: состояние постукивания по сухожилию × взаимодействие усилия, p <0,001; рис. 4B). Мы не обнаружили статистических различий в вариабельности крутящего момента между условиями TAP и NTAP (CV: среднее ± стандартное отклонение: 2,94% ± 0,35 против 2,75 ± 0,21; p = 0,291) или целевого уровня (5%: 3,14 ± 0,27%; 10%: 2,58). ± 0,25%; 15%: 2.51 ± 0,22%; 20%: 2,62 ± 0,33%; 30%: 2,99 ± 0,48%; 40%: 3,24 ± 0,37%; р = 0,169).
4B). Мы не обнаружили статистических различий в вариабельности крутящего момента между условиями TAP и NTAP (CV: среднее ± стандартное отклонение: 2,94% ± 0,35 против 2,75 ± 0,21; p = 0,291) или целевого уровня (5%: 3,14 ± 0,27%; 10%: 2,58). ± 0,25%; 15%: 2.51 ± 0,22%; 20%: 2,62 ± 0,33%; 30%: 2,99 ± 0,48%; 40%: 3,24 ± 0,37%; р = 0,169).
Рис. 3. Средний момент сгибания колена.
Испытания TAP (светлые, пунктирные линии) и испытания NTAP (темные, сплошные линии) показаны для каждого целевого уровня крутящего момента в процентах от MVC. Заштрихованная область показывает время, в течение которого испытуемым была предоставлена визуальная обратная связь.
https://doi.org/10.1371/journal.pone.0133561.g003
Чтобы определить влияние афферентной обратной связи Ia от мышц-антагонистов (четырехглавой мышцы) на амплитуду активности мышц-агонистов (подколенных сухожилий), мы сравнили среднеквадратичную амплитуду MH EMG между состояниями TAP и NTAP.Мы наблюдали тенденцию к большему процентному изменению амплитуды MH EMG во время TAP (% Δ MH: 3,57 ± 1,94%) по сравнению с условием NTAP (% Δ MH: -0,37 ± 1,81%), но эти результаты существенно не различались ( основной эффект состояния отвода сухожилия: MH, p = 0,14). Как и ожидалось, у субъектов наблюдались более значительные изменения амплитуды MH EMG с усилием (основной эффект целевого уровня:% Δ MH, p <0,001). Подобно результатам по крутящему моменту, субъекты демонстрировали большее процентное изменение амплитуды MH EMG во время более низких целевых уровней по сравнению с более высокими целевыми уровнями (Post-hoc:% Δ MH, 10%> 30%, 40%, p <0.05).
Как и ожидалось, у субъектов наблюдались более значительные изменения амплитуды MH EMG с усилием (основной эффект целевого уровня:% Δ MH, p <0,001). Подобно результатам по крутящему моменту, субъекты демонстрировали большее процентное изменение амплитуды MH EMG во время более низких целевых уровней по сравнению с более высокими целевыми уровнями (Post-hoc:% Δ MH, 10%> 30%, 40%, p <0.05).
Мы количественно оценили амплитуду рефлекса P-P, чтобы определить, различалась ли амплитуда рефлекса между целевыми уровнями. Был значительный основной эффект от целевого уровня (p = 0,003). Мы наблюдали снижение P-P рефлекторных ответов с целевого уровня 5% (1,2 ± 0,84%) до целевого уровня 40% (0,7 ± 0,19%), но апостериорные сравнения не выявили каких-либо существенных различий.
Мы также количественно определили коэффициент коактивации между RMS MH и средним RMS четырехглавой мышцы во время сокращения POST.Коэффициент совместной активации был значительно (p = 0,042) ниже для TAP (3,1 ± 0,23%) по сравнению с условием NTAP (3,4 ± 0,29%). Мы также наблюдали значительные различия в коэффициентах совместной активации между целевыми уровнями (p = 0,001). Апостериорные сравнения показали, что коэффициент коактивации для исследования 10% MVC был больше, чем соотношение из 15%, 20%, 30% и 40% (p <0,05 для всех сравнений).
Мы также наблюдали значительные различия в коэффициентах совместной активации между целевыми уровнями (p = 0,001). Апостериорные сравнения показали, что коэффициент коактивации для исследования 10% MVC был больше, чем соотношение из 15%, 20%, 30% и 40% (p <0,05 для всех сравнений).
Влияние сенсорной обратной связи антагониста Ia на низкочастотные колебания
Мы также исследовали, как обратная связь Ia посредством постукивания по сухожилию мышц-антагонистов влияет на мощность низкочастотных колебаний (<10 Гц) момента сгибания колена.В целом, изменение пиковой мощности в пределах выходного крутящего момента было на ниже (основной эффект условий отвода, p = 0,001) во время испытаний TAP (Δ% пиковая мощность, 6,19 ± 3,22%) по сравнению с испытаниями NTAP (Δ% пиковая мощность, 20,97%). ± 3,31%). Эта разница была значительно ниже (состояние отвода x усилие x взаимодействие полосы частот, p = 0,01) при 10% и 15% целевых уровнях крутящего момента MVC (рис. 5).
Рис. 5. Спектральный анализ крутящего момента.
При более низких целевых уровнях крутящего момента (10% MVC (A) и 15% MVC (B)) у субъектов наблюдались меньшие изменения пиковой мощности в пределах 3–10 Гц после возмущений постукивания сухожилий.Представленные здесь данные относятся к сегментам POST. Значения представляют собой средние значения ± SE.
https://doi.org/10.1371/journal.pone.0133561.g005
Подколенные сухожилия Изменения мощности ЭМГ
Мы заметили, что постукивание по сухожилию влияет на мощность внутри MH EMG. Мы обнаружили большее увеличение пиковой мощности во всех частотных диапазонах (13–30, 30–60 и 60–100 Гц) после постукивания по сухожилию (TAP) по сравнению с испытаниями без отводов по сухожилию (NTAP) (Δ% пиковая мощность MH — состояние отвода основной эффект, p = 0.01).
Обсуждение
Результаты этого исследования показывают, что удары по сухожилию мышцы-антагониста по-разному влияют на произвольный контроль над мышцей-агонистом на разных уровнях сокращения. Постукивание сухожилий (предположительно, группа Ia) от антагониста заставляло субъектов производить большие ошибки в выходном крутящем моменте при более низких уровнях сокращения (5%, 10% и 15% MVC). Напротив, во время более высоких уровней сокращения (20%, 30% и 40%) погрешность выходного крутящего момента существенно не различалась с отводом сухожилия надколенника и без него.Эти данные свидетельствуют о том, что при относительно низкой мышечной активности наблюдается кинестетический дефицит способности корректировать двигательную активность после сенсорных нарушений.
Постукивание сухожилий (предположительно, группа Ia) от антагониста заставляло субъектов производить большие ошибки в выходном крутящем моменте при более низких уровнях сокращения (5%, 10% и 15% MVC). Напротив, во время более высоких уровней сокращения (20%, 30% и 40%) погрешность выходного крутящего момента существенно не различалась с отводом сухожилия надколенника и без него.Эти данные свидетельствуют о том, что при относительно низкой мышечной активности наблюдается кинестетический дефицит способности корректировать двигательную активность после сенсорных нарушений.
Влияние постукивания по сухожилию антагониста на мощность двигателя агониста
Одно из объяснений увеличения ошибки крутящего момента при сгибании колена, наблюдаемой при более низких уровнях сокращения, заключается в том, что субъекты ощущали изменение длины четырехглавой мышцы после ударов по сухожилию надколенника. Удар по сухожилию надколенника вызывает рефлекс растяжения четырехглавой мышцы, вызывая сначала растяжение, а затем сокращение мышцы. Поскольку задача требовала от испытуемых поддерживать постоянный момент сгибания колена, кратковременное рефлекторно-индуцированное сокращение мышц-разгибателей колена могло быть воспринято как изменение результирующего крутящего момента. Несоответствие между сенсорной обратной связью и нисходящими моторными командами к мышце-агонисту может привести к тому, что субъекты будут воспринимать сенсорный сигнал как ошибку и чрезмерно компенсировать нарушение, чтобы удовлетворить требованиям задачи. Другие исследования продемонстрировали аналогичное поведение моторной отдачи при изменении сенсорной обратной связи.Например, во время задач согласования сил изменение сенсорной обратной связи, связанной с растяжением, посредством вибрации сухожилий вызывает завышение силы невозмущенной руки [12].
Поскольку задача требовала от испытуемых поддерживать постоянный момент сгибания колена, кратковременное рефлекторно-индуцированное сокращение мышц-разгибателей колена могло быть воспринято как изменение результирующего крутящего момента. Несоответствие между сенсорной обратной связью и нисходящими моторными командами к мышце-агонисту может привести к тому, что субъекты будут воспринимать сенсорный сигнал как ошибку и чрезмерно компенсировать нарушение, чтобы удовлетворить требованиям задачи. Другие исследования продемонстрировали аналогичное поведение моторной отдачи при изменении сенсорной обратной связи.Например, во время задач согласования сил изменение сенсорной обратной связи, связанной с растяжением, посредством вибрации сухожилий вызывает завышение силы невозмущенной руки [12].
Напротив, мы не наблюдали различий в ошибке на более высоких уровнях сокращения (20%, 30% и 40% MVC) между испытаниями с постукиванием сухожилия и без него. Поскольку не наблюдалось увеличения коактивации агониста-антагониста с увеличением уровней крутящего момента, маловероятно, что уменьшение ошибки просто связано с повышенной коактивацией антагониста.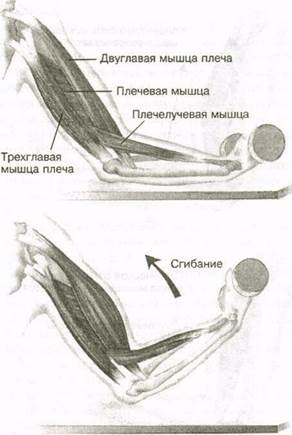 Другие исследования показали, что совместное сокращение может влиять на выходной крутящий момент [31], и в будущих исследованиях будет изучено взаимодействие совместного сжатия и входа Ia. В качестве альтернативы, возможно, что при более высоких уровнях сокращения сенсорные сигналы, полученные от постукивания по сухожилию надколенника, могут быть слишком малы по сравнению с активацией подколенных сухожилий. Это могло снизить эффективность постукивания по сухожилию надколенника для изменения активности подколенных сухожилий.
Другие исследования показали, что совместное сокращение может влиять на выходной крутящий момент [31], и в будущих исследованиях будет изучено взаимодействие совместного сжатия и входа Ia. В качестве альтернативы, возможно, что при более высоких уровнях сокращения сенсорные сигналы, полученные от постукивания по сухожилию надколенника, могут быть слишком малы по сравнению с активацией подколенных сухожилий. Это могло снизить эффективность постукивания по сухожилию надколенника для изменения активности подколенных сухожилий.
Эти данные, вероятно, предполагают, что большее количество нисходящего входа в пул мотонейронов в сочетании с восходящим входом от мышц-агонистов могло перевесить сенсорный вход от антагониста.Это уменьшит противоречивые ошибки сенсорной обратной связи, возникающие во время выполнения задания. Изменения в усилении системы Ia могут влиять на регулирование силы, при котором уменьшение ввода Ia связано с повышенной устойчивостью [32]. В текущем исследовании абсолютная величина нашего входного сигнала Ia была постоянной в каждом состоянии, а нисходящий вход увеличился (о чем свидетельствует повышенная активация MH), что, вероятно, привело к относительному снижению вклада четырехглавой мышцы Ia в выходной крутящий момент.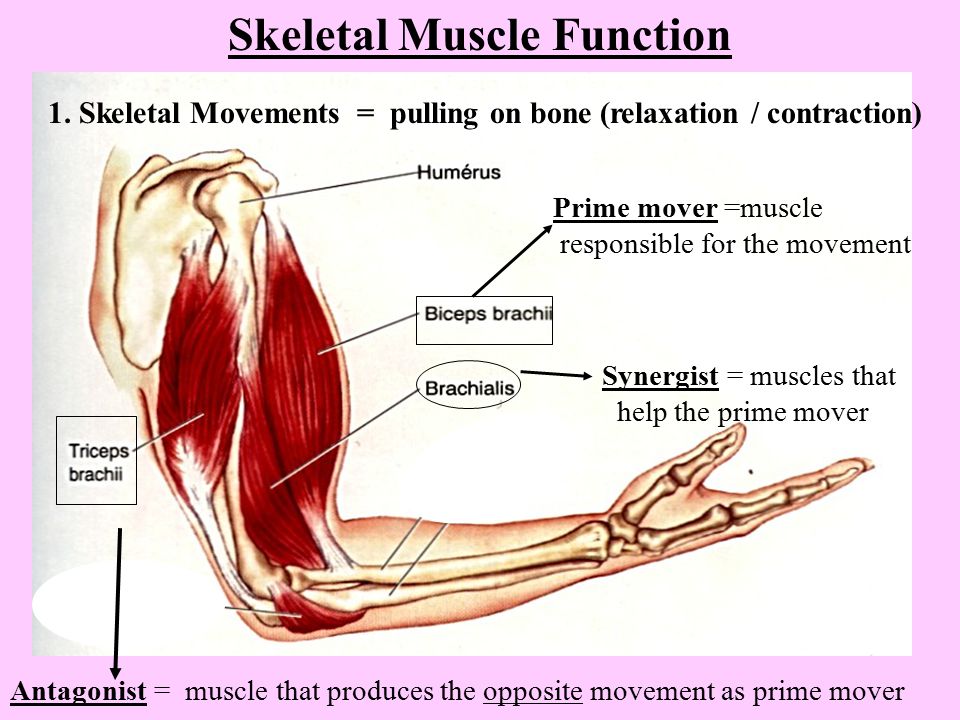 подколенных сухожилий.Хотя возможно, что при более высоких нагрузках наблюдалось снижение гамма-драйва [33] или усиление пресинаптического ингибирования Ia [34], что снизило бы относительную эффективность ввода антагониста, или меньшее сенсорное воздействие на мышцу-антагонист при более высокой нагрузке. уровни сокращений по сравнению с нижними уровнями сокращений.
