
Что такое аэробный метаболизм?
Аэробный метаболизм использует кислород для удаления энергии из глюкозы и сохраняет ее в биологической молекуле, называемой аденозинтрифосфатом (АТФ). АТФ является источником энергии человеческого организма, и распадающиеся молекулы АТФ высвобождают энергию, которая используется для различных биологических процессов, включая движение молекул через мембраны. Аэробный метаболизм также называется аэробным дыханием, клеточным дыханием и аэробным клеточным дыханием. Анаэробный метаболизм является еще одной формой метаболизма, но происходит без кислорода, но человеческое тело не способно поддерживать анаэробное дыхание в течение длительного времени, и это вызывает сильный стресс.
Первая стадия аэробного обмена называется гликолизом. Гликолиз происходит в цитоплазме клетки. Сложные сахара расщепляются на глюкозу различными ферментами, и эта глюкоза затем разлагается на две молекулы пировиноградной кислоты, также известные как пируват.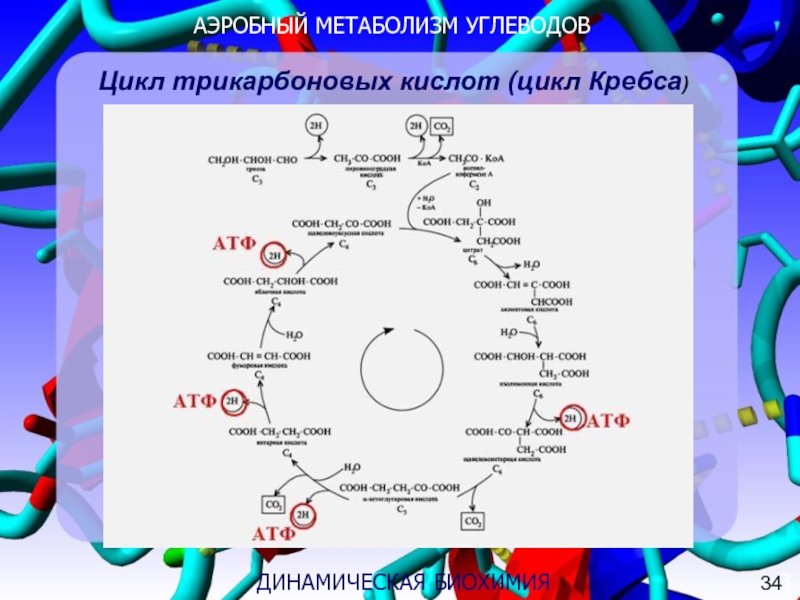
На второй стадии аэробного метаболизма, называемой циклом лимонной кислоты, две молекулы пирувата используются для создания богатых энергией восстанавливающих молекул, которые впоследствии используются в процессе дыхания. Некоторые из этих молекул могут быть преобразованы непосредственно в АТФ, если это необходимо, хотя это не всегда происходит. Вода и углекислый газ образуются как отходы этого цикла, поэтому люди вдыхают кислород и выделяют углекислый газ. Цикл лимонной кислоты, как и гликолиз, дает 2 АТФ.
Финальная стадия аэробного метаболизма называется цепью переноса электронов и происходит на внутренней мембране митохондрий. На этом этапе богатые энергией молекулы, полученные из цикла лимонной кислоты, используются для поддержания градиента положительного заряда, называемого хемиосмотическим градиентом, который используется для генерации многих молекул АТФ.
Аэробный метаболизм генерирует около 36 молекул АТФ. Анаэробное дыхание генерирует только около десяти процентов от этого количества. Использование кислорода является наиболее важным в конце цепи переноса электронов, поскольку оно помогает хемиосмотическому градиенту. Именно поэтому кислородно-зависимый метаболизм является причиной того, что митохондрии широко известны как источник энергии для организма.
ДРУГИЕ ЯЗЫКИ
Метаболизм аэробный — Справочник химика 21
 Главное условие эффективности биологических процессов метаболизма в аэротенке — наличие растворенного в воде кислорода. Для этого проводят аэрацию и перемешивают смесь воды и активного ила пневматическими, механическими или смешанного типа устройствами. [c.101]
Главное условие эффективности биологических процессов метаболизма в аэротенке — наличие растворенного в воде кислорода. Для этого проводят аэрацию и перемешивают смесь воды и активного ила пневматическими, механическими или смешанного типа устройствами. [c.101]Биохимическое потребление кислорода (БПК), используемое для оценки концентрации растворенных в воде веществ, определяется аэробным метаболизмом гетеротрофных микроорганизмов (1-я стадия БПК или углеродное БПК), а также метаболизмом автотрофных организмов (2-я стадия ВПК). Полный расход кислорода О2 в мг/л на этих двух стадиях составляет, [c.147]
При аэробном или анаэробном метаболизме организмы получают энергию в процессе окисления подложки — сахара (глюкозы) или какого-либо другого материала (битума). Это окисление с выделением энергии происходит путем перехода протонов или электронов через ряд стадий, регулируемых ферментами, до появления конечного акцептора электронов. В аэробных процессах конечным акцептором электрона или иона водорода является кислород. В анаэробных процессах таким акцептором является окисленный материал типа нитрата или сульфата. Опыт показал, что аэробный метаболизм эффективнее анаэробного, так как для роста в аэробных процессах требуется меньше материала подложки, чем в анаэробных при одинаковом количественном росте бактерий. Причиной такого явления, известного как эффект Пастера, является большее выделение энергии в процессе аэробного метаболизма. [c.186]
В анаэробных процессах таким акцептором является окисленный материал типа нитрата или сульфата. Опыт показал, что аэробный метаболизм эффективнее анаэробного, так как для роста в аэробных процессах требуется меньше материала подложки, чем в анаэробных при одинаковом количественном росте бактерий. Причиной такого явления, известного как эффект Пастера, является большее выделение энергии в процессе аэробного метаболизма. [c.186]
При работе умеренной интенсивности мышца может покрывать свои энергетические затраты за счет аэробного метаболизма. Однако при больших нагрузках, когда возможность снабжения кислородом отстает от потребности в нем, мышца вынуждена использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость расщеп-
Для завершения окисления жирных кислот ацетильные остатки молекулы ацетил-СоА, образовавшиеся в результате реакций р-окисления, должны быть окислены до двуокиси углерода и воды [14]. Цикл трикарбоновых кислот, в ходе которого осуществляется это окисление, является жизненно важной частью метаболизма почти всех аэробных организмов. Центральное место этого цикла в метаболизме обусловлено еще и тем, что ацетил-СоА образуется также в процессах катаболизма углеводов и некоторых аминокислот. [c.317]
Цикл трикарбоновых кислот, в ходе которого осуществляется это окисление, является жизненно важной частью метаболизма почти всех аэробных организмов. Центральное место этого цикла в метаболизме обусловлено еще и тем, что ацетил-СоА образуется также в процессах катаболизма углеводов и некоторых аминокислот. [c.317]
Брожение является также жизненно важным процессом и для человеческого организма. Хотя в обычных условиях наши мышцы получают вполне достаточные количества кислорода, чтобы произошло окисление пирувата и образование АТР аэробным путем, бывают обстоятельства, когда поступление кислорода оказывается недостаточным. Например, при крайнем напряжении сил, когда уже весь запас кислорода израсходован, мышечные клетки образуют лактат путем брожения. Более того, в белых мышцах рыб или домашней птицы аэробный метаболизм относительно невелик, и основным конечным продуктом оказывается L-лактат. В организме человека есть такие ткани, которые слабо снабжаются кровью, например хрусталик и роговица глаза.
 В клетках этих тканей окислительный метаболизм выражен слабо, а энергия в основном образуется при сбраживании глюкозы в лактат. [c.345]
В клетках этих тканей окислительный метаболизм выражен слабо, а энергия в основном образуется при сбраживании глюкозы в лактат. [c.345]Четвертый этап — воздействие продуктов метаболизма, образующихся в результате жизнедеятельности колоний микроорганизмов, на материал конструкции (кислотное, щелочное, окислительное и ферментативное). Несовершенные грибы (аэробные гетеротрофы) стимулируют коррозию металлов следующим образом. [c.53]
Наряду с аэробным метаболизмом углеводов мозговая ткань способна к довольно интенсивному анаэробному гликолизу. Значение этого явления [c.633]
Ряд культур дрожжей, в том числе Sa haromy es, в условиях недостаточного обеспечения среды кислородом и при наличии углеводов получают энергию путем анаэробного расщепления сахаров (гликолиз) при этом образуется этанол. Как только в среде появляется кислород, клетки дрожжей сразу переключаются на энергетически более выгодный аэробный метаболизм (Пастеровский эффект) и способны метаболизировать не только глюкозу, но и накопившийся в среде этанол.
АЭРОБНЫЙ МЕТАБОЛИЗМ ПИРУВАТА [c.343]
Среди факультативных анаэробов в условиях осуществления ими метаболизма аэробного типа также могут быть микроаэрофилы. [c.129]
ЛЮЦИИ живых существ, так и в преобразовании минералов. В результате превращения цитохромов в терминальные оксидазы и использования молекулярного кислорода в качестве акцептора электронов у бактерий стал возможным новый тип метаболизма-аэробное дыхание. [c.521]
Определение скорости окисления оказало большую помощь при изучении метаболических процессов у бактерий. Однако очевидно, что при изучении роли цикла трикарбоновых кислот в обмене веществ у бактерий полученные результаты необходимо интерпретировать с большой осторожностью. Хотя теперь признано, что в метаболизме аэробных бактерий этот цикл играет центральную роль, в конце сороковых годов считалось, что у бактерий он отсутствует.
Аэрация. Кислород необходим для метаболизма аэробных микроорганизмов, участвующих в компостировании. Аэрация может осуществляться естественной диффузией в компостируемую массу благодаря перемешиванию компоста вручную, с помощью механизмов или принудительной аэрации. Естественной диффузии часто оказывается недостаточно для адекватной аэрации на ранних стадиях процесса, что ведет к анаэробиозу в центральных зонах компостируемой массы. [c.240]
Специфические приспособления для защиты нитрогеназы от высоких концентраций О2 выработаны симбиотическими азотфиксатора-ми — клубеньковыми бактериями. Уже сами клубеньки, места активного размножения бактерий и фиксации ими N2, следует рассматривать как структуру, одним из назначений которой является ограничение доступа внутрь молекулярного кислорода. Эту же функцию выполняет содержащийся в клубеньках леггемоглобин (белок, аналогичный гемоглобину), способный активно связывать О2 и контролировать его поступление в бактероиды.
Но, как часто случается, жизнь сумела обойти это препятствие и даже обратила его в преимущество. Отходами жизнедеятельности первичных простейших организмов были такие соединения, как молочная кислота и этанол. Эти вещества намного менее энергоемки по сравнению с сахарами, но они способны высвобождать большое количество энергии, если полностью окисляются до СО2 и Н2О. В результате эволюции возникли живые организмы, способные фиксировать опасный кислород в виде Н2О и СО2, а взамен получать энергию сгорания того, что прежде было их отходами. Так появился аэробный метаболизм.
Значительный успех на этом пути снова был достигнут благодаря процессам координационной химии. Центральную роль в механизме аэробного метаболизма, который приводит к полному сгоранию органических молекул, играют цитохромы.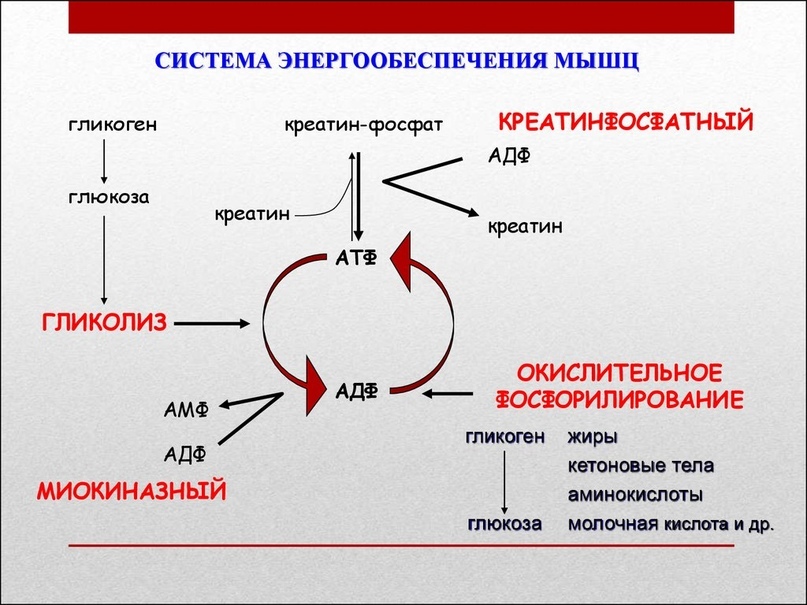 Так называются молекулы, в которых атом железа связан в комплекс с порфирином, образуя с ним гем (см. рис. 20-20), а гем связан с белком. Атом железа переходит из состояния окисления 4- 2 в + 3 и обратно в результате переноса электронов от одного компонента цепи к другому. Весь аэробный механизм представляет собой совокупность тесно связанных друг с другом окислительно-восстано-вительных реакций, окончательным результатом которых является процесс, обратный фотосинтезу [c.257]
Так называются молекулы, в которых атом железа связан в комплекс с порфирином, образуя с ним гем (см. рис. 20-20), а гем связан с белком. Атом железа переходит из состояния окисления 4- 2 в + 3 и обратно в результате переноса электронов от одного компонента цепи к другому. Весь аэробный механизм представляет собой совокупность тесно связанных друг с другом окислительно-восстано-вительных реакций, окончательным результатом которых является процесс, обратный фотосинтезу [c.257]
Аэробный метаболизм пирувата. Этот процесс выходит за рамки углеводного обмена, однако может рассматриваться как завершающая его стадия окисление продукта гликолиза—пирувата. [c.319]
Изучалось влияние аэробной микрофлоры в речной воде на скорость коррозии [7]. Появление нового деполяризатора — микроорганизмов — стимулирует процесс коррозии в связи с накоплением ионов Н 0+ в продуктах метаболизма [c. 28]
28]
Представление об основных биохимических процессах, происходящих в клетках, на примере сапрофитных микроорганизмов с аэробным типом питания [2], дает упрощенная схема метаболизма на рис. 1.2. Даже в таком упрощенном виде схема позволяет оценить многообразие и сложность внутриклеточных процессов, насчитывающих несколько тысяч реакций, в результате которых синтезируются клеточные вещества. Математическое описание всей совокупности данных реакций и использование такой модели для практических целей представляет собой чрезвычайно сложную задачу. Наряду с микробиологическими процессами, направленными на образование биомассы микроорганизмов или ценных продуктов клеточного метаболизма большую роль в БТС занимают процессы биологической очистки, протекающие с участием бактериальных клеток по следующей трофической схеме органические загрязнениям бактерии-> простейшие. В процессе биологической очистки сточных вод, содержащих органические и минеральные вещества, формируется биоценоз активного ила, включающий бактерии, простейшие и многоклеточные организмы. В процессе потребления органических загрязнений происходит интенсивный рост бактерий и ферментативное окисление органических веществ. По мере удаления из среды питательных веществ происходит эндоген- [c.10]
В процессе потребления органических загрязнений происходит интенсивный рост бактерий и ферментативное окисление органических веществ. По мере удаления из среды питательных веществ происходит эндоген- [c.10]
ДЫХАНИЕ, совокупность процессов, обеспечивающих поступление в организм атмосферного или растворенного в воде О2, использование его в окислит.-восстановит. р-циях, а также удаление из организма СО и нек-рых др. соед.-конечных продуктов обмена в-в. Играет фундам. роль в энергообеспечении и метаболизме у большинства организмов. При Д. кислород участвует гл. обр. в окислении орг. соед. с. образованием Н О или HjO (в нек-рых случаях-О ) или включается в молекулу окисляемого в-ва. Нек-рые организмы (гл. обр. мн. бактерии) могут использовать в качестве акцептора электронов не только О , но и др. соед. с высоким сродством к электрону, напр, нитраты и сульфаты. В этих случаях иногда говорят о нитратном и сульфатном Д. в отличие от аэробного (кислородного) Д. [c. 124]
124]
Ввиду исключительной важности процессов окисления для метаболизма у аэробных организмов часто бывает весьма полезно знать свободную энергию сгорания того или иного соединения. Эти данные легко получить из свободной энергии образования. Например, АОс ацетата (водн) можно найти следующим образом [c.227]
Мочевина СН4Н20 является конечным продуктом метаболизма белков в организмах животных. Допустим, что аэробные бактерии могут разлагать ее следующим образом [c.169]
Метаболизм рассматриваемого кремнезема исследовался на процессах роста диатомей путем измерения эффектов, возникающих при недостаточном содержании кремнезема в растворе. Левин обнаружил, что кремнезем не поглощался промываемыми клетками диатомей до тех пор, пока в систему не добавлялось соединение серы. Кадмий затормаживал поглощение кремнезема, возможно, потому, что связывал соединение серы. Поглощение кремнезема представляет собой также, аэробный процесс [51]. Соотношение между поглощениями изотопов и СОг [c.1016]
Соотношение между поглощениями изотопов и СОг [c.1016]
Цикл трикарбоновых кислот, являясь одним из наиболее важных циклов метаболизма аэробных организмов (бактерий, простейших, грибов, высших растений и человека), представляет собой к тому же типичный каталитический цикл. Во всех других циклах также принимают участие один или несколько первичных субстратов и по меньшей мере один регенерирующийся субсграт. Таким образом, с каталитическим циклом всегда ассоциирован метаболический путь, обеспечивающий синтез регенерирующегося субстрата. Хотя, как правило, и не требуется, чтобы такой синтез шел быстро, поскольку обычно бывает необходимо восполнить лишь небольшие потери регенерирующегося субстрата в побочных реакциях, однако используемый при этом метаболический путь обеспечивает механизм биосинтеза любых необходимых количеств любого промежуточного продукта, образующегося в ходе цикла. Так, клетки получают из цикла трикарбоновых кислот значительные количества оксалоацетата, а-кетоглутарата и сукци-нил-СоА, используя их для синтеза других клеточных компонентов. Например, аспартат и глутамат образуются непосредственно из оксалоацетата и а-кетоглутарата путем переаминирования [уравнение (8-16)]. Часто говорят, что цикл трикарбоновых кислот работает на биосинтез, однако, строго говоря, когда из цикла выводятся промежуточные продукты, работает неполный цикл. Правильнее сказать, что метаболический путь синтеза регенерирующегося субстрата и еще некоторые из ферментов цикла используются для формирования тех или иных путей биосинтеза. [c.323]
Например, аспартат и глутамат образуются непосредственно из оксалоацетата и а-кетоглутарата путем переаминирования [уравнение (8-16)]. Часто говорят, что цикл трикарбоновых кислот работает на биосинтез, однако, строго говоря, когда из цикла выводятся промежуточные продукты, работает неполный цикл. Правильнее сказать, что метаболический путь синтеза регенерирующегося субстрата и еще некоторые из ферментов цикла используются для формирования тех или иных путей биосинтеза. [c.323]
Некоторое представление о важнейших биохимических процессах в клетках сапрофитов с аэробным типом обмена веществ дает упрощенная схема метаболизма (рис. 14). [c.36]
Регуляция процессов активного транспорта, обеспечивающего поступление подавляющего большинства необходимых прокариотам веществ, происходит на уровне синтеза переносчика и его функционирования. Биосинтез белковых компонентов многих транспортных систем регулируется по типу индукции. Глюкоза, транспортная система которой у большинства прокариот конститутивна, подавляет образование транспортных систем других сахаров и ряда органических кислот путем катаболитной репрессии. Исключение составляют некоторые облигатно аэробные прокариоты, у которых транспорт органических кислот конститутивен, а индуцируемой является транспортная система глюкозы. Избыток субстрата в среде может репрессировать синтез соответствующей транспортной системы. Это особенно характерно для аминокислот. В этом случае регуляция транспорта координирована с регуляцией их последующего метаболизма. Обнаружена также регуляция транспорта по типу отрицательной обратной связи, когда субстрат, [c.124]
Исключение составляют некоторые облигатно аэробные прокариоты, у которых транспорт органических кислот конститутивен, а индуцируемой является транспортная система глюкозы. Избыток субстрата в среде может репрессировать синтез соответствующей транспортной системы. Это особенно характерно для аминокислот. В этом случае регуляция транспорта координирована с регуляцией их последующего метаболизма. Обнаружена также регуляция транспорта по типу отрицательной обратной связи, когда субстрат, [c.124]
О2 (последний не участвует в осуществляемых ими метаболических реакциях), но способные расти в его присутствии, являются по типу осуществляемого ими метаболизма облигатными анаэробами, устойчивыми к О2 внешней среды. Примером таких организмов служат молочнокислые бактерии. Многие прокариоты, относящиеся к этой же группе, приспособились в зависимости от наличия или отсутствия О2 в среде переключаться с одного метаболического пути на другой, например с дыхания на брожение, и наоборот. Такие организмы получили название факультативных анаэробов, или факультативных аэробов. Представителями этой физиологической группы прокариот являются энтеробактерии. В аэробных условиях они получают энергию в процессе дыхания. В анаэробных условиях источником энергии для них служат процессы брожения или анаэробного дыхания. [c.129]
Такие организмы получили название факультативных анаэробов, или факультативных аэробов. Представителями этой физиологической группы прокариот являются энтеробактерии. В аэробных условиях они получают энергию в процессе дыхания. В анаэробных условиях источником энергии для них служат процессы брожения или анаэробного дыхания. [c.129]
Примерно 1,5—2 10 лет назад парциальное давление Оа в атмосфере достигло 0,02—0,207о современного уровня. При этом начал возникать аэробный метаболизм, дыхание. При клеточном дыхании происходит ряд взаимосвязанных процессов синтеза биологических молекул, необходимых для жизни, и зарядка АТФ (окислительное фосфорилирование). Молекулы пищевых веществ сгорают , окисляются до СОг и НаО, причем Оа служит конечным акцептором водорода. Освобождение химической энергии из пищи происходит, грубо говоря, в трех фазах. Первая состоит в расщеплении макромолекул и молекул жиров. Из белков получаются аминокислоты, из углеводов (крахмал, гликоген)—гексо-зы, из жиров — глицерин и жирные кислоты. Из этих веществ [c.53]
Из этих веществ [c.53]
Реакция г в табл. 8-4, напротив, не может быть осуществлена системой пиридиннуклеотидов вследствие неподходящего восстановительного потенциала. Необходима более сильная окисляющая система флавинов. (Однако обратная реакция, гидрирование связи С = С, частО протекает в биологических системах с участием восстановленного пи-ридиннуклеотида.) Реакции типа г имеют важное значение в энергетическом метаболизме аэробных клеток. Так, например, первой окислительной стадией при -окислении жирных кислот (гл. 9, разд. А,1) является а,р-дегидрирование ацил-СоА-производных жирных кислот. Аналогичной реакцией, протекающей в цикле трикарбоновых кислот, является дегидрирование сукцината в фумарат [c.258]
К основным питательным веществам, используемым микроорганизмами в качестве исходного сырья для биосинтеза, следует отнести углерод, азот и фосфор. При аэробном культивировании микроорганизмов в энергетическом метаболизме клетки непосредственное участие принимает кислород, выполняя роль акцептора электронов. С участием молекулярного кислорода происходит окисление углеводородного субстрата с последовательным образованием надвинного спирта, а затем жирной кислоты. При анаэробном процессе микроорганизмы получают энергию в результате окисления, когда акцепторами электронов выступают неорганические соединения. У фототрофов (фотосинтезирующих бактерий, водорослей) в качестве источника энергии служит энергия солнечной радиации. [c.10]
С участием молекулярного кислорода происходит окисление углеводородного субстрата с последовательным образованием надвинного спирта, а затем жирной кислоты. При анаэробном процессе микроорганизмы получают энергию в результате окисления, когда акцепторами электронов выступают неорганические соединения. У фототрофов (фотосинтезирующих бактерий, водорослей) в качестве источника энергии служит энергия солнечной радиации. [c.10]
Резкое возрастание масштабов взаимодействия прокариот с О2 при функционировании метаболизма аэробного типа делает неэффективными неферментативные пути устранения Н2О2. Для разложения перекиси водорода, образующейся в больших количествах, необходимы специальные ферменты, повышающие скорость разложения Н2О2 на несколько порядков. Это обеспечивается каталазой и пероксидазой. Таким образом, в условиях активного взаимодействия клеток с О2, делающего возможным аэробную жизнь, система ферментной защиты от его токсических эффектов сформирована с участием супероксидцисмутазы, каталазы и пероксидазы в качестве необходимых компонентов (рис. 87). [c.337]
87). [c.337]
Аэротаксис. У подвижных бактерий можно определить тип метаболизма (аэробный или анаэробный) по их аэротаксическим движениям и скоплению на определенных расстояниях от края покровного стекла. В слое бактерий, помещенных между предметным и покровным стеклами, аэрофильные бактерии скапливаются у края покровного стекла или в непосредственной близости от оказавшихся в препарате пузырьков воздуха это указывает на их потребность в аэробных условиях и на то, что необходимую энергию они получают за счет дыхания (рис, 2.40). Строго анаэробные бактерии будут скапливаться в центре. Микроаэрофильные бактерии, например некоторые псевдомонады и спириллы, будут держаться на определенном расстоянии от края. С помощью бактерий, проявляющих положительный аэротаксис, Энгельману удалось продемонстрировать выделение кислорода локально освещаемыми хлоропластами зеленой во-. доросли 8р1годуга. [c.69]
Осн. пути метаболизма D-Г. 1) гликолиз и аэробное окисление до Oj и HjO, в результате к-рых образуются АТФ и др. макроэргич. соединения 2) синтез олиго- и полисахаридов 3) превращение в пентозы и др. простые сахара в пентозофосфатном цикле. О биосинтезе D-Г. см. Глюконеогенез. [c.589]
макроэргич. соединения 2) синтез олиго- и полисахаридов 3) превращение в пентозы и др. простые сахара в пентозофосфатном цикле. О биосинтезе D-Г. см. Глюконеогенез. [c.589]
Состав и соотношение форм И. (спектр И.) изменяется в зависимости от их локализации в органах и тканях организмов одного вида и даже в разных субклеточных органеллах одной и той же клетки. На спектр И. оказывает влияние разное физиол. состояние организма и патологич. процессы, происходящие в нем. Поскольку И. различаются по свои.м св-вам (оптимуму pH, активации ионами, по сродству к субстратам, ингибиторам, активаторам, кофакторам), то характер их распределения отражает регуляторные механизмы, контролирующие метаболизм. Так, напр., лактатдегидрогеназа представлена в организме человека и животных пятью формами, каждая из к-рых представляет собой тетрамер, состоящий из субъединиц двух типов (а и Р) в разных соотношениях. В сердце и печени представлена в осн. форма 04, а в мышцах-Р . Первая ингибируется избытком пировиноградной к-ты и поэтому преобладает в органах с аэробным типом метаболизма, вторая не ингибируется избытком этой к-ты и преобладает в мышцах с высоким урювнем гликолиза. О важной роли И. в тонкой регуляции метаболич. процессов свидетельствует также изменение их спектра под влиянием разл. воздействий и физиол. состояний (охлаждение, гипоксия, денервация и др.). [c.202]
О важной роли И. в тонкой регуляции метаболич. процессов свидетельствует также изменение их спектра под влиянием разл. воздействий и физиол. состояний (охлаждение, гипоксия, денервация и др.). [c.202]
Скорость окислительных стадий цикла определяется скоростью реокисления NADH в цепи переноса электронов. При некоторых условиях ее может лимитировать скорость поступления Ог. Однако в аэробных организмах она обычно определяется концентрацией ADP и (или) Р , доступных для превращения в АТР в процессе окислительного фосфорилирования (гл. 10). Если в ходе катаболизма образуется больше АТР, чем это необходимо для энергетических потребностей клетки, концентрация ADP падает до низкого уровня, выключая, таким образом, процесс фосфорилирования. Одновременно АТР, присутствующий в высоких концентрациях, действуя по принципу обратной связи, ингибирует процессы катаболизма углеводов и жиров. Это ингибирование осуществляется во многих пунктах метаболизма, часть которых показана на рис 9-3. Важным участком, на котором осуществляется такое ингибирование, является пируватдегидрогеназный комплекс (гл 8, разд К2) [19]. Другим таким участком сложит цитратсинтетаза— фермент, катализирующий первую реакцию цикла трикарбоновых кислот [20]. Правда, существуют сомнения относительно того, имеет ли такое ингибирование физиологическое значение [16]. Уровень фосфорилирования аденилатной системы может регулировать работу цикла еще и другим способом, связанным с потребностью в GDP на стадии е цикла (рис. 9-2). В митохондриях GTP в основном используется для превращения АМР в ADP. Следовательно, образование GDP зависит от АМР — соединения, которое образуется в митохондриях при использовании АТР для активации жирных кислот [уравнение (9-1)]. [c.324]
Важным участком, на котором осуществляется такое ингибирование, является пируватдегидрогеназный комплекс (гл 8, разд К2) [19]. Другим таким участком сложит цитратсинтетаза— фермент, катализирующий первую реакцию цикла трикарбоновых кислот [20]. Правда, существуют сомнения относительно того, имеет ли такое ингибирование физиологическое значение [16]. Уровень фосфорилирования аденилатной системы может регулировать работу цикла еще и другим способом, связанным с потребностью в GDP на стадии е цикла (рис. 9-2). В митохондриях GTP в основном используется для превращения АМР в ADP. Следовательно, образование GDP зависит от АМР — соединения, которое образуется в митохондриях при использовании АТР для активации жирных кислот [уравнение (9-1)]. [c.324]
Восстановление диоксиацетонфосфата в глицерофосфат происходит также в летательных мышцах насекомых по-видимому, оно представляет путь, альтернативный образованию в этих тканях молочной кислоты. Хотя превращение свободной глюкозы в глицерофосфат и пируват не дает в итоге прироста АТР, следует учесть, что в мышцах исходным материалом служит гликоген, который по сравнению со свободной глюкозой требует для затравочных реакций вдвое меньше АТР. Кроме того, дисмутация триозофосфата, приводящая к образованию глицерофосфата и пирувата, может обеспечить быструю наработку АТР при интенсивных сокращениях мощной летательной мышцы насекомого. Во время более медленной восстанпвительной фазы глицерофосфат, как полагают, снова окисляется, поступая в митохондрии этих в высокой степени аэробных клеток. Таким образом, транспортировка глицерофосфата в митохондрии служит средством доставки в митохондрии восстановительных эквивалентов, полученных от NADH. Возможно поэтому, что значение глицерофосфата для мышечного метаболизма связано в основном с его транспортной функцией, а не с участием в бысТ» ром образовании АТР. [c.349]
Кроме того, дисмутация триозофосфата, приводящая к образованию глицерофосфата и пирувата, может обеспечить быструю наработку АТР при интенсивных сокращениях мощной летательной мышцы насекомого. Во время более медленной восстанпвительной фазы глицерофосфат, как полагают, снова окисляется, поступая в митохондрии этих в высокой степени аэробных клеток. Таким образом, транспортировка глицерофосфата в митохондрии служит средством доставки в митохондрии восстановительных эквивалентов, полученных от NADH. Возможно поэтому, что значение глицерофосфата для мышечного метаболизма связано в основном с его транспортной функцией, а не с участием в бысТ» ром образовании АТР. [c.349]
Действие ПДГК очень важно в метаболизме углеводов на стадии перехода от анаэробного обмена к аэробному. В этом случае образование ацетил-КоА способствует синтезу лимонной кислоты и началу цикла Кребса. [c.38]
Для реализации биосинтеза и метаболизма необходима энергия, запасаемая в клетках в химической форме, главным образом в экзергонических третьей и второй фосфатной связи АТФ. Соответственно метаболические биоэнергетические процессы имеют своим результатом зарядку аккумулятора — синтез АТФ из АДФ и неорганического фосфата. Это происходит в процессах дыхания и фотосинтеза. Современные организмы несут память об эволюции, начавшейся около 3,5 10 лет назад. Имеются веские основания считать, что жизнь на Земле возникла в отсутствие свободного кислорода (см. 17.2). Метаболические процессы, протекающие при участии кислорода (прежде всего окислительное фосфорилирование при дыхании), относительно немногочисленны и эволюционно являются более поздними, чем анаэробные процессы. В отсутствие кислорода невозможно полное сгорание (окисление) органических молекул пищевых веществ. Тем не менее, как это показывают свойства ныне существующих анаэробных клеток, и в них необходимая для жизни энергия получается в ходе окислительно-восстановительных процессов. В аэробных системах конечным акцептором (т. е. окислителем) водорода служит Ог, в анаэробных — другие вещества. Окисление без Oj реализуется в двух путях брожения — в гликолизе и в спиртовом брожении.