подколенных сухожилий.Хотя возможно, что при более высоких нагрузках наблюдалось снижение гамма-драйва [33] или усиление пресинаптического ингибирования Ia [34], что снизило бы относительную эффективность ввода антагониста, или меньшее сенсорное воздействие на мышцу-антагонист при более высокой нагрузке. уровни сокращений по сравнению с нижними уровнями сокращений.
Возможные механизмы
Повышенная погрешность при более низких уровнях сокращения может быть связана с корковой стратегией компенсации сенсорных нарушений.Хотя у нас нет прямых доказательств кортикально связанных изменений моторного выхода после постукивания надколенника, было высказано предположение, что изменения в колебаниях моторного выхода связаны с изменениями кортикального возбуждения мотонейронов [35–37]. В текущем исследовании мы наблюдали различия в спектре мощности выходного крутящего момента в пределах 3–10 Гц между условиями TAP и NTAP на низких целевых уровнях (спектр мощности есть только для сегментов POST сокращения; Рис.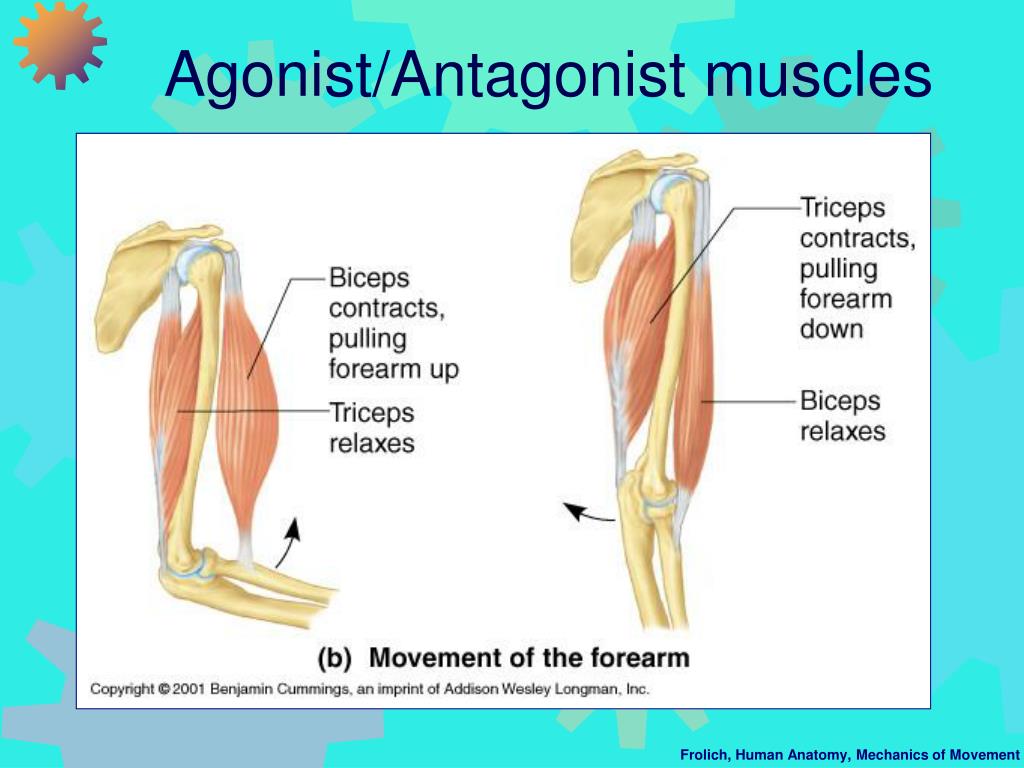 5). Во время состояния NTAP изменение от визуальной обратной связи к отсутствию визуальной обратной связи могло привести к тому, что субъекты стали меньше полагаться на корковые стратегии для поддержания правильного результата сгибания колена.Напротив, во время состояния TAP не было практически никакого изменения мощности в диапазоне 3–10 Гц, что может указывать на большее поражение коры. Tracy et al. [8] наблюдали аналогичные колебательные изменения во время зрительно-моторной изометрической задачи. Во время выполнения задания устранение визуальной обратной связи увеличивало мощность разгибателя колена в диапазоне частот 8–12 Гц, что предполагает изменение стратегии управления, включающее усиление непроизвольных процессов [8]. Точно так же другие приписывают общий драйв различиям в двигательной активности между антагонистами-агонистами мышечной активности [38,39].Тем не менее, мы не увидели какого-либо большого вклада от постукивания сухожилий мышцы-антагониста, и это может зависеть от положения сустава, которое не позволяли наши методы [38].
5). Во время состояния NTAP изменение от визуальной обратной связи к отсутствию визуальной обратной связи могло привести к тому, что субъекты стали меньше полагаться на корковые стратегии для поддержания правильного результата сгибания колена.Напротив, во время состояния TAP не было практически никакого изменения мощности в диапазоне 3–10 Гц, что может указывать на большее поражение коры. Tracy et al. [8] наблюдали аналогичные колебательные изменения во время зрительно-моторной изометрической задачи. Во время выполнения задания устранение визуальной обратной связи увеличивало мощность разгибателя колена в диапазоне частот 8–12 Гц, что предполагает изменение стратегии управления, включающее усиление непроизвольных процессов [8]. Точно так же другие приписывают общий драйв различиям в двигательной активности между антагонистами-агонистами мышечной активности [38,39].Тем не менее, мы не увидели какого-либо большого вклада от постукивания сухожилий мышцы-антагониста, и это может зависеть от положения сустава, которое не позволяли наши методы [38].
Хотя спинальные механизмы могут играть роль в ответных реакциях, мы не думаем, что наши открытия сильно связаны со спинальными механизмами. Если наблюдаемое увеличение ошибки было связано с простым суммированием входных сигналов на уровне спинного мозга, мы могли бы ожидать, что испытуемые будут недооценивать цель (производят меньшее сгибание колена), потому что рефлекс растяжения четырехглавой мышцы теоретически подавлял бы пул МН. подколенные сухожилия.
Функциональные последствия
В контрольной группе здоровых людей мы показали, что ошибка крутящего момента больше при более низких по сравнению с более высокими нагрузками в ответ на стимуляцию путей антагониста Ia. Чрезмерная компенсация момента сгибания колена и ошибка в конечном итоге могут быть стратегией стабилизации, чтобы предотвратить потерю равновесия или травму. В текущем исследовании двигательная цель состоит в том, чтобы произвести определенную степень момента сгибания колена, а рефлекторный ответ квадрицепса контрпродуктивен для цели двигательной команды. Кинестетический дефицит и ошибка при более низких нагрузках могут быть объяснены посредством визуальной информации у здоровых людей. Однако дефицит при низких нагрузках может способствовать несбалансированной активности подколенных сухожилий и четырехглавой мышцы, что, как предполагается, способствует разрывам связок колена [40].
Кинестетический дефицит и ошибка при более низких нагрузках могут быть объяснены посредством визуальной информации у здоровых людей. Однако дефицит при низких нагрузках может способствовать несбалансированной активности подколенных сухожилий и четырехглавой мышцы, что, как предполагается, способствует разрывам связок колена [40].
Что касается популяций пациентов, пациенты с инсультом имеют повышенную возбудимость Ia путей [3,4] и нарушения регуляции силы низкого уровня [41–43] во время изометрических сокращений. Хотя механизмы нарушения регуляции силы после инсульта полностью не объяснены, существуют косвенные доказательства изменения координации активности агонистов-антагонистов и увеличения вовлеченности спинномозговых путей с визуальной обратной связью и без нее.Мы прогнозируем, что эффекты, наблюдаемые в настоящем исследовании, могут быть преувеличены в популяции пациентов с инсультом и потенциально могут способствовать дефициту регуляции силы низкого уровня. Наши результаты также могут иметь значение для восстановления после травм, связанных со спортом, которые требуют выполнения упражнений с пониженным весом, но при выполнении функционально значимой задачи. В частности, слишком большая разгрузка ног может быть вредной для контроля, и могут быть пороговые уровни силы, необходимые для улучшения активного диапазона движений без нарушения реакции равновесия.Это подтверждается другими исследованиями, которые показали влияние нагрузки на кинематику и кинетику ходьбы в популяциях пациентов [44].
В частности, слишком большая разгрузка ног может быть вредной для контроля, и могут быть пороговые уровни силы, необходимые для улучшения активного диапазона движений без нарушения реакции равновесия.Это подтверждается другими исследованиями, которые показали влияние нагрузки на кинематику и кинетику ходьбы в популяциях пациентов [44].
Вклад авторов
Задумал и спроектировал эксперименты: AH TO BDS. Проведены эксперименты: ТО АХ. Проанализированы данные: ТО. Написал бумагу: TO AH BDS.
Ссылки
- 1. Янковская Е. Спинальные межнейронные системы: идентификация, многофункциональность и реконфигурации у млекопитающих. J. Physiol 2001; 15; 533 (Pt 1): 31–40.
- 2. Nielsen JB. Сенсомоторная интеграция на уровне позвоночника как основа координации мышц во время произвольных движений у людей. J App Physiol 2004; 96: 1961–1967.
- 3.
Крон С., Йонсен Л.Л., Биринг-Соренсен Ф., Нильсен Дж. Б. Возникновение взаимного облегчения разгибателей голеностопного сустава от сгибателей голеностопного сустава у пациентов с инсультом или травмой спинного мозга. Brain 2003; 126 (2): 495–507.
- 4. Морита Х., Крон С., Кристенхуис Д., Петерсен Н. Т., Нильсен Дж. Б. Модуляция пресинаптического торможения и дисинаптического реципрокного торможения Ia во время произвольного движения при спастичности. Brain 2001; 124: 826–837.
- 5. Христу Е.А., Зелент М., Карлтон Л.Г. Контроль силы больше в верхних конечностях по сравнению с нижними. J Mot Behav 2003; 35: 322–324.
- 6. Грунт И., Хантер Г. Р., Маккарри Б. Д., Болдинг М. С., Рой Д. Л., Маккарти Д. П..Возрастные и гендерные различия в устойчивости разгибания бедра и сгибания. Геронтология 2010; 56: 533–541.
- 7.
Hyngstrom AS, Kuhnen HR, Kirking KM, Hunter SK. Функциональные последствия нарушения контроля субмаксимального сгибания бедра после инсульта. Мышечный нерв 2014; 49: 225–232.
- 8. Трейси Б.Л., Диненно Д.В., Йоргенсен Б., валлийский С.Дж. Старение, зрительно-моторная коррекция и колебания силы в крупных мышцах. Med Sci Sports Exercise 2007; 39: 469–479.
- 9. Трейси Б.Л., Мехудар П.Д., Ортега Д.Д. Амплитуда изменчивости силы коррелирует в мышцах разгибателя колена и сгибателя локтя. Exp Brain Res 2007; 176: 448–464.
- 10. Baweja HS, Kwon M, Christou EA. Усиленная визуальная обратная связь усугубляет изменчивость положения у пожилых людей из-за измененной модуляции основной мышцы-агониста. Exp Brain Res 2012; 222: 355–65.
- 11. Чен Ю.Т., Пинто Нето О, де Миранда Марзулло А.С., Кеннеди Д.М., Фокс Е.Дж., Кристу Е.А.Возрастное нарушение конечной точности целенаправленных сокращений, выполняемых двумя пальцами, связано с измененной активацией синергетических мышц. Exp Gerontol 2012; 47: 519–26.
- 12. Джонс Л.А., Хантер И.В. Влияние вибрации сухожилий мышц на восприятие силы. Exp Neurology 1985; 87: 35–45.
- 13. Thompson S, Gregory JE, Proske U. Ошибки в оценке силы могут быть объяснены десенсибилизацией сухожильных органов. Exp Brain Res 1990; 79: 365–372.
- 14. Aimonetti JM, Vedel JP, Schmied A, Pagni S. Ингибирование против облегчения рефлекторной реакции идентифицированных моторных единиц разгибателей запястья на афферентные входы антагонистов сгибателей у людей. Exp Brain Res 2000; 133: 391–401.
- 15. Кудина Л.П. Влияние сухожильного рефлекса на мотонейроны мышцы-антагониста у человека. Neurophysiolgica 1976; 8: 624–32.
- 16. Онушко Т, Шмит БД.Рефлекторный ответ на наложенные двусторонние колебания бедра при травме спинного мозга человека. J Neurophysiol 2007; 98: 1849–1861.
- 17.
Мэтьюз, ПБК, Мышечные рецепторы млекопитающих и их центральное действие . 1972, Лондон: Арнольд.