Соответственно метаболические биоэнергетические процессы имеют своим результатом зарядку аккумулятора — синтез АТФ из АДФ и неорганического фосфата. Это происходит в процессах дыхания и фотосинтеза. Современные организмы несут память об эволюции, начавшейся около 3,5 10 лет назад. Имеются веские основания считать, что жизнь на Земле возникла в отсутствие свободного кислорода (см. 17.2). Метаболические процессы, протекающие при участии кислорода (прежде всего окислительное фосфорилирование при дыхании), относительно немногочисленны и эволюционно являются более поздними, чем анаэробные процессы. В отсутствие кислорода невозможно полное сгорание (окисление) органических молекул пищевых веществ. Тем не менее, как это показывают свойства ныне существующих анаэробных клеток, и в них необходимая для жизни энергия получается в ходе окислительно-восстановительных процессов. В аэробных системах конечным акцептором (т. е. окислителем) водорода служит Ог, в анаэробных — другие вещества. Окисление без Oj реализуется в двух путях брожения — в гликолизе и в спиртовом брожении. Гликолиз состоит в многостадийном расщеплении гексоз (например, глюкозы) вплоть до двух молекул пирувата (пировиноградной кислоты), содержащих по три атома углерода. На этом, пути две молекулы НАД восстанавливаются до НАД.Н и две молекулы АДФ фосфоршгируются— получаются две молекулы АТФ. Вследствие обратной реакции [c.52]
Гликолиз состоит в многостадийном расщеплении гексоз (например, глюкозы) вплоть до двух молекул пирувата (пировиноградной кислоты), содержащих по три атома углерода. На этом, пути две молекулы НАД восстанавливаются до НАД.Н и две молекулы АДФ фосфоршгируются— получаются две молекулы АТФ. Вследствие обратной реакции [c.52]
Местообитанием некоторых штаммов грамотрицательных облигатных аэробных бактерий Vitreos illa являются сильно обедненные кислородом непроточные водоемы. Чтобы получать нужное количество кислорода для роста и метаболизма, они синтезируют гемоглобиноподобное вещество, связывающее кислород окружающей среды и увеличивающее концентрацию доступного кислорода в клетке. Когда ген, кодирующий этот белок, был введен в клетки Е. соИ, в последних сразу произошли серьезные изменения повысился уровень синтеза клеточных и рекомбинантных белков, возросла эффективность протонных насосов, увеличилось количество образующегося АТР и его концентрация, особенно при низком содержании кислорода в среде. Чтобы такую стратегию можно было ис- [c.122]
Чтобы такую стратегию можно было ис- [c.122]
Эти микроорганизмы в анаэробных и аноксических условиях потребляют ацетат и пропионат и накапливают их внутриклеточпо в виде полимерных насыщенных оксикислот (ПНО) (параллельно в клетках происходит потребление гликогена). Процесс накопления обеспечивается энергией, выделяющейся при разложении полифосфатов, в результате чего образуется ортофосфат. При pH выше 8,0-8,5 образующийся фосфат может осаждаться в виде солей кальция, алюминия или других металлов. В аэробных условиях эти организмы будут расти, потребляя фосфат, накапливая его в виде полифосфатов и возобновляя таким образом запас гликогена. Основным источником энергии, необходимой для осуществления этого процесса, является окисление ПНО. Рассматриваемые микроорганизмы также способны окислять и другие органические субстраты, доступные в аэробных условиях. Рис. 3.15 иллюстрирует в упрощенном виде метаболизм ФАО. [c.137]
обмен аэробный — это.
 .. Что такое обмен аэробный?
.. Что такое обмен аэробный?- обмен аэробный
- разновидность энергетического О., при котором конечным акцептором электронов в системе окислительно-восстановительных реакций является кислород.
Большой медицинский словарь. 2000.
- обмен анаэробный
- обмен белковый
Смотреть что такое «обмен аэробный» в других словарях:
ОБМЕН ВЕЩЕСТВ — метаболизм, совокупность протекающих в живых организмах химич. превращений, обеспечивающих их рост, жизнедеятельность, воспроизведение, постоянный контакт и обмен с окружающей средой. Благодаря О. в. происходит расщепление и синтез молекул,… … Биологический энциклопедический словарь
Анаэробные организмы — Аэробные и анаэробные бактерии предварительно идентифицируются в жидкой питательной среде по градиенту концентрации O2: 1.
 Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать… … Википедия
Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать… … ВикипедияМЕТАБОЛИЗМ — или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате … Энциклопедия Кольера
Углеводы — Структурная формула лактозы содержащегося в молоке дисахарида Углеводы (сахара, сахариды) органические вещества, содержащие карбонильную гру … Википедия
Трутовые грибы — Трутовые грибы в подавляющем большинстве разрушители древесины, причем во многих случаях именно они оказываются первопричиной поражения и последующей гибели живых деревьев. Вегетативное тело гриба (мицелий) развивается и функционирует в… … Биологическая энциклопедия
Гликолиз — I Гликолиз (греч.
glykys сладкий + lysis разрушение, распад) ферментативный процесс анаэробного негидролитического расщепления углеводов (главным образом глюкозы) в клетках человека и животных, сопровождающийся синтезом аденозинтрифосфорной… … Медицинская энциклопедияИшемия — I Ишемия (ischaemia, греч. ischō задерживать + haima кровь) уменьшение кровоснабжения участка тела, органа или ткани вследствие ослабления или прекращения притока к нему артериальной крови. Ишемия может быть обусловлена Ангиоспазмом, Тромбозом,… … Медицинская энциклопедия
кислород — а; м. Химический элемент (O), газ без цвета и запаха, входящий в состав воздуха, необходимый для дыхания и горения и образующий в соединении с водородом воду. ◊ Перекрыть кислород кому л. Создать невыносимые условия жизни, работы. ◁ Кислородный,… … Энциклопедический словарь
Halomonas titanicae — ? Halomonas titanicae Научная классификация Царство: Бактерии Тип: Протеобактерии Класс: Гамма протеобактерии … Википедия
Аэробное упражнение — Необходимо проверить качество перевода и привести статью в соответствие со стилистическими правилами Википедии.
Вы можете помочь … Википедия
 Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать… … Википедия
Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать… … Википедия glykys сладкий + lysis разрушение, распад) ферментативный процесс анаэробного негидролитического расщепления углеводов (главным образом глюкозы) в клетках человека и животных, сопровождающийся синтезом аденозинтрифосфорной… … Медицинская энциклопедия
glykys сладкий + lysis разрушение, распад) ферментативный процесс анаэробного негидролитического расщепления углеводов (главным образом глюкозы) в клетках человека и животных, сопровождающийся синтезом аденозинтрифосфорной… … Медицинская энциклопедия Вы можете помочь … Википедия
Вы можете помочь … ВикипедияОграничить, чтобы победить. Как на самом деле работает мельдоний и почему его не стоило запрещать
— Что вообще такое допинг? Мне кажется, спортсмены и далекие от спорта люди понимают под этим термином разные вещи.— В какой-то момент спортивные чиновники решили выделить группу лекарственных препаратов (спортсмены тоже люди и иногда принимают лекарства), которые кардинальным образом улучшают возможности человека, воздействуя на конкретные звенья метаболизма, на мышечную массу, то есть на то, что позволяет добиться лучших результатов в спорте. Такие препараты назвали допингом, потому что с этической точки зрения не очень правильно, находясь под их влиянием, участвовать в состязаниях.
— А эти препараты используются в обычной жизни?
— Да, большая часть препаратов, относящихся к допингу, либо их аналоги используются в медицине и, собственно, именно из медицины и приходят в спорт.
— Если такие препараты нормально применять в медицине, почему этого нельзя делать спортсменам?
— В медицине они применяются для лечения того, что в организме «сломалось». Хорошо, если больной человек, используя препараты, улучшит свое состояние хотя бы до того уровня, на котором здоровый спортсмен находится изначально. Кроме того, лекарства могут быть небезопасны для здоровых людей, потому что чем сильнее лекарственный препарат, тем больше у него побочных эффектов, и длительное применение таких препаратов может приводить к кардинальным изменениям в организме спортсмена.
Хорошо, если больной человек, используя препараты, улучшит свое состояние хотя бы до того уровня, на котором здоровый спортсмен находится изначально. Кроме того, лекарства могут быть небезопасны для здоровых людей, потому что чем сильнее лекарственный препарат, тем больше у него побочных эффектов, и длительное применение таких препаратов может приводить к кардинальным изменениям в организме спортсмена.
— То есть большинство допингов — это лекарства, которые просто используются не по прямому назначению?
— Есть еще препараты в серой зоне. Они не признаны допингом, но трудно представить, что они могут использоваться для лечения каких-либо заболеваний. При этом такие вещества могут повлиять на спортивный результат. Среди этих препаратов выделяется группа метаболических препаратов — средств, которые воздействуют на те или иные звенья метаболизма клеток, то есть они воздействуют на механизмы, при помощи которых клетка получает энергию.
Мария Шарапова, пожалуй, самая известная российская спортсменка, в пробах которой обнаружен мельдоний. Фото: Бобылев Сергей/ТАСС
Фото: Бобылев Сергей/ТАССТрадиционно эти средства очень широко использовались в российском спорте, причем зачастую вообще без всякой системы, просто по принципу «нальем как можно больше всего разного». Эти вещества не были запрещены, применялись в очень высоких дозах в различных видах спорта, но по непонятным причинам два таких препарата попали в начале 2016 года в стоп-лист WADA. Один из этих препаратов — милдронат (он же мельдоний). Почему именно эти лекарства попали в список запрещенных, непонятно: они не слабее и не сильнее других метаболических препаратов, и оба действуют довольно странным образом.
— Как вообще работают метаболические препараты?
— Если очень сильно упростить (заранее прошу прощения у биохимиков), то можно объяснить их действие так. Наши клетки могут получать энергию, используя один из двух способов метаболизма — аэробный (кислородный) или анаэробный (бескислородный). Анаэробный метаболизм — это, в первую очередь, гликолиз, благодаря которому организм получает энергию за счет «расщепления» глюкозы без использования кислорода. И при анаэробных нагрузках, то есть коротких и сверхинтенсивных, результат достигается во многом за счет энергии, которую мышечные клетки получают в процессе гликолиза.
И при анаэробных нагрузках, то есть коротких и сверхинтенсивных, результат достигается во многом за счет энергии, которую мышечные клетки получают в процессе гликолиза.
Возьмем крайний вариант анаэробных нагрузок, скажем поднятие штанги, когда нужно за короткое время расщепить огромное количество энергии. Это осуществляется за счет гликолиза, потому что кислородное регулирование просто не успевает подключиться. А при активностях вроде длительной игры в футбол или бега энергия в первую очередь производится за счет кислородного способа — окислительного фосфорилирования.
При сверхинтенсивных краткосрочных нагрузках организм не может полноценно использовать наиболее эффективный кислородный путь метаболизма. Фото: Алеев Егор/ТАСС— Какой способ эффективнее?
— В клетках энергия запасается в виде химических связей в молекуле под названием АТФ, и количество молекул АТФ, которое синтезируется за счет гликолиза, значительно меньше, чем количество молекул АТФ, которое синтезируется за счет окислительного фосфорилирования. И главная концепция, такой мейнстрим спортивной фармакологии — сделать так, чтобы как можно более продуктивно задействовать именно окислительное фосфорилирование, то есть кислородную составляющую.
И главная концепция, такой мейнстрим спортивной фармакологии — сделать так, чтобы как можно более продуктивно задействовать именно окислительное фосфорилирование, то есть кислородную составляющую.
Приведу аналогию. Представьте, что у вас есть две электростанции — одна угольная (гликолиз), другая атомная (окислительное фосфорилирование). Угольную можно быстро запустить и остановить, а атомную включать долго, но она дает больше энергии.
— Есть какие-то препараты, которые могут «подстегнуть» кислородный метаболизм?
— Есть, более того, они разрешены и льются рекой, их перед соревнованиями получают и наши спортсмены, и зарубежные. Классический пример метаболической терапии — «обработка» бегунов на короткие дистанции. Им прямо в кровь вводится фосфокреатин: он помогает очень быстро насытить пул АТФ, который находится рядом с миофибриллами, и в итоге можно получить невероятный результат. Нередко эти вещества дают спортсменам и между тренировками, «на всякий случай», но смысла в этом нет никакого, улучшение результатов будет, только если принимать его непосредственно перед соревнованиями.
Предполагается, что есть и второй путь улучшить спортивные результаты: наоборот, взять и переключить все процессы с кислородных на бескислородные, они же анаэробные. Именно так работает мельдоний. Он перекрывает поступление в митохондрии жирных кислот. То есть выступает таким логистом, который говорит: «Мы лучше повезем топливо не на атомную, а на угольную электростанцию, так как она очень быстро включается». Считается, что в этом случае при коротких анаэробных нагрузках вроде поднятия штанги будет синтезироваться больше энергии за счет гликолиза. Но уверенности в этом нет, так как серьезных исследований мельдония не публиковалось.
— Если я правильно помню, создатель препарат объяснял, что у него иное действие…
— Создатель препарата объяснял, что мельдоний — это не допинг, а препарат, который предотвратит повреждение тканей сердца на фоне избыточных нагрузок. То есть это нечто вроде ограничителя скорости в машине: двигатель позволяет идти со скоростью 260 км/ч, но ограничитель не дает развить больше 200 км/ч.
Так что мельдоний, закрывая вход жирных кислот в наши энергетические станции — митохондрии, по сути ограничивает возможности спортсменов, у которых высокие аэробные нагрузки, например теннисистов, бегунов на длинные дистанции, футболистов.
Хотя в этих видах спорта на пиковых нагрузках «подключается» анаэробный путь получения энергии, определяющую роль играет именно «мощность» аэробного обмена.
— Зачем создавать препарат, который снижает физические возможности?
— Мельдоний разрабатывался для людей с ишемическими повреждениями сердца. При ишемии кислорода по определению мало, поэтому выгодней использовать гликолиз. Да, энергии при этом будет не хватать, но организм хотя бы останется жив. Кроме того, при переходе на бескислородный метаболизм будет генерироваться меньше свободных радикалов, которые повреждают клетки. В этом смысле идея препарата достаточно грамотная: перекрыть ишемизированной клетке возможность использовать «дорогой» аэробный путь метаболизма и жить, пусть и плохо, за счет более «дешевого» анаэробного. Такая же ситуация — недостаток кислорода — наблюдается при тяжелых анаэробных нагрузках, так что спортсменам его дают, видимо исходя из предположения, что мельдоний должен помочь улучшить результат, закрыв менее выгодный в такой ситуации кислородный путь.
Такая же ситуация — недостаток кислорода — наблюдается при тяжелых анаэробных нагрузках, так что спортсменам его дают, видимо исходя из предположения, что мельдоний должен помочь улучшить результат, закрыв менее выгодный в такой ситуации кислородный путь.
Хотя я не встречал ни одного качественного исследования, где было бы показано, что препараты типа мельдония действительно могут помочь, потому что человек — очень сложная система, и даже если мы видим какой-то эффект в культуре клеток, не факт, что он будет таким же у пациента. Да и в любом случае, даже при недостатке кислорода, при ишемии, действие препарата довольно слабое: это все равно что, скажем, машину, которую заправляли 98-м бензином, начнут заправлять 95-м. Разница есть, но небольшая.
— Если препарат настолько слабый, почему WADA включила его в список запрещенных?
— У меня нет разумного объяснения, почему это произошло. Препарат действительно слабый, многие незапрещенные вещества оказывают на спортивные результаты заметно больший эффект.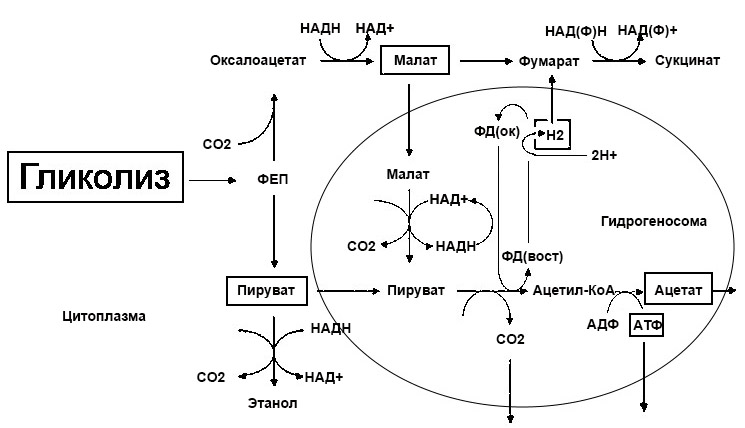 Кроме того, препарат вполне безопасен, и я не встречал серьезных побочных эффектов от его применения.
Кроме того, препарат вполне безопасен, и я не встречал серьезных побочных эффектов от его применения.
Много всего
— Есть еще какие-то разрешенные препараты, которые спортсмены принимают, чтобы улучшить результат?
— Препаратов, которые принимают спортсмены, очень много. Помимо метаболических препаратов есть и неметаболические, например те же антидепрессанты и ноотропы. Подтвержденных клинических данных, как именно они влияют на здоровых людей, тем более спортсменов, нет, но могу предположить, что правильно подобранный коктейль ноотропов вполне может обеспечить лучший результат в тех видах спорта, где нужна концентрация внимания, например в стрельбе. При этом ноотропы и многие антидепрессанты не запрещены WADA.
— Как антидепрессанты могут повлиять на спортивный результат? Веселые спортсмены быстрее добегут до финиша?
— Антидепрессанты, влияя на метаболизм такого нейромедиатора, как серотонин, могут улучшать синаптическую передачу (скорость обмена данными между нейронами — прим. «Чердака»), то есть потенциально увеличивать скорость реакции. Но, повторюсь, влияние ноотропов и антидепрессантов на здоровых людей — совершенно неизученная сфера.
«Чердака»), то есть потенциально увеличивать скорость реакции. Но, повторюсь, влияние ноотропов и антидепрессантов на здоровых людей — совершенно неизученная сфера.
— Есть данные, что спортсмены активно употребляют эти препараты?
— Да, их применяют в достаточно больших количествах, и большой вопрос, употребляет ли их спортсмен из-за наличия депрессии или потому, что хочет улучшить концентрацию внимания.
— Откуда спортивные врачи берут все эти вещества для спортсменов?
— К сожалению, в России подавляющее большинство лекарственных препаратов, включая очень серьезные средства, до сих пор можно купить без рецепта, например, в интернет-аптеках. Есть разные недобросовестные конторы, которые занимаются натуральным шаманством: подбирают спортсменам какие-то препараты, которые якобы хорошо влияют на что-то, при этом зачастую такие люди не знают даже основ фармакологии.
— Откуда они берут информацию?
— Есть вал советских работ, очень странных, выполненных без соблюдения протоколов исследования, которые проводились на советских спортсменах начиная с 70-х годов и даже раньше. Есть практика, которая передается из уст в уста. Некоторые самые продвинутые даже читают какие-то клинические статьи, но все равно все, что они делают, — это такой «самиздат» на коленке.
— Насколько близко современные спортсмены подошли к пределу своих физических возможностей? Реально без допинга добиваться таких результатов или «чистые» спортсмены попросту не смогут побить те рекорды, которые уже установлены?
Я думаю, что скорее второе. Мы достигли предела физических возможностей, и крайне маловероятно, что возможно улучшить эти результаты без препаратов поддержки, к которым относится и допинг, и различные дизайнерские лекарства, и лекарства на стадии клинических испытаний.
— То есть, если сейчас полностью запретить все допинги и препараты поддержки, зрелищный спорт закончится?
— Да, именно так. Я вижу три пути развития спорта. Первый — разработать список лекарств, которые можно применять. В этот «белый список» можно включить не только антибиотики и средства для лечения бронхиальной астмы, но и те же метаболические препараты и еще какие-то средства поддержки, при этом строго запретить применять все остальное. Любые подозрительные новые препараты тоже записывать в список запрещенных. Вторая стратегия — наоборот, разрешить спортсменам принимать абсолютно любые вещества. И третий путь — оставить все как есть. Второй вариант, конечно, самый зрелищный, но и самый негуманный. Нередко у профессиональных спортсменов, которые принимают всевозможные средства, к 40 годам развивается сердечная недостаточность: у этих еще совсем молодых людей сердечная мышца бывает в крайне тяжелом состоянии.
Я вижу три пути развития спорта. Первый — разработать список лекарств, которые можно применять. В этот «белый список» можно включить не только антибиотики и средства для лечения бронхиальной астмы, но и те же метаболические препараты и еще какие-то средства поддержки, при этом строго запретить применять все остальное. Любые подозрительные новые препараты тоже записывать в список запрещенных. Вторая стратегия — наоборот, разрешить спортсменам принимать абсолютно любые вещества. И третий путь — оставить все как есть. Второй вариант, конечно, самый зрелищный, но и самый негуманный. Нередко у профессиональных спортсменов, которые принимают всевозможные средства, к 40 годам развивается сердечная недостаточность: у этих еще совсем молодых людей сердечная мышца бывает в крайне тяжелом состоянии.
— Из этих трех стратегий та, что есть сейчас, выглядит самой слабой. Учитывая современные возможности химии и молекулярной биологии, она больше похожа на игру в поддавки…
— Да, выбирая этот путь, мы, возможно, наносим куда больший вред спортсменам. Сейчас можно чуть ли не в полуподвальных условиях синтезировать весьма «продвинутые» вещества, и такие препараты в неумелых руках могут заметно ухудшить результат, не говоря уж об общем уровне здоровья спортсменов.
Сейчас можно чуть ли не в полуподвальных условиях синтезировать весьма «продвинутые» вещества, и такие препараты в неумелых руках могут заметно ухудшить результат, не говоря уж об общем уровне здоровья спортсменов.
Ярослав Ашихмин, кандидат медицинских наук, руководитель отделения терапии Юсуповской больницы и заместитель генерального директора по медицине. Выпускник ММА имени И.М. Сеченова, автор более 75 научных публикаций, член Американской ассоциации сердца (AHA) и Европейского общества кардиологов (ESC)
Ирина Якутенко
РАЗНИЦА МЕЖДУ АЭРОБНЫМ И АНАЭРОБНЫМ МЕТАБОЛИЗМОМ | СРАВНИТЕ РАЗНИЦУ МЕЖДУ ПОХОЖИМИ ТЕРМИНАМИ — НАУКА
Аэробный против анаэробного метаболизма Клеточный метаболизм — это процесс преобразования углеводов, жиров и белков в энергию, необходимую клеткам. Во время метаболизма клетки энергия накапливается
Аэробный против анаэробного метаболизма
Клеточный метаболизм — это процесс преобразования углеводов, жиров и белков в энергию, необходимую клеткам. Во время метаболизма клетки энергия накапливается в высокоэнергетических фосфатных связях молекул аденозинтрифосфата (АТФ), которые служат энергетической валютой клеток. В зависимости от потребности в кислороде во время производства АТФ в клетке присутствуют два основных типа метаболизма; а именно аэробный и анаэробный. Из трех основных метаболических путей только гликолиз считается анаэробным метаболизмом, тогда как остальные, включая цикл лимонной кислоты (цикл Кребса) и цепь переноса электронов, рассматриваются как аэробный метаболизм.
Во время метаболизма клетки энергия накапливается в высокоэнергетических фосфатных связях молекул аденозинтрифосфата (АТФ), которые служат энергетической валютой клеток. В зависимости от потребности в кислороде во время производства АТФ в клетке присутствуют два основных типа метаболизма; а именно аэробный и анаэробный. Из трех основных метаболических путей только гликолиз считается анаэробным метаболизмом, тогда как остальные, включая цикл лимонной кислоты (цикл Кребса) и цепь переноса электронов, рассматриваются как аэробный метаболизм.
Аэробный метаболизм
Аэробный метаболизм происходит при наличии кислорода. Он происходит в митохондриях клетки и отвечает за обеспечение 90% потребности организма в энергии. Во время аэробного метаболизма весь основной субстрат, включая углеводы, жиры и белки, расщепляется и объединяется с молекулярным кислородом для производства энергии, выделяя в качестве конечных продуктов диоксид углерода и воду. В общем, окислительный метаболизм производит от 150 до 300 мл воды за 24 часа. В аэробном метаболизме участвуют два пути; цикл лимонной кислоты; который происходит в матрице митохондрий и цепи переноса электронов; который происходит в системе транспорта электронов, расположенной во внутренней мембране митохондрий.
В аэробном метаболизме участвуют два пути; цикл лимонной кислоты; который происходит в матрице митохондрий и цепи переноса электронов; который происходит в системе транспорта электронов, расположенной во внутренней мембране митохондрий.
Анаэробный метаболизм
Анаэробный метаболизм не требует кислорода для производства АТФ. Это происходит в результате гликолиза — процесса высвобождения энергии из глюкозы. Эффективность анаэробного метаболизма низкая, и количество АТФ вырабатывается низким по сравнению с аэробным метаболизмом. Гликолиз происходит в цитоплазме и не требует никаких органелл. Следовательно, это важный процесс, в котором у организмов отсутствуют митохондрии, такие как прокариоты. Конечным продуктом аэробного метаболизма является молочная кислота, которая может быть относительно вредной для организма.
Аэробный против анаэробного метаболизма
• Аэробный метаболизм требует кислорода, тогда как анаэробный метаболизм — нет.
• Анаэробный метаболизм не может продолжаться бесконечно. Напротив, аэробный метаболизм может продолжаться вечно, только в теоретических условиях.
• Углеводы, жиры и белки используются как источники аэробного метаболизма, тогда как только углеводы участвуют в анаэробном метаболизме.
• Аэробный метаболизм включает в себя деятельность от низкой до умеренной интенсивности, тогда как анаэробный метаболизм включает только деятельность с высокой интенсивностью.
• Анаэробный метаболизм происходит в цитоплазме клеток, а аэробный метаболизм — в митохондриях.
• Аэробный метаболизм производит больше энергии, чем анаэробный метаболизм, если такое же количество того же субстрата.
• Гликолиз — это анаэробный метаболический путь, тогда как цикл лимонной кислоты и цепь переноса электронов — аэробные метаболические пути.
• Аэробный метаболизм вносит больший вклад (около 90%) в снабжение энергией, в то время как анаэробный метаболизм вносит меньший вклад.
• Конечным продуктом анаэробного метаболизма является молочная кислота, тогда как продуктом аэробного метаболизма является углекислый газ и вода.
Источник изображения: любезно предоставлено http://webanatomy.net/anatomy/aerobic.webp
Публикации в СМИ
ЗАЩИТА МИОКАРДА (кардиоплегия)
Кардиоплегия — метод защиты миокарда от повреждения во время выполнения манипуляций на остановленном сердце.
• Патофизиологические этапы ишемии миокарда •• Латентный период (аэробный метаболизм за счёт запасов кислорода в виде оксигемоглобина в крови коронарных сосудов) продолжается в течение 1–15 с •• Период выживания (анаэробный гликолиз происходит при тканевом pО2 менее 5 мм рт.ст. и продолжается при +28 °С в течение 5–6 мин) •• Период обратимого повреждения (глубокие ультраструктурные изменения, после реперфузии наблюдают период оглушения от нескольких минут до нескольких часов) •• Некробиоз (при нормотермии развивается в течение 15 мин) •• Некроз (при нормотермии развивается в течение 20 мин).
• Факторы защиты миокарда •• Гипотермия (обычная температура кардиоплегического р-ра +4 °С) •• Высокая концентрация внеклеточных калия и магния (инактивация быстрых и медленных натриевых и кальциевых каналов) •• Применение -адреноблокаторов, местных анестетиков, антагонистов кальция (используют редко по причине трудной управляемости эффекта) •• Использование энергетических субстратов.
• Требования к кардиоплегии: •• Молниеносная остановка сердца •• Предупреждение повреждения коронарных артерий и миокарда самим кардиоплегическим р-ром •• Предупреждение реперфузионного синдрома •• Создание физиологических условий в период восстановления сердечной деятельности.
• Методики проведения кардиоплегии: кровяная непрерывная, кровяная прерывистая, кристаллоидная непрерывная, кристаллоидная прерывистая.
• Техника введения кардиоплегического р-ра •• Антеградное введение в корень аорты •• Антеградное введение в устья коронарных артерий (применяют во время операций на левых камерах сердца, митральном и аортальном клапанах, а также на восходящей части аорты) •• Ретроградное введение в коронарный синус (наиболее универсальная и адекватная, хотя и сложная техника) •• Введение кардиоплегического р-ра и локальная гипотермия следуют тотчас после наложения зажима на аорту.
• Восстановление сердечной деятельности обычно происходит через фибрилляцию желудочков, но иногда собственный ритм может восстанавливаться спонтанно. В любом случае этот факт не имеет прогностической значимости. Как правило, согревание занимает около 10 мин (следует ориентироваться на температуру в прямой кишке), после чего при стабильной гемодинамике снимают зажим с аорты.
Сокращения • ИК — искусственное кровообращение • ОСП — объёмная скорость перфузии
Приложение № 5. Тактика в гребном спорте.
‹‹‹ назад
Для оценки действий гребца при прохождении дистанции целесообразно рассмотреть физиологические закономерности энергообеспечения от старта до финиша.
На дистанции 2000 метров гребцы преодолевают три фазы: стартовую, середину дистанции и финишную фазу.
Со
старта гребцы начинают гонку преимущественно с более высоким темпом
гребли, чем в середине дистанции, и скорость хода более высока, чем средняя
скорость на всей дистанции.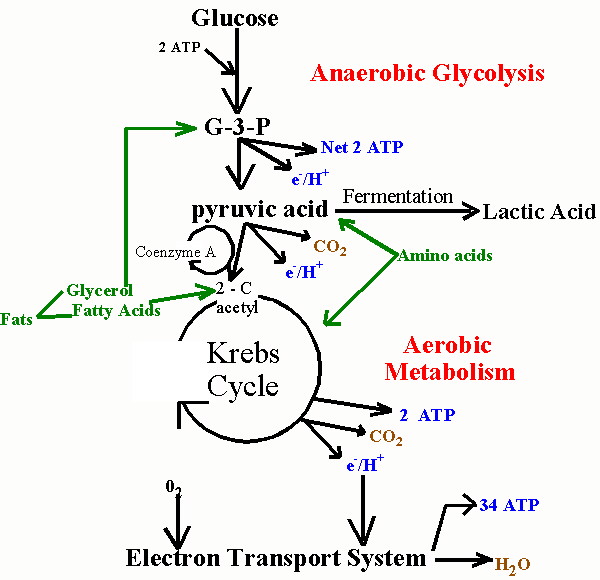 Энергия здесь извлекается из мышечных клеток в
результате распада химических связей, накопленного топливного ресурса
организма. Но мышечные клетки поставляют энергию в условиях недостаточного
количества кислорода анаэробным процессом энергообеспечения. В результате
этого процесса образуется побочный продукт – молочная кислота, большое
содержание которой вызывает болевые ощущения в мышцах спортсмена.
Энергия здесь извлекается из мышечных клеток в
результате распада химических связей, накопленного топливного ресурса
организма. Но мышечные клетки поставляют энергию в условиях недостаточного
количества кислорода анаэробным процессом энергообеспечения. В результате
этого процесса образуется побочный продукт – молочная кислота, большое
содержание которой вызывает болевые ощущения в мышцах спортсмена.
На середине дистанции спортсмен использует энергию, полученную в результате преобразования топливного ресурса организма, происходящего в присутствии кислорода – аэробный процесс. Фаза середины дистанции длятся около 4-6 минут до наступления финишной фазы. Аэробные процессы почти в 18 раз продуктивнее анаэробных и не производят ненужной молочной кислоты, ослабляющей организм. Тем не менее, анаэробный обмен веществ кратковременно обеспечивает большую мощь и позволяет поддержать высокую скорость мышечных сокращений. Как и в стартовой фазе, гребцы увеличивают темп гребли на финише, стремясь ускорить ход лодки в течение одной-двух последних минут гонки. Все это требует дополнительных энергозатрат, превышающих возможности аэробного обмена веществ, поэтому подключаются анаэробные процессы с выделением все увеличивающегося количества молочной кислоты. Человеческое тело работает как некоторый двигатель, и развивает мощность, зависящую от прилагаемого усилия, что обеспечивает перемещение лодки с определенной скоростью.
Источником энергии для мышечного сокращения является расщепление химических соединений, находящихся в клетках мышцы.
1. АТФ (аденозинтрифосфат)
2. КФ (креатинфосфат)
3. Глюкоза, содержащаяся в качестве гликогена
4. Жиры.
АТФ — это единственное соединение, которое может напрямую сообщить энергию мышечному сокращению. Так как содержащейся в мышцах АТФ достаточно для мышечных сокращений только в течение нескольких секунд, то необходимо воспроизводство АТФ. Другие соединения являются не прямыми источниками энергии, так как предоставляют энергию для ресинтеза или воспроизврлства АТФ, которое включает в себя 3 процесса:
1. АТФ/КФ реакция
2. Анаэробный гликолиз
3. Аэробный метаболизм
1. АТФ/КФ реакция.
Хранящееся в мышечных клетках КФ-соединение, сходное с АТФ. Оно может стать источником для быстрого ресинтеза АТФ, однако его количества хватит не более, чем на 20 сек. работы. Так как этот процесс происходит при отсутствии кислорода и не вызывает появление молочной кислоты, его называют алактатным анаэробным.
Несмотря на то, что КФ обеспечит гребца энергией на старте, его вклад в общую энергию, необходимую для прохождения дистанции 2000м, чрезвычайно мал.
2. Анаэробный гликолиз.
Это производство энергии с отсутствием кислорода с появлением молочной кислоты, определяется как лактатный анаэробный процесс. Это важный источник энергии на стартовом и финишном отрезках гонки.
Результатом этого процесса является производство энергии для ресинтеза АТФ посредством расщепления корбогидратов (преимущественно гликогена, содержащегося в мышечных клетках — поэтому он называется анаэробным гликолизом). Он может обеспечить энергией так же быстро, как и АТФ/КФ реакция.
Этот процесс может обеспечить значительным объемом энергии, однако расщепление гликогена и накопление молочной кислоты в мышечных клетках снижают способность мышцы к сокращению, вызывая боль в ней, что является препятствием для длительного использования этого источника энергии. Поэтому он используется преимущественно на старте и финише гонки.
Несмотря на то, что названный процесс может обеспечивать гребца энергией для двух-трех минут интенсивной работы (30-90 секунд после старта и 60-90 секунд на финише), в целом он дает только 20-25% всей необходимой для прохождения дистанции энергии.
3. Аэробный метаболизм.
Этот процесс обеспечивает около 75-80% всей необходимой для прохождения дистанции энергии. Он требует «сгорания топлива» в мышечных клетках в присутствии кислорода. Источником его является либо гликоген, либо жиры, хранящиеся в мышцах, или глюкоза и жиры, хранящиеся в других депо организма и перемещаемые мышечными клетками по кровеносным сосудам.
Так как этот процесс включает в себя большее число реакций в мышечных клетках, то он происходит значительно медленнее и требует присутствия кислорода. Отсюда совершенно ясно, какую важную роль играет транспорт кислорода в ткани и его утилизация. В продвижении кислорода воздуха к тканям участвуют три системы.
Система дыхания передает воздух в легкие. В легких кислород диффундирует из воздуха, проникая через стенки альвеол в кровь.
Система кровообращения передает кровь, насыщенную кислородом, из легких к сердцу, которое перекачивает ее по артериям к органам, наиболее нуждающимся в кислороде крови, а именно, работающим мышцам. По мере передвижения крови по артериям, последние разветвляются до капилляров, заключающих в себе отдельные мышечные волокна.
Третья система — мышечная, принимает кислород по мере того, как он проникает через стенки капилляров в мышечные клетки. Здесь кислород попадает в митохондрию (энерговырабатывающую станцию мышечных клеток) и утилизуется, производя энергию за счет переработки топливного ресурса организма.
Необходимо отметить, что аэробный процесс включает два основных:
-расщепление липидов (жиров)
-аэробный гликолиз (расщепление гликогена)
Так как расщепление жиров дает большое количество энергии, он является важным источником при тренировке, но так как его реакции очень медленны, он почти не успевает развернуться при прохождении дистанции 2000 м. Для такой дистанции характерен аэробный гликолиз и полное расщепление гликогена.
Взаимодействие этих процессов энергообеспечения на дистанции 2000 м следующее:
1. АТФ/КФ реакция — менее 5%
2. Анаэробный гликолиз — 20-25%
3. Аэробный процесс — 75-80%
Эти три процесса действуют не изолированно друг от друга во время прохождения дистанции, а наслаиваются друг на друга, обеспечивая необходимый уровень энергии на дистанции.
Более точно определить относительный вклад каждого из процессов трудно, но максимальное потребление кислорода гребцом указывает на наиболее высокий уровень аэробного обмена веществ. Это обстоятельство очень важно, так как именно аэробный метаболизм преобладает при прохождении дистанции в академической гребле.
Аэробный метаболизм — обзор
Метаболизм
Существует два основных метаболических пути использования Glc. Основным путем является гликолитический путь, который идет через пируват и ацетил-КоА в цикл Кребса или питает различные синтетические пути через его промежуточные метаболиты. Полное окисление Glc по этому пути дает десять НАДН, два восстановленных убихинона и четыре АТФ / ГТФ. Если кислорода недостаточно (анаэробные условия) для утилизации NADH, метаболизм Glc может быть прекращен на уровне лактата без чистой продукции NADH.Распад Glc посредством гликолиза и цикла Кребса является основой мышц и большинства других клеток. Пентозофосфатный цикл (гексозо-монофосфатный шунт) удаляет один углерод из Glc с каждым циклом. Этот путь особенно важен для быстрорастущих клеток, поскольку он генерирует два НАДФН (используемых для многих синтетических путей) в каждом цикле и обеспечивает рибозу для синтеза ДНК и РНК. Эритроциты также в значительной степени зависят от пентозо-фосфатного цикла в своем топливном метаболизме.
Гликолиз: Начальное фосфорилирование глюкозы катализируется гексокиназой (HK; EC2.7.1.1) на внешней мембране митохондрий. Четыре гена кодируют HK, которые присутствуют на разных уровнях в большинстве тканей. Альтернативный сплайсинг HK 4 (глюкокиназы) дает две изоформы, специфичные для печени, и специфичную для поджелудочной железы. Альтернативные промоторы избирательно реагируют на инсулин (изоформы печени) или глюкозу (изоформы поджелудочной железы). Большое количество генов и изоформ и их различные характеристики соответствуют разнообразным потребностям различных тканей, которые могут быть удовлетворены с помощью точно подобранной смеси.Продукт, глюкозо-6-фосфат, аллостерически ингибирует все эти формы. Альтернативой фосфорилированию Glc в печени является неклассическая функция фермента цинка глюкозо-6-фосфатазы (EC3.1.3.9). Фактически это многокомпонентный комплекс, внедренный в мембрану эндоплазматического ретикулума, который включает как комплексные каталитические активности, так и по крайней мере четыре различных свойства транспорта субстрата. Глюкозо-6-фосфатаза может использовать как карбамилфосфат, так и пирофосфат в качестве доноров фосфата (Nordlie et al., 1999).
Следующие шаги зависят от глюкозо-6-фосфат-изомеразы (EC5.3.1.9) и фосфофруктокиназы-1 (EC2.7.1.11). Фосфофруктокиназа активируется описанным выше регуляторным метаболитом фруктозо-2,6-бисфосфата. Полученный фруктозо-1,6-бисфосфат расщепляется на трехуглеродные молекулы фруктозобисфосфатальдолазой (альдолаза; EC4.1.2.13), ключевым регуляторным ферментом гликолиза, который активируется АМФ, АДФ и бисфосфатом фруктозы и ингибируется. последующими продуктами цитрат и АТФ.Три разных гена кодируют основные формы последнего в мышцах (альдолаза A), печени (альдолаза B) и головном мозге (альдолаза C), а дополнительные изоформы генерируются путем альтернативного сплайсинга.
Триозоизомераза (EC5.3.1.1) превращает дигидроксиацетонфосфат в глицеральдегид-3-фосфат в почти равновесной реакции. Глицеральдегид-3-фосфатдегидрогеназа / фосфорилирование (GAPDH; EC1.2.1.12): окислительная реакция существует в мышечной и печеночной формах, кодируемых разными генами.Метаболизм в пируват продолжается с фосфоглицераткиназой (EC2.7.2.3, повсеместная и специфическая для семенников формы), фосфоглицератмутазой (EC5.4.2.1, три разных изофермента для мышц, эритроцитов и других тканей), фосфопируватгидратазой (энолаза; EC4. 2.1.11, магнийзависимый, множественные изоферменты, кодируемые по крайней мере четырьмя генами) и пируваткиназа (EC2.7.1.40, множественные изоферменты из-за трех генов и альтернативного сплайсинга).
Анаэробный метаболизм: Способность продуцировать АТФ, скорее всего, будет ограничена доступностью кислорода для окислительного фосфорилирования, чем доступностью окисляемого субстрата.Это типично для интенсивных краткосрочных упражнений. Мышцы могут метаболизировать анаэробно, но с гораздо меньшим выходом энергии, чем при аэробном метаболизме. В этом случае пируват восстанавливается до лактата L-лактатдегидрогеназой (EC1.1.1.27), обеспечивая возобновление поступления окисленного НАД для продолжения гликолиза. Две молекулы АТФ и два лактата могут быть произведены анаэробно из одной молекулы глюкозы. Протоны, возникающие в результате производства лактата, увеличивают внутриклеточную кислотность и помогают вытеснить избыток лактата через котранспортер 1 протонов / монокарбоновых кислот (MCT1, SLC16A1).Лактат легко поглощается печенью, используется для синтеза Glc и снова возвращается в кровообращение и мышцы по мере необходимости. Это перемещение лактата и Glc между мышцами и печенью (цикл Кори) позволяет отдельным мышцам продолжать работать без бремени метаболических обязательств, связанных с предыдущими тренировками.
Аэробный метаболизм: Пируват транспортируется через внутреннюю митохондриальную мембрану пируваттранслоказой, где находятся ферменты окислительного метаболизма.При достаточном поступлении кислорода пируват метаболизируется комплексом пируватдегидрогеназы (EC1.2.4.1) до ацетил-КоА в реакции, продуцирующей НАДН. Многосубъединичный ферментный комплекс требует тиаминпирофосфата (связанного с субъединицами E1), липоата (связанного с субъединицами E2, дигидролипоамид S-ацетилтрансферазы; EC2.3.1.12) и FAD (связанного с субъединицами E3, дигидролипоамиддегидрогеназы; EC1.8.1 .4). Фосфорилирование серинов в субъединице E1 [пируватдегидрогеназой (липоамид)] киназой (EC2.7.1.99) инактивирует ферментный комплекс. Дефосфорилирование [пируватдегидрогеназа (липоамид)] -фосфатаза (EC3.1.3.43) снова активирует его.
Рисунок 7.4. Гликолиз включает в себя эти начальные анаэробные этапы метаболизма глюкозы
Пентозофосфатный путь: Этот альтернативный путь метаболизма Glc особенно важен для быстро делящихся тканей, поскольку он генерирует рибозо-5-фосфат, который является предшественником сахара для синтеза ДНК и РНК. и НАДФН, который используется многими путями биосинтеза.НАДФН необходим для восстановления окисленного глутатиона в эритроцитах. Снижение доступности НАДФН (типично при дефиците глюкозо-6-фосфатдегидрогеназы) увеличивает уязвимость эритроцитов к окислительному стрессу и склонность к гемолизу. Прием пиримидин-агликона дивицина с бобами Vicia fava (или воздействие их пыльцы) у людей с дефицитом глюкозо-6-фосфатдегидрогеназы вызывает окислительную модификацию гемоглобина и может вызвать у них острый гемотоксический кризис (McMillan et al., 2001).
Рисунок 7.5. Аэробная часть метаболизма глюкозы начинается с переноса пирувата в митохондрии
После фосфорилирования Glc гексокиназой (EC2.7.1.1), как описано выше, последовательные действия глюкозо-6-фосфатдегидрогеназы (EC1.1.1.49), глюконолактоназа (EC3.1.1.17) и 6-фосфоглюконатдегидрогеназа (EC1.1.1.44) генерируют пентозерибулозо-5-фосфат. Первая и третья реакции генерируют НАДФН.
Транскетолаза (EC2.2.1.1) с ковалентно связанным тиаминпирофосфатом катализирует две реакции перегруппировки. Один из них превращает два пентозофосфата (X5P и рибозо-5-фосфат) в набор соединений с семью (D-седогептулоза-7-фосфат) и тремя (глицеральдегид-3-фосфат, GAP) атомами углерода. Другой преобразует X5P плюс эритрозо-4-фосфат в глицеральдегид-3-фосфат и фруктозо-6-фосфат (F6P). Третья возможность — это перегруппировка двух молекул X5P в две молекулы GAP и одну эритрулозу (Быкова и др., 2001). Те же реакции катализируются транскетолазой 2 (Coy et al. , 1996) с различными изоформами в головном мозге и сердце, генерируемыми альтернативным сплайсингом. Варианты гена транскетолазы 2 могут быть вовлечены в патогенез синдрома Вернике-Корсакова. Трансальдолаза (EC2.2.1.2) дополняет катализируемые транскетолазой реакции перегруппировки, превращая соединения с семью и тремя атомами углерода в эритрозо-4-фосфат (4 атома углерода) и F6P (6 атомов углерода). Две дополнительные стадии, катализируемые глюкозо-6-фосфат-изомеразой (EC5.3.1.9) и глюкозо-6-фосфатаза (EC3.1.3.9) могут затем генерировать глюкозу. Альтернативно, в зависимости от статуса питания, 6-фосфофруктокиназа (фосфофруктокиназа I; EC2.7.1.11) может инициировать утилизацию посредством гликолиза.
Рисунок 7.6. Пентозофосфатный путь является основным источником НАДФН
Аэробный метаболизм против анаэробного метаболизма »Наука ABC
Метаболизм — это все химические реакции, которые происходят внутри тела. Аэробный метаболизм — это когда организм вырабатывает энергию (в форме АТФ) с помощью кислорода.Анаробный метаболизм — это когда организм вырабатывает энергию без кислорода. Аэробный метаболизм более эффективен в создании энергии, чем анаэробный метаболизм.
Хотя многие люди этого не осознают, каждый день, когда вы просыпаетесь и переживаете существование, довольно невероятен. Каждое действие нашего тела, от перекачивания крови по нашим органам и вдыхания кислорода до сгибания пальцев и подъема по лестнице, требует энергии. Для каждого живого существа приобретение, преобразование и расход энергии имеют решающее значение для выживания.Производство этой энергии посредством химических процессов в организме известно как клеточный метаболизм.
По сути, метаболизм — это все химические реакции, происходящие внутри тела. Когда мы смотрим на метаболизм на клеточном уровне, мы называем его клеточным метаболизмом. Эти реакции, в основном катализируемые ферментами, можно разделить на множество различных категорий в зависимости от того, в каком процессе они участвуют.
В этой статье мы будем говорить об энергетическом или топливном метаболизме, который представляет собой те ферментативные реакции, которые производят энергию для организма из пищи, которую мы едим.
Пища, которую мы едим — углеводы, белки и жиры, расщепляется на АТФ (аденозинтрифосфат), энергетическую валюту организма. АТФ — это молекула, которая дает телу энергию для выполнения других процессов, таких как упражнения, чтение, мысли, танцы и, в основном, жизнь.
Этот распад, который приводит к выработке энергии, может происходить двумя путями: аэробным метаболизмом или анаэробным метаболизмом. Эти две стороны метаболической монеты совершенно разные, несмотря на то, что они начинаются с одного и того же сырья, но баланс этих двух процессов необходим для выживания!
Эта товарная позиция может немного вводить в заблуждение, предполагая, что эти две формы метаболизма противоположны, хотя на самом деле они в определенном смысле тесно связаны и взаимозависимы.Ключевое различие между ними заключается в том, что аэробный метаболизм происходит в присутствии кислорода, а анаэробный — нет.
Хотя аэробный метаболизм является более медленным из двух процессов, он гораздо более распространен и важен для нашего повседневного функционирования. На долю аэробного метаболизма приходится около 90% всего клеточного метаболизма, при котором жиры и углеводы превращаются в полезную энергию в форме АТФ (аденозинтрифосфата). На аэробный метаболизм рассчитывают для производства энергии, которая может поддерживать устойчивые энергетические потребности, например.г., ходьба, бег трусцой, ежедневные нагрузки.
Превращение углеводов в энергию требуется каждый день для удовлетворения всех ваших основных функциональных потребностей. Вот почему в организме постоянно происходит аэробный метаболизм. Аэробный метаболизм также является единственным средством, с помощью которого ваше тело может извлекать энергию из жира. Аминокислоты из белков при расщеплении на топливо, что в основном происходит, когда организму не хватает углеводов или жиров, например, во время голодания или диабета, могут быть преобразованы в глюкозу, гликоген или кетоновые тела посредством процесса, называемого кетозом.
Несмотря на то, что эти два процесса медленнее, аэробный метаболизм очень эффективен и способен выдавливать 34 молекулы АТФ из одной молекулы глюкозы. Дополнительные побочные продукты включают диоксид углерода и воду. Аэробный метаболизм состоит из двух различных путей, цикла Кребса и цепи переноса электронов, которые происходят в митохондриях, энергетических фабриках клеток. Сырье, необходимое для этих двух стадий, включает воду и кислород, но стадия, предшествующая этим аэробным процессам, представляет собой не что иное, как анаэробный гликолиз, который мы объясним ниже.
В отличие от формы, описанной выше, анаэробный метаболизм не требует присутствия кислорода для преобразования сырья в энергию. Однако анаэробный гликолиз намного менее эффективен, производя только две молекулы АТФ, по сравнению с впечатляющими 34 аэробными метаболизмом.
Организм полагается на анаэробное дыхание, когда требуется внезапный прилив энергии за короткий промежуток времени. Например, представьте, что вы спринтер или штангист; ваши физические потребности обычно высоки, но только в течение ограниченного периода времени.Поскольку аэробный метаболизм занимает больше времени, организм использует анаэробный метаболизм для выработки энергии для немедленного использования из углеводов, но не жиров или белков. Начиная с одной молекулы глюкозы, процесс гликолиза разворачивается в цитоплазме клетки и не требует никаких органелл.
Кроме того, при занятиях спортом или тяжелой атлетике организму не хватает кислорода. Сердце качает кровь так быстро, как может, но недостаточно и не вовремя, чтобы удовлетворить потребности мышц или других клеток.
(Фото: YassineMrabet / Wikimedia Commons)
К сожалению, одним из побочных продуктов анаэробного метаболизма является молочная кислота, которая может вызывать усталость. Быстрое накопление молочной кислоты вызывает судороги у спортсменов, которые слишком сильно нагружают себя без должной разминки или когда организм не может сбалансировать аэробный и анаэробный метаболизм.
В организме по умолчанию используется аэробный метаболизм, поскольку он намного эффективнее анаэробного (если вы не анаэробные бактерии, кислород может вас убить).Анаэробное дыхание присутствует в чрезвычайных ситуациях, пока организм не сможет вернуться к своему первоначальному состоянию, зависящему от кислорода.
В конце концов, не такие уж и разные…
Хотя объем и разнообразие конечных продуктов различаются, аэробный и анаэробный метаболизм тесно связаны. Как упоминалось ранее, гликолиз — это первый шаг, ведущий к аэробному дыханию, и поэтому он происходит постоянно, как и аэробный метаболизм. По сути, организму необходимо «разогреться» до аэробного метаболизма жиров, поэтому он начинается с преобразования чистых углеводов в цитоплазме, а затем переходит в аэробный метаболизм.
В большинстве случаев вы не замечаете отрицательных побочных продуктов анаэробного метаболизма, потому что ваше тело может устранить побочные продукты быстрее, чем вы их произведете. Представьте, что вы совершаете долгую неторопливую прогулку по парку. Ваше тело будет подвергаться анаэробному и аэробному метаболизму, но при условии, что вы съели достаточно еды этим утром, двигаетесь в нормальном темпе и не слишком напрягаетесь, вы, вероятно, не вспотеете и не разовьетесь спазмами из-за молочной кислоты. Пока аэробные метаболические пути тела могут идти в ногу с потребностями организма, анаэробный метаболизм может отойти на второй план.Когда энергии, производимой аэробными процессами, недостаточно, анаэробные процессы берут на себя большую роль в удовлетворении спроса.
Статьи по теме
Статьи по теме
Эти два процесса явно взаимосвязаны и одинаково важны. Без аэробного дыхания нам не хватало бы постоянных источников энергии, необходимых для ходьбы, дыхания, работы, речи и вождения автомобиля. Без анаэробного дыхания наша способность действовать, например, во время сценария борьбы или бегства, была бы серьезно нарушена.В общем, мы должны быть бесконечно благодарны обеим сторонам метаболического пути и умному пути эволюции, который позволяет нам жить!
16.4: Источники топлива — Medicine LibreTexts
Цели обучения
- Узнайте о различных источниках топлива в организме человека
Человеческое тело использует углеводы, жиры и белки, содержащиеся в пище и из запасов тела, в качестве энергии. Эти важные питательные вещества необходимы независимо от интенсивности вашей активности.Если вы лежите, читаете книгу или бегаете марафон в Гонолулу, эти макроэлементы всегда необходимы вашему организму. Однако для того, чтобы эти питательные вещества использовались в качестве топлива для организма, их энергия должна быть передана в высокоэнергетическую молекулу, известную как аденозинтрифосфат (АТФ). АТФ — это непосредственный источник энергии для организма, который может вырабатываться либо в присутствии кислорода, известном как аэробный метаболизм, либо в его отсутствие в результате анаэробного метаболизма. Тип метаболизма, который преимущественно используется во время физической активности, определяется доступностью кислорода и количеством используемых углеводов, жиров и белков.
Анаэробный и аэробный метаболизм
Анаэробный метаболизм происходит в цитозоле мышечных клеток. Как видно на рисунке \ (\ PageIndex {1} \), небольшое количество АТФ вырабатывается в цитозоле без присутствия кислорода. Анаэробный метаболизм использует глюкозу как единственный источник топлива и производит пируват и молочную кислоту. Затем пируват можно использовать в качестве топлива для аэробного метаболизма. Аэробный метаболизм происходит в митохондриях клетки и может использовать углеводы, белок или жир в качестве источника топлива.Аэробный метаболизм — это гораздо более медленный процесс, чем анаэробный метаболизм, но производит большую часть АТФ.
Рисунок \ (\ PageIndex {1} \): анаэробный и аэробный метаболизм. Изображение Эллисон Калабрезе / CC BY 4.0.Продолжительность физической активности и расход топлива
Дыхательная система играет жизненно важную роль в поглощении и доставке кислорода мышечным клеткам по всему телу. Кислород вдыхается легкими и переносится из легких в кровь, где сердечно-сосудистая система направляет богатую кислородом кровь к мышцам.Затем кислород поглощается мышцами и может использоваться для выработки АТФ. Когда тело находится в состоянии покоя, сердце и легкие способны снабжать мышцы достаточным количеством кислорода для удовлетворения энергетических потребностей аэробного метаболизма. Однако во время физической активности потребности ваших мышц в энергии и кислороде увеличиваются. Чтобы обеспечить мышечным клеткам больше кислорода, у вас увеличится частота сердечных сокращений и частота дыхания. Количество кислорода, которое доставляется к тканям через сердечно-сосудистую и дыхательную системы во время упражнений, зависит от продолжительности, интенсивности и физического состояния человека.
Во время первых нескольких шагов упражнения ваши мышцы первыми реагируют на изменение уровня активности. Однако ваши легкие и сердце не реагируют так быстро, и во время этих первых шагов они не начинают увеличивать доставку кислорода. Чтобы наши тела получали энергию, необходимую на этих начальных этапах, мышцы полагаются на небольшое количество АТФ, которое хранится в мышцах в состоянии покоя. Накопленный АТФ способен обеспечить энергию всего за несколько секунд, прежде чем он истощится.Как только накопленный АТФ почти израсходован, организм прибегает к другой высокоэнергетической молекуле, известной как креатинфосфат, для преобразования АДФ (аденозиндифосфата) в АТФ. Примерно через 10 секунд запас креатинфосфата в мышечных клетках также истощается.
Примерно через 15 секунд после начала упражнения запасы АТФ и креатинфосфата расходуются в мышцах. Сердце и легкие все еще не адаптировались к возрастающей потребности в кислороде, поэтому мышцы должны начать производить АТФ за счет анаэробного метаболизма (без кислорода).Анаэробный метаболизм может производить АТФ в быстром темпе, но использует только глюкозу в качестве источника топлива. Глюкоза получается из крови мышечного гликогена. Примерно через 30 секунд анаэробные пути работают на полную мощность, но, поскольку доступность глюкозы ограничена, они не могут продолжаться в течение длительного периода времени.
По мере того, как ваше упражнение длится две-три минуты, частота сердечных сокращений и частота дыхания увеличиваются, чтобы снабдить мышцы кислородом. Аэробный метаболизм — наиболее эффективный способ производства АТФ, поскольку он производит в 18 раз больше АТФ для каждой молекулы глюкозы, чем анаэробный метаболизм.Хотя основным источником АТФ в аэробном метаболизме являются углеводы, жирные кислоты и белок также могут использоваться в качестве топлива для выработки АТФ.
Рисунок \ (\ PageIndex {2} \): Влияние продолжительности упражнений на энергетические системы. Изображение Эллисон Калабрезе / CC BY 4.0.Источники топлива для анаэробного и аэробного метаболизма будут меняться в зависимости от количества доступных питательных веществ и типа метаболизма. Глюкоза может поступать из глюкозы в крови (которая состоит из пищевых углеводов или гликогена печени и синтеза глюкозы) или мышечного гликогена.Глюкоза является основным источником энергии как для анаэробного, так и для аэробного метаболизма. Жирные кислоты хранятся в мышцах в виде триглицеридов, но около 90% запасенной энергии находится в жировой ткани. Поскольку упражнения с низкой и средней интенсивностью продолжаются с использованием аэробного метаболизма, жирные кислоты становятся основным источником топлива для тренируемых мышц. Хотя белок не считается основным источником энергии, небольшое количество аминокислот используется во время отдыха или выполнения какой-либо деятельности. Количество аминокислот, используемых для энергетического обмена, увеличивается, если общее потребление энергии из вашего рациона не соответствует потребностям в питательных веществах или если вы выполняете длительные упражнения на выносливость.Когда аминокислоты расщепляются с удалением азотсодержащей аминокислоты, оставшаяся молекула углерода может быть расщеплена на АТФ посредством аэробного метаболизма или использована для производства глюкозы. Когда упражнения продолжаются в течение многих часов, увеличивается использование аминокислот в качестве источника энергии и для синтеза глюкозы.
Рисунок \ (\ PageIndex {3} \): Источники топлива для анаэробного и аэробного метаболизма.Интенсивность физической активности и расход топлива
Интенсивность упражнений определяет вклад типа источника топлива, используемого для производства АТФ (см. Рисунок \ (\ PageIndex {2} \)).И анаэробный, и аэробный метаболизм сочетаются во время упражнений, чтобы гарантировать, что мышцы оснащены достаточным количеством АТФ для выполнения возложенных на них требований. Величина вклада каждого типа метаболизма будет зависеть от интенсивности деятельности. При выполнении упражнений низкой интенсивности используется аэробный метаболизм для обеспечения мышц достаточным количеством АТФ. Однако во время высокоинтенсивных занятий требуется больше АТФ, поэтому мышцы должны полагаться как на анаэробный, так и на аэробный метаболизм, чтобы удовлетворить потребности организма.
Во время низкоинтенсивных занятий организм использует аэробный метаболизм, а не анаэробный, потому что он более эффективен, производя большее количество АТФ. Жирные кислоты являются основным источником энергии при малоинтенсивных занятиях. При практически неограниченных запасах жира в организме малоинтенсивные занятия могут продолжаться долгое время. Наряду с жирными кислотами используется небольшое количество глюкозы. Глюкоза отличается от жирных кислот, в которых запасы гликогена могут истощаться.Когда запасы гликогена истощаются, рано или поздно наступает усталость.
Рисунок \ (\ PageIndex {4} \): Влияние интенсивности упражнений на источники топлива.Зона сжигания жира
Зона сжигания жира — это аэробная активность низкой интенсивности, при которой частота пульса составляет от 60 до 69% от максимальной частоты пульса. С другой стороны, кардиозона — это аэробная активность высокой интенсивности, которая поддерживает частоту сердечных сокращений примерно от 70 до 85% от вашей максимальной частоты сердечных сокращений. Так в какой зоне вы сжигаете больше всего жира? Технически ваше тело сжигает больший процент калорий из жира во время аэробной активности низкой интенсивности, но это еще не все.Когда вы начинаете упражнения с низкой интенсивностью, около 50% сжигаемых калорий приходится на жир, тогда как в кардиозоне только 40% приходится на жир. Однако если посмотреть на фактическое количество сожженных калорий, то при более высокой интенсивности сжигается столько же жира и гораздо больше калорий в целом.
Рисунок \ (\ PageIndex {5} \): Зона сжигания жира. Изображение Эллисон Калабрезе / CC BY 4.0.«Удар о стену» или «Удар по стене»
Если вы знакомы с видами спорта на выносливость, возможно, вы слышали о «ударе в стену» или «ударе».Эти разговорные термины относятся к крайней усталости, которая наступает примерно через 120 минут занятий спортом на выносливость, например, марафонским бегом или велоспортом на длинные дистанции. Физиология, лежащая в основе «удара о стену», означает, что мышцы израсходовали весь свой запасенный гликоген и, следовательно, зависят от других питательных веществ для удовлетворения своих энергетических потребностей. Жирные кислоты транспортируются от жировых клеток к мышцам, чтобы восполнить дефицит питательных веществ. Однако жирным кислотам требуется больше времени для преобразования в энергию, чем глюкозе, что снижает уровень производительности.Чтобы избежать «ударов о стену» или «ударов», спортсмены на выносливость загружаются углеводами за несколько дней до соревнований, что называется углеводной загрузкой. Это позволит максимально увеличить запасы гликогена в печени и мышцах спортсмена. Важно не думать, что углеводная загрузка работает для всех. Без сопутствующих тренировок на выносливость вы не увеличите запасы глюкозы. Если вы планируете пробежать пятимильную гонку для развлечения со своим другом и решите съесть большое количество углеводов в виде большого ужина из спагетти накануне вечером, избыток углеводов будет сохранен в виде жира.Следовательно, если вы не являетесь атлетом на выносливость, тренирующимся более 90 минут, углеводная загрузка не принесет пользы, а может даже иметь некоторые недостатки. Еще один способ для спортсменов избежать «удара о стену» — это употребление углеводосодержащих напитков и продуктов во время соревнований на выносливость. Фактически, на протяжении «Тур де Франс» — гонки на двадцать два дня и двадцать четыреста миль — средний велосипедист потребляет более 60 граммов углеводов в час.
Авторы и авторство
границ | Поглощение Ca2 + митохондриями и тонкая настройка аэробного метаболизма
Введение
Внутриклеточный Ca 2+ играет важную роль в качестве вторичного посредника, контролирующего как повсеместные, так и тканеспецифические процессы.Приманка различных триггеров и четко определенные пространственно-временные паттерны ответа Ca 2+ на различные стимулы гарантируют специфичность клеточного ответа (Berridge, 2001).
Внешнее перемещение протонов через внутреннюю митохондриальную мембрану (IMM), определяемое активностью цепи переноса электронов (ETC), генерирует потенциал митохондриальной мембраны (Δψ), который внутри является отрицательным. Приток Ca 2+ в активированные митохондрии зависит от электрохимического градиента (Deluca, Engstrom, 1961; Vasington, Murphy, 1962) и обеспечивается активностью Mitochondrial Calcium Uniporter (MCU), высокоселективного канала IMM. (Киричок и др., 2004; Baughman et al., 2011; Де Стефани и др., 2011). Микродомены с высокой концентрацией Ca 2+ в местах непосредственной близости между ER / SR и митохондриями, так называемые митохондриально-ассоциированные мембраны (MAM), образуются при высвобождении Ca 2+ из ER / SR ( эндоплазматический / саркоплазматический ретикулум), обеспечивая быстрое проникновение в митохондрии Ca 2+ (Rizzuto et al., 1993, 1998).
В физиологическом состоянии митохондриальный матрикс Ca 2+ регулирует активность дегидрогеназ цикла TCA, тем самым запуская генерацию восстанавливающих эквивалентов, которые питают ETC и, в конечном итоге, продукцию ATP.Напротив, в патологических условиях перегрузка митохондрий Ca 2+ вызывает открытие поры перехода проницаемости (PTP), что приводит к быстрому коллапсу Δψ и набуханию митохондрий вместе с высвобождением цитохрома с и про апоптотические факторы (рисунок 1) (Briston et al., 2017; Giorgio et al., 2018).
Рисунок 1. Физиопатологическая роль митохондриального поглощения Ca 2+ . В физиологических условиях поглощение митохондриями Ca 2+ стимулирует выработку АТФ благодаря положительной регуляции трех Ca 2+ -зависимых ферментов цикла TCA (пируватдегидрогеназа, изоцитратдегидрогеназа и оксоглутаратдегидрогеназа).В цитоплазме гликолитические реакции превращают глюкозу в пируват. Митохондриальный переносчик пирувата (MPC) определяет проникновение пирувата в митохондрии, где пируватдегидрогеназа (PDH) окисляет пируват до ацетил-КоА. Ацетил-КоА входит в цикл TCA, что приводит к образованию восстанавливающих эквивалентов, которые питают цепь переноса электронов и, в конечном итоге, производство АТФ. Напротив, в патологических условиях перегрузка митохондрий Ca 2+ может вызвать открытие митохондриальной переходной поры проницаемости (mPTP), что в конечном итоге приведет к гибели клеток.
В этом обзоре мы обсудим роль митохондриальной передачи сигналов Ca 2+ в контроле клеточного метаболизма, уделяя особое внимание органам, участвующим в регуляции системного метаболизма, включая печень, скелетные мышцы и жировую ткань. Мы также рассмотрим роль поглощения митохондриальным Ca 2+ в контроле функции сердца, органа, в котором этот аспект широко исследовался. Мы особенно обсудим исследования, основанные на генетической модуляции активности MCU, как в клеточных линиях, так и в моделях на животных.
Молекулярная идентичность и регуляция комплекса MCU
Порообразующие субъединицы
На сегодняшний день три различных белка митохондриальной мембраны были охарактеризованы как компоненты унипортера Ca 2+ , то есть MCU, MCUb и эссенциальный регулятор MCU (EMRE).