- 18. Misiaszek J.E. H-рефлекс как инструмент нейрофизиологии: его ограничения и использование для понимания функции нервной системы. Muscle Nerve 2003; 28: 144–60.
- 19. Voerman GE, Gregoric M, Hermens HJ.Нейрофизиологические методы оценки спастичности: рефлекс Хоффмана, рефлекс сухожилия и рефлекс растяжения. Disabil Rehabil 2005; 27: 33–68.
- 20. Baweja HS, Patel BK, Martinkewiz JD, Vu J, Christou EA. Удаление визуальной обратной связи изменяет мышечную активность и снижает изменчивость силы во время постоянных изометрических сокращений. Exp Brain Res 2009; 197: 35–47.
- 21. Baweja HS, Kennedy DM, Vu J, Vaillancourt DE, Christou EA.Увеличение количества визуальной обратной связи снижает изменчивость силы за счет уменьшения колебаний силы с 0–1 до 3–7 Гц. Eur J Appl Physiol 2010; 108: 935–43.
- 22.
Кеннеди DM, Христу EA. Большой объем визуальной информации усугубляет контроль силы у пожилых людей во время постоянных изометрических сокращений. Exp Brain Res 2011; 213: 351–61.
- 23. Palmieri RM, Ingersoll CD, Hoffman MA. Рефлекс Хоффмана: методологические соображения и приложения для использования в исследованиях спортивной медицины и спортивной подготовки. J Athl Train 2004; 29: 268–77.
- 24. Симонсен Э.Б., Дайре-Поульсен П. Амплитуда H-рефлекса камбаловидной мышцы человека при ходьбе и беге. J. Physiol 1999; 515: 929–39.
- 25. Бакторп М.В., Ханна Р., Пейн Т.Г., Фолланд, JP. Надежность нервно-мышечных измерений во время взрывных изометрических сокращений, с особым упором на методы нормализации электромиографии. Мышечный нерв 2012; 46: 566–76.
- 26.Бакторп М., Пейн М.Т., Фолланд, JP. Центральная усталость способствует большему снижению взрывной силы, чем максимальная сила при утомлении высокой интенсивности. Exp Physiol 2014; 99: 964–73.
- 27. Перейра Х.М., Спирс В.К., Шлиндер-Делап Б., Юн Т., Нильсон К.А., Хантер С.А. Возрастные и половые различия в устойчивости мышц сгибателей локтя с навязанной когнитивной потребностью. Eur J Appl Physiol 2015; 115: 1367–79.
- 28. Рудрофф Т., Джастис Дж. Н., Мэтьюз С., Цзо Р., Енока Р. М..Мышечная активность с соблюдением нагрузки при утомительных сокращениях мышц-разгибателей колена. Exp Brain Res 2010; 203: 307–16.
- 29. Тейлор AM, Кристу EA, Enoka RM. На колебания силы во время изометрических сокращений влияют многочисленные особенности активности двигательных единиц. J Neurophysiol 2003; 90: 1350–1361.
- 30. Браун П. Корковые двигатели в мышцах человека: волынщик и связанные с ним ритмы. Prog Neurobiol 2000; 60: 97–108.
- 31.
Billot M, Ducly J, Simoneau-Buessinger EM, Ballay Y, Martin A. Является ли совместное сокращение ответственным за снижение максимального крутящего момента в коленном суставе у пожилых мужчин? Age (Дордр) 2014; 36: 899–910.
- 32. Кришнан Ч., Аллен Э.Дж., Уильямс Г.Н. Влияние положения колена на устойчивость силы четырехглавой мышцы и стратегии активации. Мышечный нерв 2012; 43 (4): 563–73.
- 33. Хагбарт К.Э., Мейсфилд В.Г. Фузимоторная система. Adv Exp Med Biol 1995; 384: 259–70.
- 34. Hultborn H, Meunier S, Pierrot-Deseilligny E, Shindo M. Изменения пресинаптического торможения волокон Ia в начале произвольного сокращения человека. J. Physiol 1987; 389: 757–72.
- 35. Бейкер С.Н., Оливье Э., Лимон Р.Н. Когерентные колебания моторной коры головного мозга обезьяны и ЭМГ мышц руки демонстрируют модуляцию, зависящую от задачи. J Phyiol 1997; 501: 225–241.
- 36.
Фермер С.Ф., Бремнер Ф.Д., Халлидей Д.М., Розенберг-младший, Стивенс Дж.А.Частотный состав общих синаптических входов в мотонейроны изучался при произвольном изометрическом сокращении у человека. J. Physiol 1993; 470: 127–155.
- 37. Салениус С., Портин К., Каджола М., Салмелин Р., Хари Р. Кортикальный контроль активации человеческих мотонейронов во время изометрического сокращения. J Neurophsyiol 1997; 77: 3401–3405.
- 38. Billot M, Simoneau EM, Ballay Y, Van Hoecke J, Martin A. Как угол голеностопного сустава изменяет моменты антагонистов и агонистов во время максимальных усилий при тыльном и подошвенном сгибании. Scand J Med Sci Sports 2011; 21: e273 – e281.
- 39. Billot M, Duclay J, Simoneau-Buessinger EM, Ballay Y, Martin A. Является ли совместное сокращение ответственным за снижение максимального крутящего момента в коленном суставе у пожилых мужчин? Возраст 2014; 36: 899–910.
- 40.
Аленторн-Гели Э., Майер Г.Д., Сильверс Х.Дж., Самитьер Дж., Ромеро Д., Лааро-Аро С. и др. Профилактика бесконтактных повреждений передней крестообразной связки у футболистов. Часть 1: Механизмы травм и основные факторы риска. Коленная хирургия Sports Traumatol Arthosc 2009; 17: 705–29.
- 41. Hyngstrom AS, Kuhnen HR, Kirking KM, Hunter SK. Функциональные последствия нарушения контроля субмаксимального сгибания бедра после инсульта. Мышечный нерв 2014; 49 (2): 225–32.
- 42. Лодха Н., Наик С.К., Кумбс С.А., Каурау Дж. Х. Силовой контроль и степень двигательных нарушений при хроническом инсульте. Clin Neurophysiol 2010; 121 (11): 1952–61.
- 43. Лодха Н., Мисра Дж., Кумбс С.А., Кристу Е.А., Каорау Дж. Х.Повышенная вариабельность силы при хроническом инсульте: вклад модуляции силы ниже 1 Гц. 2013; PLoS One 8 (12): e83468.
- 44. Чу VW, Хорнби Т.Г., Шмит Б.Д. Восприятие нагрузок на нижние конечности у перенесших инсульт. Clin Neurophysiol 2015; 126 (2): 372–81.
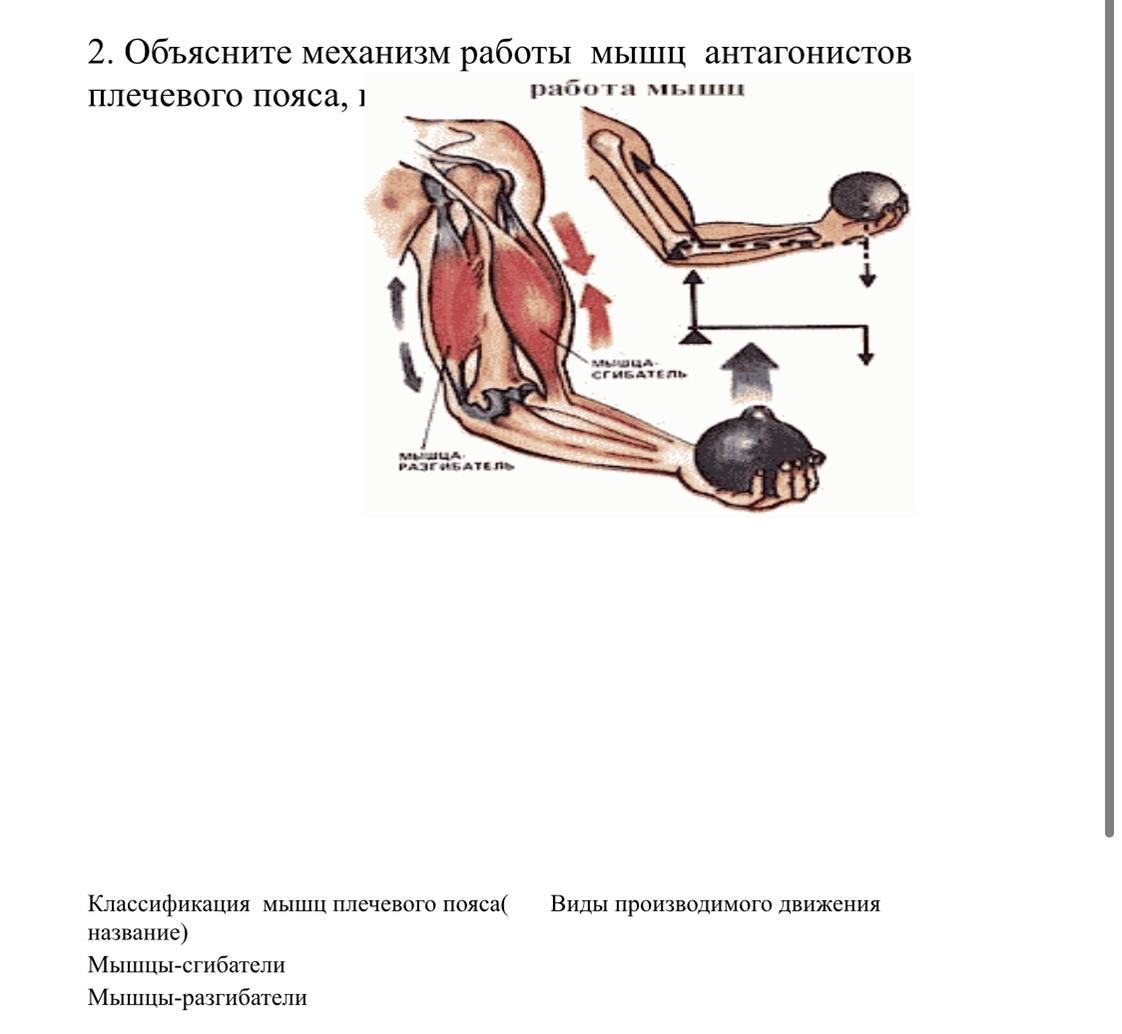 Б. Возникновение взаимного облегчения разгибателей голеностопного сустава от сгибателей голеностопного сустава у пациентов с инсультом или травмой спинного мозга. Brain 2003; 126 (2): 495–507.
Б. Возникновение взаимного облегчения разгибателей голеностопного сустава от сгибателей голеностопного сустава у пациентов с инсультом или травмой спинного мозга. Brain 2003; 126 (2): 495–507. Мышечный нерв 2014; 49: 225–232.
Мышечный нерв 2014; 49: 225–232. Джонс Л.А., Хантер И.В. Влияние вибрации сухожилий мышц на восприятие силы. Exp Neurology 1985; 87: 35–45.
Джонс Л.А., Хантер И.В. Влияние вибрации сухожилий мышц на восприятие силы. Exp Neurology 1985; 87: 35–45.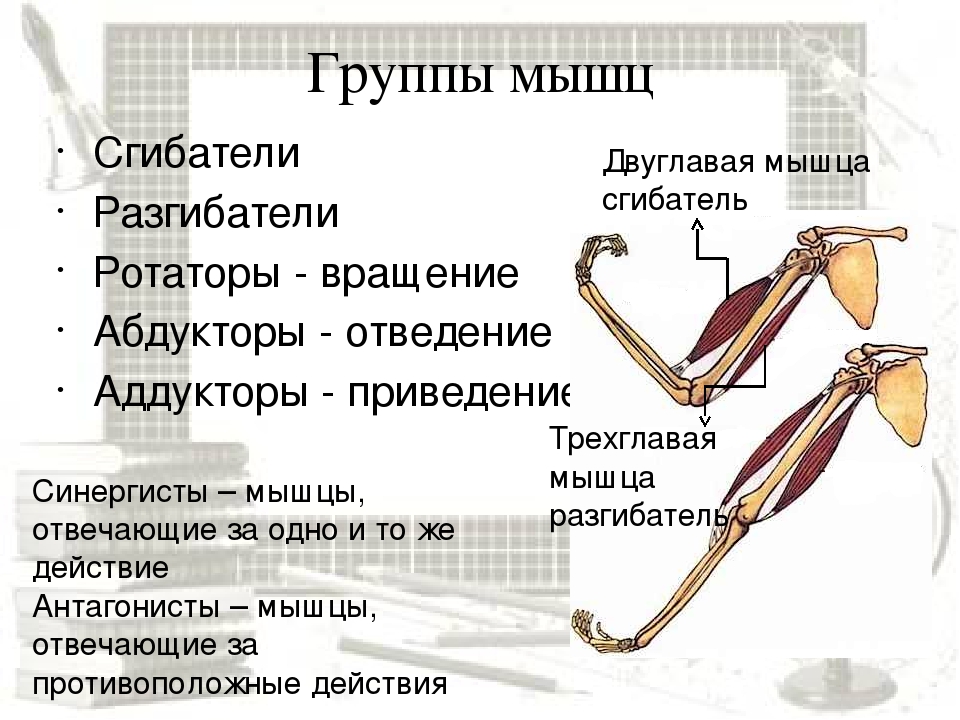 1972, Лондон: Арнольд.