Митохондриальный кальций Uniporter
Все эукариоты, за исключением дрожжей, демонстрируют хорошо законсервированный ген MCU (первоначально известный как CCDC109a ), который кодирует белок 40 кДа, состоящий из двух доменов спиральной спирали и двух трансмембранных доменов, последние разделены коротким, но высоко расположенным консервативная петля, обращенная к межмембранному пространству (IMS).Эта петля обогащена кислотными остатками, необходимыми для придания селективности Ca 2+ . Было предсказано, что MCU будет олигомеризоваться с целью формирования функционального канала, и эта гипотеза была подтверждена разделением на геле нативных митохондриальных белков, которое выдвинуло на первый план MCU-содержащий комплекс с кажущейся молекулярной массой 450 кДа (Baughman et al., 2011; De Стефани и др., 2011). Хотя с помощью молекулярно-динамического подхода было предсказано, что MCU образуют тетрамеры (Raffaello et al., 2013), первая предложенная структура MCU Caenorhabditis elegans с помощью методов ЯМР и крио-ЭМ предусматривала пентамерную сборку (Oxenoid et al., 2016). Однако в последнее время сообщалось о крио-ЭМ и / или рентгеновских структурах для микроконтроллеров грибов либо отдельно (Baradaran et al., 2018; Fan et al., 2018; Nguyen et al., 2018; Yoo et al. , 2018) или в присутствии EMRE (Wang et al., 2019). Эти исследования продемонстрировали, что комплексы микроконтроллеров грибов характеризуются тетрамерной архитектурой. Тетрамерная сборка консервативна также в MCU рыбок данио, которые, обладая более чем 90% гомологией с MCU человека, являются убедительным доказательством в пользу этой архитектуры у высших эукариот (Baradaran et al., 2018).
Митохондриальный сайленсинг унипортера кальция приводит к прекращению захвата митохондриальным Ca 2+ , в то время как его сверхэкспрессия вызывает как значительное увеличение митохондриальных транзиентов Ca 2+ в интактных клетках, так и увеличение тока Ca 2+ в митопластах. (Де Стефани и др., 2011; Чаудхури и др., 2013). Рекомбинантных MCU достаточно для формирования Ca 2+ -селективного канала в плоском липидном бислое per se (De Stefani et al., 2011), с током, сходным, хотя и не идентичным, с тем, который был зарегистрирован в экспериментах с патч-кламп изолированными митопластами (Kirichok et al., 2004). Общая активность комплекса MCU сильно варьируется среди тканей (Fieni et al., 2012). Эта особенность, вероятно, объединяет различные факторы: объем, занимаемый митохондриями в конкретном типе клеток (Fieni et al., 2012), относительные уровни экспрессии MCU по сравнению с другими сложными компонентами и / или регуляторами, включая MCUb (Raffaello et al., 2013) и MICU1 (Paillard et al., 2017) и, возможно, степенью сайтов контакта между митохондриями и ER / SR.
MCUb
ГенMCUb ( CCDC109b ) законсервирован у большинства позвоночных, хотя он отсутствует у некоторых организмов, у которых присутствует MCU (например, у растений, нематод и членистоногих). Белок MCUb с молекулярной массой 40 кДа имеет 50% гомологию последовательности с MCU, и он аналогичным образом организован: он имеет два домена со спиральной спиралью и два трансмембранных домена, разделенных петлей.Важная аминокислотная замена (E256V) в области петли, нейтрализующая отрицательный заряд, ответственна за резкое снижение проводимости канала по сравнению с MCU (Raffaello et al., 2013). Сверхэкспрессия MCUb снижает транзиенты митохондриального Ca 2+ при стимуляции агонистом в клетках, и при вставке в плоский липидный бислой ток не регистрируется. Напротив, подавление MCUb вызывает значительное увеличение поглощения митохондриальным Ca 2+ (Raffaello et al., 2013). Это связано с тем, что MCUb образует гетероолигомеры с MCU, отрицательно влияя на проникновение Ca 2+ через канал.
Что касается физиологической значимости MCUb, следует отметить, что соотношение MCU / MCUb сильно варьируется в разных тканях. Например, он выше в скелетных мышцах по сравнению с сердцем (Raffaello et al., 2013), что согласуется с различной активностью MCU в разных тканях (Fieni et al., 2012).
EMRE
EMRE представляет собой белок 10 кДа, расположенный в IMM, состоящий из трансмембранного домена, короткого N-концевого домена и высококонсервативного С-конца, обогащенного кислотными остатками (Sancak et al., 2013). Хотя в плоском липидном бислое MCU per se достаточно для генерации тока, эксперименты, проведенные на нокаутных клетках EMRE, демонстрируют, что этот небольшой белок важен для активности MCU. В клетках EMRE, как полагают, модулирует взаимодействие между областью поры и регуляторными субъединицами канала (Sancak et al., 2013). Однако в плоском липидном бислое MCU и его регуляторные субъединицы MICU1 и MICU2 обладают способностью взаимодействовать друг с другом без присутствия EMRE (Patron et al., 2014). Тем не менее, гетерологичное восстановление MCU человека в дрожжах требует EMRE, указывая тем самым, что EMRE необходим для сборки функционального канала у многоклеточных животных (Kovács-Bogdán et al., 2014). Кроме того, было продемонстрировано, что С-конец EMRE обращен к митохондриальному матриксу и образует Ca 2+ -зависимый комплекс с MICU1 и MICU2. Таким образом, EMRE будет действовать как датчик для [Ca 2+ ] по обе стороны от IMM (Vais et al., 2016). Однако в качестве альтернативы была предложена другая топология EMRE для IMM, несовместимая с ролью С-конца EMRE в качестве датчика матричного Ca 2+ (Yamamoto et al., 2016). Недавно для определения релевантности EMRE in vivo была создана модель мыши EMRE — / — (Liu et al., 2020). Исследования на этих мышах продемонстрировали, что EMRE действительно требуется для активности MCU, поскольку изолированные митохондрии EMRE — / — не накапливают Ca 2+ . Делеция EMRE приводит к почти полной эмбриональной летальности у инбредных мышей C57Bl6 / N. Однако животные EMRE — / — жизнеспособны на смешанном фоне, хотя они меньше и рождаются реже.Не было обнаружено различий в отношении основных метаболических функций у животных гибрида Bl6 / N-CD1 EMRE — / — по сравнению с контрольными животными. Действительно, удаление EMRE не повлияло ни на общее потребление кислорода телом, ни на выработку углекислого газа. Мыши EMRE — / — показали нормальную функцию скелетных мышц с точки зрения мышечной силы и способности бегать. Кроме того, животные с нокаутом демонстрировали нормальную сердечную функцию, включая отсутствие изменений в повреждении IR (ишемия-реперфузия). Таким образом, аналогично модели мыши MCU — / — , созданной на смешанном фоне (Pan et al., 2013; Murphy et al., 2014), адаптации происходят для поддержания митохондриальной активности в отсутствие функционального канала MCU. Наконец, Лю и др. (2016) показали, что в отсутствие экспрессии MICU1 делеция одного аллеля EMRE снижает поглощение митохондриями Ca 2+ , предполагая, что попытки модулировать комплекс MCU могут быть полезными при заболеваниях, характеризующихся перегрузкой Ca 2+ .
Регуляторные подразделения
Одним из важнейших аспектов накопления в митохондриях Ca 2+ является его сигмоидальный ответ на цитоплазматические уровни Ca 2+ .Другими словами, при низком цитозольном [Ca 2+ ] митохондриальное поглощение Ca 2+ незначительно, тогда как оно экспоненциально увеличивается, когда цитозольное [Ca 2+ ] превышает определенный порог (Рисунок 2). Различные уровни активности являются следствием точно настроенной модуляции открытия канала из-за присутствия регуляторных белков, расположенных в IMS, которые взаимодействуют с порообразующими субъединицами. Эти регуляторы принадлежат к семейству MICU, которое включает MICU1, MICU2 и MICU3, каждый из которых характеризуется различными паттернами экспрессии и конкретными функциями.
Рисунок 2. Схема комплекса MCU. Митохондриальный унипортер Ca 2+ (MCU) состоит из порообразующих субъединиц (например, MCU, MCUb и EMRE) и регуляторных субъединиц (например, MICU1, MICU2 и MICU3). MCU и доминантно-отрицательная изоформа MCUb собираются в тетрамерные комплексы, которые охватывают IMM. EMRE является важным регулятором MCU, белком 10 кДа, расположенным в IMM, отсутствие которого вызывает потерю активности унипортерного канала. Особенным аспектом захвата митохондриальным Ca 2+ является его сигмоидальный ответ на внемитохондриальный [Ca 2+ ].Это свойство связано с присутствием в IMS регуляторов MCU, MICU1, MICU2 и, почти исключительно в нервной системе, MICU3. При низком цитозольном уровне [Ca 2+ ] небольшое количество Ca 2+ проникает через канал. После стимуляции клеток увеличение цитозоля [Ca 2+ ] ощущается EF-руками, присутствующими как на MICU1, так и на MICU2. Следовательно, происходят конформационные изменения регуляторных субъединиц, что позволяет открыть канал, ведущий к проникновению Ca 2+ внутрь митохондрий.
Семейство регуляторов микроконтроллеров MICU
MICU1 был первым компонентом комплекса MCU, который был идентифицирован, и он был описан как критический модулятор митохондриального захвата Ca 2+ (Perocchi et al., 2010). MICU1 играет двойную роль в контроле активности MCU. С одной стороны, он держит канал закрытым, когда внемитохондриальная концентрация Ca 2+ ниже определенного порога, тем самым предотвращая непрерывное и устойчивое проникновение в митохондрии Ca 2+ , которое в конечном итоге может вызвать перегрузку Ca 2+ .С другой стороны, когда внемитохондриальная концентрация Ca 2+ превышает определенное значение, MICU1 действует как кооперативный активатор MCU, гарантируя высокую проводимость Ca 2+ (Mallilankaraman et al., 2012b; Csordás et al. , 2013; Patron et al., 2014). Таким образом, MICU1 ведет себя как привратник MCU при низких концентрациях Ca 2+ в цитозоле и обеспечивает эффективное поглощение Ca 2+ митохондриями при увеличении концентрации Ca 2+ в цитозоле (Csordás et al., 2013). Большинство мышей MICU1 — / — умирают перинатально, а те, что выживают, страдают от атаксии и мышечной слабости (Antony et al., 2016; Liu et al., 2016).
Позже были описаны два гена паралога MICU1: MICU2 (ранее назывался EFHA1) и MICU3 (ранее назывался EFHA2), которые, как и MICU1, расположены в IMS (Hung et al., 2014; Lam et al., 2015). В то время как паттерн тканевой экспрессии MICU2 аналогичен MICU1, MICU3 экспрессируется преимущественно в нервной системе (Plovanich et al., 2013; Paillard et al., 2017). MICU2 образует облигатный гетеродимер с MICU1, стабилизированный дисульфидной связью через два консервативных остатка цистеина. Во многих типах клеток стабильность MICU2 зависит от присутствия MICU1, поскольку нокдаун MICU1 вызывает значительное снижение уровней белка MICU2. Об этом сообщалось в клетках HeLa (Patron et al., 2014) и в фибробластах пациентов, несущих гомозиготную делецию MICU1 с потерей функции (Debattisti et al., 2019). Однако в скелетных мышцах MICU1 — / — , а также в скелетных мышцах MICU1 — / — уровни белка MICU2 не были затронуты (Antony et al., 2016; Дебаттисти и др., 2019).
Гетеродимеры MICU1-MICU2 регулируют активность комплекса MCU из-за присутствия доменов EF-hand. Связывание Ca 2+ с этими доменами запускает конформационные модификации, приводящие к открытию канала. Однако сущность сродства Ca 2+ к доменам EF-hand все еще остается открытым вопросом. Измерения, выполненные с помощью калориметрии изотермического титрования, показывают, что Kd взаимодействия MICU1 / Ca 2+ находится в диапазоне от 4 до 40 мкМ (Wang et al., 2014; Vecellio Reane et al., 2016). Однако измерения, основанные на собственной флуоресценции триптофана, зарегистрировали Kd ~ 300 нМ (Kamer et al., 2017). Конечно, разные методы в разных экспериментальных условиях могли дать неоднородные результаты, что указывает на то, что этот вопрос требует дальнейшего прояснения. Что касается роли MICU2, электрофизиологические исследования, проведенные на плоском липидном бислое, продемонстрировали, что MICU2 ингибирует активность MCU при низком уровне цитозоля [Ca 2+ ], таким образом выступая в качестве подлинного привратника канала (Patron et al., 2014).
Наконец, стехиометрия регуляторов MCU внутри комплекса все еще остается в значительной степени неизвестной. Мультимеры MICU1 претерпевают молекулярную перестройку во время стимуляции Ca 2+ (Waldeck-Weiermair et al., 2015). Кроме того, Paillard et al. (2017) продемонстрировали, что соотношение MICU1 / MCU объясняет различные регуляторные свойства комплекса MCU в разных тканях. Например, высокое соотношение MICU1 / MCU, присутствующее в печени, обеспечивает высокую кооперативную активацию канала и одновременно увеличивает порог активации.Соответственно, небольшие изменения транзиентов цитозольного Ca 2+ в гепатоцитах недостаточны для того, чтобы привести к митохондриальному накоплению Ca 2+ , что действительно требует устойчивого увеличения цитозольного Ca 2+ . Другой сценарий происходит в сердце, в котором низкое соотношение MICU1 / MCU позволяет митохондриям поглощать Ca 2+ даже при низком цитозольном уровне [Ca 2+ ]. В этом случае переходные процессы Ca 2+ от биений к ударам декодируются микроконтроллером с низким уровнем стробирования канала и низкой степенью кооперативности, тем самым инициируя интегративное накопление Ca 2+ (Paillard et al., 2017).
Было приложено много усилий для оценки молекулярной динамики комплекса MICU1 – MICU2. В частности, Цзя и соавторы разрешили структуру MICU2 человека, чтобы исследовать взаимодействия между MICU1 и MICU2 как в форме, связанной с аро, так и в форме Ca 2+ . Во взаимодействии MICU1 – MICU2 участвуют различные остатки в зависимости от связывания Ca 2+ с мотивами EF-hands. В апо-форме критическими аминокислотами для образования комплекса являются Glu242 в MICU1 и Arg352 в MICU2, тогда как в связанном с Ca 2+ состоянии Phe383 в MICU1 взаимодействует с Glu196 в MICU2 (Wu et al., 2019). Совсем недавно структуры митохондриального унипортерного голокомплекса кальция человека, полученные методом криоэлектронной микроскопии, были разрешены в состояниях, активированных апо и Ca 2+ . Это исследование продемонстрировало, что один гетеродимер MICU1-MICU2 способен блокировать канал, образованный 4 MCU и 4 субъединицами EMRE. В соответствии с предыдущими отчетами структурный анализ подтвердил существование различных конформаций в соответствии с [Ca 2+ ]. В частности, это исследование демонстрирует, что, в то время как при низком [Ca 2+ ] MICU1 покрывает поры, при высоком [Ca 2+ ] MICU1 перемещается от поверхности MCU к краю тетрамера MCU-EMRE, таким образом позволяя открывать поры (Fan et al., 2020).
Несколько лет назад наша лаборатория охарактеризовала альтернативный вариант сплайсинга MICU1, а именно MICU1.1 (Vecellio Reane et al., 2016). MICU1.1 экспрессируется только в скелетных мышцах на более высоких уровнях по сравнению с MICU1 и в головном мозге. По сравнению с MICU1, MICU1.1 содержит дополнительный экзон, кодирующий четыре аминокислоты (EFWQ). MICU1.1 связывает Ca 2+ более эффективно по сравнению с MICU1, а гетеродимер MICU1.1-MICU2 активирует канал при более низком [Ca 2+ ], чем гетеродимер MICU1-MICU2.Эти особенности особенно важны для скелетных мышц, где MICU1.1 гарантирует устойчивую продукцию АТФ. Однако то, как домен EFWQ влияет на аффинность связывания Ca 2+ и структуру белка, все еще исследуется.
MICU3, как уже упоминалось выше, специфически экспрессируется в головном мозге (Plovanich et al., 2013). MICU3 образует опосредованные дисульфидной связью димеры с MICU1, но не с MICU2, и действует как активатор активности MCU (Patron et al., 2019). Соответственно, подавление MICU3 в первичных кортикальных нейронах нарушает передачу сигналов Ca 2+ , предполагая роль в контроле функции нейронов.Таким образом, нейроны экспрессируют как димеры MICU1 – MICU2, которые гарантируют низкий порочный цикл Ca 2+ в условиях покоя, так и димеры MICU1-MICU3, которые снижают порог открытия MCU, обеспечивая митохондриальный захват Ca 2+ даже в присутствии быстрые и небольшие цитозольные Ca 2+ переходные процессы. Недавно было продемонстрировано, что MICU3 участвует в метаболической гибкости нервных окончаний. MICU3, настраивая чувствительность комплекса MCU к Ca 2+ , позволяет пресинаптическим митохондриям поглощать Ca 2+ в ответ на небольшие изменения в цитозоле [Ca 2+ ] и, таким образом, поддерживать синтез АТФ в аксонах (Ashrafi и другие., 2020).
Другие предполагаемые модуляторы MCU
Другие митохондриальные белки были идентифицированы как предполагаемые модуляторы активности каналов MCU. Вкратце, MCUR1, ранее известный как CCDC90a , был описан как модулятор MCU, поскольку его молчание снижает накопление Ca 2+ в митохондриях и базальный матрикс митохондрий [Ca 2+ ] в клетках HEK293T (Mallilankaraman et al., 2012а). Однако Субридж и др. продемонстрировали, что MCUR1 также играет критическую роль в сборке комплекса V, и его подавление резко снижает потенциал митохондриальной мембраны (Paupe et al., 2015). Таким образом, является ли MCUR1 прямым модулятором MCU или, скорее, он неспецифически притупляет движущую силу для захвата митохондриальным Ca 2+ , все еще обсуждается. Наконец, SLC25A23 является членом семейства переносчиков растворенных веществ, которые транспортируют Mg-ATP / Pi через IMM (Hoffman et al., 2014). Мутации в доменах EF-hand SLC25A23 снижают накопление митохондриального Ca 2+ , что позволяет предположить, что он может играть роль в контроле активности MCU.
Ca
2+ Мишени аэробного метаболизма: регуляция митохондриальных дегидрогеназМитохондриальный Ca 2+ играет различные роли, влияя на энергетический обмен.Он способствует регулированию челночных систем через IMM, которые обеспечивают транспорт нуклеотидов, метаболитов и кофакторов (см. Обзор Rossi et al., 2019). Кроме того, недавно было предложено, что внутри матрицы Ca 2+ непосредственно модулирует активность АТФ-синтазы (Territo et al., 2000) и ETC (Glancy et al., 2013).
Что наиболее важно, митохондриальное поглощение Ca 2+ способствует регуляции производства энергии, влияя на активность митохондриальных дегидрогеназ (рис. 3).Более подробно, FAD-связанная глицеринфосфатдегидрогеназа (GPDH) в IMS и пируватдегидрогеназа (PDH), изоцитратдегидрогеназа (IDH) и оксоглутаратдегидрогеназа (OGDH) в матриксе прямо или косвенно контролируются митохондриальным Ca 2+ ( Дентон, 2009).
Рисунок 3. Ca 2+ -зависимая регуляция митохондриальных дегидрогеназ. (A) Митохондриальный Ca 2+ косвенно регулирует пируватдегидрогеназный комплекс (PDC), который окисляет пируват до ацетил-КоА.PDC состоит из нескольких копий трех компонентов: пируватдекарбоксилазы (E1), дигидролипоатацетилтрансферазы (E2) и дигидролипоатдегидрогеназы (E3). В ответ на увеличение митохондрий [Ca 2+ ], Ca 2+ -зависимая изоформа пируватдегидрогеназной фосфатазы 1 (PDP1) дефосфорилирует E1, тем самым запуская активацию PDC. (B) Ca 2+ непосредственно активирует изоцитратдегидрогеназу (IDH). Отношение АТФ / АДФ отрицательно регулирует чувствительность связывания Ca 2+ с IDH. (C) Оксоглутаратдегидрогеназа (OGDH) представляет собой мультиферментный комплекс, подобный PDC. Он состоит из субъединиц дигидролипоамидсукцинилтрансферазы (E2, зеленый на рисунке), оксоглутаратдекарбоксилазы (E1, розовый на рисунке) и дигидролипоамиддегидрогеназы (E3, желтый на рисунке). Связывание Ca 2+ напрямую регулирует активность фермента.
Глицеринфосфатдегидрогеназа, связанная с FAD (GPDH)
Шаттл глицеринфосфатазы состоит из митохондриальной FAD-связанной глицеринфосфатдегидрогеназы и цитозольной NAD-зависимой глицеринфосфатдегидрогеназы.Этот челнок переносит восстанавливающие эквиваленты от НАДН в цитозоле к ФАДН 2 в митохондриальном матриксе и, в конечном итоге, к ЭТЦ. Связанный с FAD GPDH является трансмембранным белком IMM и обладает сайтами связывания для глицеринфосфата и Ca 2+ , обращенными к IMS, таким образом определяя цитоплазматическую концентрацию обеих этих молекул (Cole et al., 1978; Garrib and McMurray, 1986 ). В частности, два домена EF-hand ответственны за Ca 2+ -зависимую активацию фермента (MacDonald and Brown, 1996).
Пируватдегидрогеназа (ПДГ)
Комплекс пируватдегидрогеназы (PDC) млекопитающих имеет молекулярную массу около 8 МДа и содержит несколько копий трех компонентов, которые катализируют превращение пирувата в ацетил-КоА. Пируватдекарбоксилаза (E1) и дигидролипоатдегидрогеназа (E3) присоединены к центральному ядру, образованному дигидролипоатацетилтрансферазой (E2) (Hiromasa et al., 2004). Пируватдекарбоксилаза представляет собой тетрамер, состоящий из двух различных субъединиц (α2 и β2), катализирующих необратимую стадию реакции.PDC играет решающую роль в контроле клеточного метаболизма и, соответственно, подлежит тонкой регуляции. Ацетил-КоА и НАДН, конечные продукты окислительной реакции, ингибируют активность PDC. Активность PDC также ингибируется обратимым фосфорилированием трех сайтов на субъединице E1 (Denton, 2009), катализируемым пируватдегидрогеназными киназами (PDK) и реверсируемым пируватдегидрогеназными фосфатазами (PDP). В частности, PDP1 и PDP2 представляют собой две изоформы, принадлежащие к семейству 2C / PPM, с молекулярной массой ∼55 кДа и Mg 2+ -зависимой каталитической субъединицей (Teague et al., 1982; Карпова и др., 2003). Ca 2+ -зависимая активация PDP1 приводит к дефосфорилированию PDH и, в конечном итоге, к превращению пирувата в ацетил-КоА (Teague et al., 1982; Turkan et al., 2004). Примечательно, что Ca 2+ активирует PDP1, но не PDP2, механизм, который все еще исследуется. Хотя последовательность PDP1 содержит предполагаемый мотив EF-руки (Lawson et al., 1993), кристаллическая структура PDP1 выявила, что этот домен не участвует в связывании Ca 2+ (Vassylyev and Symersky, 2007).Дальнейшие исследования, проведенные с использованием очищенного PDP1, показали, что сайт связывания Ca 2+ может находиться на границе раздела между PDP1 и E2 с Kd, близким к 1 мкМ (Turkan et al., 2004).
Изоцитратдегидрогеназа (IDH)
Этот фермент цикла TCA катализирует декарбоксилирование изоцитрата до α-кетоглутарата. IDH представляет собой октамер с массой около 320 кДа, и каждая единица октамера состоит из трех различных субъединиц (Nichols et al., 1993, 1995). Увеличение соотношений АТФ / АДФ и НАДН / НАД + ингибирует фермент IDH, свойство, которое присуще двум другим Ca 2+ -зависимым дегидрогеназам, пируватной и α-кетоглутаратдегидрогеназам.Чувствительность связывания Ca 2+ с IDH контролируется соотношением АТФ / АДФ. Уменьшение отношения АТФ / АДФ увеличивает связывание Ca 2+ с IDH, что, в свою очередь, приводит к снижению Km для изоцитрата (Rutter and Denton, 1988, 1989).
Оксоглутаратдегидрогеназа (OGDH)
Оксоглутаратдегидрогеназа катализирует реакцию цикла TCA, которая превращает α-кетоглутарат в сукцинил-КоА. Его тетрамерная структура имеет некоторое сходство с PDC. Он характеризуется ядром, состоящим из множества субъединиц дигидролипоамидсукцинилтрансферазы (E2), к которым присоединены субъединицы 2-оксоглутаратдекарбоксилазы (E1) и дигидролипоамиддегидрогеназы (E3).Увеличение соотношений сукцинил-КоА / КоА и НАДН / НАД + ингибирует OGDH. В отличие от PDC, OGDH не регулируется событиями фосфорилирования. Связывание Ca 2+ с OGDH приводит к снижению Km для α-кетоглутарата, подобно тому, что происходит для IDH (Denton, 2009).
Поглощение митохондриального Ca
2+ регулирует метаболизм в печениСистемная метаболическая адаптация к изменениям питания и изменениям физической активности необходима для энергетического гомеостаза.В частности, несколько механизмов и различных органов вовлечены в тонкую модуляцию метаболизма глюкозы. Среди них печень играет решающую роль из-за ее способности накапливать глюкозу во время еды и выпускать ее в кровоток во время голодания. Несколько сигналов контролируют баланс между хранением гликогена, гликогенолизом и глюконеогенезом. Инсулин отвечает за накопление гликогена, тогда как гипергликемические гормоны, включая глюкагон и адреналин, ответственны за глюконеогенез и гликогенолиз.Благодаря плодотворной работе лаборатории Кэбболда, динамика одноклеточного Ca 2+ гепатоцитов крысы выявила колебательный паттерн цитозольного Ca 2+ при гормональной стимуляции (Woods et al., 1986, 1987). Впоследствии был охарактеризован пространственно-временной паттерн транзиентов Ca 2+ и расшифрована соответствующая регуляция Ca 2+ -чувствительных белков и митохондриальных ферментов (Hajnóczky et al., 1995; Robb-Gaspers et al., 1998; обзор см. В Bartlett et al., 2014). В частности, Hajnóczky et al. (1995) продемонстрировали, что цитозольные колебания Ca 2+ транслируются в митохондриальные повышения Ca 2+ , чтобы стимулировать аэробный метаболизм. Более подробно, в то время как медленное увеличение концентрации цитозольного Ca 2+ недостаточно для активации митохондриального метаболизма, высвобождение Ca 2+ из ЭР после открытия рецептора InsP3 запускает быстрое цитозольное колебание Ca 2+ и, как следствие, митохондриальный Ca 2 + освоение.Таким образом, увеличение цитозольного [Ca 2+ ] из-за утечки Ca 2+ из хранилищ отличается от таковых из-за активации рецепторов InsP3, последние из которых способны запускать митохондриальный ответ. Каждый митохондриальный пик [Ca 2+ ], запускаемый определенными частотами цитозольных колебаний Ca 2+ , сопровождается максимальной активацией чувствительных к Ca 2+ митохондриальных дегидрогеназ.
Чтобы проверить роль поглощения митохондриальным Ca 2+ в метаболизме печени, была разработана гепатоцит-специфическая модель мыши MCU — / — (Tomar et al., 2019). MCU — / — гепатоциты демонстрировали истощение митохондриального матрикса Ca 2+ , сопровождающееся последующим нарушением дыхательной способности. Интересно, что в гепатоцитах MCU — / — наблюдается накопление липидов. Механически инактивация AMPK была указана как триггер этого события. Делеция MCU задерживает цитозольный клиренс Ca 2+ , и это увеличивает активность Ca 2+ -зависимой фосфатазы PP4. В свою очередь, PP4 дефосфорилирует AMPK.Примечательно, что специфической для печени делеции AMPK достаточно, чтобы вызвать накопление липидов в печени. Кроме того, была произведена модель мыши MCU с функцией усиления функции «нокаут» (Tomar et al., 2019). В этой модели гепатоциты демонстрировали повышенное поглощение митохондриями Ca 2+ и респираторную способность, AMPK был активен, а уровни триацилглицерина в печени были снижены. Следует отметить, что гепатоциты со сверхэкспрессией MCU были устойчивы к накоплению липидов при помещении в среду с высоким содержанием глюкозы.
Регенеративная способность печени имеет чрезвычайно важное патофизиологическое значение, и недавние данные показывают, что динамика Ca 2+ принимает участие в этом процессе.После частичной гепатэктомии повышение содержания Ca 2+ в цитозоле способствует переходу регенерирующих гепатоцитов через пролиферативное состояние, позволяя рост печени. Однако в печени MICU1 — / — повышение цитозольного Ca 2+ , вызванное частичной гепатэктомией, вызывает перегрузку митохондрий Ca 2+ и, в конечном итоге, открытие PTP. Это вызывает усиление провоспалительной фазы и блокировку перехода через пролиферативную фазу, что приводит к нарушению восстановления печени (Antony et al., 2016).
Митохондрии Ca
2+ Передача сигналов в контроле функции скелетных мышцФизиология мышц в значительной степени зависит от двух внутриклеточных органелл: SR для хранения и высвобождения Ca 2+ и митохондрий для синтеза АТФ (Franzini-Armstrong and Jorgensen, 1994). Скелетным мышцам необходим АТФ для управления как актомиозиновым поперечным мостиком, так и цитозольной буферизацией Ca 2+ во время сокращения, последнее обеспечивается АТФ-зависимой помповой активностью SERCA.Семенная работа продемонстрировала, что митохондрии мышечных трубок крыс усиливают в 4-6 раз цитозольные [Ca 2+ ] увеличения, запускаемые различными стимулами (Brini et al., 1997). Это наблюдение выдвинуло на первый план ключевую роль митохондрий как важных игроков в динамической регуляции передачи сигналов Ca 2+ в скелетных мышцах, в соответствии с сообщенным близким сопоставлением ER и митохондрий, которое обеспечивает эффективное поглощение Ca 2+ (Rizzuto et al. ., 1993, 1998). Электронно-микроскопические исследования продемонстрировали присутствие связывающих органеллы белков, расположенных на стыке между SR и митохондриями, которые создают физическое сцепление между этими двумя органеллами (Boncompagni et al., 2009), позволяя быстро переносить SR-митохондрии Ca 2+ . Ранние исследования продемонстрировали увеличение NADH / NAD + во время перехода от состояния покоя к рабочим мышцам, предполагая, что повышение внутриклеточного Ca 2+ способствует метаболизму митохондрий в скелетных мышцах (Sahlin, 1985; Duboc et al., 1988; Кунц, 2001). Недавно было показано, что ингибирование RyR1, то есть снижение поглощения митохондриальным Ca 2+ , запускает снижение потребления кислорода, связанного с АТФ, указывая на контроль аэробного метаболизма митохондриями [Ca 2+ ] ( Диас-Вегас и др., 2018). Важно отметить, что гликолитические и окислительные мышцы различаются по объему митохондрий и метаболическим свойствам. В частности, медленно сокращающиеся волокна полагаются на окислительное фосфорилирование, демонстрируя большую устойчивость к усталости, в то время как быстро сокращающиеся волокна в основном полагаются на гликолиз как на источник АТФ.
Важность митохондриального поглощения Ca 2+ в физиологии скелетных мышц демонстрируется фенотипом заболевания пациентов, несущих мутации в гене MICU1. Мутации с потерей функции вызывают миопатию, трудности в обучении и экстрапирамидные двигательные расстройства (Logan et al., 2014). У других пациентов с гомозиготной делецией экзона 1 MICU1 наблюдаются утомляемость, вялость и слабость (Lewis-Smith et al., 2016). Чтобы выяснить, являются ли мышечные дефекты следствием нарушения функции мышечного MICU1 или, скорее, вторичными по отношению к нейрональным нарушениям, была разработана мышиная модель MICU1 — / — скелетных мышц (Debattisti et al., 2019). Эти мыши характеризуются мышечной слабостью, утомляемостью и повышенной восприимчивостью к повреждающим сокращениям, что указывает на то, что потеря MICU1 в мышцах является причиной фенотипа заболевания.Кроме того, фармакологическое подавление захвата митохондриального Ca 2+ молекулами, нацеленными на MICU1, нарушает рост мышечных трубок (Di Marco et al., 2020).
ДелецияMCU в модели MCU — / — на мышах вызывает значительное нарушение способности к физической нагрузке (Pan et al., 2013). В изолированных митохондриях MCU — / — мускулатуры покоящегося матрикса уровни Ca 2+ снижены на 75% по сравнению с митохондриями дикого типа, а зависимое от Ca 2+ фосфорилирование PDH повышено.Эти данные соответствуют роли митохондриального накопления Ca 2+ в регулировании производства АТФ, необходимого для поддержания здоровой функциональности мышц.
Дальнейшие исследования были выполнены с помощью мышечной модуляции экспрессии MCU (Mammucari et al., 2015). В частности, сверхэкспрессия и подавление MCU вызывают гипертрофию и атрофию мышц, соответственно. Контроль массы скелетных мышц с помощью митохондриальной модуляции Ca 2+ происходит из-за активации двух основных гипертрофических путей скелетных мышц, PGC-1α4 и IGF1-AKT / PKB.Удивительно, но эти эффекты не зависят от контроля аэробного метаболизма, о чем свидетельствуют различные свидетельства. Во-первых, активность PDH, хотя и является дефектной в мышцах с молчанием MCU, не затрагивалась в мышцах со сверхэкспрессией MCU, и, во-вторых, гипертрофия была сопоставима как в окислительных, так и в гликолитических мышцах (Mammucari et al., 2015).
Мышечно-специфическая мышь с нокаутом MCU (skMCU — / — ), помимо подтверждения роли MCU в поддержании мышечной массы, помогла выяснить вклад митохондриального захвата Ca 2+ в метаболизм мышц.Кроме того, эта животная модель показала, как MCU-зависимый окислительный метаболизм скелетных мышц системно влияет на доступность субстрата (Gherardi et al., 2018). Метаболически мыши skMCU — / — характеризовались дефектным окислением глюкозы, что проявлялось в повышении содержания лактата в крови и снижении скорости потребления кислорода (OCR) миофибриллами в среде с высоким содержанием глюкозы. Поглощение глюкозы скелетными мышцами было увеличено, так же как и содержание гликогена, что свидетельствует о неизменной чувствительности к инсулину, но неэффективном использовании глюкозы.
Несмотря на снижение мышечной силы и работоспособности во время упражнений на утомление, производительность мышц не сильно ухудшилась. Это было связано с увеличением окисления жирных кислот (ЖК), которое в значительной степени способствовало базальному OCR (Gherardi et al., 2018). Вторая модель MCU KO, специфичная для скелетных мышц, характеризовалась, с одной стороны, пониженной производительностью спринтерских упражнений, с другой стороны, улучшенной работоспособностью мышц в условиях утомления, где преобладает метаболизм ЖК (Kwong et al., 2018). Из-за измененной утилизации глюкозы в мышцах печень и жировая ткань отреагировали усиленным катаболизмом. Глюконеогенез в печени, липолиз и, следовательно, циркулирующие кетоновые тела были увеличены. Кроме того, липолиз жировой ткани был усилен, что привело к снижению жировой массы тела как во взрослом возрасте (Gherardi et al., 2018), так и при старении (Kwong et al., 2018). Основным триггером этой метаболической перестройки является снижение активности пируватдегидрогеназы, которая жестко контролируется митохондриальным Ca 2+ , как показывают генетические эксперименты (Gherardi et al., 2018).
Соответственно, мышь с нокаутом, специфичным для скелетных мышц митохондриального носителя пирувата (MPC), поразительно напоминала фенотип мыши skMCU — / — . В скелетных мышцах делеция MPC снижает окисление глюкозы, несмотря на повышенное поглощение глюкозы и производство лактата. Это вызвало усиленное окисление ЖК и циклирование Кори с общим увеличением расхода энергии. Таким образом, ограниченная в скелетных мышцах делеция MPC ускоряла потерю жировой массы, когда мышей снова кормили диетой после ожирения, вызванного диетой с высоким содержанием жиров (Sharma et al., 2019). В целом эти исследования подтверждают модель, в которой нарушение митохондриального захвата Ca 2+ и ингибирование митохондриального проникновения пирувата приводят к аналогичным дефектам окисления глюкозы, что приводит к перекрыванию локальной и системной метаболической адаптации. Эти данные указывают на нарушение окисления пирувата как на основной фактор наблюдаемой перестройки метаболизма.
Сердечная энергия необходима для правильной работы сердца
В последнее десятилетие вклад митохондриального захвата Ca 2+ в регуляцию сердечной энергетики был глубоко исследован.Pozzan и соавторы продемонстрировали, что митохондриальные транзиенты Ca 2+ происходят от удара к удару и участвуют в формировании систолических пиков Ca 2+ в кардиомиоцитах новорожденных. Присутствие высоких микродоменов [Ca 2+ ], генерируемых в контактах SR / митохондрий, позволяет митохондриальному поглощению Ca 2+ во время систолы, что приводит к значительной буферизации пиков цитозольного Ca 2+ . Ca 2+ в конечном итоге высвобождается обратно в цитоплазму во время диастолы из-за присутствия обменника Na 2+ / Ca 2+ и антипортера Ca 2+ / H + .Кроме того, генетическая модуляция MCU, таким образом, изменение митохондриального накопления Ca 2+ , напрямую влияет на амплитуду цитоплазматических колебаний Ca 2+ в кардиомиоцитах новорожденных (Drago et al., 2012). Однако, происходит ли этот сценарий во взрослых кардиомиоцитах и в сердце, в котором было высказано предположение, что вход в митохондрии Ca 2+ не зависит от цитозольного порога Ca 2+ , все еще остается предметом дискуссий (Boyman et al. ., 2014).
За последние несколько лет было создано множество моделей in vivo на мышах для исследования роли митохондриального захвата Ca 2+ в контроле функции сердца. Согласно конститутивным и кардиоспецифическим моделям мышей MCU — / — , MCU незаменим для биоэнергетики митохондрий в сердце на исходном уровне (Pan et al., 2013; Kwong et al., 2015). На сердечную функцию мышей с конститутивным нокаутом MCU не влияет β-адренергическая стимуляция, что предполагает возможность долгосрочной адаптации (Holmström et al., 2015). Однако у индуцибельных кардиоспецифичных мышей с нокаутом MCU во время реакции борьбы или бегства требуется увеличение выработки митохондриального АТФ для поддержки повышенного энергопотребления увеличенной частоты сердечных сокращений (Kwong et al., 2015; Luongo et al. , 2015). Кроме того, сердца, экспрессирующие доминантно-отрицательные MCU или сверхэкспрессирующие MCUb, обработанные β-адренергическим агонистом изопротеренолом, демонстрируют нарушение ускорения сердечного ритма (Wu et al., 2015; Lambert et al., 2019).В условной модели трансгенной мыши MCUb, специфичной для сердца, биоэнергетика митохондрий была нарушена из-за повышенных уровней фосфорилирования ПДГ, что привело к снижению максимального дыхания и резервной емкости. Нарушение отзывчивости сердца было подчеркнуто тем фактом, что эти мыши не выжили после повреждения IR. Однако длительная индукция экспрессии MCUb сопровождалась восстановлением сердечной функции и уменьшением размера инфаркта, скорее всего, из-за компенсаторных механизмов, запускающих дефосфорилирование ПДГ и, следовательно, активацию (Lambert et al., 2019). Кроме того, в сердцах этих мышей наблюдали изменение стехиометрии комплекса MCU. Уровни MCU, MICU1 и MICU2 были увеличены, несмотря на снижение ассоциации MICU1 и MICU2 с комплексом MCU (Lambert et al., 2019).
Индуцибельная кардиоспецифическая модель MCU — / — на мышах была дополнительно исследована для оценки сердечных функций ex vivo (Altamimi et al., 2019). Неожиданно изолированное сердце MCU — / — показало улучшенную сердечную функцию даже на исходном уровне, которая сохранялась после стимуляции изопротеренолом.Таким образом, специфическое для сердца ингибирование захвата Ca 2+ митохондриями препятствует сердечному ответу на β-адренергическую стимуляцию in vivo (Kwong et al., 2015; Luongo et al., 2015; Wu et al., 2015), но а не ex vivo (Altamimi et al., 2019). Это говорит о том, что адренергический ответ всего тела по-другому влияет на работу сердца по сравнению с прямой стимуляцией. Детальный анализ предпочтения энергетических субстратов показал, что сердца с удаленными MCU зависят от повышенного окисления ЖК в ответ на β-адренергическую стимуляцию.Окисление ЖК коррелирует со снижением уровней малонил-КоА и с повышенным ацетилированием фермента β-окисления β-гидроксиацил-КоА дегидрогеназы (β-HAD) (Altamimi et al., 2019).
Основным участником митохондриального оттока Ca 2+ , необходимого для поддержания митохондриального гомеостаза Ca 2+ , является митохондриальный обменник Na + / Ca 2+ (NCLX), который кодируется Slc8b1. ген (Palty et al., 2010). Недавняя работа помогла выяснить последствия нарушения оттока Ca 2+ на сердечную функцию.Условная кардиомиоцит-специфическая делеция Slc8b1 вызывает внезапную смерть у большинства животных из-за острой дисфункции миокарда и молниеносной сердечной недостаточности (Luongo et al., 2017). Сердца с нокаутом Slc8b1 демонстрируют дезорганизацию саркомеров, некротическую гибель клеток и фиброз, сопровождающиеся дилатацией левого желудочка и снижением функции. Механически активация mPTP была ответственна за фенотип, о чем свидетельствует тот факт, что скрещивание этих мышей с циклофилин-D (CypD) -нуль-мышами спасает сердечную функцию и выживает.Напротив, кардиоспецифические трансгенные сердца NCLX характеризовались повышенной способностью к оттоку, пониженной активацией mPTP и были защищены от ИК-повреждения и ишемической сердечной недостаточности.
Кроме того, нарушения активности пируватдегидрогеназы в сердце вызывают метаболическую дисфункцию и, как следствие, кардиомиопатию. Patel и соавторы создали мышей с нокаутом PDH, специфичных для сердца и скелетных мышц (H / MS-PDCKO) (Sidhu et al., 2008). Активность PDC отсутствует как в скелетных мышцах, так и в сердце этих животных.Все мыши H / MS-PDCKO умерли через 7 дней после отъема на кормовой диете, и смерть была вызвана нарушением систолической функции левого желудочка, демонстрируя, что последующее увеличение скорости гликолиза было недостаточным для производства АТФ. требуется для нормальной сердечной функции. Напротив, когда мышей H / MS-PDCKO отнимали от диеты с высоким содержанием жиров, они выживали в течение многих месяцев. Однако даже в этом случае у этих мышей развилась гипертрофия левого желудочка, когда они вернулись на диету. Эти данные предполагают, что активность PDC обеспечивает метаболическую гибкость в сердце.Кроме того, Gopal et al. (2018) создали индуцибельную кардиоспецифичную модель нокаута ПДГ. У этих мышей отслеживали использование субстрата и сердечную функцию. Авторы наблюдали метаболическую перестройку, которая включает уменьшение окисления глюкозы в миокарде и увеличение окисления ЖК. Этот метаболический переключатель не изменил систолическую функцию, однако он нарушил диастолическую функцию, вызывая кардиомиопатический фенотип, подобный диабету или ожирению.
Эти данные предполагают, что, как и в скелетных мышцах, снижение митохондриального захвата Ca 2+ и нарушение окисления пирувата усиливают β-окисление в сердце.Недавняя работа Wescott et al. (2019) указывает, что, хотя митохондрии [Ca 2+ ] регулируют вход пирувата и глутамата в цикл TCA, окисление липидов не контролируется митохондриями [Ca 2+ ]. Таким образом, в контексте митохондриального нарушения Ca 2+ жирные кислоты легко окисляются для поддержания потребности в энергии.
Ca
2+ Сигнализация играет центральную роль в гомеостазе жировой тканиИзучение роли гомеостаза Ca 2+ в биологии жировой ткани представляет собой относительно новую область исследований.Различные данные свидетельствуют о том, что нарушение регуляции Ca 2+ может влиять на функцию адипоцитов. Соответственно, было продемонстрировано, что внутриклеточная концентрация Ca 2+ регулирует липидный метаболизм адипоцитов, таким образом контролируя адипогенез. Shi et al. (2000) показали, что цитозольный [Ca 2+ ] играет двухфазную регуляторную роль в дифференцировке адипоцитов: с одной стороны, увеличение цитозольного [Ca 2+ ] подавляет ранние стадии адипогенеза у мышей; с другой стороны, он способствует поздним стадиям дифференцировки жировой ткани.Наиболее важно то, что повышенное содержание внутриклеточного [Ca 2+ ] в адипоцитах, по-видимому, способствует инсулинорезистентности пожилых пациентов с гипертонической болезнью и ожирением. Поразительно, что 1 месяц терапии нитрендипином, блокатором каналов Ca 2+ , снизил артериальное давление, снизил уровень инсулина в плазме после стимуляции глюкозой до контрольных значений и восстановил нормальное поглощение глюкозы адипоцитами (Byyny et al., 1992).
В частности, в коричневой жировой ткани (BAT) ранние исследования изолированных адипоцитов продемонстрировали, что стимуляция норэпинефрином мобилизует внутриклеточные хранилища Ca 2+ (Connoly et al., 1984), что позже подтвердили последующие исследования (Lee et al., 1993; Leaver, Pappone, 2002; Nakagaki et al., 2005; Chen et al., 2017). Однако роль передачи сигналов Ca 2+ в термогенезе BAT оставалась в значительной степени неизученной до недавнего времени, когда Kcnk3, канал K + , присутствующий на плазматической мембране коричневых адипоцитов, был идентифицирован как негативный регулятор термогенеза (Chen et al. , 2017). В частности, уровни цитозольного Ca 2+ при адренергической стимуляции повышаются в коричневых адипоцитах с дефицитом Kcnk3, что позволяет предположить, что Kcnk3 ограничивает индуцированный NE цитозольный [Ca 2+ ] приток, что приводит к снижению продукции цАМФ, таким образом ослабляя липолиз и термогенное дыхание.
Хорошо известно, что окислительный метаболизм и, в целом, митохондриальная биоэнергетика изменяются при метаболическом синдроме, ожирении и диабете 2 типа (Heinonen et al., 2019), и что нормализация функции митохондрий может восстановить чувствительность к инсулину. Например, лечение розиглитазоном, агонистом PPARγ и хорошо зарекомендовавшим себя лекарством от диабета 2 типа (Lebovitz, 2019), вызывает усиление митохондриального дыхания и окисление ЖК в адипоцитах мышей ob / ob (Wilson-Fritch et al., 2004) и индуцирует экспрессию структурных белков митохондрий и клеточных антиоксидантных ферментов (Rong et al., 2007). Тем не менее, хотя вклад митохондриального метаболизма в гомеостаз жировой ткани хорошо известен, роль митохондриальной передачи сигналов Ca 2+ все еще в значительной степени не исследована. Недавно была исследована связь между активностью MCU и развитием ожирения и диабета (Wright et al., 2017). В адипоцитах 3T3-L1 инсулинорезистентность сопровождалась увеличением экспрессии MCU и MICU1 и митохондриальным захватом Ca 2+ .Соответственно, экспрессия компонентов комплекса MCU была увеличена в висцеральной жировой ткани (VAT) пациентов с ожирением, как преддиабетических, так и диабетических, по сравнению с худыми людьми. Интересно, что после бариатрической хирургии экспрессия компонентов комплекса MCU в НДС пациентов вернулась к физиологическим уровням, что свидетельствует о том, что митохондриальное накопление Ca 2+ играет активную роль в этой патологии. Сверхэкспрессия Mcub, хотя и недостаточна для устранения дефекта в инсулино-стимулированном поглощении глюкозы инсулинорезистентными клетками, восстанавливала продукцию ROS до нормального уровня.Кроме того, сверхэкспрессия Mcub снижает экспрессию TNF-альфа и IL-6, что указывает на возможную роль митохондриального захвата Ca 2+ в воспалении, связанном с ожирением (Wright et al., 2017). Недавно была охарактеризована BAT-специфичная мышь с нокаутом MCU (Flicker et al., 2019). Несмотря на полное отсутствие митохондриального поглощения Ca 2+ , не наблюдалось никаких эффектов с точки зрения толерантности к холоду и ожирения, вызванного диетой, что позволяет предположить, что MCU незаменимы для термогенеза и метаболизма BAT.Эти последние два исследования показывают, что MCU в жировой ткани играет разные роли в патологии и физиологии. Однако необходимы дальнейшие исследования, чтобы прояснить различную роль накопления митохондриального Ca 2+ в метаболически различных компартментах жировой ткани как в здоровом, так и в болезненном состоянии.
Дальнейшее понимание роли передачи сигналов Ca 2+ в регуляции метаболизма жировой ткани получено в исследованиях, проведенных на Drosophila melanogaster .В этой животной модели жировое тело, которое считается эквивалентом печени, жировой ткани, кроветворной и иммунной систем млекопитающих, определяет доступность питательных веществ и контролирует метаболические реакции (Hotamisligil, 2006). Что касается передачи сигналов Ca 2+ , Stim, мембранный белок ER, участвующий в накопительном входе Ca 2+ (SOCE), был идентифицирован с помощью скрининга РНКи среди генов, влияющих на ожирение у мух (Baumbach et al., 2014) . Подавление стимуляции в жировой ткани снижает уровни цитозольного Ca 2+ и приводит к увеличению общего ожирения.Это связано с удаленной активацией орексигенного короткого нейропептида F в головном мозге (sNPF). Наряду с гиперфагией ось Stim-sNPF контролирует метаболизм липидов, что приводит к гипертрофии жировой ткани. Дальнейшие доказательства роли гомеостаза Ca 2+ в метаболизме липидов D. melanogaster получены из исследований мутантов рецептора InsP3, которые страдают ожирением и демонстрируют измененное использование FA (Subramanian et al., 2013). Кроме того, Хуанг и соавторы показали, что dSeipin, который играет центральную роль в метаболизме липидов, взаимодействует с dSERCA, модулируя цитозольный [Ca 2+ ] (Bi et al., 2014). Мутации в dSeipin вызывают снижение активности SERCA и, как следствие, снижение уровней ER Ca 2+ , что приводит к нарушению накопления жира. Более того, у мутантов dSeipin митохондриальный захват Ca 2+ снижен, вызывая снижение активности цикла TCA и накопление цитрата, ключевого компонента липогенеза. Дефект накопления липидов в мутантных жировых клетках dSeipin устраняется восстановлением митохондрий [Ca 2+ ] или восстановлением нормальных уровней цитрата (Ding et al., 2018).
Заключительные замечания
Примерно через 10 лет после идентификации генов, кодирующих фундаментальный регулятор MCU (например, MICU1, Perocchi et al., 2010) и сам MCU (Baughman et al., 2011; De Stefani et al., 2011), большая часть собрана информация о функции канала и последствиях его нарушения. Большинство исследований основано на генетических манипуляциях с компонентами комплекса MCU в определенных субклеточных компартментах или на фенотипе заболевания пациентов, несущих мутации в генах, связанных с MCU.Недавно исследования, проведенные в метаболически активных тканях, подчеркнули важность MCU для обеспечения метаболической гибкости в использовании окисляемых субстратов. Отсутствие поступления Ca 2+ вызывает нарушение окислительного метаболизма в печени, сердце, скелетных мышцах и жировой ткани и изменение схемы утилизации субстрата, которая определяет повышенное окисление жирных кислот в сердце и скелетных мышцах, а также накопление липидов в печени. Дальнейшая работа прольет свет на важность захвата митохондриальным Ca 2+ и окисления пирувата в важнейших вопросах метаболизма, а также на перекрестное взаимодействие между различными органами.
Взносы авторов
Рукопись написалиGG и CM. HM подготовил цифры. RR способствовал критическому анализу. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Исследования в нашей лаборатории поддерживаются финансированием Французской ассоциации мышечной дистрофии (22493 для CM), Министерства исследований Италии (Prin 2015W2N883_003 для CM), Итальянской ассоциации исследований рака (AIRC) (IG18633 для RR), Университета из Падуи (грант STARS @ UNIPD WiC 2017 для RR), Итальянской ассоциации телемарафонов (GGP16029 для RR), Министерства здравоохранения Италии (RF-2016-02363566 для RR) и Фонда Карипаро (для RR).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Альтамими, Т. Р., Карви, К. Г., Уддин, Г. М., Фукусима, А., Квонг, Дж. К., Молькентин, Дж. Д. и др. (2019). Специфический для сердца дефицит митохондриального унипортера кальция увеличивает окисление жирных кислот и функциональный резерв. J. Mol.Клетка. Кардиол. 127, 223–231. DOI: 10.1016 / j.yjmcc.2018.12.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энтони, А.Н., Пайяр, М., Моффат, К., Юскевичуте, Э., Корренти, Дж., Болон, Б. и др. (2016). Регулирование поглощения Са2 + митохондриями с помощью MICU1 определяет выживание и регенерацию тканей. Нат. Commun. 7: 10955. DOI: 10.1038 / ncomms10955
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ашрафи, Г., де Хуан-Санс, Дж., Фаррелл, Р. Дж., И Райан, Т. А. (2020). Молекулярная настройка аксонального митохондриального унипортера Ca2 + обеспечивает метаболическую гибкость нейротрансмиссии. Нейрон 105, 678.e5–687.e5. DOI: 10.1016 / j.neuron.2019.11.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барадаран, Р., Ван, К., Силичиано, А. Ф., и Лонг, С. Б. (2018). Крио-ЭМ структуры унипортеров кальция митохондрий грибов и многоклеточных животных. Природа 559, 580–584. DOI: 10.1038 / s41586-018-0331-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бартлетт П.Дж., Гасперс, Л. Д., Пьеробон, Н., Томас, А. П. (2014). Кальцийзависимая регуляция гомеостаза глюкозы в печени. Cell Calc. 55, 306–316. DOI: 10.1016 / j.ceca.2014.02.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Baughman, J.M., Perocchi, F., Girgis, H.S., Plovanich, M., Belcher-Timme, C.A., Sancak, Y., et al. (2011). Интегративная геномика определяет MCU как важный компонент митохондриального унипортера кальция. Природа 476, 341–345.DOI: 10.1038 / природа10234
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Баумбах Дж., Хуммель П., Бикмайер И., Ковальчик К. М., Франк М., Кнорр К. и др. (2014). Скрининг дрозофилы in vivo определяет поступление кальция, управляемое хранилищами, как ключевой регулятор ожирения. Cell Metab. 19, 331–343. DOI: 10.1016 / J.CMET.2013.12.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берридж, М. Дж. (2001). Универсальность и сложность кальциевой сигнализации. Новартис Найдено. Symp. 239, 52–64; обсуждение 6, 150–159.
Google Scholar
Би, Дж., Ван, В., Лю, З., Хуанг, X., Цзян, К., Лю, Г. и др. (2014). Seipin способствует накоплению жира в жировой ткани через ER Ca2 + -ATPase SERCA. Cell Metab. 19, 861–871. DOI: 10.1016 / j.cmet.2014.03.028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бонкомпаньи, С., Росси, А.Э., Микарони, М., Безноусенко, Г.В., Полищук, Р.С., Дирксен, Р. Т. и др. (2009). Митохондрии связаны с запасами кальция в поперечно-полосатых мышцах посредством регулируемых в процессе развития связывающих структур. Mol. Биол. Cell 20, 1058–1067. DOI: 10.1091 / mbc.e08-07-0783
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бойман, Л., Чикандо, А. К., Уильямс, Г. С. Б., Хайраллах, Р. Дж., Кеттлвелл, С., Уорд, К. В. и др. (2014). Движение кальция в сердечных митохондриях. Biophys. J. 107, 1289–1301.DOI: 10.1016 / j.bpj.2014.07.045
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брини М., Де Джорджи Ф., Мурджа М., Марсо Р., Массимино М. Л., Кантини М. и др. (1997). Субклеточный анализ гомеостаза Са2 + в первичных культурах миотрубок скелетных мышц. Mol. Биол. Cell 8, 129–143. DOI: 10.1091 / mbc.8.1.129
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бристон, Т., Робертс, М., Льюис, С., Пауни, Б., Staddon, J.M., Szabadkai, G., et al. (2017). Митохондриальная проницаемость переходной поры: чувствительность к открытию и механистическая зависимость от доступности субстрата. Sci. Отчет 7: 10492. DOI: 10.1038 / s41598-017-10673-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Byyny, R. L., Verde, M., Lo, Lloyd, S., Mitchell, W., and Draznin, B. (1992). Цитозольный кальций и инсулинорезистентность у пожилых пациентов с гипертонической болезнью. Am. J. Hypertens. 5, 459–464. DOI: 10.1093 / ajh / 5.7.459
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, Ю., Цзэн, X., Хуанг, X., Сераг, С., Вульф, К. Дж., И Шпигельман, Б. М. (2017). Перекрестное взаимодействие между KCNK3-опосредованным ионным током и адренергической передачей сигналов регулирует термогенез жировой ткани и ожирение. Ячейка 171, 836.e13–848.e13. DOI: 10.1016 / j.cell.2017.09.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коул, Э.С., Лепп, К.А., Холохан П. Д. и Фонди Т. П. (1978). Выделение и характеристика флавин-связанной глицерин-3-фосфатдегидрогеназы из митохондрий скелетных мышц кролика и сравнение с ферментом из мозга кролика. J. Biol. Chem. 253, 7952–7959.
Google Scholar
Конноли, Э., Нанберг, Э. и Недргаард, Дж. (1984). Na + -зависимая, альфа-адренергическая мобилизация внутриклеточного (митохондриального) Ca2 + в коричневых адипоцитах. Eur. J. Biochem. 141, 187–193.DOI: 10.1111 / j.1432-1033.1984.tb08173.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Csordás, G., Golenár, T., Seifert, E.L., Kamer, K.J., Sancak, Y., Perocchi, F., et al. (2013). MICU1 контролирует как пороговую, так и кооперативную активацию митохондриального унипортера Ca2 +. Cell Metab. 17, 976–987. DOI: 10.1016 / j.cmet.2013.04.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Де Стефани, Д., Рафаэлло, А., Тирдо, Э., Сабо, И., и Риццуто, Р. (2011). Белок внутренней мембраны в сорок килодальтон является митохондриальным унипортером кальция. Природа 476, 336–340. DOI: 10.1038 / природа10230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дебаттисти В., Хорн А., Сингх Р., Зайферт Э. Л., Хогарт М. В., Мазала Д. А. и др. (2019). Нарушение регуляции митохондриального захвата Ca2 + и восстановления сарколеммы лежат в основе мышечной слабости и истощения у пациентов и мышей, лишенных MICU1. Cell Rep. 29, 1274.e – 1286.e. DOI: 10.1016 / j.celrep.2019.09.063
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Делука, Х. Ф., и Энгстрем, Г. У. (1961). Поглощение кальция митохондриями почек крысы. Proc. Natl. Акад. Sci. США 47, 1744–1750.
Google Scholar
Ди Марко, Г., Валлезе, Ф., Журде, Б., Бергсдорф, К., Стурлезе, М., Де Марио, А. и др. (2020). Высокопроизводительный скрининг выявляет соединения, нацеленные на MICU1. Cell Rep. 30, 2321.e6–2331.e6. DOI: 10.1016 / j.celrep.2020.01.081
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Диас-Вегас, А. Р., Кордова, А., Валладарес, Д., Льянос, П., Идальго, К., Герарди, Г. и др. (2018). Повышение содержания кальция в митохондриях, индуцированное RyR1 и IP 3 активация R-канала после деполяризации мембраны регулирует метаболизм скелетных мышц. Фронт. Physiol. 9: 791. DOI: 10.3389 / fphys.2018.00791
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дин, Л., Ян, X., Тиан, Х., Лян, Дж., Чжан, Ф., Ван, Г., и др. (2018). Сейпин регулирует гомеостаз липидов, обеспечивая кальций-зависимый митохондриальный метаболизм. Эмбо Дж. 37: e97572. DOI: 10.15252 / embj.201797572
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Драго И., Де Стефани Д., Риццуто Р. и Поццан Т. (2012). Поглощение митохондриального Ca2 + способствует буферизации пиков цитоплазматического Ca2 + в кардиомиоцитах. Proc. Natl. Акад. Sci. США 109, 12986–12991.DOI: 10.1073 / pnas.1210718109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Duboc, D., Muffat-Joly, M., Renault, G., Degeorges, M., Toussaint, M., and Pocidalo, J. J. (1988). NADH-лазерная флуориметрия in situ быстро- и медленно сокращающихся мышц крыс при столбняке. J. Appl. Physiol. 64, 2692–2695. DOI: 10.1152 / jappl.1988.64.6.2692
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фан, К., Фан, М., Орландо, Б. Дж., Фастман, Н.M., Zhang, J., Xu, Y., et al. (2018). Рентгеновские и крио-ЭМ структуры митохондриального унипортера кальция. Природа 559, 575–579. DOI: 10.1038 / s41586-018-0330-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Fan, M., Zhang, J., Tsai, C.-W., Orlando, B.J., Rodriguez, M., Xu, Y., et al. (2020). Структура и механизм митохондриального унипортерного холокомплекса Са2 +. Природа 582, 129–133. DOI: 10.1038 / s41586-020-2309-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фиени, Ф., Ли, С. Б., Ян, Ю. Н., Киричок, Ю. (2012). Активность митохондриального унипортера кальция сильно различается между тканями. Нат. Commun. 3: 1317. DOI: 10.1038 / ncomms2325
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фликер, Д., Санджак, Ю., Мик, Э., Голдбергер, О., и Мутха, В. К. (2019). Изучение in vivo роли митохондриального унипортера кальция в биоэнергетике бурого жира. Cell Rep. 27, 1364.e5–1375.e5. DOI: 10.1016 / J.CELREP.2019.04.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Францини-Армстронг, К., Йоргенсен, А. О. (1994). Структура и развитие блоков сцепления E-C в скелетных мышцах. Annu. Rev. Physiol. 56, 509–534. DOI: 10.1146 / annurev.ph.56.030194.002453
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гарриб А. и МакМюррей В. К. (1986). Очистка и характеристика глицерин-3-фосфатдегидрогеназы (флавин-связанной) из митохондрий печени крыс. J. Biol. Chem. 261, 8042–8048.
Google Scholar
Gherardi, G., Nogara, L., Ciciliot, S., Fadini, G.P., Blaauw, B., Braghetta, P., et al. (2018). Потеря митохондриального унипортера кальция изменяет метаболизм скелетных мышц и предпочтение субстрата. Cell Death Differ. 26, 362–381. DOI: 10.1038 / s41418-018-0191-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джорджио В., Го Л., Бассо К., Петронилли В. и Бернарди П.(2018). Кальций и регуляция перехода митохондриальной проницаемости. Cell Calc. 70, 56–63. DOI: 10.1016 / j.ceca.2017.05.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Глэнси, Б., Уиллис, У. Т., Чесс, Д. Дж., И Балабан, Р. С. (2013). Влияние кальция на каскад окислительного фосфорилирования в митохондриях скелетных мышц. Биохимия 52, 2793–2809. DOI: 10.1021 / bi3015983
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гопал, К., Альмутаири, М., Аль Батран, Р., Итон, Ф., Ганди, М., и Ашер, Дж. Р. (2018). Специфическая делеция пируватдегидрогеназы снижает скорость окисления глюкозы и вызывает диастолическую дисфункцию. Фронт. Кардиоваск. Med. 5:17. DOI: 10.3389 / fcvm.2018.00017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хайноцкий, Г., Робб-Гасперс, Л. Д., Зейтц, М. Б., и Томас, А. П. (1995). Расшифровка колебаний цитозольного кальция в митохондриях. Cell 82, 415–424.