1972, Лондон: Арнольд. Большой объем визуальной информации усугубляет контроль силы у пожилых людей во время постоянных изометрических сокращений. Exp Brain Res 2011; 213: 351–61.
Большой объем визуальной информации усугубляет контроль силы у пожилых людей во время постоянных изометрических сокращений. Exp Brain Res 2011; 213: 351–61. Exp Physiol 2014; 99: 964–73.
Exp Physiol 2014; 99: 964–73.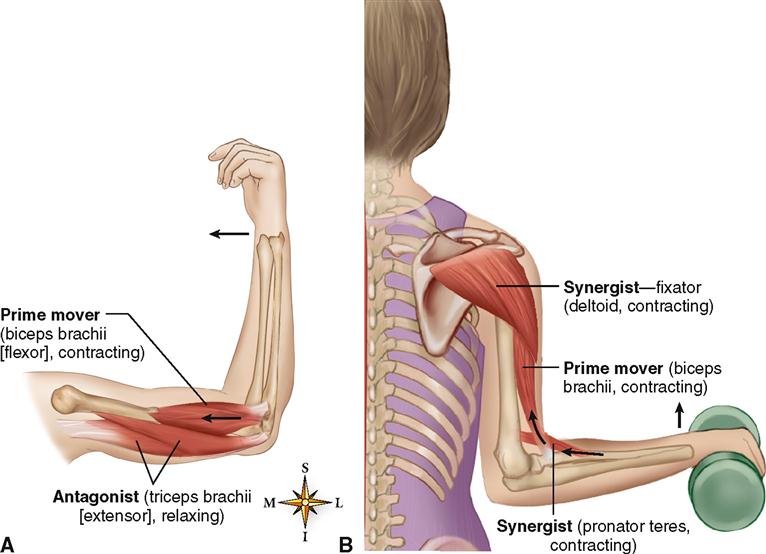
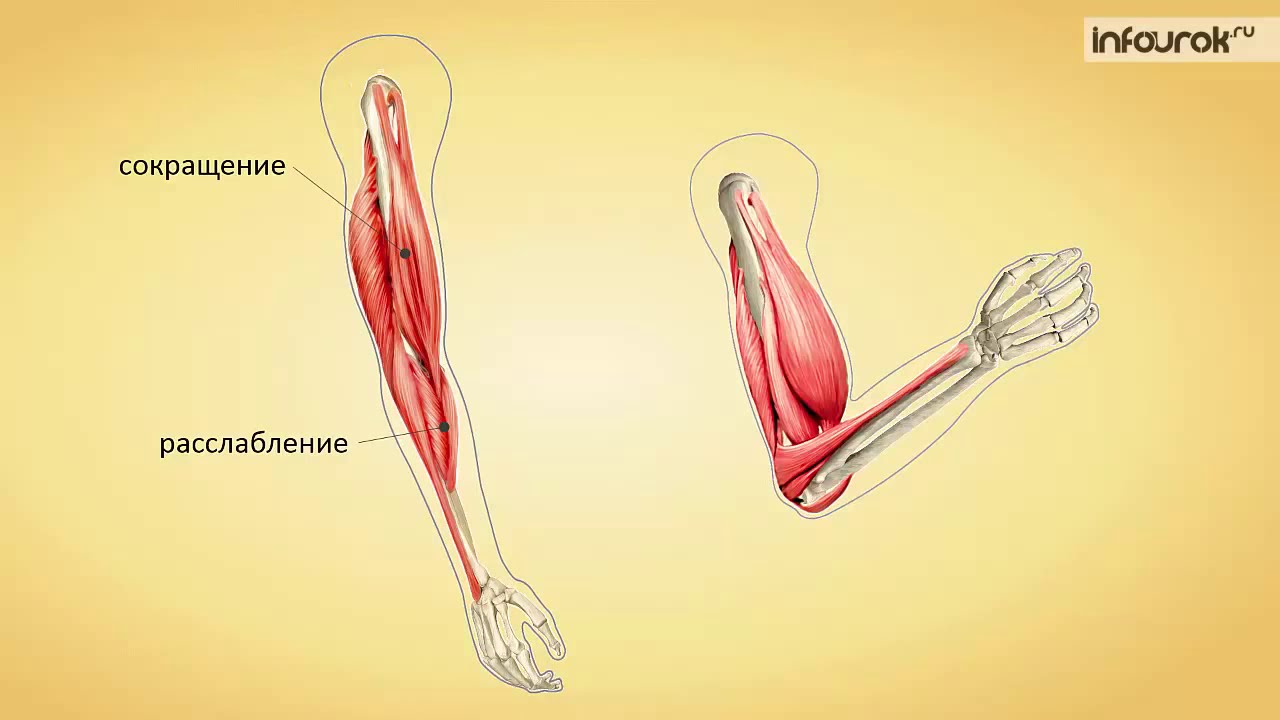
 Коленная хирургия Sports Traumatol Arthosc 2009; 17: 705–29.
Коленная хирургия Sports Traumatol Arthosc 2009; 17: 705–29.Адаптация мышц-агонистов, сопровождающаяся атрофией мышц-антагонистов в задних конечностях мышей после тренировки сокращения-растяжения-сокращения | BMC Musculoskeletal Disorders

Педерсен Б.К., Салтин Б. Физические упражнения как лекарство — данные о назначении физических упражнений в качестве терапии при 26 различных хронических заболеваниях. Scand J Med Sci Sports. 2015; 25 Дополнение 3: 1–72.
Артикул PubMed Google ученый
Тровато Ф.М., Имбеси Р., Конвей Н., Кастрогиованни П. Морфологические и функциональные аспекты скелетных мышц человека. J Funct Morphol Kinesiol. 2016; 1 (3): 289–302.
Артикул Google ученый
Musumeci G, Imbesi R, Trovato FM, Szychlinska MA, Aiello FC, Buffa P, Castrogiovanni P. Важность серотонина (5-HT) и его предшественника l-триптофана для гомеостаза и функции скелетных мышц у крыс. Морфологическое и эндокринологическое исследование. Acta Histochem. 2015; 117 (3): 267–74.
CAS Статья PubMed Google ученый
Musumeci G, Castrogiovanni P, Coleman R, Szychlinska MA, Salvatorelli L, Parenti R, Magro G, Imbesi R.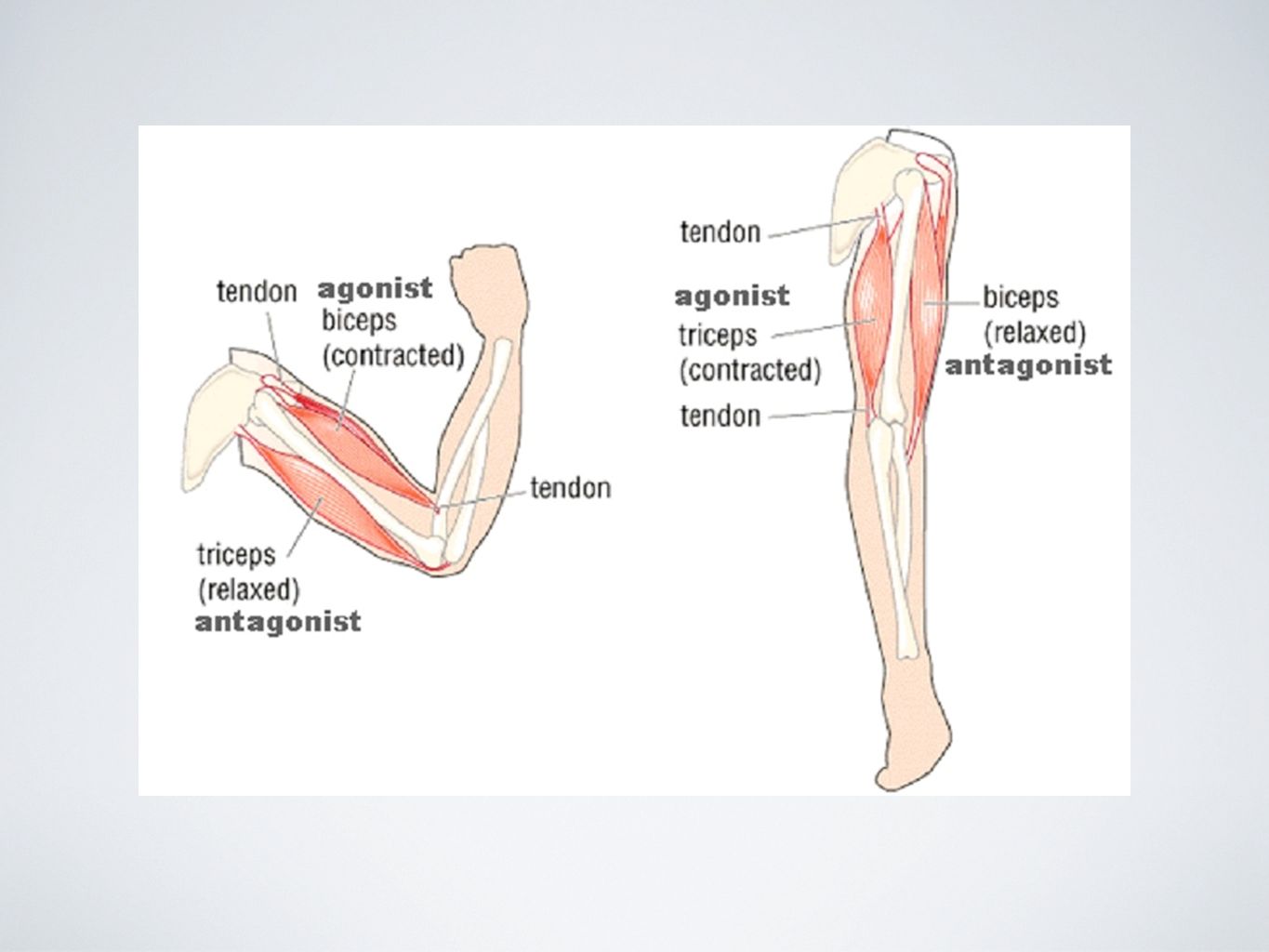 Сомитогенез: от сомита до скелетных мышц. Acta Histochem. 2015; 117 (4–5): 313–28.
Сомитогенез: от сомита до скелетных мышц. Acta Histochem. 2015; 117 (4–5): 313–28.
CAS Статья PubMed Google ученый
Naci H, Ioannidis JP. Сравнительная эффективность упражнений и медикаментозных вмешательств на исходы смертности: метаэпидемиологическое исследование. BMJ. 2013; 347: f5577.
Артикул PubMed PubMed Central Google ученый
Рейдер EP, Миллер Г.Р., Четлин Р.Д., Вирт О., Бейкер Б.А. Произвольная поднятие тяжестей у крыс способствует адаптации за счет производительности и морфологии мышц до набора мышечной массы. Environ Health Insights. 2014; 8 Дополнение 1: 1–9.
Артикул PubMed PubMed Central Google ученый
Giddings CJ, Gonyea WJ. Морфологические наблюдения, подтверждающие гиперплазию мышечных волокон у кошек после выполнения упражнений с поднятием тяжестей. Анат Рек.1992. 233 (2): 178–95.
Анат Рек.1992. 233 (2): 178–95.
CAS Статья PubMed Google ученый
Алвей С.Е., Грумбт WH, Гоньеа В.Дж., Стрэй-Гундерсен Дж. Контрасты в мышцах и миофибриллах у элитных бодибилдеров мужского и женского пола. J Appl Physiol. 1989. 67 (1): 24–31.
CAS PubMed Google ученый
Larsson L, Tesch PA. Плотность волокон двигательных единиц в чрезвычайно гипертрофированных скелетных мышцах человека.Электрофизиологические признаки гиперплазии мышечных волокон. Eur J Appl Physiol Occup Physiol. 1986. 55 (2): 130–6.
CAS Статья PubMed Google ученый
MacDougall JD, Sale DG, Elder GC, Sutton JR. Ультраструктурные характеристики мышц у элитных пауэрлифтеров и бодибилдеров. Eur J Appl Physiol Occup Physiol. 1982. 48 (1): 117–26.
CAS Статья PubMed Google ученый

Klitgaard H, Brunet A, Maton B, Lamaziere C, Lesty C, Monod H. Морфологические и биохимические изменения в мышцах старых крыс: эффект от более широкого использования. J. Appl Physiol (1985). 1989. 67 (4): 1409–17.
CAS Google ученый
Vaczi M, Tihanyi J, Hortobagyi T., Racz L, Csende Z, Costa A, Pucsok J. Механические, биохимические и электромиографические реакции на краткосрочную эксцентрично-концентрическую тренировку разгибателей коленного сустава у людей. J Strength Cond Res.2011; 25 (4): 922–32.
Артикул PubMed Google ученый
Бейкер Б.А., Cutlip RG. Повреждение скелетных мышц в сравнении с адаптацией к старению: новый взгляд на запутанные парадигмы. Exerc Sport Sci Rev.2010; 38 (1): 10–6.
Артикул PubMed Google ученый
Китаока Ю., Наказато К., Огасавара Р. Комбинированные эффекты тренировок с отягощениями и ограничения калорий на митохондриальные белки слияния и деления в скелетных мышцах крыс.J Appl Physiol. 2016; 121 (3): 806–10.