Google Scholar
Хейнонен, С., Йокинен, Р., Риссанен, А., и Пиетилайнен, К. Х. (2019). Метаболизм митохондрий белой жировой ткани в условиях здоровья и ожирения. Obes. Версия 21: e12958. DOI: 10.1111 / obr.12958
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хиромаса Ю., Фудзисава Т., Асо Ю. и Рош Т. Е. (2004). Организация ядер комплекса пируватдегидрогеназы млекопитающих, образованного E 2 и E 2 плюс E 3-связывающим белком, и их способности связывать компоненты E 1 и E 3. J. Biol. Chem. 279, 6921–6933. DOI: 10.1074 / jbc.M308172200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hoffman, N.E., Chandramoorthy, H.C., Shanmughapriya, S., Zhang, X.Q., Vallem, S., Doonan, P.J., et al. (2014). SLC25A23 увеличивает поглощение митохондриальным Ca2 +, взаимодействует с MCU и вызывает гибель клеток, опосредованную окислительным стрессом. Mol. Биол. Cell 25, 936–947. DOI: 10.1091 / mbc.E13-08-0502
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хольмстрём, К.M., Pan, X., Liu, J. C., Menazza, S., Liu, J., Nguyen, T. T., et al. (2015). Оценка сердечной функции у мышей, лишенных митохондриального унипортера кальция. J. Mol. Клетка. Кардиол. 85, 178–182. DOI: 10.1016 / j.yjmcc.2015.05.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hung, V., Zou, P., Rhee, H.-W., Udeshi, N.D., Cracan, V., Svinkina, T., et al. (2014). Протеомное картирование митохондриального межмембранного пространства человека в живых клетках с помощью ратиометрической метки APEX. Mol. Ячейка 55, 332–341. DOI: 10.1016 / j.molcel.2014.06.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Камер, К. Дж., Грабарек, З., и Мутха, В. К. (2017). Высокоаффинное кооперативное связывание Ca 2+ с помощью MICU 1– MICU 2 служит переключателем для унипортера. Embo Rep. 18, 1397–1411. DOI: 10.15252 / embr.201643748
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Карпова Т., Данчук С., Колобова Е., и Попов, К. М. (2003). Характеристика изоферментов фосфатазы пируватдегидрогеназы: значение для регуляции активности пируватдегидрогеназы. Biochim. Биофиз. Протеомика белков Acta 1652, 126–135. DOI: 10.1016 / j.bbapap.2003.08.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ковач-Богдан, Э., Санчак, Ю., Камер, К. Дж., Плованич, М., Джамбекар, А., Хубер, Р. Дж. И др. (2014). Восстановление митохондриального унипортера кальция в дрожжах. Proc. Natl. Акад. Sci. США 111, 8985–8990. DOI: 10.1073 / pnas.1400514111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кунц, В. С. (2001). Контроль окислительного фосфорилирования в скелетных мышцах. Biochim. Биофиз. Acta Bioenerg. 1504, 12–19. DOI: 10.1016 / S0005-2728 (00) 00235-8
CrossRef Полный текст | Google Scholar
Kwong, J. Q., Huo, J., Bround, M. J., Boyer, J. G., Schwanekamp, J. A., Ghazal, N., et al.(2018). Митохондриальный унипортер кальция лежит в основе предпочтения метаболического топлива в скелетных мышцах. JCI Insight 3: e121689. DOI: 10.1172 / jci.insight.121689
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Квонг, Дж. К., Лу, X., Коррелл, Р. Н., Шванекамп, Дж. А., Вагноцци, Р. Дж., Сарджент, М. А., и др. (2015). Митохондриальный унипортер кальция избирательно сопоставляет метаболический выход с острым сократительным стрессом в сердце. Cell Rep. 12, 15–22.DOI: 10.1016 / j.celrep.2015.06.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лам, С.С., Мартелл, Дж. Д., Камер, К. Дж., Деринк, Т. Дж., Эллисман, М. Х., Мутха, В. К. и др. (2015). Направленная эволюция APEX2 для электронной микроскопии и бесконтактного мечения. Нат. Методы 12, 51–54. DOI: 10.1038 / nmeth.3179
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ламберт, Дж. П., Луонго, Т. С., Томар, Д., Джадия, П., Гао, Э., Zhang, X., et al. (2019). MCUB регулирует молекулярный состав митохондриального унипортерного канала кальция, чтобы ограничить перегрузку митохондрий кальцием во время стресса. Тираж 140, 1720–1733. DOI: 10.1161 / CIRCULATIONAHA.118.037968
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоусон, Дж. Э., Ниу, X.-D., Браунинг, К. С., Тронг, Х., Ле, Ян, Дж. И др. (1993). Молекулярное клонирование и экспрессия каталитической субъединицы бычьей пируватдегидрогеназы фосфатазы и сходство последовательности с протеинфосфатазой 2C. Биохимия 32, 8987–8993. DOI: 10.1021 / bi00086a002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ливер, Э. В., и Паппоне, П. А. (2002). β-Адренергическое усиление высвобождения Ca 2+ эндоплазматического ретикулума в бурых жировых клетках. Am. J. Physiol. Physiol. 282, C1016 – C1024. DOI: 10.1152 / ajpcell.00204.2001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли С.С., Нуччителли Р. и Паппоне П.А. (1993).Адренергически активированный Ca2 + увеличивается в коричневых жировых клетках: эффекты блокировки Ca2 +, K + и K. Am. J. Physiol. Physiol. 264, C217 – C228. DOI: 10.1152 / ajpcell.1993.264.1.C217
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Льюис-Смит, Д., Камер, К. Дж., Гриффин, Х., Чайлдс, А.-М., Писден, К., Титов, Д., и др. (2016). Гомозиготная делеция в MICU1 проявляется утомляемостью и летаргией в детстве. Neurol. Genet. 2: e59. DOI: 10.1212 / NXG.0000000000000059
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, Дж. К., Лю, Дж., Холмстрём, К. М., Менацца, С., Паркс, Р. Дж., Фергюссон, М. М. и др. (2016). MICU1 служит молекулярным привратником для предотвращения перегрузки митохондрий кальцием in vivo. Cell Rep. 16, 1561–1573. DOI: 10.1016 / j.celrep.2016.07.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, Дж. К., Сидер, Н. К., Гораши, Н. С., Уиллингем, Т.Б., Паркс, Р. Дж., Сан, Дж. И др. (2020). EMRE важен для митохондриальной активности унипортера кальция на мышиной модели. JCI Insight 5: e134063. DOI: 10.1172 / jci.insight.134063
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Логан К. В., Сабадкай Г., Шарп Дж. А., Парри Д. А., Торелли С., Чайлдс А.-М. и др. (2014). Мутации с потерей функции в MICU1 вызывают расстройство мозга и мышц, связанное с первичными изменениями митохондриальной передачи сигналов кальция. Нат. Genet. 46, 188–193. DOI: 10,1038 / нг.2851
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Луонго, Т. С., Ламберт, Дж. П., Гросс, П., Нвокеди, М., Ломбарди, А. А., Шанмугхаприя, С. и др. (2017). Митохондриальный обменник Na + / Ca2 + необходим для гомеостаза и жизнеспособности Ca2 +. Nature 545, 93–97. DOI: 10.1038 / nature22082
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Луонго, Т.С., Ламберт, Дж.П., Юань, А., Чжан, X., Гросс, П., Сонг, Дж. И др. (2015). Митохондриальный унипортер кальция согласовывает снабжение энергией с нагрузкой на сердце во время стресса и модулирует переход проницаемости. Cell Rep. 12, 23–34. DOI: 10.1016 / j.celrep.2015.06.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Макдональд, М. Дж., И Браун, Л. Дж. (1996). Повторное исследование кальциевой активации митохондриальной глицеринфосфатдегидрогеназы. Arch. Biochem. Биофиз. 326, 79–84. DOI: 10.1006 / abbi.1996.0049
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маллиланкараман К., Карденас К., Дунан П. Дж., Чандрамурти Х. К., Ирринки К. М., Голенар Т. и др. (2012a). MCUR1 является важным компонентом захвата митохондриальным Ca2 +, который регулирует клеточный метаболизм. Нат. Cell Biol. 14, 1336–1343. DOI: 10.1038 / ncb2622
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маллиланкараман, К., Doonan, P., Cárdenas, C., Chandramoorthy, H.C., Müller, M., Miller, R., et al. (2012b). MICU1 является важным привратником для MCU-опосредованного поглощения Ca2 + митохондриями, который регулирует выживаемость клеток. Ячейка 151, 630–644. DOI: 10.1016 / j.cell.2012.10.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маммукари К., Герарди Г., Зампаро И., Рафаэлло А., Бонкомпаньи С., Чемелло Ф. и др. (2015). Митохондриальный унипортер кальция контролирует трофику скелетных мышц in vivo. Cell Rep. 10, 1269–1279. DOI: 10.1016 / j.celrep.2015.01.056
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерфи, Э., Пан, X., Нгуен, Т., Лю, Дж., Холмстрем, К. М., и Финкель, Т. (2014). Нерешенные вопросы из анализа мышей с отсутствием экспрессии MCU. Biochem. Биофиз. Res. Commun. 449, 384–385. DOI: 10.1016 / j.bbrc.2014.04.144
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Накагаки, И., Сасаки, С., Яхата, Т., Такасаки, Х., и Хори, С. (2005). Цитоплазматические и митохондриальные уровни Са2 + в коричневых адипоцитах. Acta Physiol. Сканд. 183, 89–97. DOI: 10.1111 / j.1365-201X.2004.01367.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Nguyen, N.X., Armache, J.-P., Lee, C., Yang, Y., Zeng, W., Mootha, V.K., et al. (2018). Крио-ЭМ структура грибкового митохондриального унипортера кальция. Природа 559, 570–574. DOI: 10.1038 / s41586-018-0333-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Николс, Б.Дж., Холл, Л., Перри, А. К. и Дентон, Р. М. (1993). Молекулярное клонирование и выведенные аминокислотные последовательности гамма-субъединиц НАД (+) — изоцитратдегидрогеназ крысы и обезьяны. Biochem. J. 295 (Pt 2), 347–350. DOI: 10.1042 / bj2950347
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Николс, Б. Дж., Перри, А. К., Холл, Л., и Дентон, Р. М. (1995). Молекулярное клонирование и выведенные аминокислотные последовательности альфа- и бета-субъединиц НАД (+) — изоцитратдегидрогеназы млекопитающих. Biochem. J. 310 (Pt 3), 917–922. DOI: 10.1042 / bj3100917
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Oxenoid, K., Dong, Y., Cao, C., Cui, T., Sancak, Y., Markhard, A. L., et al. (2016). Архитектура митохондриального унипортера кальция. Природа 533, 269–273. DOI: 10.1038 / природа17656
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пайяр, М., Чордаш, Г., Санда, Г., Голенар, Т., Дебаттисти, В., Барток А. и др. (2017). Тканеспецифическое митохондриальное декодирование цитоплазматических сигналов Ca 2+ контролируется стехиометрией MICU1 / 2 и MCU. Cell Rep. 18, 2291–2300. DOI: 10.1016 / j.celrep.2017.02.032
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Палти Р., Сильверман В. Ф., Хершфинкель М., Капорале Т., Сенси С. Л., Парнис Дж. И др. (2010). NCLX является важным компонентом митохондриального обмена Na + / Ca2 +. Proc. Natl. Акад.Sci. США 107, 436–441. DOI: 10.1073 / pnas.09107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пан, X., Лю, Дж., Нгуен, Т., Лю, К., Сан, Дж., Тэн, Ю. и др. (2013). Физиологическая роль митохондриального кальция выявлена на мышах, лишенных митохондриального унипортера кальция. Нат. Cell Biol. 15, 1464–1472. DOI: 10.1038 / ncb2868
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Покровитель, М., Чеккетто, В., Raffaello, A., Teardo, E., Vecellio Reane, D., Mantoan, M., et al. (2014). MICU1 и MICU2 точно настраивают митохондриальный унипортер Ca2 +, оказывая противоположные эффекты на активность MCU. Mol. Cell 53, 726–737. DOI: 10.1016 / j.molcel.2014.01.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Патрон, М., Гранатьеро, В., Эспино, Дж., Риццуто, Р., и Де Стефани, Д. (2019). MICU3 — тканеспецифический усилитель поглощения кальция митохондриями. Cell Death Differ. 26, 179–195. DOI: 10.1038 / s41418-018-0113-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пауп В., Прудент Дж., Дасса Э. П., Рендон О. З. и Шубридж Э. А. (2015). CCDC90A (MCUR1) является фактором сборки цитохром-с-оксидазы, а не регулятором митохондриального унипортера кальция. Cell Metab. 21, 109–116. DOI: 10.1016 / j.cmet.2014.12.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перокки, Ф., Гохил, В. М., Гирджис, Х. С., Бао, X. Р., МакКомбс, Дж. Э., Палмер, А. Е. и др. (2010). MICU1 кодирует митохондриальный белок руки EF, необходимый для поглощения Ca2 +. Природа 467, 291–296. DOI: 10.1038 / nature09358
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Плованич, М., Богорад, Р. Л., Санджак, Ю., Камер, К. Дж., Стритматтер, Л., Ли, А. А. и др. (2013). MICU2, паралог MICU1, находится в митохондриальном унипортерном комплексе, чтобы регулировать обработку кальция. PLoS One 8: e55785. DOI: 10.1371 / journal.pone.0055785
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рафаэлло А., Де Стефани Д., Саббадин Д., Тирдо Э., Мерли Г., Пикард А. и др. (2013). Митохондриальный унипортер кальция представляет собой мультимер, который может включать доминантно-отрицательную порообразующую субъединицу. Embo J. 32, 2362–2376. DOI: 10.1038 / emboj.2013.157
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Риццуто, Р., Брини, М., Мурджа, М., и Поццан, Т. (1993). Микродомены с высоким содержанием Ca2 +, близкие к IP3-чувствительным каналам, которые воспринимаются соседними митохондриями. Наука 262, 744–747.
Google Scholar
Риццуто, Р., Пинтон, П., Кэррингтон, В., Фэй, Ф. С., Фогарти, К. Э., Лифшиц, Л. М. и др. (1998). Тесные контакты с эндоплазматическим ретикулумом как детерминанты митохондриальных ответов Ca2 +. Science 280, 1763–1766. DOI: 10.1126 / science.280.5370.1763
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Робб-Гасперс, Л.Д., Бернетт, П., Раттер, Г. А., Дентон, Р. М., Риццуто, Р., и Томас, А. П. (1998). Интеграция сигналов цитозольного кальция в метаболические ответы митохондрий. Embo J. 17, 4987–5000. DOI: 10.1093 / emboj / 17.17.4987
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rong, J. X., Qiu, Y., Hansen, M. K., Zhu, L., Zhang, V., Xie, M., et al. (2007). Митохондриальный биогенез жировой ткани подавляется у мышей, получавших диету db / db и с высоким содержанием жиров, и улучшается с помощью розиглитазона. Диабет 56, 1751–1760. DOI: 10.2337 / db06-1135
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Росси А., Пиццо П. и Филади Р. (2019). Кальций, митохондрии и клеточный метаболизм: функциональный треугольник в биоэнергетике. Biochim. Биофиз. Acta Mol. Cell Res. 1866, 1068–1078. DOI: 10.1016 / j.bbamcr.2018.10.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Раттер Г. А. и Дентон Р. М. (1988).Регулирование NAD + -связанной изоцитратдегидрогеназы и 2-оксоглутаратдегидрогеназы ионами Ca2 + в митохондриях сердца крысы, проницаемых для толуола. Взаимодействие с регуляцией адениновых нуклеотидов и соотношением НАДН / НАД +. Biochem. J. 252, 181–189.
Google Scholar
Раттер, Г. А., и Дентон, Р. М. (1989). Связывание ионов Са2 + с НАД + -изоцитратдегидрогеназой сердца свиньи и комплексом 2-оксоглутаратдегидрогеназы. Biochem. J. 263, 453–462.DOI: 10.1042 / bj2630453
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Санджак Ю., Маркхард А. Л., Китами Т., Ковач-Богдан Э., Камер К. Дж., Удеши Н. Д. и др. (2013). EMRE является важным компонентом митохондриального унипортерного комплекса кальция. Science 342, 1379–1382. DOI: 10.1126 / science.1242993
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарма, А., Оонтонпан, Л., Шелдон, Р. Д., Раукхорст, А.J., Zhu, Z., Tompkins, S.C., et al. (2019). Нарушение поглощения митохондриального пирувата скелетными мышцами изменяет метаболизм глюкозы, что приводит к похуданию всего тела. eLife 8: e45873. DOI: 10.7554 / eLife.45873
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ши, Х., Халворсен, Й.-Д., Эллис, П. Н., Уилкисон, У. О., Земель, М. Б. (2000). Роль внутриклеточного кальция в дифференцировке адипоцитов человека. Physiol. Геномика 3, 75–82. DOI: 10.1152 / физиолгеномика.2000.3.2.75
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сидху, С., Гангасани, А., Коротчкина, Л. Г., Сузуки, Г., Фаллаволлита, Дж. А., Канти, Дж. М. и др. (2008). Дефицит тканеспецифического комплекса пируватдегидрогеназы вызывает гипертрофию сердца и внезапную смерть самцов мышей, отлученных от груди. Am. J. Physiol. Heart Circ. Physiol. 295, H946 – H952. DOI: 10.1152 / ajpheart.00363.2008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Субраманян, М., Метя, С. К., Садаф, С., Кумар, С., Швудке, Д., и Хасан, Г. (2013). Измененный гомеостаз липидов у мутантов рецептора InsP3 дрозофилы приводит к ожирению и гиперфагии. Dis. Модель. Мех. 6, 734–744. DOI: 10.1242 / dmm.010017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тиг, В. М., Петтит, Ф. Х., Ву, Т. Л., Сильберман, С. Р., и Рид, Л. Дж. (1982). Очистка и свойства фосфатазы пируватдегидрогеназы из сердца и почек крупного рогатого скота. Биохимия 21, 5585–5592. DOI: 10.1021 / bi00265a031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Террито, П. Р., Мута, В. К., Френч, С. А., и Балабан, Р. С. (2000). Са 2+ активация окислительного фосфорилирования митохондрий сердца: роль F 0 / F 1 -АТФазы. Am. J. Physiol. Physiol. 278, C423 – C435. DOI: 10.1152 / ajpcell.2000.278.2.C423
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Томар, Д., Jaña, F., Dong, Z., Quinn, W.J., Jadiya, P., Breves, S.L. и др. (2019). Блокада MCU-опосредованного захвата Ca2 + нарушает метаболизм липидов за счет PP4-зависимого дефосфорилирования AMPK. Cell Rep. 26, 3709.e7–3725.e7. DOI: 10.1016 / J.CELREP.2019.02.107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тюркан А., Хиромаса Ю. и Рош Т. Е. (2004). Образование комплекса каталитической субъединицы изоформы 1 фосфатазы пируватдегидрогеназы (PDP1c) и домена L2 образует сайт связывания Ca 2+ и захватывает PDP1c в качестве мономера †. Биохимия 43, 15073–15085. DOI: 10.1021 / bi048901y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вайс, Х., Маллиланкараман, К., Мак, Д.-О. D., Hoff, H., Payne, R., Tanis, J.E., et al. (2016). EMRE — это матричный Ca2 + -сенсор, который управляет воротами митохондриального унипортера Ca2 +. Cell Rep. 14, 403–410. DOI: 10.1016 / j.celrep.2015.12.054
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Васингтон, Ф.Д. и Мерфи Дж. В. (1962). Поглощение ионов Са митохондриями почек крысы и его зависимость от дыхания и фосфорилирования. J. Biol. Chem. 237, 2670–2677.
Google Scholar
Васылев Д. Г., Сымерский Дж. (2007). Кристаллическая структура пируватдегидрогеназы фосфатазы 1 и ее функциональное значение. J. Mol. Биол. 370, 417–426. DOI: 10.1016 / j.jmb.2007.05.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вечеллио Реане, Д., Vallese, F., Checchetto, V., Acquasaliente, L., Butera, G., De Filippis, V., et al. (2016). Вариант сплайсинга MICU1 придает высокую чувствительность митохондриальному механизму захвата Са 2+ скелетными мышцами. Mol. Ячейка 64, 760–773. DOI: 10.1016 / j.molcel.2016.10.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Waldeck-Weiermair, M., Malli, R., Parichatikanond, W., Gottschalk, B., Madreiter-Sokolowski, C.T., Klec, C., et al. (2015). Перегруппировка мультимеров MICU1 для активации MCU контролируется исключительно цитозольным Ca (2.). Sci. Отчет 5: 15602. DOI: 10.1038 / srep15602
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, L., Yang, X., Li, S., Wang, Z., Liu, Y., Feng, J., et al. (2014). Структурные и механистические сведения о регуляции MICU1 поглощения митохондриального кальция. EMBO J. 33, 594–604. DOI: 10.1002 / embj.201386523
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, Y., Nguyen, N. X., She, J., Zeng, W., Yang, Y., Bai, X., et al. (2019). Структурный механизм EMRE-зависимого стробирования митохондриального унипортера кальция человека. Ячейка 177, 1252.e13–1261.e13. DOI: 10.1016 / j.cell.2019.03.050
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вескотт, А. П., Као, Дж. П. Ю., Ледерер, В. Дж., И Бойман, Л. (2019). Производство кальций-чувствительного АТФ под действием напряжения митохондриями. Нат. Метаб. 1, 975–984. DOI: 10.1038 / s42255-019-0126-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уилсон-Фритч, Л., Nicoloro, S., Chouinard, M., Lazar, M. A., Chui, P. C., Leszyk, J., et al. (2004). Ремоделирование митохондрий в жировой ткани, связанное с ожирением и лечение розиглитазоном. J. Clin. Инвестировать. 114, 1281–1289. DOI: 10.1172 / JCI21752
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вудс, Н. М., Катбертсон, К. С., и Кобболд, П. Х. (1986). Повторяющиеся преходящие повышения уровня свободного кальция в цитоплазме в гепатоцитах, стимулированных гормонами. Природа 319, 600–602.DOI: 10.1038 / 319600a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вудс, Н. М., Катбертсон, К. С., и Кобболд, П. Х. (1987). Колебания концентрации свободного кальция в цитоплазме в отдельных гепатоцитах крысы, вызванные агонистами. Cell Calcium 8, 79–100. DOI: 10.1016 / 0143-4160 (87)
-8
CrossRef Полный текст | Google Scholar
Райт, Л. Е., Вечеллио Реане, Д., Милан, Г., Террин, А., Ди Белло, Г., Беллиголи, А. и др. (2017).Повышенный митохондриальный унипортер кальция в адипоцитах лежит в основе митохондриальных изменений, связанных с инсулинорезистентностью. Am. J. Physiol. Метаб. 313, E641 – E650. DOI: 10.1152 / ajpendo.00143.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wu, W., Shen, Q., Lei, Z., Qiu, Z., Li, D., Pei, H., et al. (2019). Кристаллическая структура MICU 2 дает представление о связывании Ca 2+ и образовании гетеродимера MICU 1-MICU 2. Embo Rep. 20: e47488.DOI: 10.15252 / embr.201847488
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wu, Y., Rasmussen, T. P., Koval, O. M., Joiner, M.-L. А., Холл, Д. Д., Чен, Б. и др. (2015). Митохондриальный унипортер контролирует учащение пульса «борьба или бегство». Нат. Commun. 6: 6081. DOI: 10.1038 / ncomms7081
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ямамото Т., Ямагоши Р., Харада К., Кавано М., Минами Н., Идо Ю. и др.(2016). Анализ структуры и функции EMRE в системе экспрессии дрожжей. Biochim. Биофиз. Acta Bioenerg. 1857, 831–839. DOI: 10.1016 / j.bbabio.2016.03.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yoo, J., Wu, M., Yin, Y., Herzik, M.A., Lander, G.C., and Lee, S.-Y. (2018). Крио-ЭМ структура митохондриального унипортера кальция. Наука 361, 506–511. DOI: 10.1126 / science.aar4056
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Влияние температуры на аэробный метаболизм: к механистическому пониманию реакции эктотермов на изменяющуюся окружающую среду | Журнал экспериментальной биологии
Температура оказывает сильное влияние на химические и биохимические реакции; таким образом, понимание механизмов, которые используют организмы, чтобы справиться с тепловыми изменениями, было в центре внимания области биохимической адаптации с момента ее появления (Hochachka, 1965, 1967; Somero et al., 1968; Сомеро и Хочачка, 1969, 1971; Хочачка, Сомеро, 1968, 1973, 2002). За последние 50 лет был достигнут значительный прогресс в понимании биохимических основ термической адаптации (Cossins and Bowler, 1987; Angilletta, 2009), особенно на уровне отдельных белков (Somero, 2004; Fields et al., 2015). . Однако многие вопросы остаются без ответа даже на биохимическом уровне, и нам все еще не хватает полного механистического понимания влияния температуры на биологические процессы на разных уровнях организации и набора адаптаций, которые организмы используют, чтобы справиться с этими эффектами (Somero, 2012 ).Эти вопросы становятся все более важными, потому что изменение климата, вызванное деятельностью человека, изменяет модели средних и экстремальных температур по всему миру, что приводит к изменениям в биогеографическом распределении видов (Parmesan and Yohe, 2003; Perry et al., 2005; Parmesan, 2006). .
Самый простой способ описать влияние температуры на скорость биохимических, физиологических или поведенческих процессов — это построить кривую тепловых характеристик (TPC, рис.1) (Schulte et al., 2011). Многие исследования задокументировали формы TPC на разных уровнях биологической организации и таксонов. Мета-анализ этих данных (Dell et al., 2013) показывает, что TPCs имеют тенденцию быть одномодальными и смещены влево с тремя отдельными областями (Dell et al., 2011, 2013): (1) фаза роста при повышении температуры; (2) фаза плато, которая охватывает тепловой оптимум ( T opt ) для признака; и (3) фаза крутого спада при более высоких температурах.
Рис.1.
Кривая тепловых характеристик (TPC). TPC суммируют влияние температуры на скорость биологических процессов. Критическими особенностями TPC являются крутизна увеличения скорости с температурой, термический оптимум для процесса ( T opt ) и термическая ширина, то есть диапазон температур, в котором скорость процесса максимальна. .
Рис. 1.
Кривая тепловых характеристик (TPC). TPC суммируют влияние температуры на скорость биологических процессов.Критическими особенностями TPC являются крутизна увеличения скорости с температурой, термический оптимум для процесса ( T opt ) и термическая ширина, то есть диапазон температур, в котором скорость процесса максимальна. .
Однако внутри этой общности существует огромное количество различий в форме TPC. Эта изменчивость частично является результатом адаптивных различий между таксонами, а также может быть вызвана нейтральной изменчивостью и различными типами пластичности, включая эпигенетические эффекты, пластичность развития и акклиматизацию (Schulte et al., 2011), а также методологическими проблемами, такими как скорость изменения температуры во время экспериментального определения TPC (Cossins and Bowler, 1987).
На механистическом уровне фаза роста TPC часто описывается как вызванная термодинамическими эффектами, которые можно описать с помощью уравнения Аррениуса (Arrhenius, 1915): (1) где k — скорость реакции, — это предэкспоненциальный множитель (который постоянен при биологически значимых температурах), E a — энергия активации реакции, R — газовая постоянная и T — температура в кельвинах.Уравнение Аррениуса предсказывает, что фаза нарастания TPC должна иметь экспоненциальную форму, если преобладают простые термодинамические эффекты. Преобразование в натуральный логарифм уравнения Аррениуса приводит к следующему: (2) Если построить график ln k против 1/ T в кельвинах (на графике Аррениуса), будет получена прямая линия с наклоном, равным — E a R −1 до температур чуть ниже T opt , если процесс следует уравнению Аррениуса.Когда TPC достигает значения T opt , наклон графика Аррениуса должен иметь значительный разрыв в точке, называемой температурой точки излома Аррениуса (ABT). Если присутствует более одного ABT или данные показывают нелинейную зависимость от температуры, то TPC не следует простым предсказаниям Аррениуса (Cossins and Bowler, 1987).Список сокращений и символов
ABT
Температура излома по Аррениусу
C P
CT макс. м
MR
MMR
MTE
метаболическая теория экологии
OCLTT
Кислород и ограниченная емкость 9128 9111 901 901 901 912 9129 opt
TPC
Кривая тепловых характеристик
V макс.
Δ G ‡
Gib1 энергия активации
Хотя уравнение Аррениуса можно использовать для объяснения формы фазы нарастания TPC, оно не может объяснить наиболее очевидную особенность TPC для биологических процессов: наличие максимума (при T opt ) резким снижением скорости.Это снижение обычно объясняется денатурацией белков при высоких температурах. Однако даже для отдельных белков этот учет представляет собой чрезмерное упрощение процессов, которые формируют TPC, и неясно, достаточно ли этих факторов при рассмотрении сложных биологических характеристик, вызванных взаимодействием многих белков и процессов на нескольких уровнях биологической организации (Prosser, 1973; Cossins and Bowler, 1987; Knies and Kingsolver, 2010).
Хотя наше понимание лежащих в основе биохимических и физиологических механизмов, которые формируют TPC, является неполным, эмпирические данные о формах TPC в настоящее время используются для разработки моделей прогнозирования реакции видов, популяций и сообществ на изменение климата (например,грамм. см. Kordas et al., 2011; Амарасекаре и Сэвидж, 2012; Dell et al., 2014; Гилберт и др., 2014). Превращение этих эмпирически основанных моделей в истинные причинно-следственные механистические модели (Helmuth et al., 2005) потребует понимания ближайших и конечных причин вариаций формы TPC. Таким образом, цель этого обзора — изучить наше текущее понимание лежащих в основе биохимических и физиологических механизмов, которые формируют TPC для аэробного метаболизма в ответ на резкие изменения температуры, чтобы представить перспективы механизмов физиологической пластичности и эволюционной адаптации, которые организмы могут использовать. чтобы справиться с этими эффектами.
Большинство биохимических реакций в значительной степени зависят от температуры, но есть несколько причин сосредоточить внимание на аэробном метаболизме, размышляя о роли, которую температура играет в формировании распределения и численности видов. Аэробный метаболизм настолько важен для животных, что его назвали «огнем жизни» (Kleiber, 1961). Большинство животных зависят от присутствия кислорода, по крайней мере, на некоторых этапах своей жизни, и даже животные, выработавшие стратегии жизни в бескислородных средах обитания, обычно являются лишь факультативными анаэробами (Tielens et al., 2002; Mentel, Martin, 2010), причем полные анаэробы являются редким исключением из этого правила (Kristensen, 2002; Danovaro et al., 2010; Mentel, Martin, 2010). Как и большинство биохимических реакций, реакции аэробного метаболизма напрямую зависят от температуры окружающей среды (Hochachka and Somero, 2002), поэтому у организмов должны быть механизмы, чтобы справиться с этими эффектами, чтобы сохранить выработку энергии при любых температурах. Таким образом, основной причиной, по которой следует сосредоточить внимание на аэробном метаболизме, является его важность в обеспечении энергией, необходимой для жизни животных.
Вторая важная причина для сосредоточения внимания на аэробном метаболизме заключается в том, что недавние теоретические синтезы были сосредоточены на метаболических характеристиках как ключевых участниках реакции организмов на изменение климата. В частности, как метаболическая теория экологии (MTE) (Brown et al., 2004), так и гипотеза термостойкости, ограниченной кислородом и емкостью (OCLTT) (Pörtner, 2001, 2002a, b, 2010; Pörtner and Knust, 2007; Claireaux and Lefrançois, 2007; Pörtner and Farrell, 2008) ставят тепловую зависимость процессов скорости метаболизма в центр своего синтеза.Таким образом, понимание механистической основы тепловой зависимости аэробного метаболизма необходимо для оценки обоснованности этих теорий.
MTE построен на идее о том, что метаболизм обеспечивает механистическую основу для центральной объединяющей теории экологии (Brown et al., 2012), и он был широко принят экологами, заинтересованными в прогнозировании реакции видов на изменение климата (Whitefield , 2004; Vasseur, McCann, 2005; O’Connor et al., 2009, 2011; Диллон и др., 2010). MTE основан на более ранних работах (West et al., 1997; Gillooly et al., 2001), в которых была предпринята попытка предоставить механистическую основу для прогнозирования скорости метаболизма с учетом влияния размера тела и температуры. MTE распространил эти идеи на уровни биологической организации от индивидуальной до популяций, сообществ и экосистем.
Многие из лежащих в основе механистических аспектов MTE, которые основывались на свойствах фрактальных сетей, подвергались критике (Clarke, 2004, 2006; Clarke, Fraser, 2004; Cyr and Walker, 2004; O’Connor et al., 2007; Martínez del Rio, 2008), и в результате MTE можно было бы более точно рассматривать как эмпирическую, а не механистическую теорию (Martínez del Rio, 2008; Price et al., 2012). В частности, MTE делает предположение, что биологические процессы будут соответствовать предсказаниям уравнения Аррениуса с экспоненциальной фазой роста, и исходит из этого предположения, чтобы делать прогнозы о реакции видов на изменение климата. Одна из целей этого обзора — спросить, оправдано ли это предположение.
Гипотеза OCLTT использует другой взгляд на роль аэробных процессов в определении реакции организмов на изменение климата, но при этом подчеркивает их важность. Вместо того, чтобы смотреть на скорость метаболизма как таковую, OCLTT фокусируется на аэробном объеме или разнице между базальным и максимальным потреблением кислорода, которая представляет собой энергию, доступную организму для выполнения важных функций, таких как рост, передвижение и размножение (Pörtner, 2001, 2002a, b; Пёртнер и Кнут, 2007; Клеро и Лефрансуа, 2007; Пёртнер и Фаррелл, 2008).Эта теория параллельна ранним работам Фреда Фрая и его учеников (Fry, 1947; Fry and Hart, 1948; Brett, 1971), которые предположили, что ограничения аэробных возможностей ограничивают способность организма выполнять различные экологические функции. OCLTT, который также связывает аэробный объем с работоспособностью и физической подготовкой всего организма, предоставил дополнительную механистическую основу для этой идеи, учитывая различные основные факторы, которые могут вступать в игру в разных точках кривой аэробного размаха.
Последняя ключевая причина сосредоточения внимания на аэробном метаболизме связана с сравнительными и эволюционными исследованиями вариаций аэробных метаболических характеристик.Например, многочисленные исследования документально подтвердили корреляцию между аэробными метаболическими функциями и градиентами широты и высоты (Angilletta et al., 2002; Fontanillas et al., 2005; Pörtner, 2006; Hassanin et al., 2009; Cheviron and Brumfield, 2009; Scott et al., 2011; Toews et al., 2014; Stager et al., 2014). Аналогичным образом, многие исследования продемонстрировали, что вариации аэробного метаболизма, измеряемые на уровне тканей и всего организма, связаны с различиями в производительности и физической форме или коррелятами физической подготовки (Pörtner and Knust, 2007; Farrell et al., 2008; Eliason et al., 2011; Зера, 2011; White et al., 2012; Rauhamäki et al., 2014). Кроме того, генетическая изменчивость на уровне митохондрий связана с различиями в приспособленности (Rand, 2001; Ballard and Whitlock, 2004; Blier et al., 2013). Вместе эти данные убедительно свидетельствуют о том, что аэробный метаболизм находится под сильным отбором окружающей среды и, вероятно, играет важную роль в формировании реакции видов на изменение климата.
Из-за сложности воздействия температуры на процессы на клеточном и организменном уровнях теории, такие как MTE, обычно предполагают, что TPC на более высоких уровнях биологической организации формируются под влиянием температуры на ключевой базовый процесс на биохимический уровень.Однако даже на уровне белка многое еще предстоит узнать о механизмах, формирующих TPC. Классическая модель для объяснения формы TPC для отдельных белков приписывает форму эффектам Аррениуса при повышении температуры в сочетании с денатурацией белка при высоких температурах. Одна из основных проблем этой модели заключается в том, что денатурация — это процесс, зависящий от времени; чем дольше фермент находится при высокой температуре, тем больше вероятность его денатурирования, и, поскольку денатурация необратима, более длительное время анализа приводит к большей денатурации и более низкому кажущемуся T opt .Другая проблема классической модели заключается в том, что наблюдаемое значение T opt белка может быть значительно ниже температуры денатурации белка, по крайней мере, для некоторых ферментов (Feller and Gerday, 2003). Например, рассмотрим миофибриллярную АТФазу киллифиса Fundulus heteroclitus , который имеет температуру разрыва Аррениуса около 12 ° C (Sidell et al., 1983), но не денатурирует до ~ 40 ° C.
Существуют две связанные модели, которые были предложены для учета недостатков классической модели.«Модель равновесия» предполагает, что ферменты проходят обратимо инактивированную форму перед денатурацией (Petersen et al., 2004; Daniel et al., 2010; Daniel and Danson, 2010). Увеличение доли обратимо инактивированной формы при повышении температуры постепенно сводит на нет увеличение скорости реакции, вызванное термодинамическими эффектами, что приводит к снижению активности фермента и характерной формы TPC. Модель Шарпа – Скулфилда (Sharpe, DeMichele, 1977; Schoolfield et al., 1981) аналогична, но постулирует, что обратимо инактивированные формы встречаются как при высоких, так и при низких температурах. Обе эти модели лучше соответствуют наблюдаемым данным по сравнению с классической моделью. Они также согласуются с предположениями о том, что обратимо денатурированные промежуточные соединения могут объяснять стабилизирующие эффекты осмолитов при высоких температурах (Winzor et al., 1992; Hall et al., 1995; Winzor and Jackson, 2006).