Комбинированные эффекты тренировок с отягощениями и ограничения калорий на митохондриальные белки слияния и деления в скелетных мышцах крыс.J Appl Physiol. 2016; 121 (3): 806–10.
Cutlip RG, Baker BA, Geronilla KB, Mercer RR, Kashon ML, Miller GR, Murlasits Z, Alway SE. Хроническое воздействие сокращений растяжения-сокращения приводит к адаптации скелетных мышц у молодых крыс и дезадаптации у старых крыс. Appl Physiol Nutr Metab. 2006. 31 (5): 573–87.
Артикул PubMed Google ученый
Рейдер Е.П., Лайнер К.Н., Трискит А.М., Четлин Р.Д., Энси Дж., Бейкер Б.А.Возрастная адаптация мышц после хронических сокращений растяжения-сокращения у крыс. Aging Dis. 2015; 7 (1): 1–13.
Рейдер Е.П., Наимо М.А., Лайнер К.Н., Трискит А.М., Четлин Р.Д., Энси Дж., Бейкер Б.А. Укрепление скелетных мышц у старых крыс после высокоинтенсивной тренировки на сокращение-растяжение. Rejuvenation Res. 2016. впереди печати. DOI: 10.1089 / rej.2016.1816.
2016. впереди печати. DOI: 10.1089 / rej.2016.1816.
Такао К., Миякава Т. Геномные ответы на моделях мышей в значительной степени имитируют воспалительные заболевания человека.Proc Natl Acad Sci U S. A. 2015; 112 (4): 1167–72.
CAS Статья PubMed Google ученый
Бейкер Б.А., Холландер М.С., Кашон М.Л., Cutlip RG. Влияние истощения глутатиона и возраста на работоспособность и морфологию скелетных мышц после воздействия хронического сокращения-сокращения растяжения. Eur J Appl Physiol. 2010. 108 (3): 619–30.
CAS Статья PubMed Google ученый
Ingalls CP, Warren GL, Lowe DA, Boorstein DB, Armstrong RB. Дифференциальные эффекты анестетиков на сократительную функцию скелетных мышц in vivo у мышей. J. Appl Physiol (1985). 1996. 80 (1): 332–40.
CAS Google ученый

Borde R, Hortobagyi T, Granacher U. Взаимосвязь между дозой и реакцией тренировок с отягощениями у здоровых пожилых людей: систематический обзор и метаанализ. Sports Med. 2015; 45 (12): 1693–720.
Артикул PubMed PubMed Central Google ученый
Бейкер Б.А., Холландер М.С., Мерсер Р.Р., Кашон М.Л., Cutlip RG. Адаптивные сокращения растяжения-укорачивания: снижение регенерационной способности с возрастом. Appl Physiol Nutr Metab. 2008. 33 (6): 1181–91.
Артикул PubMed Google ученый
Бейкер Б.А., Мерсер Р.Р., Геронилла КБ, Кашон М.Л., Миллер Г.Р., Cutlip RG. Стереологический анализ морфологии мышц после воздействия повторяющихся циклов растяжения-сокращения на модели крысы.Appl Physiol Nutr Metab. 2006. 31 (2): 167–79.
Артикул PubMed Google ученый
Бейкер Б. А., Мерсер Р.Р., Геронилла КБ, Кашон М.Л., Миллер Г.Р., Cutlip RG. Влияние количества повторов на производительность мышц и гистологический ответ. Медико-спортивные упражнения. 2007. 39 (8): 1275–81.
Артикул PubMed Google ученый
Виллемс М.Э., Штаубер В.Т. Влияние силовых тренировок на мышечную усталость и восстановление у интактных крыс.Медико-спортивные упражнения. 2000. 32 (11): 1887–93.
CAS Статья PubMed Google ученый
Виллемс М.Э., Штаубер В.Т. Усталость и восстановление длинных и коротких мышц после эксцентрической тренировки. Медико-спортивные упражнения. 2002. 34 (11): 1738–43.
Артикул PubMed Google ученый
Sinacore DR, Coyle EF, Hagberg JM, Holloszy JO. Гистохимические и физиологические корреляты изменений восстановления после теста на усталость, вызванных тренировкой и снижением тренированности. Phys Ther. 1993. 73 (10): 661–7.
Phys Ther. 1993. 73 (10): 661–7.
CAS PubMed Google ученый
Смит Л.Р., Мейер Г., Либер Р.Л. Системный анализ биологических сетей в функции скелетных мышц. Wiley Interdiscip Rev Syst Biol Med. 2013; 5 (1): 55–71.
CAS Статья PubMed Google ученый
Оюка Э.О. Роль киназы кальция и АМФ в регуляции митохондриального биогенеза и уровней GLUT4 в мышцах.Proc Nutr Soc. 2004. 63 (2): 275–8.
CAS Статья PubMed Google ученый
Стоппани Дж., Хильдебрандт А.Л., Сакамото К., Камерон-Смит Д., Гудиер Л.Дж., Нойфер П.Д. АМФ-активированная протеинкиназа активирует транскрипцию генов UCP3 и HKII в скелетных мышцах крысы. Am J Physiol Endocrinol Metab. 2002. 283 (6): E1239–1248.
CAS Статья PubMed Google ученый
Хьюз С.М., Чи М.М., Лоури О.Н., Гундерсен К. Миогенин вызывает сдвиг активности фермента от гликолитического к окислительному метаболизму в мышцах трансгенных мышей. J Cell Biol. 1999; 145 (3): 633–42.
CAS Статья PubMed PubMed Central Google ученый
Zhu LN, Ren Y, Chen JQ, Wang YZ. Влияние миогенина на типы мышечных волокон и ключевые метаболические ферменты у мышей-переносчиков генов и миобластов C2C12. Ген. 2013; 532 (2): 246–52.
CAS Статья PubMed Google ученый
Чен Д., Ван И, Чин Э.Р. Активация стрессовой реакции эндоплазматического ретикулума в скелетных мышцах мышей с боковым амиотрофическим склерозом G93A * SOD1. Front Cell Neurosci. 2015; 9: 170.
PubMed PubMed Central Google ученый
Гробет Л., Пироттин Д., Фарнир Ф., Понселе Д., Ройо Л.Дж., Брауэрс Б., Кристиан Э., Десмехт Д., Койньуль Ф., Кан Р., Жорж М.Регулирование массы скелетных мышц путем постнатальной мышечной инактивации гена миостатина. Бытие. 2003. 35 (4): 227–38.
CAS Статья PubMed Google ученый
Nishi M, Yasue A, Nishimatu S, Nohno T., Yamaoka T., Itakura M, Moriyama K, Ohuchi H, Noji S. Миссенс-мутантный миостатин вызывает гиперплазию без гипертрофии в мышцах мыши. Biochem Biophys Res Commun. 2002. 293 (1): 247–51.
CAS Статья PubMed Google ученый
Кларк Д.Л., Кларк Д.И., Хоган Е.К., Крошер К.А., Дилгер А.С. Повышенная экспрессия инсулиноподобного фактора роста 2 может вносить вклад в гипермышечный фенотип мышей без миостатина. Гормона роста IGF Res. 2015; 25 (5): 207–18.
CAS Статья PubMed Google ученый
Van Laere AS, Nguyen M, Braunschweig M, Nezer C, Collette C, Moreau L, Archibald AL, Haley CS, Buys N, Tally M, Andersson G, Georges M, Andersson L. Регуляторная мутация в IGF2 вызывает основной эффект QTL на рост мышц у свиней.Природа. 2003. 425 (6960): 832–6.
Артикул PubMed Google ученый
Gardan D, Gondret F, Van den Maagdenberg K, Buys N, De Smet S, Louveau I. Липидный метаболизм и клеточные особенности скелетных мышц и подкожной жировой ткани у свиней, различающихся генотипом IGF-II. Domest Anim Endocrinol. 2008. 34 (1): 45–53.
CAS Статья PubMed Google ученый
Йошихара Т., Мачида С., Куросака Ю., Какиги Р., Сугиура Т., Наито Х. Иммобилизация вызывает накопление ядер HDAC4 в скелетных мышцах крысы. J Physiol Sci. 2016; 66 (4): 337–43.
CAS Статья PubMed Google ученый
Chopard A, Hillock S, Jasmin BJ. Молекулярные события и сигнальные пути, участвующие в атрофии, вызванной неиспользованием скелетных мышц, и влияние контрмер. J Cell Mol Med. 2009. 13 (9B): 3032–50.
Артикул PubMed PubMed Central Google ученый
Griffin CA, Kafadar KA, Pavlath GK. MOR23 способствует регенерации мышц и регулирует клеточную адгезию и миграцию. Dev Cell. 2009. 17 (5): 649–61.
CAS Статья PubMed PubMed Central Google ученый
Pichavant C, Pavlath GK. Частота и тяжесть ветвления миофибрилл при регенерации и старении.Скелетная мышца. 2014; 4: 9.
Артикул PubMed PubMed Central Google ученый
Хайдс Дж., Стэнтон В. Мышечный дисбаланс среди элитных австралийских футболистов по правилам: продольное исследование изменений в размере мышц туловища. J Athl Train. 2012; 47 (3): 314–9.
PubMed PubMed Central Google ученый
Стр. Сенсомоторная тренировка: «глобальный» подход к тренировке равновесия.J Bodyw Mov Ther. 2006. 10 (1): 77–84.
Артикул Google ученый
Каллен Д.М., Бойл Дж.Дж., Силберт П.Л., певица ЛЮ, певица КП. Инъекции ботулинического токсина для облегчения реабилитации синдромов мышечного дисбаланса в спортивной медицине. Disabil Rehabil. 2007. 29 (23): 1832–9.
CAS Статья PubMed Google ученый
Kim SB, You JS, Kwon OY, Yi CH.Пояснично-тазовые кинематические характеристики гольфистов с ограниченным вращением бедра. Am J Sports Med. 2015; 43 (1): 113–20.
Артикул PubMed Google ученый
Ramaswamy KS, Palmer ML, van der Meulen JH, Renoux A, Kostrominova T.Y, Michele DE, Faulkner JA. Боковая передача силы нарушена в скелетных мышцах дистрофических мышей и очень старых крыс. J Physiol. 2011; 589 (Pt 5): 1195–208.
CAS Статья PubMed PubMed Central Google ученый
Dubinska-Magiera M, Jablonska J, Saczko J, Kulbacka J, Jagla T., Daczewska M. Вклад малых белков теплового шока в развитие и функцию мышц. FEBS Lett. 2014. 588 (4): 517–30.
CAS Статья PubMed Google ученый
Hirai H, Verma M, Watanabe S, Tastad C, Asakura Y, Asakura A. MyoD регулирует апоптоз миобластов посредством опосредованной микроРНК подавляющей регуляции Pax3. J Cell Biol. 2010. 191 (2): 347–65.
CAS Статья PubMed PubMed Central Google ученый
Bahrami BF, Ataie-Kachoie P, Pourgholami MH, Morris DL. p70 Киназа рибосомного протеина S6 (Rps6kb1): обновление. J Clin Pathol. 2014. 67 (12): 1019–25.
Артикул Google ученый
Harada H, Andersen JS, Mann M, Terada N, Korsmeyer SJ. Киназа p70S6 сигнализирует о выживании и росте клеток, инактивируя проапоптотическую молекулу BAD.Proc Natl Acad Sci U S. A. 2001; 98 (17): 9666–70.
CAS Статья PubMed PubMed Central Google ученый
Кэролан Б., Кафарелли Э. Адаптации коактивации после изометрической тренировки с отягощениями. J. Appl Physiol (1985). 1992. 73 (3): 911–7.
CAS Google ученый
Geertsen SS, Lundbye-Jensen J, Nielsen JB. Усиленное центральное облегчение реципрокного торможения антагонистами в начале тыльного сгибания после взрывной силовой тренировки.J. Appl Physiol (1985). 2008. 105 (3): 915–22.
Артикул Google ученый
Tang H, Veldman MB, Goldman D. Характеристика мышечно-специфического энхансера в промоторе MuSK человека показывает важную роль миогенина в контроле регуляции зависимых от активности генов. J Biol Chem. 2006. 281 (7): 3943–53.
CAS Статья PubMed Google ученый
Шиотани А., Флинт П.В. Состав тяжелых цепей миозина в мышцах гортани крыс после денервации. Ларингоскоп. 1998. 108 (8 Pt 1): 1225–9.
CAS Статья PubMed Google ученый
Slimani L, Micol D, Amat J, Delcros G, Meunier B, Taillandier D, Polge C, Bechet D, Dardevet D, Picard B, Attaix D, Listrat A, Combaret L. Ухудшение передней большеберцовой мышцы атрофия мышц во время восстановления после иммобилизации коррелирует с увеличением площади соединительной ткани, протеолизом и апоптозом.Am J Physiol Endocrinol Metab. 2012; 303 (11): E1335–1347.
CAS Статья PubMed Google ученый
Stevens JE, Pathare NC, Tillman SM, Scarborough MT, Gibbs CP, Shah P, Jayaraman A, Walter GA, Vandenborne K. Относительный вклад активации мышц и размера мышц в момент подошвенного флексора во время реабилитации после иммобилизации. J Orthop Res. 2006. 24 (8): 1729–36.
Артикул PubMed Google ученый
Legerlotz K, Matthews KG, McMahon CD, Smith HK. Паралич, вызванный ботулиническим токсином, приводит к замедлению состава изоформ тяжелой цепи миозина и снижению содержания тайтина в икроножной мышце молодых крыс. Мышечный нерв. 2009. 39 (4): 472–9.