Однако как модель равновесия, так и модель Шарпа – Скулфилда могут представлять чрезмерное упрощение высокодинамичных структур белков (Shukla et al., 2015). Недавние биофизические данные свидетельствуют о том, что белки представляют собой совокупность молекул с большим количеством конформаций, называемых микросостояниями, которые могут различаться по каталитической активности или связыванию субстрата. (Henzler-Wildman, Kern, 2007; Wrabl et al., 2011). Белки могут переключаться между этими состояниями в различных временных масштабах, и вероятность переключения изменяется в зависимости от температуры.Таким образом, вместо того, чтобы моделировать эффекты температуры как изменение доли «нативных» и обратимо или необратимо «денатурированных» белков, более вероятно, что изменения температуры вызывают сдвиги в ландшафте вероятностей, связывающем множество функционально различных микросостояний. Пока эти идеи не будут полностью интегрированы в наше понимание влияния температуры на функцию белков, мы не сможем достичь полностью механистического описания формы TPC на уровне даже одного фермента.
Другая модель, объясняющая влияние температуры на активность белка, теория скорости макромолекул (Hobbs et al., 2013), основана на основах статистической термодинамики и, в частности, на теории переходных состояний (Eyring, 1935). Он сильно отличается от других моделей, поскольку не требует денатурации фермента для учета формы TPC для отдельных белков (Schipper et al., 2014). Чтобы понять теорию, необходимо подумать о стадиях, посредством которых фермент катализирует превращение субстрата в продукт. Во время этого преобразования молекула субстрата проходит через переходное состояние, которое имеет более высокую свободную энергию, чем у субстрата или продукта. Разница в свободной энергии между начальным и переходным состояниями называется энергией Гиббса активации (Δ G ‡ ). Фермент ускоряет скорость реакции, стабилизируя переходное состояние и, таким образом, уменьшая изменение свободной энергии, связанное с образованием переходного состояния, что снижает энергетический барьер активации (ΔG ‡ ).Одно из центральных предположений теории переходного состояния состоит в том, что ΔG ‡ не зависит от температуры. Это предположение хорошо подходит для реакций малых молекул в водных растворах, но реакции, катализируемые ферментами, включают взаимодействия между субстратом и сложным макромолекулярным ферментом, который может претерпевать большие изменения теплоемкости ( C P ) во время катализа. Теория скорости макромолекул использует фундаментальные уравнения статистической термодинамики, чтобы показать, что большая Δ C P , связанная с макромолекулярными реакциями, приводит к температурной зависимости Δ G ‡ (LiCata and Liu, 2011).
Если Δ G ‡ зависит от температуры и особенно если Δ C P большое и отрицательное, то математически легко показать, что формы TPC для ферментов должны иметь четкую форму T opt с последующим снижением активности фермента с повышением температуры без необходимости вызывать денатурацию фермента. Точно так же, когда Δ C P велико и отрицательно, термическая чувствительность будет выше при низких температурах — закономерность, которая не соответствует предсказаниям уравнения Аррениуса, но часто наблюдается для биохимических реакций, в том числе связанных с аэробными реакциями. метаболизм (Хочачка, Гуппи, 1987).Вероятно, что Δ C P варьируется среди ферментов, что приведет к различиям в форме TPC для разных ферментов. Таким образом, эта теория обеспечивает механистическую основу формы TPC и ее вариаций среди белков.
Хотя они предполагают различные механизмы для объяснения влияния температуры на активность белка, все эти модели фокусируются на влиянии температуры на максимальную активность ( V max или k cat ).При В max обеспечивается уровень насыщения субстрата, и скорость реакции в первую очередь ограничивается скоростью превращения субстрата в продукт, а не связыванием субстрата. Но это не отражает биологическую реальность, поскольку концентрации субстрата in vivo низкие. Это наблюдение подчеркивает важность другого критического параметра классической кинетики ферментов — константы Михаэлиса ( k m ), которая является индикатором взаимодействия между ферментом и его субстратом.Константы Михаэлиса также зависят от температуры, при этом повышение температуры обычно вызывает увеличение видимой k m (Somero, 1995; Somero et al., 1996; Hochachka and Somero, 2002). Такое влияние температуры на k m , как полагают, связано с изменениями в конформации белка (то есть сдвигами между микросостояниями белка), которые влияют на способность белка связываться с субстратом и катализировать превращение в продукт.
В целом, организмы из более теплых мест обитания, как правило, имеют ферменты, которые функционируют при более высоких температурах, чем организмы из более холодных мест обитания, и такая адаптация к высокой температуре имеет такую цену, что эти ферменты не работают так же хорошо при низких температурах (Somero, 1995; Винтроде, Арнольд, 2001; Поля, 2001).С механистической точки зрения этот образец обычно интерпретируется как компромисс между гибкостью и стабильностью, так что высокая структурная стабильность, необходимая для выдерживания высоких температур, приводит к снижению гибкости, что ограничивает каталитическую способность при более низких температурах (Fields, 2001). . Интересно, однако, что можно использовать in vitro, направленную эволюцию для «конструирования» ферментов, обладающих как превосходной высокотемпературной стабильностью, так и высокой каталитической активностью при более низких температурах (Wintrode and Arnold, 2001; Arnold et al., 2001; Дин и Торнтон, 2007; Kaltenbach and Tokuriki, 2014), что предполагает, что компромисс между стабильностью и гибкостью не является биофизическим ограничением. Но почему же тогда очевидный компромисс между этими свойствами настолько распространен в природе?
Есть по крайней мере два возможных объяснения этого наблюдения. Во-первых, недостаток природных ферментов с такими свойствами может быть результатом нейтральных процессов. Существует очень мало возможных структур, которые обеспечивают как стабильность при высоких температурах, так и хорошую каталитическую активность при низких температурах, поэтому большинство мутаций будет иметь тенденцию давать ферменты, лишенные этих двойных свойств (Wintrode and Arnold, 2001; Harms and Thornton, 2013).В отсутствие строгого отбора по обоим свойствам случайная мутация будет иметь тенденцию удалять эти ферменты из популяции. В качестве альтернативы могут существовать функциональные компромиссы, связанные с поддержанием как стабильности, так и гибкости, которые не полностью учитываются в лабораторных экспериментах по эволюции. Например, белки, полученные в лаборатории, которые стабильны при высоких температурах и активны при низких температурах, часто имеют очень низкие значения k m по сравнению с белками, полученными естественным путем (Wintrode and Arnold, 2001; Arnold et al., 2001). Возможно, что эти изменения в свойствах связывания субстрата вредны в естественной окружающей среде, что объясняет отсутствие белков, полученных естественным путем, как с высокой термостабильностью, так и с высокой каталитической активностью при низких температурах.
Существует также значительная вариация среди белков в восходящем наклоне TPC, но менее ясно, имеет ли эта вариация постоянная связь с местом обитания.Недавний мета-анализ восходящего наклона TPC для ферментов от бактерий, адаптированных к холоду, умеренным и высоким температурам, не смог найти какой-либо устойчивой связи между этими уклонами и температурой среды обитания (Elias et al., 2014), хотя исследования на других организмах показали сделал это (Dahlhoff et al., 1991; Feller and Gerday, 2003). Точно так же эмпирические данные, сравнивающие стенотермы и эвритермы, иногда (например, Hardewig et al., 1999), но не всегда (Swimmer et al., 2004), выявляли пониженный наклон ферментов из эвритермических организмов.Таким образом, трудно сделать вывод, что отбор сильно влияет на изменение восходящего наклона TPCs для белков.
Десятилетия элегантных исследований с использованием как внутривидовых (например, Place and Powers, 1979), так и межвидовых сравнений (например, Dahlhoff and Somero, 1993; Somero et al., 1996) убедительно доказывают, что k m демонстрирует термический приспособление. Например, многочисленные исследования показали, что значения k m ферментов эвритермических организмов имеют тенденцию быть менее термически зависимыми, чем значения ферментов стенотермальных организмов.Точно так же существует общая закономерность сохранения k m , так что значения k m гомологичных ферментов организмов из разных местообитаний схожи при оценке при соответствующей рабочей температуре для данного вида (Somero, 1995). ; Поля, 2001). Таким образом, любое всестороннее рассмотрение влияния температуры на биологические процессы должно учитывать не только влияние температуры на активность фермента, но также влияние температуры на взаимодействия фермент-субстрат.С точки зрения теории скорости макромолекул, это могло бы включать включение термина, улавливающего эффекты большого Δ C P , которые возникают во время связывания субстрата, что еще предстоит включить в теорию.
Вышеупомянутое обсуждение было сосредоточено в первую очередь на ферментах, которые действуют в водной среде клетки, но многие из основных ферментов, участвующих в аэробном метаболизме, встроены в митохондриальную мембрану.На свойства мембран, такие как текучесть, сильно влияет температура (Cossins, 1983; Hazel, 1995), и эти изменения текучести мембран могут иметь прямое влияние на активность встроенных в мембрану белков (Somero, 2011). В самом деле, по крайней мере в некоторых случаях нарушение липид-белковых взаимодействий было предположено как причина вызванного высокой температурой отказа митохондриального дыхания (например, O’Brien et al., 1991). Взаимодействия между белками и их мембранным окружением могут приводить к сложной форме TPC этих белков и могут влиять на такие свойства, как k m .Это наблюдение указывает на важность локальной среды вокруг белка и является предостережением как для белков, встроенных в мембрану, так и для цитозольных белков. Это также указывает на более общее предостережение, связанное с измерениями TPC на изолированных ферментах, поскольку измеренные активности in vitro часто не совпадают с измеренными in vivo (Wright et al., 1992; Teusink et al., 2000).
Очевидно, что еще многое предстоит узнать о факторах, формирующих TPC, даже для отдельных белков, особенно тех, которые участвуют в аэробном метаболизме, а понимание механизмов еще менее развито на более высоких уровнях биологической организации, таких как сложные биохимические сети и интактные клетки.Были проведены некоторые измерения влияния температуры на аэробный метаболизм на клеточном уровне у животных (Somero and DeVries, 1967; Hoskins and Aleksiuk, 1973; Jorjani and Ozturk, 1999), и некоторые наборы данных хорошо согласуются с уравнением Аррениуса, а другие не надо. Аналогичные закономерности наблюдаются при измерении потребления кислорода изолированными митохондриями (van den Thillart and Modderkolk, 1978; Somero et al., 1996; Weinstein and Somero, 1998; Hardewig et al., 1999; Abele et al., 2002; Somero, 2002). ; Джонстон и др., 1994; Fangue et al., 2009; Hilton et al., 2010; Ифтикар и др., 2010, 2014; Ифтикар и Хикки, 2013). Например, на рис. 2 показаны ТРС потребления кислорода изолированными митохондриями из F. heteroclitus , анализируемые в условиях насыщения субстрата. В этом случае данные хорошо согласуются с предсказаниями уравнения Аррениуса, но акклиматизация к низким температурам может привести к существенным разрывам в TPC (Fangue et al., 2009).
Рис.2.
TPC для скорости потребления кислорода митохондриями эвритермального киллифа Fundulus heteroclitus . Максимальные скорости АДФ-фосфорилирования потребления кислорода (состояние III) с пируватом в качестве субстрата. Митохондрии выделяли из печени рыб, акклиматизированных к 25 ° C (данные Fangue et al., 2009). Значения являются средними ± стандартное отклонение.
Рис. 2.
TPC для скорости потребления кислорода митохондриями эвритермального киллифа Fundulus heteroclitus . Максимальные скорости АДФ-фосфорилирования потребления кислорода (состояние III) с пируватом в качестве субстрата. Митохондрии выделяли из печени рыб, акклиматизированных к 25 ° C (данные Fangue et al., 2009). Значения являются средними ± стандартное отклонение.
Данные, которые соответствуют уравнению Аррениуса, часто объясняются предположением, что форма TPC для аэробного метаболизма на клеточном уровне определяется тепловой зависимостью одного основного процесса, например, лимитирующего шага в биохимической сети. (Corkrey et al., 2012). Точно так же случаи, в которых в TPC обнаруживаются значительные разрывы (ABT) ниже T opt , часто приписывают эффектам температурно-индуцированных изменений в структуре мембраны, которые влияют на функцию митохондриальной цепи переноса электронов (O ‘ Brien et al., 1991), и поэтому эти объяснения также имеют тенденцию сосредотачиваться на отдельных ключевых процессах и их взаимодействии с клеточной средой. Однако неясно, оправдано ли предположение, что один ключевой процесс определяет форму TPC для аэробного метаболизма на клеточном уровне.
Аэробный метаболизм — это многоступенчатый процесс, включающий сложные биохимические сети как в цитоплазме, так и в митохондриях. Контроль потока через такие биохимические сети, вероятно, будет определяться на нескольких этапах, а не с помощью одного процесса «ограничения скорости» (Kacser and Burns, 1973; Fell, 1997; Bruggeman and Westerhoff, 2007). Почему тогда аэробный метаболизм в целом когда-либо демонстрирует поведение, подобное Аррениусу, в ответ на повышение температуры, а не всегда демонстрирует прерывания? Возможно, что все этапы управления потоком имеют сходные тепловые зависимости в данном организме (Chaui-Berlinck et al., 2004). В поддержку этой идеи анализ анаэробного гликолиза в дрожжах ( Saccharomyces cerevisiae ) предполагает, что активности всех гликолитических ферментов у этого вида имеют сходные температурные зависимости (Cruz et al., 2012). Однако другие исследования дали другие результаты (Postmus et al., 2012). Еще меньше информации доступно для ферментов, участвующих в аэробном метаболизме, но, по крайней мере, в некоторых исследованиях наблюдались различия в наклоне TPC для разных митохондриальных ферментов (Lenaz et al., 1972; Водтке, 1976; Ирвинг и Ватсон, 1976). Даже если отдельные ферменты в биохимической сети имеют разные тепловые зависимости, все равно можно будет наблюдать относительно плавную TPC для процесса в целом, если большое количество этапов совместно контролируют весь процесс и нет основной температуры. промежутки между оптимумами лежащих в основе кривых. На данный момент у нас просто нет достаточных данных о характеристиках биохимических сетей, чтобы различать эти возможности.
Анализ, обсужденный выше, был попыткой смоделировать системные свойства биологических сетей с использованием данных об активности белков, измеренных in vitro . In vivo поток через метаболические пути чаще всего контролируется изменением концентраций субстратов, продуктов или аллостерических эффекторов, что называется «метаболической регуляцией» (ter Kuile and Westerhoff, 2001; Suarez and Moyes, 2012).В самом деле, исследования тепловой зависимости гликолитического потока у дрожжей показывают, что метаболическая регуляция является доминирующей силой в определении реакции метаболического потока на повышение температуры (Postmus et al., 2008). Рассмотрение этой точки зрения на метаболический контроль предполагает, что невозможно разработать всеобъемлющее механистическое понимание того, как аэробный метаболизм реагирует на температуру, просто рассматривая влияние температуры на отдельные белки без учета роли субстратов, продуктов и аллостерических эффекторов, а также того, как на них влияет температура.Понимание этого процесса потребует дополнительных эмпирических исследований влияния температуры на метаболизм, активность ферментов и метаболиты на клеточном уровне.
Технологические достижения теперь позволяют использовать высокопроизводительные методы, позволяющие одновременно оценивать несколько биологических процессов. Например, этот подход был применен к влиянию температуры на рост Escherichia coli (Chang et al., 2013), определяя синтез кофактора как наиболее лимитирующий процесс при высокой температуре. Такие подходы «системной биологии» в сочетании с аналитическими методами, такими как иерархический анализ (ter Kuile and Westerhof, 2001), обещают распутать механизмы, посредством которых температура влияет на сложные метаболические сети и неповрежденные клетки (Bordbar et al., 2014). Однако очень мало исследований изучали влияние температуры на этом уровне биологической организации, и эта область представляет собой плодотворное направление для будущих исследований как острого воздействия температуры на аэробный метаболизм, так и реакции, которую клетки используют, чтобы справиться с этим. эффекты.
TPC для аэробного метаболизма на уровне организма были охарактеризованы для многих видов (например, Clarke and Johnston, 1999; Sokolova and Pörtner, 2003; Giomi and Pörtner, 2013). На рис. 3 показан пример стандартной скорости метаболизма (RMR) высокоэвритермального киллифа, Fundulus heteroclitus (Healy and Schulte, 2012). Этот TPC соответствует общей форме TPC, показанной на рис.1, но в этом случае фазу спада кривой невозможно измерить, потому что рыба достигает своего критического теплового максимума (CT max ) при температурах лишь немного выше, чем показано здесь (Fangue et al., 2006). На основании наблюдения, что прокариоты и одноклеточные эукариоты могут переносить значительно более высокие температуры, чем те, которые переносятся многоклеточными животными, было высказано мнение, что уровни организации выше, чем у одиночной клетки, должны иметь решающее значение для определения верхних температурных пределов интактных животных (Pörtner, 2002a). ; Сторч и др., 2014). Среди предложенных механизмов — отказ нервных процессов системного уровня, таких как регуляция вентиляции и кровообращения мозгом (Lagerspetz, 1974), отказ нервных процессов на клеточном уровне, таких как проводимость потенциала действия или синаптическая передача (Gladwell et al., 1976; Cossins et al., 1977; Rosenthal and Bezanilla, 2000; Robertson, 2004; Miller and Stillman, 2012), неспособность поддерживать мышечный мембранный потенциал (Hosler et al., 2000), нарушение связанного с мембраной процессы в более общем плане (Hulbert, 2003), нарушение сердечной функции (Pörtner and Farrell, 2008; Eliason et al., 2011) и сбой на уровне митохондрий (Pörtner et al., 2000; Pörtner, 2002a, b; Iftikar, Hickey, 2013).
Рис. 3.
TPC для рутинного аэробного метаболизма всего животного в Fundulus heteroclitus . Обычная скорость метаболизма (RMR) — это скорость потребления кислорода, когда рыба спокойно отдыхает в респирометре, но не обязательно при минимальном уровне активности (данные Healy and Schulte, 2012).Значения представляют собой средние значения ± s.e.m.
Рис. 3.
TPC для рутинного аэробного метаболизма всего животного в Fundulus heteroclitus . Обычная скорость метаболизма (RMR) — это скорость потребления кислорода, когда рыба спокойно отдыхает в респирометре, но не обязательно при минимальном уровне активности (данные Healy and Schulte, 2012). Значения представляют собой средние значения ± s.e.m.
Тем не менее, сложные взаимозависимости аэробного метаболизма затрудняют определение точного уровня биологической организации, на которой происходит сбой, и предполагают, что вероятно участие комбинации факторов.Например, сердечная или нервная недостаточность на биохимическом или клеточном уровне может привести к нарушению подачи кислорода к тканям, что ограничит скорость аэробного метаболизма на уровне всего организма. Точно так же отказ ключевых контролирующих процессов в головном мозге может вызвать снижение сердечной или вентиляционной способности, ограничивая поступление кислорода и вызывая митохондриальную недостаточность и коллапс аэробного метаболизма. Таким образом, аэробный метаболизм может дать сбой в ответ на резкие высокие температуры из-за сбоя любого из процессов, участвующих в установлении либо предложения, либо потребности в кислороде на нескольких уровнях организации, а цикл положительной обратной связи между этими процессами может привести к быстрой и катастрофической системе. неудача при резком воздействии высоких температур.
Второй важной особенностью TPC для аэробного метаболизма на уровне организма является форма кривой при повышении потребления кислорода до T opt (рис. 3). Наклон этой фазы подъема различается у разных видов (Dell et al., 2011) и, возможно, среди признаков (Amarasekare and Savage, 2012), но вопрос о том, происходит ли это изменение согласованным образом между организмами, адаптированными к разным температурам, остается вопросом дискуссии более 50 лет (Scholander et al., 1953; Рао и Баллок, 1954; Кларк и Джонстон, 1999). Проверка предсказания Аррениуса экспоненциальной фазы роста имеет важное значение, потому что теории, такие как MTE, предполагают, что метаболизм может быть смоделирован на основе уравнения Аррениуса. К сожалению, проверить это предсказание непросто, поскольку неясно, действительно ли измерения скорости метаболизма на уровне организма имеют достаточное разрешение, чтобы различать альтернативные модели формы этого наклона. Например, используя обширный набор данных по скорости метаболизма костистых рыб, Кларк и Джонстон (1999) показали, что взаимосвязь между скоростью метаболизма в состоянии покоя и температурой может быть одинаково хорошо описана с помощью нескольких различных аппроксимаций кривой.Аналогичным образом, некоторые исследования продемонстрировали закономерности в тепловой зависимости аэробного метаболизма, которые могут быть адекватно описаны уравнением Аррениуса (Алексюк, 1971; Стаму и др., 1995; Ли и др., 2003; Кларк и Джонстон, 1999; Дальхофф и др.) Somero, 1993), в то время как другие нет (Giomi and Pörtner, 2013; некоторые классические примеры для рыб см. Fry and Hart, 1948; Beamish, 1964). В целом, эмпирические данные показывают, что ТПК для аэробного метаболизма на уровне организма могут, но не всегда, соответствовать предсказаниям уравнения Аррениуса, и что могут быть различия в этих формах между стандартной и максимальной скоростью метаболизма (Pörtner, 2002a). , б).На уровне организма существуют сложные гомеостатические механизмы, вовлекающие нервную и эндокринную системы, которые могут влиять на формирование реакции скорости метаболизма на повышение температуры, что может изменить форму TPC в различных временных масштабах (Schulte et al., 2011). Таким образом, имеющиеся данные не соответствуют предположениям таких моделей, как MTE. Это наблюдение поднимает более общий вопрос о том, действительно ли мы ожидаем, что сложный процесс на уровне организма, такой как аэробный метаболизм, будет соответствовать предсказаниям теории, которая была создана для объяснения поведения отдельных химических реакций.Фактически, распространение теории, разработанной для индивидуальных реакций, даже на уровень клеток или биохимических сетей, может быть проблематичным.
Хотя теории, такие как MTE и OCLTT, используют влияние температуры на аэробный метаболизм как фундаментальный строительный блок своих прогнозов о воздействии изменения климата, это дает лишь частичную картину влияния температуры на метаболизм.Потребление кислорода является хорошим показателем потока через митохондриальную цепь переноса электронов, но не обязательно является хорошим показателем производства АТФ при разных температурах из-за влияния температуры на протекание протонов через внутреннюю мембрану митохондрий. Утечка протонов позволяет протонам перемещаться через внутреннюю мембрану митохондрий по путям, отличным от АТФ-синтазы, тем самым рассеивая движущую силу протона и стимулируя потребление кислорода без образования АТФ. Значительный объем данных свидетельствует о том, что термочувствительность генерации АТФ и утечки протонов различается (Hardewig et al., 1999; Hilton et al., 2010; Ифтикар и др., 2010, 2014; Iftikar and Hickey, 2013) из-за увеличения проницаемости для протонов из-за изменений текучести мембран при более высоких температурах (Seebacher et al., 2010; Zukiene et al., 2010). В результате утечка протонов представляет собой увеличивающуюся долю потребления кислорода при повышении температуры (Hilton et al., 2010; Seebacher et al., 2010; Iftikar and Hickey, 2013), вызывая снижение эффективности производства митохондриального АТФ с повышением температуры.Эти эффекты учитываются в OCLTT, но в основном игнорируются MTE.
Также важно учитывать потенциально вредные побочные эффекты митохондриального дыхания. Поток через цепь переноса электронов приводит к образованию активных форм кислорода (АФК), которые могут повредить клеточные макромолекулы, такие как липиды, белки и ДНК (Murphy, 2009; Tomanek, 2015). АФК также действуют как мощные сигнальные молекулы внутри клетки (Finkel, 2011) и, таким образом, могут играть роль в регуляции различных клеточных процессов.Производство АФК митохондриями увеличивается с повышением температуры (Abele et al., 2002), что может влиять на реакцию организмов на изменение температуры (Heise et al., 2006; Blier et al., 2013).
Сосредоточение внимания на аэробном метаболизме с точки зрения потребления кислорода также имеет тенденцию подчеркивать важность снабжения энергией и потенциально недооценивает роль путей, вовлеченных в потребность в энергии (Hofmeyr and Cornish-Bowden, 2000).Анализ метаболического контроля предполагает, что большая часть контроля над потоком через пути доставки АТФ находится в потребности клетки в АТФ (Koebmann et al., 2002), а процессы спроса на АТФ зависят от температуры. Если процессы спроса более термочувствительны, чем процессы, участвующие в энергоснабжении, то изменения в активности этих процессов спроса будут формировать TPC для аэробного метаболизма. Таким образом, полное понимание реакции организмов на изменение климата требует понимания влияния температуры как на энергоснабжение, так и на спрос на энергию.
Концепция OCLTT явно решает эту проблему согласования спроса и предложения, выдвигая гипотезу о том, что в конечном итоге несоответствие между потребностью тканей в кислороде и способностью поставлять кислород сердечно-сосудистой системой ограничивает функцию при высоких температурах (Pörtner and Knust , 2007). OCLTT предполагает, что это несоответствие между спросом и предложением наиболее ярко проявляется в потере аэробных возможностей при высоких температурах, и что это снижение аэробных возможностей напрямую и причинно связано со снижением таких характеристик производительности, как рост и репродуктивный успех.Эта связь была убедительно продемонстрирована для ряда видов рыб и морских беспозвоночных (Pörtner, 2001, 2010; Eliason et al., 2011). Однако существует ограниченная поддержка аэробных возможностей, ограничивающих максимальную термостойкость у эктотерм, дышащих воздухом (Fobian et al., 2014), и особенно у таксонов, таких как насекомые (Klok et al., 2004; Stevens et al., 2010). Кроме того, недавние исследования различных видов рыб показали, что, по крайней мере, у некоторых видов, аэробный диапазон все еще может быть высоким при температурах, при которых нарушается рост и размножение, что позволяет предположить, что тепловые ограничения аэробной способности не могут быть прямой причиной. ограничений приспособленности у этих видов (Healy and Schulte, 2012; Clark et al., 2013а; Gräns et al., 2014; Норин и др., 2014). В результате этих расходящихся наблюдений в настоящее время в литературе ведутся очень оживленные дискуссии относительно актуальности OCLTT для прогнозирования реакции видов на изменение климата (Clark et al., 2013a, b; Pörtner and Giomi, 2013 ; Фаррелл, 2013).
Многие наборы данных, к которым успешно применялась OCLTT, имеют TPC для стандартной скорости метаболизма (SMR) и максимальной скорости метаболизма (MMR) формы, показанной на рис.4А. В этих TPC MMR достигает предела при более низкой температуре, чем SMR, что приводит к снижению аэробного объема при относительно низкой температуре (рис. 4B). Однако возможны и другие схемы. На рис. 4C показана общая форма TPC для SMR и MMR на основе данных для F. heteroclitus (Healy and Schulte, 2012). В этом случае как SMR, так и MMR увеличиваются экспоненциально, по существу параллельно, до температур, близких к максимальной температуре, которую может переносить (даже остро) организмом, что приводит к широкой кривой аэробного размаха (рис.4D). Сравнение двух типов кривых аэробного прицела, показанных на рис. 4, служит напоминанием о том, что аэробный прицел является составной чертой, возникающей в результате влияния температуры на SMR и MMR, и эти показатели, вероятно, будут находиться при различных ограничениях и реагировать независимо. к выбору.
Рис. 4.
Два основных типа аэробной кривой размаха у рыб. (A) RMR (обычная скорость метаболизма) и MMR (максимальная скорость метаболизма), где MMR, скорее всего, ограничивается подачей кислорода при умеренных температурах.(B) Кривая аэробного охвата (MMR-RMR) для данных в A. (C) RMR и MMR, где нет доказательств того, что MMR ограничивается подачей кислорода, за исключением экстремальных температур. (D) Кривая аэробного размаха (MMR – RMR) для данных в C.
Рис. 4.
Два основных типа кривой аэробного размаха у рыб. (A) RMR (обычная скорость метаболизма) и MMR (максимальная скорость метаболизма), где MMR, скорее всего, ограничивается подачей кислорода при умеренных температурах. (B) Кривая аэробного объема (MMR-RMR) для данных в A.(C) RMR и MMR, если нет доказательств того, что MMR ограничивается подачей кислорода, за исключением экстремальных температур. (D) Кривая аэробного объема (MMR – RMR) для данных в C.
У видов с кривыми аэробного размаха, аналогичными показанным на рис. 4D, мало доказательств ограничения подачи кислорода при повышении температуры, а также нет четкой связи между положением кривой аэробного размаха и температурами, которые максимизируют физическую форму. -связанные черты, такие как рост (Healy and Schulte, 2012; Donelson et al., 2014; Хан и др., 2014; Норин и др., 2014; Gräns et al., 2014). Напротив, у этих видов рост и развитие имеют тепловые оптимумы значительно ниже верхнего края окна аэробного прицела T opt . Например, наша работа над F. heteroclitus показывает, что при температурах, когда и RMR, и MMR все еще экспоненциально растут с температурой, а аэробный объем максимален, рыбам трудно поддерживать массу тела во время длительной акклиматизации (Healy and Schulte, 2012).Эти данные указывают на ограничения способности поглощать, обрабатывать или ассимилировать достаточное количество питательных веществ для поддержания высокой скорости метаболизма, типичной для высоких температур. Подобные несоответствия между скоростью усвоения питательных веществ и скоростью метаболизма при повышении температуры наблюдались у различных видов (Lemoine and Burkepile, 2012; Rall et al., 2010; Alcaraz et al., 2014), что свидетельствует о наличии и несоответствие спроса, которое не обязательно отражается в потере общего аэробного объема.Это означает, что у этих организмов может быть затруднительно установить прямую связь между кривой аэробного размаха и физической подготовкой.
Аналогичное разъединение происходит при попытке связать закономерности, полученные из кривой аэробного прицела, и другие особенности жизненного цикла. Например, тепловое окно для воспроизводства часто намного уже, чем тепловое окно для активности (Fry, 1971), и, таким образом, температура, при которой максимальный аэробный объем, достигаемый во время тренировки, начинает снижаться, не всегда может использоваться в качестве показателя температур, которые компромисс фитнеса.Точно так же отказ сложных процессов на клеточном уровне, таких как клеточный цикл, происходит при умеренных температурах (van der Have, 2002), что может быть причиной того, что окна толерантности для развития уже, чем окна толерантности взрослых. В конечном итоге, форма TPC для аэробного метаболизма на уровне организма, вероятно, будет зависеть от процессов спроса и предложения, действующих на нескольких уровнях организации, и должна быть продуктом ряда « решений » распределения энергии, принимаемых организмом в условиях сложные комбинации биотических и абиотических факторов, характеризующие природную среду.Многие из этих вопросов рассматриваются в OCLTT, но связь между этими решениями о распределении энергии и аэробными масштабами не всегда проясняется в рамках теории или неправильно интерпретируется теми, кто пытается ее применить. Таким образом, основная дискуссия вокруг OCLTT сосредоточена не на механистических основах теории, а скорее на предположении, что аэробный объем является адекватным предиктором этих проблем. Фактически, различные концепции того, что подразумевается под термином аэробный прицел, могут лежать в основе некоторых дискуссий по поводу этой концепции (Clark et al., 2013а, б; Пёртнер и Джоми, 2013; Фаррелл, 2013). Разнообразие моделей, показанных на рис. 4, указывает на то, что наиболее подходящим является широкий взгляд на метаболизм, который включает в себя множество факторов, которые могут влиять на предложение и спрос на энергию в разных временных масштабах (например, Соколова и др., 2012; Соколова, 2013). способ рассмотрения взаимосвязи между аэробным метаболизмом и температурой на уровне организма.
Клеточные клейма выявляют ограниченный аэробный метаболизм на пределе температур
[Примечание редакции: ответы автора на первый раунд рецензирования приводятся ниже.]
Внимательно прочитав комментарии и предложения рецензентов, мы провели эксперименты и изменили рукопись, чтобы полностью решить проблемы, поднятые тремя рецензентами, как подробно описано в сопроводительном письме с опровержением. В частности, теперь была рассмотрена первая критика, высказанная рецензентами №1 и №3 относительно отсутствия прямых доказательств ограничения аэробного метаболизма на предельных температурах. Как показано на новом рисунке 4A, измерения дыхания показывают, что аэробный метаболизм действительно снижается как за нижний, так и за верхний температурные пределы.Более того, мы также обнаружили, что эмбрионы, лишенные отдельных компонентов митохондриальной дыхательной цепи, меньше подвержены влиянию температурных пределов, чем температурный диапазон (новый рисунок 4B-D). Благодаря этим новым открытиям наша работа впервые предоставляет убедительные доказательства того, что ограниченный аэробный метаболизм является ключевой особенностью температурных ограничений в организме наземных многоклеточных животных. Более того, мы изменили текст, чтобы сделать его более доступным для более широкой аудитории, уточнив представление и обсуждение работы, тем самым устраняя вторую главную критику рецензентов.