CAS Статья PubMed Google ученый
Дрейк Дж. К., Уилсон Р. Дж., Ян З. Молекулярные механизмы митохондриальной адаптации скелетных мышц к тренировкам с физическими нагрузками. FASEB J. 2016; 30 (1): 13–22.
CAS Статья PubMed Google ученый
Ham DJ, Kennedy TL, Caldow MK, Chee A, Lynch GS, Koopman R. Цитруллин не предотвращает истощение или слабость скелетных мышц у мышей с гипсовой повязкой конечностей. J Nutr. 2015. 145 (5): 900–6.
CAS Статья PubMed Google ученый
Ham DJ, Murphy KT, Chee A, Lynch GS, Koopman R. Введение глицина снижает истощение скелетных мышц на мышиной модели раковой кахексии.Clin Nutr. 2014. 33 (3): 448–58.
CAS Статья PubMed Google ученый
Гилберт К.Л., Стокс К.А., Холл Г.М., Томпсон Д. Реакция гормона роста на 3 различных цикла упражнений у мужчин от 18 до 25 и от 40 до 50 лет. Appl Physiol Nutr Metab. 2008. 33 (4): 706–12.
CAS Статья PubMed Google ученый
Pesta D, Thaler A, Hoppel F, Macek C, Schocke M, Burtscher M.Влияние 10-недельной обычной программы силовых тренировок на работу мышц голени у мальчиков-подростков по сравнению со взрослыми. J Sports Med Phys Fitness. 2014; 54 (2): 147–53.
CAS PubMed Google ученый
Вирт О., Грегори Е.В., Катлип Р.Г., Миллер Г.Р. Контроль и количественная оценка произвольной работы крыс в поднятии тяжестей. J. Appl Physiol (1985). 2003. 95 (1): 402–12.
Артикул Google ученый
Повышение общих входов агонистов-агонистов и агонистов-антагонистов, связанных с трудностями при стоянии, обусловлено кортикоспинальными и подкорковыми входами соответственно
Кришнамурти В., Гудман С., Зациорский В. и Латаш М. Л. Мышечная синергия во время смещения центра давления стоячими людьми: определение мышечных режимов. Biol. Киберн. 89 , 152–161 (2003).
Артикул Google ученый
Кришнамурти В., Латаш М. Л., Шольц Дж. П. и Зациорский В. М. Мышечные режимы при смещении центра давления стоячими людьми: эффект нестабильности и дополнительная поддержка. Exp. Brain Res. 157 , 18–31 (2004).
Артикул Google ученый
Имагава, Х., Хагио, С. и Кузаки, М. Синергетическая совместная активация в многонаправленном контроле позы у людей. J. Electromyogr. Кинезиол. 23 , 430–437 (2013).
Артикул Google ученый
Danna-Dos-Santos, A. et al. .Управление несколькими мышцами при двуногом положении: подход к анализу ЭМГ-ЭМГ. Exp. Brain Res. 232 , 75–87 (2014).
Артикул Google ученый
Danna-Dos-Santos, A. et al. . Влияние визуальной информации на управление несколькими мышцами во время спокойной позиции: подход спектрального анализа. Exp. мозг Res. 233 , 657–669 (2015).
Артикул Google ученый
Semmler, J. G. Синхронизация двигательных единиц и нервно-мышечные функции. Exerc. Sport Sci. Ред. 30 , 8–14 (2002).
Артикул Google ученый
Винтер Д. А. Биомеханика и моторный контроль движений человека . (Джон Вили и сыновья, 2009 г.).
Винтер, Д. А., Принс, Ф., Франк, Дж. С., Пауэлл, К. и Забжек, К. Ф. Единая теория относительно баланса A / P и M / L в спокойной позиции. J. Neurophysiol. 75 , 2334–2343 (1996).
CAS Статья Google ученый
Амиридис, И. Г., Хацитаки, В., Арабатци, Ф. Модификации статического контроля позы у людей, вызванные возрастом. Neurosci. Lett. 350 , 137–140 (2003).
CAS Статья Google ученый
Ватанабэ, Т., Сайто, К., Ishida, K., Tanabe, S. & Nojima, I. Координация мышц подошвенных сгибателей во время двуногих и одноногих положений у молодых и пожилых людей. Exp. мозг Res . 1–11 (2018).
Ян, В. К., Ченг, К. Х., Ван, Х. К., Лин, К. Х. и Хсу, В. Л. Мульти-мышечная координация во время сложной стойки. Eur. J. Appl. Physiol. 115 , 1959–1966 (2015).
Артикул Google ученый
Ривз, Н. П., Нарендра, К. С. и Холевицки, Дж. Стабильность позвоночника: шесть слепых и слон. Clin. Биомех. 22 , 266–274 (2007).
Артикул Google ученый
Грюнеберг, К., Блум, Б. Р., Онеггер, Ф. и Аллум, Дж. Х. Дж. Влияние искусственно увеличенной жесткости бедер и туловища на контроль равновесия у человека. Exp. мозг Res. 157 , 472–485 (2004).
Артикул Google ученый
Бунстра, Т. У. и Брейкспир, М. Нейронные механизмы межмышечной когерентности: значение для исправления поверхностной электромиографии. J. Neurophysiol. 107 , 796–807 (2011).
Артикул Google ученый
Де Лука, К. Дж. И Эрим, З. Общее побуждение в двигательных единицах синергетической пары мышц. J. Neurophysiol. 87 , 2200–2204 (2002).
Артикул Google ученый
Гроссе П., Кэссиди М. Дж. И Браун П. Анализ частоты ЭЭГ-ЭМГ, МЭГ-ЭМГ и ЭМГ-ЭМГ: физиологические принципы и клиническое применение. Clin. Neurophysiol. 113 , 1523–1531 (2002).
CAS Статья Google ученый
Обата, Х., Абэ, М. О., Масани, К. и Накадзава, К. Модуляция между двусторонними ногами и внутри односторонних мышечных синергистов изменения постуральной мышечной активности с развитием и старением. Exp. мозг Res. 232 , 1–11 (2014).
Артикул Google ученый
Papegaaij, S., Taube, W., Hogenhout, M., Baudry, S. & Hortobágyi, T. Возрастное снижение моторного коркового торможения во время стояния в различных сенсорных условиях. Фронт. Aging Neurosci. 6 , 126 (2014).
PubMed PubMed Central Google ученый
Nandi, T., Fisher, B.E., Hortobágyi, T. & Salem, G.J. Увеличение медиолатерального покачивания стоя связано с увеличением кортикоспинальной возбудимости и уменьшением ингибирования и облегчения M1. Походка 60 , 135–140 (2018).
Артикул Google ученый
Tokuno, C. D., Taube, W. & Cresswell, A. G. Повышенный уровень моторной корковой возбудимости во время контроля человеческого положения. Acta Physiol. 195 , 385–395 (2009).
CAS Статья Google ученый
Ассоциация, G. A. Всемирной медицинской ассоциации W. M. Хельсинкская декларация: этические принципы медицинских исследований с участием людей. J. Am. Coll. Вмятина. 81 , 14 (2014).
Google ученый
Хеббал, Г. В. и Майсорекар, В. Р. Оценка некоторых задач, используемых для определения руки и ноги. Percept. Mot. Навыки 102 , 163–164 (2006).
Артикул Google ученый
Яо Б., Салениус С., Юэ Г. Х., Браун Р. В. и Лю Дж. З. Влияние выпрямления поверхностной ЭМГ на анализ мощности и когерентности: исследование ЭЭГ и МЭГ. J. Neurosci. Методы 159 , 215–223 (2007).
Артикул Google ученый
Майерс, Л. Дж. и др. . Выпрямление и нелинейная предварительная обработка сигналов ЭМГ для кортико-мышечного анализа. J. Neurosci. Методы 124 , 157–165 (2003).
CAS Статья Google ученый
Постон Б., Данна-душ Сантос, А., Хесунатадас, М., Хамм, Т. М. и Сантелло, М. Распределение коррелированных нервных сигналов в мышцах рук при захвате трехзначных чисел, не зависящее от силы. Дж. Нейрофизиол 104 , 1141–1154 (2010).
Артикул Google ученый
Холлидей, Д. М. и др. . Структура для анализа смешанных временных рядов / данных точечных процессов — теория и применение для изучения физиологического тремора, разрядов отдельных моторных единиц и электромиограмм. Прог. Биофиз. Мол. Биол. 64 , 237–278 (1995).
CAS Статья Google ученый
Розенберг, Дж. Р., Амджад, А. М., Бриз, П., Бриллинджер, Д. Р. и Халлидей, Д. М. Подход Фурье к идентификации функциональной связи между цепочками спайков нейронов. Прог. Биофиз. Мол. Биол. 53 , 1–31 (1989).
CAS Статья Google ученый
Нортон, Дж. А. и Горассини, М. А. Изменения корковой межмышечной когерентности, сопровождающие улучшение двигательных навыков при неполном повреждении спинного мозга. J. Neurophysiol. 95 , 2580–2589 (2006).
Артикул Google ученый
Амджад, А. М., Халлидей, Д. М., Розенберг, Дж. Р. и Конвей, Б. А. Расширенный тест на различие когерентности для сравнения и комбинирования нескольких независимых оценок когерентности: теория и применение к изучению двигательных единиц и физиологического тремора. J. Neurosci. Методы 73 , 69–79 (1997).
CAS Статья Google ученый
Гарсия-Массо, X., Пеллисер-Ченолл, М., Гонсалес, Л. М. и Тока-Эррера, Дж. Л. Сложность задачи по контролю осанки влияет на контроль нескольких мышц во время спокойного стояния. Exp. мозг Res. 234 , 1977–1986 (2016).
Артикул Google ученый
Boonstra, T. W. и др. . Мышечные сети: анализ связности ЭМГ-активности во время постурального контроля. Sci. Отчетность 5 , 17830 (2015).
ADS CAS Статья Google ученый
Фергюсон, К. Дж. Праймер для определения размера эффекта: руководство для клиницистов и исследователей. Prof. Psychol. Res. Практик. 40 , 532 (2009).
Артикул Google ученый
Ричардсон, Дж. Т. Э. Квадрат эта и частичный квадрат эта как меры величины эффекта в образовательных исследованиях. Educ. Res. Ред. 6 , 135–147 (2011).
Артикул Google ученый
Лакенс, Д. Расчет и отчет о размерах эффекта для облегчения накопления научных данных: практическое руководство по t-тестам и ANOVA. Фронт. Psychol. 4 , 1–12 (2013).
Артикул Google ученый
Фармер, С. Ф., Бремнер, Ф. Д., Холлидей, Д. М., Розенберг, Дж. Р. и Стивенс, Дж. А. Частотный состав общих синаптических входов в мотонейроны, изученный во время произвольного изометрического сокращения у человека. J. Physiol. 470 , 127–155 (1993).
CAS Статья Google ученый
Нильсен, Дж. Б. и др. . Снижение общего мотонейронального драйва на пораженной стороне при ходьбе у пациентов с гемиплегическим инсультом. Clin. Neurophysiol. 119 , 2813–2818 (2008).
CAS Статья Google ученый
Мима, Т. и Халлетт, М. Кортико-мышечная когерентность: обзор. J. Clin. Neurophysiol. 16 , 501 (1999).
CAS Статья Google ученый
Шницлер А., Тиммерманн Л. и Гросс Дж. Физиологические и патологические колебательные сети в двигательной системе человека. J. Physiol. 99 , 3–7 (2006).
Google ученый
Холлидей, Д. М., Конвей, Б. А., Фармер, С. Ф. и Розенберг, Дж. Р. Использование электроэнцефалографии для изучения функциональной связи между корковой активностью и электромиограммами во время произвольных сокращений у людей. Neurosci. Lett. 241 , 5–8 (1998).
CAS Статья Google ученый
Boonstra, T. W. и др. . Низкие дозы алкоголя снижают частоту 10-15 Гц, воздействующую на двусторонние мышцы ног при спокойном стоянии. J. Neurophysiol. 100 , 2158–2164 (2008).
CAS Статья Google ученый
Дегани, А. М., Леонард, К. Т. и Данна-дос-Сантос, А. Использование анализа межмышечной когерентности в качестве нового подхода к обнаружению возрастных изменений постуральной мышечной синергии. Neurosci. Lett. 656 , 108–113 (2017).
CAS Статья Google ученый
Саффер М., Киемель Т. и Джека Дж. Анализ когерентности мышечной активности в спокойной позе. Exp. мозг Res. 185 , 215–226 (2008).
Артикул Google ученый
Sozzi, S., Honeine, J. L., Do, M. C. & Schieppati, M.Активность мышц ног во время тандемной стойки и контроль баланса тела во фронтальной плоскости. Clin. Neurophysiol. 124 , 1175–1186 (2013).
Артикул Google ученый
Дидериксен, Дж. Л. и др. . Когерентность поверхностной ЭМГ и общего синаптического входа для моторных нейронов. Frontiers in Human Neuroscience 12 , 207 (2018).
Артикул Google ученый
Boonstra, T. W. и др. . Связанные с утомлением изменения в синхронизации двигательных единиц четырехглавой мышцы ног и поперек ног. J. Electromyogr. Кинезиол. 18 , 717–731 (2008).