Мы отправляем вам полные комментарии всех трех рецензентов. В общем, все согласны с тем, что работа очень интересная. Однако у них есть два ключевых критических замечания. Во-первых, в работе конкретно не рассматривается механистическая связь между изменениями размера / формы эмбриона, тепловыми эффектами и аэробным метаболизмом. Во-вторых, запутывается изложение и обсуждение работы. Обсуждая, отклонять ли рукопись или поощрять ее пересмотр, рецензенты пришли к выводу, что для получения данных, которые позволят отреагировать на первую критику, потребуется более трех месяцев.Однако, если у вас уже есть какой-то дополнительный механизм, решающий первый пункт, мы, конечно, рассмотрим опровержение.
Рецензент № 1 :
Резюме : В этой статье исследуется, как эмбрионы двух родственных видов нематод реагируют на изменения температуры. Авторы исследуют 35 особенностей одноклеточных зигот в ответ на температуру и определяют те особенности, которые ведут себя по-разному в температурных пределах животных. В частности, они обнаружили, что эмбрионы имеют тенденцию увеличивать отношение поверхности к объему на предельных температурах.Предыдущие теоретические исследования показали, что изменения аэробной способности лежат в основе теплового стресса. В соответствии с этой гипотезой авторы обнаружили, что истощение ключевых метаболических ферментов также вызывает увеличение отношения поверхности к объему. Основываясь на этой корреляции, авторы делают вывод, что «аэробный метаболизм ограничен как нижними, так и верхними тепловыми пределами».
Критика :
1) Основной тезис данной работы основан на корреляции. Нет прямых доказательств того, что аэробный метаболизм ограничивается температурными пределами.Казалось бы, можно провести простые эксперименты, чтобы более прямо продемонстрировать эту гипотезу. Например, становятся ли эмбрионы, истощенные по ключевым ферментам аэробного метаболизма, гиперчувствительностью к изменениям температуры? Без дополнительных экспериментов, которые проверяют гипотезу напрямую, данные слишком предварительные .
Мы благодарим рецензента за то, что он поднял этот очень важный вопрос. Действительно, в нашем первоначальном представлении отсутствовали прямые доказательства того, что аэробный метаболизм ограничен температурными пределами.Эта критическая проблема теперь решена путем измерения скорости дыхания эмбрионов C. elegans дикого типа от 9 ° C до 28 ° C. Как мы теперь показываем на рис. 4A, этот новый эксперимент показал, что дыхание экспоненциально увеличивается в пределах теплового диапазона, как и предсказывается кинетикой типа Аррениуса (Arrhenius, 1915). Кроме того, мы обнаружили, что дыхание снижается как ниже нижнего, так и выше верхнего теплового предела. Эти результаты показывают, что аэробная способность действительно снижается за пределами обоих тепловых пределов по сравнению с тепловым диапазоном.
Важно также, что, как было предложено рецензентом, мы дополнительно оспорили наши выводы, теперь измеряя жизнеспособность эмбрионов в широком диапазоне температур при истощении ключевых белков аэробного метаболизма (субъединицы комплекса I, III и V митохондриальной дыхательной цепи). Мы рассудили, что если аэробная способность используется в меньшей степени для производства энергии за пределами обоих тепловых пределов, чем в пределах теплового диапазона, то нарушение митохондриальной активности также должно иметь меньшее влияние на тепловых пределах, чем в тепловом диапазоне.Примечательно, что мы действительно обнаружили, что эмбриональная летальность была снижена до температурных пределов по сравнению с температурным диапазоном у эмбрионов atp-2 (RNAi), cyc-1 (RNAi) и nuo-1 (RNAi) (Рисунок 4B -D). Эти результаты предлагают сильную экспериментальную поддержку представлению о том, что способность митохондриальной дыхательной цепи действительно снижена за пределы обоих температурных пределов.
Все эти критические открытия теперь представлены и обсуждаются в нашей рукописи в разделе «Результаты», озаглавленном «Клеточные признаки температурных пределов повторяются при нарушении аэробного метаболизма» .
2) Представление данных часто сбивает с толку. Например, Рисунок 1 сообщает о размере эмбриона (вы имеете в виду объем?) И форме эмбриона (длина / ширина). Отношение площади поверхности к объему, которое в Обсуждении представляется наиболее важным параметром, на рисунке не представлено. В результате очень сложно оценить значимость различий / выводов .
Приносим извинения за недостаточно ясное представление данных.Мы приложили большие усилия, чтобы полностью переписать рукопись, чтобы данные были представлены более четко. На рис. 2-3 мы указываем площадь эмбриона, которую мы используем в качестве показателя размера эмбриона с учетом симметрии вращения эмбрионов C. elegans (см. Материалы и методы). В предыдущей версии рукописи отношения площади к объему не указывались, потому что они были просто оценены, а не измерены, как описано в Материалах и методах первоначального представления. В новой версии рукописи мы уменьшаем важность отношения поверхности к объему и вместо этого сосредотачиваемся на характерных клеточных изменениях в температурных пределах, а именно на уменьшении размера эмбриона ниже нижнего теплового предела и удлинении эмбриона выше верхнего теплового предела. оба из которых были измерены напрямую.См. Раздел «Результаты», озаглавленный «Клеточные признаки температурных пределов резюмируются при нарушении аэробного метаболизма».
3) Кажется, что данные не всегда полностью объясняются в тексте. Авторы заявляют, что размер эмбриона уменьшается ниже нижнего теплового предела (от 12 до 10 градусов), но на самом деле размер эмбриона резко увеличивается (увеличение на 25%) между 14 и 12 градусами, а затем незначительно снижается между 12 и 10 градусами ( Рис. 2E ). Это поведение вверх-вниз не обсуждается .
Мы благодарим рецензента за поднятие этого важного вопроса. Как мы теперь объясним лучше (см. Введение, третий абзац и раздел результатов «Размер и форма зародыша чувствительны к температурным пределам » ), тот факт, что размер тела меньше при более высоких температурах в пределах теплового диапазона, является обычным среди эктотермов и имеет было придумано правило размера и температуры (Atkinson, 1994; Forster et al., 2012). Такое уменьшение размера с повышением температуры считается адаптивной реакцией на сохранение аэробной способности за счет увеличения отношения поверхности к объему и, таким образом, потенциального поступления кислорода за счет облегченной диффузии (Atkinson et al., 2006).
В предыдущей версии рукописи мы сосредоточили наше внимание исключительно на обращении этого правила «температура-размер» ниже нижнего теплового предела, о котором ранее сообщалось только у протистов и у Drosophila (Karan et al., 1998 ; Atkinson et al., 2003), и предположили, что это обусловлено ограниченным аэробным метаболизмом ниже нижнего теплового предела. Новая версия рукописи сохраняет этот фокус, помещая его в контекст правила «температура-размер», описанного выше, тем самым проясняя то, что могло проявиться как неожиданное поведение на нижнем тепловом пределе.
4) Авторы не описывают, как эмбрионы были помещены на предметные стекла для визуализации. Если эмбрионы были помещены под покровное стекло, давление покровного стекла могло повлиять на их размер и / или форму. В таком случае различия между различными условиями могут отражать различия в сопротивлении яичной скорлупы давлению покровного стекла, а не фактические различия в размере .
Этот момент действительно заслуживает дальнейшего пояснения. Как мы теперь объясняем в первом абзаце материалов и методов, эмбрионы препарировали в среде 1x M9, закаленной при температуре культивирования, помещали на предметные стекла, помещали под покровное стекло и отображали с помощью покадровой ДИК-микроскопии.Принимая во внимание переполненную компрессионную среду матки у интактного животного и, кроме того, что для всех образцов применялась одна и та же процедура монтажа при всех температурах, мы предполагаем, что наблюдаемые изменения в тепловом отклике размера и формы эмбриона при заданных температурах не являются следствием к порядку монтажа.
Рецензент № 2 :
Это интересная статья, посвященная наблюдениям за функциональными изменениями ограничения температуры у эмбрионов Caenorhabditis elegans . Этот нетрадиционный способ решения этой проблемы будет стимулировать сообщество биологов животных. Тем не менее, текст следует изменить и сократить жаргон, чтобы сделать его более доступным. Некоторые проблемы могут быть обычно выражены таким образом в мире нематод, но заслуживают объяснения, например почему и как отношение поверхности к объему связано с аэробным метаболизмом?
Мы благодарим рецензента за признание нашего нетрадиционного подхода к решению этой проблемы, а также считаем, что наши результаты будут стимулировать сообщество биологов животных в целом.
Это написание во введении немного неточно: биогеографический диапазон относится к тепловому окну вида, то есть к верхнему и нижнему пределу. Этот основной тепловой диапазон используется не полностью, но составляет основу используемого диапазона .
Мы согласны с рецензентом в том, что наш текст был несколько неточным, и теперь четко упоминаем во введении, что другие факторы также определяют фактический эксплуатируемый диапазон:
«Частично в результате организмы имеют тенденцию распространяться в океане и на суше в зависимости от широты, а также глубины и высоты, хотя другие элементы, такие как доступность пищи и света, также играют роль в формировании предпочтительных мест обитания (Pörtner et al., 2006; Пёртнер, 2002; Prasad et al., 2011) ».
Результаты и обсуждение, третий абзац: неверно, что начало анаэробного метаболизма характеризует первую линию теплового ограничения. Процитированные статьи Пёртнера и его коллег относятся к так называемой концепции термостойкости, ограниченной кислородом и емкостью (OCLTT, ср. Pörtner 2010 J. exp Biol.). Самый ранний предел — это потеря производительности, вызванная несоответствием мощности подачи кислорода и связанных с этим затрат, а также гипоксемия, ограничивающая производительность, оставляя меньше энергии для e.грамм. рост (Пёртнер и Кнуст, 2007, наука). Таким образом, температурные ограничения приводят к началу анаэробного метаболизма. Эти находки недавно были рассмотрены в различных областях организма (Storch et al., 2014, Global Change Biology), что должно предостеречь авторов относительно некоторых из их утверждений о прокариотах и придать их обоснование более ясной форме .
Мы признаем, что наше письмо было слишком упрощенным в попытке удовлетворить широкую читательскую аудиторию, таким образом теряя часть своего смысла.Теперь мы должны четко упомянуть, что несоответствие в предложении и спросе кислорода определяет первый тепловой предел (Pörtner, 2002; Pörtner et al., 2006). Теперь мы также подробно описываем эксперименты как на водных организмах, так и на прокариотах, чтобы лучше направить читателя через то, что было достигнуто на данный момент, и что еще требует дальнейшей проверки перед нашей работой. См. Введение:
“ Постулируется, что подача кислорода играет роль в установлении температурных ограничений в многоклеточных организмах.[…] Даже в прокариоте E. coli (Morrison and Shain, 2008), предполагая, что ограничение энергии может быть общей чертой, которая характеризует жизнь на границе температурного диапазона ».
В конце введения: MASROS выглядит как дополнительный продукт OCLTT, следует упомянуть эту ссылку .
Действительно, MASROS основан на OCLTT, и теперь мы сначала объясним OCLTT во Введении, и только потом опишем, как он уступает MASROS. См. Введение:
«Размер тела уменьшается с повышением температуры в подавляющем большинстве эктотерм (« правило температуры-размера ») (Аткинсон, 1994; Форстер и др., 2012), тем самым увеличивая соотношение поверхности к объему и, следовательно, потенциально доступность кислорода.(…) Это привело к предположению, что изменения в размере клеток в ответ на изменения температуры в пределах температурного диапазона являются адаптивными реакциями для сохранения аэробной способности, что было названо гипотезой MASROS (Поддержание аэробных возможностей — регулирование подачи кислорода) (Аткинсон и др., 2006) ».
В эмбрионах снабжение кислородом может быть ограничено диффузионными ограничениями спроса и предложения .
Результаты и обсуждение, седьмой абзац: моделирование влияния гипоксемии на аэробный объем путем снижения содержания АТФ — отличный способ ответить на этот вопрос.Мне не хватало оценок изменения уровня АТФ? Возможны ли другие эффекты? При каких условиях подачи кислорода концентрация АТФ изменится in vivo?
Это действительно важный момент. Измерения уровней АТФ уже были выполнены на экстрактах взрослых червей, что выявило снижение уровня АТФ в ~ 2-5 раз в условиях, которые нарушают дыхательную цепь митохондрий (Dillin et al., 2002). Мы пытались провести аналогичные измерения на эмбриональных экстрактах, но наблюдали сбивающую с толку недостаточную воспроизводимость.Частично это может быть связано с тем фактом, что экстракты по необходимости получают из несинхронизированных эмбрионов (поскольку крупномасштабная синхронизация в этой системе невозможна) и что уровни АТФ могут значительно различаться между ранней и поздней стадиями эмбриогенеза.
Как мы сейчас обсуждаем в разделе «Результаты и обсуждение», действительно возможны и другие эффекты, включая изменения цитозольного pH или увеличение количества активных форм кислорода.
Насколько нам известно, неизвестно, при каких условиях кислорода уровни АТФ будут изменяться in vivo.Хотя это интересный момент, который заслуживает дальнейшего изучения, мы считаем, что он выходит за рамки основной области нашей рукописи.
Результаты и обсуждение, четвертый абзац: не следует говорить о тепловых ограничениях до их фактического определения. Является ли плато клеточного цикла ранним ограничением (например, пределом пежуса?)
Результаты и обсуждение, шестой абзац: то же самое, вам следует использовать все эти данные позже, чтобы вписать их в концептуальную основу ограничения .
Мы определяем температурные пределы как границы температурного диапазона, в котором вылупляется> 90% эмбрионов. Принимая во внимание новые измерения дыхания, представленные на рисунке 4A, мы полагаем, что эти предельные температуры очень похожи на пределы pejus, определенные другими (Pörtner, 2002).
Далее по тексту мы используем температурные пределы, определенные тестами на жизнеспособность эмбрионов, и исследуем различные клеточные функции в этих пределах. Интересно, что мы обнаружили, что тепловая реакция отдельных клеточных функций изменяется точно при предельных температурах, определенных тестами на жизнеспособность эмбрионов (см. Рисунок 1A).Теперь об этом упоминается в нашей рукописи в разделе «Результаты».
Это оставляет мою критику по поводу пробелов в интерпретации этих интересных данных. Изучение концепции OCLTT может дать основу и терминологию, необходимую для построения последовательной истории с учетом различных уровней теплового ограничения. Это также может побудить по-новому взглянуть на некоторые из их данных и на то, как их лучше всего интегрировать .
Мы сердечно благодарим рецензента за поддержку в этом направлении.В результате мы переписали всю рукопись, используя структуру и терминологию гипотез OCLTT и MASROS, и полагаем, что теперь текст стал более последовательным, а данные и интерпретации более надежными.
Рецензент № 3 :
Хотя это исследование предоставляет новые количественные наблюдения за термочувствительностью раннего эмбрионального развития, основные выводы статьи лишь слабо подтверждаются экспериментальными данными. Более того, представление и интерпретация данных, а также размещение наблюдений в их общем контексте часто неясны .
Мы согласны с рецензентом в том, что ключевые выводы, сделанные из экспериментов, представленных в первоначальной документации, требуют дальнейшей проверки. Как мы упоминали ранее в этом опровержении (см. Ответ на пункт 1 от обозревателя № 1), мы провели как измерения дыхания, так и тесты потомства у эмбрионов с нарушенным аэробным метаболизмом в широком диапазоне температур. Полученные данные убедительно подтверждают мнение о том, что аэробный метаболизм действительно ограничен температурными пределами.Мы также позаботились о том, чтобы данные были четко представлены и что наши интерпретации подробно обсуждались на всем протяжении новой интерпретации рукописи.
1) Связь между температурным воздействием на размер / форму эмбриона и аэробным метаболизмом явно не проверяется. Экспериментальные данные (RNAi) наводят на размышления, но не приводится ни одного эксперимента (или цитирования в литературе), который сообщил бы о температурно-зависимых изменениях аэробного метаболизма у эмбрионов. Следовательно, хотя данные по РНКи наводят на размышления, они явно слишком предварительные, чтобы поддерживать сильные выводы, сделанные .
Мы благодарим рецензента за предложение этого важного эксперимента, который также был предложен рецензентом №1. Мы любезно просим рецензента № 3 обратиться к ответу на пункт 1 рецензента № 1, где мы подробно объясняем, как была решена эта проблема.
2) Фактические данные обсуждаются недостаточно. Например, ряд измеренных фенотипов демонстрирует нелинейные температурные реакции, включая уменьшение размера эмбриона при более низких температурах (например, Рисунок 2E, 3E ).У C. elegans размер эмбриона кажется уменьшенным только при сравнении обработки 10C и 12C; Напротив, при сравнении всех температур от 12 ° C до 10 ° C размер эмбриона кажется увеличенным. Эти нелинейные реакции, на мой взгляд, несовместимы с гипотезой об аэробном метаболизме — однако эти экспериментальные данные не обсуждаются подробно .
Мы подтверждаем, что данные в пределах температурного диапазона, в частности, в отношении размера эмбриона, не обсуждались в достаточной степени в первоначальном представлении.У обозревателя №1 был аналогичный комментарий, к которому мы обращались выше (см. Ответ на его / его пункт 3). Как мы объясняем в этом абзаце, такие нелинейные реакции на самом деле очень подтверждают гипотезу об аэробном метаболизме.
3) Температурные пределы определяются как верхняя и нижняя температуры, при которых вылупление эмбрионов снижается до ~ 90%, но неясно, что приводит к этому снижению на 10%, например если этому способствуют изменения в раннем эмбриональном развитии. Более того, для данных, полученных за пределами температурных пределов, неясно, наблюдаются ли какие-либо изменения (например,грамм. форма эмбриона) способствуют увеличению эмбриональной смертности .
Этот момент понятен: мы действительно не знаем, какая стадия эмбрионального развития наиболее чувствительна к температурным пределам, а также не можем установить, вносят ли изменения, которые мы идентифицировали во время первого клеточного цикла, наблюдаемую летальность. Теперь мы явно указываем это ограничение в наших Заключениях. Интересно, однако, что мы обнаружили, что тепловая реакция отдельных клеточных характеристик изменялась точно при предельных температурах, определенных тестами на жизнеспособность эмбрионов, что указывает на минимально сильную корреляцию.
[Также: температурный диапазон не является общим термином, но должен быть явно связан с измеряемым фенотипом, например эмбриональная жизнеспособность, плодовитость, выживаемость. У данного генотипа будут разные термические пределы для этих разных фенотипов] .
Мы полностью согласны с рецензентом в том, что температурный диапазон не является общим термином, но всегда должен быть связан с каким-либо фенотипом. В нашей рукописи мы связали это с жизнеспособностью эмбриона. После определения этого диапазона в начале раздела «Результаты» мы затем спросили, легко ли наблюдаются клеточные изменения при тех предельных температурах, которые определены тестами на жизнеспособность эмбрионов, и неожиданно обнаружили, что это так для некоторых (см. Рисунки 2-3).
4) Данные сосредоточены на средних изменениях фенотипов развития, хотя наблюдаемые температурные эффекты (как и многие стрессоры в целом) сильно увеличивают дисперсию фенотипов при экстремальных температурах (например, дебафферинг). Это не анализируется и не обсуждается должным образом.
Мы благодарим рецензента за выделение этого момента. Фактически, мы представили данные в виде коробчатых диаграмм, чтобы убедиться, что среднее значение, дисперсия и любой потенциальный перекос в данных будут четко видны.Мы не обсуждали это в нашей рукописи, потому что мы проверили, как до подачи заявки, что дисперсия клеточных характеристик не увеличивалась за пределы теплового диапазона по сравнению с тепловым диапазоном. Это также можно увидеть в цветных прямоугольниках на рисунках 2-3, представляющих данные за пределами тепловых ограничений. Теперь это упоминается в легенде на Рисунке 2.
5) Эволюционный контекст, представленный для этого исследования, немного наивен, иногда ошибочен (например, первые два предложения Введения).Например, тепловая адаптация редко бывает видоспецифической чертой, но существует значительная генетическая изменчивость тепловых реакций внутри вида. Это важно также в контексте данного исследования: C. briggsae демонстрирует обширные генетические вариации в верхних температурных пределах репродуктивной способности с генетически отличными тропическими и умеренными кладами ( Prasad et al. 2011 ). [Кроме того, , C. briggsae встречается во многих холодных регионах по всему миру, ср. Результаты и обсуждение, последняя строка первого абзаца] .
Мы согласны с рецензентом в том, что изначально предоставленный эволюционный контекст был несколько наивным. Мы полностью осознаем, что тепловая адаптация, как правило, не является видоспецифической чертой, и что вступительные предложения в нашем первоначальном сообщении были чрезмерно упрощенными. Мы перефразировали первый абзац нашего введения, чтобы лучше учесть множество факторов, которые могут определять тепловой диапазон организма. Теперь он гласит:
.«Все организмы живут в пределах определенного температурного диапазона, за пределами которого рост и плодовитость снижаются (Pörtner et al., 2006). Частично в результате организмы имеют тенденцию распространяться в океане и на суше в соответствии с широтой, а также глубиной и высотой, хотя другие элементы, такие как доступность пищи и света, также играют роль в формировании предпочтительных мест обитания (Pörtner et al., 2006 ; Pörtner, 2002; Prasad et al., 2011). Несмотря на свою важность, механизмы, устанавливающие тепловые ограничения, остаются не до конца понятыми ».
Мы благодарим рецензента за то, что он отметил, что C. briggsae также встречается во многих холодных регионах, и поэтому наш текст был слишком общим.Мы немного изменили текст, чтобы отреагировать на этот момент (Результаты и обсуждение):
“ Таким образом, мы обнаружили, что температурные пределы для C. elegans составляли 12 ° и 25 ° C (Рисунок 1A), а для C. briggsae — 14 ° и 27 ° C (Рисунок 1B), в соответствии с тем фактом, что C. briggsae обычно обитает в более теплом климате, чем C. elegans (Prasad et al., 2011) ».
Аналогичным образом обсуждение литературы о влиянии температуры на аэробный метаболизм и т. Д.недостаточно ясно, чтобы следовать логике авторов .
Мы приносим свои извинения за путаницу, вызванную предыдущей версией рукописи, и искренне надеемся, что с этой новой версией теперь легче следовать нашей логике. Убедительно просим рецензента обратиться к нашим новым разделам «Введение» и «Результаты».
[Примечание редакции: ответы автора на повторную рецензию приводятся ниже.]
В целом, в рукописи представлено множество измерений (например, размер, форма, жизнеспособность), но как они интегрируются и как они связаны с аэробным метаболизмом и температурными пределами, остается в основном неясным.
Ниже мы резюмируем, как, на наш взгляд, наши выводы образуют связную историю. Мы также дополнительно прояснили эту последовательность во всем исправленном тексте рукописи.
1) Изменения размера и формы зародыша наблюдаются за пределами температурных пределов у эмбрионов C. elegans и C. briggsae .
2) Аэробный метаболизм, измеряемый как дыхание, снижен за пределы обоих температурных пределов у эмбрионов C. elegans .
3) Чтобы проверить, являются ли наблюдаемые изменения размера и формы за пределами тепловых ограничений результатом снижения аэробной способности (т.е. является ли пункт 1 выше результатом пункта 2 выше), мы проанализировали клеточные особенности в пределах температурного диапазона у эмбрионов, нарушенных для функции митохондриальной дыхательной цепи. Мы обнаружили, что и размер, и форма эмбриона изменяются в этих условиях, что подтверждает гипотезу о том, что такие изменения за пределами температурных пределов у дикого типа действительно отражают снижение аэробного метаболизма.
4) Эксперименты с дыханием показывают, что аэробный метаболизм снижается на пределе температур.Одной этой части данных недостаточно, чтобы различить сценарий, в котором энергетические потребности эмбриона не удовлетворяются из-за недостаточного аэробного метаболизма, и сценарий, в котором эти потребности фактически удовлетворяются в некоторой степени, несмотря на снижение дыхания, возможно, из-за того, что другие метаболические пути не выполняются. используется в большей степени. Интересно, что эти два сценария предсказывают разные исходы у эмбрионов с нарушением функции митохондриальной дыхательной цепи. Если аэробный метаболизм становится недостаточным за пределами температурного диапазона, то дальнейшее нарушение митохондриальной активности должно иметь большее влияние на тепловых пределах, чем на тепловом диапазоне.Напротив, если энергетические потребности могут быть удовлетворены, по крайней мере, до некоторой степени, несмотря на снижение дыхания за пределы теплового диапазона, то дальнейшее нарушение митохондриальной активности должно иметь меньшее влияние на тепловых пределах, чем на тепловом диапазоне. Важно отметить, что мы обнаружили, что последнее имеет место, демонстрируя, что аэробный метаболизм ограничен температурными пределами, как показано на Рисунке 4B-D. Мы тщательно переписали соответствующий раздел в отредактированной рукописи, чтобы еще больше прояснить этот важный момент (раздел «Результаты и обсуждение», подраздел, озаглавленный «Клеточные признаки температурных пределов повторяются при нарушении аэробного метаболизма»).
У животных, получавших РНКи, прямых измерений частоты дыхания не проводилось.
Следуя предложению автора обзора, мы решили проверить, что дыхание снижено у эмбрионов с нарушенной функцией митохондриальной дыхательной цепи. Поскольку АТФ-2 и NUO-1 являются частью комплексов V и I, соответственно, дыхание может все еще наблюдаться после их истощения, несмотря на нарушение дыхательной цепи митохондрий (см. Braeckman et al. 2009), так что измерения дыхания не обязательно говорят об этом. в этих случаях вместо этого дыхание должно быть уменьшено при нарушении функции комплекса III, как у эмбрионов cyc-1 (RNAi) .Как и ожидалось, мы действительно обнаружили, что дыхание у таких эмбрионов заметно снижено, составляя в среднем 56% ± 13% от уровня эмбрионального дыхания дикого типа в этих экспериментальных условиях (t-критерий p-значение <10 -3 ) . Эти новые результаты представлены в разделе «Результаты и обсуждение», в разделе «Клеточные признаки температурных пределов резюмируются при нарушении аэробного метаболизма» (см. Также обновленные соответствующие материалы и методы в подразделе , , озаглавленном «Измерение эмбрионального дыхания»).
Остается неясным, в какой степени аэробный метаболизм и эмбриональная летальность причинно связаны у животных веса .
Этот момент хорошо усвоен, и теперь мы явно указываем это ограничение в Результатах и Обсуждении (в подразделе, озаглавленном «Клеточные признаки температурных пределов повторяются при нарушении аэробного метаболизма»).
Статистический анализ новых данных кажется недостаточным, и экспериментальный план требует дополнительных пояснений.
Мы благодарим рецензента за обнаружение этого упущения; извиняюсь по этому поводу. Фактически мы провели анализ F-теста (как описано в дополнительном файле 1 для других измерений) и теперь добавили соответствующие p-значения в подраздел «Клеточные признаки температурных пределов повторяются при нарушении аэробного метаболизма» (F- значение p теста <10 -4 и значение p теста F <10 -10 ниже нижнего и выше верхнего теплового предела соответственно).
Мы также расширили текст в разделе «Материалы и методы», чтобы лучше объяснить экспериментальный план экспериментов с дыханием.
Авторы показывают, что эмбрионы на температурном пределе менее чувствительны к истощению митохондриальной активности РНКи, чем животные на тепловом диапазоне. На верхнем пределе температурного диапазона данные показывают, что истощение РНКи двух митохондриальных ферментов (но, что любопытно, не третьего) не влияет на жизнеспособность.Нет никаких средств контроля, показывающих, что истощение РНКи одинаково хорошо работает при всех температурах и для всех трех генов, поэтому возможно, что это странное поведение связано с неполным истощением при разных температурах и / или для разных генов.
Чтобы решить эту потенциальную проблему, мы провели тесты потомства при различных температурах в качестве показателя эффективности истощения с использованием РНКи против AIR-1, серин / треонинкиназы, необходимой для сборки веретена (Hannak et al. 2001), процесс неизвестен. быть связано с метаболическим статусом.Как показано ниже (см. Панель A), мы обнаружили, что air-1 (RNAi) был 100% эмбрионально летальным при 12 ° C, 20 ° C и 24 ° C, как и предполагалось в предыдущей работе (Hannak et al. 2001 ). Чтобы титровать фенотип, мы выполнили двойную РНКи путем смешивания бактерий, экспрессирующих дцРНК против air-1 , с бактериями, экспрессирующими дцРНК против gfp в соотношении 1: 3. Важно отметить, что в этом случае мы обнаружили, что летальность была выше при 12 ° C и при 24 ° C, чем при 20 ° C (см. Панель B). Следовательно, фенотип РНКи на самом деле сильнее для air-1 в направлении нижнего и верхнего теплового предела, чем при 20 ° C, что указывает на то, что результаты, которые мы обнаружили при нацеливании на компоненты дыхательной цепи митохондрий (рис. 4B-D рукописи), являются не из-за общего температурно-зависимого ответа РНКи.Эти результаты показаны на Рисунке 4 — в приложении 2 и представлены в разделе «Материалы и методы».
Кроме того, неясна логика, использованная для интерпретации этих экспериментов. Казалось бы, если дыхание становится ограничивающим на тепловом пределе, то эмбрионы на пределе должны стать более чувствительными, а не менее, к истощению дыхательных компонентов. Исходя из этого рассуждения, новые данные предполагают, что на самом деле дыхание не является этапом, ограничивающим скорость на тепловом пределе.
Мы согласны с рецензентом в том, что аэробный метаболизм не ограничивается тепловыми пределами (как мы писали в самом первоначальном сообщении), и приносим свои извинения за то, что, вероятно, не дали достаточно ясности по этому вопросу в нашем последнем сообщении. Как мы объяснили в пункте 4 выше, наши данные подтверждают механизм, посредством которого аэробный метаболизм ограничен (а не ограничен) в температурных пределах. Как уже упоминалось выше, мы значительно изменили основной текст, чтобы сделать его более ясным (раздел «Результаты и обсуждение» в подразделе, озаглавленном «Клеточные признаки температурных пределов, приводящие к нарушению аэробного метаболизма»).
Кроме того, рецензента 1 все еще беспокоила возможность того, что изменения в составе яичной скорлупы у эмбрионов, выращенных при разных температурах, могут привести к тому, что эмбрионы будут казаться больше или меньше из-за разной устойчивости к давлению покровного стекла, используемого для визуализации. Мы были бы признательны, если бы вы рассмотрели эту возможность при обсуждении ваших данных.
Мы обновили Материалы и методы, чтобы прямо указать на эту возможность.
https: // doi.org / 10.7554 / eLife.04810.014Разница между аэробным и анаэробным метаболизмом
Автор: Admin
Аэробный против анаэробного метаболизма
Клеточный метаболизм — это процесс преобразования углеводов, жиров и белков в энергию, необходимую клеткам. Во время метаболизма клеток энергия накапливается в высокоэнергетических фосфатных связях молекул аденозинтрифосфата (АТФ), которые служат энергетической валютой клеток.В зависимости от потребности в кислороде во время производства АТФ в клетке присутствуют два основных типа метаболизма; а именно аэробный и анаэробный. Из трех основных метаболических путей только гликолиз считается анаэробным метаболизмом, тогда как остальные, включая цикл лимонной кислоты (цикл Кребса) и цепь переноса электронов, считаются аэробным метаболизмом.
Аэробный метаболизм
Аэробный метаболизм происходит при наличии кислорода. Он происходит в митохондриях клетки и отвечает за обеспечение 90% потребности организма в энергии.Во время аэробного метаболизма весь основной субстрат, включая углеводы, жиры и белки, расщепляется и объединяется с молекулярным кислородом для производства энергии с выделением углекислого газа и воды в качестве конечных продуктов. В общем, окислительный метаболизм производит от 150 до 300 мл воды за 24 часа. В аэробном метаболизме участвуют два пути; цикл лимонной кислоты; который происходит в матриксе митохондрий и цепи переноса электронов; который происходит в системе транспорта электронов, расположенной во внутренней митохондриальной мембране.
Анаэробный метаболизм
Анаэробный метаболизм не требует кислорода для производства АТФ. Это происходит в результате гликолиза — процесса высвобождения энергии из глюкозы. Эффективность анаэробного метаболизма низкая, и количество АТФ вырабатывается низким по сравнению с аэробным метаболизмом. Гликолиз происходит в цитоплазме и не требует никаких органелл. Следовательно, это важный процесс, в котором у организмов отсутствуют митохондрии, такие как прокариоты.Конечным продуктом аэробного метаболизма является молочная кислота, которая может быть относительно вредной для организма.
Аэробный и анаэробный метаболизм
• Аэробный метаболизм требует кислорода, тогда как анаэробный метаболизм — нет.
• Анаэробный метаболизм не может продолжаться бесконечно. Напротив, аэробный метаболизм может продолжаться вечно, только в теоретических условиях.
• Углеводы, жиры и белки используются в качестве источников аэробного метаболизма, в то время как только углеводы участвуют в анаэробном метаболизме.
• Аэробный метаболизм включает в себя деятельность от низкой до умеренной интенсивности, тогда как анаэробный метаболизм включает только деятельность с высокой интенсивностью.
• Анаэробный метаболизм происходит в цитоплазме клеток, а аэробный метаболизм — в митохондриях.
• Аэробный метаболизм производит больше энергии, чем анаэробный метаболизм, если такое же количество того же субстрата.
• Гликолиз — это анаэробный метаболический путь, тогда как цикл лимонной кислоты и цепь переноса электронов — аэробные метаболические пути.
• Аэробный метаболизм вносит больший вклад (около 90%) в снабжение энергией, в то время как анаэробный метаболизм вносит меньший вклад.