CAS Статья Google ученый
Маколи, Дж. Х. и Марсден, К. Д. Физиологический и патологический тремор и ритмический центральный моторный контроль. Мозг 123 , 1545–1567 (2000).
Артикул Google ученый
Ватанабэ, Т., Сайто, К., Исида, К., Танабе, С. и Нодзима, И. Возрастное снижение способности модулировать общий входной сигнал для двусторонних и односторонних сгибателей подошвы во время прямой позы. Наклонять. Frontiers in Human Neuroscience 12 , 254 (2018).
Артикул Google ученый
Papegaaij, S., Baudry, S., Négyesi, J., Taube, W. & Hortobágyi, T. Интракортикальное торможение в камбаловидной мышце снижается во время контроля вертикального положения как у молодых, так и у пожилых людей. Eur. J. Appl. Physiol. 116 , 959–967 (2016).
Артикул Google ученый
Nandi, T. et al. . В положении стоя кортикоспинальная возбудимость пропорциональна скорости КС, тогда как возбудимость M1 зависит от участника. Фронт. Гм. Neurosci . 12 (2018).
Слобунов С., Халлетт М., Цао К. и Ньюэлл К. Модуляция корковой активности в результате произвольного направления постурального колебания: исследование ЭЭГ. Neurosci. Lett. 442 , 309–13 (2008).
CAS Статья Google ученый
Фец, Э. и Чейни, П. Д. Постспайковое облегчение мышечной активности передних конечностей кортикомотонейрональными клетками приматов. J. Neurophysiol. 44 , 751–772 (1980).
CAS Статья Google ученый
Cheney, P. D., Kasser, R. & Holsapple, J. Взаимное влияние отдельных кортикомотонейрональных клеток на активность мышц-разгибателей и сгибателей запястья у приматов. Brain Res. 247 , 164–168 (1982).
CAS Статья Google ученый
В чем разница между мышцами-агонистами и антагонистами?
Когда мы говорим о мышцах-агонистах и антагонистах, мы говорим о ролях, которые конкретные мышцы играют в диапазоне движений.Эти два термина описывают, в частности, мышцы, которые обеспечивают силу при движении, будь то простая ходьба или если вы выполняете тяжелую физическую активность.
Знание роли этих мышц в упражнениях — особенно в случае концентрических, эксцентрических и изометрических сокращений — может помочь вам более целенаправленно подходить к вашей тренировочной программе, которая более эффективно приближает вас к вашим целям в фитнесе. Но что такое мышцы-агонисты и антагонисты и какова их роль в движении мышц? Давайте рассмотрим эти концепции более подробно.
Вместо описания конкретных мышечных групп эти два термина описывают роли, которые любая функционирующая мышца играет в движении:
- с описанием группы, создающей силу, и
- другой, описывающий мышцы в расслабленном состоянии во время движения.
Агонист против мышц-антагонистов
Что такое мышцы-агонисты?Мышцы-агонисты — это мышцы, которые во время любого движения прикладывают силу, которая позволяет этому движению произойти.При ходьбе именно мышцы ног обеспечивают подъем и движение вперед.
Поскольку именно эти мышцы обеспечивают силу, можно легко предположить, что это всегда будут мышцы, которые концентрически сокращаются (укорачиваются) во время движения. Однако это не всегда так. Мышца-агонист — это мышца, которая обеспечивает основное движение и, как таковая, может быть концентрическим, эксцентрическим или изометрическим сокращением.
Чтобы прояснить ситуацию, давайте посмотрим, как работает мышца-агонист при сгибании бицепса:
Мышцы-агонисты при сгибании бицепса
Во время сгибательной части сгибания бицепса (в то время как бицепс представляет собой концентрическое сокращение) сами бицепсы являются агонистами.Это потому, что это мышцы, обеспечивающие силу для движения, следовательно, именно они задействованы во время упражнения.
Однако когда рука вытягивается, бицепсы расслабляются, а трицепсы сокращаются эксцентрически. В этом положении сила исходит от трицепсов, что делает их мышцами-агонистами в этом сценарии.
Что такое мышцы-антагонисты?С другой стороны, мышцы-антагонисты — это мышцы, которые никак не сокращаются во время движения.Это мышцы в состоянии покоя во время выполнения движения. Таким образом, они не будут мешать работе мышц-агонистов. Следовательно, эти мышцы всегда противостоят мышцам-агонистам.
Мышца-антагонист не всегда находится в расслабленном состоянии; однако это зависит от типа выполняемого движения. Иногда мышцы-агонисты могут замедлять или останавливать движение.
Мышцы-антагонисты при сгибании бицепса
Это противоположное соотношение между мышцами-агонистами и антагонистами можно довольно четко увидеть в сгибаниях бицепса, где вместо полного расслабления мышцы-агонисты в движении используются для стабилизации руки, когда она поднимает вес.
Когда рука согнута, трицепсы действуют как антагонисты во время сгибания бицепса. Когда он разгибается, бицепсы действуют как антагонисты. В обоих состояниях мышцы-антагонисты не обеспечивают силы для движения, поскольку они активируются для стабилизации руки на каждом этапе движения.
Это служит для защиты суставов, обеспечивая достаточное напряжение для борьбы с действием силы тяжести, особенно при снижении веса, не снижая эффективности мышц-агонистов, мешая им.
Обладая такими специальными знаниями в своем наборе, вы сможете добиться лучших результатов для себя и других во время тренировок. Есть еще много чего узнать!
Почему бы не погрузиться в захватывающий мир профессионального фитнеса, начав свою карьеру в фитнесе с сертификата персонального обучения? Здесь, в Trifocus Fitness Academy, мы предлагаем ряд специализированных курсов, которые имеют международную аккредитацию и могут быть пройдены онлайн в соответствии с вашим расписанием.
Свяжитесь с нами сегодня или посетите наш веб-сайт, чтобы узнать больше о наших предложениях о курсах фитнеса.
Агонизм и антагонизм мышц плечевого сустава: подход SEMG
Автор делится 10-летними клиническими доказательствами использования поверхностной электромиографии для восстановления здорового и функционального диапазона движений — на примере плеча.
Страницы 41-44
Плечевой суставДля физиотерапевтов и врачей-реабилитологов плечевой сустав является одним из самых сложных суставов тела.Девятнадцать разных мышц имеют разные компоненты и вместе участвуют в любом заданном движении. Плечевой сустав и мышцы переходят из положения четвероногих в положение двуногого, свободно свисая в положениях стоя, лежа или лежа. В вертикальном положении мышцы плеча модулируют и поддерживают нейтральное положение шеи и головы. Хотя плечи в значительной степени независимы друг от друга, они обычно работают в тандеме. Однако каждое плечо может выдерживать независимое действие одновременно.Они поддерживают и передают импульс мышцам локтя и косвенно дистальным миотическим единицам запястья и кисти. 1
Плечевой сустав состоит из нескольких анатомических компонентов. В то время как эти компоненты существуют в четвероногом положении, они переходят в функцию для двуногого положения. Ряд мышц охватывают плечевой сустав, а некоторые мышцы перекрывают строго анатомическое определение положения на плече:
- верхняя область : поднимающая лопатка, надостная мышца, средняя дельтовидная мышца, верхняя трапеция, coracobrachialis
- передняя область : передняя дельтовидная, большая и малая грудные мышцы
- задняя область : подлопаточная мышца, задняя дельтовидная мышца, средняя и нижняя трапеции
- задняя область, латеральная сторона : infraspinatus, teres major и minor, latissimus dorsi
- задняя область, медиальная сторона : большой и малый ромбовидные кости
- нижняя область : передняя зубчатая мышца.
Классически плечевой сустав имеет несколько сегментов движения, которые вместе составляют диапазон движения (ROM):
- похищение
- приведение
- переднее сгибание
- боковое сгибание
- заднее сгибание
- внутреннее вращение
- внешнее вращение.
Правильное понимание физических принципов импульса, инерции и векторной активности имеет первостепенное значение для понимания ROM.Феномен совместной активации или совместного сокращения иллюстрируется наличием активных потенциалов низкого уровня в мышце покоя, в то время как гомологичная контралатеральная мышца активна и движется. 2 У здорового человека потенциалы активной амплитуды движения во время движения мышцы одной конечности не встречаются с какими-либо активными потенциалами в гомологичной мышце другой конечности, когда эта конечность находится в состоянии покоя. Сбалансированное соотношение между мышцами сустава способствует нормальному функционированию, например, способности выполнять движения в течение длительного периода времени без усталости и боли.
Если хотя бы одна мышца сустава не работает, эта мышца будет влиять на функцию всего сустава, ограничивая движение, использование энергии, сопротивления и силы. Следовательно, добровольно или непроизвольно, этот сустав может стать недостаточно задействованным («расколотым»), а контралатеральный сустав будет иметь защитную защиту и станет чрезмерно задействованным. Если наблюдается более значительный дисбаланс, чрезмерно загруженный сустав может в конечном итоге стать дисфункциональным и вызвать утомляемость, триггерные точки и боль.
В этой статье рассматривается, как поставщик PM&R может восстановить здоровую функцию и ROM у пациента, перенесшего миофасциальную травму, которая привела к мышечной боли и триггерных точках. В дальнейшем объем работ будет сосредоточен на использовании поверхностной электромиографии (SEMG) для лечения боли при миофасциальных дисфункциях, а также при острой и хронической боли от травм на основе клинического опыта автора в течение 10 лет. 1-9
Динамическое тестированиеSEMG обычно выполняется менее чем за 15 минут (семь движений по 90 секунд каждое).(Источник: 123RF)
Восстановление здорового функционирования с помощью SEMGМетоды
Мышцы плеча можно тренировать в течение нескольких дней после операции после снятия швов или когда мышцам больше не грозит разрыв. Физиотерапию следует начинать постепенно: сначала задействовать мышцы и движения, требующие меньше энергии, и постепенно переходить ко всем движениям. Тренировку нужно проводить без дополнительного сопротивления (помимо силы тяжести).Он может прогрессировать, добавляя сопротивление по мере переносимости и, в конечном итоге, до уровня эргономических или спортивных потребностей.
SEMG динамическое тестирование, тестирование сустава с помощью классической ROM, состоит из повторяющихся мышечных движений, выполняемых с минимальным уровнем усилия (активности и отдыха) с помощью классической ROM любого сустава. Данные обычно собираются в единицах среднеквадратичного (RMS) микровольт и учитываются только тогда, когда коэффициенты вариации (CV) во время движения и покоя составляют 10% или меньше. 6 Использование динамического тестирования SEMG позволяет не только найти амплитудные потенциалы, лежащие в основе концепции, но также и статистический коэффициент корреляции. 6 Результаты, как положительные, так и отрицательные, формируют основу для агонистических и антагонистических ценностей и отношений (см. Врезку « Clinical Refresher: Agonism vs Antagonism and the Shoulder »). 5
Обучение может начинаться только с биологической обратной связи SEMG, а затем проводиться в сочетании с другими методами, всегда переходя от «простого» к «сложному».«Конечная цель — оптимальное функционирование пациента. 3-5
Динамическое тестированиеSEMG неинвазивно, утомительно или болезненно. Обычно тестирование занимает менее 15 минут; в плече есть семь движений, и проверка любого движения обычно занимает 90 секунд. 7 Тестирование лучше всего проводить квалифицированным клиницистом или под своим наблюдением с использованием оборудования SEMG, которое включает статистический пакет. Статистический пакет должен включать возможность оценивать среднюю (или среднюю) амплитуду во время мышечной активности и отдыха, а также параметры стандартного отклонения, коэффициента вариации и регрессионного анализа.Все эти параметры необходимы для оценки статистики, лежащей в основе амплитудной области. Тестирование может проводиться в частотной области, при этом средняя частота является параметром выбора. Тестирование, лежащее в основе данной статьи, проводилось в амплитудной области.
Большинство статистических пакетов позволяют считывать результаты в положительном элементе (не в необработанном SEMG), который является результатом преобразования Фурье положительных и отрицательных результатов амплитуды в только положительные значения.Только тестирование, которое может показать параметры средних значений, коэффициент вариации, стандартное отклонение и, при необходимости, регрессионный анализ, совместимо с требованиями правила доказательства Дауберта, необходимыми для проверки достоверности и научной ценности тестирования.
Клинические доказательства
Авторские исследования динамического тестирования SEMG были основаны на примерно 6800 плечевых мышцах примерно 850 пациентов, протестированных с помощью классических сегментов ROM движения, упомянутых выше, в соответствии с установленными протоколами. 2,5-7 Две из 19 плечевых мышц, подлопаточная и коракобрахиальная, не могли быть протестированы, однако, из-за их глубокого расположения (в настоящее время электроды SEMG не показывают согласованных показаний, если исследуемые мышцы глубже 1,5 см). Данные были собраны у согласившихся пациентов с одинаковым количеством пациентов мужского и женского пола. Возраст варьировался от 21 до 75, и данные не различались в зависимости от пола или возраста. 2,5,6
Данные отражают результаты только для бессимптомных мышц.Значения амплитудного потенциала (среднеквадратичное значение микровольт) обрабатывали статистически для определения коэффициентов корреляции. Положительные коэффициенты корреляции представляют агонистические отношения, а отрицательные значения представляют антагонистические отношения. 5 Эти исследования показали, что мышцы активны во время любого сегмента движения; во время какого-либо движения не было видно «молчаливых» мышц.
Общая средняя активность, представленная в виде амплитудных потенциалов, составила 29,2 мкВ RMS. Эта амплитуда отмечает самый высокий потенциал активности для всех суставов и средних сегментов движения, протестированных с помощью SEMG при минимальных произвольных сокращениях (MVC), подробно описанных ниже.Таким образом, можно сделать вывод, что плечевой сустав является самым высоким суставом с точки зрения использования мышечной энергии, раннего потенциала утомления от чрезмерного использования и последующей боли. 6
В порядке убывания использования энергии мышцы плечевого сустава работают следующим образом:
- пожимание плечами
- похищение
- боковое сгибание
- внешнее вращение
- заднее сгибание
- внутреннее вращение
- переднее сгибание
- приведение. 6
Общий принцип гомеостаза гласит, что чем меньше энергии используется мышцами для выполнения задачи, тем меньше вероятность того, что у них возникнет повторяющаяся или чрезмерная усталость, боль и / или дисфункция. В плечевом суставе 19 мышц напрямую участвуют в любом движении, поэтому вероятность утомления меньше, чем в других суставах.
Данные динамического исследования автора SEMG неизменно показывают, что все мышцы, которые соприкасаются с данным суставом, активны во время любого вектора движения этого сустава.Активность подтверждается наличием амплитудных потенциалов, которые варьируются от мышцы к мышце и от движения к движению. 6 Учитывая любую последовательность из семи движений, мышцы, которые последовательно движутся в одном и том же векторном направлении во время последовательности, следует рассматривать как агонистические или синергетические. Если они склонны действовать в противоположном направлении, они считаются антагонистическими.
Общие расчеты межмышечных отношений плечевых мышц показаны в Таблице I.Обобщение взаимосвязей показывает, что 137 являются агонистическими, а 102 — антагонистическими. Неравные числа возникают из-за того, что некоторые значения регрессии были слишком близки к нулю, чтобы их можно было считать положительными или отрицательными. 6
Минимальные произвольные сокращения
Динамическое тестирование SEMG через ROM было выполнено автором с наименьшим общим знаменателем усилий — MVC. 7 Такое использование минимального количества энергии не способствует чрезмерной нагрузке на мышцы с такими последствиями, как усталость и боль.Результаты коэффициента корреляции между мышцами плеча на уровне MVC могут стать разными, когда для любого конкретного движения требуется определенное усилие. Однако по мере оптимизации движений мышечное усилие станет меньше, и оптимальное использование мышц плеча может начать напоминать то, что было в исходной MVC.
Общая инграмма или гипотетическое постоянное изменение в мозге, объясняющее наличие памяти (след), будет отличаться для каждой функции плеча с разными коэффициентами корреляции.Цель состоит в том, чтобы отформатировать инграммы, чтобы уменьшить общее усилие действия и, следовательно, избежать усталости и боли.
ЗаключениеСпециалисты по физической медицине и реабилитации занимаются отдельными поврежденными и дисфункциональными мышцами. Поставщикам услуг необходимо понимать ожидаемые «нормальные» ценности и отношения, чтобы продолжить процесс реабилитации. Процесс оптимального функционирования, будь то с точки зрения эргономики или легкой атлетики, может потребовать дальнейшей тонкой настройки и может даже больше зависеть от количественной оценки ожидаемых значений SEMG.Понимание каждой мышцы с точки зрения ее агонистических и антагонистических отношений, как описано слева, может считаться необходимым для картирования этого процесса тонкой настройки.
Клиническое освежение: агонизм против антагонизма и плечоАгонизм , или синергизм, относится к положительной взаимосвязи в сокращении (концентрическом или эксцентрическом) двух или более мышц, которые относятся к данному суставу, на всем протяжении данного набора движений, например диапазона движений. Антагонизм относится к обратной зависимости. Мышца-антагонист может стабилизировать или изменить движение агониста, а мышца-антагонист не отдыхает, пока агонист сокращается. Эти отношения отображены в 17 мышцах плеча, протестированных следующим образом: *
- Передняя дельтовидная мышца:
- агонист : средняя дельтовидная мышца, нижняя трапеция, большая грудная мышца, малая грудная мышца, передняя зубчатая мышца, малая круглая мышца, инфраспинатус, малая ромбовидная мышца, надостная мышца, верхняя трапеция
- антагонистический : широчайшая мышца спины, средняя трапеция, большая круглая мышца, задняя дельтовидная мышца, поднимающая лопатка, большой ромбовидный элемент
- Средняя дельтовидная мышца:
- агонист : нижняя трапеция, средняя трапеция, поднимающая лопатка, малый ромбовидная мышца, надостная мышца, верхняя трапеция
- антагонистические : широчайшая мышца спины, большая грудная мышца, малая грудная мышца, передняя зубчатая мышца, большая круглая мышца, малая круглая мышца, задняя дельтовидная мышца, большая ромбовидная мышца
- Задняя дельтовидная мышца:
- агонист : широчайшая мышца спины, средняя трапеция, передняя зубчатая мышца, большая ромбовидная мышца, верхняя трапеция
- антагонистический : передняя дельтовидная, средняя дельтовидная, нижняя трапеция, большая грудная мышца, малая грудная мышца, инфраспинатус, леватор лопатки, малый ромбовидная мышца, надостная мышца
- Большая грудная мышца:
- агонист : передняя дельтовидная мышца, широчайшая мышца спины, малая круглая мышца, инфраспинатус, малая грудная мышца, передняя зубчатая мышца
- антагонистическая : средняя дельтовидная, нижняя трапециевидная, средняя трапециевидная, задняя дельтовидная, леватор лопатки, большой ромбовидный, малый ромбовидный, надостной, верхний трапециевидный.
- Малая грудная мышца:
- агонист : малая круглая мышца, инфраспинатус, передняя зубчатая мышца
- антагонистическая : задняя дельтовидная мышца, поднимающая лопатка, большой ромбовидный элемент, надостная мышца, верхняя трапеция, большая круглая мышца
- Трапеция верхняя:
- агонист : передняя дельтовидная, средняя дельтовидная, широчайшая мышца спины, нижняя трапеция, средняя трапеция, передняя зубчатая мышца, малая круглая мышца, задняя дельтовидная мышца, поднимающая лопатка, надостная мышца
- антагонист : большая грудная, малая грудная, инфраспинатус, большой ромбовидный, малый ромбовидный
- Средняя трапеция:
- агонист : средняя дельтовидная, широчайшая мышца спины, нижняя трапеция, задняя дельтовидная мышца, поднимающая лопатку, большой ромбовидный, надостной, верхний трапециевидный сустав
- антагонистический : передняя дельтовидная, широчайшая мышца спины, малая круглая мышца, инфраспинатус, малая ромбовидная мышца
- Трапеция нижняя:
- агонистический : передняя дельтовидная, средняя дельтовидная, малая круглая мышца, инфраспинатус, леватор лопатки, большой ромб, малый ромб, надостная мышца, верхняя трапеция, средняя трапеция, передняя зубчатая мышца, большая круглая мышца
- антагонистический : широчайшая мышца спины, задняя дельтовидная мышца, большая грудная мышца, малая грудная мышца
- Надостной мышцы:
- агонист : передняя дельтовидная, средняя дельтовидная, нижняя трапеция, средняя трапеция, малая круглая мышца, леватор лопатки
- антагонистические : широчайшая мышца спины, большая грудная мышца, малая грудная мышца, передняя зубчатая мышца, задняя дельтовидная мышца, подостная мышца, большая ромбовидная мышца, малая ромбовидная мышца
- Поддостная мышца:
- агонист : передняя дельтовидная мышца, широчайшая мышца спины, нижняя трапеция, большая грудная мышца, малая грудная мышца, передняя зубчатая мышца, малая круглая мышца
- антагонистическая : средняя дельтовидная, средняя трапециевидная, задняя дельтовидная
- Ромбовидный мажор:
- агонист : широчайшая мышца спины, нижняя трапеция, средняя трапеция, малая круглая мышца, задняя дельтовидная мышца
- антагонист : передняя дельтовидная, средняя дельтовидная, большая грудная мышца, малая грудная мышца, инфраспинатус, леватор лопатки
- Ромбовидный минор:
- агонист : малая круглая мышца, большая ромбовидная мышца, инфраспинатус, передняя дельтовидная мышца, средняя дельтовидная мышца, нижняя трапеция, средняя трапеция, передняя зубчатая мышца
- антагонистический : задняя дельтовидная мышца, поднимающая лопатка, широчайшая мышца спины, большая грудная мышца, малая грудная мышца
- Круг мажор:
- агонист : широчайшая мышца спины, нижняя трапеция, большая грудная мышца, передняя зубчатая мышца
- антагонистический : передняя дельтовидная, средняя дельтовидная, средняя трапеция, малая грудная мышца
- Круг минор:
- агонист : передняя дельтовидная мышца, широчайшая мышца спины, нижняя трапеция, большая грудная мышца, малая грудная мышца, задняя дельтовидная мышца, инфраспинатус, большой ромбовидный элемент, малый ромб, верхняя надостная мышца трапеции
- антагонистический : средняя дельтовидная мышца, средняя трапеция, передняя зубчатая мышца, поднимающая лопатку
- Широчайшая мышца спины:
- агонист : малая круглая мышца, задняя дельтовидная мышца, инфраспинатус, большая ромбовидная мышца, верхняя трапеция, большая грудная мышца, малая грудная мышца, передняя зубчатая мышца, большая круглая мышца
- антагонистический : передняя дельтовидная, средняя дельтовидная, поднимающая лопатка, малый ромбовид, надостная мышца, нижняя трапеция, средняя трапеция
- Передняя зубчатая мышца:
- агонист : большая круглая мышца, малая круглая мышца, задняя дельтовидная мышца, инфраспинатус, малый ромбовидная мышца, верхняя трапеция, передняя дельтовидная мышца, широчайшая мышца спины, нижняя трапеция, большая грудная мышца, малая грудная мышца
- антагонистический : поднимающая лопатка, надостная мышца, средняя дельтовидная мышца, средняя трапеция
- Лопатка, поднимающая лопатку:
- агонист : средняя дельтовидная, нижняя трапеция, средняя трапеция
- антагонистический : передняя дельтовидная, широчайшая мышца спины, большая грудная мышца, малая грудная мышца, передняя зубчатая мышца, малая круглая мышца, задняя дельтовидная мышца, подостая мышца
* Коэффициенты корреляции для каждой мышцы можно найти в ссылке 6.
Последнее обновление: 2 августа 2019 г.
Выгорание врача: взгляд старика
Что делает мышца-антагонист? | Live Healthy
Лиза Мэлони Обновлено 21 мая 2019 г.
Когда вы выполняете какое-либо упражнение, мышца, которая заставляет движение, то есть та, которая выполняет работу и устает, называется агонистом. Это легко запомнить, потому что, когда вы «чувствуете ожог», это может показаться своего рода агонией.Но ни одна мышца не работает в полной изоляции, и для каждой мышцы, производящей движение, есть другая мышца, которая производит противоположное движение в том же суставе. Эта мышца известна как антагонист. Если он сократится одновременно с агонистом, ваш сустав никогда не двинется. Вместо этого мышца-антагонист обычно удлиняется, позволяя двигаться по мере сокращения противоположной мышцы-агониста.
Совет
Когда мышца сокращается для движения сустава — например, двуглавая мышца сгибает ваш локоть — это называется агонистом.Антагонист — это группа мышц, противоположная активной мышце. В этом примере антагонист — трицепс.
Рассмотрим трицепс и бицепс
Трицепс и бицепс являются одними из лучших примеров для понимания мышц-агонистов / антагонистов. Каждый из них является основным двигателем противоположных движений локтя: ваша двуглавая мышца сгибает вашу руку в локте, а трицепс разгибает руку в локте. Итак, когда вы делаете сгибания на бицепс, ваш бицепс является мышцей-агонистом, потому что он обеспечивает основное движение.Ваш трицепс — мышца-антагонист.
Если вы выполняете обратное движение, выполняя разгибания на трицепс или отжимание с сопротивлением, ваш трицепс становится мышцей-агонистом, потому что он обеспечивает движение, в то время как ваш бицепс становится мышцей-антагонистом, которая пассивно удлиняется, так что ваш трицепс может двигать локтевым суставом.
ID Другие пары мышц
У каждой скелетной мышцы вашего тела есть антагонист, который выполняет противоположное движение в том же суставе. Например, ваши грудные мышцы или мышцы груди качают вместе руки перед вами; ваши мышцы спины являются антагонистами этого движения, потому что, если бы им представилась возможность, они оттянули бы ваши руки назад.Твои квадрицепсы на передней части бедра вытягивают ногу в колене; В этом случае ваши подколенные сухожилия являются антагонистами, которые пассивно поддерживают это движение. Если бы роли мышц поменялись местами и ваши подколенные сухожилия были агонистами, используемыми для сгибания вашей ноги в колене, ваши квадрицепсы стали бы мышцами-антагонистами.
Когда антагонист сгибается
Хотя, как правило, мышца-антагонист пассивно удлиняется, позволяя мышце-агонисту выполнять свою работу, есть несколько заметных исключений.Первый — это изометрические сокращения, при которых ваши мышцы сокращаются, но не производят движения в суставах. Некоторые примеры этого включают сидение у стены, удержание высокой или низкой планки или стояние в положении «вверх» при подъеме на носки.